
Submitted:
31 August 2023
Posted:
01 September 2023
You are already at the latest version
Abstract
Creating transgenic insects is a key technology in insect genetics and molecular biology. A widely used instrument in insect transgenesis is the piggyBac transposase, resulting in essentially ran-dom genomic integrations. In contrast, site-specific recombinases allow the targeted integration of the transgene construct into a specific genomic target site. Both strategies, however, often face limitations due to low transgenesis rates. We aimed to enhance transgenesis rates by utilizing a capped mRNA as a source of transposase or recombinase instead of a helper plasmid. A system-atic comparison of transgenesis rates in Aedes mosquitoes, as models for hard to transform in-sects, showed that suppling piggyBac transposase as mRNA increased the average transfor-mation efficiency in Aedes aegypti from less than 5% with the plasmid source to about 50% with mRNA. Similar high transformation activity was observed in Ae. albopictus with pBac mRNA. No efficiency differences between plasmid and mRNA were observed in recombination experi-ments. Furthermore, a codon-optimized version of piggyBac transposase delivered as plasmid didn’t improve the transformation efficiency in Ae. aegypti or the agricultural pest D. suzukii. We believe that the use of mRNA has strong potential for enhancing pBac transformation efficiencies in other mosquitoes and important agricultural pests such as tephritids.
Keywords:
Insect transgenesis
; Aedes
; Drosophila suzukii
; tephritids
; transformation efficiency
; recombination efficiency
; piggyBac transposase
; helper plasmid
; capped mRNA
1. Introduction
Creating transgenic insects is a rapidly advancing technology with various applications in insect genetics and molecular biology. For example, insect transgenesis can address fundamental research questions like uncovering the functions of specific genes and developing genetically modified strains with unique features that can be used in agricultural pest or vector control applications. These applications have been accelerated and become more widespread due to the discovery of CRISPR genome editing and its successful application to many insect species. However, the efficiency of integrating DNA constructs via CRISPR-HDR is often low and requires a sequenced genome of the target species. Therefore, transposon-based transformation still is a standard and widely used method to create transgenic insects because it is not dependent on the availability of a high-quality genome sequence. Many different class II transposable elements (DNA transposons) have been investigated for their suitability as a tool for insect transformation since their initial use more than 40 years ago [1-5]. The piggyBac (pBac) transposase has proven to be particularly important as a universal tool for insect transgenesis. It has been applied to a wide variety of insect orders and species [6-11], including Aedes and Anopheles mosquitoes [12-17]. Class II transposable elements naturally comprise the transposase gene flanked by inverted repeats. The transposase enzyme recognizes the inverted repeats to catalyze the excision of the gene from the genome and insert the gene at a new position (a cut-and-paste mechanism). Insect transformation by transposable elements traditionally uses a bipartite system consisting of a helper plasmid encoding the transposase gene and a donor plasmid encoding the transgene construct flanked by the inverted repeat sequences of the transposable element. As a result, only the transgene construct is inserted in the genome. In contrast, the nucleic acid sequence encoding the transposase gene on the plasmid is not inserted into the genome. It gets lost after a few cell divisions in the developing embryo, ensuring the stability of the genomic integration.
Both plasmids are injected into the posterior pole of preblastoderm insect embryos to achieve transformation in the germline. Transposases commonly have very short recognition sequences (for example, TTAA for pBac transposase), which leads to essentially random integration of the encoded transgene construct into the target genome. This characteristic has proven a versatile tool to uncover gene functions due to insertional mutagenesis effects or enhancer traps [18, 19]. However, insertional mutagenesis can reduce the transformed lines’ fitness, thereby reducing their utility. In contrast, site-specific recombination (SSR) systems can avoid this problem by targeting the integration of the transgene construct into a previously characterized “landing site” or “docking site” by a mechanism called recombinase-mediated cassette exchange (RMCE). RMCE landing-sites lines are usually created by transposon-mediated random genomic integration of a landing-site construct, typically consisting of a fluorescent marker flanked by recombination sites for an appropriate recombinase enzyme. The lines are then characterized for their viability and the integration site, and the best line(s) can be used for SSR experiments. SSR systems used in insects so far include the phiC31 integrase from the phage phiC31 [20], the Cre recombinase from the Escherichia coli phage P1 [21], and the Flipase from the yeast Saccharomyces cerevisiae [22]. Similar to pBac, these recombinases can be provided as a helper plasmid into the early embryo, together with a donor plasmid encoding the gene construct of interest flanked by the recombination sequences compatible with the respective sequences at the genomic landing site.
In most cases of insect transgenic experiments, the transgenic insect is the basic product needed for the downstream experiments or a whole research project. Two measures of how efficiently a transgenic insect can be created are the transposition and recombination efficiencies. These efficiencies are ideally calculated as the number of observed independent transgenic events divided by the number of fertile G0 adults resulting from the injection. If the latter is unavailable due to experiment design, the minimal efficiency can be calculated as the number of independent transgenic events divided by the total number of G0 adults. In Drosophila melanogaster and some other species, the transposition and recombination efficiencies are usually sufficiently high that the transgenesis step does not represent a substantial barrier to progress [23-26]. However, in other insect species, transgenesis rates are often low [7]. For example, transgenesis rates mostly range between 0% and less than 10% in mosquitoes [7, 12-14, 27-31]. Low efficiencies of 1-5 % were also routinely obtained in our laboratory for pBac-mediated transposition and Cre- and phiC31-RMCE in the yellow fever mosquito, Aedes aegypti, leading to a low number of transgenic lines per injection. No line was often obtained despite hundreds or thousands of injected embryos and tens of thousands of offspring screened for the presence of the transgene construct. Thus, creating transgenic lines can become a significant bottleneck, especially in non-model organisms.
Higher transposition and recombination efficiencies in Ae. aegypti and other mosquito species would facilitate the application of transgenic work in this taxon of medically important insects. A widely used transgenesis tool in mosquitoes is the helper plasmid for pBac-mediated transformation driving transposase expression via the D. melanogaster heat shock promoter 70 (hsp70) [9]. The same promoter drives recombinase expression in Cre and phiC31 helper plasmids used in our lab [24]. One cause for the low efficiencies observed with these helper plasmids could be a low transcriptional activity of the exogenous D. melanogaster hsp70 promoter in Aedes. Additionally, a low activity of the enzyme itself could be responsible. Transcription-related effects could be solved by injection of capped mRNA instead of the helper plasmid. To test this hypothesis, we systematically compared the efficiency of hsp70 helper plasmids to that of in vitro transcribed, capped mRNA for pBac transposase, phiC31 integrase, and Cre recombinase in Ae. aegypti, and we also tested pBac and Cre mRNA efficiencies in the congeneric species Ae. albopictus. For pBac transposase, an alternative solution could also be using a hyperactive version of the pBac enzyme (IPB7) selected in a S. cerevisiae mutagenesis screen [32]. In the initial study, the hyperactive version of pBac did not improve the transposition rate in Ae. aegypti and D. melanogaster in genetic transformations [33]. However, a systematic comparison between the original pBac transposase and IPB7 under the control of the D. melanogaster hsp70 promoter achieved up to 15-fold higher transformation rates with IPB7 in Tribolium castaneum, D. melanogaster, and Ceratitis capitata ([34]; this study uses IPB7 under the name ihyPBase). We therefore decided to test this hyperactive pBac transposase again in Ae. aegypti and additionally in Drosophila suzukii (Spotted Wing Drosophila, SWD) embryos by performing comparative injections with both helper plasmids.
The data presented here confirm the results by Wright et al. [33] regarding the efficiency of the hyperactive pBac plasmid on the transposition rate in Ae. aegypti. IPB7 also doesn’t improve the transposition rates in D. suzukii. The effect of mRNA on the transformation efficiency seems to be enzyme-dependent. While using Cre or phiC31 mRNA didn’t improve the recombination efficiencies in the tested mosquito species, using pBac mRNA resulted in an average more than 30-fold increase in transposition efficiency compared to the helper plasmid in Ae. aegypti. The results in Ae. albopictus indicated that pBac mRNA activity is similarly high as in Ae. aegypti. We attribute these differences in mRNA efficiency between pBac-mediated transposition and Cre- or phiC31-mediated recombination to the completely different reaction dynamics. For pBac transposition, there are potentially millions of possible genomic integration sites. On the contrary, for RMCE, only one recombination site exists in the genome.
The substantial improvement of transposition efficiencies observed with pBac mRNA in the two Aedes species makes it a promising strategy to apply to other mosquito species for which low transformation rates with pBac plasmid were observed. Moreover, injections of mRNA as transposase source could be considered for other challenging to transform insect species, and also for other transposable elements like Minos, Hobo, or Hermes.
2. Results
2.1. ihypBase helper plasmid doesn’t increase the transposition efficiencies in Ae. aegypti and D. suzukii embryos
To test if the original hyperactive version of the pBac transposase with the insect codon usage under the D. melanogaster hsp70 promoter (ihyPBase) [34] results in higher transposition efficiencies in the yellow fever mosquito, Ae. aegypti, embryos of the Higgs White Eye (HWE) strain were injected with varying concentrations of the phsp-ihyPBase helper plasmid in combination with two different donor plasmids, including a variation of helper-donor ratios. In parallel, the same injections were performed with the standard phsp-pBac plasmid [9]. Moreover, the results were compared to the transposition efficiencies obtained with the phsp-pBac plasmid in our laboratory over the years, which had as well been performed at various helper concentrations and helper/donor ratios in the attempt to optimize phsp-pBac-mediated transformation efficiencies (Table 1, Table S1). ihyPBase helper concentrations were chosen lower than the phsp-pBac concentrations typically used in expectance of higher transformation rates as observed in T. castaneum, D. melanogaster, and C. capitata [34].
In the three injections with the ihyPBase plasmid, the larval hatch rate was about 7-8% lower than primarily observed in phsp-pBac injections (between 10-24%) (Table 1). However, this difference was insignificant (p = 0.081, single factor ANOVA). No noticeable difference could be observed for the adult emergence rate, and also the fertility of the G0 families was comparable with both helper plasmids (Table 1 and Figure S1a - d; p = 0.175, and p = 0.647, single factor ANOVA). The average transformation efficiency in our laboratory with the phsp-pBac plasmid over the years was 1.46% (13 experiments in total, including seven experiments with no transgenic event). In this set of parallel injections with the ihyPBase plasmid and phsp-pBac, no transgenic event was recovered (a total of more than 2200 injected embryos each, and 23000 screened G1 for ihyPBase, and 34000 screened G1 for phsp-pBac).
Shown are the injection data using the standard phsp-pBac piggyBac helper plasmid (pBac) collected over six years and injections performed using the phsp-HypB plasmid (HypB) containing the insect codon optimized hyperactive pBac. Injections 4, 14, and 15, as well as 5, 6, and 16 (printed in bold) were performed in parallel with the same donor plasmid and eggs from the same female cohort. “Injected embryos” represents the number of black embryos 24 h post injection; “hatch rate” = no. larvae / no. black eggs (%); “eclosion rate” = no. adults / no. larvae (%); “no. transg. events” is the number of independent transposition events observed; “transf. eff.” = the minimal transformation efficiency, calculated as: number of independent transpositions events/ total number of fertile G0 adults. The actual transformation efficiency can be higher, because in group backcrosses of G0 individuals, the number of infertile G0 isn’t known. Therefore, all G0 are included in the calculation; n.a. = not applicable; n.d. = not determined; avg = average; * this data has been published before in Häcker et al. 2017 [35].
The same ihyPBase helper plasmid was tested for transposition efficiency in the agricultural pest D. suzukii and compared to previous injections with the standard phsp-pBac helper. In D. suzukii, the standard injection mix concentration for the phsp-pBac helper over the years was predominantly 200 ng/µl helper and 500 ng/µl donor plasmid. These concentrations were also used for ihyPBase injections in two experiments. Moreover, both helper plasmids were mixed at equal concentrations in three independent injection experiments (Table 2). As in Ae. aegypti, the ihyPBase didn’t improve the transformation efficiency compared to the standard phsp-pBac plasmid. We also did not observe differences in development of injected embryos between phsp-pBac and ihyPBase injections (Table 2, Figure S1e - g).
