Submitted:
15 September 2023
Posted:
18 September 2023
You are already at the latest version
Abstract
Brassicaceae are well known as non-arbuscular mycorrhizal (AM) host plants. This study was aimed to examine effects of two Brassica; cabbage (Brassica oleracea L. cv. cabitata) and mustard (B. juncea Coss.) on three AM species (Acaulospora, Entrophospora and Glomus) and to examine effect of root incorporation into soil on root AM infectivity in maize growing in iron toxicity soil. Experiment 1, cabbage and mustard crop reduced spore density of Acaulospora, but there was no effect on spore density of Entrophospora. While, mustard crop had more effect on Glomus spore density than cabbage crop. Experiment 2, AM spore from two conditions; without and with 6 weeks of root residues (cabbage, mustard and maize) incorporation was examined for its infectivity in maize roots. At D21, without root incorporated, the infectivity of Acaulospora, Entrophospora and Glomus were not different. By contrast, cabbage and mustard root incorporation depressed infec-tivity of Acaulospora but, there was no effect on Entrophospora. Whereas, cabbage root incorporation only depressed infectivity of Glomus. However, AM infectivity was recovered at D42. These results suggested that Brassica root incorporated into soil was the actual effect on the viability of AM spore resulting in reduction of AM infectivity at the early stage.
Keywords:
arbuscular mycorrhizal fungi
; iron toxicity soil
; non-Host plant
1. Introduction
Brassicaceae tissues contain significant quantities of the thioglucoside compounds as glu-cosinolates (GSLs). GSLs are hydrolysed by the myrosinase (β-thioglucoside glucohydrolases) enzyme to release a range of hydrolysis products including nitriles, oxazolidinethiones, thiocyanates and various forms of volatile isothiocyanates (ITCs). This hydrolysis products, in particular the ITCs, are known to have wide biocidal effects including phytotoxic [1], nematicidal [2] and fungicidal [3]. According to brassica green manures and rotation crops have been reported to suppress pest and disease organisms [4,5].
Plants belonging to Brassicaceae are well known as non-AM host plants. Many studies have been reported that the previous Brassica crop reduced AM root colonization and spore number in the subsequent crop. For example, Brassica rotation crop in Australia was reported that previous crop with canola (Brassica napus) reduced AM colonization in the subsequent AM-host (wheat) crop [6]. Similarly, decreasing in AM spore number and root colonization of sunflower and maize were occurred when growing after white mustard (B. alba L.) [7,8]. The recent study has been concluded that indolic glucosinolate production pathway in non-host Brassicaceae appears to play the role in reducing AM root colonization [9].
This study aimed to examine effects of Brassica on AM species and to determine effects of Brassica root residues incorporated into soil on the infectivity of AM fungi in maize roots. Three indigenous AM fungal species in red soil with iron toxicity; Acaulospora, Entrophospora and Glomus and two non-AM host plants; cabbage (B. oleracea L. cv. cabitata) and green mustard (B. juncea Coss.) were chosen. These AM species were dominant species in this soil and the previous study showed that they were sensitivity to Brassica crop [10]. The two Brassica have different types of ITC.
2. Materials and Methods
The study was consisted with 2 experiments as following. Experiment 1 was to apply three AM fungal species, Acaulospora, Entrophospora and Glomus to cabbage (B. oleracea cv. cabitata), mustard (B. juncea Coss.) and maize (Zea mays L., cv. suwan 4452 as AM host plant) in sterilized soil. The sterilized soil adding spore of AM fungi and leaving bare fallow was used as a control treatment. These 3 species of AM fungi were chosen because of their sensitivity to Brassica. Experiment 2 was to examine effects of root residues (from sub-experiment 1) incorporated into soil on viability of AM fungal spore to colonize in maize roots.
- Soil preparation
Soil sample belongs to Pak Chong soil series: clay-loam, kaolinitic, isohyperthermic, Typic Paleustults. The soil was collected at the depth of 0-15 cm (14° 38΄ N, 101° 19′ E, elevation 354 m above sea level, National corn and sorghum research centre, Thailand). The soil physical properties were clay soil with reddish brown (2.5YR 6/6). The soil chemical properties were pH 6.3 (1:1 soil:H2O), soil organic matter 23.5 g kg-1 (Walkley and Black method), available phosphorus (P) 18 mg kg-1 (Bray II) and extractable iron (Fe) 512 mg kg-1 (NH4OAc, pH 7.0). The soil was allowed to air dry, crushed with a mallet, roots removed by hand, well mixed and then sterilized twice by autoclave at 121 °C for 15 min.
- Experimental design
Experiment 1 Pot experiment was undertaken in completely randomized design with 3 replications consisting of factorial combinations of 3 AM fungal species (Acaulospora, Entrophospora and Glomus) and 4 crop regimes (fallow, cabbage, mustard and maize).
