
Submitted:
28 August 2023
Posted:
29 August 2023
You are already at the latest version
Abstract
Ribotoxin-like proteins (RL-Ps) are specific ribonucleases found in mushrooms that are able to cleave a single phosphodiester bond located in the Sarcin-Ricin Loop (SRL) of the large rRNA. The cleaved SRL interacts differently with some ribosomal proteins (P-stalk). This action blocks protein synthesis because the damaged ribosomes are unable to interact with elongation factors.
Here, the amino acid sequence of eryngitin 3 and 4, RL-Ps isolated from Pleurotus eryngii fruiting bodies, was determined to: i) obtain structural information on this specific ribonuclease family from edible mushrooms; and ii) explore the structural determinants which justify their different biological and antipathogenic activities. Indeed, eryngitin 3 (exhibited higher toxicity with respect to eryngitin 4 against tumoral cell lines and model fungi. Structurally, eryngitin 3 and 4 consist of 132 amino acids, most of them identical and exhibit a single free cysteinyl residue. The amino acidic differences between the two toxins are: i) an additional phenylalanyl residue at the N-terminus of eryngitin 3, not retrieved in eryngitin 4; and ii) an additional arginyl residue at C-terminus of eryngitin 4, not retrieved in eryngitin 3. The 3D-models of eryngitins show slight differences at the N- and C-terminal regions. In particular, the positive electrostatic surface at C-terminal of eryngitin 4 is due to the additional arginyl residue not retrieved in eryngitin 3. This additional positive charge could interfere with the binding to the SRL (substrate) or with some ribosomal proteins (P-stalk structure), during substrate recognition.
Keywords:
amino acid sequence
; green mold
; king trumpet mushroom
; MALDI-ToF
; ribotoxin-like proteins (RL-Ps)
; Sarcin-Ricin Loop
1. Introduction
Ribotoxin-like proteins (RL-Ps) are a family of specific basidiomycetes ribonucleases which catalyse endonucleolytic cleavage of large ribosomal RNA at level of a specific site located in the sarcin-ricin loop (known as SRL), leading to protein synthesis inhibition and cell death [1,2]. The specific endonucleolytic action is ascertained by Endo’s assay, through the detection of a characteristic fragment of ~450 nucleotides known as α-fragment, when these enzymes are incubated with ribosomes [3]. RL-Ps have been identified and purified from various edible mushroom fruiting bodies such as Cyclocybe aegerita (previously known as Agrocybe aegerita) [2], Boletus edulis [4], Agaricus bisporus [5] and two Pleurotus species (P. ostreatus [6] and P. eryngii [7]), most of them are of considerable commercial interest, being a source of high-quality nutrients [8].
Structurally, RL-Ps are monomeric proteins (~15-kDa), with basic pI (usually > 9.0) as displayed by computational analyses [2]. Furthermore, the residues involved in the catalytic action are two aspartyl and one histidinyl residues, which constitute the enzymatic catalytic triad [9,10]. In addition, these enzymes contain a single cysteinyl residues at the N-terminal region, implicated in the binding with metal ions (e.g.: Zn+2) and protein stability [9,11].
The cytotoxic effect of RL-Ps likely consists of an irreversible damage to the ribosome (in particular, the large subunit), which becomes unable to bind the elongation factors, with consequent arrest of protein synthesis. Indeed, when SRL is damaged, the consequent structural variations also affect the P-stalk, usually necessary to allow the correct interaction of elongation factors with ribosomes, by enhancing ribosome GTPase activity [12,13]. The structural variations due to the damage at level of SRL are confirmed by studies on different toxin families targeting SRL, such as ribosome inactivating proteins from plants (N-glycosylase enzymes) [14,15], Shiga-Like toxins from bacteria (analogues to plant N-glycosylases) [16,17] and ribotoxins, specific extracellular ribonucleases isolated from ascomycetes fungi [18,19]. These studies, highlight that the variation in the interactions between the P-stalk and these toxins is important to facilitate the recruitment of toxins on the ribosomes, modulating their toxicity [20].
Most of RL-Ps exhibit cytotoxic effects towards several human malignant cell lines, while ageritin, the prototype of RL-Ps, isolated from C. aegerita fruiting bodies, also exhibits antifungal activity [2,21,22] and cytotoxic effect towards insect Sf9 cell line [7]. In this framework, it is reasonable to consider RL-Ps involvement in fungi self-defence mechanisms, while a possible physiological role must be further investigated.
Finally, several bioinformatics approaches performed to find homologous proteins in basidiomycetes showed that the gene of these enzymes is conserved [10,23]. In particular, the RL-Ps gene retrieved for ageritin [24] and ostreatin (RL-P isolated from P. ostreatus) [6] consists of a coding sequence (CDS) for a pre-protein (pre-form), exhibiting an additional N- and C-terminal amino acid region, not retrieved in native proteins, likely involved in the sorting or controlling their toxicity in the fungi that express these toxins [24].
In this scenario, to obtain more information on the biological and antipathogenic activities as well as structural features of RL-Ps family, we decided to further analyse eryngitin 3 and 4 (~15-kDa), novel RL-Ps recently isolated from P. eryngii fruiting bodies (known as king trumpet mushroom or ‘Cardoncelli’ in Italian) [7]. Based on the significant differences found in the biological and antipathogenic activities of the two isolated proteins, we carried out an extensive structural characterization to determine the primary structure of both RL-Ps. Finally, the 3D-models of both toxins (obtained by AlphaFold prediction) were used to give a possible explanation on the biological differences found between eryngitin 3 and 4.
2. Results and discussion
2.1. Purification of eryngitin 3 and 4
Eryngitin 3 and 4 purification was performed by applying the procedure reported in Landi et al. (2022) [7]. The homogeneity of purified proteins was assessed by SDS-PAGE analysis with and without reducing agent (Figure 1a).
Aliquots of eryngitin 3 and 4 in the presence of reducing agent (lanes 1 and 2, Figure 1a) showed a single protein band with an electrophoretic migration of ~15-kDa, while without reducing agent, the presence of a main protein band of ~15-kDa and an additional faint protein band of ~30-kDa (lanes 3 and 4, Figure 1a), for both proteins, is confirmed. Considering the presence of a single free cysteinyl residue responsible for dimeric forms under denaturing conditions in ostreatin, a homologous enzyme previously isolated from P. ostreatus fruiting bodies [6], SDS-PAGE analysis was carried out without reducing agent after treatment with 4-vynylpyridine, a chemical agent able to alkylate free cysteinyl residues. In this condition, the disappearance of the ~30-kDa protein band is evident for both eryngitins (lanes 5 and 6, Figure 1a). In light of this, both eryngitins likely have at least one free cysteinyl reactive residue, as previously reported for ostreatin and ageritin, the latter, prototype of RL-Ps isolated from C. aegerita fruiting bodies [2].
2.2. Assessment of biological and antipathogenic activities of eryngitin 3 and 4
In a previous work, the ability of eryngitin 3 and 4 to release the α-fragment, hallmark of RL-Ps, and their cytotoxic effect towards both human (HUVEC) and insect (Sf9) cell lines, were ascertained [7]. These enzymes were able to release the α-fragment and exhibit low in vitro cytotoxicity against the cell lines tested. In this framework, considering their possible involvement in fungal self-defence mechanisms, a further enzymatic characterization (i.e.: in vitro IC50 determination and α-fragment release) as well as the evaluation of in vitro biological and antipathogenic activities on model cells/organisms were carried out [25].
The determination of the IC50 (concentration inhibiting 50% of protein synthesis) of eryngitin 3 and 4 by in vitro assay was evaluated using a rabbit reticulocytes lysate system. Considering that the α-fragment (Endo’s assay) release is only a qualitative result, IC50 determination is important to obtain quantitative information necessary to compare the different toxins. In particular, eryngitin 3 and 4 exhibit an IC50 of 0.3 ng/mL (20.7 pM) and 160 ng/mL (11.0 nM), respectively (Figure 2a). Thus, under the same experimental conditions, the IC50 value of eryngitin 3 is ~530-fold lower compared to that of eryngitin 4, although both toxins release the α-fragment. Moreover, the IC50 value of eryngitin 3 is 6.5-fold lower than that of ageritin (IC50= 133 pM), prototype of RL-Ps isolated from C. aegerita fruiting bodies [1] and 11.3-fold lower than that of ostreatin (IC50= 234 pM), a homologous RL-P isolated from P. ostreatus fruiting bodies [6]. Subsequently, to investigate whether eryngitin 3 and 4 had specific ribonuclease action on the SRL, we carried out the Endo’s assay on yeast (Saccharomyces cerevisiae L.) and mealworm insect (Tenebrio molitor L.) ribosomes. As shown in Figure 2b,c, when ribosomes were incubated with eryngitin 3 and 4, the characteristic α-fragment, identical to that produced by the ribotoxin α-sarcin [26], was produced. This clearly indicates that eryngitin 3 and 4 exhibit a specific ribonuclease action on ribosomes from different sources. On the other hand, this suggests that these proteins could play an insecticidal and fungicidal role, as has been suggested for ribotoxins [18,19,21,26].
In addition, to obtain more information on their cytotoxic effects against different human tumour cell lines, eryngitin 3 and 4 were tested on both HeLa and COLO 320 cell lines and the IC50 values (concentration of protein causing 50% of cell death) of eryngitin 3 and 4 are reported in Table S1. As displayed in Figure 3a, the most sensitive were HeLa cells, with IC50 values from 3.0 to 230 nM after 72 h of treatment, while COLO 320 cells have values between 420 to 1700 nM after 72 h of treatment (Figure 3b). The lowest IC50 values for both cell lines refer to eryngitin 3, which is 77-fold and 4-fold more toxic than eryngitin 4, when incubated with Hela and COLO 320 cells, respectively. In previous works, the RL-Ps cytotoxicity was attributed to their ability to trigger the apoptotic pathway [2] that may be linked to signalling through the ribotoxic stress response or to inhibition of protein synthesis. Therefore, we first wanted to see whether, after endocytosis, eryngitin 3 was able to reach the cytosol and inactivate the ribosomes. Thus, we analysed the rRNA from HeLa cells treated with eryngitin 3 for 48 h. Figure 3c shows that the RL-P displayed 28S rRNA ribonuclease activity on cell ribosomes, as indicated by the release of the α-fragment, demonstrating that eryngitin 3 was able to reach and damage the ribosomes. Second, in order to ascertain the involvement of caspase mediated apoptosis in eryngitin 3 and 4 mediated cell death, we tested the sensitivity to the pan-caspase inhibitor Z-VAD for HeLa cells, or the cleavage of chromosomal DNA into oligonucleosomal fragments for COLO 320 cells.