Shown are the injection data using the standard phsp-pBac helper plasmid (pBac) (exp. Ds 1-18), and injections performed using phsp-HypB plasmid (hyPB) with insect codon optimized hyperactive pBac helper (Ds 22, 23) or a combination of both (Ds 19-21). The injections for AH443 [24], V220-223 [36], V146, V183-V185, V188, V213, V215, V226-229, V250-251 [37], V209 [38], and V265 [39], were performed across more than 10 years. All injected eggs were counted. “hatch rate” = no. larvae / no. injected eggs (%); “fertile eclosion rate” = no. fertile G0 adults/ no. hatched larvae (%); “no. transg. lines” is the number of independent transgenic lines obtained (all G0 were backcrossed individually, except injection Ds 18; here independent lines were identified from each family and confirmed by segregation analysis); “transf. eff.” = the transformation efficiency, calculated as: number of independent transgenic events / total number of fertile G0 adults; n.d. = not determined; avg = average.
2.2. pBac mRNA boosts the transposition rate in Ae. aegypti
To assess the efficiency of in vitro transcribed, capped mRNA as a pBac helper source, pBac mRNA injections were performed with six different donor plasmids (V19, V96, V97, V368, V369, V370) ranging from 7 to almost 12 kb in plasmid size and 3.6 to 6.5 kb in insert size. The minimal transformation efficiencies of these injections were compared to the phsp-pBac helper plasmid injections performed over the years in our laboratory with various donor plasmids (Table 3 and Table S2). Of the six donor plasmids used in pBac mRNA injections, V19 had previously been injected with the pBac helper plasmid. Moreover, the V370 donor plasmid used in mRNA injections is identical to the V258 donor injected with pBac helper plasmid except for an additional attB recombination site in V370. As all the plasmid helper and mRNA helper experiments were conducted at different time points, with different HWE female cohorts, by different experimenters, and some in a different laboratory (V369, V370), we also performed side-by-side injections with the phsp-pBac plasmid or pBac mRNA together with the V368 donor plasmid into embryos collected from the same batch of HWE females.
Injection numbers across all experiments ranged from 150 to more than 1000 injected embryos per experiment for the helper plasmid, and 110 to less than 600 embryos for the helper mRNA (Table 3 and Table S2). In helper plasmid injections, pBac and donor concentrations varied between 160 and 400 ng/µl, and 150 and 600 ng/µl, respectively. In mRNA injections, the donor concentration was kept constant at 300 ng/µl, and the mRNA concentration varied between 100 and 300 ng/µl. G0 injection survivors were backcrossed individually or in groups of up to 15 individuals, depending on the number of G0 survivors and the current insectary capacity, and the offspring (G1) screened for fluorescent marker expression. For all mRNA injections and three of the pBac plasmid injections (exp. 2, 3, 9), a subset of positive G1 of each G0 family was analyzed for the transgene copy number in the genome via droplet digital PCR (ddPCR). In case of single integration events identified by ddPCR, inverse PCR was performed to determine and distinguish between genomic integration sites. Some individuals were sacrificed for the transformation event analysis only after individual backcrossing and successful mating. The minimum number of independent genomic integration events per G0 family was determined by summing up the number of G1 with a different copy number or integration sites. A copy number greater than one was counted as one independent event in this analysis. The complete set of data for the integration event analysis is summarized in Table S3.
Data from 13 injection experiments using the phsp-pBac helper plasmid over six years and nine injection experiments using pBac capped mRNA over five years are displayed. Exp. 1-13 are identical to the ones shown in Table 1. Exp. 12, 13, 21, and 22 (printed in bold) were performed in parallel with eggs from the same female cohort; Injections 14 and 15 were performed in the WT Orlando laboratory strain, and all other injections in the Higgs White Eye strain; “Injected embryos” represents the number of black embryos 24 h post injection; “hatch rate” = no. larvae / no. black eggs (%); “eclosion rate” = no. adults / no. larvae (%); “no. transg. events” is the number of independent transposition events observed (multiple genomic integrations in one individual were counted as one event); “no. of transg. events / G0” = the maximum number of independent transposition events detected in a G0 founder individual as determined by ddPCR and iPCR analysis of positive G1. If the number is given as ‘≥ number’, then only a subset of positive G1 was molecularly analyzed, and additional independent events might not have been detected; “transf. eff.” = the minimal transformation efficiency, calculated as: number of independent transgenic events/ total number of G0 adults. The transformation efficiency in several families is assumed to be higher (see text); n.a. = not applicable; n.d. = not determined; avg = average; * detailed numbers of all transgenic events identified in each G0 family and calculations see Tables S2 and S3; ** transformation efficiency of > 100% results from single G0 founder individuals producing more than one independent transposition event (as determined by ddPCR and iPCR); *** this data has been published before in Häcker et al. 2017 [35].
Across all injections, we observed a lower but not significantly different larval hatch rate in the mRNA helper injections (Figure 1a; p = 0.06619, one-factor ANOVA), which seemed independent of the size of the injected donor construct. There was no difference in the G0 adult eclosion rate and in the fertility of the G0 families between plasmid and mRNA helper (Figure 1b, c; p = 0.2340 and p = 0.6204, respectively, one-factor ANOVA). However, using the mRNA helper, the transposition efficiency increased from an average of 1,49% with the helper plasmid to more than 50% on average (Figure 1g, p =1.646E-06). These numbers refer to the calculated minimal transformation efficiency, i.e., the number of obtained independent transgenic events divided by the total number of adult G0 survivors of each injection. Different independent events within one G0 family as determined by digital PCR and inverse PCR were counted separately. The actual transformation efficiency, however, in several cases might have been even higher than the numbers reported here for the following reasons: i) G0 injection survivors were not always backcrossed individually (Table S2). Thus, in some cases, more than one positive G0 could have been in a positive pool, resulting in more than one independent transgenic event, which ddPCR and inverse PCR wouldn’t have necessarily identified as only small numbers of G1 individuals per family were assessed. This applies especially to the V369 and V370 injections (exp. 16, 17), the only mRNA injection experiments where the G0 were exclusively backcrossed in large groups (12 individuals per cage, see Table S3). Moreover, of several G0 families, the offspring was not molecularly characterized for copy number and integration sites. ii) All emerged G0 adults were counted for the calculation, including sterile individuals, which on average, were about 40% (Table S4).
A frequent observation with pBac mRNA injections were multiple transgene construct integrations in one individual, ranging from two to 18 integrations at the higher mRNA concentration, and two to 14 integrations at the lower concentration in the G1 animals analyzed by ddPCR. Different integration frequencies were observed between the offspring of different G0 founders and within the offspring of single-founder G0 families, including individuals with single integrations at different genomic locations as determined by inverse PCR. On the contrary, from all phsp-pBac plasmid injections, only one individual with two integrations of a transgene construct was obtained. For the detailed results, see Tables S2 and S3.
2.3. Preliminary data indicate high transposition rates with pBac mRNA in Ae. albopictus
The efficiency of capped pBac mRNA was also assessed for Ae. albopictus. In this experiment, no control injections with pBac helper plasmid were performed, as the primary purpose was to create landing site lines for RMCE. G0 survivors were exclusively backcrossed in groups of 10-15 individuals, and the positive G1 offspring was not further analyzed molecularly to identify different transposition events within a G0 family. Nevertheless, this experiment indicated a similarly high transposition rate in Ae. albopictus as in Ae. aegypti, as 15 of the 19 G0 families produced positive G1 (Table 4). This percentage of positive families is comparable to the Ae. aegypti mRNA injection experiments #16 and 17, where G0 were backcrossed in a similar scheme (mostly groups of 12). These group backcrosses produced a similar rate of positive families, i.e., 11 of 12 (exp. 16) and five of six families (exp. 17) (Table 3, Table S2). If only the number of positive families is counted as independent transposition events in these experiments, this results in a transformation efficiency of 7%, i.e., in the same range as observed for the Ae. albopictus mRNA injection. We, therefore, assume that the actual transformation efficiency in Ae. albopictus was much higher.
Displayed are data from one injection experiment using pBac capped mRNA in Ae. albopictus. “no. injected embryos” total number of injected embryos; “hatch rate” = no. larvae / total no of injected eggs (%); “eclosion rate” = no. adults / no. larvae (%); “no. transg. events” is the number of independent transposition events observed; “transf. eff.” = the minimal transformation efficiency, calculated as: number of independent transgenic events/ total number of G0 adults. The actual transformation efficiency might be higher (see text). For more details in Ae. albopictus transformation data, please see Table S5. * In this plasmid, the D. melanogaster PUbCFP cassette was replaced with an Ae. aegypti PUbeCFP cassette from pSL1180-HR-PUbeCFP (Addgene plasmid 47917).
2.2. Capped mRNA doesn’t improve Cre- or phiC31-RMCE efficiencies in Aedes.
Based on the strong positive effect of capped mRNA on pBac transformation efficiencies, we subsequently tested the efficiency of capped mRNA in phiC31- and Cre-RMCE experiments in direct comparison to the respective helper plasmid.
For phiC31-RMCE, comparative injections were performed at two different helper/donor concentrations, 150/300 ng/µl and 300/500 ng/µl in Ae. aegypti. Concentrations were chosen to be in the same range as concentrations used in phiC31-RMCE experiments in two different landing site lines previously performed in our laboratory (Table S6). They were also comparable to the pBac mRNA concentrations. Five independent injections were performed for the lower concentration, two with the phiC31 helper plasmid and three with the phiC31 mRNA. For the higher concentration, two replicates each were performed (Table 5). No significant differences in the larval hatch rate, adult eclosion rate, or G0 fertility were observed between helper plasmid and mRNA injections (p = 0.109, p = 0.181, p = 0.456, respectively, statistics done across all concentrations; see also Fig. S2;). Based on the numbers obtained with pBac mRNA, one transgenic event would have been expected for about every 50th injected embryo, assuming a similar efficiency. However, the overall recombination efficiency was very low. Only in one of the three replicates with mRNA at the lower helper/donor concentration, we observed one RMCE event. In comparison, we identified one donor plasmid integration event in one of the two replicates with helper plasmid at the higher helper/donor concentration (Table 5). The corresponding recombination efficiencies within the replicates were 10 and 6.45%, respectively, and 3.34 and 3.23% across the replicates.
The efficiencies obtained with the helper plasmid were in the same range as those observed in the previous experiments, performed with the same or different donor constructs into different landing site lines (Table S6, exp. I - V).
Data from comparative injection experiments are displayed at two different helper/donor concentrations into the same landing site line; “no. injected embryos” represents the number of black embryos 24 h post injection; “hatch rate” = no. larvae / no. black eggs (%); “eclosion rate” = no. adults / no. larvae (%), “no. transg. events” is the number of independent recombination events observed; “recomb. eff.” is the minimal recombination efficiency, calculated as: number of transgenic events/ total number of G0 adults. The recombination efficiency can be higher, as in group backcrosses of G0, the number of infertile G0 is unknown. All positive individuals obtained showed the RMCE phenotype. * one additional family with transient donor phenotype; parental phenotype in next generation; ** one family with transient donor phenotype; parental phenotype in next generation
In phiC31-RMCE injections, the positive effect of the mRNA could not be observed when mRNA concentrations similar to the pBac transformations were used. Thus, concentrations were increased for Cre-RMCE injections to 450 ng/µl helper and 350 ng/µl donor. Helper plasmid and mRNA injections were performed in three independent replicates each. Similar to the phiC31 experiments, only one plasmid or mRNA replicates produced recombination events. In one of the plasmid helper injections, one family produced an RMCE phenotype, and another one offspring with a donor plasmid integration phenotype, corresponding to an overall minimum recombination efficiency of 2.44% (0.8% across all replicates, Table 6). In the mRNA injections, a single family produced offspring with an integration phenotype, corresponding to a minimum recombination efficiency of 3% (1% across all replicates). For complete injection, crossing, and screening data, see Table S7.
Data from comparative injection experiments are displayed at two different helper/donor concentrations into the same landing site line; “no. injected embryos” represents the number of black embryos 24 h post injection; “hatch rate” = no. larvae / no. black eggs (%); “eclosion rate” = no. adults / no. larvae (%); “no. transg. events” is the number of independent recombination events observed; “recomb. eff.” is the minimal recombination efficiency, calculated as: number of transgenic events/ total number of G0 adults. The recombination efficiency can be higher, as in group backcrosses of G0, the number of infertile G0 is unknown. * 1 integration, one RMCE phenotype; ** integration phenotype.
We also performed Cre-RMCE injections in Ae. albopictus into two different landing site lines. In one line (17A1), we tested injecting with either 150 ng/µl phsp CRE or 190 ng/µl helper mRNA. From 34 fertile families of G0 founders injected with helper plasmid, one showed expression of the donor transgene in addition to the original landing site marker, indicating an integration event. This translates into a recombination efficiency with Cre helper plasmid of 2.27 % (Table S8). None of the 46 fertile families made of G0 founders injected with helper mRNA showed expression of the transgene.