Sterilized plastic pots, 27 cm diameter at the top, 17 cm diameter at the bottom and 25 cm in height, were prepared as follows: a lower 13 cm deep layer of 3 kg of autoclaved soil was added and overlain with a 10 cm layer of mixture of 1 kg of soil AM inoculum (containing ca. 18000, 24000 and 36000 spores of Acaulospora, Entrophospora and Glomus, respectively) and 2 kg of autoclaved soil.
Plant seeds were sterilized by soaking in 0.5% sodium hypochloride solution for 10 min followed by rinsing several times with sterilized water. Seeds were placed onto the pot soil and then covered with ca. 2 cm layer of sterilized soil. Fertilizer was applied on the planting day. Nitrogen (N) was applied as urea at the rate of 1.9 g urea per pot (on a soil weight basis, equivalent to 210 kg N ha-1) on soil surface. P was applied as triple super phosphate (TSP, 0-46-0) by surface banding on one side of the pot at the rate of 0.70 g TSP per pot (equivalent to 32.75 kg P ha-1). Zinc (Zn) fertilizer was applied at the rate of 0.38 g of Zn per pot as Zn-EDTA (equivalent to 30.4 kg Zn ha-1). After seed emergence, 10 days after planting (D), seedlings were thinned to 3 plants per pot and grown under greenhouse conditions (35-40 °C). Pots were watered by spraying distilled water over the surface as required. Weeds and insects were removed by hand. No other chemicals were applied.
At D70, the roots were removed from the soil as much as possible and then washed carefully with tap water. Cleaned roots were placed in a sealed plastic bag and stored immediately in an ice box before moving to laboratory. The root sample was determined for fresh weight and then divided equal parts. The first root fraction was cut into 1-cm pieces by sterilized scissor and then stored in a sealed plastic bag at 4°C for using in Experiment 2. The remaining root fraction was determined for dry weight, AM colonization and ITCs content. The root fraction for ITC analysis was frozen immediately at -20 °C until required.
The soil without the roots was mixed thoroughly and subsampled ca. 200 g per pot for determining AM spore number. The rest of soil was placed in a sealed plastic bag and then stored at room temperature for two days before being used as soil sample in experiment 2.
Experiment 2 The soil sample from Experiment 1 was divided into equal parts (for with/without root incorporation) and then placed in sterilized plastic pots. The root fraction from Experiment 1 which was stored in sealed plastic bag at 4 °C for 2 days, was added into soil pot in treatment with root incorporation. The root was mixed throughout the soil with a sterilized spatula. This process was also undertaken with maize soil from Experiment 1. Distilled water was added to pots to field capacity (30% w/v) and then a transparent plastic bag was placed over the pot without being sealed. The pots were maintained at room temperature for 6 weeks. Maximum and minimum temperatures of incubation room were 32±3 and 25±3 °C, respectively (during late wet season, from September to October, 2022).
After soil-root incubation period, three maize seeds were planted in each pot. After seeds emergence (D10), maize was thinned to one plant per pot. Maize was grown in greenhouse conditions (35-40 °C) during the early dry season, from November to December, 2022. Fertilizer was applied on the planting day. N was applied as urea at the rate of 0.9 g urea per pot (equivalent to 210 kg N ha-1) on soil surface. P was applied as triple super phosphate (TSP, 0-46-0) by surface banding on one side of the pot at the rate of 0.35 g TSP per pot (equivalent to 32.75 kg P ha-1). Zinc (Zn) fertilizer was applied at the rate of 0.19 g of Zn per pot as Zn-EDTA (equivalent to 30.4 kg Zn ha-1). Pots were watered by spraying distilled water over the surface as required. Weeds and insects were removed by hand. No other chemicals were applied. At D21 and D42, each of 3 replications was harvested for determining AM colonization.
- Data collection
In Experiment 1, root sample was determined for dry weight, AM colonization and ITC contents and soil sample was determined for spore number. In Experiment 2, determination of AM colonization in maize root was undertaken.
- Determination of AM spore number
The soil samples were left to air-dry for determining the total AM spore number by the wet sieving and decanting method [11] followed by sucrose centrifugation [12].
- Measurement of AM root colonization
AM colonization was done by cleared in 10% KOH solution (w/v), stained with trypan blue (C. I. 23850, Ajex Finechem) [13], determined by making slides and viewing the roots with a compound microscope [14] and then the percentage of AM colonization was calculated by the method of Trouvelet [15].
