
Submitted:
05 September 2023
Posted:
05 September 2023
You are already at the latest version
Abstract
Catalase can decompose hydrogen peroxide in plants under controlled and stressed condi-tions. In a stressed environment, the increase in hydrogen peroxide production makes catalase activity a major defense system for plant growth. The current study was conducted to evaluate the catalase activity of pre-isolated and identified bacterial strains Bacillus aryabhattai (AN30), Bacillus megaterium (AN24), Bacillus megaterium (AN31) and Bacillus megaterium (AN35) and their potential for rice seedlings growth promotion. These strains were characterized for quantita-tive catalase, urease, siderophores, and exopolysaccharides production using LB media. Sub-sequently, the effectiveness of these strains was checked by quantifying the catalase activity in the rhizosphere of rice seedlings. Furthermore, the catalase activity was quantified in the roots and shoots of rice seedlings. Plant growth parameters were also studied in a growth room trial. The strain AN24 showed the maximum catalase activity (1.36 mol cm-1), urease ac-tivity (1.35 mol cm 1), exopolysaccharides (4.20 µg mL-1), and siderophores (2.32%) production in LB media. All tested strains showed significantly higher catalase activity in soil compared to control. Among sole application, strain AN24 showed better results; however, consortium application of strains AN24+AN30+AN35+AN31 showed the maximum increase in catalase ac-tivity in the rhizosphere of rice seedlings. Moreover, consortium application of AN24+AN30+AN31+AN35 resulted in the highest increase in catalase activity, dry biomass of shoots and roots, shoot and root length of rice seedlings. The results showed that a consortium of these Bacillus strains with catalase activity has greater potential to enhance an antioxidant defense system and growth promotion of rice seedlings. However, further experimentation under natural conditions is required before using these strains as potential bioinoculants for improving the rice growth and yield.
Keywords:
Antioxidative enzymes
; Bacillus sp.
; Catalase activity
; Exopolysaccharide production
; Reactive oxygen species
1. Introduction
Enzymes are mainly derived from crops, animals, and microbes covalently attached, linked, and incorporated inside soil debris microcapsules [1]. They are also used for reactive species decomposition. In plants, reactive species formation is the by-product of normal metabolic processes [2]. Hydrogen peroxide is the most active reactive specie, formed in peroxisomes under stress conditions [3], and also during metabolic processes [4]. It performs a dual function for plants: at optimum level, it can act as signaling molecule for plant growth [5] while its higher concentration damages the plant biomolecules and ultimately causes death of plants [2].
Catalase is an important intercellular enzyme, found in aerobic and facultative anaerobic microorganisms [6]. It is a heme containing enzyme belonging to the oxido-reductase family, a widely used industrial enzyme with the highest turnover rate [7] and well-defined reaction system [8], offering multiple diagnostic and therapeutic applications [9]. Its function is to remove hydrogen peroxide in plants by breaking it into water and oxygen and also to protect the cells [10]. For plants, it can serve as defense molecule and help them to grow [11]. It is also used to detoxify the hydrogen peroxide produced in various processes such as electron transport chain, photo respiratory oxidation and fatty acids oxidation [3].
Rice is a staple crop that follows the C3 photosynthetic pathway (Calvin cycle). Chloroplasts and stromules occupy 95 percent of the cell periphery in rice cells. While peroxisomes and mitochondria, which are important for photorespiration in the cell, are lined up along the chloroplast wall [12]. In the C3 pathway, hydrogen peroxide is produced because of photorespiration [13] which can affect crop growth, yield, and physiology. Here catalase enzyme’s presence is important to protect them. In addition, it is studied in dicotyledonous plants [14], but there are few studies on monocotyledonous plants such as rice [15]. During rice germination process, catalase enzyme is crucial for the hydrolysis of seed’s endosperm, which allows its development providing necessary energy for root and shoot development. This interesting anatomy of rice mesophyll cells with catalase enzyme is most important in searching for photorespiratory CO2 to improve Rubisco's carboxylation capability for CO2 refixation [16]. Therefore, catalytic induction and catalase production can improve plant establishment and growth of the rice plant [17]. The catalase enzyme activity is also directly related to the production of organic acid that lowers the pH and improves the availability of nutrients [18]. The application of citric acid significantly increased catalase activity in rice [18]. Citric acid, as a supplement, increases catalase enzyme activity, which reduces the damaging effect of reactive oxygen species. According to a recent study, exogenic utilization of citric acid increases the activity of antioxidant enzymes, demonstrating the plant's defense mechanism against oxidative stress [19].
Microbial enzymes can be used to overcome problems caused by various environmental conditions that can damage the physiology of plants, therefore microbial enzymes have replaced plant enzymes because of genetic operation [20]. For resisting pathogen attack, plant growth promoting rhizobacteria (PGPR)-mediated immune systemic resistance (ISR) provides defense mechanism through enzyme production (i.e., catalase) and release of allelopathic compounds [21]. Bacteria have two types of catalase enzymes, one which catalyzes only H2O2 and the other also has peroxidase activity. Catalase catalyzes the transformation of H2O2 to water and oxygen in bacteria [6]. It is also the defensive mechanism for bacteria against stress [6]. It may also contribute to various cellular processes such as cell expansion, variation, and metabolite production [22]. However, bacteria are a diverse group of microorganisms that can grow under harsh environmental condition. Therefore bacterial strains with catalase activity can be helpful to improve plant growth under changing climatic conditions [23]. They are mostly favored due to their higher yield, economic viability, product optimization and alteration and fast growth of microbes in an inexpensive media. Under changing climate, the role of rhizobacteria having ability to produce stress enzymes, hormones, exopolysaccharides and siderophores becomes crucial which can play important role for solubilizing the fixed organic and inorganic nutrient sources [24].
Plants need an optimum level of hydrogen peroxide for their growth and therefore produce a specific amount of catalase for their protection. Various strategies are used to eliminate hydrogen peroxide in plants. One of them is the catalase production by microbes that are able to maintain the level of hydrogen peroxide in plants. Keeping in view, the importance of catalase, rice, and rhizobacterial Bacillus strains, we hypothesized that the production of catalase ability of Bacillus strains can support plants and bacteria and to maintain the growth and physiology of rice under normal conditions. The objective of the study was to evaluate the potential of pre-isolated Bacillus strains for quantitative catalase, urease, exopolysaccharides, and siderophores production ability. The effectiveness of these strains was also evaluated for the promotion of rice seedlings growth under controlled conditions in a growth room trial.
2. Results
2.1. Biochemical characterization of Bacillus strains
The biochemical characterization of tested strains is presented in Figure 1. All strains show different behaviour in terms of catalase activity, urease activity, siderophores and exopolysaccharide (EPS) production in broth culture. Statisitacal analysis revealed that catalase (Figure 1a) and urease (Figure 1b) activity were similar in strains AN31 and AN35. Moreover, EPS (Figure 1c) and siderophores (Figure 1d) production were statistically different in all tested strains. However, the strain AN24 showed the maximum catalase activity (1.36 mole cm-1), urease activity (1.35 mole cm-1), EPS production (13.5 µg mL-1) and siderophores production (23.0 µg mL-1).
2.2. Effect of catalase-producing Bacillus strains to improve catalase activity in soil and plant
The efficacy of Bacillus strains with catalase activity were evaluated for improvement in catase activity in soil and plant (shoot and root). The tested strains significantly improved catalase activity in soil and plant (Figure 2). In sole inoculation, the Bacillus strain AN24 gives the maximum increase in catalase activity in soil (24%), shoot (27%) and root (30%) as compared to uninoculated control. However, co-inoculation performed better results than sole inoculation. Consortium application of AN24+AN35+AN31+AN30 resulted in the maximum increase of 43, 51, and 55% in catalase activity in soil, shoot and root, respectively.