In a second line (1A3), the landing site integrated 1.3 Mb upstream of the Nix locus as determined via inverse PCR, and marker fluorescence was only observed in males. Thus, all injected embryos were heterozygous for the landing site. Notably, in this line, the Ae. aegypti PUbeCFP expression was not visible, but PCR could confirm the presence of the sequence. This line was only injected with 150 ng/µl phsp-CRE, and one showed expression of the donor plasmid fluorescence marker from a total of six fertile families. PCR of the animals that showed transgene expression suggested both excision of the original PUbeCFP sequence and multiple, potentially tandem, integrations of the plasmid, i.e., non-canonical recombination events.
3. Discussion
Producing transgenic insects can be time-consuming because transgenesis efficiency can be very low in many insect orders and species. This is particularly true for Aedes mosquitoes using piggyBac transformations and recombination in cassette exchange experiments (RMCE). In many experiments published to date, the phsp-pBac helper plasmid [9] was used as a source of pBac transposase, with the D. melanogaster hsp70 promoter to drive transposase expression. Minimum transposition rates in these publications are reported to be between 0 and 4 % [12, 30, 31, 35]. This transposition rate doesn’t improve when the embryos are heat-shocked shortly after injection [30]. A large meta-analysis of pBac transformation efficiencies reports an average of 7% for Ae. aegypti (46 experiments, 3314 total G0 adults, 239 independent transgenic lines) and 1.6% for Ae. albopictus (10 experiments, 5339 G0 adults, 89 independent lines).
The hyperactive version (ihyPBase) of pBac transposase helper plasmid didn’t yield transgenic lines in Ae. aegypti in three independent experiments with two different constructs and more than 170 G0 adults. These injections used similar injection numbers and a similar size of the transgene constructs as in Eckermann et al., but at lower donor and helper plasmid concentrations than the ones used by these authors (300 ng/µl and 500 ng/µl, respectively) [34]. Moreover, Eckermann et al. performed individual backcrosses of all G0, while we did group backcrosses. This might have masked the presence of transgenic G0 in case those were less competitive than the non-transgenic G0. On the other hand, at similar efficiencies as the ones observed by Eckermann et al. a few transgenic events should still have occurred. Instead, our results confirm the experiments by Wright et al., who also didn’t obtain transposition events with ihyPBase in Ae. aegypti. However, we did not observe the G0 sterility reported by Wright et al. for both, Ae. aegypti and D. melanogaster [33]. This might be attributed to the approximately two-fold lower ihyPBase helper concentrations or the absence of a heat shock treatment of the injected embryos [33] in our study. ihyPBase seemed to result in lower larval hatch rates than the standard phsp-pBac plasmid in our injections. However, the three injections performed with ihyPBase are not enough to assess if there is an actual effect. In D. suzukii, the ihyPBase helper plasmid yielded similar transposition efficiencies as the standard pBac helper plasmid. There were also no differences in the other life parameters between the ihyPBase and standard pBac helper plasmid injections in D. suzukii.
We hypothesized that the D. melanogaster hsp70 promoter does not function well in Aedes mosquitoes, causing low transposase levels. Therefore, we performed pBac-mediated transformations using in vitro transcribed, capped mRNA. Comparing the results of 13 injections with the pBac helper plasmid and nine injections with capped mRNA in Ae. aegypti, the transposition efficiencies using capped mRNA on average went up more than 30-fold, from 0-5% to around 50%, some even higher. Thus, while hundreds of embryos had to be injected to obtain a transgenic line using the pBac helper plasmid, with the mRNA on average, a few dozen embryo injections were sufficient. These data are based on the results obtained with two different Ae. aegypti strains, Higgs White Eye, and Orlando, six different donor plasmids, three different mRNA concentrations, and different injection personnel. It is important to note that the transposition efficiencies reported in this work are minimal efficiencies, as the original purpose of the experiments was not to determine transposition efficiencies. Therefore, G0 adults were backcrossed in groups to a certain extent in each injection. The actual efficiencies will consequently be higher, especially in mRNA injections # 16 and 17 (V370 and V369 donor, respectively), where all G0 individuals were backcrossed in large groups, and only a fraction of the positive offspring was analyzed molecularly. All other mRNA injections provide a more accurate transposition efficiency estimate because most G0 were backcrossed individually or in small groups. However, for all efficiency calculations, the total number of G0 was considered, not the number of fertile G0. Taking all individually backcrossed G0 from all pBac injections and assessing their fertility, it can be estimated that the average G0 fertility was about 57% for both mRNA helper and plasmid helper injections. The observed G0 infertility rate of mostly 35-45% matches well with published numbers for Ae. aegypti [30,31].
The results of the capped pBac mRNA injections in Ae. albopictus strongly suggest that capped mRNA has similarly high activity in Ae. albopictus as in Ae. aegypti. Even though the experiment didn’t include individual G0 backcrosses and the positive G1 were not further assessed for integration copy number and integration sites, the high percentage of positive families is similar to that obtained in the Ae. aegypti injections #16 and 17, where we used a very similar G0 group backcrossing scheme (mostly 12 G0 per cage). Such a high percentage of positive families was not observed in any pBac helper plasmid injection in Ae. aegypti. If only the number of positive families is considered as the number of independent events for the transformation efficiency calculation of the Ae. aegypti injections #16 and 17, the transformation efficiency would be between 6 and 7%, i.e., identical to the one obtained for Ae. albopictus using this calculation.
One important effect of the mRNA-mediated transformation was that many G1 individuals contained more than one genomic copy of the transgene construct, as determined by ddPCR. Therefore, with pBac mRNA injections, it is recommended to do single backcrosses of at least some of the positive G1. Analyzing the copy number after line establishment is necessary to identify lines with a single integration event. One approach to minimize the frequency of multiple genomic insertions would be to titrate the mRNA concentration used in injections to optimize the fraction of G1 carrying only one genomic integration. We did not observe an apparent correlation between the mRNA concentration and the max. number of integrations per individual or the relative frequency of G1 with more than one integration. However, a precise analysis of a possible correlation was not possible, as in mRNA injections #16 and 17, the positive individuals were group backcrossed until G2. Molecular analysis of integration events was only performed with generation G3. The two generations of backcrossing integrations might have segregated, consistent with the overall low number of integration events per individual observed in these two injections compared to the other experiments.
Transformation efficiencies with mRNA observed in our hands were substantially higher than the numbers reported for Aedes mosquitoes in the literature [7, 14, 27]. Specifically, in one large-scale experiment (6000 embryos), Labbe et al. injected a pBac helper plasmid and helper mRNA mix (200 ng/µl and 300 ng/µl, respectively) into Ae. albopictus, resulting in a transformation efficiency of 2.2-3.6% [14]. This estimate of transformation efficiency incorporates a correction for an assumed 30-50% G0 infertility. Haghighat-Khah et al. injected 700 ng/µl helper mRNA in Ae. aegypti embryos, resulting in only 1% transposition efficiency (two experiments totaling 3000 embryos) [27]. Only one in nine independent lines resulting from their injections showed more than one transgene integration. We can only speculate about the reason for the differences in transposition efficiencies between published work and our study. Based on our observations with 100 and 300 ng/µl of pBac mRNA in the injection mix, it is possible that at 700 ng/µl, barely any germline cells with only one or very few integrations arose. Instead, most transformed germ cells had so many integrations that the resulting zygotes were not viable. Additionally, the strain’s genetic background and the number of generations the strain had been cultivated in the lab could influence the transformation efficiency. While we injected into HWE and Orlando laboratory strains, the experiments by Labbe et al. and Haghighat-Khah et al. were performed in strains of Malaysian origin [14, 27].
Injections using phiC31 capped mRNA for RMCE in Ae. aegypti have been published before. In three different landing site line injections, using 730 ng/µl mRNA helper in the injection mix, the authors obtained no recombination event, once obtained only a donor plasmid integration event, and once obtained an RMCE event. Both of the latter events occurred at an efficiency between 4 and 5 % [27]. Our study included parallel helper plasmid injections into embryos from the same female cohorts to directly compare the recombination efficiencies with plasmid helper and mRNA helper. Based on the high transposition efficiencies observed with pBac mRNA, it was unexpected that efficiencies for RMCE experiments were not increased using mRNA as a recombinase source. This result occurred for phiC31 and Cre recombinase, where even higher mRNA concentrations were used (Table 5, 6). The phiC31-RMCE efficiency of 10% achieved in one of the injections is in the same range as the published efficiencies [27], although at about 5-fold lower mRNA concentration.
The reasons for the lack of an mRNA effect in RMCE experiments in Ae. albopictus and Ae. aegypti are currently not known. We can exclude the inactivity of the injected mRNA due to degradation, because for every injected slide of embryos, the leftover injection mix from the needle was recovered and run on a gel to confirm mRNA integrity. One big difference between pBac-mediated transposition and Cre- or phiC31-mediated recombination is the lower frequency of sites in the genome at which recombination events can occur. pBac transposase uses TTAA sequences in the genome for insertion, which are predicted to occur every 256 base-pairs. In contrast, the recombination sites for Cre and phiC31 don’t occur naturally in Aedes genomes. Thus, the genomes of our landing site lines contain exactly one position where recombination can take place, and the rate-limiting step might be the likelihood of the enzyme and donor plasmid being present at this position simultaneously. This likelihood can be increased by increasing the amount of injected mRNA and donor plasmid. However, this would be in the range of a maximum two- to three-fold increase as the viscosity of the nucleic acid solution limits the injectability.
Moreover, experiments published previously by us and others show that in case of Cre recombinase, the amount of Cre enzyme in the commonly used injection concentrations is not limiting. In genomic excision experiments, excision rates between 25% and 100% were obtained when both recombination sites were close [27, 35, 40]. Furthermore, the high efficiency of the excisions also excludes the possibility that heterochromatic factors silence the landing site constructs. A reason for the missing effect of the mRNA in RMCE injections might be the limited likelihood of co-localizing all three components for RMCE (landing site, enzyme, donor plasmid) at the same time at a single site in the genome. On the other hand, the same situation applies to other insect species, such as D. melanogaster, D. suzukii, and Anastrepha suspensa, in which RMCE efficiencies between 10 and 20% were achieved (single experiments, [24, 26, 41, 42]).
Nevertheless, for Cre-RMCE, we obtained the first one-step RMCE published so far in Aedes aegypti. In previous experiments, we only achieved two-step Cre-RMCE, obtaining an integration line first, from which, by injection of only Cre helper plasmid, the complete RMCE event was obtained [35].
4. Materials and Methods
4.1. Insect rearing
Aedes aegypti wild-type strains and transgenic lines were reared in an insectary at constant conditions of 27°C, 70% RH, and a 12:12 hrs light:dark cycle. Larvae were fed on Tetra TabiMin fish food pellets. The adult mosquito diet was sterile-filtered 10% (w/v) sucrose solution. Moreover, adult females were fed once per week with pig blood purchased from a butcher shop. Aedes aegypti laboratory strains used in the experiments were the Orlando wild type strain and the Higgs White Eye (HWE) strain (a spontaneous white eye mutant strain of the Rexville D strain from Puerto Rico [43]).
A lab colony of Aedes albopictus was established with pupae and larvae collected from an auto-salvage yard in Manassas, Virginia, in 2018. Animals were reared under standard laboratory conditions at 21-26°C, 80% RH, 16 h light:8 h dark for three generations prior to injections for pBac-mediated transformation [44]. Larvae were fed on a MWF schedule with 1 ml of a larval food slurry consisting of 1 liter DI water, 120 g dog food (Nutro Ultra Small Breed Puppy, Nutro Products Inc., Franklin, TN, USA), and 40 g frozen brine shrimp (Sally’s Frozen Brine Shrimp, San Francisco Bay Brand, Newark, CA, USA) [45]. Adult females were provisioned with organic raisins (Newman’s Own, Westport, CT, USA) to allow ad libitum sugar feeding. They were allowed to blood feed on a human host for egg production. The Georgetown University IRB has determined that mosquito blood feeding is not human research and does not require IRB approval; however, the blood feeding protocol has been approved by the Georgetown University OHS office.
The wild-type D. suzukii USA strain and transgenic lines were maintained at 25°C and 55–60% humidity with a 12 h photoperiod. Flies were briefly anesthetized with CO2 for screening and to set up crosses.