- ITC analysis
The Brassica root fraction was determined the types of ITCs. Briefly, 5 g of fresh weight were de-frosted, finely chopped and then placed in 50 ml centrifuge tube. The 0.1 M CaCl2 and ether, each of 5 ml, were added into the centrifuge tube. The tube was shaken at 100 rpm for 30 min and then centrifuged at 239 g for 5 min. The supernatant was removed and placed at 4 °C prior to analysis. The extraction process was conducted twice producing ca. 10 ml ether extractant. The samples were identified for the types of ITCs by GC/MS. The analysis was undertaken by using a Thermo Scientific (ITO 900) equipped with a mass spectrophotometer detector. The column was a 30m x 0.25 mm x 0.25 μm (TR-5ms Thermo®). The temperature program was set at initial 35 °C for 3 min, at ramp 1 (12 °C min-1) 96 °C and ramp 2 (18°C min-1) 240 °C for 6 min. The post run condition was set at 300 °C with 5 min hold time. The sample was injected splitless by AI/AS 3000 autosampler at an oven temperature of 50 °C. The carrier gas used was helium at flow rate 1 ml min-1 and velocity 30 cm s-1. 2-phenylethyl ITC was used as standard.
- Statistical analysis
Percentage of AM root colonization was transformed by arc-sine for analyzing with ANOVA. All data were checked for normal distribution. Subsequently, data were subjected to analysis of variance with the SPSS. ANOVA was used to determine the main effects of AM fungal species and crop regimes and their two-way interactions. Duncan’s Multiple Range Test at P < 0.05 % was used for post hoc testing.T tests were used to compare means of two data sets.
3. Results
3.1. Experiment 1
- Spore density
AM species responded variously to Brassica (cabbage and mustard) crop. Mustard had more effect on spore density of Glomus than cabbage. Glomus spore density under mustard crop was lower (P<0.001) than those under cabbage crop by 16% (Figure 1). However, effect of cabbage or mustard on spore density of Acaulospora and Entrophospora did not significantly differ (Figure 1).
Brassica tended to reduce spore density but it was less severe than in the fallow treatment and maize tended to promote AM spore number. Compared to the start of cropping, the spore density of all species (Acualospora, Entrophospora and Glomus) decreased (P<0.001, 0.007, 0.002) by 61, 56 and 42%, respectively, under fallow (Figure 2, a-c). The spore density of Acaulospora declined (P=0.024, 0.025) by 33 and 35% under cabbage and mustard, respectively, (Figure 2, b-c), but was not changed under maize (Fig. 2, a). By contrast, spore number of Entrophospora was not affected by either cabbage or mustard but it increased (P=0.035) under maize by 19% (Figure 2, b). Furthermore, Glomus spore number fell (P<0.001, 0.001) under cabbage and mustard by 33 and 40%, respectively, but it increased (P=0.019) under maize by 20% (Figure 2, c).
- AM colonization
A few entry points of infection were observed in cleared and stained cabbage and mustard roots under the compound microscope. Colonization ranged from 1 to 3% in cabbage and mustard roots, to 34, 20 and 67%, respectively, in maize inoculated with Acaulospora, Entrophospora and Glomus. The symbiotic association with maize was characterized by numerous arbuscules and vesicles in root cortical cells, but these structures were absent in roots of the two Brassica.
- Root dry matter and ITC content of Brassica roots
Root dry matter of cabbage was higher (P<0.001) than of mustard. The dry matter of cabbage and mustard roots were 28.4±4.7 and 19.5±3.2 g pot-1 (n=12), respectively. Three ITC compounds were presented in cabbage roots, namely; Ally ITC, Propyl ITC and 2-Phenylethyl ITC. While, there were two ITC compound in mustard roots; Ally ITC and 2-Phenylethyl ITC. However, concentration of 2-Phenylethyl ITC in cabbage and mustard roots were 45±8 and 27±11 µg g-1 root fresh weight (n=4), respectively. Nitrile compounds were also presented; Butanennitrile, Pentanenitrile and Benzenepropanenitrile in cabbage, but Acrylonitrile and Benzenepropanenitrile in mustard.