2.3. Organic acid determination in bacterial culture
High performance liquid chromatography (HPLC) analyses depicted the production of organic acids by tested strains in liquid culture (Figure 3). Production of citric acid, pyruvic acid and oxalic acid was observed by all tested strains. Maximum pyruvic acid (43 µg mL-1) was produced by strain AN30. The strain AN24 produced 10 and 125 µg mL-1 of melonic acid and succinic acid, respectively. Whereas, other tested strains did not produce melonic acid and succinic acid. Fumeric acid was observed by strain AN31 (64 µg mL-1) and AN35 (120 µg mL-1). Bacterial cultures of tested strains were analyzed for phenolic compounds. The HPLC analysis revealed that the only strain which produced galic acid (0.61 µg mL-1) was AN24. Whereas, quercetin was produced by all the tested strains. However, the maximum (2.24 µg mL-1) quercitin was produced by strain AN35 followed by strain AN31 with 2.01 µg mL-1.
2.4. Plant growth promotion by the application of catalase-producing Bacillus strains
The application of Bacillus strains having catalase activity significantly improved the growth and root morphology of rice seedlings (Table 1). Among sole inoculation, the strain AN24 showed the maximum of 28, 25 and 35% increase in shoot length, root length and seedling dry weight, respectively. However, consortium application of AN24+AN35+AN31+AN30 showed even better results. This treatment showed the maximum increase in shoot length (46%), root length (55%) and seedling dry weight (56 %) as compared to uninoculated control. Similarly, the maximum root surface area, root diameter and root volume was observed by consortium application of AN24+AN35+AN31+AN30 which showed 33, 62 and 46% increase in above mentioned parameters, respectively.
2.5. Effect of Bacillus strains with catalase activity on soil biological properties
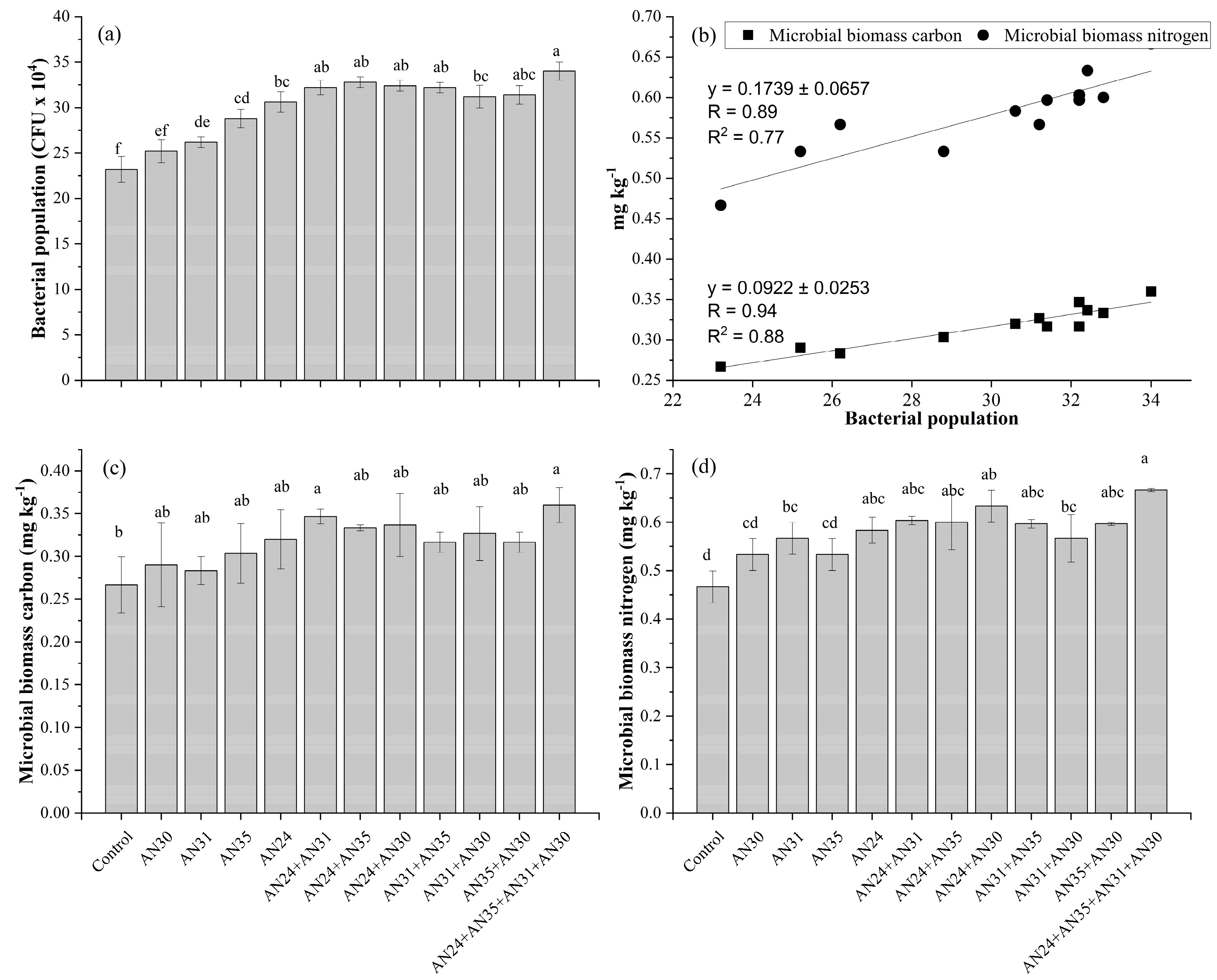
The catalase-producing Bacillus strains effect on soil biological properties is explained in (Figure 4). All the applied treatments showed significant increase in bacterial population in rhizospheric soil, however co-inoculation and consortium application performed better than sole inoculation (Figure 4a). Among sole inoculation, the maximum increase was observed (24%) in bacterial population by strain AN24. The highest increase (32%) was observed by consortium application of AN24+AN35+AN31+AN30 followed by co-inoculation of AN24+AN35 and AN24+AN31 which showed a 29.3% increase in bacterial population in rhizospheric soil. Microbial biomass carbon (MBC) (Figure 4c) and microbial biomass nitrogen (MBN) (Figure 4d) was significantlty different in applied treatments in a comparison with uninoculated control. Whereas, sole and co-inoculation showed statistically non-significant results than each other. However, consortium application of AN24+AN35+AN31+AN30 showed significantly higher results than all applied treatments where 26 and 30% increase in microbial biomass carbon and nitrogen, respectively, was observed. Regression and correlation between bacterial population and microbial biomass carbon and nitrogen showed straight line which indicated the direct relation between tested parameters (Figure 4b). A positive relation between bacterial population and microbial biomass carbon was recorded with a Pearson correlation coefficient of R = 0.94 and a linear regression coefficient of R² = 0.88. Furthermore, bacterial population and microbial biomass carbon showed a direct relation at R = 0.89) and R² = 0.77.
Figure 5.
Effect of Bacillus strains with catalase activity on soil biological properties. Bars having the same letter(s) did not differ from one another statistically at p≤0.05.
Figure 5.