4.2. In vitro transcription (IVT) of pBac, phiC31, and Cre mRNA for injections
4.2.1. Production of the IVT templates
pBac IVT template was obtained by PCR on the phsp-pBac plasmid [9], using the forward primer P1269 (5’GAAACTAATACGACTCACTATAGGGAGAGCCGCCACatgggtagttctttagacgatg; upper case letters represent T7 initiation sequence and linker) and the reverse primer P1270 (5’cttattagtcagtcagaaacaac). The PCR reaction contained 2 ng plasmid DNA, 500 nM of each primer, 200 µM of each dNTP, 1x Q5 reaction buffer, and 1 µl Q5 Polymerase (NEB) in a final volume of 100 µl. The reaction was run in a Biorad C1000 Touch Thermal Cycler (initial denaturation at 98°C for 30 sec, 30 x [98°C for 10 sec; 51°C for 20 sec; 72°C for 1 min] followed by the final elongation at 72°C for 2 min).
Cre and phiC31 IVT templates were obtained by PCR on plasmids AH445 (phsp-Cre) and AH444 (phsp-phiC31), respectively. Primers were P2203 (5’GAAACTAATACGACTCACTATAGGGAGAGCCGCCACatgtccaatttactgaccgtacacc) and P2204 (5’gctaatcgccatcttccagcag) for Cre, and P1630 (5’GAAACTAATACGACTCACTATAGGGAGAGCCGCCACatggacacgtatgccggtgcttac) and P1631 (5’ctaggccgctacgtcttcggtgc) for phiC31. The PCR reaction contained 10 ng plasmid DNA, 500 nM of each primer, 100 µM of each dNTP, 1x Platinum Taq reaction buffer, 1.25 mM MgCl2, and 1 µl Platinum Taq DNA Polymerase (Life Technologies) in a final volume of 50 µl. The reaction was run in a Biorad C1000 Touch Thermal Cycler (initial denaturation at 95°C for 2 min, 35 x [94°C for 30 sec; 59°C for 30 sec; 72°C for 2 min] followed by the final elongation at 72°C for 10 min).
The PCR products were analyzed and purified by 1% agarose gel electrophoresis and gel extracted with the ZymoClean Gel DNA Recovery kit according to the manufacturer’s instructions.
4.2.2. In vitro transcription reaction and mRNA purification
IVT was performed using the NEB HiScribe T7 Arca mRNA kit (#2060S) according to the manufacturer’s instructions, using 800 - 1000 ng IVT template. mRNA was purified using the MegaClear Transcription clean-up kit (AM1908) according to the manufacturer’s instructions choosing elution option 1 (50 µl elution solution on the column, incubate at 65°C for 5 min) performed twice, and including the optional Ammonium Acetate precipitation. mRNA quality was analyzed by agarose gel electrophoresis, and mRNA was stored in 5 µl aliquots at -80°C until use.
4.3. Preparation of injection mixes
Transposase, recombinase-encoding helper plasmids, or in vitro transcribed, capped mRNA were mixed with the corresponding donor plasmids at the final concentrations specified in Table 1, Table 2, Table 3, Table 4 and Table 5 in RNAse-free 1x embryonic injection buffer (5 mM KCl, 0.1 mM NaPO4, pH 6.8). To remove particles and dust that could clog the injection needles, the injection mixes were centrifuged at 13000 rpm for 30 min at 4°C. The supernatant was taken carefully without disturbing a possible pellet and stored in 5 µl aliquots at -80°C until further use.
4.4. Embryonic microinjections of Ae. aegypti
Injections for pBac-mediated transformation: Ae. aegypti transgenic lines were created by injecting preblastodermal embryos of the HWE strain with the phsp-pBac helper plasmid [9] or pBac in vitro transcribed, capped mRNA and a donor plasmid at varying concentrations (see Table 1 and 3), in 1x embryonic injection buffer (EIB; 5 mM KCl, 0.1 mM NaPO4, pH 6.8). Injected embryos were kept moist for two days to allow completion of embryonic development before transferring to water with some TabiMin fish food for hatching. Survivors were sexed in the pupal stage and backcrossed to HWE in small groups or individually. G1 offspring was collected for 3-6 gonotrophic cycles and screened for the presence of the transgenic marker (DsRed or eGFP fluorescent protein) at the larval or pupal stage. Positive G1 were again backcrossed individually or in groups to establish transgenic lines.
Injections for recombinase-mediated cassette exchange: preblastoderm embryos of the Ae. aegypti landing site lines for phiC31-RMCE (lines V19-M2M1, V19-M26M3m2; V19 landing site construct = attP_3xP3-eGFP_attPrev) or Cre-RMCE (line V3-M30M1; V3 landing site construct = FRT_3xP3-DsRed_FRT3_loxN_3xP3-FRT5_AmCyan-lox2272-loxP_attP220rev) were injected with the recombinase helper plasmid or in vitro transcribed, capped recombinase mRNA and a donor plasmid at varying concentrations (see Table 4 and Table 5), in 1x EIB. Further rearing and crossing were identical to pBac injections described above.
4.5. Embryonic microinjections of Ae. albopictus
4.5.1. Preparation of donor plasmid
To generate the modified AH452 construct (pXL-BACII_FRT_3xP3DsRed_FRT3_loxN-PUbeCFP-lox2272), the D. melanogaster PUbCFP cassette from AH452 [27, 35, 40] was replaced with an Ae. aegypti PUb-eCFP cassette from pSL1180-HR-PUbeCFP (Addgene plasmid 47917). One fragment from the AH452 plasmid was amplified using primers Vector_F (5’ CTAAATTGTAAGCGTTAATATTTTG 3‘) and Vector_R (5’ATAACTTCGTATAAGGTATACT 3 ‘), and a second fragment was amplified using primers SG_frag_2_Forward (5’ CTGGATCATAACTTCGTATAGGATACTTTATACGAAGTTATAACTCGAC 3‘) and SG_frag_2_Reverse (5‘ ACGCTTACAATTTAGGTGGCACTTTTCGGGGAAATGT 3‘). A third fragment containing the Ae. aegypti PUb-eCFP cassette was amplified from pSL1180-HR-PUbeCFP using primers SG_frag_1_Forward (5’ CTTATACGAAGTTATTATCTTTACATGTAGCTTGTGCATTGAATCCAA 3‘) and SG_frag_1_Reverse (5‘ GAAGTTATGATCCAGACATGATAAGATACATTGATGAGTTTGG 3‘). All PCR bands were gel excised and purified using a QIAquick Gel Extraction Kit (QIAGEN, Germantown, MD, USA). Bold sequences in SG_frag_1_Forward and Vector_R, in SG_frag_1_Reverse and SG_frag_1_Forward, in Vector_R and SG_frag_1_Reverse, and Vector_F and SG_frag_2_Reverse represent complementary sequences. The complementary ends of each fragment were then annealed in a single reaction using an In-Fusion Cloning Kit (Takara, Ann Arbor, MI, USA).
4.5.2. Injections for pBac-mediated transformation
Aedes albopictus transgenic lines were created by injecting preblastodermal embryos with pBac in vitro transcribed, capped mRNA (300 ng/µl) and the donor plasmid pXL-BACII_FRT_3xP3DsRed_FRT3_loxN-PUbeCFP-lox2272 (150 ng/µl) in 1x EIB. Injected embryos were kept moist for two days to allow completion of embryonic development before transferring to water with three droplets of larval food slurry (described in Ae. albopictus insect rearing above). Survivors were sexed at the pupal stage and backcrossed to WT individuals in small groups of 10-15. G1 offspring were collected for 2-3 gonotrophic cycles and screened for the presence of the transgenic marker (DsRed or eCFP fluorescent protein) at the larval or pupal stage. Positive G1 were again backcrossed individually to establish transgenic lines.
4.5.3. Injections for recombinase-mediated cassette exchange
Pre-blastoderm embryos of an Ae. albopictus landing site line for Cre-RMCE (line 17A1; landing site construct = pXL-BACII_FRT_3xP3DsRed_FRT3_loxN-PUbeCFP-lox2272) were injected with either the recombinase helper plasmid (150 ng/µl) or with in vitro transcribed, capped recombinase mRNA (190 ng/µl) and a donor plasmid (250 ng/µl) in 1x EIB. Survivors were sexed at the pupal stage and backcrossed to WT individuals (see the Manassas, VA collected population described above) individually or in small groups of 1-3. G1 offspring were collected for 2-3 gonotrophic cycles and screened for the presence of the transgenic marker (3xP3AmCyan) at the larval or pupal stage. Positive G1 individuals were backcrossed individually.
An additional Ae. albopictus transgenic line (1A3) with a sex-linked landing site (the landing-site integrated 1.3 Mb upstream of the Nix locus as determined via inverse PCR and was only observed in males) was injected with recombinase helper plasmid (150 ng/µl) and a donor plasmid (250 ng/µl) in 1x EIB. Survivors were sexed at the pupal stage and backcrossed to WT individuals individually or in small groups of 4-12. G1 offspring were collected for two gonotrophic cycles and screened for the transgenic marker (3xP3AmCyan) at the larval or pupal stage. Positive G1 were again backcrossed individually.
4.6. Embryonic microinjections of D. suzukii
Germline transformation with piggyBac constructs was carried out as previously described [11]. A mixture of the piggyBac donor construct (500 or 700 ng/µl) and the phsp-pBac or the phsp-HypB transposase helper (200 or 300 ng/µl) was injected into WT embryos. In a third series of experiments, the two helper plasmids were combined at 200 ng/µl each with 500 ng/µl donor plasmid. G0 adults were individually (unless otherwise stated) crossed to WT flies, and G1 flies were screened for fluorescence. Segregation tests were conducted by outcrossing the transformants to WT flies, and transgenic lines were established from single G1-positive adults.
4.7. Copy number variation (CNV) and linkage analysis of transgene integrations by droplet digital PCR
The number of genomic integrations of the transgene cassettes in Ae. aegypti was analyzed by ddPCR probing for the eGFP or DsRed marker genes. The reference gene was mEF1, a one-copy gene in the Ae. aegypti genome. ddPCR was performed with the Biorad QX200 and Auto-DG System. 20 µl CNV reactions contained 20-100 ng Ae. aegypti genomic DNA, 1x ddPCR Supermix for probes (Biorad #1863010), 1x primer-probe mix target gene (FAM-labeled), 1x primer-probe mix reference gene (HEX-labeled), and 1U EcoRI (NEB: EcoRI cuts within the transgene constructs but not within the PCR amplicons). Droplets were generated with the Automated Droplet Generator. PCR cycling conditions (deep well block) were: 95°C for 10 min, 40x [94°C for 30 sec, 60°C for 1 min], 98°C for 10 min, 4°C hold. The ramp rate was 2°C /sec. Primer and probes were prepared and stored as a 20x primer-probe mix consisting of 18 µM each of forward and reverse primer and 5 µM probe (the final concentration of primers and probe in the reaction was 900 nM and 250 nM, respectively). Primers and probe used for target gene EGFP were: EGFP-for (P106) = 5’caaagaccccaacgagaagc, EGFP-rev (P108) = 5’gtccatgccgagagtgatcc, EGFP-probe = 5’ FAM-cgatcacatggtcctgctgg-BHQ1. Primers and probe used for target gene DsRed were: DsRed-for (P49) = 5’gatccacaaggccctgaagc, DsRed-rev (P50) = 5’gctccacgatggtgtagtcc, DsRed-probe = 5’ FAM-tcgttgtgggaggtgatgtc-BHQ1. Primers and probe used for reference gene mEF1 were: mEF-for (P63) = 5’tccggtttgcctacgatacc, mEF-rev (P64) = 5’actgggcagttgtactcacg, mEF-probe = 5’ HEX-tcgggaatgggtgaattgca-BHQ1.
The distribution of positive and negative droplets in each well was analyzed individually and the threshold corrected manually if necessary.
For linkage analysis, the CNV experiment was once conducted with restriction digest and once without and the results were compared. If two transgene cassettes are linked on the same chromosome, the copy number obtained from the undigested reaction is approximately ½ of the digested reaction. Linkage analysis requires cautious preparation of genomic DNA to avoid shearing forces. Depending on the extent of shearing and the distance of the two integration sites, the value obtained from undigested DNA can converge towards the value of the digested sample.