In particular, HeLa cells were pretreated and maintained in 100 µM Z-VAD for 48 h, and cell viability was determined in the presence of different concentrations of eryngitin 3 and 4. As shown in Figure 3a, the presence of Z-VAD increased cell viability from 11 to 54% in 3.1 M eryngitin 3-treated cells and from 29 to 64% in 3.1 M eryngitin 4-treated cells. Moreover, the typical apoptotic morphological features were also visualized microscopically when HeLa cells were incubated with eryngitin 3 or eryngitin 4 for 48 h (Figure S1). On the other hand, when COLO 320 cells were treated for 72 h with 300 nM of eryngitin 3, the breakdown of nuclear DNA into oligonucleosomal fragments was clearly observed (Figure 3d). Altogether, our data suggested that the apoptotic pathway, was involved in cell death mediated by ribonucleolytic activity of eryngitin 3 and 4, as already proved for other RL-Ps [2].
Finally, we evaluate the effects of eryngitin 3 and 4 on the growth of the green mold Penicillium digitatum. This ascomycete fungus is responsible for the postharvest decay of citrus, causing a significant reduction in citrus quality and marketable yield [27]. As shown in Figure 4, eryngitin 3 and 4 exhibit a growth inhibitory effect on P. digitatum in a concentration dependent manner.
In particular, 60, 30 and 12 µg/mL of eryngitin 3 resulted in 84%, 76% and 66% growth inhibition, respectively, after 84 h of growth (Figure 4a). On the contrary, eryngitin 4 showed less ability to inhibit P. digitatum growth. Indeed, fungal growth was inhibited by 53%, 36% and 18% after 84 h using 98, 49 and 20 µg/mL of eryngitin 4, respectively, which are ~1.6-fold higher concentrations with respect to those of eryngitin 3 (Figure 4b). Furthermore, the toxicity of eryngitins on mycelial growth of P. digitatum was also visualized microscopically by observing alterations of hyphal morphology (Figure 4c).
Taken together, both biological and antipathogenic data evidence a more pronounced toxic effect of eryngitin 3 compared to eryngitin 4, although both enzymes are able to release the α-fragment, making necessary a complete structural characterization to justify their functional differences.
2.3. Relative molecular masses of eryngitin 3 and 4 with and without alkylation
In order to achieve the relative molecular masses (Mr) of native eryngitin 3 and 4, as well as their cysteinyl residue/s content, Mr of both native and alkylated proteins after RP-HPLC desalting was determined by MALDI-ToF MS analyses.
The chromatographic profiles of native eryngitin 3 and 4 are reported in Figure 1b,c, respectively. RP-HPLC highlights a heterogeneity for eryngitin 3 preparation, not detected by SDS-PAGE analysis, characterized by the elution of two-overlapping protein peaks (Figure 1b; component a ~68%, component b ~32%) and a single protein peak for eryngitin 4 (Figure 1c), confirming for the latter the homogeneity found by SDS-PAGE analysis. Moreover, considering the difficulty in separating the two components of eryngitin 3 by RP-HPLC after several attempts of elution conditions, we decided to directly analyse both component a and b of eryngitin 3 in mixture after RP-HPLC as well as the single peak of eryngitin 4 by MALDI-ToF MS. Typical MALDI-ToF MS spectra of eryngitin 3 mixture and eryngitin 4 are shown in Figure 5a,b, respectively.
The mixture of two-overlapping eryngitin 3 peaks (component a and b) contained two experimental Mr, 14447.86 ([M+H+]+) and 14302.18 ([M+H+]+), while a single Mr of 14454.98 ([M+H+]+) was retrieved for eryngitin 4. In particular, a difference of ~146-Da (14447.86 - 14302.18 = ~146-Da) between component a and b was found in eryngitin 3 mixture. Subsequently, the experimental Mr of eryngitin 3 and 4 were acquired after reduction and alkylation with 4-vinylpyridine, and the related MALDI-ToF MS spectra are displayed in Figure 5c,d. In these conditions, experimental Mr of both components detected in eryngitin 3 mixture (component a and b after RP-HPLC) and the Mr of eryngitin 4 differed of ~105-Da comparing to the experimental Mr of native proteins. This additional molecular mass of ~105-Da corresponds to the chemical modification of a single cysteinyl residue to pyridyl-cysteine, in the presence of 4-vinylpyridine [28]. In addition, similar results were obtained also when the proteins were directly subjected to alkylation with 4-vinylpyridine without preliminary reduction. Indeed, both eryngitin 3 components and eryngitin 4 exhibit an additional molecular mass of ~105-Da (data not shown).
Overall, these results suggest that both components of eryngitin 3 as well as eryngitin 4 contain a single free cysteinyl residue susceptible to S-pyridylethylation (Δ-mass = 105-Da). This structural characterization suggests that the covalent dimeric forms retrieved in SDS-PAGE without reducing agent are due to the formation of interchain disulphide bonds under denaturing conditions.
2.4. N-terminal amino acid determination and search in fungal data base genome
In order to clarify the structural features which could explain the differences in biological and antipathogenic activities of eryngitin 3 and 4, we decided to get the amino acid sequence of both proteins. Therefore, to optimize the information reported in the fungal genomics resource ‘MycoCosm’ on-line database [29], the following strategy was carried out: i) N-terminal amino acid sequence determination by using Edman degradation; ii) search of similar proteins in P. eryngii genome by using N-terminal sequence as query; iii) enzymatic cleavage with endoproteinases (i.e.: trypsin, endoproteinase Glu-C, chymotrypsin, and pepsin) followed by MALDI-ToF MS analysis of the resulting peptides; and iv) mapping of the obtained peptides with homologous reference protein.
Automatic Edman degradation of eryngitin 4 and 3 mixture after RP-HPLC provided a single N-terminal sequence for eryngitin 4 and two different amino acid sequences for eryngitin 3 (the two N-terminal amino acid sequences were assigned on the basis of their different amount) [30]. N-terminal amino acid sequences up to 20 residues are reported in Data report 1.

Data report 1. N-terminal amino acid sequences of alkylated eryngitin 4 and eryngitin 3 obtained by automatic Edman degradation. Eryngitin 3 exhibit two amino acid sequences (a and b) assigned on the basis of their different amount.
These data show that most of amino acid residues constituting eryngitin 4 N-terminal sequence and the two N-terminal sequences of eryngitin 3 are identical. In particular, the N-terminal of eryngitin 4 and the N-terminal amino acid sequence present in less amount (30%; component b) in eryngitin 3 mixture are identical, while the N-terminal sequence present in higher amount (70%; component a) in eryngitin 3 mixture has an additional phenylalanyl residue at the N-terminal region.
Subsequently, the N-terminal amino acid sequence of eryngitin 3 (1-FGEVTQNYPS KELASKAACT-20) was used as query for protein sequence identification in P. eryngii genome by using BlastP tool. The algorithm identified a hypothetical protein BDN71DRAFT_1455417 (AC: KAF9489889.1; 162 amino acid residues) (Figure S2), which showed the same N-terminal amino acid sequence of both eryngitin 3 and 4. In this framework, we decided to use the amino acid sequence of this hypothetical protein as a reference protein for the determination of eryngitin 4 and 3 amino acid sequences.
2.5. Determination of eryngitin 4 amino acid sequence
Considering the high homogeneity of eryngitin 4, we firstly determined its amino acid sequence, by MALDI-ToF MS analysis followed by peptide mapping strategy using the hypothetical protein BDN71DRAFT_1455417 as reference protein (162 amino acid residues; Figure S2), which exhibits 100% identity with the first 20 amino acid residues of alkylated eryngitin 4.
In light of this, a first trypsin mapping on alkylated eryngitin 4 was carried out. Tryptic peptides molecular masses retrieved in digested samples are reported in Table S2, whereas their sequence positions are mapped in Figure 6a. This set of data provided for 67.8% of reference protein (99 out of 146 amino acids, considering Gly17 and Ala162 as first and last amino acid residues at N- and C-terminus of hypothetical protein BDN71DRAFT_1455417, respectively, (Figure S2). Notably, no tryptic peptide mapping of the reference protein the C-terminal region was found. Thus, for sequence completion and/or verification, we decided to perform a complete peptide mapping by MALDI-ToF mass spectrometry, with new peptides sets of alkylated eryngitin 4 derived from endoproteinase Glu-C, chymotrypsin and pepsin digestion. These new peptides sets are reported in Table S2, and mapped in Figure 6a. Overall, all obtained peptides overlap the entire amino acid sequence of the reference protein from Gly17 to Arg148. Thus, the results exclude that native eryngitin 4 has the amino acid regions at the N- (position 1-16) and C-terminal (position 149-162) regions, like the reference protein, confirming that eryngitin 4 consists of 132 amino acids (Figure 6a).
Finally, the agreement between the experimental mass of the native protein (Figure 5b; 14454.98 Da; [M+H+]+) and the calculated one, on the basis of the amino acid sequence (14455.38 Da) confirms the correctness of amino acid sequence determination. Moreover, eryngitin 4 preserves the catalytic triad of RL-Ps: Asp62, Asp 64 and His75 assigned by homology, considering the experimental data reported for ageritin, prototype of RL-Ps family [9,10].
2.6. Determination of eryngitin 3 amino acid sequence
The same strategy reported for the determination of eryngitin 4 amino acid sequence was carried out to acquire also the primary structure of eryngitin 3, considering that the principal component of eryngitin 3 (component a, ~70%) has as first/additional amino acid, a phenylalanyl residue at N-terminus, as retrieved by Edman degradation, present also at the N-terminal region of reference protein (hypothetical protein BDN71DRAFT_1455417), Figure S2.
Tryptic mapping of alkylated eryngitin 3 (Mr peptides set reported in Table S3, and sequence positions are mapped in Figure 6b), provided for 58.2% of the reference protein (85 out of 146 amino acids, considering Phe16 and Ala162 as the first and last amino acid residue, at N- and C-terminus of the hypothetical protein BDN71DRAFT_1455417, respectively, (Figure S2). Notably, this first data set highlights: i) the absence of tryptic peptides to map the C-terminal region of the reference protein; and ii) the presence of a T1 peptide with a Mr of 1798.25 ([M+H+]+) to overlap the sequence at position 1-16, confirming the additional phenylalanyl residue at the N-terminus of eryngitin 3 (Figure 6b). Moreover, a peptide with an experimental Mr of 1122.59 (Table S3), alternative N-terminus starting with a glycinyl residue (position 2-11 in eryngitin 3, N-terminal peptide of component b), was also detected, confirming that eryngitin 3 is a mixture of two proteins which differ in amino acid at the N-terminus. Indeed, the molecular weight of a phenylalanyl residue is 147.18 Da, corresponding to the molecular weight difference found between component a and b of eryngitin 3 detected after separation by RP-HPLC and MALDI ToF MS analysis (Figure 5c; 14447.86 - 14302.18 = ~146 Δ-mass).
However, for eryngitin 3 sequence completion and/or verification, a complete peptide mapping by MALDI-ToF mass spectrometry, with new peptides set of alkylated eryngitin 3 derived from endoproteinase Glu-C, chymotrypsin and pepsin digestion, was performed. The novel different sets of peptides obtained are reported in Table S3 and mapped in Figure 6b. Overall, all peptides obtained overlap with the entire amino acid sequence of the reference protein, from Phe16 to Gly147 (Figure S2). Thus, the results achieved show that eryngitin 3 lacks the additional amino acid regions at the N- (position 1-15) and C-terminus (position 148-162) of the reference protein (Figure S2), while the component retrieved after RP-HPLC analysis differs only by the presence (component a, ~70%) or absence (component b, ~30%) of a phenylalanyl residue at the N-terminus.