3.2. Experiment 2
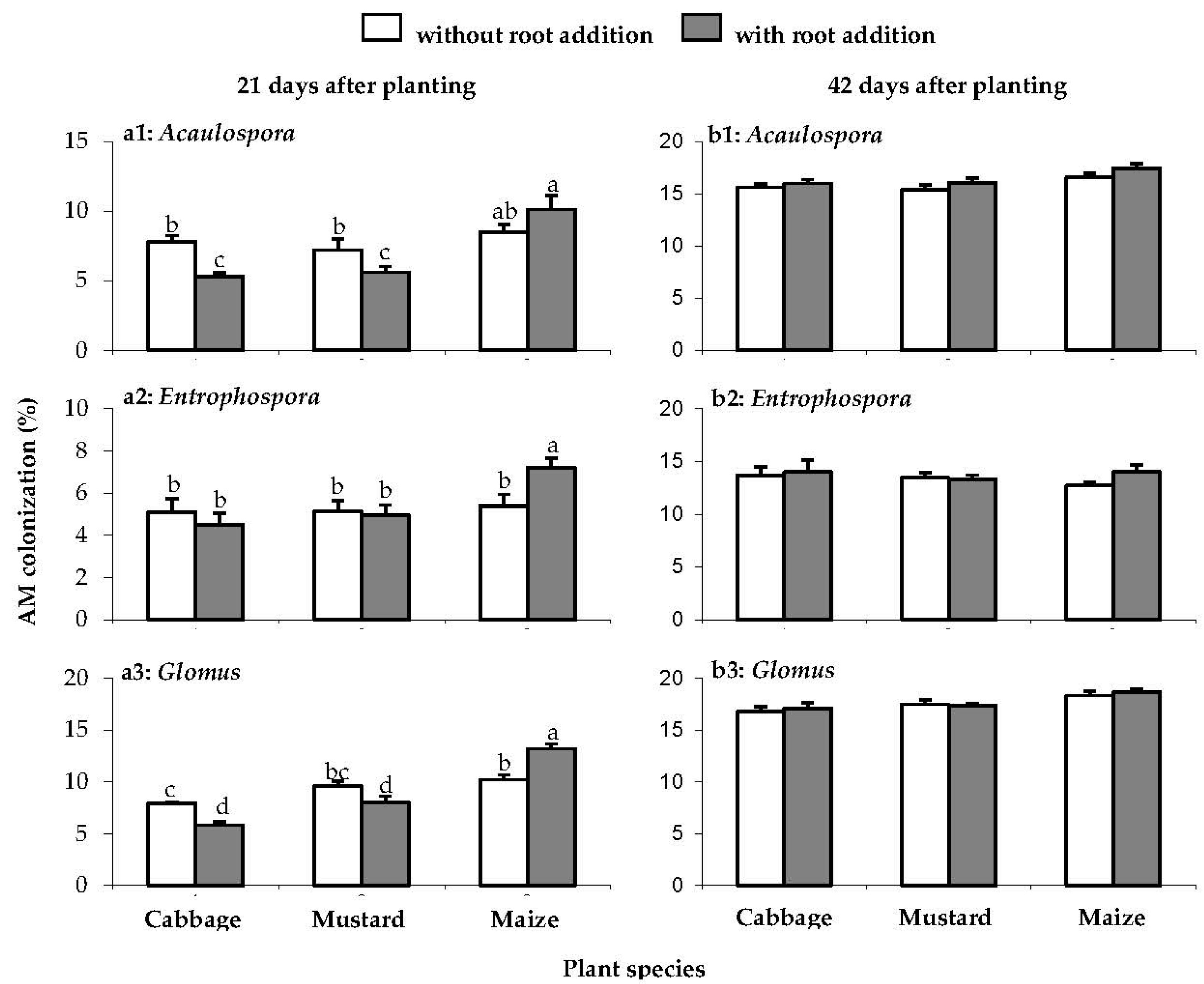
Incorporation with Brassica root residues into soil depressed the viability of AM spore resulting in reduction of (P<0.001) AM infectivity in maize roots. At D21, Acaulospora spores from cabbage and mustard root incorporated treatment was lower infectivity in maize roots by 32 and 22%, respectively when compared to percentage of colonization in the same treatment but without root incorporated (Figure 3, a1). Infectivity of Glomus decreased by 27% in cabbage root incorporated treatment but not in mustard root incorporated (Figure 3, a3). Whereas, Brassica root incorporated did not have effect on infectivity of Entrophospora (Figure 3, a2). By contrast, AM spore from maize root incorporated was higher colonization of Acaulospora, Entrophospora and Glomus by 18, 30 and 27%, respectively when compared to the same treatment but without root incorporated (Figure 3, a1-3).
The infectivity of AM spore from treatment without Brassica or maize incorporated did not differ (Figure 3, a1-3). At D21, root colonization by Acaulospora (Figure 3, a1), Entrophospora (Figure 3, a2) and Glomus (Figure 3, a3) in maize roots were 7.8±0.4, 5.2±0.2 and 9.2±0.7%, respectively (Figure 3, a1-3).
Previous crop with Brassica delayed AM root colonization in the subsequent maize crop. At D21, incorporation with Brassica roots decreased infectivity of AM spores in maize root when compared to the infectivity of AM spore from maize root incorporated treatment (Figure 3, a1-3). However, AM infectivity was recovered at D42. The percentage of AM root colonization did not differ between AM species (Fig. 3, b). Root colonization by Acaulospora (Figure 3, b1), Entrophospora (Figure 3, b2) and Glomus (Figure 3, b3) were 16.2±0.3, 13.5±0.2 and 17.6±0.3%, respectively.