4.8. Analysis of genomic integration sites by inverse PCR
Genomic locations of the transgene constructs were determined by inverse plus nested PCR according to the following protocol: genomic DNA (600 ng) was digested with 4 U MspI in a 20 µl reaction for 1 hr at 37°C. Digested DNA was immediately precipitated, pelleted, and re-dissolved in 50 µl TE buffer. The complete amount of MspI-digested DNA (50 µl) was used for the subsequent self-ligation reaction in 350 µl total volume containing 1x T4 Ligation Buffer and 2 µl T4 DNA ligase (NEB, 400,000 U/ml) overnight at 16°C. Ligated DNA was precipitated and dissolved in 50 µl. 3 µl DNA was PCR amplified in 20 µl containing 1x Phusion Flash High-Fidelity Polymerase Mastermix (Thermo Scientific F548S) and 500 nM of each primer. Cycling conditions for touchdown inverse PCR (iPCR) were: 98°C for 10 sec, 5x [98°C for 1 sec; Tm + 5°C for 5 sec, reduced by 2°C per cycle; 72°C for 1 min], 30x [98°C for 1 sec; Tm + 5°C for 5 sec; 72°C for 1 min], 72°C for 1 min, 12°C hold. The annealing temperature was adjusted for each primer pair.
The iPCR reaction was either directly purified by agarose gel electrophoresis and sent to sequencing, or diluted 1:100, and 1 µl used for the (semi-) nested PCR. (Semi-) nested PCR reactions were identical to the iPCR reactions. Cycling conditions for (semi-) nested PCR (nPCR) were: 98°C for 10 sec, 30x [98°C for 1 sec, Tm minus 5°C for 5 sec, 72°C for 1 min], 72°C for 1 min, 12°C hold. Primers for probing the 3’ piggyBac integration site were mfs12 (5’CCTCGATATACAGACCGATAAAACAC)/P139 (5’CTTTTATCGAATTCCTGCAGC) (iPCR) and mfs34 (5’CGTACGTCACAATATGATTATCTTTCTAGG)/P139 (nPCR), and for probing the 5’ piggyBac integration site mfs10 (5’ACGACCGCGTGAGTCAAAATGACG)/ mfs11 (5’ATCAGTGACACTTACCGCATTGACA) (iPCR) and mfs10/mfs31 (5’CGACTGAGATGTCCTAAATGCACAG) (nPCR).
4.9. Transformation efficiency calculation
Minimal transformation efficiency was calculated as the number of obtained independent transgenic lines divided by the total number of adult G0 survivors of each injection. The actual transformation efficiency in several cases was higher than the numbers reported here for several reasons. All emerged G0 adults were counted for efficiency calculation, including sterile individuals who are often excluded from such calculations; this was necessary to avoid bias in efficiency calculation between the individual backcross and the group backcross experiments, where sterile individuals would not have been detected. Moreover, not all potentially present independent integration events were detected because i) only the individuals with CN = 1 were analyzed for their genomic integration site by inverse PCR. Thus, potentially different integration events in individuals with CN > 1 were not detected; ii) only a subset of positive G1 offspring of most G0 families was individually backcrossed and analyzed, while the rest was group backcrossed and kept as backup; iii) in some experiments with lots of positive G0 families not all families were further characterized (V19, V369, V370); iv) some G1 found dead in cages did not produced sufficient quality DNA for digital and inverse PCR. Finally, it has to be noted that individual backcrossing in V369 and V370 injections was only started at the G3 generation after two generations of group backcrosses of positive individuals. Not all positive G1 were used for backcrossing. Some integration events might have been lost.
4.10. Statistics
Differences in fitness parameters and transgenesis efficiencies between phsp-pBac and phsp-HypB injections or helper plasmid and helper mRNA injections were analyzed with single factor ANOVA, at alpha = 0.05.
5. Conclusions
piggyBac mRNA strongly increased the piggyBac transformation efficiencies in Ae. aegypti and Ae. albopictus. The use of mRNA probably circumvents inefficient transcription from pBac helper plasmids. In contrast, RMCE efficiencies could not be improved in the two tested Aedes species. Here, the limiting factor might not be the recombinase availability in the first place but rather the more complex reaction dynamics.
Our results show that using pBac mRNA could be a promising approach to improve transformation efficiencies in other insects, especially if low efficiency is suspected to be caused by low transposase expression from the helper plasmid. Moreover, it could also be applied to other transposases such as Hermes, Minos, or Hobo. Therefore, the use of transposase mRNA has the potential to make a change in the field of insect transformation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S2: Comparative pBac helper plasmid and pBac mRNA injection data in Ae. aegypti. Data from 13 injection experiments using the phsp-pBac helper plasmid over six years and nine injection experiments using capped pBac mRNA over five years are displayed. Exp. 1-13 are identical to the ones shown in Suppl. Table S1. Exp. 12, 13, 21, and 22 were performed in parallel with eggs from the same female cohort. Injections 14 and 15 were performed in the WT Orlando laboratory strain and all other injections in the Higgs White Eye strain. Both V368 plasmid helper injections (# 12, 13) yielded just 14 adult survivors versus 34 from the mRNA helper injections. Therefore, a larger experiment might have produced transgenic offspring from the plasmid injections. It can still be said with confidence, however, that the transposition efficiency is much lower than that using mRNA, as with an average of one transgenic event per three G0 adults, at a similar efficiency to the mRNA injections, the plasmid injections should have resulted in five to seven independent transgenic events for these 14 G0. “no. injected embryos” represents the number of black embryos 24 h post injection; “hatch rate” = no. larvae / no. black eggs (%); “eclosion rate” = no. adults / no. larvae (%), “no. single/group crossed” shows the number of G0 backcrossed individually or in groups, respectively. “group size” refers to the number of G0 individuals per family; “no. transgenic events” refers to the number of independent transposition events observed, which can be higher than the number of transgenic families if more than one founder per family was positive or/and if more than one independent event occurred in a G0 individual. In the case of the mRNA injections, these numbers are given as minimal numbers, as in none of the families all positive G1 offspring were analyzed molecularly. Therefore, additional independent transformation events might have been missed. This is especially the case for the V369 and V370 injections, where G0 were backcrossed mostly in large groups, and only a small subset of G3 of some of the positive families analyzed molecularly; “total number G1 of transg. fam.” refers to the total number of G1 screened across all positive families; “no. pos. G1 of transg. fam.” refers to the number of positive G1 across all positive families within one injection experiment; “G1 with multiple integrations” = given is the number of G1 individuals with more than one donor construct integration out of the total number of G1 analyzed molecularly; “max. no. integr. / individual” = max. number of donor construct integrations detected within one individual by ddPCR; “transf. eff.” is the minimal transformation efficiency, calculated as: number of independent transgenic events/ total number of G0 adults. The transformation efficiency in several families is assumed to be higher (see text). n.a. = not applicable; n.d. = not determined; avg = average; M = male; F = female. Table S3: Integration event analysis in pBac mRNA injections in Ae. aegypti. The detailed backcrossing schemes and integration event analysis are shown for the positive G1, G2, and G3 generations of the pBac mRNA injections. The integration copy number was determined by ddPCR. Subsequently, the genomic integration position for individuals with only one integration was determined by inverse PCR. Individuals with more than one integration (by ddPCR) were not further investigated by iPCR for the genomic integration site. For the injections of the donor plasmids V369 and V370, the individual backcrossing of positive individuals was started at the G3 generation due to limited rearing capacity in the G1 and G2 generations. The positive G1 and G2 individuals were backcrossed to the parental WT line (HWE) as male or female groups. This most likely is the reason why in these two injections, only one to two copies of the donor plasmid per individual were counted by ddPCR, as integrations located on different chromosomes or with sufficient distance on the same chromosome will have segregated in the G1 and G2 generation backcrosses. Table S4: Determination of the fertility rate of microinjected Ae. aegypti G0 adults. Across all injections with pBac helper plasmid or mRNA, the experiments with single G0 backcrosses were identified. If three or more G0 were individually backcrossed in one experiment, it was included in this table to determine the G0 fertility in the individual backcrosses. The information regarding the total number of individually backcrossed G0, the number of fertile individual G0 founders, and the percentage of fertile individual founders is boxed. The percentage of fertile G0 individuals was calculated as no. of fertile single founder families / no. indiv. backcrossed G0 adults * 100. Table S5: pBac mRNA injection data in Ae. albopictus. Displayed are detailed data from an injection experiment using capped pBac mRNA. “no. injected embryos” represents the number of black embryos 24 h post injection; “hatch rate” = no. larvae / no. black eggs (%); “eclosion rate” = no. adults / no. larvae (%), “no. single/group crossed” shows the number of G0 backcrossed individually or in groups, respectively. “group size” refers to the number of G0 individuals per family; “no. transgenic events” refers to the number of independent transposition events observed, which can be higher than the number of transgenic families if more than one founder per family was positive or/and if more than one independent event occurred in a G0 individual. These numbers are given as minimal numbers, as the positive G1 offspring were not analyzed molecularly. Therefore, additional independent transformation events were likely missed, because G0 individuals were backcrossed in large groups; “total number G1 of transg. fam.” refers to the total number of G1 screened across all positive families; “no. pos. G1 of transg. fam.” refers to the number of positive G1 across all positive families within one injection experiment; “G1 with multiple integrations” = given is the number of G1 individuals with more than one donor construct integration out of the total number of G1 analyzed molecularly; “max. no. integr. / individual” = max. number of donor construct integrations detected within one individual by ddPCR; “transf. eff.” is the minimal transformation efficiency, calculated as: number of independent transgenic events/ total number of G0 adults. The transformation efficiency in several families is assumed to be higher (see above). n.a. = not applicable; n.d. = not determined; avg = average; M = male; F = female. Table S6: Comparison of phiC31-RMCE injection data in Ae. aegypti using phsp-phiC31 helper plasmid or capped phiC31 mRNA. Displayed are data from comparative injection experiments at two different helper/donor concentrations into the same landing site line (LSL) (exp. 1 -9); previous phiC31-RMCE injections with the helper plasmid (exp I – V) had been performed in different landing site lines and at different helper/donor concentrations (400/600 and 200/300 ng/µl). The landing site line used in exp. I – III turned out to have two linked, non-segregating landing site construct integrations, as determined by ddPCR after the injections had been performed. Therefore, in some individuals, we found an RMCE event at one landing site and an integration event at the other landing site. These were counted here only as one recombination event per individual. The LSL used for exp. IV and V had only one genomic integration site; “no. injected embryos” represents the number of black embryos 24 h post injection; “hatch rate” = no. larvae / no. black eggs (%); “eclosion rate” = no. adults / no. larvae (%), “no. transg. events” is the number of independent recombination events observed; “recomb. eff.” is the minimal recombination efficiency, calculated as: number of transgenic events/ total number of G0 adults. The recombination efficiency can be higher, as in group backcrosses of G0, the number of infertile G0 is unknown. M = male; F = female; a) RMCE phenotype; one additional family with transient donor phenotype that showed parental phenotype in next generation; b) both integration events; c) one family with transient donor phenotype that showed parental phenotype in next generation; d) all injected eggs counted; e) mixed larvae either from the 400/600 or 200/300 injection; f) 2 RMCE and 2 integration events; g) 1 RMCE and 1 integration event. Table S7: Comparison of Cre-RMCE injection data in Ae. aegypti using phsp-Cre helper plasmid or capped Cre mRNA. Data from comparative injection experiments are displayed at two different helper/donor concentrations into the same landing site line; “no. injected embryos” represents the number of black embryos 24 h post injection; “hatch rate” = no. larvae / no. black eggs (%); “eclosion rate” = no. adults / no. larvae (%); “no. transg. events” is the number of independent recombination events observed; “recomb. eff.” is the minimal recombination efficiency, calculated as: number of transgenic events/ total number of G0 adults. The actual recombination efficiency can be higher, as in group backcrosses of G0, the number of infertile G0 isn’t known; M = male; F = female. Table S8: Comparison of Cre-RMCE injection data in Ae. albopictus using phsp-Cre helper plasmid or capped Cre mRNA. Displayed are data from comparative injection experiments. Plasmid helper injections were performed into two different landing site lines. Exp. 1 was performed with LSL line 1A3, where the landing-site construct integrated 1.3 Mbp upstream of the Nix locus. Thus, all injected embryos were heterozygous for the landing site. Exp. 2 was performed with LSL line 17A1; “no. injected embryos” represents the number of black embryos 24 h post-injection; “hatch rate” = no. larvae / no. black eggs (%); “eclosion rate” = no. adults / no. larvae (%); “total number G1 of transg. fam.” refers to the total number of G1 screened across all positive families; “no. pos. G1 of transg. fam.” refers to the number of positive G1 across all positive families within one injection experiment; “no. transg. events” is the number of independent recombination events observed; “recomb. eff.” is the minimal recombination efficiency, calculated as: number of transgenic events/ total number of G0 adults. The recombination efficiency can be higher, as in group backcrosses of G0, the number of infertile G0 isn’t known; M = male; F = female. Figure S1: Effect of phsp-pBac helper plasmid and phsp-hyPB helper plasmid in Ae. aegypti and D. suzukii. Box and whisker plots (exclusive median) are displayed on the data presented in Table 1 and Table 2 for Ae. aegypti larval hatch rate (a), G0 adult eclosion rate (b), G0 fertility (percentage of fertile G0 families in each experiment) (c), and the minimal transformation efficiency (%) (d), as well as D. suzukii larval hatch rate (e), G0 adult eclosion rate (f),) and the minimal transformation efficiency (%) (g). The horizontal line represents the median, the cross (x) represents the mean; n.s. = no significant difference (p > 0.05, single factor ANOVA). Figure S2: Effect of phsp-phiC31 helper plasmid and phiC31 mRNA injections in Ae. aegypti on larval hatch rate (a), G0 adult eclosion rate (b), G0 fertility (c), and the minimal transformation efficiency (%) (d). Results from injections at low (150/300 ng/µl, exp. 1-5, Table 5) and high (300/500 ng/µl, exp. 6-9, Table 5) helper/donor concentrations were combined for data analysis. Box and whisker plots (exclusive median) are displayed in the data presented in Table 4. The horizontal line represents the median, the cross represents the mean; n.s. = no significant difference (p > 0.05, single factor ANOVA). .
