
Submitted:
28 August 2023
Posted:
30 August 2023
You are already at the latest version
Abstract
The large American liver fluke (Fascioloides magna) is considered an invasive trematode parasite in Europe. Its origin dates back before the Cretaceous-Paleogene Mass Extinction, after which it survived at least three population bottlenecks, and successful host switches before it arrived in Europe. The authors review the evolutionary history of F. magna, the distribution by its ancient proboscidean hosts and the probable drivers of switch to the white-tailed deer (Odocoileus virginianus). The review collects knowledge on the biology of intermediate hosts, which help understand the factors that influence the epidemiology of F. magna in aquatic ecosystems. The authors demonstrate the adaptation potential of the parasite by data from both the North American and the European endemics. Finally, the study calls attention to the epidemiological risk of human induced global change, with a special interest to the invasive snail species, Pseudosuccinea columella.
Keywords:
Fascioloides magna
; proboscidea
; host switch
; bottleneck
; virulence
; Pseudosuccinea columella
1. Introduction
The large American liver fluke (Fascioloides magna) belongs to the Digenean subclass of Trematode class. With Protofasciola robusta, Fasciolopsis buski, Fasciola hepatica, F. gigantica, F. nyanzae, Tenuifasciola tragelaphi, and Fascioloides jacksoni, it is a member of Fasciolidae family (Figure 1) [1]. This taxon consists of several endoparasites with human and animal health importance.Their development is indirect, which means their life cycle needs an intermediate host. The fluke possesses both female and male genitals, they are hermaphrodites [2,3] The F. magna can parasitise mostly cervids, less frequently other ruminants and sometimes suids, horses and rodents; contrary to Fasciola hepatica, F. gigantica, and F. buski, the zoonotic relevance of F. magna has not proved [2].
Figure 1.
European representatives of the Fasciolidae family: Fasciola hepatica (in the middle) and Fascioloides magna. Besides the size, “shoulders” differentiate F. hepatica from F. magna.
Figure 1.
European representatives of the Fasciolidae family: Fasciola hepatica (in the middle) and Fascioloides magna. Besides the size, “shoulders” differentiate F. hepatica from F. magna.

The first description of the species was published in 1875 by Bassi. That work was based on the discovery of an isolated population of the parasite in the Regional Park La Mandria near Turin, Italy [3,4]. After this first European detection, several authors described the species during the last decades of the 19th century on the North American continent, and its American origin was confirmed [4,5].
In its final hosts, F. magna causes mild to severe lesions of the liver parenchyma, which affects hepatic function, thus metabolism of the body and performance of the host [6,7,8]. In its life cycle, amphibian snails of the Lymnaeidae family play the role of the intermediate host [5,9,10], therefore the distribution of the parasite depends on freshwater habitats where its intermediate hosts live (Figure 2) [11].
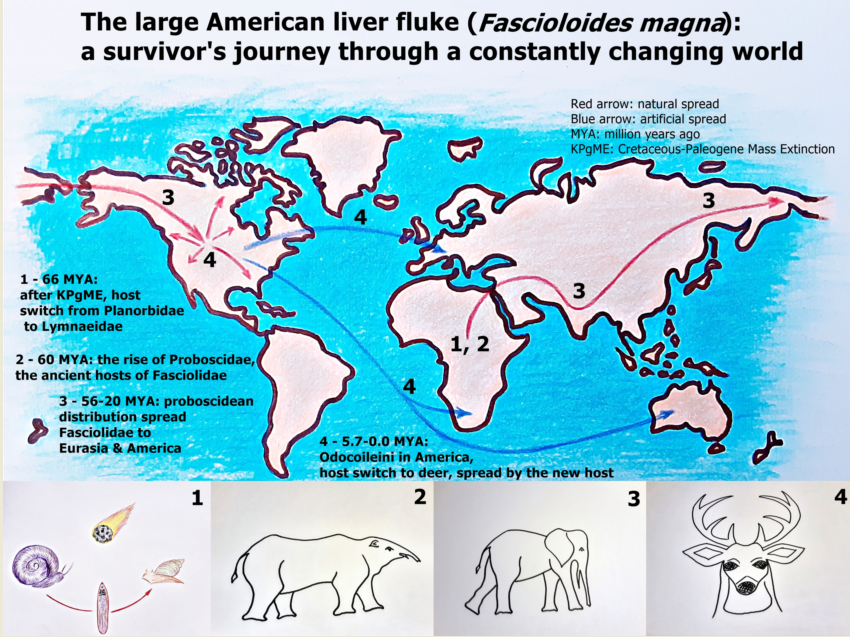
At present, F. magna occupies natural habitats in North and Central America, Europe, South Africa, and Australia [2,3,4]. From its original North American habitats, it travelled to other continents by transport of infected hosts (mostly cervids). The history, especially the European history of the large American liver fluke demonstrates the central role of human activities in the spread of this exotic parasite. For now, F. magna became an important element of global change [12]. Our review attempts to demonstrate those evolutionary, epidemiological, and pathological features, which contributed to its successful distribution.
Figure 2.
Lentic freshwater ecosystem with shallow shores and rich aquatic plant community, which provide ideal habitat for lymnaeid snails (small picture).
Figure 2.
Lentic freshwater ecosystem with shallow shores and rich aquatic plant community, which provide ideal habitat for lymnaeid snails (small picture).

2. Evolutionary history of the large American liver fluke and its relatives
2.1. The Cretaceous-Paleogene Boundary
Little evidence is available on the distribution and host specificity of the early Digenean trematodes [13]. It is hypothesised that before adaptation to vertebrate hosts, these trematodes parasitised molluscs [11]. Some excavations revealed that Digenean trematodes can be found in dinosaur coprolites and mollusc fossils from the Upper (Late) [14] and Lower (Early) [15,16] Cretaceus period; nevertheless Cretaceous-Paleogene Mass Extinction (KPgME) around 66 million years ago (MYA) swept away most host species with their parasites [13].
The most probable cause of KPgME was the Chicxulub asteroid, which ejected enormous amounts of dust, ash, soot, and other aerosols at its impact. The outcome was prolonged sunlight screening, which resulted in months of global blackout. The direct consequence was the extremely cold “impact winter” lasting for a decade. The exaggerated cooling destabilised all trophic levels in the biosphere, which resulted in extinction of most species [17,18]. For parasites, host switch might have been an option to survive KPgME, as it is confirmed in feather lice of birds [19]. In these hardly tolerable climatic conditions, freshwater microhabitats with higher thermal inertia could provide refugia for certain species. The detritus of these swamps and ponds supplied nutrients to the survivors. In freshwater habitats, the biodiversity loss was barely 10%, contrary to the terrestrial 90% [17]. The recovery from this climatic catastrophe needed about 30 years [17,18].
The seismic effect of the asteroid impact generated changes in the volcanic activity of the Deccan Traps resulting in increased CO2 outgassing [20,21]. Due to this extensive volcanism, the greenhouse effect augmented, and a drastic global warming began, which contributed to the formation of the early Cenozoic ecosystem [18,22]. The massive carbon release to the atmosphere finally led to the Paleocene-Eocene Thermal Maximum (PETM) around 56 MYA characterised by global warming, reduced latitudinal temperature gradients, ice-free poles, and ocean acidification [23].
The emergence of the Fasciolidae family dates back to 90-100 MYA, after the fragmentation of Gondwanaland into Africa and South America [24]. Based on molecular genetic analysis, it is confirmed that the lineages of Fasciolopsis and Fasciola diverged cca. 88.1 MYA, long before the KPgME [25]. The recent intermediate hosts of the Fasciolidae family are gastropod species of Basommatophora suborder evolved in the Carboniferous period. Based on the internal transcribed spacers of the ribosomal DNA (ITS-1 and ITS-2) and the mitochondrial 16 S ribosomal DNA, it is confirmed that Lymnaeidae snails, the most important intermediate hosts of Fasciolida family originated during the Jurassic period [26], while Lymnaeidae and Planorbidae families diverged 250 MYA [27], long before KPgME, therefore these snails are also survivors of KPgME.
Both Fasciolidae and their basommatophoran snail hosts evolved before the extreme winter caused by the Chicxulub asteroid impact. They also avoided extinction later, during the extreme hot climate of PETM. Presumably, their characteristic habitat, freshwater ponds, and swamps, mitigated the temperature extremities and provided nutrients for the snails and thus their parasites. The harsh climate change caused aridification of inland habitats, therefore surface water cover decreased resulting in habitat fragmentation [17].
Among these circumstances, isolated subpopulations, living in ponds, creeks, ditches, and shallow lakes almost worldwide, sustained the survivor species in small subpopulations [28]. Extreme habitat fragmentation jeopardises the sustainability of the subpopulations, thus the whole population itself because if a population decreases below the effective population size, its maintenance is questionable. The ability of self-fertilisation reduces the effective population size, whereas hypothetically only one mature individual can multiply itself [11,29]. Several taxa of Basommatophora possess the ability of self-fertilisation, which assures the survival of the population after large-scale losses and contributes to the invasive ability of certain species [30,31].
The gene flow between the subpopulations of the habitat fragments is indispensable in the survival of the species. Lymnaeid snails are very small animals with limited moving ability, therefore they cannot travel very far under their own power. However, these species are maintained during the rapidly changing KPgM, moreover they spread worldwide from their fragmented refugia [26]. The historical spread of Lymnaeid snails is confirmed by the finding of a fossil Galba sp. from the Cretaceous period on an island 1500 km far from the mainland. The migration route cannot be reconstructed; however, the authors presume that an ancient bird should have transported it on its body surface or within its intestinal tract through the Indian Ocean [32]. The same phenomenon is observed in the European distribution of F. magna, whereas ungulate wildlife can transport snails by mud stuck on hair [33]. This route can contribute to the translocation of a limited number of snails. Population level spread of these small gastropods can occur in lotic habitats in the flow direction [34], which is generated by strong current, e.g., by stormwater runoff [35].
The phenotypic plasticity, which helped these mollusc taxa to survive and spread during the climatic extremities of the Cretaceous-Paleogene Boundary (KPgB), is conspicuous recently. These snail taxa can be found in temperate zone floodplains [3,4,5,33], high altitude mountain lakes in the Alps [36] and in the Andean areas of South America [37], and in the tropics [38,39,40].
2.2. Distribution by the Proboscidea order
Within the intensely changing environment between the KPgB and PETM, the Fasciolid parasites met their ancient final hosts, the early proboscideans [1,24]. After the loss of the late Cretaceous megafauna, the ancient mammals began to diversify and filled the ecological niches left by the extinct non-avian dinosaurs [41]. The order of Proboscidea was one of the earliest known modern placental orders in the Late Paleocene, cca. 60 MYA in Africa [42]. The oldest fossils of ancient elephant relatives can be traced back to 45-60 million years [42,43,44]. Due to the drastically increasing temperature, tropical and subtropical forest environments became the dominant habitats globally and the primitive proboscideans lived in swamp ecosystems and fed on freshwater vegetation in riverine or swampy settings [27,41] such as the Lymnaeid snails at that time [28].
The following 40 million years in the Cenozoic Era witnessed the evolutionary history of proboscideans. During this time period, the order diversified and spread almost all over the world by three major radiations [25,43]. The first occurred during the Palaeocene Eocene Boundary (56 MYA), with the diversification and intra-African spread of primitive proboscideans [42,43]. The second radiation took place during the Oligocene early Miocene period when the supervening connection of Africa to Eurasia facilitated the faunal interchange between the two continents. This eventuated 25-20 MYA and was termed Proboscidean Datum Event (PDE), which resulted in rapid global dispersal of the order, by which it reached the Americas [43,45]. After the PDE, the proboscideans became one of the most diverse taxa of the paleofauna [45]. The last radiation took place during the late Miocene/early Pliocene and resulted in the diversification of Elephantidae, the ancestors of the extant two genera, Loxodonta and Elephas [43].
The phylogenetic analyses of the Fasiolidae family support the hypothesis that proboscidean dispersal spread these parasites globally. The basal extant member of the taxon, Protofaciola robusta, parasitises the small intestine of the African bush elephant (Loxodonta africana) and utilises Planorbid snails as intermediate hosts. The next most basal member, Fasciolopsis buski, is also an intestinal parasite with planorbid intermediate host [1]. The whole genome analysis of Fasciolidae revealed that the clade comprising Fasciola and Fascioloides genera split from the ancient lineage between 65 MYA and 55.9 MYA, during the rapid diversification of the Proboscidea order. This split also meant a switch between both habitats (from the intestine to the liver) and intermediate hosts (from Planorbidae to Lymnaeidae) [1,24,25].
The closest relative of F. magna is Fascioloides jacksoni [46], which parasitises the liver of the Indian elephant (Elephas maximus). This fact suggests that these sibling species spread by their primary definitive hosts, which were proboscideans [47,48]. A comparative study of karyotypes and chromosomal location of rDNA genes in F. magna and F. hepatica suggested that Fasciola is the younger genus within the liver pathogen clade of Fasciolidae [49]. Based on the molecular genetic investigations, fossil records, and recent epidemiological and pathological findings, it is very probable that the Fasciola genus evolved as an adaptation to the emerging taxon of ruminants [24]. Though, a recent phylogenetic investigation hypothesised that the oldest member of Fasciola genus is F. nyanzae, therefore the first hosts of this genus was hippopotamuses and subsequent fasciola species jumped to ruminants by an intermediary host, which could be wild boar (Sus scrofa) [50].