Effect of Bacillus strains with catalase activity on soil biological properties. Bars having the same letter(s) did not differ from one another statistically at p≤0.05.
3. Discussion
The growth of plants stimulated by use of microbial inoculants for healthier food and improves sustainability to food supply [25]. Bacteria produce siderophores and EPS, which are high molecular weight polymers and iron scavengers that help bind soil particles and improve soil aggregation. The catalase and urease enzymes are essential elements for the hydrolysis of endosperm in seed and boost the energy level of root and shoot development. The catalytic rhizobacterial strains play important role in several microbial processes independent of oxidative stress, removes hydrogen peroxide (H2O2) from the cells, therefore catalase production by Bacillus strains proved significant for hydrogen peroxide detoxification in plants or in microbes for their survival [26]. In the present study, the Bacillus strains showed positive results for production of catalase with high catalytic activity and removal of hydrogen peroxide (H2O2) [27]. Microbes with catalase production have different mechanisms: breakdown of organic matter, promotion of plant growth and protection against reactive oxygen species [27].
The identified rhizobacterial strains showed a significant increase in urease activity in rice seedlings, suggesting that these Bacillus strains enhance nitrogen utilization to plants and promote root growth [28]. It has been reported that rhizobacterial strains such as B. pasteurii and B. megaterium potentially enhance hydrolysis of urea and stimulate urease activity [29]. In addition to nutrient solubility, Bacillus strains increase the amount of siderophore and have high affinity to iron. Microbial siderophores are iron-scavenging, low molecular weight ligands that are produced in the case of iron deficiency [30]. Moreover, the Bacillus strains exhibited variable growth with increasing EPS concentration and improved root and shoot growth with root adhering ability in soil. Our results are consistent with those of various Bacillus strains that produce EPS such as Bacillus licheniformis and Bacillus subtilis. These strains have higher molecular weight polymers such as proteins, carbohydrates, and other organic compounds [31]. The EPS thus released by bacteria to trigger the plant and are associated with the formation of an adaptive mechanisms under stressful conditions.
Plant growth promoting rhizobacterial Bacillus strains having catalase activity also showed organic acids production. In the soil, organic acid production increased the hydrogen ion concentration by solubilizing nutrients which improved plant growth [32]. The results were similar to those in previous literature [19]. Citric acid significantly increased catalase activity, demonstrating its protective function against oxidative stress in crops. In our study, all Bacillus strains produced organic acids like succinic acid, pyruvic acid, citric acid, gallic acid, oxalic acid, fumaric acid and quercetin. Similarly, our study was also in line with Mumtaz et al. [33] who reported about Bacillus sp. strain ZM20, Bacillus cereus and Bacillus subtilis strain ZM63 produced citric acid, acetic acid, formic acid, lactic acid, isovaleric acid, succinic acid, formic acid, and isobutyric acid.
The identified Bacillus strains were inoculated with rice seeds and tested for their effects in plant growth promotion and root development. The inoculation with rhizobacterial Bacillus strains improved root colonization, root surface area, root diameter, dry biomass and root/shoot length in current study. The strains showed different responses and maximum growth with the inoculation of catalase producing Bacillus strains in rice seedlings. However, a consortium of Bacillus strains (AN24+ AN35+ AN30 +AN31) showed the maximum growth promotion. Our study was also consistent with previous research that these rhizobacterial bacillus strains improve the length of plant roots, thus increasing water and nutrients uptake, growth and seedling physiology [34]. Another study also proved catalase production by Bacillus strains and its possible influence on plant growth [35]. This percentage increase in plant growth parameters is due to the inoculaned rice seedlings stimulating the growth-promoting abilities of the bacterial isolates. Our study showed that the microbial catalytic activity significantly improved the catalase activity in both, soil and plant. Maximum catalase activity was observed by the consortium application of AN24+ AN35+ AN30 +AN31. The catalase enzyme is closely linked with the rhizosphere of rice seedlings and enzymes released by roots, its expansion and tolerance mechanisms [36]. Soil enzymes can positively influence the plant growth through catalase, involving various biochemical reactions that improve soil health and plant growth [37]. Furthermore, the present study demonstrated that the use of catalytic Bacillus strains enhanced soil chemical and biological properties. The microbial biomass carbon, microbial population in rhizosphere and microbial biomass nitrogen significantly improved by the inoculation of catalytic strains. It is well documented that the seed inoculation significantly increased bacterial population [38]. The bacterial population improved the amounts of nitrate-nitrogen, ammonium-nitrogen, and microbial biomass carbon [39]. This was also reported by Iqbal et al. [40]. The treatments applied in our study showed a remarkable improvement in microbial carbon biomass and act as substrate for nutrient availability in soils, hence improving their biochemical properties and organic matter.
4. Materials and Methods
4.1. Quantitative characterization of Bacillus strains:
In the present study, the following pre-isolated and identified Bacillus strains AN24 (Bacillus megaterium), AN35 (Bacillus megaterium), AN30 (Bacillus aryabhattai) and AN31 (Bacillus megaterium) were used, provided by the Soil Microbiology and Biotechnology Laboratory, Department of Soil Science, the Islamia University of Bahawalpur. These strains have plant growth promoting traits and are well characterized for this purpose, and the ability to produce indole-3-acetic acid (IAA), catalase, urease, exopolysacchride and siderophore production ability has been positively reported [32]. The current study has been planned for quantitative characterization of strains for catalase activity and their role in plant growth promotion.
The Bacillus strains were assessed for catalase activity in broth culture as studied by Beers et al. [41] and Hildebrandt et al. [42]. A nutrient broth medium (1 liter of distilled water containing 3 g yeast extract, 10 g peptone, and 1g NaCl, (pH 7.2) was used. The overnight grown bacterial strains having 0.6 OD600 were further inoculated in nutrient broth medium and incubated in a rotary shaker at 30 °C for 24 h. To get crude extracts, cultures were centrifuged at 4500 rpm for 10 min. Then, 1.5 mL of a 30 mM H2O2 solution was added to 0.1 mL of extract in 2.7 mL of buffer (50 mM Na2HPO4/NaH2PO4). To calculate the quantitative catalase production, a spectrophotometer (Model Carry 60, Agilent Technologies Santa Clara, CA) absorbance was recorded at 240 nm.
4.2. Biochemical quantification of strains
The Bacillus strains were tested for their urease producing ability for the growth promotion by following the method of Burbank et al. [43]. The overnight grown bacterial isolate was added to Christensen’s urea broth [44] and placed in an incubate at 30 ± 1 °C in rotary shaker for 24 hours. Urease buffer was prepared by adding 1 mg EDTA, 50 mg HEPES and 20 g urea /L, cooled at 4 °C before use. Then, 2 µL of Christensen’s urea bacterial culture was added in 1800 µL urease buffer(1 litter buffer prepared by adding 1 mg EDTA, 50 mg HEPES and 20 g urea and cooled at 4 °C before use) in microtubes placed in ice both followed by the addition of sodium hypochlorite solution (17.5 g NaOH, 59.45 g Na2HPO4 and 200 mL bleach per litter solution) and 0.2 mL of phenol nitroprusside (70 g phenol and 0.34 g nitroprusside per litter).Mixture was gently shaken by inverting tubes and incubated at 30 ± 1 °C for 20 min. Ammonia standards, i.e 1, 2, 3, 4, 5, 10, 15, and 20 µg mL-1 were prepared by using ammonium chloride (NH4Cl). The urease buffer solution was also added to the standards. Urease activity in standards solutions and samples were measured at 640 nm using spectrophotometer (Model Carry 60, Agilent Technologies Santa Clara, CA). Concentration of urease in samples were calculated through comparison with standard curve.