Author Contributions
IH and MFS designed the study in Ae. aegypti, MH and PA designed the study in Ae. albopictus, YY designed the study in D. suzukii, IH and TR performed the pBac transformation experiments in Ae. aegypti, IH, TR, DMC, and HS performed RMCE experiments in Ae. aegypti, MH performed pBac transformation and RMCE experiments in Ae. albopictus, YY performed pBac transformation experiments in D. suzukii; IH, TR, MH, YY, DMC, and HS analyzed and interpreted the data; IH prepared the original draft of the manuscript, and all authors reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
Open Access funding was enabled and organized by Projekt DEAL. This work was supported by the Emmy Noether program of the German Research Foundation (SCHE 1833/1-1 to MFS), the German-Israeli Project Cooperation of the German Research Foundation (SCHE 1833/7-1 to MFS), the European Union’s Horizon Europe Research and Innovation Programme REACT (grant agreement number 101059523 to MFS), the Temporary Positions for Principal Investigators Module of the German Research Foundation (YA 502/3-1 to YY), and the US National Institutes of Health grant R01AI132409 and the Davis Family Endowment to PA and MH.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this published article and its supplementary information files.
Acknowledgments
We thank Bashir Hosseini for excellent technical assistance with the D. suzukii embryonic microinjections, and Robert Harrell II and Channa Aluvihare for performing all embryonic microinjections in Ae. albopictus.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bingham, P.M.; Kidwell, M.G.; Rubin, G.M. The molecular basis of P-M hybrid dysgenesis: the role of the P element, a P-strain-specific transposon family. Cell 1982, 29, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Rubin, G.M.; Spradling, A.C. Genetic transformation of Drosophila with transposable element vectors. Science 1982, 218, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Rubin, G.M.; Spradling, A.C. Vectors for P element-mediated gene transfer in Drosophila. Nucleic Acids Res 1983, 11, 6341–6351. [Google Scholar] [CrossRef] [PubMed]
- O'Brochta, D.A.; Atkinson, P.W. Transformation systems in insects. Methods MolBiol 2004, 260, 227–254. [Google Scholar]
- O'Brochta, D.A.; Sethuraman, N.; Wilson, R.; Hice, R.H.; Pinkerton, A.C.; Levesque, C.S.; et al. Gene vector and transposable element behavior in mosquitoes. J Exp Biol 2003, 206, 3823–3834. [Google Scholar] [CrossRef] [PubMed]
- Handler, A.M. Use of the piggyBac transposon for germ-line transformation of insects. Insect Biochem Mol Biol 2002, 32, 1211–1220. [Google Scholar] [CrossRef]
- Gregory, M.; Alphey, L.; Morrison, N.I.; Shimeld, S.M. Insect transformation with piggyBac: getting the number of injections just right. Insect Mol Biol 2016, 25, 259–271. [Google Scholar] [CrossRef]
- Allen, M.L.; Handler, A.M.; Berkebile, D.R.; Skoda, S.R. piggyBac transformation of the New World screwworm, Cochliomyia hominivorax, produces multiple distinct mutant strains. Medical and Veterinary Entomology 2004, 18, 1–9. [Google Scholar] [CrossRef]
- Handler, A.M.; Harrell, R.A. Germline transformation of Drosophila melanogaster with the piggyBac transposon vector. Insect Molecular Biology 1999, 8, 449–457. [Google Scholar] [CrossRef]
- Handler, A.M.; McCombs, S.D.; Fraser, M.J.; Saul, S.H. The lepidopteran transposon vector, piggyBac, mediates germ-line transformation in the Mediterranean fruit fly. Proceedings of the National Academy of Sciences, USA 1998, 95, 7520–7525. [Google Scholar] [CrossRef]
- Schetelig, M.F.; Handler, A.M. Germline transformation of the spotted wing drosophilid, Drosophila suzukii, with a piggyBac transposon vector. Genetica 2013, 141, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Kokoza, V.; Ahmed, A.; Wimmer, E.A.; Raikhel, A.S. Efficient transformation of the yellow fever mosquito Aedes aegypti using the piggyBac transposable element vector pBac[3xP3-EGFPafm]. Insect Biochem Mol Biol 2001, 31, 1137–1143. [Google Scholar] [CrossRef]
- Nolan, T.; Bower, T.M.; Brown, A.E.; Crisanti, A.; Catteruccia, F. piggyBac-mediated germline transformation of the malaria mosquito Anopheles stephensi using the red fluorescent protein dsRED as a selectable marker. Journal of Biological Chemistry 2002, 277, 8759–8762. [Google Scholar] [CrossRef] [PubMed]
- Labbé, G.M.; Nimmo, D.D.; Alphey, L. piggybac- and PhiC31-mediated genetic transformation of the Asian tiger mosquito, Aedes albopictus (Skuse). PLoS neglected tropical diseases 2010, 4, e788. [Google Scholar] [CrossRef] [PubMed]
- Perera, O.P.; Harrell, R.; Handler, A.M. Germ-line transformation of the South American malaria vector, Anopheles albimanus, with a piggyBac/EGFP transposon vector is routine and highly efficient. Insect Molecular Biology 2002, 11, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, F.G.; Oliveira, S.B.; Rocha, B.C.; Moreira, L.A. Germline transformation of Aedes fluviatilis (Diptera:Culicidae) with the piggyBac transposable element. Mem Inst Oswaldo Cruz 2006, 101, 755–757. [Google Scholar] [CrossRef]
- Grossman, G.L.; Rafferty, C.S.; Clayton, J.R.; Stevens, T.K.; Mukabayire, O.; Benedict, M.Q. Germline transformation of the malaria vector, Anopheles gambiae, with the piggyBac transposable element. Insect Molecular Biology 2001, 10, 597–604. [Google Scholar] [CrossRef]
- O'Brochta D.A.; Alford R.T.; Pilitt K.L.; Aluvihare C.U.; Harrell R.A.; 2nd. piggyBac transposon remobilization and enhancer detection in Anopheles mosquitoes. Proc Natl Acad Sci USA 2011, 108, 16339–16344. [CrossRef]
- O'Brochta D.A.; Pilitt K.L.; Harrell R.A.; 2nd, Aluvihare C.; Alford R.T. Gal4-based enhancer-trapping in the malaria mosquito Anopheles stephensi. G3 (Bethesda) 2012, 2, 1305–1315. [CrossRef]
- Thorpe, H.M.; Smith, M.C. In vitro site-specific integration of bacteriophage DNA catalyzed by a recombinase of the resolvase/invertase family. Proceedings of the National Academy of Sciences, USA 1998, 95, 5505–5510. [Google Scholar] [CrossRef]
- Siegal, M.L.; Hartl, D.L. Application of Cre/loxP in Drosophila. Site-specific recombination and transgene coplacement. Methods in Molecular Biology 2000, 136, 487–495. [Google Scholar]
- Andrews, B.J.; Beatty, L.G.; Sadowski, P.D. Site-specific recombination of the yeast plasmid two-micron circle: intermediates in the binding process. Basic Life Sciences 1986, 40, 407–424. [Google Scholar] [PubMed]
- Horn, C.; Handler, A.M. Site-specific genomic targeting in Drosophila. Proceedings of the National Academy of Sciences, USA 2005, 102, 12483–12488. [Google Scholar] [CrossRef] [PubMed]
- Schetelig, M.F.; Handler, A.M. A Functional Comparison of the 3xP3 Promoter by Recombinase-Mediated Cassette Exchange in Drosophila and a Tephritid Fly, Anastrepha suspensa. G3 (Bethesda) 2013, p. [Google Scholar] [CrossRef]
- Schetelig, M.F.; Scolari, F.; Handler, A.M.; Kittelmann, S.; Gasperi, G.; Wimmer, E.A. Site-specific recombination for the modification of transgenic strains of the Mediterranean fruit fly Ceratitis capitata. Proc Natl Acad Sci U S A 2009, 106, 18171–18176. [Google Scholar] [CrossRef] [PubMed]
- Schetelig, M.F.; Yan, Y.; Zhao, Y.; Handler, A.M. Genomic targeting by recombinase-mediated cassette exchange in the spotted wing drosophila, Drosophila suzukii. Insect Mol Biol 2019, 28, 187–195. [Google Scholar] [CrossRef]
- Haghighat-Khah, R.E.; Scaife, S.; Martins, S.; St John, O.; Matzen, K.J.; Morrison, N.; et al. Site-specific cassette exchange systems in the Aedes aegypti mosquito and the Plutella xylostella moth. PLoS One 2015, 10, e0121097. [Google Scholar] [CrossRef]
- Amenya, D.A.; Bonizzoni, M.; Isaacs, A.T.; Jasinskiene, N.; Chen, H.; Marinotti, O.; et al. Comparative fitness assessment of Anopheles stephensi transgenic lines receptive to site-specific integration. Insect Mol Biol 2010, 19, 263–269. [Google Scholar] [CrossRef]
- Meredith, J.M.; Basu, S.; Nimmo, D.D.; Larget-Thiery, I.; Warr, E.L.; Underhill, A.; et al. Site-specific integration and expression of an anti-malarial gene in transgenic Anopheles gambiae significantly reduces Plasmodium infections. PLoS One 2011, 6, e14587. [Google Scholar] [CrossRef]
- Lobo, N.F.; Hua-Van, A.; Li, X.; Nolen, B.M.; Fraser, M.J., Jr. Germ line transformation of the yellow fever mosquito, Aedes aegypti, mediated by transpositional insertion of a piggyBac vector. Insect Mol Biol 2002, 11, 133–139. [Google Scholar] [CrossRef]
- Adelman, Z.N.; Jasinskiene, N.; Vally, K.J.; Peek, C.; Travanty, E.A.; Olson, K.E.; et al. Formation and loss of large, unstable tandem arrays of the piggyBac transposable element in the yellow fever mosquito, Aedes aegypti. Transgenic Res 2004, 13, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Yusa, K.; Zhou, L.; Li, M.A.; Bradley, A.; Craig, N.L. A hyperactive piggyBac transposase for mammalian applications. Proc Natl Acad Sci USA 2011, 108, 1531–1536. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.A.; Smith, R.C.; Li, X.; Craig, N.L.; Atkinson, P.W. IPB7 transposase behavior in Drosophila melanogaster and Aedes aegypti. Insect Biochem Mol Biol 2013, 43, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Eckermann, K.N.; Ahmed, H.M.M.; KaramiNejadRanjbar, M.; Dippel, S.; Ogaugwu, C.E.; Kitzmann, P.; et al. Hyperactive piggyBac transposase improves transformation efficiency in diverse insect species. Insect Biochem Mol Biol 2018, 98, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Häcker, I.; Harrell II, R.A.; Eichner, G.; Pilitt, K.L.; O'Brochta, D.A.; Handler, A.M.; et al. Cre/lox-recombinase-mediated cassette exchange for reversible site-specific genomic targeting of the disease vector, Aedes aegypti. Sci Rep 2017, 7, 43883. [Google Scholar] [CrossRef]
- Schwirz, J.; Yan, Y.; Franta, Z.; Schetelig, M.F. Bicistronic expression and differential localization of proteins in insect cells and Drosophila suzukii using picornaviral 2A peptides. Insect Biochem Mol Biol 2020, 119, 103324. [Google Scholar] [CrossRef]
- Schetelig, M.F.; Schwirz, J.; Yan, Y. A transgenic female killing system for the genetic control of Drosophila suzukii. Sci Rep 2021, 11, 12938. [Google Scholar] [CrossRef]
- Yan, Y.; Schwirz, J.; Schetelig, M.F. Characterization of the Drosophila suzukii beta2-tubulin gene and the utilization of its promoter to monitor sex separation and insemination. Gene 2021, 771, 145366. [Google Scholar] [CrossRef]
- Yan, Y.; Kobayashi, Y.; Huang, C.; Liu, B.; Qian, W.; Wan, F.; et al. Highly Efficient Temperature Inducible CRISPR-Cas9 Gene Targeting in Drosophila suzukii. Int J Mol Sci 2021, 22, p. [Google Scholar] [CrossRef]
- Jasinskiene, N.; Coates, C.J.; Ashikyan, A.; James, A.A. High efficiency, site-specific excision of a marker gene by the phage P1 cre-loxP system in the yellow fever mosquito, Aedes aegypti. Nucleic Acids Res 2003, 31, e147. [Google Scholar] [CrossRef]
- Ahmed, H.M.M.; Heese, F.; Wimmer, E.A. Improvement on the genetic engineering of an invasive agricultural pest insect, the cherry vinegar fly, Drosophila suzukii. BMC Genet 2020, 21, 139. [Google Scholar] [CrossRef] [PubMed]
- Wimmer, E.A. Insect transgenesis by site-specific recombination. Nature Methods 2005, 2, 580–582. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.R.; Mitchell, C.J. Genetic selection of a flavivirus-refractory strain of the yellow fever mosquito Aedes aegypti. Am J Trop Med Hyg 1991, 45, 399–407. [Google Scholar] [CrossRef]
- Batz, Z.A.; Clemento, A.J.; Fritzenwanker, J.; Ring, T.J.; Garza, J.C.; Armbruster, P.A. Rapid adaptive evolution of the diapause program during range expansion of an invasive mosquito. Evolution 2020, 74, 1451–1465. [Google Scholar] [CrossRef] [PubMed]
- Armbruster, P.; Hutchinson, R.A.; Linvell, T. Equivalent inbreeding depression under laboratory and field conditions in a tree-hole-breeding mosquito. Proc Biol Sci 2000, 267, 1939–1945. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of pBac helper plasmid and pBac mRNA injections in Ae. aegypti on larval hatch rate (a), G0 adult eclosion rate (b), G0 adult fertility (based on individually backcrossed G0 across all pBac injection experiments (Table S4 for numbers) (c), the number of injected embryos per transposition event (d), the number of screened G1 per transposition event (e), the percentage of transgenic G1 per positive G0 family (f), and the minimal transformation efficiency (%) (g). Box and whisker plots (exclusive median) are displayed based on the data presented in Table 3 and Table S2. The horizontal line represents the median; the cross (x) represents the mean; ns = no significant difference, p > 0.05; * = significant difference, p ≤ 0.05; *** = significant difference, p ≤ 0.001.