2.3. The fall of the proboscideans and the dawn of the ruminants
After the PETM about 34 MYA, a temperature falling and aridification process started. During the Miocene (23.03 - 5.333 MYA), the climate began to cool drastically, which induced changes in the ecosystem. The cooler and, in mid-latitudes, dry and seasonal climate supported grasslands to spread. The early fossil record of grasses dates back to Late Cretaceous; however, until the Miocene, grasses did not form extensive grassland ecosystems [41]. The previously woodland-dominated vegetation turned to more open, savanna-type environments [51,52].
This period coincided with the Messinian Event 6.0 - 5.3 MYA when the Mediterranean dried up and the world’s oceans became less saline [53]. Just after that, the Miocene-Pliocene Boundary (MPB, 5.3 MYA) marks a transition from a late Miocene cooling trend to the early to middle Pliocene warm period, when the Northern Hemisphere was largely ice-free and atmospheric CO2 concentrations were comparable to present-day levels [54]. This interim warming during the global cooling trend turned to the subsequent glacials and led to the unstable Pleistocene with extended arid glaciations, sea level changes, fluctuating atmospheric carbon dioxide and shifting vegetation [53]. During this volatile geological time of MPB, the two sister species of the genus Fasciola, F. hepatica and F. gigantica, diverged [25].
The alteration in vegetation brought dramatic change in nutrient supply of the megafauna. Those taxa could reach evolutionary success, which could adapt to utilisation of grasses as nutrient sources (grazers, GR), while those herbivores, which depended on woodland plants (browsers, BR) began to decline [45]. The diversification of GR and the decline of BR initiated during the Miocene and accelerated through the MPB. This process was more rapid in North America than the rest of the world [52].
Among these conditions, ruminants began to diversify extensively through adaptation to more heterogeneous habitats and seasonal climates; and they colonised a large range of biomes [51]. In the meanwhile, specialist browsers declined in species richness due to the reduced primary production and loss of forest habitats. Proboscideans are mostly browser herbivores, which means that they depend on woody and non-woody dicotyledonous plants, leaves and barks of trees as food source [41,53]. Owing to the enhanced competition with other ungulate clades that evolved specialist grazing ecomorphs, the extinction of the proboscidean species accelerated around 8 MYA, after the onset of grassland-dominated habitats [45,52].
The global temperature and the consequential vegetation shift cannot explain the replacement of proboscideans with grazing-adapted ungulates [41,51]. A paleontological study confirmed that adaptation to grazing lifestyle could be detected in late gomphotheres, a proboscidean taxon, which went extinct in the late Pleistocene [55]. However, another study highlighted that proboscidean fossils from the middle Miocene to the late Pleistocene showed enamel hypoplasia, which can be regarded as the sign of chronic stress, though the reason cannot be specified [56].
Multiple extinction events occurred during the Late Pleistocene, mainly over the last 100,000 years with a peak between 12,000 and 10,000 years ago [41,52]. The most severely affected fauna was large-bodied herbivores above 1,000 kg body weight. Especially in the Americas, where 100% of this sized species became extinct [52]. This era of natural history is characterised by human presence. A consensus is emerging that climate change and early human activities went together in contributing to biodiversity loss of megaherbivore fauna [41]. Top-down effects of human predation, landscape modification, especially controlled use of fire affected the proboscidean populations of the palearctic regions [52,55].
In contrast with proboscideans, the ruminants proved the real winners of the climatic fluctuations of the last 45 million years. After the Eocene-Oligocene cooling and aridification event 34 MYA, and especially during the Miocene Climatic Optimum cca. 18–15 MYA the ruminants diversified and successfully adapted to the heterogeneous habitats and climate of the Earth [51].
2.4. Switch to the cervid host
Within the Ruminantia suborder, Bovidae and Cervidae families evolved in Asia during the Miocene and started colonising a large range of ecosystems [51,57]. By analysing two mitochondrial protein-coding genes and two nuclear introns for 25 species of deer, an investigation estimates that the Cervidae family originated in the Late Miocene, 7.7–9.6 MYA and the common ancestor of the American Odocoileini is dated between 4.2 and 5.7 MYA. The fossil records of American cervid species also supported the molecular findings that Odocoileini entered the American continent during the MPB, while other cervid taxa reside in America probably as a result of a recent dispersal event, cca. a million years later, during the Early Pleistocene [57] or at a much later time, during the presence of the last landbridge between Eurasia and America between 10,000 and 70,000 years ago [58].
The host switch of F. magna from proboscideans to cervids presumably eventuated in North America [1]. In Eurasia, a large number of potential host species were available and neither the proboscidean loss was so severe as in the Americas [24,51,52]. It is very probable that a drastic population collapse of the American proboscidean host species forced the host shift to the sympatric cervid species [1], which could be the ancestor of the recent white-tailed deer (Odocoileus virginianus).
On the North American continent, white-tailed deer were the most abundant host species for thousands of years. It is estimated that before the arrival of the European settlers, between 23 and 40 million white-tailed deer inhabited North America. Due to its extensive hunting, cca. 500,000 animals remained by the late 1800s. During the 20th century, 41 states made a law for the protection of the species, therefore the currently estimated population is between 14 and 20 million individuals in the United States [3].
On the North American continent, F. magna has five focuses: (1) the Great Lakes region; (2) the Gulf coast, lower Mississippi, and southern Atlantic seaboard; (3) northern Pacific coast; (4) the Rocky Mountain trench; and (5) northern Quebec and Labrador [59]. Though potential habitats, appropriate intermediate and final host species exist even between the main focuses, considerable expansion cannot be detected [5]. Phylogenetic analysis of F. magna strains originating from different American focuses confirmed that most populations share haplotypes with low genetic difference, however the eastern and western populations show genetic distance [60,61]. The patchy distribution and the contradictory genetic overlap of the geographically separated subpopulations are suggested to be the consequence of the final host’s population loss, which forced the partial switch to alternative hosts belonging also to the Cervidae family. This phenomenon is frequently detected in host-parasite systems during host extinction when the parasite prefers host-switching to species which are phylogenetically close to its original host [62].
3. Recent European distribution
The European history of F. magna proves substantially eventful. After the first introduction, which remained a local endemic in La Mandria, later immigration events resembled a territorial conquest. Importation of infected white-tailed deer and wapitis from endemic areas is supposed to be the main route of disease introduction to Europe [3,4,5]. The parasite seemed to switch to Eurasian cervid species without impediment, presumably due to the phylogenetic proximity of the previous and the new hosts [3,62]. The exact beginning of the process can hardly be determined, notwithstanding two further focuses formed in Central Eastern Europe during the 20th century [3,4,5]. One of those emerged in Czechia and expanded to Poland or vice versa, whereas Poland also imported wapitis from the United States during the 19th century [63].
The other focus formed in the floodplain forests of the Danube basin. The first cases were detected in Slovakia by the Hungarian border [64], then a year later, on the opposite side of the border river Danube in Hungary [65]. This ecosystem provides ideal conditions for continuous spread through the continent by the river’s flow direction [3,4,5,66,67].
In Croatia, F. magna occurred around the turn of the millennium in the Kopacki Rit Nature Park at the confluence of the Danube and the Drava [68,69,70]. Some years later, during the 2007-2008 hunting season, its presence was reported from Serbia, near Belgrade at the Danube-Sava confluence [71,72]. In Bavaria, a suspicious case was detected in 2009 near the Czech border, then the endemic was confirmed later, in 2011 [73]. The relevance of tributary streams of the Danube as potential expansion routes is confirmed several times. In Slovakia, the Vah/Vág and the Hron/Garam open gates to spread opposite of the flow direction [67]. By the Croatian-Hungarian border river Drava, the same phenomenon can be observed [69,70,74,75].
Molecular investigation of the mitochondrial genome of F. magna confirmed that the North American population possesses a higher level of genetic diversity [4,61,76], while the European populations belong to two phylogenetic lineages. The investigations revealed that the population of the Italian focus originated from the northern Pacific coast, while the two other European focuses acquired infection from South Carolina; the Gulf coast, lower Mississippi, and southern Atlantic seaboard focus [4,5].
4. European final hosts
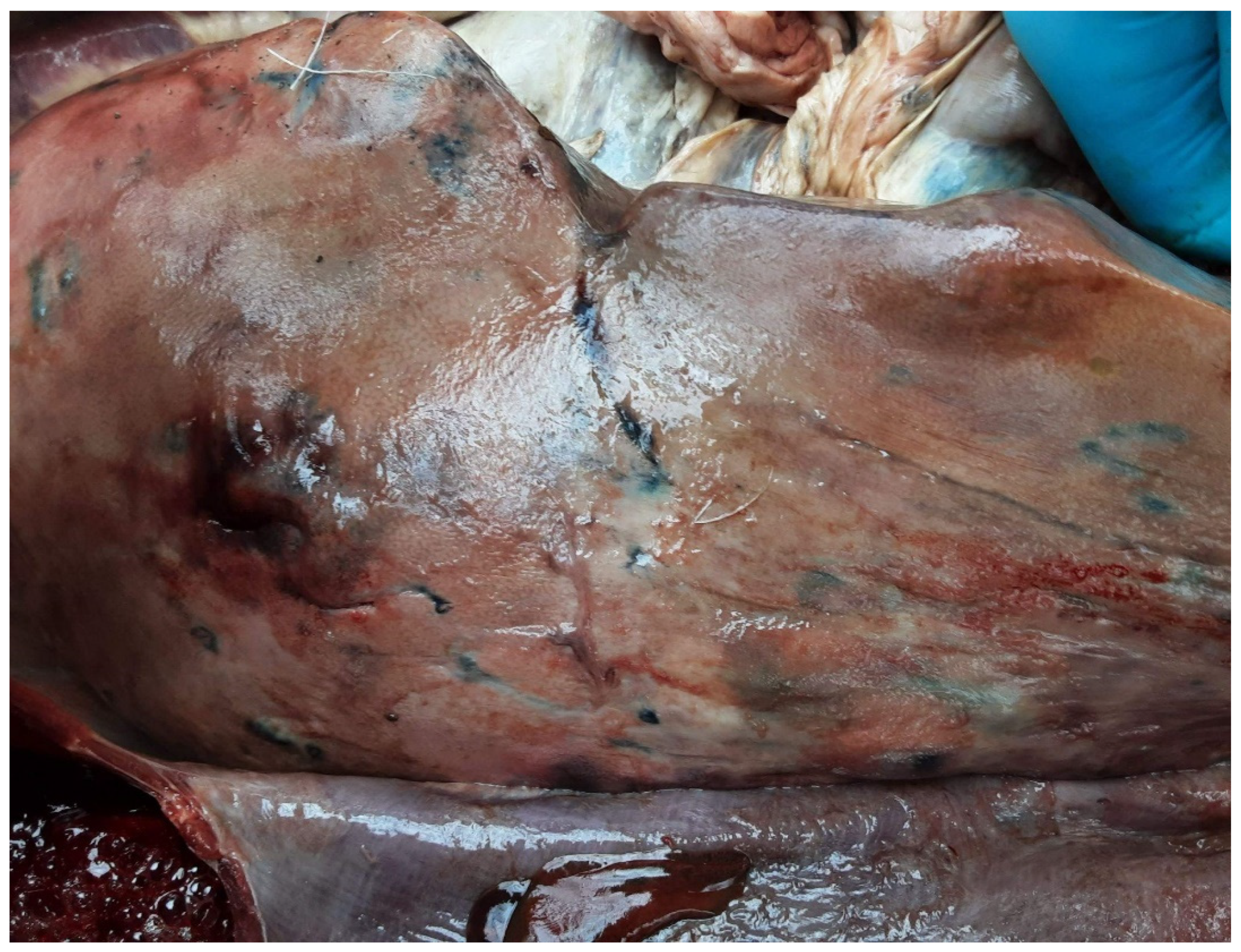
The final hosts of F. magna are sorted into three categories, based on the fluke development and the lesions in the host’s body. After entering the small intestines, the cercariae penetrate the intestinal wall, begin their migration through the abdominal cavity towards the liver. Within the liver parenchyma they are looking for potential mating partners causing characteristic lesions of larval migration. These lesions are brownish stripes (Figure 3), which contain haematin, a product of red blood cell disintegration as a consequence of tissue and bloodvein damage caused by the juvenile parasites migrating through the parenchyma [5,8].
Figure 3.
Brownish stripes on the surface of an enlarged liver as a sign of juvenile fluke’s migration.
Figure 3.
Brownish stripes on the surface of an enlarged liver as a sign of juvenile fluke’s migration.
The definitive hosts behave in the parasite’s life cycle very similarly to the white-tailed deer, the native American final host of the large American liver fluke. When the parasite finds a partner or gives up further searching, it terminates its migration. As a result of the parasite migration, the excessive hepatocyte damage causes fibroblast proliferation, thus progressive fibrogenesis, which leads to the formation of thick-walled pseudocyst (Figure 4) around the parasites [6]. The wall of the pseudocyst contains connective tissue and smooth muscle elements, while inside the cyst, brownish-greenish detritus and 1-3 parasites can be found [3,5,6]. Histopathological investigation of the remnant parenchyma demonstrates a general chronic inflammation, infiltration by macrophages and eosinophils, degeneration of hepatocytes, wall thickening in both blood veins and bile ducts [6,8]. Some of the bile ducts are incorporated into the pseudocysts, thus the mature flukes can shed eggs through this route [5]. In definitive hosts, prepatent period is at least 30 weeks, and the lifespan of the flukes can reach five years [3]. In Europe, red deer (Cervus elaphus), fallow deer (Dama dama) are confirmed to be definitive hosts [3,5], while the assessment of roe deer (Capreolus capreolus) has been varying recently [74,75].