4. Discussion
Brassica reduced AM density of three fungi; Acaulospora, Entrophospora and Glomus. However, it remains unknown how some AM spores disappeared without a host crop. Surprisingly, almost nothing has been published on this topic and there remains a need for detailed studies on spore longevity in field soils. However, one possible explanation is that ITCs could affect immature AM spores causing them to break down before the spore walls have matured. Anthony et al. [9] demonstrated that AM fungi failed to penetrate to roots of non-host Brassicales. The laboratory studies of Schreiner and Koide [16,17] showed that living roots of B. nigra and B. kaber inhibited the germination of Glomus spore. The pot with split-root system of Vierheilig et al. [18] also reported that two non-AM host plants (B. nigra and Beta vulgaris L.) inhibited colonization by G. mosseae in cucumber. From these previous studies, ITCs releasing from Brassica would be the primary effect on AM growth. Although, Brassica living roots do not actively release large amounts of ITCs because GSLs and myrosinase are compartmentalized in the cell. As long as this separation exists, ITC can be released only from injured cells where the spatial separation of GSLs and myrosinase is destroyed [19]. Rumberger and Marschner [20] reported that intact living canola roots continuously release a little amount of ITC via the outermost cell layers in rhizosphere, while the majority of root cells remain intact. This would be interesting to follow up in the future particularly where brassicas are being incorporated into the cropping cycle as biofumigants.
Brassica root residues incorporation into soil was the actual effect on the viability of AM spore resulting in reduction of AM infectivity. In Experiment 2, AM spore form two conditions; without and with root residues (cabbage, mustard and maize) incorporation was examined for its ability to colonize in maize roots. The infectivity of Acaulospora, Entrophospora and Glomus of treatment without cabbage or mustard or maize incorporated were not different. These results showed previous crop with either AM host or non-AM host did not have effect on the AM infectivity if the root residues were removed. By contrast, when cabbage or mustard root was incorporated, it depressed AM infectivity. This might be due to larger amounts of ITCs can be released by the breakdown of the cells during decomposition of dead plant material or even faster by incorporating green plant material into the soil. Many studies have been reported that previous Brassica crop reduced AM root colonization and spore number in subsequent crop [6,7,8].
Effect of Brassica root incorporated on AM fungal species was varied. Cabbage and mustard root incorporation depressed infectivity of Acaulospora, whereas, there was no effect on infectivity of Entrophospora. However, cabbage root incorporation only depressed infectivity of Glomus. This might be due to different types of ITC in root tissues of cabbage and mustard and ITC differ in their toxicity.
Three ITCs presented in cabbage roots, but there were two ITCs in mustard roots. 2-Phenylethyl ITC which have been reported as an effective biocide was presented 9.6±1.0 and 5.6±0.7 μg g-1 fresh weight of cabbage and mustard, respectively. Another possible explanation might be larger amount of cabbage root dry matter. Thus, cabbage would release more ITCs to soil than mustard. Furthermore, Sarwar et al. [21] found that 2-phenylethyl ITC (released from aromatic GSL) has been shown to be significantly more toxic to fungi than 2-Propenyl ITC (released from aliphatic GSL). ITCs produced by hydrolysis of aromatic GSLs such as 2-Phenylethyl is generally less volatile than aliphatic types and may therefore persist for longer in the soils [22].
Incorporation with Brassica roots into soils delayed AM colonization at the early stage of infection. However, at D42, AM colonization did not differ between AM species. Incorporation with either Brassica or maize did not have effect on AM infectivity in maize roots. This result was in accordance with the study of Gavito and Miller [23] who observed the maize following canola had significantly lower AM colonization for up to 62 days after planting after that the colonization was equal to maize following AM host species, bromegrass (Bromus inermis Leys.) and alfalfa (Medicago sativa L.). These observations suggest that AM populations can be built up and the inhibitory effects of a non-AM host crop can be reversed after cropping with AM host crop.
5. Conclusions
Incorporation of Brassica roots into soil was the primarily impact on AM development. Different Brassica various responded to AM fungal species. This may be useful in reducing the abundance of ineffective AM species in the field.