Figure 1.
Effect of pBac helper plasmid and pBac mRNA injections in Ae. aegypti on larval hatch rate (a), G0 adult eclosion rate (b), G0 adult fertility (based on individually backcrossed G0 across all pBac injection experiments (Table S4 for numbers) (c), the number of injected embryos per transposition event (d), the number of screened G1 per transposition event (e), the percentage of transgenic G1 per positive G0 family (f), and the minimal transformation efficiency (%) (g). Box and whisker plots (exclusive median) are displayed based on the data presented in Table 3 and Table S2. The horizontal line represents the median; the cross (x) represents the mean; ns = no significant difference, p > 0.05; * = significant difference, p ≤ 0.05; *** = significant difference, p ≤ 0.001.
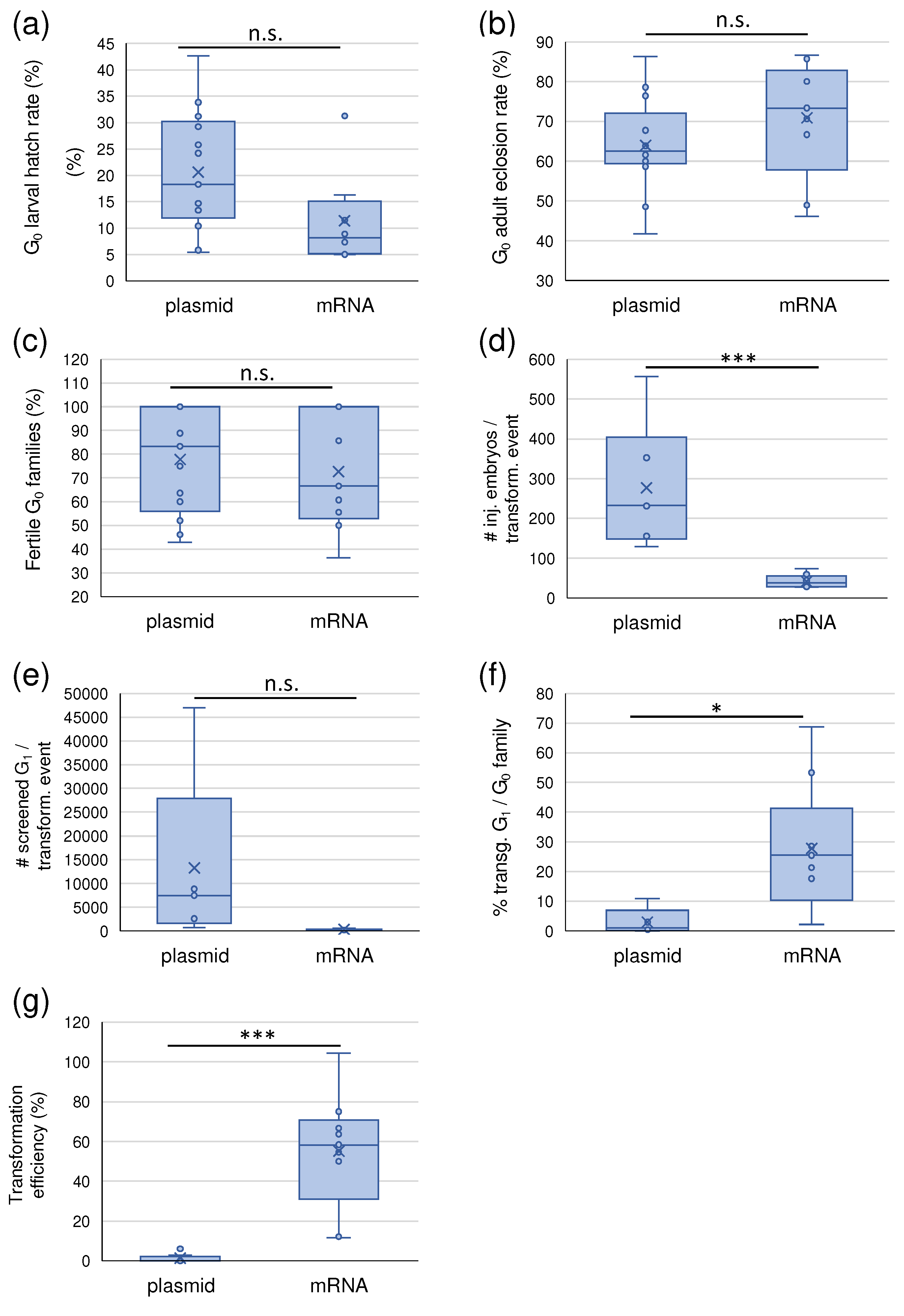

Table 1.
Transformation efficiencies in Ae. aegypti using the standard piggyBac helper or ihypBase helper plasmids.
Table 1.
Transformation efficiencies in Ae. aegypti using the standard piggyBac helper or ihypBase helper plasmids.
| Exp. no | helper template | [helper/ donor] (ng/µl) | donor construct | donor plasmid (insert) size (bp) | no. injected embryos | larval hatch rate (%) | adult eclosion rate (%) | total no. G0 families | % fertile G0 families | no. transg. G0 families | total no. G1 screened | no. transg. events | transf. eff. (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | pBac | 300/150 | AH452 * | (5267) 8649 | 623 | 29.21 | 78.57 | 8 | 100.00 | 4 | n.d. | 4 | 2.80 |
| 2 | pBac | 200/500 | V3 | (3545) 6911 | 234 | 31.20 | 86.30 | 25 | 52.00 | 1 | 8830 | 1 | 1.25 |
| 3 | pBac | 400/600 | V285 | (5180) 8682 | 705 | 25.82 | 64.29 | 18 | 88.89 | 2 | 14966 | 2 | 1.71 |
| 4 | pBac | 300/500 | V286 | (5600) 9102 | 680 | 13.38 | 41.76 | 15 | 60.00 | 0 | 8772 | 0 | 0.00 |
| 5 | pBac | 160/185 | V258 | (6190) 9690 | 922 | 10.41 | 37.50 | 18 | 83.33 | 0 | 6631 | 0 | 0.00 |
| 6 | pBac | 400/600; 228/342 | V258 | (6190) 9690 | 599 | 24.21 | 34.48 | 11 | 100.00 | 0 | 18721 | 0 | 0.00 |
| 7 | pBac | 300/150 | V257 | (7088) 10587 | 1113 | 42.59 | 76.37 | 26 | 100.00 | 2 | 93998 | 2 | 0.55 |
| 8 | pBac | 200/500 | V19 | (3630) 7163 | 257 | 18.29 | 63.83 | 9 | 100.00 | 2 | 5185 | 2 | 6.67 |
| 9 | pBac | 300/500 | V19 | (3630) 7163 | 192 | 33.85 | 61.54 | 11 | 63.64 | 0 | 2679 | 0 | 0.00 |
| 10 | pBac | 200/500 | V19 | (3630) 7163 | 399 | 13.53 | 61.11 | 13 | 46.15 | 0 | 1313 | 0 | 0.00 |
| 11 | pBac | 200/500 | V19 | (3630) 7163 | 462 | 14.72 | 48.53 | 7 | 100.00 | 2 | 1389 | 2 | 6.06 |
| 12 | pBac | 300/300 | V368 | (5835) 11097 | 257 | 5.84 | 60.00 | 7 | 42.86 | 0 | 502 | 0 | 0.00 |
| 13 | pBac | 100/300 | V368 | (5835) 11097 | 148 | 5.41 | 62.50 | 4 | 75.00 | 0 | 382 | 0 | 0.00 |
| avg | 1.46 | ||||||||||||
| 14 | HypB | 100/200 | V286 | (5600) 9102 | 594 | 8.08 | 75.00 | 12 | 75.00 | 0 | 9382 | 0 | 0.00 |
| 15 | HypB | 200/200 | V286 | (5600) 9102 | 730 | 8.63 | 79.37 | 17 | 70.59 | 0 | 7604 | 0 | 0.00 |
| 16 | HypB | 200/200 | V258 | (6190) 9690 | 895 | 6.82 | 65.57 | 13 | 69.23 | 0 | 5199 | 0 | 0.00 |
| avg | 0.00 |
Table 2.
Transformation efficiencies in D. suzukii with the standard piggyBac helper or ihypBase helper plasmids.
Table 2.