In dead-end hosts, the parasite may reach its sexual maturity similarly to definitive hosts. However, the host's body reacts differently than definitive hosts. The wall of the pseudocyst is very thick, and no exit is created by bile duct incorporation, therefore, in dead-end hosts, the parasites are ambushed, and eggs cannot fulfil their destiny. These hosts generally do not show severe clinical signs of fascioloidosis [4,77]. Dead-end hosts can shed eggs when a mechanical trauma injures the pseudocyst and the eggs can reach the bile ducts, thus the intestinal tract and the environment [3]. In Europe, wild boar (Sus scrofa) is the most important dead-end host in natural habitats. It is suggested that hunting offals containing pseudocysts with eggs can serve as a risk for parasite spread, especially if these remnants are utilised by carnivores [4]. In the Croatian territory of F. magna distribution, Konjevic et al. experienced that wild boar can develop thinner-walled pseudocyst, similarly to definitive hosts [78]. Among farm animals, cattle, pigs, and horses can play the role of dead-end hosts [3,5]. In heavily infected American areas, decreased performance and high rate of abattoir liver condemnation are observed in pasture reared cattle [4]. In Europe, a similar phenomenon has not been confirmed yet [3].
Figure 4.
Extremely large thick-walled pseudocyst in a red deer’s (Cervus elaphus) liver parenchyma containing huge amount of greyish-brownish detritus.
Figure 4.
Extremely large thick-walled pseudocyst in a red deer’s (Cervus elaphus) liver parenchyma containing huge amount of greyish-brownish detritus.

Neither aberrant hosts contribute to the maintenance of F. magna. In their body, the parasite cannot reach sexual maturity. After the infection, a very excessive wandering of juveniles occurs, which causes severe tissue damage in both body cavities. The haematin containing greyish-brownish stripes of migration tracks can be seen in the liver, under the serosa of the visceral organs and the abdominal cavity, and also in the lungs and under the pleura. The tissue degeneration causes multiple organ failure, which leads to the death of the animal before the flukes mature, mostly in 4-6 months after infection. Pseudocyst formation in aberrant hosts cannot be detected [3,5]. Among the aberrant host, mostly smaller sized ruminant species, such as mouflon (Ovis aries musimon), sheep, goat [5], and chamois (Rupicapra rupicapra) [78], can be found.
4.1. The special role of roe deer
The roe deer (Capreolus capreolus) was considered an aberrant host, whereas a severe, frequently fatal course of infection could be observed in this host, especially after the emergence of F. magna in a new area. In these cases, necropsy investigation of the carcasses did not detect pseudocyst formation [3,5,12] and egg production [66] in roe deer. Currently, increasing amounts of evidence confirm that in roe deer, pseudocyst development [79], egg production [74,75] and egg shedding [80] are common phenomena in endemic areas of Europe.
Among potential definitive hosts in Europe, roe deer is the only one with a solitary lifestyle. These animals live, feed and drink in a limited territory, they encounter their conspecifics during the mating season, thus the acquisition of infection is incidental [74,79]. In different areas, a very diverse prevalence can be detected. In Czechia, 70-80%, in Slovakia, 60% [5], in Hungary, 36.7% [75], and in Poland, 13.6% [66] prevalence was confirmed in roe deer. This also supports the hypothesis that roe deer data cannot reflect the real epidemiological situation in a region [79]. Different roe deer territories represent different habitats with varying epidemiological risk of F. magna infection [74].
However, roe deer, also belonging to the Capreolinae subfamily, is a closer relative to white-tailed deer than any other Old World Cervidae [57]. Considering that parasites’ host switch can occur most commonly between species in phylogenetic proximity [62], roe deer was expected to serve as an efficient host for F. magna to fulfil its life cycle. The higher mortality rate is presumed to be the consequence of the small body-size of this species, which barely stands the damage caused by large-sized parasites. This hypothesis is supported by the fact that fallow deer, the next smallest Eurasian cervid species, also show a significantly high loss during F. magna infection, though it is considered a definitive host based on its epidemiologic role and the pathological course in the liver parenchyma [3].
However, in areas where the infection has been established for years, a host-parasite co-evolutionary process is observable. In the Croatian distribution area, after 20 years of presence, the investigated 34 roe deer showed chronic infection and the signs of reinfection in 20%. This means that a considerable proportion of the infected population does not die from fascioloidosis [74]. A very similar phenomenon is observed in a Hungarian area where the infection was detected first a few years before. In this investigation, the researchers found exclusively chronically infected roe deer in almost 40% of the examined 60 carcasses. Moreover, a considerable population loss of the study area was not detected as a result of F. magna emergence [75]. Both investigated endemics are part of the Danube basin distribution of F. magna, which had been occupied for more than two decades [5,65].
The very beginning of the European F. magna invasion was characterised by a strong impact on naive cervid populations. Reduced reproduction performance and trophy quality increased natural loss were observed after the first emergence of the parasite [68,81]. After two decades of attendance in Central Eastern Europe, F. magna appeared to lose its virulence. In an investigation in Bavaria 2019-2020, Sommer et al. determined a 5.9% prevalence in red deer (N=640), of which 76.3% was mild infection. However, in wild boars, fallow deer and roe deer, no infected individuals were detected by investigating 60 carcasses totally. The authors concluded that very low prevalence of the parasite in the examined red deer population is attributable to the high proportion of mild, thus barely noticeable cases. Based on their findings, the researchers suggested that F. magna was underdetected in its settled endemics [8].
At the European occurrence of F. magna, roe deer was evaluated as an aberrant host, whereas this small-sized cervid suffered heavily from the parasite. Fatal cases were common, and serious population dynamic impact was detected [3,4,5]. The recent studies on roe deer confirmed that the parasite and its host got into balance [74,75]. Since the first detection of the parasite in the Danube basin, only some generations of roe deer were born, and due to the territorial lifestyle of roe deer, the encounter between the populations of the host and the parasite was not as extensive as in the case of red deer. [74,79]. The adaptation of roe deer populations to the new parasite is less probable than vice versa. In a coevolutionary process, both the host and the parasite affect each other's population size, which alters the polymorphism in both populations. These cycles of coevolution are more detectable in parasites rather than in the hosts [82]. Those roe deer populations, which were surveyed in the 2020s, showed no significant loss, thus gene frequency alteration. It is suggested that the virulent strains of F. magna became the victims of the arms race between the host and the parasite whereas they were lost together with the hosts they killed [75]. In its invasive range, a parasite must surrender its virulence for the favour of population maintenance. This phenomenon is especially characteristic for generalist parasites, which are highly adaptive to heterogenous environments [83].
5. Intermediate hosts of F. magna
Current intermediate hosts of F. magna are referred to the gastropod suborder Basommatophora comprising more than 300 species in five families. The most abundant families of this taxon are Lymnaeidae, Planorbidae, and Physidae involving 90% of all species within Basommatophora [26]. The Planorbidae family is the largest, comprising 250 species. Lymnaeidae and Physidae families have cca. 100 and 80 species, respectively [84]. These families possess several intermediate host species for other digenean Trematodes with human health and veterinary relevance [26].
On the American continent, six snail species proved to serve as an intermediate host of the large American liver fluke, i.e., Lymnaea modicella, L caperata, Galba (syn. Lymnaea) bulimoides, Fossaria (syn. Lymnaea) parva, Stagnicola (syn. Lymnaea) palustris nuttalliana and Pseudosuccinea columella [3,4,5,9,10]. However, in Europe, only two species, the Galba truncatula and Radix labiata/peregra are confirmed to be able to acquire infection and produce viable cercariae in natural circumstances [4,9,67].
The identification of susceptible intermediate host species presents significant difficulties. On the one hand, infection prevalence in snail populations even within confirmed endemics is very low, less than 1% [12,85,86]. On the other hand, the appropriate morphological identification of snails is almost impossible, and their phylogenetic classification changes time and again [26,87,88].
During the history of malacological research, 1143 species of the Lymnaeidae family were described, however fewer than 100 species remained after synonymisation of identical ones. Most genus names, except for Radix, are based on phenotypic resemblance instead of sound evolutionary and phylogenetic considerations [26]. Within the Radix genus, R. peregra and R. labiata proved to be conspecifics, based on karyotype analysis [89] and molecular genetic investigation of internal transcribed spacers, ITS-2 sequence [77,87,88]. Among the proven American intermediate host snails, Lymnaea modicella and Fossaria (syn. Lymnaea) parva are suspected to correspond to G. truncatula species [26]. Recently, molecular methods have been developed for proper species identification of potential intermediate host snails [37,90,91].
Based on the two nuclear internal transcribed spacers of the ribosomal DNA (ITS-1 and ITS-2) and the mitochondrial 16 S ribosomal DNA of 38 lymnaeid species, Correa et al. demonstrated that Lymnaeidae family comprises three separate phylogenetic clades, of which C1 originated on the American continent and incorporates the genus Galba. The two other clades are both Old World originated; C2 is Eurasian; while C3 is Australian-Indopacific originated and the latter incorporates Radix genus, with the most important intermediate hosts of F. gigantica. This investigation proved that the recently Eurasian distributed G. truncatula diverged from its New World sister species 145-200 MYA [26].
5.1. Susceptibility of the intermediate hosts
Among the known intermediate host species, the susceptibility and cercaria producing efficiency are not identical. In Europe, G. truncatula provides a more suitable environment for development of the flukes than R. labiata, however the latter is more abundant and can tolerate acidic environments, therefore its distribution is wider [3,4,5,9,77]. In the case of F. hepatica, the ability of G. truncatula and R. labiata to act as an intermediate host and produce viable cercariae proved similar [93]; though, a previous study confirmed the opposite [94]. In two further potential lymnaeid hosts, Radix ovata and Stagnicola palustris, the larval development breaks off and the snail perishes [3]. Though, in S. palustris, F. hepatica can complete its development [95].
Within a certain species, individual differences can be observed, as bigger sized, well-developed snails can produce more cercaria [3,10,12], and older snails are more resistant to the infection [3,5], while experimental infection of snails with miracidia originating from a different region resulted in slower cercaria production [5].
The success of infection is also affected by environmental factors. In Austrian habitats, stony soil with poor vegetation is a place where the prevalence of infection is the lowest, however the snail density is also very low. In the same investigation, the researchers found that the infection reaches its peak in June and July when the water temperature is between 20.6-20.8 oC, though the optimum for the miracidia (Figure 5) to infect snails is 22-25 oC [86]. At very high summer temperature, the snails aestivate reducing their metabolism to avoid exsiccation and heat shock. Therefore, in hot weather, the density of detectable snails reduces. As a consequence, spring born snails begin the reproduction during the fall, and their offspring overwinters and reproduces by the next May-June [3].
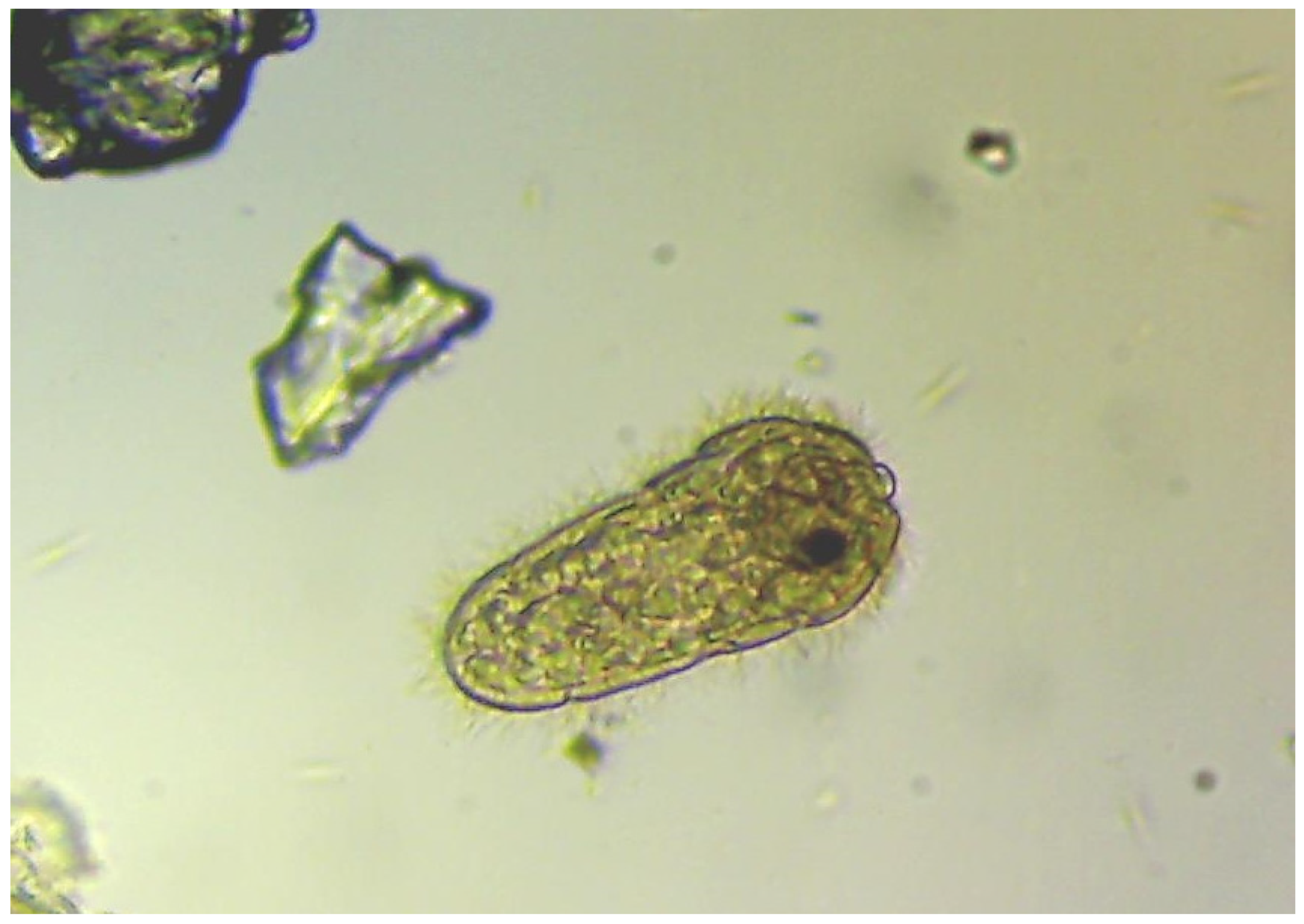
Figure 5.