The bacterial isolates were further tested for siderophore production in Chrome Azurol S (CAS) media [45]. For this purpose, 72.9 mg of hexadecyltrimethylammonium bromide (HDTMA) and 10 mg of FeCl3.6H2O were dissolved in 40 mL distilled water whereas 60.5 mg of CAS was dissolved in 50 mL distilled water in a separate tube. To prepare CAS media, mix both solutions and make volume 100 mL with distilled water. The overnight grown bacterial strains (0.1 mL) having 0.6 OD600 were inoculated in sterilized Kings B broth (99.9 mL) and incubate for 48 hours at 30 °C. After this, centrifuge the culture at 4500 rpm for 15 minutes, collect the supernatant. By mixing 0.5 mL supernatant with 0.5 mL CAS reagent, the color was developed, after 20 minutes of incubation optical density was measured on a spectrophotometer (Model Carry 60, Agilent Technologies Santa Clara, CA) at 630 nm. Siderophore was calculated as percent siderophore unit (psu) [46].
The bioassay developed by Ashraf et al. [47] was used to determine the synthesis of exopolysaccharide. For the EPS bioassay, bacterial culture was added to RCV broth medium, which was then incubated for 72 hours at 30 ±°C under shaking (100 rpm). The culture was centrifuged for 20 minutes at 1000 rpm to separate the supernatant in a tube. In order to form EPS crystals, the extracted supernatant was combined (1:3) with cooled acetone and allowed overnight at 4 °C. The solution was centrifuged at 4500 rpm for 15 minutes and the supernatant discarded. To remove water, the remaining EPS pellets were lyophilized and then dissolved in a 1:5 solution of phenol and sulfuric acid. To measure the absorbance at 640 nm a spectrophotometer (Model Carry-60, Agilent Technologies Santa Clara, CA) was used. By comparison with a standard curve prepared with glucose standards with concentrations of 1, 2, 3, 4, 5, 10, 15, and 20 g mL-1, the EPS concentration was determined. [48].
4.3. Determination of organic acids
Organic acids produced in bacterial culture were determined as studied by Butsat et al. [49]. The bacterial isolates (100 µL) inoculated in test tubes that contains DF minimal broth (14.9 mL) and incubate for 72 hours at 28 °C. After this, centrifuged the cultures at 10,000 rpm, mixed the supernatant with HPLC-grade methanol in 1:1 ratio. By using HPLC, the organic acids were measured along with standards (Shimadzu, Japan).
4.4. In vitro rice growth promotion characterization of catalase producing Bacillus strains
A loop full of respective bacterial strain (AN24, AN30, AN31 and AN35) was dipped in 100 mL of sterilized Luria Bertani (LB) liquid medium separately [50] and incubated at 100 rpm at 30 ± 2 °C in a shaking incubator (S19R-2, Sheldon Manufacturing, Cornelius, OR97113, USA). After 48 hours of incubation, bacterial broth culture has 1.2 OD600 was used as inoculum. The rice seed were disinfected by dipping in 0.2% solution of HgCL2 for 30 seconds followed by one minute dipping in ethanol (95%) and thoroughly rinsed for 5 times with sterilized water. Then, disinfected seeds were dipped in respective bacterial culture for 30 minutes before sowing. For co-inoculation the equal volume of both cultures was mixed, and rice seeds were dipped in that mixture. Ten inoculated rice seeds were sown in plastic jars, filled with 600 g of sterilized sandy loam soil that was characterized before filing in jars (Table 2). Five replications of each treatment were arranged in completely randomized design (CRD) and placed in a growth room which wase set as 12 hours light (1000 flux) with 35 ± 1 °C temperature and 12 hours dark with 25 ± 2 °C temperature. Humidity was also adjusted at 60-70% throughout experiment. After germination, plant population was maintained @ five plants per jar. Half strength Hoagland solution (20 mL) was used after alternate days to fulfill the water requirement. After 35 days of sowing, rice seedlings were uprooted and growth parameters (dry biomass of shoots and roots, length of shoots and roots, root volume, root surface area and root diameter) were measured. Plant samples were analyzed for root colonization and catalase activity in roots and shoots.
4.5. Postharvest soil analysis
After harvesting rice seedlings, soil samples were collected and tested for microbial biomass nitrogen and carbon as well as for catalase activity. Through the use of the serial dilution and pour plate technique, the bacterial population was identified [51].
The catalase activity in soil was determined by potassium permanganate titration [52,53]. For this purpose, 2 g freshly collected soil samples was dissolved in 40 ml of sterilized distilled cooled water followed by adding (3 %) of 1.5 mL of H2O2 (3 %) and mixed by shaking for 30 minutes on orbital shaker. Soil particles and impurities were removed by filtration of solution using sterile Whatman filter paper number 1. Then, titrated a 25 mL filtrate against a 0.1 M potassium permanganate solution until the end point was colorless. The units of catalase activity are represented by the volume of the 0.1 M potassium permanganate solution used for the titration. The breakdown of 0.1 moles of H2O2 in one minute is represented by one unit (1 U) of catalase activity.
4.6. Statistical analysis:
Five replications were maintained in all experiments for better comparison of results. A linear model completely randomized design (CRD) was used for statistical analysis by using one-way analysis of variance. Statistics 8.1 (Informer Technologies, Inc., CA 90045, USA) was used to perform multiple comparison of means at par 5% probability level. Linear regression and correlation analysis between bacterial population in soil and microbial biomass carbon and nitrogen was done by Origin 2018. Scattered diagram plotted for comparisons of organic acids released by tested strains.
5. Conclusions
Catalase-producing Bacillus strains, i.e. Bacillus megaterium AN35, Bacillus megaterium AN24, Bacillus megaterium AN31 and Bacillus aryabhattai, have several plant growth promoting characters in terms of urease, exopolysaccharide, and siderophore production. The sole, as well as co-inoculation of catalase producing Bacillus strains improved the growth of rice seedlings. The outcome showed that a consortium of Bacillus strains with catalase activity has more potential to promote antioxidant defense system in plants and promoted growth of rice seedlings. Thus, consortium of catalytic bacterial strains having plant growth promotion traits can be used as bioinoculants to improve growth and physiology of crops.
Author Contributions
Conceptualization, data collection, methodology, formal analysis, writing (original draft), S.W.Z.S., Z.I., and M.A.; project supervision, management, formal analysis, writing, S.S., Z.A.Z., and M.A.; and funding procurement, writing (review and editing), M.A., and T.H.H (Thomas H. Hilger). The published version of the manuscript has been read and approved by all authors.
Funding
The Higher Education Commission (HEC) of Pakistan provided financial support for this work under the National Research Program for Universities project number "7140/Pujnab/NRPU/R&D/HEC/2017." The German Academic Exchange Service (DAAD) supported the exchange between the scientists of the participating Pakistani and German universities within the framework of the project "Green Revolution 2.0" (Project No. 57524035).
Data Availability Statement
The raw data presented in this study are available and will be available on editor’s request.