Transformation efficiencies in D. suzukii with the standard piggyBac helper or ihypBase helper plasmids.
| Exp. no. | helper template | [helper/ donor] (ng/µl) | donor construct | donor plasmid (insert) size (bp) | no. injected eggs | no. hatched larvae | hatch rate (%) | no. fertile adults | fertile eclosion rate (%) | no. transg. lines | transf. eff. (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ds 1 | pBac | 200/500 | AH443 | (9191) 12576 | 75 | n. d. | n. d. | 25 | n.d. | 4 | 16.00 |
| Ds 2 | pBac | 200/500 | V220 | (7865) 11249 | 1601 | 231 | 14.43 | 65 | 28.14 | 1 | 1.54 |
| Ds 3 | pBac | 200/500 | V221 | (7867) 11252 | 443 | 102 | 23.02 | 9 | 8.82 | 0 | 0.00 |
| Ds 4 | pBac | 200/500 | V146 | (7118) 10503 | 481 | 173 | 35.97 | 20 | 11.56 | 2 | 10.00 |
| Ds 5 | pBac | 200/500 | V183 | (7583) 10968 | 753 | 305 | 40.50 | 43 | 14.10 | 5 | 11.63 |
| Ds 6 | pBac | 200/500 | V184 | (9438) 12823 | 640 | 167 | 26.09 | 17 | 10.18 | 0 | 0.00 |
| Ds 7 | pBac | 200/500 | V185 | (8493) 11878 | 802 | 346 | 43.14 | 53 | 15.32 | 6 | 11.32 |
| Ds 8 | pBac | 200/500 | V188 | (10347) 13732 | 631 | 285 | 45.17 | 55 | 19.30 | 1 | 1.82 |
| Ds 9 | pBac | 200/500 | V213 | (10059) 13443 | 538 | 107 | 19.89 | 27 | 25.23 | 0 | 0.00 |
| Ds 10 | pBac | 200/500 | V215 | (8204) 11589 | 626 | 173 | 27.64 | 29 | 16.76 | 1 | 3.45 |
| Ds 11 | pBac | 200/500 | V226 | (8163) 12054 | 410 | 51 | 12.44 | 9 | 17.65 | 1 | 11.11 |
| Ds 12 | pBac | 200/500 | V227 | (10018) 13909 | 493 | 122 | 24.75 | 12 | 9.84 | 1 | 8.33 |
| Ds 13 | pBac | 200/500 | V228 | (8607) 12498 | 310 | 97 | 31.29 | 13 | 13.40 | 0 | 0.00 |
| Ds 14 | pBac | 200/500 | V250 | (9072) 12963 | 378 | 129 | 34.13 | 28 | 21.71 | 0 | 0.00 |
| Ds 15 | pBac | 200/500 | V229 | (10434) 14325 | 339 | 92 | 27.14 | 26 | 28.26 | 1 | 3.85 |
| Ds 16 | pBac | 200/500 | V251 | (10927) 14818 | 413 | 70 | 16.95 | 10 | 14.29 | 0 | 0.00 |
| Ds 17 | pBac | 200/500 | V265 | (9952) 13337 | 520 | 124 | 23.85 | 32 | 25.81 | 0 | 0.00 |
| avg | 4.65 | ||||||||||
| Ds 18 | pBac | 300/700 | V265 | (9952) 13337 | 582 | 238 | 40.89 | 77 | 32.35 | 5 | 6.49 |
| Ds 19 | pBac + hyPB | 200+200/500 | V221 | (7867) 11252 | 1457 | 271 | 18.60 | 42 | 15.50 | 1 | 2.38 |
| Ds 20 | pBac + hyPB | 200+200/500 | V222 | (7861) 11246 | 1898 | 401 | 21.13 | 34 | 8.48 | 0 | 0.00 |
| Ds 21 | pBac + hyPB | 200+200/500 | V223 | (7865) 11249 | 488 | 82 | 16.80 | 30 | 36.59 | 2 | 6.67 |
| avg | 3.02 | ||||||||||
| Ds 22 | hyPB | 200/500 | V209 | (5538) 8922 | 1081 | 252 | 23.31 | 37 | 14.68 | 2 | 5.41 |
| Ds 23 | hyPB | 200/500 | V265 | (9952) 13337 | 98 | 32 | 32.65 | 8 | 25.00 | 0 | 0.00 |
| avg | 2.70 |
Table 3.
Transformation data in Ae. aegypti using the phsp-pBac helper plasmid or capped pBac mRNA as piggyBac source.
Table 3.
Transformation data in Ae. aegypti using the phsp-pBac helper plasmid or capped pBac mRNA as piggyBac source.
| Exp. no | helper template | [helper/ donor] (ng/µl) | donor construct | donor plasmid (insert) size (bp) | no. injected embryos | hatch rate (%) | eclosion rate (%) | total no. G0 families | no. fertile G0 families | no. transg. G0 families | total no. G1 screened | no. transg. events * | no. transg. events/ G0 | transf. eff. (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | plasmid | 300/150 | AH452 *** | (5267) 8649 | 623 | 29.21 | 78.57 | 8 | 8 | 4 | n.d. | 4 | 1 | 2.80 |
| 2 | plasmid | 200/500 | V3 | (3545) 6911 | 234 | 31.20 | 86.30 | 25 | 13 | 1 | 8830 | 1 | 1 | 1.59 |
| 3 | plasmid | 400/600 | V285 | (5180) 8682 | 705 | 25.82 | 64.29 | 18 | 16 | 2 | 14966 | 2 | 1 | 1.71 |
| 4 | plasmid | 300/500 | V286 | (5600) 9102 | 680 | 13.38 | 41.76 | 15 | 9 | 0 | 8772 | 0 | n.a. | 0.00 |
| 5 | plasmid | 160/185 | V258 | (6190) 9690 | 922 | 10.41 | 67.71 | 18 | 15 | 0 | 6631 | 0 | n.a. | 0.00 |
| 6 | plasmid | 400/600; 228/342 | V258 | (6190) 9690 | 599 | 24.21 | 58.62 | 11 | 11 | 0 | 18721 | 0 | n.a. | 0.00 |
| 7 | plasmid | 300/150 | V257 | (7088) 10587 | 1113 | 42.59 | 76.37 | 26 | 26 | 2 | 93998 | 2 | n.d. | 0.55 |
| 8 | plasmid | 200/500 | V19 | (3630) 7163 | 257 | 18.29 | 63.83 | 9 | 9 | 2 | 5185 | 2 | 1 | 6.67 |
| 9 | plasmid | 300/500 | V19 | (3630) 7163 | 192 | 33.85 | 61.54 | 11 | 7 | 0 | 2679 | 0 | n.a. | 0.00 |
| 10 | plasmid | 200/500 | V19 | (3630) 7163 | 399 | 13.53 | 61.11 | 13 | 6 | 0 | 1313 | 0 | n.a. | 0.00 |
| 11 | plasmid | 200/500 | V19 | (3630) 7163 | 462 | 14.72 | 48.53 | 7 | 7 | 2 | 1389 | 2 | 1 | 6.06 |
| 12 | plasmid | 300/300 | V368 | (5835) 11097 | 257 | 5.84 | 60.00 | 7 | 3 | 0 | 502 | 0 | n.a. | 0.00 |
| 13 | plasmid | 100/300 | V368 | (5835) 11097 | 148 | 5.41 | 62.50 | 4 | 3 | 0 | 382 | 0 | n.a. | 0.00 |
| avg | 1.49 | |||||||||||||
| 14 | mRNA | 182/300 | V96 | (5278) 8813 | 113 | 11.50 | 46.15 | 6 | 3 | 1 | 370 | 4 | 4 | 66.67 |
| 15 | mRNA | 182/300 | V97 | (4307) 7841 | 298 | 5.03 | 73.33 | 11 | 4 | 2 | 514 | 7 | ≥ 2 - ≥5 | 63.64 |
| 16 | mRNA | 300/300 | V370 | (5870) 11131 | 576 | 31.25 | 86.67 | 12 | 12 | 11 | 4388 | > 19 | ≥1 | 12.18 |
| 17 | mRNA | 300/300 | V369 | (6420) 11684 | 520 | 16.35 | 80.00 | 6 | 6 | 5 | 2173 | > 8 | ≥1 | 11.76 |
| 18 | mRNA | 300/300 | V19 | (3630) 7163 | 527 | 8.92 | 48.94 | 28 | 17 | 10 | 1813 | ≥ 24 | 1 - ≥5 | 104.35 ** |
| 19 | mRNA | 100/300 | V19 | (3630) 7163 | 310 | 5.48 | 70.59 | 9 | 6 | 2 | 1195 | ≥ 7 | ≥3 - ≥4 | 58.33 |
| 20 | mRNA | 100/300 | V19 | (3630) 7163 | 646 | 1.61 | 80.00 | 7 | 7 | 3 | 1030 | 4 | 1 - 2 | 50.00 |
| 21 | mRNA | 300/300 | V368 | (5835) 11097 | 449 | 7.35 | 66.67 | 18 | 10 | 5 | 1014 | ≥ 12 | ≥2 - 4 | 54.55 |
| 22 | mRNA | 100/300 | V368 | (5835) 11097 | 281 | 4.98 | 85.71 | 7 | 6 | 5 | 870 | ≥ 9 | 1 - ≥3 | 75.00 |
| avg | 49.02 |
Table 4.
Transformation data in Ae. albopictus using capped mRNA as piggyBac source.
| Exp. no | helper template | [helper/ donor] (ng/µl) | donor construct | donor plasmid (insert) size (bp | no. injected embryos | hatch rate (%) | eclosion rate (%) | total no. G0 families | no. fertile G0 families | no. transg. G0 families | total G1 screened | no. transg events | no. transg. events/ G0 | transf. eff. (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | mRNA | 300/150 | AH452 modified * | (5205) 7845 | 721 | 39.53 | 83.51 | 19 | 19 | 15 | 7049 | ≥ 15 | ≥ 1 | 6.3 % |
Table 5.
phiC31-RMCE injection data in Ae. aegypti using phsp-phiC31 helper plasmid or capped phiC31 mRNA.
Table 5.
phiC31-RMCE injection data in Ae. aegypti using phsp-phiC31 helper plasmid or capped phiC31 mRNA.
| Exp. no | helper template | [helper/ donor] (ng/µl) | donor construct | donor plasmid (insert) size (bp | no. injected embryos | hatch rate (%) | eclosion rate (%) | total no. G0 families | no. fertile G0 families | no. transg. G0 families | total G1 screened | no. transg. events | recomb. eff. (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | phsp-phiC31 | 150/300 | V101 | (4404) 8216 | 510 | 3.14 | 75.00 | 9 | 6 | 0 | 1690 | 0 | 0 |
| 2 | phsp-phiC31 | 150/300 | V101 | (4404) 8216 | 237 | 7.59 | 61.11 | 5 | 5 | 0 | 2851 | 0 | 0 |
| 3 | mRNA | 150/300 | V101 | (4404) 8216 | 502 | 4.38 | 59.09 | 11 | 4 | 0 | 534 | 0 | 0 |
| 4 | mRNA | 150/300 | V101 | (4404) 8216 | 335 | 3.28 | 54.55 | 4 | 2 | 0 | 458 | 0 | 0 |
| 5 | mRNA | 150/300 | V101 | (4404) 8216 | 361 | 3.32 | 83.33 | 7 | 4 | 1 * | 318 | 1 | 10 |
| 6 | phsp-phiC31 | 300/500 | V101 | (4404) 8216 | 443 | 2.71 | 91.67 | 9 | 4 | 0 | 1059 | 0 | 0 |
| 7 | phsp-phiC31 | 300/500 | V101 | (4404) 8216 | 302 | 13.25 | 77.50 | 11 | 9 | 2 | 6068 | 2 | 6.45 |
| 8 | mRNA | 300/500 | V101 | (4404) 8216 | 412 | 2.18 | 100.00 | 9 | 9 | 0 | 6250 | 0 | 0 |
| 9 | mRNA | 300/500 | V101 | (4404) 8216 | 248 | 13.71 | 73.53 | 10 | 6 | 0 ** | 3650 | 0 | 0 |
Table 6.
Cre-RMCE injection data in Ae. aegypti using phsp-Cre helper plasmid or capped Cre mRNA.
| Exp. no | helper template | [helper/ donor] (ng/µl) | donor construct | donor plasmid (insert) size (bp | no. injected embryos | hatch rate (%) | eclosion rate (%) | total no. G0 families | no. fertile G0 families | no. transg. G0 families | total G1 screened | no. transg. events | recomb. eff. (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | phsp-Cre | 450/350 | V20 | (1297) 4997 | 765 | 18.69 | 57.34 | 13 | 12 | 2 * | 9283 | 2 | 2.44 |
| 2 | phsp-Cre | 450/350 | V20 | (1297) 4997 | 165 | 6.67 | 72.73 | 4 | 1 | 0 | 390 | 0 | 0 |
| 3 | phsp-Cre | 450/350 | V20 | (1297) 4997 | 397 | 1.26 | 80.00 | 3 | 1 | 0 | 137 | 0 | 0 |
| 4 | mRNA | 450/350 | V20 | (1297) 4997 | 413 | 10.90 | 73.33 | 4 | 1 | 1 ** | 2419 | 1 | 3.03 |
| 5 | mRNA | 450/350 | V20 | (1297) 4997 | 230 | 5.22 | 58.33 | 3 | 3 | 0 | 996 | 0 | 0 |
| 6 | mRNA | 450/350 | V20 | (1297) 4997 | 348 | 5.75 | 80.00 | 12 | 9 | 0 | 4926 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