Acknowledgments
We thank Thammasat University Center of Excellence in Agriculture Innovation Centre through Supply Chain and Value Chain and department of Agricultural Technology, Faculty of Science and Technology, Thammasat University for providing experimental and laboratory facility. This work was supported by the Thailand Science Research and Innovation Fundamental Fund.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vercampt, H.; Koleva, l.; Vassilev, A.; Vangronsveld, J.; Cuypers, A. Short-term phytotoxicity in Brassica napus (L.) in response to pre-emergently applied metazachlor: A microcosm study. Environ. Toxicol. Chem. 2017, 36, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Charles, K.; Agathar, K.; Ronald, M.; Cosmas, P.; Ignitius, M.; Blessing, M. Nematicidal effects of brassica formulations against root knot nematodes (Meloidogyne javanica) in tomatoes (Solanum lycopersicum L.). Pak. J. Phytopathol. 2015, 27, 109–114. [Google Scholar]
- Ashiq, S.; Edwards, S.; Watson, A.; Blundell, E.; Back, M. Antifungal effect of brassica tissues on the mycotoxigenic cereal pathogen Fusarium graminearum. Antibiotics 2022, 11, 1249. [Google Scholar] [CrossRef] [PubMed]
- Larkin, R.P.; Lynch, R.P. Use and effects of different Brassica and other rotation crops on soilborne diseases and yield of potato. Horticulturae 2018, 4, 37. [Google Scholar] [CrossRef]
- Villalta, O.; Wite, D.; Riches, D.A.; Guiano, J.; Chandolu, V.; Scoble, C.; Donald, C.; Porter, I.J.; Mattner, S.W. The concentration of 2-propenyl glucosinolate in biofumigant crops influences their anti-fungal activity (In-Vitro) against soil-borne pathogens. J. Agri. Chem. Environ. 2016, 5, 38–45. [Google Scholar] [CrossRef]
- Bakhshandeh, S.; Corneo, P.E.; Mariotte, P.; Kertesz, M.A. Effect of crop rotation on mycorrhizal colonization and wheat yield under different fertilizer treatments. Agric. Ecosyst. Environ. 2017, 247, 130–136. [Google Scholar] [CrossRef]
- Karasawa, T.; Kasahara, Y.; Takebe, M. Variable response of growth and arbuscular mycorrhizal colonization of maize plants to preceding crop in various types of soils. Biol. Fertil. Soils 2001, 33, 286–293. [Google Scholar] [CrossRef]
- Karasawa, T.; Kasahara, Y.; Takebe, M. Differences in growth responses of maize to preceding cropping caused by fluctuation in the population of indigenous arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2002, 34, 851–857. [Google Scholar] [CrossRef]
- Anthony, M.A.; Celenza, J.L.; Armstrong, A.; Frey, S.D. Indolic glucosinolate pathway provides resistance to mycorrhizal fungal colonization in a non-host Brassicaceae. Ecosphere 2020, 11, e03100. [Google Scholar] [CrossRef]
- Poomipan, P.; Suwanarit, A.; Suwanarit, P.; Nopamornbodi, O.; Dell, B. Reintroduction of a native Glomus to a tropical Ultisol promoted grain yield in maize after fallow and restored the density of arbuscular mycorrhizal fungal spores. J. Plant Nutr. Soil Sci. 2011, 174, 257–268. [Google Scholar] [CrossRef]
- Gerdeman, J.W.; Nicolson, T.H. Spores of mycorrhizal Endogone extractable from soil by wet sieving and decanting. Trans. Br. Mycol. Soc. 1963, 46, 235–244. [Google Scholar] [CrossRef]
- Daniels, B.A.; Skipper, H.D. Methods for the recovery and quantitative estimation of propagules from soil. In Method and Principles of Mycorrhizal Research; Schenck, N.C., Ed.; Amer. Phytopath. Soc.: Minnesota, USA, 1982; pp. 29–35. [Google Scholar]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi. New Phytol. 1970, 124, 481–488. [Google Scholar]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Trouvelet, A.; Kough, J.L.; Gianinazzi, V.P. Measure du taux de mycorrhization VA d’un systeme radiculaire. In Physiological and Genetical Aspects of Mycorrhiza; Gianninazzi, V.P., Giannazzi, S., Eds.; LNRA: Paris, 1985; pp. 217–221. [Google Scholar]
- Schreiner, R.P.; Koide, R.T. Antifungal compounds from the roots of mycotrophic and non-mycotrophic plant species. New Phytol. 1993, 123, 99–105. [Google Scholar] [CrossRef]
- Schreiner, R.P.; Koide, R.T. Mustards, mustard oils and mycorrhizas. New Phytol. 1993, 123, 107–113. [Google Scholar] [CrossRef]
- Vierheilig, H.; Lerat, S.; Piché, Y. Systemic inhibition of arbuscular mycorrhiza development by root exudates of cucumber plants colonized by Glomus mosseae. Mycorrhiza 2003, 13, 167–170. [Google Scholar] [CrossRef]
- Bones, A.M.; Rossiter, J.T. The myrosinase-glucosinolate system, itsorganization and biochemistry. Physiol. Plant 1996, 97, 194–208. [Google Scholar] [CrossRef]
- Rumberger, A.; Marschner, P. 2-Phenylethylisothiocyanate concentration and microbial community composition in the rhizosphere of canola. Soil Biol. Biochem. 2003, 35, 445–452. [Google Scholar] [CrossRef]
- Sarwar, M.; Kirkegaard, J.A.; Wong, P.T.W.; Desmarchelier, J.M. Biofumigation potential of brassicas. Plant Soil 1998, 201, 103–112. [Google Scholar] [CrossRef]
- Brown, P.D.; Morra, M.J. Control of soil-borne plant pests using glucosinolate-containing plants. Adv. Agron. 1997, 61, 167–231. [Google Scholar]
- Gavito, M.E.; Miller, M.H. Changes in mycorrhiza development in maize induced by crop management practices. Plant Soil 1998, 198, 185–192. [Google Scholar] [CrossRef]
Figure 1.