Acknowledgments
The Soil Microbiology and Biotechnology Laboratory, Department of Soil Science, Faculty of Agriculture and Environment, The Islamia University of Bahawalpur, Pakistan, provided the facilities needed to perform this research effectively, and the authors would like to thank them for their assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Thangaraj, B.; Solomon, P.R. Immobilization of lipases–A review. Part I. Enzyme Immobilization. Chem. Bio. Eng. Reviews. 2019, 6, 157–166. [Google Scholar] [CrossRef]
- Nandi, A.; Yan, L.J.; Jana, C.K.; Das, N. Role of Catalase in Oxidative Stress- and Age- Associated Degenerative Diseases. Oxid. Med. Cell. Longev. 2019, http. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, S.; Liu, Y.; Ma, C. Redox regulated peroxisome homeostasis. Redox Biol. 2015, 4, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Simanjuntak, E.; Zulham. Superoxide dismutase (SOD) and free radical. Jurnal Keperawatan dan Fisioterapi (JKF) 2020, 2, 124–129. [Google Scholar] [CrossRef]
- Afzal, S.; Chaudhary, N.; Singh, N. K. Role of soluble sugars in metabolism and sensing under abiotic stress. Springer, Cham Plant Grow Regul. 2021, 305-334.
- Johnson, L. A.; Hug, L. A. Distribution of reactive oxygen species defense mechanisms across domain bacteria. Free Radic. Biol. Med. 2019, 140, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Stress-triggered redox signaling: What’s in pROSpect? Plant Cell Environ. 2016, 39, 951–964. [Google Scholar] [CrossRef]
- Gupta, A.; Tiwari, A.; Ghosh, P.; Arora, K.; Sharma, S. Enhanced lignin degradation of paddy straw and pine needle biomass by combinatorial approach of chemical treatment and fungal enzymes for pulp making. Bioresour. Technol. 2023, 368, 128314. [Google Scholar] [CrossRef]
- Chen, C. H. Xenobiotic Metabolic Enzymes: Bioactivation and Antioxidant Defense, Cham: Switzerland, Springer, 2020; pp. 221-234.
- Ighodaro, O. M.; Akinloye, O. A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alexandria J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Falade, A.O.; Mabinya, L.V.; Okoh, A.I.; Nwdo, U.U. Studies on peroxidase production and detection of Sporotrichum thermophile-like catalase-peroxidase gene in a Bacillus species isolated from Hogsback Forest reserve, South Africa. Heliyon 2019, 5, e03012. [Google Scholar] [CrossRef]
- Schluter, U.; Bouvier, J.W.; Guerreiro, R.; Malisic, M.; Kontny, C.; Westhoff, P.; Weber, A. P. M. Brassicaceae display diverse photorespiratory carbon recapturing mechanisms. Bio Rxiv. 2022. [CrossRef]
- Eisenhut, M.; Roell, M.S.; Weber, A. P. Mechanistic understanding of photorespiration paves the way to a new green revolution. New Phytol. 2019, 223, 1762–1769. [Google Scholar] [CrossRef] [PubMed]
- Fernie, A. R.; Bauwe, H. Wasteful, essential, evolutionary stepping stone? The multiple personalities of the photorespir-atory pathway. The Plant J. 2020, 102, 666–677. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C. P.; Filippis, D. L.; Gul, A.; Hasanuzzaman, M. M.; Ozturk, V.; Altay, M.; Lao, T. Oxidative stress and antioxidant metabolism under adverse environmental conditions: a review. Bot. Rev. 2021, 87, 421–466. [Google Scholar] [CrossRef]
- Busch, F. A. Photorespiration in the context of Rubisco biochemistry, CO2 diffusion and metabolism. Plant J. 2020, 101, 919–939. [Google Scholar] [CrossRef]
- Zhong, Q.; Hu, H.; Fan, B.; Zhu, C.; Chen, Z. Biosynthesis and roles of salicylic acid in balancing stress response and growth in plants. Inter. J. Mol. Sci. 2021, 22, 11672. [Google Scholar] [CrossRef]
- Khatun, M.R.; Mukta, R.H.; Islam, M.A.; Hud, A.N. Insight into Citric Acid-Induced Chromium Detoxification in Rice (Oryza Sativa. L). Int. J. Phytoremediation 2019, 21, 1234–1240. [Google Scholar] [CrossRef] [PubMed]
- Tahjib, U.A.M.; Zahan, M. I.; Karim, M. M.; Imran, S.; Hunter, C. T.; Islam, M. S.; Murata, Y. Citric acid-mediated abiotic stress tolerance in plants. Int. J. Mol. Sci. 2021, 22, 7235. [Google Scholar] [CrossRef]
- Choudhary, D. K; Kasotia, A; Jain, S; Vaishany, A; Kumari, S; Sharma, K.P; Varma, A. Bacterial-mediated tolerance and resistance to plants under abiotic and biotic stresses. J. Plant Grow Regul. 2016, 35, 276–300. [Google Scholar] [CrossRef]
- Hussain, S.; Khan, F.; Cao, W.; Wu, L.; Geng, M. Seed priming alters the production and detoxifcation of reactive oxygen intermediates in rice seedlings grown under sub-optimal temperature and nutrient supply. Front. Plant Sci. 2016, 7, 439 https://doi org/103389/fpls201600439. [Google Scholar] [CrossRef]
- Bibián, M. E.; Pérez-Sánchez, A.; Mejía, A.; Barrios, G. J. Penicillin and cephalosporin biosyntheses are also regulated by reactive oxygen species. Appl. Microbiol. Biotechnol. 2020, 104, 1773–1783 http://. [Google Scholar] [CrossRef]
- Taniguchi, I.; Yoshida, S.; Hiraga, K.; Miyamoto, K.; Kimura, Y.; Oda, K. Biodegradation of PET. Current status and application aspects. Acs Catal. 2019, 9, 4089–4105. [Google Scholar] [CrossRef]
- Chauhan, A.; Siani, R.; Sharma, J.C. Plant growth promoting rhizobacteria and their biological properties for soil enrichment and growth promotion. J. Plant Nutr. 2021, 45, 273–299. [Google Scholar] [CrossRef]
- Harman, G.; Khadka, R.; Doni, F.; Uphoff, N. Benefits to plant health and productivity from enhancing plant microbial symbionts. Front. Plant Sci. 2021, 11, 610065 [Google Scholar] [CrossRef]. [Google Scholar] [CrossRef]
- Babiker, B. M.; Ahmed A., E.; Ibrahim, H.M. Isolation & identification of catalase producing Bacillus spp: A comparative study. Int. J. Adv Res. 2017, 4, 1206–1211. [Google Scholar]
- Philibert, T.; Rao, Z.; Yang, T.; Zhou, J.; Huang, G.; Irene, K.; Samuel, N. Heterologous expression and characterization of a new heme-catalase in Bacillus subtilis 168. J. Ind. Microbiol. Biotechnol. 2016, 43, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Sun, B. O.; Bai, Z.; Bao, L.; Xue, L.; Zhang, S.; Wei, Y. Bacillus subtilis biofertilizer mitigating agricultural ammonia emission and shifting soil nitrogen cycling microbiomes. Environ. Int. 2020, 144, 105989. [Google Scholar] [CrossRef]