Free-swimming miracidium after hatching: besides cercaria, this developmental stage of Fascioloides magna can be found in the environment when ambient temperature is between 22-25 oC.
Figure 5.
Free-swimming miracidium after hatching: besides cercaria, this developmental stage of Fascioloides magna can be found in the environment when ambient temperature is between 22-25 oC.
5.2. The course of infection in intermediate hosts
In the body of the intermediate host, an asexual multiplication takes place. The miracidia, which penetrate the foot or the mantel region of the snail, after reaching the pulmonary sac, transform to sporocysts and produce mother rediae. Mother rediae migrate to the hepatopancreas and rear daughter rediae, which leave the mother rediae and the process is repeated again. The cercariae emerge in the daughter rediae and leave the snail’s body [2,3]. This process lasts for 6-7 weeks [12]. As a result of this asexual multiplication, from one miracidium 164-210 [10] or even 1000 [59] cercariae could be produced. The cercariae occur in the environment between May and October allowing the infection of final hosts in this period of the year [96].
Being infected with trematode larvae, the snails can show the signs of functional disorders of the kidney, the gonads, and the digestive glands [97]. Mostly they decrease or even stop reproduction. In pre-adult snails, the infection can completely destroy the genitals [3]. In adult snails, or in more resilient species, e.g., P. columella, the reproductive disorder can be temporal [9]. In pre-adult snails, the development pattern changes, as both parasitic gigantism and decreased shell growth can be observed [5,10]. As a consequence of extensive immune defence, the snails deplete their reserves, which resulted in population decrease some weeks after the infection [9].During the immunological activities, the snails’ haemocytes with their pattern recognition receptors, such as C-type lectins a
nd toll-like receptors (TLR) play the central role. These cells can phagocyte trematode larvae. By recognition of pathogen associated molecular pattern (PAMPs), C-type lectins mediate the immunological process, which enhance phagocytosis (Alba, 2019) and modulate humoral response [11]. In the case of trematodes, the PAMPs are carbohydrates, e.g., mannose, which take place on the surface of the sporocyst [98]. The hemocytes’ TLRs, the primary modulators of the innate immune response, initiate the proinflammatory signalling pathways, which results in acute phase response and redox killing contributing to an effective cellular cytotoxic immune response [38].
The efficiency of this process shows differences both intra and interspecifically. Pseudosuccinea columella, which confirmed to be an intermediate host of F. magna in America, tolerates the infection with the parasite [9,99], while the members of the Galba genus can survive the experimental infection in 30-60% depending on mainly the snails’ size [9,10]. The intraspecific resistance against trematodes was studied in P. columella. In some regions of Cuba, a F. hepatica resistant strain was detected. These snails have some morphological alteration, a different pattern of the mantle; however only 0.17% genetic difference can be identified. The resistant populations are highly adaptive to the environment as well. They can live in soft water with acidic pH where the mollusc diversity of the ecosystem is low, which mitigates the competition with other snail populations. Their resistance to F. hepatica needed a fitness cost, whereas they possess lower fecundity rates, delayed egg hatching, thus diminished net reproduction rate. The rearrangement of the resources from perivitelline fluid production to immune functions is supposed to be in the background. The F. hepatica resistant P. columella strain did not prove to be resistant to other trematode larvae [38].
Rondelaud et al. experimentally infected G. truncatula snails originated from four French habitats. They investigated the survival rate and cercaria production after F. magna infection. They confirmed that significantly different survival rates can be detected both between the populations and between the certain individuals. The authors ascertained that small-sized individuals can produce lesser cercariae after infection with single miracidium than their larger-sized counterparts [10].
On the American continent, the intermediate hosts of F. magna seem more resilient to the infection, presumably due to the long-lasting co-evolution with the parasite; contrary to their European counterparts, which were given only decades for adaptation [3]. On the other hand, observations with F. hepatica prove that parasite infection affects the local snail populations most severely while immigrants’ survival rate is higher [100].
5.3. Self-fertilisation: a trade-off between population persistence and population health
The worldwide distribution of basommatophoran snails is a result of successful recovery after population losses. The most important European intermediate host of F. magna, the G. truncatula has the ability to reconstitute the population by less than 10 survivors [101]. This extremely low effective population size is due to the high, about 90%, selfing rate of the species [34,102]. By self-fertilisation, the population surrenders the benefits of outcrossing mating strategy risking genetic load. Due to the extensive inbreeding, invasive populations of these snail species have lower genetic diversity than their native populations in their original distribution territories [103,104].
The recurrent bottleneck effects decrease the genetic variability of the concerned populations. This is supposed to cause vulnerability to the environmental factors, however G. truncatula reached a wide-spread distribution with its heterozygote deficiency [29,34]. A study investigating Galba truncatula confirmed that a very low, 2% rate of outcrossing did not result in demographic consequences of genetic load in this species [105]. The same phenomenon can be observed in other invasive species, such as water hyacinth (Eichhornia syn. Pontederia crassipes) and P. columella, of which the invasive populations possess significantly depleted genetic diversity [29,106].
On the other hand, increased susceptibility to pathogens can be detected as a result of inbreeding. Highly inbred snail populations show exaggerated predisposition to trematode infection, therefore higher infection prevalence can be observed in them, which results in higher infection risk in final hosts [11,29]. Galba truncatula, and P. columella, a lymnaeid snail with similar selfing ability, prove more effective host for parasites than the locally distributed non-invasive snail species [29,106]. A comparable phenomenon can be noticed in Biomphalaria pfeifferi, in Senegal. A dam construction, which altered the water salinity of the Senegal River, selected a highly inbred population of B. pfeifferi. Before the dam construction, only 1% of the snails belonged to this species. After the completion of the barrages, 70% of the snails were B. pfeifferi. Whereas most snails within the affected river section showed high susceptibility to parasite infection, Schistosoma mansoni trematode began a population expansion in the region, causing human health risk [107].
5.4. Environmental demands of snail hosts
The geographical distribution of the potential snail host can determine the occurrence of trematode parasites [11], therefore knowledge on environmental demands is indispensable to epidemiological research. The most relevant requirements of lymnaeid snails have not changed since the KPgB, for 66 MYA [17,28]. These snails prefer lentic or slowly flowing freshwater habitats with easily warming water by the shallow shorelines [108,109]. However, the generalist members of the taxon, e.g., G. truncatula, R. labiata, can be found in Alpine regions as well [36,88,110]. They prefer increased availability of nutrients, which is assured by progressive eutrophication (Figure 6) as a consequence of human disturbance [36,108]. They hardly tolerate acidic pH [4] because this condition, especially below pH 6, disturbs the shell development. However, in high calcium supply, the disadvantageous effect of acidic pH is mitigated [111]. Excessive water pollution, especially with high levels of heavy metals are avoided by snails, and the free-swimming stages of trematodes are also very sensitive to these pollutants [112].
Figure 6.
Conspicuous signs of eutrophication in a fishpond: diverse swampy vegetation due to the thick silt layer and the whole surface is covered by duckweeds (Lemnoidae) as a result of agricultural pollution, thus nutrient availability. This type of habitat is appropriate for Lymnaeidae, therefore agricultural activity can increase the risk of fascioloidosis in European cervid populations.
Figure 6.
Conspicuous signs of eutrophication in a fishpond: diverse swampy vegetation due to the thick silt layer and the whole surface is covered by duckweeds (Lemnoidae) as a result of agricultural pollution, thus nutrient availability. This type of habitat is appropriate for Lymnaeidae, therefore agricultural activity can increase the risk of fascioloidosis in European cervid populations.

Water flow velocity has a central role in habitat suitability. Any factor that slows down the water flow in a lotic freshwater habitat can affect the population size of snails. This phenomenon could be observed in Africa after the construction of dams, which expanded the suitable habitat of planorbid snails, increasing the risk of human schistosomiasis caused by Schistosoma mansoni, a parasitic trematode [113,114]. The same issue is discussed in Slovakia and Hungary because of the construction of the Gabčikovo-Nagymaros hydroelectric dam, after which the F. magna occurred within the most affected river section [61]. On the other hand, stormwater runoffs (Figure 7) after heavy rainfalls can remove even half of the macroinvertebrates from the riparian zones of lotic ecosystems [35], which reduce the local number of potential intermediate hosts of trematodes.
Figure 7.
Stormwater runoff by the Drava River, a Fascioloides magna endemic area. These events can remove even half the snail populations from the riparian zone of the river and can contribute to the snail and parasite spreading in the flow direction.
Figure 7.
Stormwater runoff by the Drava River, a Fascioloides magna endemic area. These events can remove even half the snail populations from the riparian zone of the river and can contribute to the snail and parasite spreading in the flow direction.

Dam constructions do not affect exclusively the water flow but can create artificial barriers in the way of migration of aquatic fauna, which can also contribute to the invigoration of host-parasite systems. Diama Dam on Senegal River, after its completion inhibited the migration of a predator prawn, Macrobrachium vollenhovennii. This prawn consumes pond snails, thus their population decrease resulted in planorbid snail population expansion in prawn-free river sections, which increased the risk of human schistosomiasis in the region [113].
Agricultural cultivation, especially the extensive use of irrigation channels elevates the nutrition level of freshwater habitats, and climate change amplifies this disadvantageous effect of agriculture [115]. In Europe, this phenomenon can be observed in the Ruhr River water reservoir system. Within the interconnected freshwater ecosystem, the water flow velocity is very low, the water temperature is mild, the sediment accumulation and phytoplankton production are increased, thus eutrophication progressively runs. In these conditions, a huge Radix auricularia population is established providing a stable environment for a diverse trematode community with both cyprinid and anatid parasites among them [108,109]. In tropical environments, the ambivalent impact of irrigation channels, both the advantage for agriculture and the disadvantage for health, is more conspicuous. Perennial irrigation channel use in Egypt strongly correlates with schistosomiasis incidence in humans [114].
In both lentic and lotic habitats, the floating aquatic plants escalate water flow restriction, thus sediment accumulation, and consequently intensifies the eutrophication process (Figure 8) [116]. This phenomenon is widely investigated in African freshwaters where the water hyacinth (Eichhornia syn. Pontederia crassipes) is an aquatic ecosystem invader originating from South America. This plant creates a dense, mat-forming vegetation, allowing muddy substrate to accumulate around, which provides food, shelter, and oviposition site for Biomphalaria sudanica, the intermediate host of human schistosomiasis [117,118]. In Europe, Hydrocharis laevigata, which is also a South American invader plant, began to spread in non-freezing water habitats. The floating rosettes of the plant block the light, thus fosters anoxic conditions, which contribute to progressive eutrophication, and diversity loss of native flora and fauna [116]. Due to the climate warming, invasive plant species can extremely accelerate the eutrophication process in European aquatic ecosystems [115,119].
Figure 8.
Floating aquatic plants, such as Nymphoides peltata in the picture, accelerate eutrophication in lotic habitats by flow restriction and sediment accumulation. Some of these plants are invasive out of their natural range and global warming can enhance their distribution.
Figure 8.
Floating aquatic plants, such as Nymphoides peltata in the picture, accelerate eutrophication in lotic habitats by flow restriction and sediment accumulation. Some of these plants are invasive out of their natural range and global warming can enhance their distribution.

5.5. Human-mediated distribution of epidemiological risk of snails
In the EU, 96 invasive aquatic plant species are recorded, of which 30% has considerable impact on the ecosystem. The main route through invasive plants entering the European ecosystem is ornamental trade [119]. Most invaded countries in Europe are France, Italy, Germany, and Hungary, with almost 100 invasive plant species in their aquatic ecosystems [115].