Finally, the agreement between experimental Mr of the native protein (component a, 14447.86 Da; component b, 14302.18 Da) and that calculated from the amino acid sequence (component a, 14446.36 Da; component b, 14299.19) confirm the correctness of the analysis.
2.7. Features of eryngitin 3 and 4 gene
The amino acid sequence of the hypothetical protein BDN71DRAFT_1455417 (AC: KAF9489889.1) was obtained by analysing P. eryngii ATCC 90797 genome [31]. In particular, the genomic sequence (831 bp) is reported in MycoCosm database as “Pleery1_1455417” ID gene. The transcript sequence (730 bp) contains the coding sequence (CDS; 489 bp) of the hypothetical protein KAF9489889.1, showing additional N- and C-terminal amino acid sequences with respect to both native eryngitins, and two introns (position 165-216 and 249-297) at 5’ transcript region (Figure S3).
The translation of this transcript covers the entire structure of both purified eryngitins (132 amino acids) including two additional amino acid sequences of ~15 and ~14 at the N- and C-terminus, respectively, compared to purified proteins, Figure S3. This result is consistent with previous data obtained for both ostreatin [6] and ageritin [24], RL-Ps isolated from P. ostreatus and C. aegerita fruiting bodies, respectively, confirming that these toxins are likely synthesized as preforms, which includes both amino acid regions (signal peptides), involved in toxin regulation or subcellular localization, and subsequently removed by specific proteases [32].
2.8. Sequence comparison between eryngitin 3 and 4 and other well-characterized RL-Ps
The alignment between eryngitin 4 and eryngitin 3 as well as ostreatin (RL-P from P. ostreatus fruiting bodies) and ageritin (RL-P from C. aegerita fruiting bodies) was displayed in Figure S4. They are well-characterized RL-Ps, and all amino acid sequences conserve the following invariant amino acid residues: i) three amino acids forming the catalytic site (Asp69, Asp71 and His82, numbering referred to the alignment consensus sequence); ii) one cysteinyl residue (Cys19) at the N-terminal region; and iii) three tryptophanyl residues (W130, W132 and W133) at the C-terminal region. The latter is a structural feature of this protein family, as already highlighted by previous homology studies using ageritin as a query in protein databases searching [2]. Furthermore, the identity between eryngitins and ostreatin, isolated from the same genus (Pleurotus) is 97.7% (~98.5% similarity), while the identity between eryngitins and ageritin (Cyclocybe genus) is 40.1% (~45.4% similarity). This observation highlights the structural differences among RL-Ps found in edible mushrooms belonging to different basidiomycetes genera, which exhibit peculiar enzymatic action/toxicity towards several cell lines or fungal model systems, such as Penicillium digitatum [2,5].
2.9. Modeling the 3D structure of eryngitin 4 and 3
In order to gain insight into the structural differences in the biological and antipathogenic activities of both eryngitins, a structure prediction study was carried out by using AlphaFold software based on artificial intelligence. As expected, the superimposed 3D models of eryngitin 4 and 3 exhibit the same protein fold (Figure 7a), considering that the two proteins are constituted by the same amino acid sequence, apart from a single residue at both the C-terminus [additional arginyl residue for eryngitin 4 (Arg132); Figure 6a] and the N-terminus [additional phenylalanyl residue for eryngitin 3 (Phe1); Figure 6b].
Moreover, the graphic of ‘Local Distance Difference Test’ (lDDT), a superposition-free score used to evaluate the local distance differences of all atoms in a model [34], displays for both eryngitins models a value higher than 90, except for positions 62-78, and the first N- and C-terminus residues (Figure S5). These 3D models consist of an extended antiparallel β-sheet with an interface in which the catalytic triad (Asp62, Asp 64 and His75, numbering referred to eryngitin 4) is conserved and an opposite interface resting on four principal α-helixes (α-helix Lys10 – Ala21; α-helix Glu93 – Ala97; α-helix P100 – Ile114; and α-helix P119 – Thr128; numbering referred to eryngitin 4), which form the bulk of the main protein fold. In particular, it is interesting to note that the main region with evident low IDDT (50< IDDT<70, positions 62-78), corresponds to the catalytic site of these enzymes, likely confirming that the catalytic site of enzymes is usually characterised by intrinsic conformational freedom necessary to adapt to the substrate [35]. Slight structural differences between eryngitin 4 and 3 are evident only at the N-terminal and C-terminal regions, as shown by electrostatic surface representation for the two different 3D models (Figure 7b for eryngitin 4 and Figure 7c for eryngitin 3). Indeed, an accentuated positive electrostatic surface was found at the C-terminal region for eryngitin 4, due to the presence of an additional arginyl residue, while a slightly negative electrostatic surface was found at the N-terminal region of eryngitin 3, due to the presence of an additional phenylalanyl residue.
This observation could explain the functional differences retrieved between the two proteins. Indeed, as highlighted by several structural/functional studies, analogous toxins such as: i) Shiga-like toxins (bacterial toxins) [36,37]; ii) ribotoxins (ascomycetes toxins) [38]; and iii) ribosome inactivating proteins (plant toxins) [36,39], sharing the same target (SRL) of eryngitins, exhibit different functional action due to their interaction and/or binding capacity with both SRL and nearby macromolecules [40]. In particular, the macromolecules which can interfere or enhance the binding between these toxins and SRL are some ribosomal proteins, generally forming the ribosomal stalk [20,37], a specific ribosomal structure, required for the binding of elongation factors to the ribosomes during protein translation [41,42]. In this contest, it can be hypothesized that slight electrostatic surface changes at the N- and C-terminal regions could justify the different functional features of the two eryngitins, by altering their interaction with SRL, the site where they exploit their specific ribonuclease activity.
In addition, some experimental data led us to consider that the involvement of slight structural variations at the N- and C-terminal regions may account for the differences in biological and antipathogenic activities of both eryngitins. Indeed, although the two enzymes share the same structural fold, eryngitin 3 exhibits a higher thermal stability (Tm= ~89.7 °C) compared to eryngitin 4 (Tm= ~83.1 °C) [7]. Furthermore, characterization of the ageritin isoform, named Met-ageritin (homologous to eryngitins) with an additional N-terminal methionyl residue compared to ageritin, showed that this single additional amino acid residue at the N-terminus, drastically decreased Met-ageritin enzymatic action [22]. Finally, the single additional positive charge of eryngitin 4 due to the arginyl residue at the C-terminus affects its physicochemical characteristics, considering a longer retention time of eryngitin 4 compared to eryngitin 3 during the last purification step (ion exchange chromatography, which separates proteins on the basis of charge differences) [7].
3. Conclusions
Knowledge of protein primary structure may contribute to understanding biological differences in enzymatic and functional activities of proteins.
In light of this, we determined the primary structure of eryngitin 3 and 4, two ribotoxin-like proteins isolated from P. eryngii fruiting bodies. Despite both enzymes are able to release the α-fragment (hallmark of RL-Ps from edible mushrooms) when incubated with ribosomes from different sources (yeast, and mealworm insect) they exhibit different biological and antipathogenic activities. Eryngitin 3 and 4 consist of 132 amino acid residues with a single free cysteinyl residue. Most of the residues between the two toxins are identical, considering that the two proteins have the same amino acid sequence, apart from a single residue at both N-terminus (additional phenylalanyl residue for eryngitin 3) absent in eryngitin 4 and C-terminus (additional arginyl residue for eryngitin 4) absent in eryngitin 4. Finally, the predicted models of the two toxins by AlphaFold highlight that the additional positive charge retrieved from eryngitin 4 at the C-terminal region could interfere with the binding to the substrate (SRL) or with some ribosomal proteins (e.g.: P-stalk), during substrate recognition, providing a possible explanation for their different toxicity.
Further studies will be necessary to understand the function of eryngitin 3 and 4 (both constitutively expressed in P. eryngii fruiting bodies) likely with a different action such as self-defence (i.e.: the more toxic eryngitin 3) or for a physiological role (i.e.: the less toxic eryngitin 4).
4. Material and Methods
4.1. Chemicals and reagents
The chemicals used in this work are previously reported [1,6,7] and most of them are obtained from Sigma-Aldrich Solutions (Merk Life Science, Milan, Italy). Endoproteinases (trypsin TPCK-treated, chymotrypsin, endoproteinase Glu-C and pepsin) for protein digestion were purchased from Sigma-Aldrich Solutions. Century™-Plus RNA Markers were purchased from Fisher Scientific (Madrid, Spain). Potato dextrose agar and Potato dextrose broth media were purchased from Sigma-Aldrich (Madrid, Spain). The RPMI 1640 medium, fetal bovine serum (FBS), penicillin, streptomycin and trypsin were purchased from GIBCO BRL (Barcelona, Spain). The Z-VAD-fmk (pan-caspase inhibitor carbobenzoxy-valyl-alanyl-aspartyl-[O-methyl]-fluoromethylketone) named ZVAD was purchased from R&D Systems (Abingdon, UK). GelRed was purchased from Biotium Inc. (Hayward, CA, USA).
The mealworms (Tenebrio molitor L.) were bought at local markets. The strain of Penicillium digitatum (Pers.) Sacc. was obtained by the Spanish Type Culture Collection (CECT), Valencia, Spain. COLO 320 (human colon adenocarcinoma) and HeLa (human cervix epitheloid carcinoma) cell lines used in this study were obtained from the European Collection of Cell Cultures (ECACC). and grown in RPMI 1640 medium (GIBCO BRL, Barcelona, Spain) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin and 0.1 mg/mL streptomycin under 5% CO2 at 37°C.
4.2. Proteins purification
The purification of eryngitin 3 and 4 for biological assessment and structural characterization was achieved as previously reported [7]. Briefly, P. eryngii fruiting bodies extract was subjected to acetic acid precipitation at pH 4.0, and soluble proteins were separated, exploiting the differences in both protein size (size exclusion chromatography) and charge (cation exchange chromatography) [2].
4.3. Analytical procedures
Purity and integrity of eryngitin 3 and 4 were determined by SDS-PAGE with a Mini-Protean II mini-gel apparatus (Bio-Rad, Milan, Italy) using 6% stacking and 15% separation polyacrylamide gel; a precision plus protein kit (Bio-Rad, Hercules, CA, USA) was used for reference proteins. Protein concentration was determined by Pierce BCA Protein Assay kit (Thermo Fisher Scientific, Rodano, Italy), using BSA as standard.
4.4. Assays of cell-free protein synthesis
The effect of eryngitin 3 and 4 on protein synthesis was determined through a coupled transcription-translation in vitro assay using a rabbit reticulocytes lysate system (Promega, Alcobendas, Madrid, Spain) as previously reported [43]. Data represent the mean of three experiments in triplicate.