- Anbu, P.; Kang, C. H.; Shin, Y.J.; So, J.S. Formations of calcium carbonate minerals by bacteria and its multiple applications. Springer plus 2016, 5, 1–26. [Google Scholar] [CrossRef]
- Dar, A.A; Pan, B.; Qin, J.; Zhu, Q.; Lichtfouse, E.; Usman, M.; Wang, C. A review on sustainable ferrate oxidation: Reaction chemistry, mechanisms and applications to eliminate micro pollutant (pharmaceuticals) in wastewater. Environ. Pollut. 2021, 290, 117957. [Google Scholar] [CrossRef] [PubMed]
- Morcillo, R.; Manzanera, M. The Effects of Plant-Associated Bacterial Exopolysaccharides on Plant Abiotic Stress Tolerance. Metabolites 2021, 11, 337. [Google Scholar] [CrossRef]
- Naseer, I.; Ahmad, M.; Hussain, A.; Jamil, M. Potential of zinc solubilizing Bacillus strains to improve rice growth under axenic conditions. Pak. J. Agric. Sci. 2020, 57, 1057–1071. [Google Scholar]
- Mumtaz, M.Z.; Barrya, K.M.; Bakera, A.L.; Nicholsb, D.S.; Ahmad, M.; Zahir, Z.A.; Britza, M.L. Production of lactic and acetic acids by Bacillus sp. ZM20 and Bacillus cereus following exposure to zinc oxide: A possible mechanism for Zn solubilization. Rhizosphere 2019, 12, 1–10. [Google Scholar] [CrossRef]
- Timmusk, S.; Behers, L.; Muthoni, J.; Muraya, A.; Aronsson, A.C. 2017. Perspectives and challenges of microbial application for crop improvement Front. Plant. Sci. 2017, 8, 49. [Google Scholar] [PubMed]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Singh, B.K. Plamt-microbiome interactions: from community to plant health. Nat. Rev. Microbial. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Q.; Lu, X.B.; Li, Z.H.; Tian, C.Y.; Song, J. The role of root-associated microbes in growth stimulation of plants under saline conditions. Land Degrad. Dev. 2021, 32, 3471–3486. [Google Scholar] [CrossRef]
- Jian, S.; Li, J.; Chen, J.I.; Wang, G.; Mayes, M.A.; Dzantor, K.E.; Hui, D.; Luo, Y. Soil extracellular enzyme activities, soil carbon and nitrogen storage under nitrogen fertilization: A meta-analysis. Soil Biol. Biochem. 2016, 101, pp.32–43. [Google Scholar] [CrossRef]
- Hussain, A.; Zahir, Z.A.; Ditta, A.; Tahir, M.U.; Ahmad, M.; Mumtaz, M.Z.; Hayat, K.; Hussain, S. Production and implication of bio-activated organic fertilizer enriched with zinc-solubilizing bacteria to boost up maize (Zea mays L.) production and biofortification under two cropping seasons. Agronomy 2020, 10, 39 [CrossRef]. [Google Scholar] [CrossRef]
- Lukashe, N.S.; Mupambwa, H.A.; Green, E.; Mnkeni, P.N.S. Inoculation of fy ash amended vermicompost with phosphate solubilizing bacteria (Pseudomonas fuorescens) and its infuence on vermi-degradation, nutrient release and biological activity. Waste Manag. 2019, 84, 14–22 [CrossRef]. [Google Scholar] [CrossRef]
- Iqbal, Z.; Bushra, Hussain, A. ; Dar, A.; Ahmad, M.; Wang, X.; Brtnicky, M.; Mustafa, A. Combined Use of Novel Endophytic and Rhizobacterial Strains Upregulates Antioxidant Enzyme Systems and Mineral Accumulation in Wheat. Agronomy 2022, 12, 551 https:// doiorg/103390/agronomy12030551. [Google Scholar] [CrossRef]
- Beers. R. F.; Sizer, I. W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J Biol Chem. 1952, 195, 133–140. [Google Scholar] [CrossRef]
- Hildebrandt, G.; Roots, I. Reduced nicotinamide adenine dinucleotide phosphate (NADPH)-dependent formation and breakdown of hydrogen peroxide during mixed function oxidation reactions in liver microsomes. Arch. Biochem. Biophy. 1975, 171, 385–397. [Google Scholar] [CrossRef]
- Burbank, M.B.; Weaver, T. J.; Williams, B.C.; Crawford, R. L. Urease activity of ureolytic bacteria isolation from six soils in which calcite was precipitated by indigenous bacteria, Geomicrobiol. J. 2012, 29, 389–395. [Google Scholar] [CrossRef]
- Cappuccino, J.G.; Sherman, N. Microbiology. Laboratory manual (8th edition,) Pearson. ISBN. 2002; Volume 13, 978-0805325782.
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Payne, S.M. Iron acquisition in microbial pathogenesis. Trends Microbiol. 1993, 1, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Hasnain, S.; Berge, O.; Mahmood, T. Inoculating wheat seedlings with exopolysaccharide-producing bacteria restricts sodium uptake and stimulates plant growth under salt stress. Biol. Fertil. Soils 2004, 40, 157–162. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Butsat, N.; Weerapreeyakul, N.; Siriamornpun, S. Change in the phenolic acids and antioxidant activity in Thai rice husk at fie growth stages during grain development. J. Agric. Food Chem. 2009, 57, 4566–4571. [Google Scholar] [CrossRef]
- Bertani, G. Studies on Lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. Bacteriol. 1952, 62, 293–300. [Google Scholar] [CrossRef]
- Wollum II, A.G. Cultural methods for soil microorganisms. In: Methods of Soil Analysis: Chemical and Microbial Properties. A.L. Page, R.H. Miller and D.R. Keeney (Eds.), ASA and SSSA Publ. Madison, WI, USA. 1982; pp. 718-802.
- Cao, J.; Ji, D.; Wang, C. Interaction between earthworms and arbuscular mycorrhizal fungi on the degradation of oxytetracycline in soils. Soil Biol. Biochem. 2015, 90, 283–292. [Google Scholar] [CrossRef]
- Li, Q.; Liang, J.H.; He, Y.Y.; Hu, Q.J.; Yu, S. Effect of land use on soil enzyme activities at karst area in Nanchuan, Chongqing Southwest China. Plant Soil Environ. 2014, 60, 15–20. [Google Scholar] [CrossRef]
Figure 1.
Biochemical characterization of Bacillus strains for catalase, urease,siderophore and Exopolysaccharide (EPS) production. Bars having the same letter(s) did not differ from one another statistically at p≤0.05.
Figure 1.
Biochemical characterization of Bacillus strains for catalase, urease,siderophore and Exopolysaccharide (EPS) production. Bars having the same letter(s) did not differ from one another statistically at p≤0.05.