The spore number of Acaulospora, Entrophospora and Glomus under fallow, cabbage, mustard and maize in Experiment 1. Bars are means ± SE (n = 3). Bars with common letters are not significantly different at P < 0.05 (DMRT).
Figure 1.
The spore number of Acaulospora, Entrophospora and Glomus under fallow, cabbage, mustard and maize in Experiment 1. Bars are means ± SE (n = 3). Bars with common letters are not significantly different at P < 0.05 (DMRT).
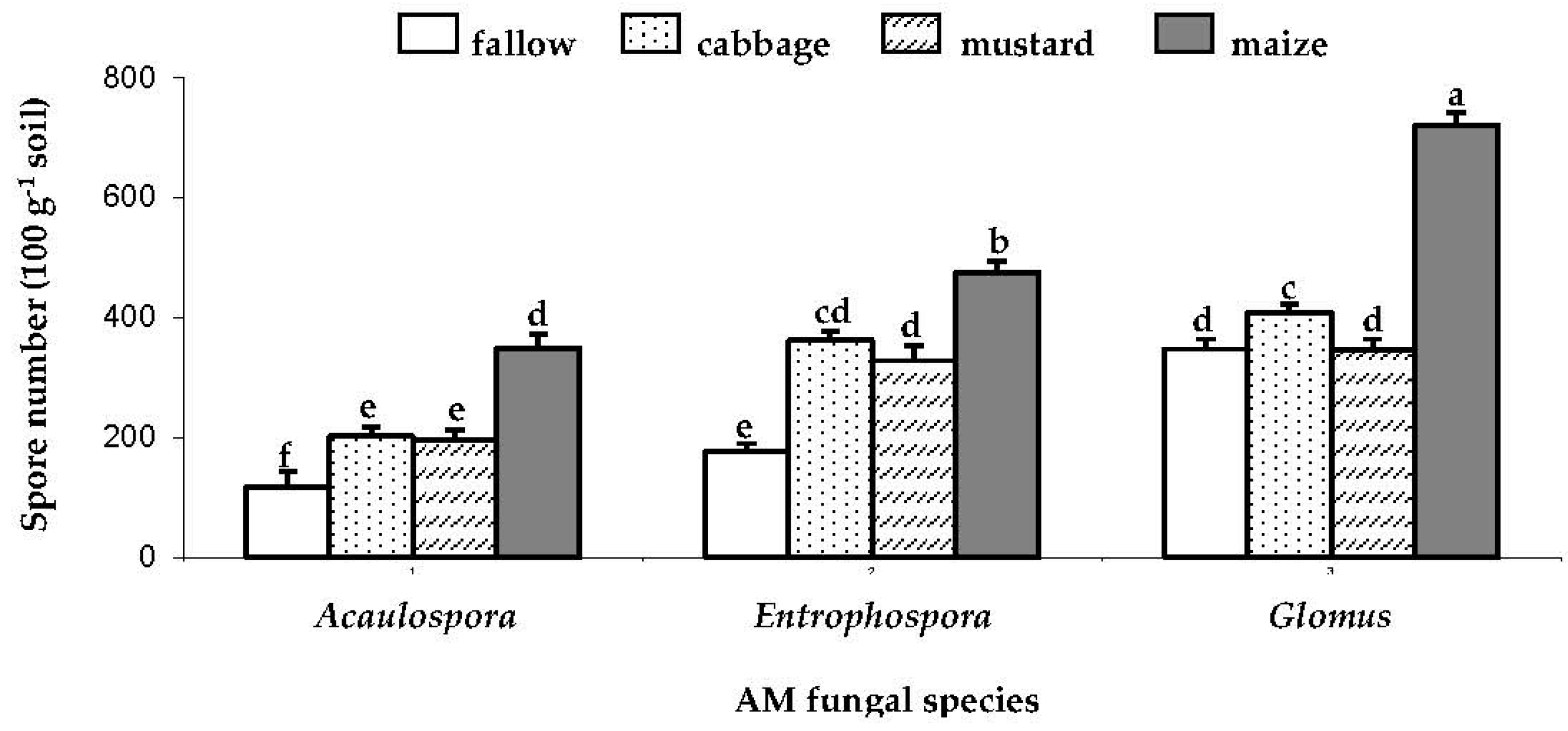
Figure 2.
The spore number of Acaulospora (a), Entrophospora (b) and Glomus (c) at the start and the end of Experiment 1. Bars are means ± SE (n=3). Non- significant differences at P < 0.05 by t test are shown by “ns”.