4.5. Ribonucleolytic activity on yeast and mealworm ribosomes
The specific activity of ribotoxins, manifested by the release of α-fragment from rRNA after enzymatic action on yeast and mealworm ribosomes, was detected as described elsewhere [1,21,44]. The 30,000 g supernatants (partial purified ribosomes, named S30) from yeast and mealworm (used as substrates) were obtained as described elsewhere [25,43,44,45].
4.6. Cell viability assays
Cell viability was determined with a colorimetric assay based on the cleavage of tetrazolium salt WST-1 to formazan by mitochondrial dehydrogenases in viable cells, as described elsewhere [43,45]. The concentration of toxins causing a 50% reduction in viability (IC50) was calculated by linear regression analysis. Eryngitin 3 and 4 toxicity was also evaluated using HeLa cells pre-treated with 100 µM of the pan-caspase inhibitor Z-VAD. The reagent was added to cells 3 h before toxin administration, and cell viability was determined in the presence of different toxin concentrations.
4.7. RNA extraction from HeLa cells
HeLa cells (1 x 106/plate) were incubated for 72 h in presence of 30 nM eryngitin 3. After treatment, cells were harvested by centrifugation at 1000 ×g for 5 min. The pellets were lysed, and the RNA was isolated following the instruction of the RNeasy Mini Kit (Qiagen GmbH, Hilden, Germany). The RNAs were subjected to electrophoresis at 16 mA in a 7.0 M urea/5% (w/v) polyacrylamide gel for 120 min and stained with GelRed nucleic acid staining [43].
4.8. DNA fragmentation analysis
COLO 320 cells (1 x 106/plate) were incubated for 72 h in the presence of eryngitins (300 nM). After treatment, cells were harvested by centrifugation (1000x g for 5 min). The pellets were lysed in 50 mM Tris•Cl, pH 8.0, containing 10 mM EDTA and 0.5% SDS, and the DNA was isolated following the manufacturer’s instructions [Genomic Prep Cells and Tissue DNA Isolation Kit (GE Healthcare, Madrid, Spain)]. DNA electrophoresis was carried out as previously reported [43].
4.9. Antifungal activity
4.10. Reduction and S-pyridylethylation
S-pyridylethylation (cysteinyl residues alkylation with 4-vinylpyridine [28]) with and without reducing agent was performed as previously reported [46]. Native or alkylated desalted proteins were obtained by RP-HPLC using a BioBasic-4 column (150 × 4.6 mm, 5-μm particle size; Thermo Fisher Scientific) at 25 °C. Solvent used are: milliQ water containing 0.1 % TFA (solvent A); acetonitrile containing 0.1 % TFA (solvent B). Proteins elution was performed using a linear gradient of solvent A and solvent B, from 5% to 65% of solvent B over 60 min (flow rate of 1.0 mL/min), monitoring the absorbance at 214 nm.
4.11. Automatic N-terminal Edman degradation
N-terminal sequence of the proteins was obtained by using automated Edman degradation performed on a Shimadzu PPSQ 33B sequencer (Shimadzu Italia S.r.l., Milan, Italy) at the service of ‘Protein/Peptide sequencing’, Institute of Biosciences and Bioresources (IBBR-CNR, Naples, Italy). Proteins were firstly alkylated with 4-vinylpyridine, desalted by RP-HPLC (paragraph 2.10), freeze-dried, and loaded on the sequencer.
4.12. Protein digestion for MALDI-ToF analysis
Enzymatic hydrolyses (i.e.: trypsin TPCK-treated, chymotrypsin, endoproteinase Glu-C and pepsin) of eryngitin 3 and 4 (50 µg) were achieved by dissolving the alkylated proteins in appropriate buffer. Specific conditions for the different proteases are listed below:
trypsin: buffer 50 mM (NH4)HCO3, containing 10% acetonitrile; enzyme/substrate 1:50 (w/w) final ratio added in three steps [the first at 0 h (E/S: 1:200); the second at 4 h (E/S: 1:100) and the third at 12 h (overnight, E/S: 1:50); 16 h total]; incubation at 37 °C [23,30];
endoproteinase Glu-C: buffer Na-phosphate 50 mM, pH 7.8, containing 10% acetonitrile; enzyme/substrate 1:25 (w/w) final ratio added in three steps [the first at 0 h (E/S: 1:100); the second at 4 h (E/S: 1:50) and the third at 12 h (overnight, E/S: 1:25); 16 h total]; incubation at 25 °C [23,30];
4.13. MALDI-ToF mass spectrometry analyses
4.14. Sequence analyses
Amino acid sequence identical to N-terminal sequence of eryngitin 3 and 4 was retrieved by using BlastP (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE=Proteins). To this aim, eryngitin 3 N-terminal amino acid sequences were used as query by setting non-redundant protein sequences (nr) database and Fungi (taxid:4751) as organism. Sequences alignment was performed using ClustalW tool available on-line (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_clustalw.html). Similarity/identity matrix and Logo were obtained using the Sequence Identity and Similarity (SIAS) tool (http://imed.med.ucm.es/Tools/sias.html) and WebLogo-3 (https://weblogo.threeplusone.com/create.cgi), respectively. Information on hypothetical protein BDN71DRAFT_1455417 (GenBank AC: KAF9489889.1) annotation is reported at the following link (https://mycocosm.jgi.doe.gov/cgi-bin/dispGeneModel?db=Pleery1&id=1455417) and is referred to the nucleotide sequence present in P. eryngii ATCC 90797 genome [31], named ‘jgi|Pleery1|1455417’.
3D structural models of eryngitin 3 and 4 were obtained using AlphaFold [49] available in UCSF ChimeraX software [50,51]. Studies on electrostatic superficial regions and graph representations were performed by using UCSF Chimera suite version 1.16 (https://www.cgl.ucsf.edu/chimera/) [52].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
A.D.M.: Conceptualization, Writing original draft, Methodology and Funding acquisition. S.R., N.L. and A.Cl.: Protein purification, analytical methods for amino acid sequence determination. S.R., R.R., and A.Ch: Mass Spectrometry analyses. N.L., P.V.P and M.S.: Structural computational analyses. L.C., R.I. and J.M.F.: Biological assays (cell-free protein synthesis, ribonucleolytic activity, cell viability, antifungal activity). S.R. and A.D.M.: Review and Editing. All authors are in agreement with the final submitted manuscript data.
Funding
this work was supported by the research project PRR.AP026.014 - PNRR - IR0000009 - Title: ‘Potentiating the Italian Capacity for Structural Biology Services in Instruct-ERIC’. Acronym: ITACA.SB. Announcement: ‘MUR D.D. 0003264 del 28/12/2021’ as well as grants from the “Ministero dell’Università e della Ricerca” (MUR; Prin 2022 PNRR, entitled “Direct or indirect effects of protein synthesis inhibitor enzymes from edible plants and mushrooms on intestinal health and microbiota”, Prot. P2022YERKR). In addition, this work was supported by grant VA033G19 (Consejería de Educación, Junta de Castilla y León) to the GIR ProtIBio.
Acknowledgments
This study was made possible by care and abnegation of all participants, despite the absence of dedicated funds and chronic difficulties afflicting the Italian scientific community.
Conflicts of Interest
The authors have no conflict of interest to declare. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Landi, N.; Pacifico, S.; Ragucci, S.; Iglesias, R.; Piccolella, S.; Amici, A.; Di Giuseppe, A.M.A.; Di Maro, A. Purification, characterization and cytotoxicity assessment of Ageritin: The first ribotoxin from the basidiomycete mushroom Agrocybe aegerita. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Ragucci, S.; Landi, N.; Russo, R.; Valletta, M.; Pedone, P.V.; Chambery, A.; Di Maro, A. Ageritin from Pioppino Mushroom: The Prototype of Ribotoxin-Like Proteins, a Novel Family of Specific Ribonucleases in Edible Mushrooms. Toxins (Basel) 2021, 13, 263–279. [Google Scholar] [CrossRef]
- Endo, Y.; Huber, P.W.; Wool, I.G. The ribonuclease activity of the cytotoxin alpha-sarcin. The characteristics of the enzymatic activity of alpha-sarcin with ribosomes and ribonucleic acids as substrates. J. Biol. Chem. 1983, 258, 2662–2667. [Google Scholar] [CrossRef] [PubMed]
- Landi, N.; Ragucci, S.; Culurciello, R.; Russo, R.; Valletta, M.; Pedone, P.V.; Pizzo, E.; Di Maro, A. Ribotoxin-like proteins from Boletus edulis: structural properties, cytotoxicity and in vitro digestibility. Food Chem. 2021, 359, 129931. [Google Scholar] [CrossRef] [PubMed]
- Ragucci, S.; Hussain, H.Z.F.; Bosso, A.; Landi, N.; Clemente, A.; Pedone, P.V.; Pizzo, E.; Di Maro, A. Isolation, Characterization, and Biocompatibility of Bisporitin, a Ribotoxin-like Protein from White Button Mushroom (Agaricus bisporus). Biomolecules 2023, 13, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Landi, N.; Ragucci, S.; Russo, R.; Valletta, M.; Pizzo, E.; Ferreras, J.M.; Di Maro, A. The ribotoxin-like protein Ostreatin from Pleurotus ostreatus fruiting bodies: Confirmation of a novel ribonuclease family expressed in basidiomycetes. Int. J. Biol. Macromol. 2020, 161, 1329–1336. [Google Scholar] [CrossRef]
- Landi, N.; Grundner, M.; Ragucci, S.; Pavšič, M.; Mravinec, M.; Pedone, P.V.; Sepčić, K.; Di Maro, A. Characterization and cytotoxic activity of ribotoxin-like proteins from the edible mushroom Pleurotus eryngii. Food Chem. 2022, 396, 133655. [Google Scholar] [CrossRef]
- Wang, M.; Zhao, R. A review on nutritional advantages of edible mushrooms and its industrialization development situation in protein meat analogues. J. Future Foods 2023, 3, 1–7. [Google Scholar] [CrossRef]
- Ruggiero, A.; García-Ortega, L.; Moreira, M.; Ragucci, S.; Landi, N.; Di Maro, A.; Berisio, R. Binding and enzymatic properties of Ageritin, a fungal ribotoxin with novel zinc-dependent function. Int. J. Biol. Macromol. 2019, 136, 625–631. [Google Scholar] [CrossRef]
- Tayyrov, A.; Azevedo, S.; Herzog, R.; Vogt, E.; Arzt, S.; Lüthy, P.; Müller, P.; Rühl, M.; Hennicke, F.; Künzler, M. Heterologous Production and Functional Characterization of Ageritin, a Novel Type of Ribotoxin Highly Expressed during Fruiting of the Edible Mushroom Agrocybe aegerita. Appl. Environ. Microbiol. 2019, 85, e01549–e01519. [Google Scholar] [CrossRef]
- Lampitella, E.; Landi, N.; Oliva, R.; Ragucci, S.; Petraccone, L.; Berisio, R.; Di Maro, A.; Del Vecchio, P. Conformational stability of ageritin, a metal binding ribotoxin-like protein of fungal origin. Int. J. Biol. Macromol. 2022, 221, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Wahl, M.C.; Möller, W. Structure and function of the acidic ribosomal stalk proteins. Curr. Protein Pept. Sci. 2002, 3, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Feng, S.; Kumar, V.; Ero, R.; Gao, Y.-G. Structure of EF-G–ribosome complex in a pretranslocation state. Nat. Struct. Mol. Biol. 2013, 20, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Stirpe, F. Ribosome-inactivating proteins: from toxins to useful proteins. Toxicon 2013, 67, 12–16. [Google Scholar] [CrossRef]
- Grela, P.; Szajwaj, M.; Horbowicz-Drożdżal, P.; Tchórzewski, M. How Ricin Damages the Ribosome. Toxins (Basel) 2019, 11, 541–556. [Google Scholar] [CrossRef]
- Robert, A.; Wiels, J. Shiga Toxins as Antitumor Tools. Toxins (Basel) 2021, 13, 690–704. [Google Scholar] [CrossRef]
- Rudolph, M.J.; Davis, S.A.; Tumer, N.E.; Li, X.P. Structural basis for the interaction of Shiga toxin 2a with a C-terminal peptide of ribosomal P stalk proteins. J. Biol. Chem. 2020, 295, 15588–15596. [Google Scholar] [CrossRef]
- Lacadena, J.; Alvarez-García, E.; Carreras-Sangrà, N.; Herrero-Galán, E.; Alegre-Cebollada, J.; García-Ortega, L.; Oñaderra, M.; Gavilanes, J.G.; Martínez del Pozo, A. Fungal ribotoxins: molecular dissection of a family of natural killers. FEMS Microbiol. Rev. 2007, 31, 212–237. [Google Scholar] [CrossRef]
- Herrero-Galán, E.; García, E.; Carreras-Sangrà, N.; Lacadena, J.; Alegre-Cebollada, J.; Martínez-del-Pozo, A.; Oñaderra, M.; Gavilanes, J. Fungal ribotoxins: structure, function and evolution. In Microbial Toxins: Current Research and Future Trends., Proft, T., Ed. Caister Academic Press,: Norfolk, UK, 2009; pp. 167–187.