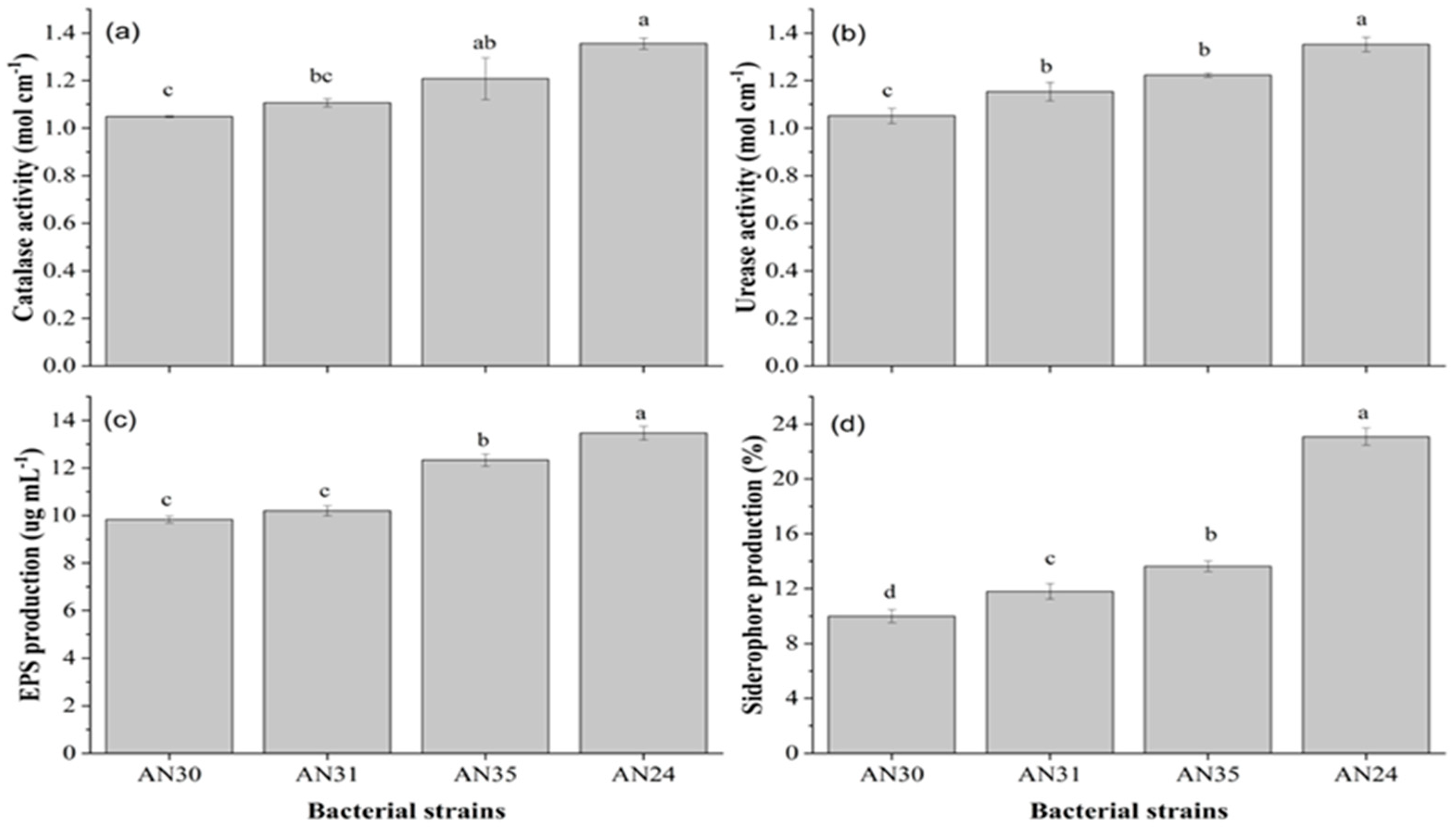
Figure 2.
Effect of sole and combined application of Bacillus strains having catalase activity to improve catalase activity in soil, shoot and root. Bars having the same letter(s) did not differ from one another statistically at p≤0.05.
Figure 2.
Effect of sole and combined application of Bacillus strains having catalase activity to improve catalase activity in soil, shoot and root. Bars having the same letter(s) did not differ from one another statistically at p≤0.05.
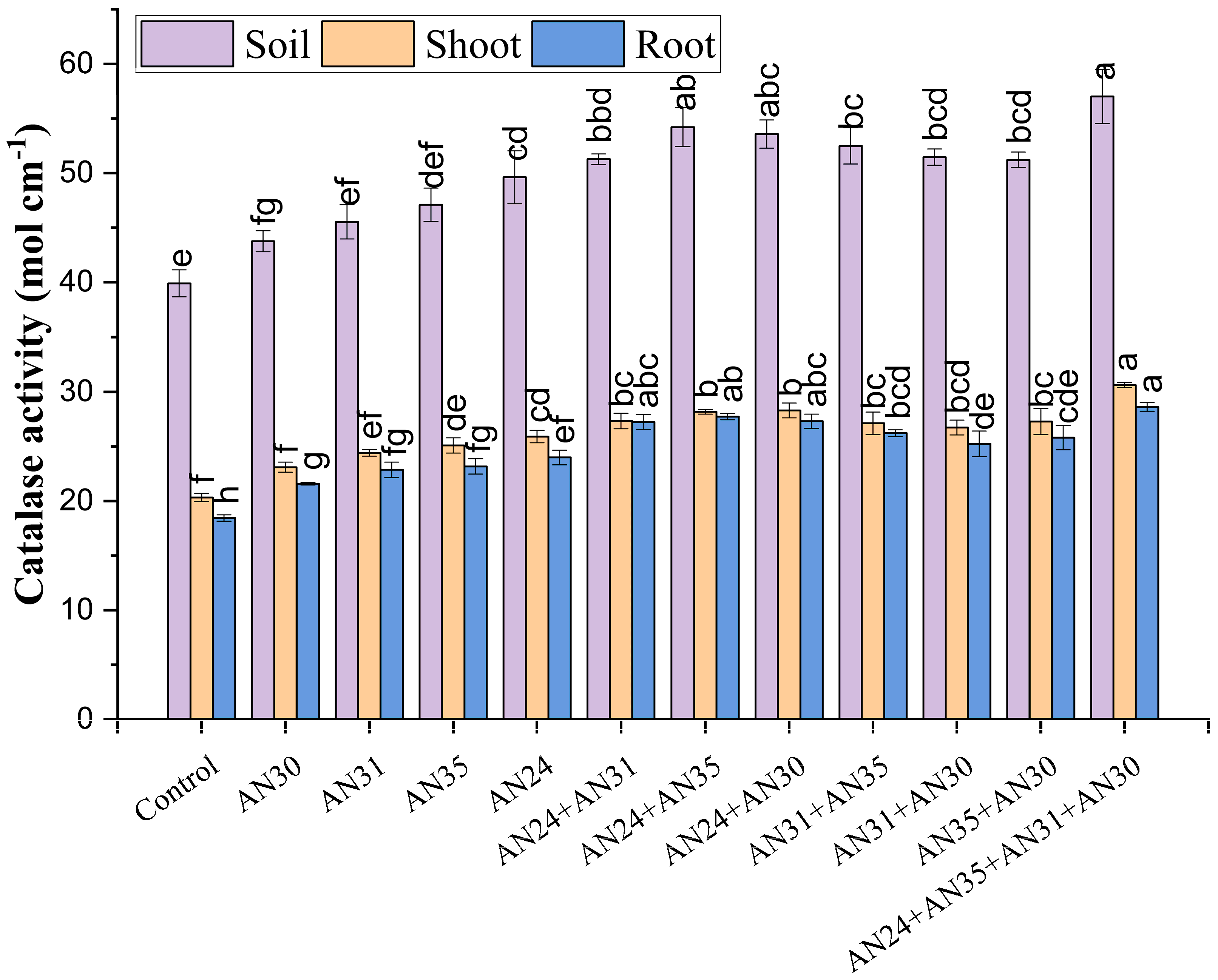
Figure 3.
Scatter diagram showing the variation in the secretion of organic acid by the tested strains in liquid culture.
Figure 3.
Scatter diagram showing the variation in the secretion of organic acid by the tested strains in liquid culture.