Figure 2.
The spore number of Acaulospora (a), Entrophospora (b) and Glomus (c) at the start and the end of Experiment 1. Bars are means ± SE (n=3). Non- significant differences at P < 0.05 by t test are shown by “ns”.
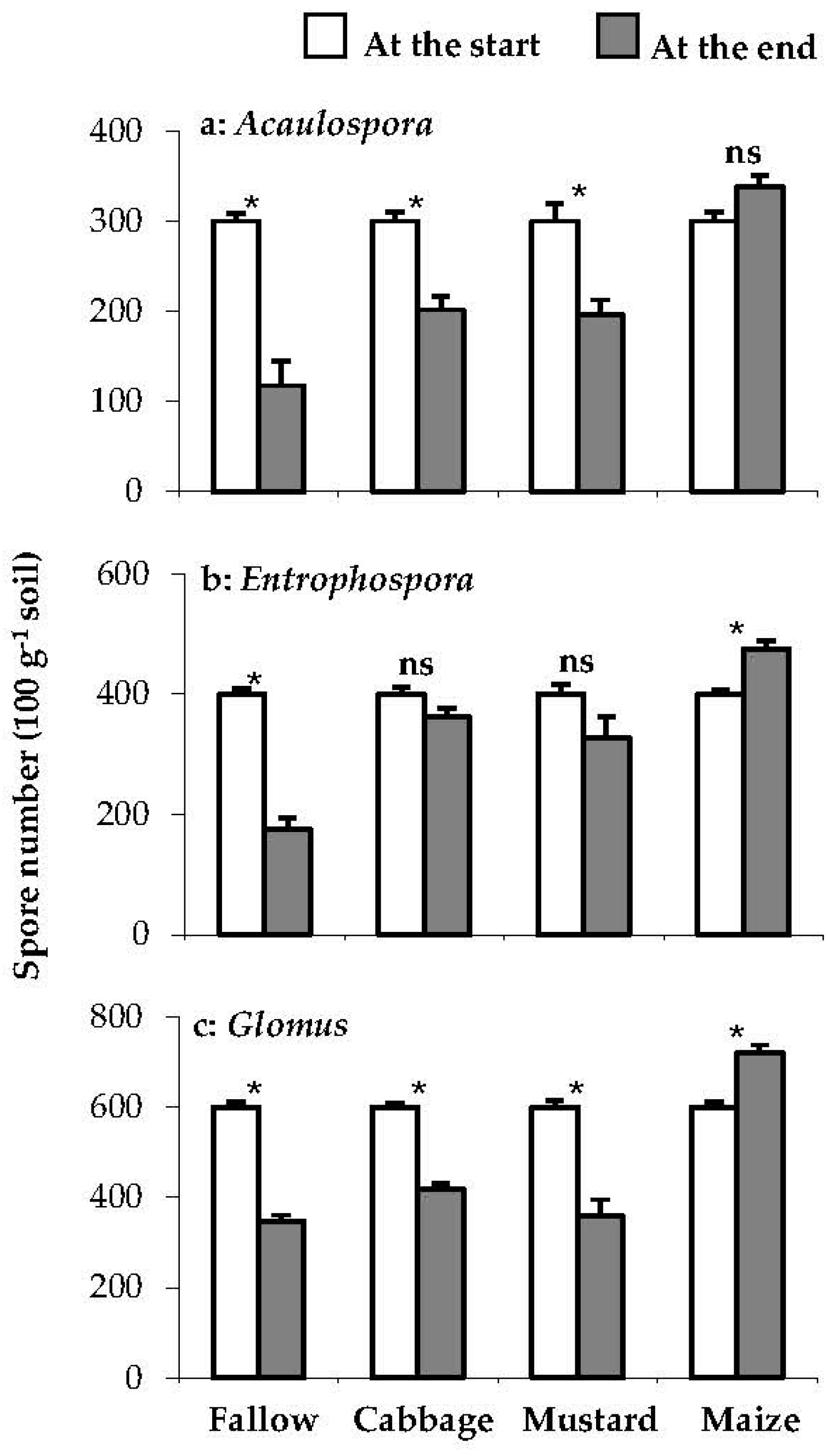
Figure 3.
The AM colonization of Acaulospora (a1, b1), Entrophospora (a2, b2) and Glomus (a3, b3) in maize roots in Experiment 2 at D21 (left) and D42 (right). Bars are means ± SE (n = 3). Bars with common letters are not significantly different at P < 0.05 (DMRT).
Figure 3.
The AM colonization of Acaulospora (a1, b1), Entrophospora (a2, b2) and Glomus (a3, b3) in maize roots in Experiment 2 at D21 (left) and D42 (right). Bars are means ± SE (n = 3). Bars with common letters are not significantly different at P < 0.05 (DMRT).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