- Choi, A.K.; Wong, E.C.; Lee, K.M.; Wong, K.B. Structures of eukaryotic ribosomal stalk proteins and its complex with trichosanthin, and their implications in recruiting ribosome-inactivating proteins to the ribosomes. Toxins (Basel) 2015, 7, 638–647. [Google Scholar] [CrossRef]
- Citores, L.; Ragucci, S.; Ferreras, J.M.; Di Maro, A.; Iglesias, R. Ageritin, a Ribotoxin from Poplar Mushroom (Agrocybe aegerita) with Defensive and Antiproliferative Activities. ACS Chem. Biol. 2019, 14, 1319–1327. [Google Scholar] [CrossRef]
- Ragucci, S.; Landi, N.; Russo, R.; Valletta, M.; Citores, L.; Iglesias, R.; Pedone, P.V.; Pizzo, E.; Di Maro, A. Effect of an additional N-terminal methionyl residue on enzymatic and antifungal activities of Ageritin purified from Agrocybe aegerita fruiting bodies. Int. J. Biol. Macromol. 2020, 155, 1226–1235. [Google Scholar] [CrossRef] [PubMed]
- Landi, N.; Ragucci, S.; Russo, R.; Pedone, P.V.; Chambery, A.; Di Maro, A. Structural insights into nucleotide and protein sequence of Ageritin: a novel prototype of fungal ribotoxin. J. Biochem. (Tokyo) 2019, 165, 415–422. [Google Scholar] [CrossRef]
- Baglivo, I.; Ragucci, S.; D'Incecco, P.; Landi, N.; Russo, R.; Faoro, F.; Pedone, P.V.; Di Maro, A. Gene Organization, Expression, and Localization of Ribotoxin-Like Protein Ageritin in Fruiting Body and Mycelium of Agrocybe aegerita. Int. J. Mol. Sci. 2020, 21, 7158–7173. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, R.; Citores, L.; Ragucci, S.; Russo, R.; Di Maro, A.; Ferreras, J.M. Biological and antipathogenic activities of ribosome-inactivating proteins from Phytolacca dioica L. Biochim. Biophys. Acta 2016, 1860, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Citores, L.; Iglesias, R.; Ragucci, S.; Di Maro, A.; Ferreras, J.M. Antifungal Activity of α-Sarcin against Penicillium digitatum: Proposal of a New Role for Fungal Ribotoxins. ACS Chem. Biol. 2018, 13, 1978–1982. [Google Scholar] [CrossRef]
- Wang, Z.; Sui, Y.; Li, J.; Tian, X.; Wang, Q. Biological control of postharvest fungal decays in citrus: a review. Crit. Rev. Food Sci. Nutr. 2022, 62, 861–870. [Google Scholar] [CrossRef]
- Friedman, M. Application of the S-pyridylethylation reaction to the elucidation of the structures and functions of proteins. J. Protein Chem. 2001, 20, 431–453. [Google Scholar] [CrossRef]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F. , et al. MycoCosm portal: gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014, 42, D699–D704. [Google Scholar] [CrossRef]
- Di Maro, A.; Chambery, A.; Carafa, V.; Costantini, S.; Colonna, G.; Parente, A. Structural characterization and comparative modeling of PD-Ls 1-3, type 1 ribosome-inactivating proteins from summer leaves of Phytolacca dioica L. Biochimie 2009, 91, 352–363. [Google Scholar] [CrossRef]
- Ruiz-Dueñas, F.J.; Barrasa, J.M.; Sánchez-García, M.; Camarero, S.; Miyauchi, S.; Serrano, A.; Linde, D.; Babiker, R.; Drula, E.; Ayuso-Fernández, I. , et al. Genomic Analysis Enlightens Agaricales Lifestyle Evolution and Increasing Peroxidase Diversity. Mol. Biol. Evol. 2021, 38, 1428–1446. [Google Scholar] [CrossRef]
- Nielsen, H.; Tsirigos, K.D.; Brunak, S.; von Heijne, G. A Brief History of Protein Sorting Prediction. Protein J. 2019, 38, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Kupfer, D.M.; Drabenstot, S.D.; Buchanan, K.L.; Lai, H.; Zhu, H.; Dyer, D.W.; Roe, B.A.; Murphy, J.W. Introns and splicing elements of five diverse fungi. Eukaryot. Cell 2004, 3, 1088–1100. [Google Scholar] [CrossRef] [PubMed]
- Mariani, V.; Biasini, M.; Barbato, A.; Schwede, T. lDDT: a local superposition-free score for comparing protein structures and models using distance difference tests. Bioinformatics 2013, 29, 2722–2728. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.K. Enzymes: An integrated view of structure, dynamics and function. Microb. Cell Fact. 2006, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Tumer, N.E.; Li, X.P. Interaction of ricin and Shiga toxins with ribosomes. Curr. Top. Microbiol. Immunol. 2012, 357, 1–18. [Google Scholar] [CrossRef]
- McCluskey, A.J.; Bolewska-Pedyczak, E.; Jarvik, N.; Chen, G.; Sidhu, S.S.; Gariépy, J. Charged and hydrophobic surfaces on the a chain of shiga-like toxin 1 recognize the C-terminal domain of ribosomal stalk proteins. PLoS One 2012, 7, e31191. [Google Scholar] [CrossRef]
- Pérez-Cañadillas, J.M.; Santoro, J.; Campos-Olivas, R.; Lacadena, J.; Martínez del Pozo, A.; Gavilanes, J.G.; Rico, M.; Bruix, M. The highly refined solution structure of the cytotoxic ribonuclease alpha-sarcin reveals the structural requirements for substrate recognition and ribonucleolytic activity. J. Mol. Biol. 2000, 299, 1061–1073. [Google Scholar] [CrossRef]
- Shi, W.W.; Mak, A.N.; Wong, K.B.; Shaw, P.C. Structures and Ribosomal Interaction of Ribosome-Inactivating Proteins. Molecules 2016, 21, 1588–1600. [Google Scholar] [CrossRef]
- Schlünzen, F.; Wilson, D.N.; Tian, P.; Harms, J.M.; McInnes, S.J.; Hansen, H.A.; Albrecht, R.; Buerger, J.; Wilbanks, S.M.; Fucini, P. The binding mode of the trigger factor on the ribosome: implications for protein folding and SRP interaction. Structure 2005, 13, 1685–1694. [Google Scholar] [CrossRef]
- Diaconu, M.; Kothe, U.; Schlünzen, F.; Fischer, N.; Harms, J.M.; Tonevitsky, A.G.; Stark, H.; Rodnina, M.V.; Wahl, M.C. Structural basis for the function of the ribosomal L7/12 stalk in factor binding and GTPase activation. Cell 2005, 121, 991–1004. [Google Scholar] [CrossRef]
- Liljas, A.; Sanyal, S. The enigmatic ribosomal stalk. Q Rev. Biophys. 2018, 51, e12. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, R.; Russo, R.; Landi, N.; Valletta, M.; Chambery, A.; Di Maro, A.; Bolognesi, A.; Ferreras, J.M.; Citores, L. Structure and Biological Properties of Ribosome-Inactivating Proteins and Lectins from Elder (Sambucus nigra L.) Leaves. Toxins (Basel) 2022, 14, 611–639. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, R.; Citores, L.; Ferreras, J.M. Ribosomal RNA N-glycosylase Activity Assay of Ribosome-inactivating Proteins. Bio Protoc. 2017, 7, e2180. [Google Scholar] [CrossRef] [PubMed]
- Citores, L.; Ragucci, S.; Russo, R.; Gay, C.C.; Chambery, A.; Di Maro, A.; Iglesias, R.; Ferreras, J.M. Structural and functional characterization of the cytotoxic protein ledodin, an atypical ribosome-inactivating protein from shiitake mushroom (Lentinula edodes). Protein Sci. 2023, 32, e4621. [Google Scholar] [CrossRef] [PubMed]
- Di Maro, A.; Ferranti, P.; Mastronicola, M.; Polito, L.; Bolognesi, A.; Stirpe, F.; Malorni, A.; Parente, A. Reliable sequence determination of ribosome- inactivating proteins by combining electrospray mass spectrometry and Edman degradation. J. Mass Spectrom. 2001, 36, 38–46. [Google Scholar] [CrossRef]
- Keil, B. ; Springer-Verlag Springer Science & Business Media, 2012; pp. 335.
- Walker, V.; Taylor, W.H. Ovalbumin digestion by human pepsins 1, 3 and 5. Biochem. J. 1978, 176, 429–432. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A. , et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef]
- Goddard, T.D.; Huang, C.C.; Meng, E.C.; Pettersen, E.F.; Couch, G.S.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Meeting modern challenges in visualization and analysis. Protein Sci. 2018, 27, 14–25. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera -a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
Figure 1.