With ornamental aquatic plants, also snails are imported mostly unintentionally [67], thus the aquarium hobby introduces 41% of the exotic snail species. In most cases, imported exotic snails do not increase epidemiological risk, whereas they have 80% lower parasite richness than native molluscs [120], which can be explained by Enemy-Release Hypothesis. Hosts can lose their native parasites establishing their invasive range [103]. However, the imported molluscs can generate significant epidemiological risk [40,121,122].
Pseudosuccinea columella is a natural intermediate host of F. magna in North America [9,99]. This snail possesses a considerable invasive capacity, whereas it can be found in 22 countries worldwide [39], in various habitat types from tropical climate to high altitudes with very low temperature [39,121]. Initially, it occurred in botanical gardens in Europe. It is supposed to be imported by ornamental aquatic plants. The first wild population of the species was recorded between 2004-2006, in France by the Lot River [123]. By this time, P. columella occurred in different European countries, such as Spain, Portugal, Greece [9], Romania, Italy [39]; Austria, Latvia, Czechia [121] and in Hungary [121,124].
In France, where the first wild population was detected, P. columella aggressively colonised freshwater ecosystems, mainly interconnected water reservoir systems. It can tolerate acidic soil and cool habitats. In competition with native lymnaeid snails, P. columella overwhelms both G. truncatula and Omphiscola glabra. The population decrease of native snails is more rapid in G. truncatula than in O. glabra [121].
Most populations of P. columella are highly susceptible to trematode infections. Fasciola hepatica [39] and F. gigantica [39,40]; F. magna [9,99], and Fasciola nyanzae [122] can complete their life cycle utilising P. columella as intermediate host. In an experimental infection with F. hepatica, 100% infection prevalence could be reached in P. columella [123].
In an Egyptian agricultural environment, irrigation channels are occupied by water hyacinth, which provides ideal habitat for P. columella. As a result, P. columella became the most dominant species in that aquatic ecosystem, almost half of the snail individuals belonged to this species. After the local parasite, F. gigantica colonised the invasive species, a spill-back mechanism could be observed. The incidence of F. gigantica increased after the settlement of the exotic snail [40]. A very similar phenomenon was noticed in Lake Kariba where also the water hyacinth provided ideal conditions for P. columella invasion. After strengthening of the snail population, the hippopotamus population of the lake faced a severe Fasciola nyanzae infection, which affected their population dynamics due to the native parasite spill-back by the exotic vector [122].
Pseudosuccinea columella can extremely quickly adapt to allopatric parasite strains. Pankrác et al. infected Oregon originated snails with Czech F. magna miracidia experimentally. As a result, 74% of the snails became infected [9]. In comparison, experimental infection of G. truncatula proved successful in 31-81% [10], while in natural circumstances, the F. magna larvae can be found in 0.02-0.03% of the G. truncatula [12,85,86]. In allopatric challenge trials, infection of G. truncatula with F. magna miracidia is less efficient than the encounter of sympatric hosts and parasites [5].
6. Conclusion
Advantageous host replacements and environmental adaptation characterised the evolutionary history of the Fasciolidae family. The change in the intermediate host, from Planorbidae family to Lymnaidae, happened just after the KPgME, which was caused by a climatic catastrophe [1,24,25]. Among those extreme circumstances, the fittest and most abundant potential hosts should be chosen for successful survival. Considering that Planorbidae family is also a very successful taxon with plenty species, worldwide distribution [84], and ability of selfing, thus population recovery after loss [125], it is supposed that shift to the Lymnaeidae family is settled locally in those small habitat fragments where the parasite could reach only lymnaeid snails. Though, this random host switch proved advantageous.
The high adaptation capacity of F. magna to potential intermediate hosts can be realised currently. Dreyfuss et al. investigated a Colombian originated Galba sp. by experimental infection with allopatric, Czech F. magna and French F. hepatica through five successive snail generations. They confirmed that during that relatively short period, F. magna could adapt to the new host well, whereas both snail survival and cercaria production increased from generation to generation. In the case of F. hepatica, this rapid adaptation could not be observed. [126].
The adaptation to highly invasive snail species guarantees the survival and spread of the parasite.The excessive spread of P. columella in Europe might open new gates for further distribution of F. magna. This snail can tolerate a wider range of environmental factors than native pond snails, such as G. truncatula [121]. Progressive eutrophication, which is enhanced by poor landscape management, e.g., ill-considered construction of irrigation channels [40] or unintentional import of invasive aquatic plants with ecological impact [122], provides suitable habitats for P. columella. The ability of invasive species to facilitate each other in their invasive range is investigated in a non-native fish and non-native snail interconnection. The authors ascertained that the common presence of invasive species has a stronger impact on the native ecosystem than if they occurred separately. The non-native elements of the flora and fauna can alter the community structure via alteration of the food web and facilitate each other in the ecosystem. This phenomenon is the invasional meltdown [127].
Exotic aquatic plants can change the flow velocity, the sediment deposition, the primary production of the ecosystem [116,119], thus the living conditions of potential intermediate hosts of trematodes [40]. The extensive agricultural activity as a risk factor for F. magna infection is confirmed [67]. The increased nutrient content of water enhances the growth of pondweeds, which provide food and shelter for snails. The slow current in water reservoir systems also threatens with population expansion of snails [108]. The aggressive colonisation of such interconnected aquatic systems by P. columella was witnessed in France [121] where the species was detected in a natural ecosystem firstly in Europe [123]. In its invasive range, P. columella successfully displaces G. truncatula [121]. The coexistence of F. magna and P. columella in a natural habitat has not been studied, yet.
In the final hosts, Fasciolidae can always be found in the most wide-spread and abundant taxa. During the golden era of Proboscideans, cca. 60-8 MYA, the parasite is supposed to spread by proboscidean radiation [1]. The decline of this host taxon and the rise of the ruminants, during the last 20 million years, promoted the host switch [24]. The opportunistic host selection of the Fasciolidae is conspicuous. Among their hosts, elephants, hippos, other ungulates, and even humans appear. It is hypothesised that this odd mixture of current definitive hosts is the result of wide-spread extinction of the ancient hosts, which forced the parasite to shift to alternative hosts [1].
The large American liver fluke shows very good adaptive capacity. During its evolutionary history, all known host switches proved unimpeded. The jump from proboscideans to Odocoilinae is supported by indirect evidence, however during the rapidly changing Pliocene when the megafauna of the Americas decreased, Odocoilinae were the most abundant ungulate taxa [1]. Much later, when the European settlers extirpated the white-tailed deer from the vast majority of its North American territories, the parasite sustained in subpopulations of its native host, however it adapted to further host species [5].
The smart adaptation to new host populations can be seen recently both in North America and in Europe. Invasion of new territories always causes some negative effects in the host population while the host and the parasite get accustomed to each other. In a North American endemic area, severe clinical signs and considerable rate of loss could be observed in a freshly infected moose population while the previously infected populations showed mild signs of the infection [81]. In Europe, the populations of large-sized deer species adapted to the parasite in some years after the emergence [12]. The most spectacular habituation could be seen in the roe deer species. At the onset of the European invasion, this species was assessed to be an aberrant host of F. magna whereas almost all animals died in 6 months after the infection [3,5]. Two decades later, chronically infected roe deer individuals could be detected in two independent investigations in the Drava basin [74,75].
The ability of self-fertilisation is also a characteristic of outstanding adaptive capacity. The selfing ability helps species to survive population bottlenecks during severe loss or occupation of new distribution areas [75]. Though trematodes are mostly outcrossing animals, self-fertilisation can be detected in 2% in F. hepatica [128]. In its invasion range, a parasite must use an alternative strategy to assure the maintenance of the population even in the lack of potential mating partner [81]. By selfing, the parasite temporarily gives up the benefits of sexual reproduction in favour of potential expansion. In an investigation carried out within a new distribution area of F. magna, the researchers found 23.5% proportion of selfing in red deer specimens. The eggs originating from selfing did not show significant fitness loss, whereas embryonic development and hatching ability of inbred eggs were similar to outcrossers [75].
In the European distribution area, a pathological variation in wild boar, a dead-end host, could be observed. By investigating 12 wild boars in Croatia, the researchers found a large pseudocyst with a thin wall in one carcass. This pseudocyst formation allows the flukes to survive for a longer time, mature and produce eggs. In this case, the wild boar, which at present is evaluated to be a dead-end host, might contribute to the maintenance, and spread of the parasite in the future [78].
In host-parasite interactions the two parties continue a trench warfare trying to defeat each other. Meanwhile they change each other’s population size, the numbers of both hosts and parasites fluctuate over time [82]. In the case of F. magna, considerable population dynamic effect on the host populations could be seen within the freshly infected areas. The mutual adaptation between the host and the parasite eventuated in years after the onset of the invasion [8,81]. As if the parasite would cause the absolutely necessary minimal damage to the host population for the sake of subsistence.
From the view of final hosts, F. magna can be considered an environmentally transmitted parasite, which does not depend on host survival. Based on the Curse of the Pharaoh hypothesis, this type of parasites just ‘sit-and-wait’ in the environment during the population decrease of hosts as it can be observed in the case of Bacillus anthracis [129]. However, the environmental survival of F. magna is not as long as of B. anthracis. Besides, the real environmental phase of the life cycle is very short. Only the free-swimming miracidia and cercariae live de facto in the environment. The longest stage out of the final host takes place in the intermediate hosts, which must also face the harmful effects of the parasite [3]. Moreover, in the life cycle of F. magna, a highly self-fertilising snail taxon [29,34,102,106] meets the mostly outcrosser parasite [75]. By obligate selfing, the host loses the genetic variation, which impedes the adaptation ability during the warfare with its parasite. According to the Red Queen hypothesis, this genetic alteration can lead to the extinction of the host population [130].
Despite extreme inbreeding in Basommatophora taxon, intermediate host species are not threatened by trematode burden. Though invasive populations, which are genetically more monomorphic, are more susceptible to the trematode infection than native populations [29,106]. However, this phenomenon does not lead to collective extinction of snails and their trematode parasites. On the contrary, it increases the epidemiological risk for the final hosts [39,40,107]. Arakelyan et al. investigated a parthenogenetic lizard population, which showed a similar parasite load as the outcrossing population despite its lower genetic variability. The authors concluded that the superiority of sexual reproduction cannot prevail if a large difference exists between the dispersal rates of the hosts and the parasites, and self-fertilising strains possess extra resistance to the parasite [131].
Highly virulent parasites exert considerable influence on the population dynamics of their hosts, therefore high prevalence of infection always causes population decline [132]. On the other hand, if the parasite meets a heterogenous host population, significantly less virulent parasites will evolve during the coevolutionary process [133]. In complex communities, where multiple parasites and the presence of predators affect the population dynamics of the host, the accommodation can be more rapid [83]. The parasite must balance between the transmission success and virulence, thus reproduction success [129].
In the evolutionary history of F. magna, more host population declines and bottlenecks, such as KPgB climatic catastrophe, proboscidean extinction, white-tailed deer population collapse, and small-scale translocation to Europe, eventuated. None of this host loss is confirmed to be due to the effect of the parasite. However, it survived all these disadvantageous events owing to its opportunistic adaptation to different types of environmental alterations. The large American liver fluke has the ability to utilise new final and intermediate hosts, and the determination to adjust its virulence to the hosts tolerance. These features are supposed to support the parasite to occupy new habitats, final and intermediate hosts. For efficient forecasting of this process, further research should be necessary with the following aims: to determine potential intermediate hosts (1); to determine if F. magna and P. columella share any ecosystem in Europe (2); if yes, assessment of epidemiological effect of mutual occurrence of the two non-native species through the invasion meltdown phenomenon (3); to survey the alteration of disease course in different types of hosts, especially in wild boars to assess the epidemiological consequence of a potential change in pathology of fascioloidosis (4).
Author Contributions
“Conceptualization, Á.C.; literature review, Á.C., T.H., G.N.; writing—original draft preparation, Á.C., T.H..; writing—review and editing, G.N..; visualization, Á.C..; supervision, G.N. All authors have read and agreed to the published version of the manuscript.” The authors confirm that they all have contributed substantially to the work reported.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lotfy, W.M.; Brant, S.V.; DeJong, R.J.; Le, T.H.; Demiaszkiewicz, A.; Rajapakse, R.J.; Perera, V.B.; Laursen, J.R.; Loker, E.S. Evolutionary origins, diversification, and biogeography of liver flukes (Digenea, Fasciolidae). Am. J. Trop. Med. Hyg. 2008, 79, 248. [CrossRef]
- Kassai, T. (1999). Digenea. In Veterinary Helminthology. Publisher: Butterworth-Heinemann, Oxford, United Kingdom, 1999.