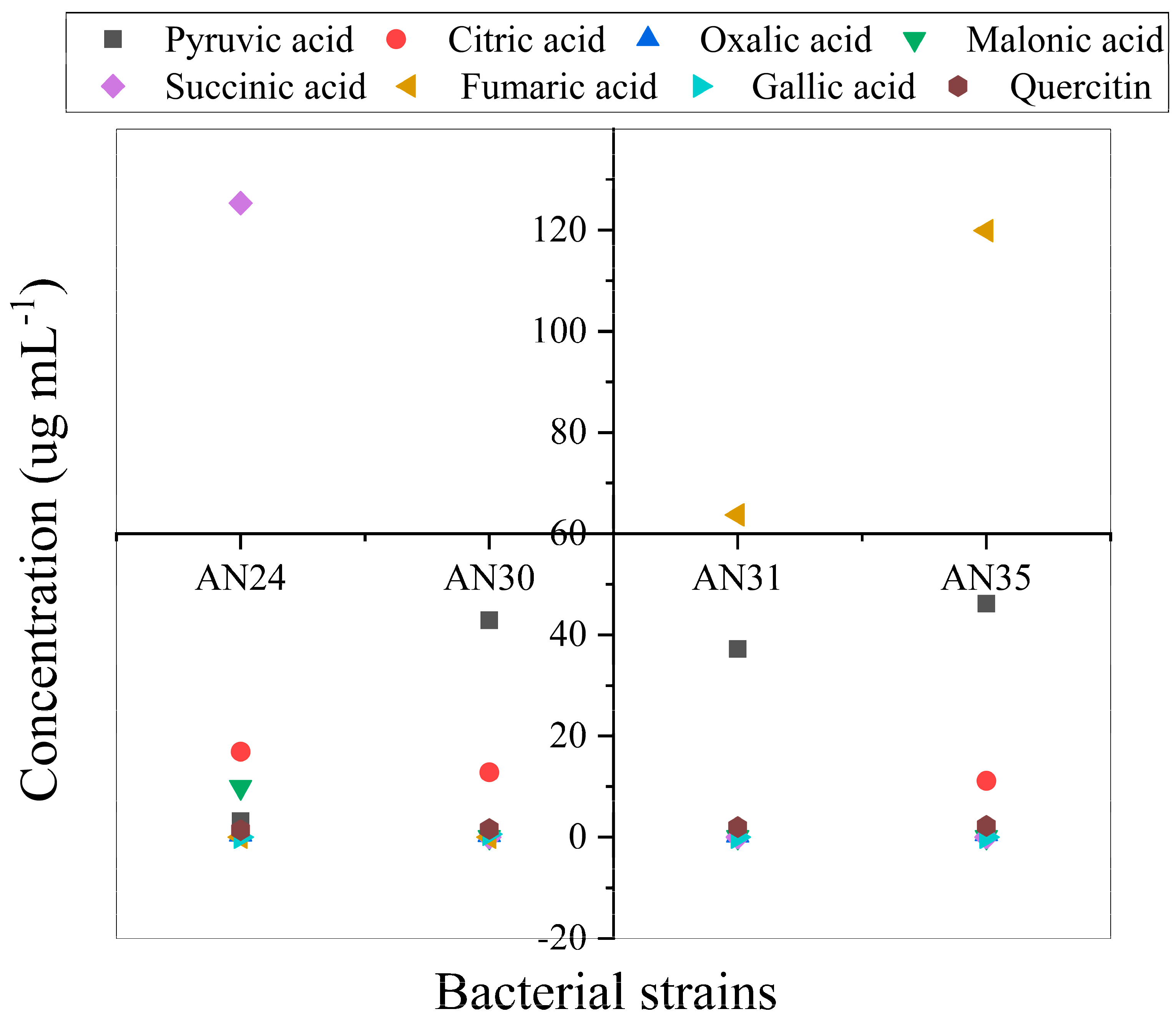

Table 1.
Effectiveness of Bacillus strains with catalase enzyme to improve growth and root morphology of rice seedling.
Table 1.
Effectiveness of Bacillus strains with catalase enzyme to improve growth and root morphology of rice seedling.
| Treatment | Shoot length (cm) | Root length (cm) | Dry weight (g seedling-1) | Root surface area (cm2) | Root diameter (mm) | Root volume (cm3) |
|---|---|---|---|---|---|---|
| Control | 6.5 ± 0.08f | 4.3 ± 0.12f | 0.51 ± 0.01f | 9.8 ± 0.06d | 0.53 ± 0.03e | 0.37 ± 0.09c |
| AN30 | 7.1 ± 0.18e | 4.9 ± 0.11e | 0.59 ± 0.02de | 10.1 ± 0.15d | 0.57 ± 0.03de | 0.39 ± 0.05ac |
| AN31 | 7.9 ± 0.16de | 5.2 ± 0.16de | 0.61 ± 0.01de | 10.9 ± 0.06cd | 0.60 ± 0.06de | 0.40 ± 0.06bc |
| AN35 | 8.0 ± 0.32de | 5.5 ± 0.10de | 0.58 ± 0.01e | 11.3 ± 0.25cd | 0.63 ± 0.03cde | 0.43 ± 0.03ac |
| AN24 | 8.3 ± 0.23cd | 5.7 ± 0.05cd | 0.64 ± 0.01cd | 11.8 ± 0.09bc | 0.63 ± 0.03cde | 0.44 ± 0.04ac |
| AN24+AN31 | 8.8 ± 0.09b | 6.1 ± 0.10b | 0.67 ± 0.03bc | 12.5 ± 0.83b | 0.77 ± 0.03abc | 0.48 ± 0.02ac |
| AN24+AN35 | 8.8 ± 0.16b | 6.1 ± 0.16b | 0.71 ± 0.01b | 12.4 ± 0.27b | 0.67 ± 0.03cde | 0.47 ± 0.03ac |
| AN24+AN30 | 8.7 ± 0.05bc | 5.8 ± 0.09bc | 0.68 ± 0.00bc | 12.3 ± 0.24b | 0.80 ± 0.06ab | 0.47 ± 0.03ac |
| AN31+AN35 | 8.5 ± 0.09bc | 5.8 ± 0.09bc | 0.68 ± 0.01bc | 12.0 ± 0.94bc | 0.63 ± 0.03de | 0.47 ± 0.03ac |
| AN31+AN30 | 8.8 ± 0.08bc | 5.9 ± 0.08bc | 0.66 ± 0.03bc | 11.9 ± 0.73bc | 0.70 ± 0.06bcd | 0.51 ± 0.01ab |
| AN35+AN30 | 8.4 ± 0.16bc | 5.8 ± 0.09bc | 0.67 ± 0.02bc | 12.0 ± 0.58bc | 0.70 ± 0.06bcd | 0.47 ± 0.03ac |
| AN24+AN35+AN31+AN30 | 9.4 ± 0.26a | 6.6 ± 0.10a | 0.80 ± 0.02a | 13.0 ± 0.25a | 0.87 ± 0.12a | 0.54 ± 0.07a |
| LSD (p≤0.05) | 0.5161 | 0.3056 | 0.0562 | 1.3950 | 0.1589 | 0.1317 |
Means having the same letter(s) do not differ from each other statistically at p≤0.05.
Table 2.
Physio-chemical characteristics of soil:.
| Analysis | Unit | Values |
|---|---|---|
| Textural class | Sandy Loam | |
| ECe | dS m-1 | 1.59 ± 0.0233 |
| pH | 7.91 ± 0.0581 | |
| Saturation percentage | % | 31.73 ± 0.2333 |
| Organic matter | % | 0.58 ± 0.0088 |
| Phosphorus | mg kg-1 | 3.02 ± 0.0722 |
| Potassium | mg kg-1 | 75.50 ± 1.0408 |
| Nitrogen | % | 0.03 ± 0.0006 |
Data is presented as means of three replicates with standard errors.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