Purification of eryngitin 3 and 4. (a), SDS-PAGE analysis of eryngitin 3 and 4 isolated from P. eryngii fruiting bodies. SDS-PAGE was carried out in 15% polyacrylamide separating gel. M, molecular markers. A total of 3.0 µg protein was loaded. Eryngitin 3 and 4 were analysed: i) under reducing conditions (+SH; lanes 1 and 2, respectively); ii) without reducing agent (-SH; lanes 3 and 4, respectively); and iii) after alkylation with 4-vinylpyridine without reducing agent (-SH; lanes 5 and 6, respectively). (b) and (c), RP-HPLC profiles of native eryngitin 3 and 4, respectively. Typically, 100 µg of protein were injected. For a and b component information in panel B, see paragraph 3.3 in main text.
Figure 1.
Purification of eryngitin 3 and 4. (a), SDS-PAGE analysis of eryngitin 3 and 4 isolated from P. eryngii fruiting bodies. SDS-PAGE was carried out in 15% polyacrylamide separating gel. M, molecular markers. A total of 3.0 µg protein was loaded. Eryngitin 3 and 4 were analysed: i) under reducing conditions (+SH; lanes 1 and 2, respectively); ii) without reducing agent (-SH; lanes 3 and 4, respectively); and iii) after alkylation with 4-vinylpyridine without reducing agent (-SH; lanes 5 and 6, respectively). (b) and (c), RP-HPLC profiles of native eryngitin 3 and 4, respectively. Typically, 100 µg of protein were injected. For a and b component information in panel B, see paragraph 3.3 in main text.
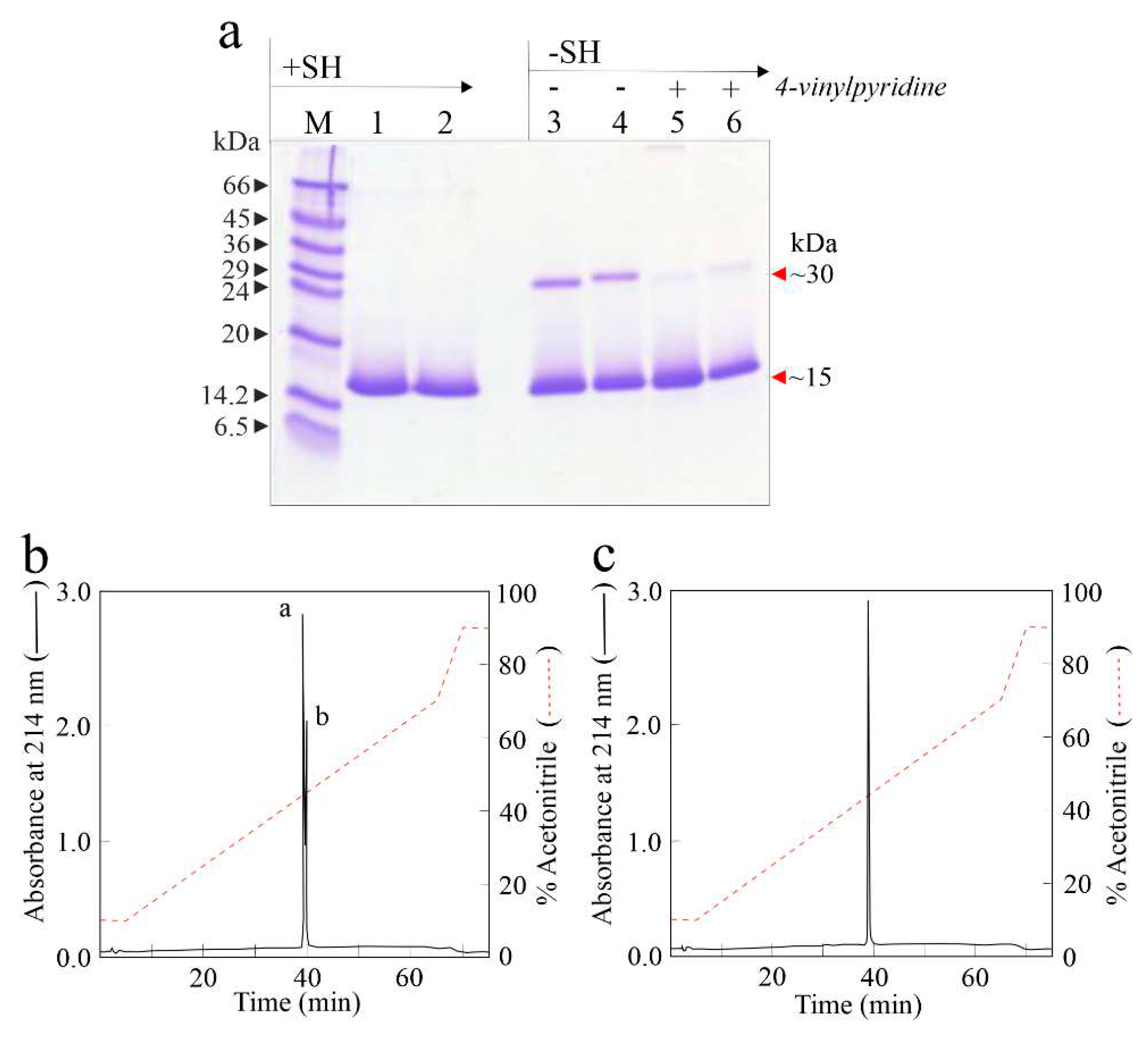
Figure 2.
Biological activities of eryngitin 3 and 4. (a), effect of eryngitin 3 (black circles) and eryngitin 4 (white circles) on protein synthesis. Translation assays were carried out using a cell free system, as indicated in Material and methods. (b) and (c), ribonucleolytic action of eryngitin 3 (lane 1 in panel b and lane 2 in panel c) and eryngitin 4 (lane 2 in panel b and lane 3 in panel c) in yeast and mealworm ribosomes, respectively, compared to that of α-sarcin (lane 3 in panel b and lane 1 in panel c). Each lane contained 5.0 µg (b) or 3.0 µg (c) of RNA isolated from ribosomes untreated (C, control) or treated with the toxin. The red arrows indicate the RNA α-fragment released as a result of the action of toxins. Numbers indicate the size of the markers (M) in nucleotides.
Figure 2.
Biological activities of eryngitin 3 and 4. (a), effect of eryngitin 3 (black circles) and eryngitin 4 (white circles) on protein synthesis. Translation assays were carried out using a cell free system, as indicated in Material and methods. (b) and (c), ribonucleolytic action of eryngitin 3 (lane 1 in panel b and lane 2 in panel c) and eryngitin 4 (lane 2 in panel b and lane 3 in panel c) in yeast and mealworm ribosomes, respectively, compared to that of α-sarcin (lane 3 in panel b and lane 1 in panel c). Each lane contained 5.0 µg (b) or 3.0 µg (c) of RNA isolated from ribosomes untreated (C, control) or treated with the toxin. The red arrows indicate the RNA α-fragment released as a result of the action of toxins. Numbers indicate the size of the markers (M) in nucleotides.
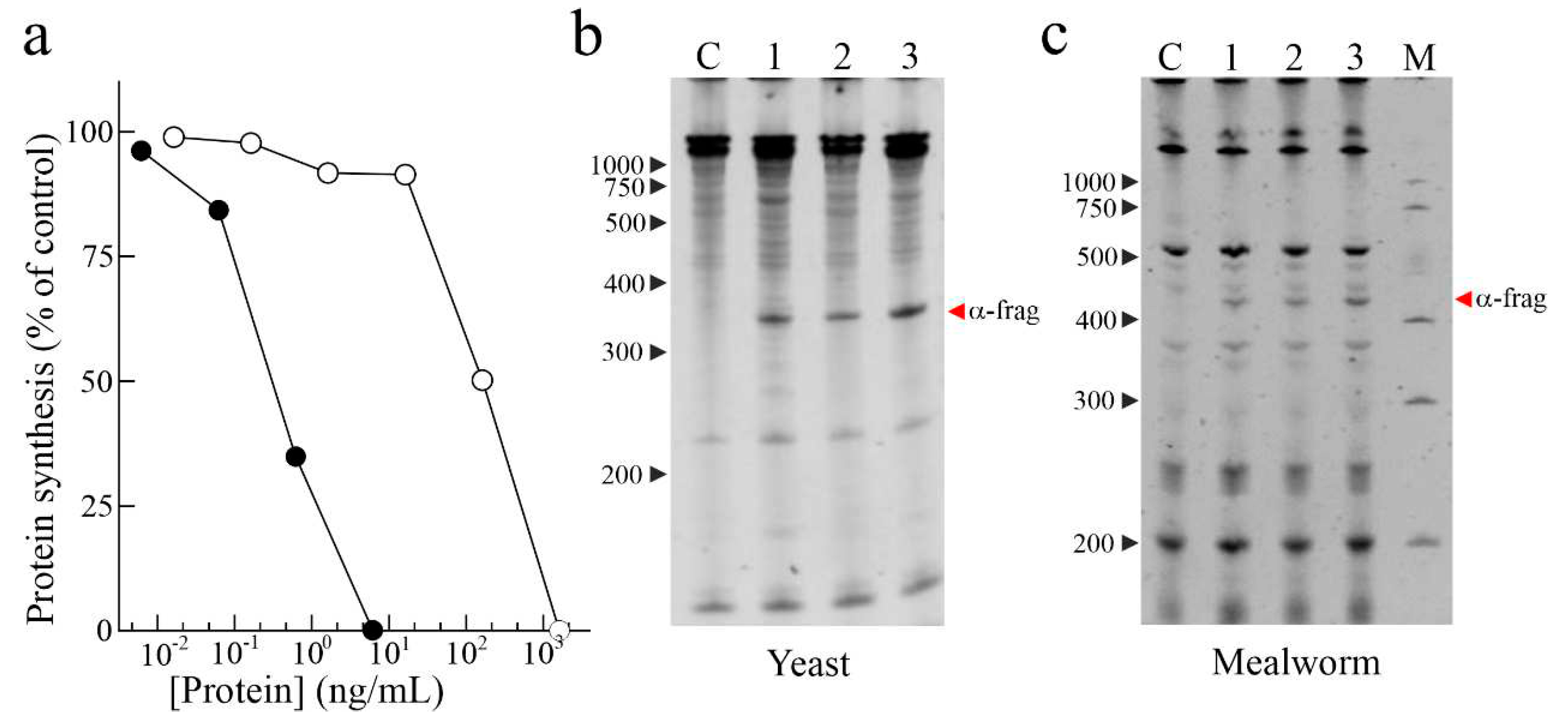
Figure 3.