- Malcicka, M. Life history and biology of Fascioloides magna (Trematoda) and its native and exotic hosts. Ecol. Evol. 2015, 5, 1381-1397. [CrossRef]
- Juhász, A; Stothard, J.R. The giant liver fluke in Europe: A review of Fascioloides magna within cervids and livestock with considerations on an expanding snail-fluke transmission risk. Adv. Parasit. 2022, 119, 223-257. [CrossRef]
- Králová-Hromadová, I.; Juhásová, L.; Bazsalovicsová, E. The giant liver fluke, Fascioloides magna: past, present and future research. Springer International Publishing: Heidelberg, Germany, 2016. [CrossRef]
- Marinković, D.; Kukolj, V.; Aleksić-Kovačević, S.; Jovanović, M.; Knežević, M.The role of hepatic myofibroblasts in liver cirrhosis in fallow deer (Dama dama) naturally infected with giant liver fluke (Fascioloides magna). BMC Vet. Res. 2013, 9, 1-6. [CrossRef]
- Šimonji, K.; Konjević, D.; Bujanić, M.; Rubić, I.; Farkaš, V.; Beletić, A.; Grbavac, L.; Kuleš, J. Liver proteome alterations in red deer (Cervus elaphus) infected by the giant liver fluke Fascioloides magna. Pathogens 2022, 11, 1503. [CrossRef]
- Sommer, M.F.; Drdlicek, J.; Müller, M.; Thelemann, A.; Just, F.T. Fascioloides magna and other liver parasites in cloven-hoofed game from northeastern Bavaria, Germany: occurrence and pathological findings with special emphasis on red deer (Cervus elaphus). Eur. J. Wildl. Res. 2022, 68, 73. [CrossRef]
- Pankrác, J.; Novobilský, A.; Rondelaud, D.; Leontovyč, R.; Syrovátka, V.; Rajský, D.; Horák, P.; Kašný, M. Effect of Fascioloides magna (Digenea) on fecundity, shell height, and survival rate of Pseudosuccinea columella (Lymnaeidae). Parasit. Res. 2016, 115, 3119-3125. [CrossRef]
- Rondelaud, D.; Novobilský, A.; Höglund, J.; Kašný, M.; Pankrác, J.; Vignoles, P.; Dreyfuss, G. Growth rate of the intermediate snail host Galba truncatula influences redial development of the trematode Fascioloides magna. J. Helmint. 2014, 88, 427-433. [CrossRef]
- Lockyer, A.E.; Jones, C.S.; Noble, L.R.; Rollinson, D. Trematodes and snails: an intimate association. Can. J. Zool. 2004, 82, 251-269. [CrossRef]
- Sattmann, H.; Hörweg, C.; Gaub, L.; Feix, A.S.; Haider, M.; Walochnik, J.; Rabitsch, W.; Prosl, H. Wherefrom and whereabouts of an alien: the American liver fluke Fascioloides magna in Austria: an overview. Wien. Klin. Wochenschr. 2014, 126, 23. [CrossRef]
- Farrell, M.J.; Park, A.W.; Cressler, C.E.; Dallas, T.; Huang, S.; Mideo, N.; Morales-Castilla, I.; Davies, T. J.; Stephens, P. The ghost of hosts past: impacts of host extinction on parasite specificity. Phil. Trans. Roy. Soc. B 2021, 376, 20200351. [CrossRef]
- Rogers, R.R.; Rogers, K.A.C.; Bagley, B.C.; Goodin, J.J.; Hartman, J.H.; Thole, J.T.; Zatoń, M. Pushing the record of trematode parasitism of bivalves upstream and back to the Cretaceous. Geology 2018, 46, 431-434. [CrossRef]
- Barrios-de Pedro, S.; Osuna, A.; Buscalioni, Á. D. Helminth eggs from early cretaceous faeces. Sci. Rep. UK 2020, 10, 18747. [CrossRef]
- Poinar, G.; Boucot, A.J. Evidence of intestinal parasites of dinosaurs. Parasitology 2006, 133, 245-249. [CrossRef]
- Kaiho, K.; Oshima, N.; Adachi, K.; Adachi, Y.; Mizukami, T.; Fujibayashi, M.; Saito, R. Global climate change driven by soot at the K-Pg boundary as the cause of the mass extinction. Sci. Rep. UK 2016, 6, 28427. [CrossRef]
- Chiarenza, A.A.; Farnsworth, A.; Mannion, P.D.; Lunt, D.J.; Valdes, P.J.;; Morgan, J.V.; Allison, P.A. Asteroid impact, not volcanism, caused the end-Cretaceous dinosaur extinction. Proc. National Acad. of Sci. 2020, 117, 17084-17093. [CrossRef]
- de Moya, R.S.; Allen, J.M.; Sweet, A.D.; Walden, K.K.; Palma, R.L.; Smith, V.S.; Cameron, S.L.; Valim, M.P.; Galloway, T.D.; Weckstein, J.D.; Johnson, K.P. Extensive host-switching of avian feather lice following the Cretaceous-Paleogene mass extinction event. Commun. Biol. 2019, 2, 445. [CrossRef]
- Renne, P.R.; Sprain, C.J.; Richards, M.A.; Self, S.; Vanderkluysen, L.; Pande, K. State shift in Deccan volcanism at the Cretaceous-Paleogene boundary, possibly induced by impact. Science 2015, 350, 76-78. [CrossRef]
- Sprain, C.J.; Renne, P.R.; Vanderkluysen, L.; Pande, K.; Self, S.; Mittal, T. The eruptive tempo of Deccan volcanism in relation to the Cretaceous-Paleogene boundary. Science, 2019, 363, 866-870. [CrossRef]
- Hull, P.M.; Bornemann, A.; Penman, D.E.; Henehan, M.J.; Norris, R.D.; Wilson, P.A.; Blum, P.; Alegret, L.; Batenburg, S.J.; Zachos, J. C. +26 authors. On impact and volcanism across the Cretaceous-Paleogene boundary. Science 2020, 367, 266-272. [CrossRef]
- Turner, S. K. Constraints on the onset duration of the Paleocene–Eocene Thermal Maximum. Philos. T. Roy. Soc. A 2018, 376, 20170082. [CrossRef]
- Mas-Coma, S.; Valero, M.A.; Bargues, M.D. Fasciola, lymnaeids and human fascioliasis, with a global overview on disease transmission, epidemiology, evolutionary genetics, molecular epidemiology and control. Adv. Parasit. 2009, 69, 41-146. [CrossRef]
- Choi, Y.J.; Fontenla, S.; Fischer, P.U.; Le, T.H.; Costábile, A.; Blair, D.; Brindley P.J.; Tort, J. F.; Cabada, M. M.; Mitreva, M. Adaptive radiation of the flukes of the family Fasciolidae inferred from genome-wide comparisons of key species. Molec. Biol. Evol. 2020, 37, 84-99. [CrossRef]
- Correa, A.C.; Escobar, J.S. Durand, P.; Renaud, F.; David, P.; Jarne, P.; Pointier J.-P.; Hurtrez-Boussès, S. Bridging gaps in the molecular phylogeny of the Lymnaeidae (Gastropoda: Pulmonata), vectors of fascioliasis. BMC Evol. Biol. 2010, 10, 1-12. [CrossRef]
- Liu, A.G.; Seiffert, E.R.; Simons, E.L. Stable isotope evidence for an amphibious phase in early proboscidean evolution. Proc. Natl. Acad. Sci. U.S.A. 2008, 105, 5786-5791. [CrossRef]
- Los Huertos, M. Coevolution of biota, geology, and climate. In Ecology and Management of Inland Waters: A Californian Perspective with Global Applications. Elsevier: Amsterdam, Netherlands, 2020. [CrossRef]
- Lounnas, M.; Correa, A.C.; Vázquez, A.A.; Dia, A.; Escobar, J.S.; Nicot, A.; Arenas, J.; Ayaqui, R.; Dubois, M.P.; Gimenez, T.; Gutierrez, A.; Gonzalez-Ramirez, C.; Noya, O.; Prepelitchi., L.; Uribe, N.; Wisnivesky-Colli, C.; Yong, M.; David, P.; Loker, E.S.; Jarne, P.; Pointier, J.P.; Hurtrez-Boussès, S. Self-fertilization, long-distance flash invasion and biogeography shape the population structure of Pseudosuccinea columella at the worldwide scale. Molec. Ecol. 2017, 26, 887-903. [CrossRef]
- Neubauer, T.A.; Georgopoulou, E. Extinction risk is linked to lifestyle in freshwater gastropods. Divers. Distrib. 2021, 27, 2357-2368. [CrossRef]
- Palasio, R.G.S.; Chiaravalloti-Neto, F.; Tuan, R. Distribution of genetic diversity of neotropical Biomphalaria (Preston 1910) (Basommatophora: Planorbidae) intermediate hosts for schistosomiasis in Southeast Brazil. Front. Trop. Dis. 2023, 4, 1143186. [CrossRef]
- Yu, T.; Neubauer, T.A.; Jochum, A. First freshwater gastropod preserved in amber suggests long-distance dispersal during the Cretaceous Period. Geological Magazine, 2021, 158(7), 1327-1334. [CrossRef]
- Juhász, A.; Majoros, G. A proposed ectochory of Galba truncatula snails between wallow sites enhances transmission of Fascioloides magna at Gemenc, in Hungary. Int. J. Parasitol. Parasites Wildl. 2023, 21, 2019-223. [CrossRef]
- Hurtrez-Boussès, S.; Hurtrez, J.E.; Turpin, H.; Durand, C.; Durand, P.; De Meeüs, T.; Meunier, C.; Renaud, F. Hydrographic network structure and population genetic differentiation in a vector of fasciolosis, Galba truncatula. Infect. Genet. Evol. 2010, 10(2), 178-183. [CrossRef]
- McCabe, D.J. Rivers and streams: life in flowing water. Nature Education Knowledge. 2010, 3(10), 19.
- Sturm, R. Aquatic molluscs in high mountain lakes of the Eastern Alps (Austria): Species-environment relationships and specific colonization behaviour. Chin. J. Oceanol. Limnol. 2012, 30(1), 59. [CrossRef]
- Bargues, M.D.; Artigas, P.; Khoubbane, M.; Ortiz, P.; Naquira, C.; Mas-Coma, S. Molecular characterisation of Galba truncatula, Lymnaea neotropica and L. schirazensis from Cajamarca, Peru and their potential role in transmission of human and animal fascioliasis. Parasites Vectors, 2012, 5, 1-16. [CrossRef]
- Alba, A.; Vázquez, A.A.; Sánchez, J.; Lounnas, M.; Pointier, J.P.; Hurtrez-Boussès, S.; Gourbal, B. Patterns of distribution, population genetics and ecological requirements of field-occurring resistant and susceptible Pseudosuccinea columella snails to Fasciola hepatica in Cuba. Sci. Rep. 2019, 9(1), 14359. [CrossRef]
- Ngcamphalala, P.I.; Malatji, M.P.; Mukaratirwa, S. Geography and ecology of invasive Pseudosuccinea columella (Gastropoda: Lymnaeidae) and implications in the transmission of Fasciola species (Digenea: Fasciolidae)–a review. J. Helminthol. 2022, 96, e1. [CrossRef]
- Grabner, D.S.; Mohamed, F.A.; Nachev, M.; Meabed, E.M.; Sabry, A.H.A.; Sures, B. Invasion biology meets parasitology: a case study of parasite spill-back with Egyptian Fasciola gigantica in the invasive snail Pseudosuccinea columella. PloS One, 2014, 9(2), e88537. [CrossRef]
- Saarinen, J. The palaeontology of browsing and grazing. In The Ecology of Browsing and Grazing II, 1st ed.; Gordon, I. J.; Prins, H. H. T, Eds.; Springer Nature: Cham, Switzerland AG, 2019. [CrossRef]
- Gheerbrant, E. Paleocene emergence of elephant relatives and the rapid radiation of African ungulates. Proc. Natl. Acad. Sci. U.S.A. 2009, 106(26), 10717-10721. [CrossRef]
- Baleka, S.; Varela, L.; Tambusso, P.S.; Paijmans, J.L.; Mothé, D.; Stafford, T.W.; Farina, R.A.; Hofreiter, M. Revisiting proboscidean phylogeny and evolution through total evidence and palaeogenetic analyses including Notiomastodon ancient DNA. iScience, 2022, 25(1), 103559. [CrossRef]
- Tabuce, R.; Sarr, R.;, Adnet, S.; Lebrun, R.; Lihoreau, F.; Martin, J.E.; Sambou, B.; Thiam, M.; Hautier, L. Filling a gap in the proboscidean fossil record: a new genus from the Lutetian of Senegal. J. Paleontol. 2020, 94(3), 580-588. [CrossRef]
- Cantalapiedra, J.L.; Sanisidro, O.; Zhang, H.; Alberdi, M.T.; Prado, J.L.; Blanco, F.; Saarinen, J. The rise and fall of proboscidean ecological diversity. Nat. Ecol. Evol. 2021, 5(9), 1266-1272. [CrossRef]