Cytotoxic effect of eryngitin 3 and 4 against human malignant cells. (a) and (b), effect of eryngitin 3 or eryngitin 4 on the viability of HeLa and COLO 320 cells, respectively. Cells were grown in RPMI 1640 medium and incubated with different eryngitins concentrations for 48 h and 72 h, and cell viability was evaluated by a colorimetric assay. To investigate the effect of Z-VAD on the viability of HeLa cells, the cells were preincubated for 3 h with Z-VAD and then incubated with different concentrations of eryngitin 3 or 4 for 48 h, and cell viability was evaluated. Data represent the mean ± SD of two experiments performed in duplicate. (c), ribonucleolytic action of eryngitin 3 on RNA from HeLa cells. Each lane contained 1.0 μg of RNA isolated from either untreated cells (C, control) or cells incubated with 30 nM of α-sarcin (lane 1) or eryngitin 3 (lane 2) for 72 h. Red arrow indicates the α-fragment released as a result of specific ribonuclease action. Numbers indicate the size of the standards (M) in nucleotides. (d), effect of eryngitin 3 on internucleosomal DNA fragmentation. COLO 320 cells were incubated in the absence (lane 1) or presence of 0.3 µM of eryngitin 3 (lane 2) for 72 h. The DNA was isolated, and 4.0 µg was electrophoresed, as indicated in Material and methods. The numbers indicate the corresponding size of the standards (M) (λDNA HindIII/EcoRI) in Kb.
Figure 3.
Cytotoxic effect of eryngitin 3 and 4 against human malignant cells. (a) and (b), effect of eryngitin 3 or eryngitin 4 on the viability of HeLa and COLO 320 cells, respectively. Cells were grown in RPMI 1640 medium and incubated with different eryngitins concentrations for 48 h and 72 h, and cell viability was evaluated by a colorimetric assay. To investigate the effect of Z-VAD on the viability of HeLa cells, the cells were preincubated for 3 h with Z-VAD and then incubated with different concentrations of eryngitin 3 or 4 for 48 h, and cell viability was evaluated. Data represent the mean ± SD of two experiments performed in duplicate. (c), ribonucleolytic action of eryngitin 3 on RNA from HeLa cells. Each lane contained 1.0 μg of RNA isolated from either untreated cells (C, control) or cells incubated with 30 nM of α-sarcin (lane 1) or eryngitin 3 (lane 2) for 72 h. Red arrow indicates the α-fragment released as a result of specific ribonuclease action. Numbers indicate the size of the standards (M) in nucleotides. (d), effect of eryngitin 3 on internucleosomal DNA fragmentation. COLO 320 cells were incubated in the absence (lane 1) or presence of 0.3 µM of eryngitin 3 (lane 2) for 72 h. The DNA was isolated, and 4.0 µg was electrophoresed, as indicated in Material and methods. The numbers indicate the corresponding size of the standards (M) (λDNA HindIII/EcoRI) in Kb.
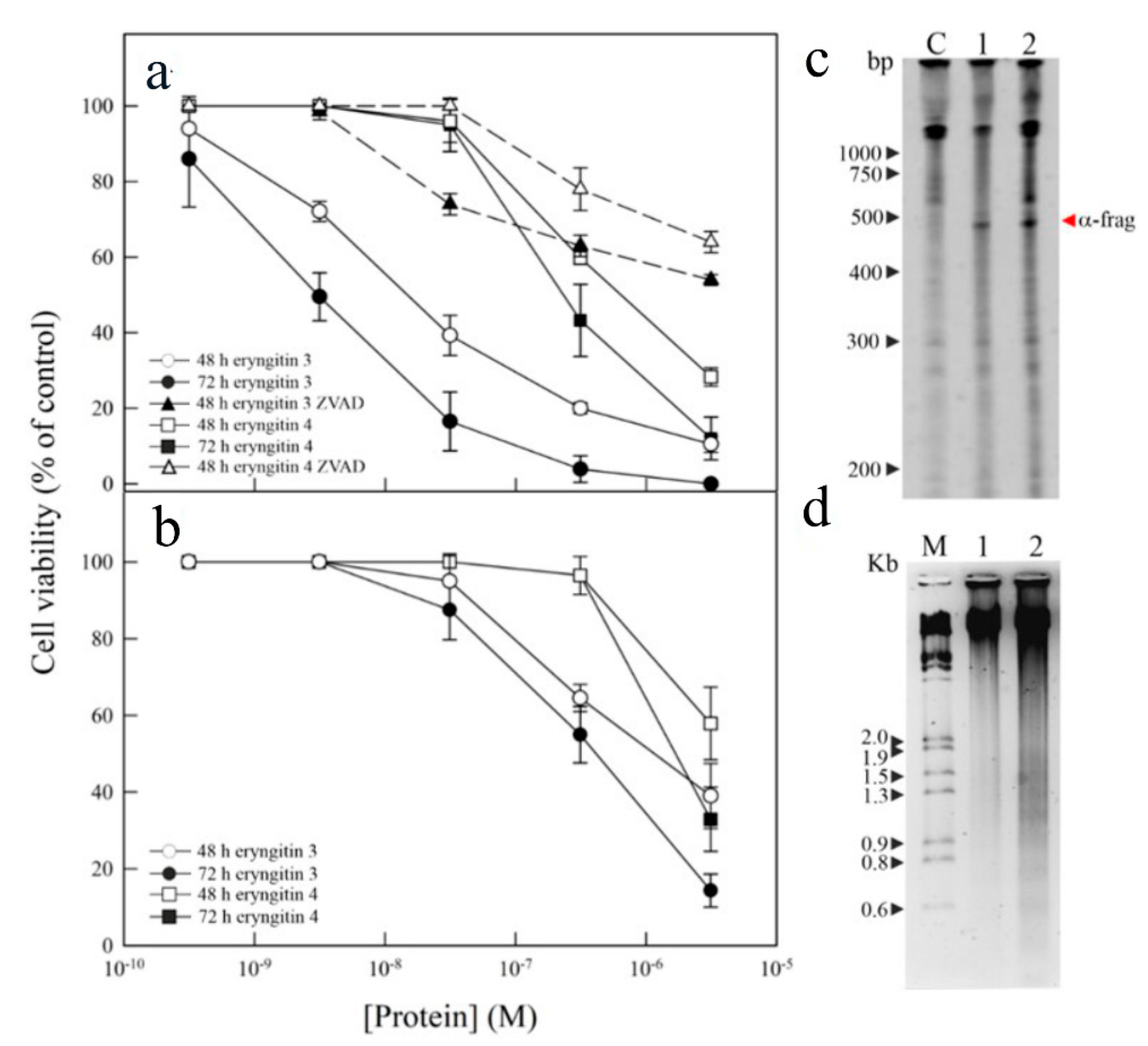
Figure 4.
Antifungal activity. The effect of eryngitin 3 (a) and 4 (b) against Penicillium digitatum was measured in a microtiter plate bioassay. Conidia of P. digitatum were grown in Potato Dextrose Broth (PDB) at 28 oC, in the presence of different eryngitins concentrations. Fungal growth was followed for 84 h and measured as an increase in absorbance at 650 nm. The curves represent the buffer control or different amounts (µg/mL) of both toxins. (c), Morphological changes in P. digitatum mycelium exposed to eryngitins. The P. digitatum mycelium was grown in the absence (control) or presence of 60 μg/mL eryngitin 3 or 98 μg/mL eryngitin 4. After incubation for 48 h, samples were visualized using light microscopy at 200× magnification.
Figure 4.
Antifungal activity. The effect of eryngitin 3 (a) and 4 (b) against Penicillium digitatum was measured in a microtiter plate bioassay. Conidia of P. digitatum were grown in Potato Dextrose Broth (PDB) at 28 oC, in the presence of different eryngitins concentrations. Fungal growth was followed for 84 h and measured as an increase in absorbance at 650 nm. The curves represent the buffer control or different amounts (µg/mL) of both toxins. (c), Morphological changes in P. digitatum mycelium exposed to eryngitins. The P. digitatum mycelium was grown in the absence (control) or presence of 60 μg/mL eryngitin 3 or 98 μg/mL eryngitin 4. After incubation for 48 h, samples were visualized using light microscopy at 200× magnification.
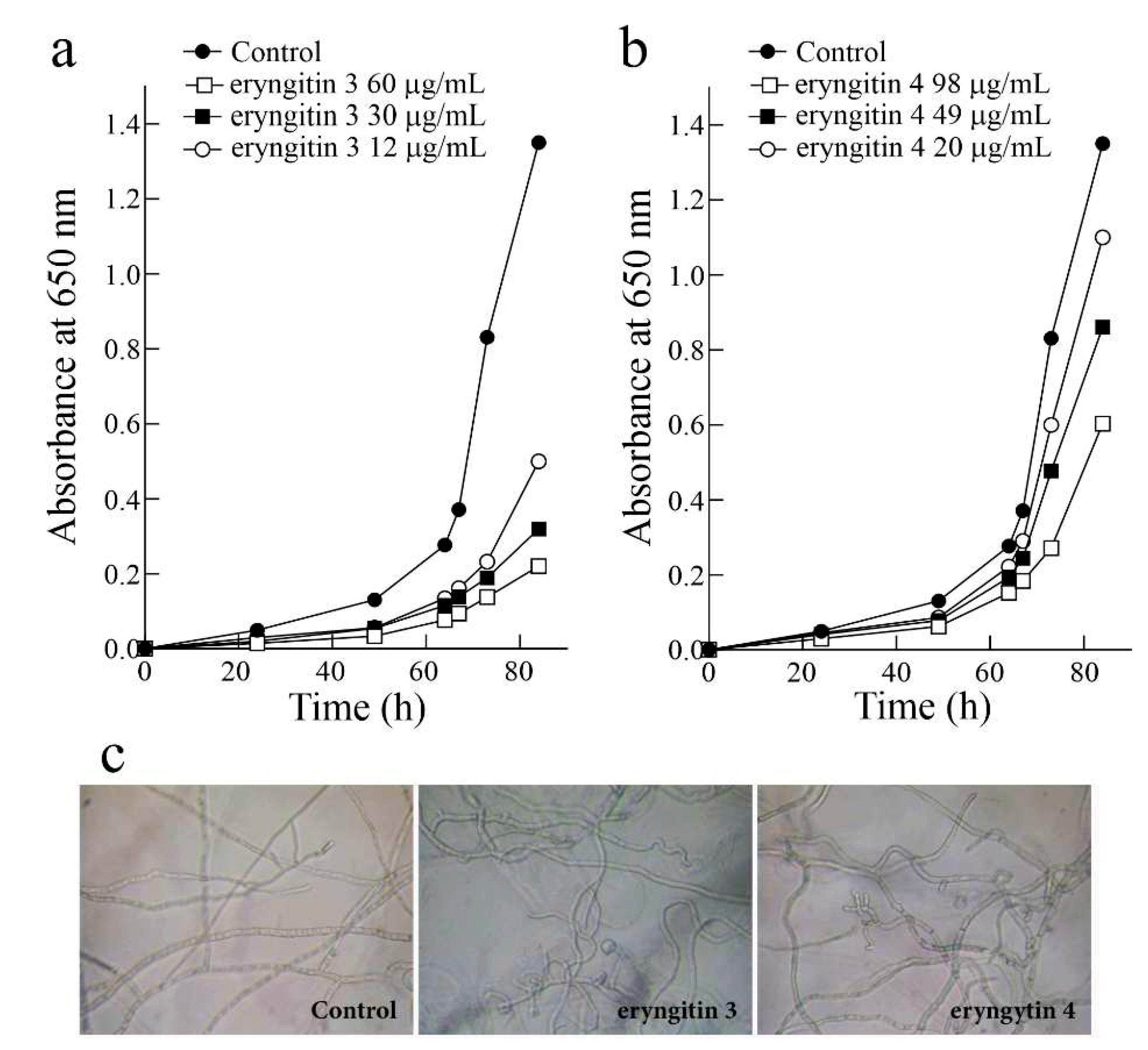
Figure 5.