- Le, T.H.; Pham, K.L.T.; Doan, H.T.T.; Le, T.X.; Nguyen, K.T.; Lawton, S.P. Description and phylogenetic analyses of ribosomal transcription units from species of Fasciolidae (Platyhelminthes: Digenea). J. Helminthol. 2020, 94, e136. [CrossRef]
- Heneberg, P. Phylogenetic data suggest the reclassification of Fasciola jacksoni (Digenea: Fasciolidae) as Fascioloides jacksoni comb. nov. Parasitol. Res. 2013, 112, 1679-1689. [CrossRef]
- Rajapakse, R.P.V.J.; Pham, K.L.T.; Karunathilake, K.K.; Lawton, S.P.; Le, T.H. Characterization and phylogenetic properties of the complete mitochondrial genome of Fascioloides jacksoni (syn. Fasciola jacksoni) support the suggested intergeneric change from Fasciola to Fascioloides (Platyhelminthes: Trematoda: Plagiorchiida). Infect. Genet. Evol. 2020, 82, 104281. [CrossRef]
- Reblánová, M.;Špakulová, M.; Orosová, M.; Bazsalovicsová, E.; Rajský, D. A description of karyotype of the giant liver fluke Fascioloides magna (Trematoda, Platyhelminthes) and a review of Fasciolidae cytogenetics. Helminthologia, 2010, 47, 69-75. [CrossRef]
- Bargues, M.D.; Halajian, A.; Artigas, P.; Luus-Powell, W.J.; Valero, M.A.; Mas-Coma, S. Paleobiogeographical origins of Fasciola hepatica and F. gigantica in light of new DNA sequence characteristics of F. nyanzae from hippopotamus. Front. Vet. Sci. 2022, 9, 990872. [CrossRef]
- Mennecart, B.; Dziomber, L.; Aiglstorfer, M.; Bibi, F.; DeMiguel, D.; Fujita, M.; Kubo, M.O.; Laurens, F.; Meng, J.; Métais, G.; Müller, B.; Ríos, M.; Rössner, G. E.; Sánchez, I. M.; Schulz, G.; Wang, S.; Costeur, L. Ruminant inner ear shape records 35 million years of neutral evolution. Nat. Commun. 2022, 13(1), 7222. [CrossRef]
- Codron, D. Evolution of large mammal herbivores in savannas. In Savanna Woody Plants and Large Herbivores, 1st ed.; Scogings, P. F.; Sankaran, M., Eds.; John Wiley & Sons Ltd., Chichester, West Sussex, UK, 2019. [CrossRef]
- Sheil, D. Dangerous giants?—Large herbivores, forest feedbacks and climate tipping points. Russ. j. ecosyst. ecol. 2020, 5(3), 1-33. [CrossRef]
- Ao, H.; Rohling, E.J.; Zhang, R.; Roberts, A.P.; Holbourn, A.E.; Ladant, J.B.; Dupont-Nivet, G.; Kuhnt, W.; Zhang, P.; Wu, F.; Dekkers, M.J.; Liu, Q.; Liu, Z.; Xu, Y.; Poulsen, C.J.; Licht, A.; Sun, Q.; Chiang, J.C.H.; Liu, X.; Wu, G.; Ma, C.; Zhou, W.; Jin, Z.; Li, X.; Li, X.; Peng, X.; Qiang, X.; An, Z. Global warming-induced Asian hydrological climate transition across the Miocene–Pliocene boundary. Nat. Commun. 2021, 12(1), 6935. [CrossRef]
- Pérez-Crespo, V.A.; Prado, J.L.; Alberdi, M.T.; Arroyo-Cabrales, J.; Johnson, E. Feeding ecology of the gomphotheres (Proboscidea, Gomphotheriidae) of America. Quat. Sci. Rev. 2020, 229, 106126. [CrossRef]
- Ameen, M.; Khan, A.M.; Ahmad, R.M.; Ijaz, M.U.; Imran, M. Tooth marker of ecological abnormality: The interpretation of stress in extinct mega herbivores (proboscideans) of the Siwaliks of Pakistan. Ecol. Evol. 2022, 12, e9432. [CrossRef]
- Gilbert, C.; Ropiquet, A.; Hassanin, A. Mitochondrial and nuclear phylogenies of Cervidae (Mammalia, Ruminantia): systematics, morphology, and biogeography. Mol. Phylogenet. Evol. 2006, 40(1), 101-117. [CrossRef]
- Polziehn, R.O.; Strobeck, C. Phylogeny of wapiti, red deer, sika deer, and other North American cervids as determined from mitochondrial DNA. Mol. Phylogenet. Evol. 1998, 10(2), 249-258. [CrossRef]
- Pybus, M.J. Liver flukes. In Parasitic Diseases of Wild Mammals; Samuel, W.M., Pybus, M.J., Kocan, A.A., Eds.; Iowa State Press: Ames, IA, USA, 2021; pp. 121–149. [CrossRef]
- Parker, A.; David, A.A. Genetic characterization of the giant liver fluke, Fascioloides magna (Platyhelminthes: Fasciolidae) from the Adirondack Region of Northern New York. Acta Parasitol. 2021, 66, 259-263. [CrossRef]
- Králová-Hromadová, I.; Bazsalovicsová, E.; Štefka, J.; Špakulová, M.; Vávrová, S.; Szemes, T.; Tkach, V.; Trudgett, A.; Pybus, M. Multiple origins of European populations of the giant liver fluke Fascioloides magna (Trematoda: Fasciolidae), a liver parasite of ruminants. Int. J. Parasitol. 2011, 41(3-4), 373-383. [CrossRef]
- Engelstädter, J.; Fortuna, N. Z. The dynamics of preferential host switching: Host phylogeny as a key predictor of parasite distribution. Evolution, 2019, 73(7), 1330-1340. [CrossRef]
- Demiaszkiewicz, A.W.; Kuligowska, I.; Pyziel, A.M.; Lachowicz, J.; Kowalczyk, R. Extension of occurrence area of the American fluke Fascioloides magna in south-western Poland. Ann. Parasitol. 2015, 61(2), 93-96.
- Rajský, D.; Patus, A.; Bukovjan, K. Prvýnález Fascioloides magna Bassi, 1875 na Slovensku (in Slovakian). Slov. Vet. Čas. 1994, 19, 29–30.
- Majoros, G.; Sztojkov, V. Appearance of the large American liver fluke Fascioloides magna (Bassi, 1875) (Trematoda: Fasciolata) in Hungary. Parasitologia Hungarica 1994, 27, 27-38.
- Houszka, M.; Piekarska, J.; Podkowik, M.; Gorczykowski, M.; Bania, J. Morphology and molecular study of Fascioloides magna–a growing threat to cervids (Cervidae) in Poland. J. Vet. Res. 2016, 60(4), 435-439. [CrossRef]
- Juhásová, L.; Králová-Hromadová, I.; Zeleňáková, M.; Blišťan, P.; Bazsalovicsová, E. Transmission risk assessment of invasive fluke Fascioloides magna using GIS-modelling and multicriteria analysis methods. Helminthologia, 2017, 54(2), 119-131. [CrossRef]
- Marinculić, A.; Džakula, N.; Janicki, Z.; Hardy, Z.; Lučinger, S.; Živičnjak, T. Appearance of American liver fluke (Fascioloides magna, Bassi, 1875) in Croatia. Vet. Arh. 2002, 72(6), 319-325.
- Slavica, A.; Florijančić, T.; Janicki, Z.; Konjević, D.; Severin, K.; Marinculić, A.; Pintur, K. Treatment of fascioloidosis (Fascioloides magna, Bassi, 1875) in free ranging and captive red deer (Cervus elaphus L.) at eastern Croatia. Vet. Arh. 2006, 76(Suppl.), 9-18.
- Konjević, D.; Janicki, Z.; Calmels, P.; Jan, D.S.; Marinculić, A.; Šimunović, M.; Pavlak, M.; Krapinec, K.; Poljak, Z. Evaluation of factors affecting the efficacy of treatment against Fascioloides magna in wild red deer population. Vet Ital, 54(1), 2018, 33-39. [CrossRef]
- Trailović, S.M.; Marinković, D.; & Kulišić, Z. Diagnosis and therapy of liver fluke (Fascioloides magna) infection in fallow deer (Dama dama) in Serbia. J. Wildl. Dis. 2016, 52(2), 319-326. [CrossRef]
- Mirčeta, J.; Pelić, M.; Božić, B.; Petrović, J.; Urošević, M.; Stankov, B.; Bugarski, D. (2018). Prevalence of the giant liver fluke (Fascioloides magna, Bassi, 1875) in red deer (Cervus elaphus) in the region of floodplain forests of northern Serbia. Arch. Vet. Med. 2018, 11(1), 17-26. [CrossRef]
- Plötz, C.; Rehbein, S.; Bamler, H.; Reindl, H.; Pfister, K.; Scheuerle, M. C. Fascioloides magna–epizootiology in a deer farm in Germany. Berl. Munch. Tierärzt. Wochensch. 2015, 128(5/6), 111-182.
- Konjević, D.; Bujanić, M.; Beck, A.; Beck, R.; Martinković, F.; Janicki, Z. First record of chronic Fascioloides magna infection in roe deer (Capreolus capreolus). Int. J. Parasitol. Parasites Wildl. 2021, 15, 173-176. [CrossRef]
- Halász, T.; Tari, T.; Nagy, E;, Nagy, G; Csivincsik, Á. (2023). Hatchability of Fascioloides magna eggs in cervids. Pathogens, 2023, 12(5), 741. [CrossRef]
- Králová-Hromadová, I.; Špakulová, M.; Horáčková, E.; Turčeková, L.; Novobilský, A.; Beck, R.; Koudela, B.; Marinculic, A.; Rajský, D.; Pybus, M. Sequence analysis of ribosomal and mitochondrial genes of the giant liver fluke Fascioloides magna (Trematoda: Fasciolidae): intraspecific variation and differentiation from Fasciola hepatica. J. Parasitol. 2008, 94(1), 58-67. [CrossRef]
- Leontovyč, R.; Košťáková, M.; Siegelová, V.; Melounová, K.; Pankrác, J.; Vrbová, K.; Horák, P.; Kašný, M. (2014). Highland cattle and Radix labiata, the hosts of Fascioloides magna. BMC Vet. Res, 2014, 10, 1-8. [CrossRef]
- Konjević, D.; Bujanić, M.; Erman, V.; Kurilj, A.G.; Živičnjak, T.; Severin, K.; Tomić, S.; Martinković, F. New data on wild boar (Sus scrofa L.) a dead-end host for large American liver fluke (Fascioloides magna). Helminthologia, 2017, 54(1), 77-80. [CrossRef]
- Demiaszkiewicz, A.W.; Kowalczyk, R.; Filip, K.J.; Pyziel, A. M. Fascioloides magna: a parasite of roe deer in Bory Zielonogórskie (in Polish). Med. Weter. 2018, 74(4), 257-260. [CrossRef]
- Filip-Hutsch, K.; Pyziel-Serafin, A.M.; Hutsch, T.; Bulak, K.; Czopowicz, M.; Merta, D.; Kobielski, J.; Demiaszkiewicz, A.W. The occurrence of Fascioloides magna (Bassi, 1875) in the wild cervid population in the Lower Silesian Wilderness–epidemiological and pathological aspects. J. Vet. Res. 66(3), 381-387. [CrossRef]
- Shury, T.K.; Pybus, M.J.; Nation, N.; Cool, N.L.; & Rettie, W.J. Fascioloides magna in moose (Alces alces) from Elk Island National Park, Alberta. Vet. Pathol. 2019, 56(3), 476-485. [CrossRef]
- Živković, D.; John, S.; Verin, M.; Stephan, W.; Tellier, A. Neutral genomic signatures of host-parasite coevolution. BMC Evol. Biol. 2019, 19(1), 230. [CrossRef]
- Buckingham, L.J.; Ashby, B. Coevolutionary theory of hosts and parasites. J. Evol. Biol. 2022, 35, 205–224.
- Saadi, A.J.; Davison, A.; Wade, C.M. Molecular phylogeny of freshwater snails and limpets (Panpulmonata: Hygrophila). Zool. J. Linn. Soc. 2020, 190(2), 518-531. [CrossRef]
- Hörweg, C.; Prosl, H.; Wille-Piazzai, W.; Joachim, A.; Sattmann, H. Prevalence of Fascioloides magna in Galba truncatula in the Danube backwater area east of Vienna, Austria. Wien Tierarztl Monat, 2011, 98, 261-7.
- Haider, M.; Hörweg, C.; Liesinger, K.; Sattmann, H.; & Walochnik, J. Recovery of Fascioloides magna (Digenea) population in spite of treatment programme? Screening of Galba truncatula (Gastropoda, Lymnaeidae) from Lower Austria. Vet. Parasitol. 2012, 187(3-4), 445-451. [CrossRef]
- Schniebs, K.; Peter, G.; Vinarski, M.V.; Hundsdoerfer, A.K. Intraspecific morphological and genetic variability in the European freshwater snail Radix labiata (Rossmaessler, 1835)(Gastropoda: Basommatophora: Lymnaeidae). Contrib. Zool. 2013, 82(1), 55-68. [CrossRef]
- Salgado, S.Q.; Soriano, J.L.; Glöer, P. First report of Radix labiata (Rossmässler, 1835)(Gastropoda: Lymnaeidae) in Aragon (NE Spain). J. Conchol, 2016, 40, 657-678.
- Garbar, А.; Komіushіn, А.V.. Karyotypes of European species of Radix (Gastropoda: Pulmonata: Lymnaeidae) and their relevance to species distinction in the genus. Malacologia, 2003, 45(1), 141-148.
- Alda, P.; Lounnas, M.; Vázquez, A.A.; Ayaqui, R.; Calvopiña, M.; Celi-Erazo, M.; Dillon, R.T.; Jr, Jarne, P.; Loker, E.S.; Muñiz Pareja, F.C.; Muzzio-Aroca, J.; Nárvaez, A.O.; Noya, O.; Robles, L.M.; Rodríguez-Hidalgo, R.; Uribe, N.; David, P.; Pointier, J.P.; Hurtrez-Boussès, S. A new multiplex PCR assay to distinguish among three cryptic Galba species, intermediate hosts of Fasciola hepatica. Vet. Parasitol. 2018, 251, 101-105. [CrossRef]
- Bargues, M.D.; Mas-Coma, S. Phylogenetic analysis of lymnaeid snails based on 18S rDNA sequences. Mol. Biol. Evol. 1997, 14(5), 569-577. [CrossRef]
- Davis, C.N.; Tyson, F.; Cutress, D.; Davies, E.;, Jones, D.L.; Brophy, P.M.; Prescott, A.; Rose, M.T.; Williams, M.; Williams, H.W.; Jones, R.A. Rapid detection of Galba truncatula in water sources on pasture-land using loop-mediated isothermal amplification for control of trematode infections. Parasites Vectors 2020, 13, 1-11. [CrossRef]
- Caron, Y.; Righi, S.; Lempereur, L.; Saegerman, C.; Losson, B. An optimized DNA extraction and multiplex PCR for the detection of Fasciola sp. in lymnaeid snails. Vet. Parasitol. 2011, 178(1-2), 93-99. [CrossRef]
- Caron, Y.; Lasri, S.; Losson, B. Fasciola hepatica: an assessment on the vectorial capacity of Radix labiata and R. balthica commonly found in Belgium. Vet. Parasitol. 2007, 149(1-2), 95-103. [CrossRef]
- Novobilský, A.; Kašný, M.; Beran, L.; Rondelaud, D.; Höglund, J. Lymnaea palustris and Lymnaea fuscus are potential but uncommon intermediate hosts of Fasciola hepatica in Sweden. Parasites Vectors 2013, 6, 1-10. [CrossRef]
- Kasny, M.; Beran, L.; Siegelova, V.; Siegel, T.; Leontovyc, R.; Berankova, K.; Pankrac, J.; Kostakova M.; Horak, P. (2012). Geographical distribution of the giant liver fluke (Fascioloides magna) in the Czech Republic and potential risk of its further spread. Vet. Med. (Praha) 2012, 57(2), 101-109. [CrossRef]
- Sindou, P.; Rondelaud, D.; Barthe, D. Fasciola hepatica L.: comparative studies of tissue lesions in seven lymnaeid species submitted to individual monomiracidial exposures at hatching. Bull. de la Soc. Zool. de France 1990, 115(4), 331-340.
- Georgieva, K.; Georgieva, L.; Mizinska-Boevska, Y.; Stoitsova, S.R. Study of surface carbohydrates in Galba truncatula tissues before and after infection with Fasciola hepatica. Mem Inst Oswaldo Cruz, 2016, 111, 475-483. [CrossRef]
- Krull, W.H. The intermediate hosts of Fasciola hepatica and Fascioloides magna in the United States. North American Veterinarian 1934, 15(12), 13-17.
- Correa, A.C.; De Meeûs, T.; Dreyfuss, G.; Rondelaud, D.; Hurtrez-Boussès, S. Galba truncatula and Fasciola hepatica: Genetic costructures and interactions with intermediate host dispersal. Infect. Genet. Evol. 2017, 55, 186-194. [CrossRef]
- Chapuis, E.; Trouve, S.; Facon, B.; Degen, L.; Goudet, J. High quantitative and no molecular differentiation of a freshwater snail (Galba truncatula) between temporary and permanent water habitats. Mol. Ecol. 2007, 16(16), 3484-3496. [CrossRef]
- Alda, P.; Lounnas, M.; Vázquez, A.A., Ayaqui, R.; Calvopiña, M.; Celi-Erazo, M.; Dillon R.T.; Jr, González Ramírez, L.C.; Loker, E.S.; Muzzio-Aroca, J.; Nárvaez, A.O.; Noya, O.; Pereira, A.E.; Robles, L.M.; Rodríguez-Hidalgo, R.; Uribe, N.; David, P.; Jarne, P.; Pointier, J.P.; Hurtrez-Boussès, S. Systematics and geographical distribution of Galba species, a group of cryptic and worldwide freshwater snails. Mol. Phylogenetics Evolut. 2021, 157, 107035. [CrossRef]
- Ebbs, E.T.; Loker, E.S.; Brant, S.V. Phylogeography and genetics of the globally invasive snail Physa acuta Draparnaud 1805, and its potential to serve as an intermediate host to larval digenetic trematodes. BMC Evol. Biol. 2018, 18(1), 1-17. [CrossRef]
- Kengne-Fokam, A.C.; Nana-Djeunga, H.C.; Djuikwo-Teukeng, F.F.; Njiokou, F. Analysis of mating system, fecundity, hatching and survival rates in two Schistosoma mansoni intermediate hosts (Biomphalaria pfeifferi and Biomphalaria camerunensis) in Cameroon. Parasites Vectors 2016, 9, 1-9. [CrossRef]
- Burgarella, C.; Gayral, P.; Ballenghien, M.; Bernard, A.; David, P.; Jarne, P.; Correa, A.; Hurtrez-Bousses, S.; Escobar, J.; Galtier, N.; Glémin, S. Molecular evolution of freshwater snails with contrasting mating systems. Mol. Biol. Evol. 2015, 32(9), 2403-2416. [CrossRef]
- Vázquez, A.A.; Alba, A.; Alda, P.; Vittecoq, M.; Hurtrez-Boussès, S. On the arrival of fasciolosis in the Americas. Trends Parasitol. 2022, 38(3), 195-204. [CrossRef]
- Campbell, G.;, Noble, L.R.; Rollinson, D.; Southgate, V.R.; Webster, J.P.; Jones, C.S. Low genetic diversity in a snail intermediate host (Biomphalaria pfeifferi Krass, 1848) and schistosomiasis transmission in the Senegal River Basin. Mol. Ecol. 2010, 19(2), 241-256. [CrossRef]
- Soldánová, M.; Selbach, C.; Sures, B.; Kostadinova, A.; Pérez-del-Olmo, A. Larval trematode communities in Radix auricularia and Lymnaea stagnalis in a reservoir system of the Ruhr River. Parasites Vectors 2010, 3, 1-13. [CrossRef]
- Selbach, C.; Soldánová, M.; Feld, C.K.; Kostadinova, A.; Sures, B. Hidden parasite diversity in a European freshwater system. Sci. Rep. 2020, 10(1), 2694. [CrossRef]
- Roldán, C.; Begovoeva, M.; López-Olvera. J.R.; Velarde, R.; Cabezón, Ó.; Molinar Min, A.R.; Pizzato, F.; Pasquetti, M.; Fernández Aguilar, X.; Mentaberre, G.; Serrano, E.;, Puig Ribas, M.; Espunyes, J.; Castillo-Contreras, R.; Estruch, J.; Rossi, L. Endemic occurrence of Fasciola hepatica in an alpine ecosystem, Pyrenees, Northeastern Spain. Transbound.Emerg. Dis. 2021, 68(4), 2589-2594. [CrossRef]
- Skowrońska-Ochmann, K.; Cuber, P.; Lewin, I. The first record and occurrence of Stagnicola turricula (Held, 1836)(Gastropoda: Pulmonata: Lymnaeidae) in Upper Silesia (Southern Poland) in relation to different environmental factors. Zoo. Anz. J. Comp. Zool. 2012, 251(4), 357-363. [CrossRef]
- Outa, J.O.; Sattmann, H.; Köhsler, M.; Walochnik, J.; Jirsa, F. Diversity of digenean trematode larvae in snails from Lake Victoria, Kenya: First reports and bioindicative aspects. Acta Trop. 2020, 206, 105437. [CrossRef]
- Sokolow, S.H.; Jones, I.J.; Jocque, M.; La, D.; Cords, O.; Knight, A.; Lund, A.; Wood, C.L.; Lafferty, K.D.; Hoover, C.M.; Collender, P.A.; Remais, J V.; Lopez-Carr, D.; Fisk, J.; Kuris, A.M.; De Leo, G.A. Nearly 400 million people are at higher risk of schistosomiasis because dams block the migration of snail-eating river prawns. Philos. Trans. R. Soc. B, Biol. Sci. 2017, 372(1722), 20160127. [CrossRef]
- Barakat, R.M. Epidemiology of schistosomiasis in Egypt: travel through time. J. Adv. Res. 2013, 4(5), 425-432. [CrossRef]
- Hrivnák, R.; Medvecká, J;, Baláži, P.; Bubíková, K.; Oťaheľová, H.; Svitok, M. Alien aquatic plants in Slovakia over 130 years: historical overview, current distribution and future perspectives. NeoBiota 2019, 49, 37–56. [CrossRef]
- Garcia-Murillo, P. Hydrocharis laevigata in Europe. Plants, 2023, 12(4), 701. [CrossRef]
- Abe, E.M.; Guan, W.; Guo, Y.H.; Kassegne, K., Qin, Z.Q.; Xu, J.; CHen, J.-H.; Ekpo, U.F.; Li, S.-Z.; Zhou, X.N. Differentiating snail intermediate hosts of Schistosoma spp. using molecular approaches: fundamental to successful integrated control mechanism in Africa. Infect. Dis. Poverty, 2018, 7(02), 6-18. [CrossRef]
- Plummer, M.L. Impact of invasive water hyacinth (Eichhornia crassipes) on snail hosts of schistosomiasis in Lake Victoria, East Africa. EcoHealth, 2005, 2, 81-86. [CrossRef]
- Hussner, A.. Alien aquatic plant species in European countries. Weed Res. 2012, 52(4), 297-306. [CrossRef]
- Preston, D L.; Crone, E.R.; Miller-ter Kuile, A.; Lewis, C.D.; Sauer, E.L.; Trovillion, D.C. Non-native freshwater snails: a global synthesis of invasion status, mechanisms of introduction, and interactions with natural enemies. Freshw. Biol. 2022, 67(2), 227-239. [CrossRef]
- Vignoles, P., Dreyfuss, G.; Rondelaud, D. Consequences of invasion by Pseudosuccinea columella on the dynamics of native lymnaeids living on the acid soils of central France. Molluscan Res. 2018, 38(4), 287-295.
- Schols, R.; Carolus, H.; Hammoud, C.; Muzarabani, K.C;, Barson, M.; Huyse, T. Invasive snails, parasite spillback, and potential parasite spillover drive parasitic diseases of Hippopotamus amphibius in artificial lakes of Zimbabwe. BMC Biol. 2021, 19, 1-21. [CrossRef]
- Pointier, J.P. Coustau, C. Rondelaud, D. Theron, A. Pseudosuccinea columella (Say 1817) (Gastropoda, Lymnaeidae), snail host of Fasciola hepatica: first record for France in the wild. Parasitol. Res. 2007, 101(5), 1389-1392. [CrossRef]
- Varga, A.; Lőkkös, A. (2021) Mollusc fauna of the Lake Hévíz (in Hungarian). .Soosiana 2021, 35, 3-18.
- Njiokou, F.; Mouafo, J.B.; Teukeng, F.; Njine, T.; Ekobo, A.S.; Jarne, P. The influence of self-fertilization and pairing on life-history traits in the freshwater snail Bulinus forskalii (Gastropoda, Planorbidae). Acta Trop. 2000, 76(2), 159-167. [CrossRef]
- Dreyfuss, G.; Vignoles, P.; Rondelaud, D. Relationships between the distribution of Galba truncatula (Gastropoda: Lymnaeidae) climatic conditions and the altitude of municipalities in Haute Vienne (France). In Ann. Limnol. 2018, 54, 19-28). [CrossRef]
- Crone, E.R.; Sauer, E.L.; Preston, D.L.. Non-native fish facilitate non-native snails and alter food web structure in experimental pond communities. Funct. Ecol. 2023, 37(4), 947-958. [CrossRef]
- Beesley, N.J.; Williams, D.J.; Paterson, S.; Hodgkinson, J. Fasciola hepatica demonstrates high levels of genetic diversity, a lack of population structure and high gene flow: Possible implications for drug resistance. Int. J. Parasitol. 2017, 47, 11–20. [CrossRef]
- Turner, W C.; Kamath, P.L.; Van Heerden, H.; Huang, Y.H.; Barandongo, Z.R.; Bruce, S.A.; Kausrud, K. The roles of environmental variation and parasite survival in virulence–transmission relationships. Royal Soc. Open Sci. 2021, 8(6), 210088. [CrossRef]
- Morran, L.T;, Schmidt, O.G.; Gelarden, I A.; Parrish, R.C.; Lively, C.M. Running with the Red Queen: host-parasite coevolution selects for biparental sex. Science, 2011, 333(6039), 216-218. [CrossRef]
- Arakelyan, M.; Harutyunyan, T.; Aghayan, S.A.; Carretero, M.A.. Infection of parthenogenetic lizards by blood parasites does not support the “Red Queen hypothesis” but reveals the costs of sex. Zoology, 2019, 136, 125709. [CrossRef]
- Majewska, A.A.; Sims, S.; Schneider, A.; Altizer, S.; Hall, R.J. Multiple transmission routes sustain high prevalence of a virulent parasite in a butterfly host. Proc. R. Soc. B, 2019, 286(1910), 20191630. [CrossRef]
- White, P.S.; Choi, A.; Pandey, R.; Menezes, A.; Penley, M.; Gibson, A.K.; de Roode, J.; Morran, L. Host heterogeneity mitigates virulence evolution. Biol. Lett. 2020, 16, 20190744. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