MALDI-ToF mass spectra of native and alkylated eryngitin 3 and 4. (a) and (b), MALDI-ToF spectra of native eryngitin 3 and 4, respectively. (c) and (d), MALDI-ToF spectra of alkylated eryngitin 3 and 4, respectively. [M+H+]+ and [M+2H+]2+ correspond to the single and doubly charged ions, respectively.
Figure 5.
MALDI-ToF mass spectra of native and alkylated eryngitin 3 and 4. (a) and (b), MALDI-ToF spectra of native eryngitin 3 and 4, respectively. (c) and (d), MALDI-ToF spectra of alkylated eryngitin 3 and 4, respectively. [M+H+]+ and [M+2H+]2+ correspond to the single and doubly charged ions, respectively.
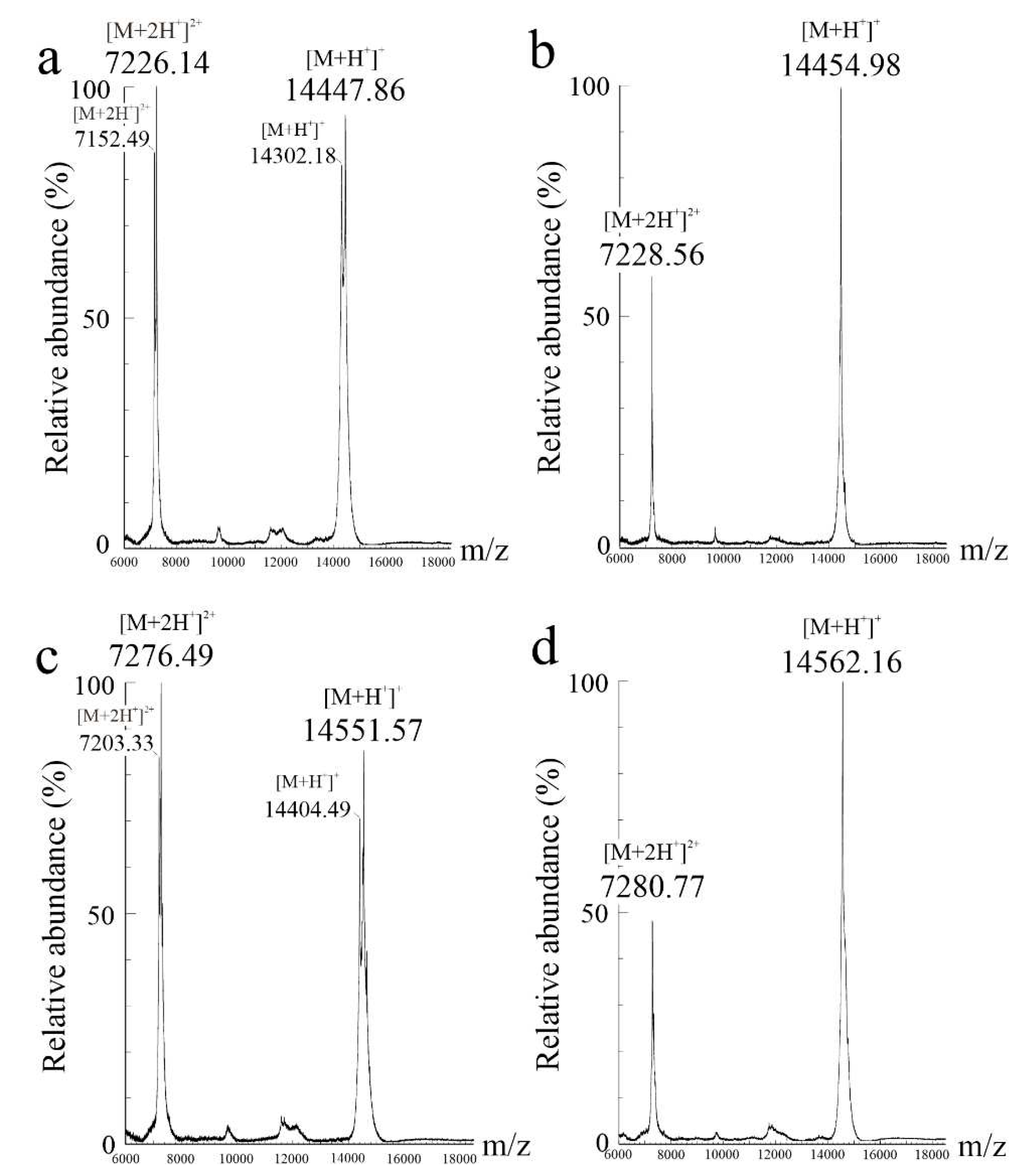
Figure 6.
Amino acid sequence of eryngitin 4 and 3. The overlapping peptides obtained by digestions of alkylated eryngitin 4 (a) and eryngitin 3 (b) used to confirm the amino acid sequence of uncharacterized protein from Pleurotus eryngii ATCC 90797 (AC: KAF9489889.1) are reported. Grey, N- and C-terminal additional amino acid residues found in reference protein retrieved in P. eryngii genome (Figure S1). The standard one-letter code was used for the amino acid residues. Catalytic residues are highlighted in red, free cysteinyl residues are in violet. Abbreviations: C, GC, P and T, chymotrypsin, endoproteinase Glu-C, pepsin and tryptic peptides, respectively.
Figure 6.
Amino acid sequence of eryngitin 4 and 3. The overlapping peptides obtained by digestions of alkylated eryngitin 4 (a) and eryngitin 3 (b) used to confirm the amino acid sequence of uncharacterized protein from Pleurotus eryngii ATCC 90797 (AC: KAF9489889.1) are reported. Grey, N- and C-terminal additional amino acid residues found in reference protein retrieved in P. eryngii genome (Figure S1). The standard one-letter code was used for the amino acid residues. Catalytic residues are highlighted in red, free cysteinyl residues are in violet. Abbreviations: C, GC, P and T, chymotrypsin, endoproteinase Glu-C, pepsin and tryptic peptides, respectively.
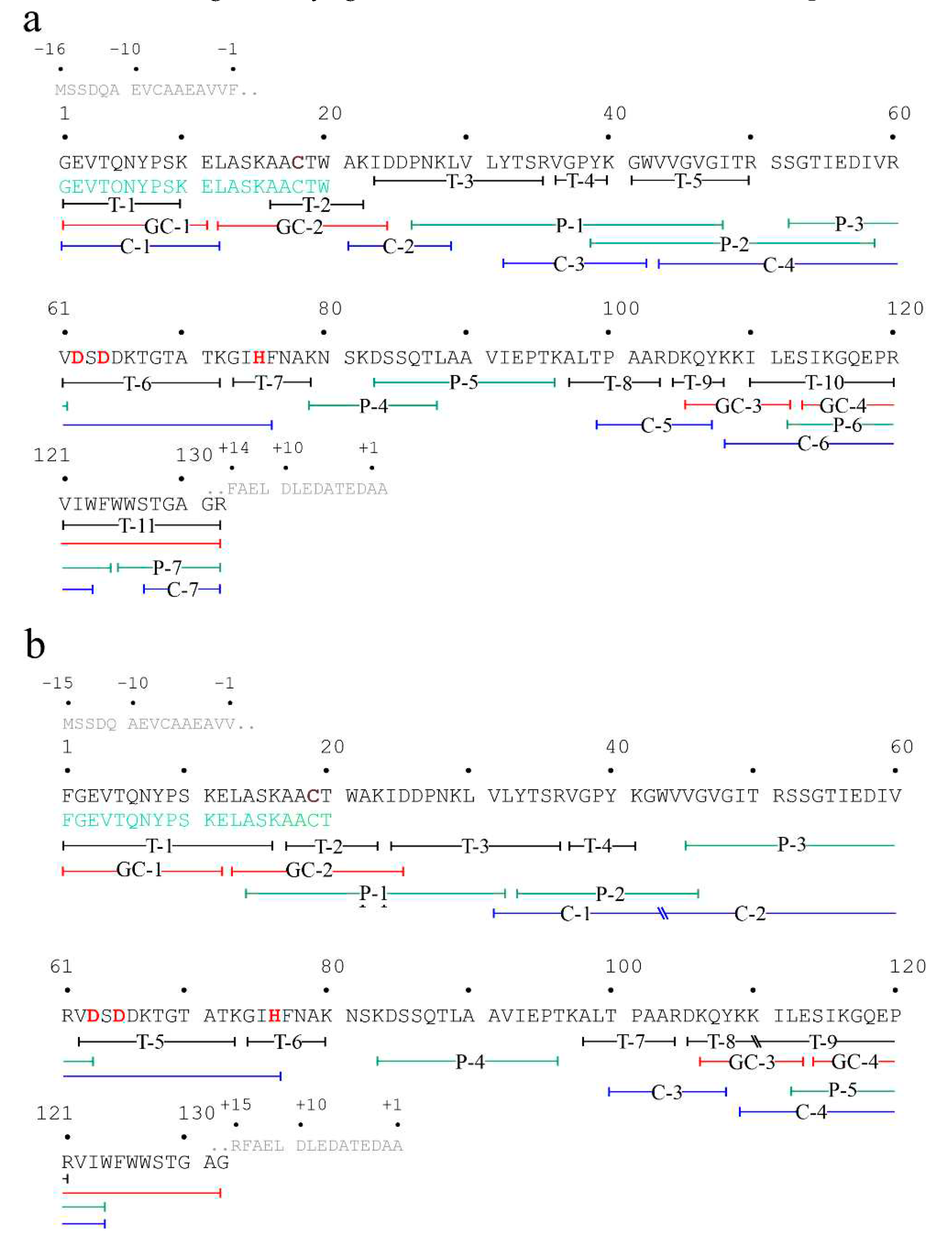
Figure 7.
Structural features of eryngitin 3 and 4 by 3D-molecular model. (a), superimposed 3D molecular model of eryngitin 4 (cyan ribbon) and eryngitin 3 (orange ribbon) obtained by AlphaFold software. Stick representation highlights the catalytic triad (Asp62, Asp64 and His75, numbering referred to eryngitin 4 amino acid sequence). (b) and (c), electrostatics surfaces of eryngitin 4 and 3, respectively. Blue and red show positive and negative surface regions, respectively. Yellow and green arrows highlight the N- and C-terminal region of the two proteins, respectively. The disposition of 3D models in (b) and (c) is in the same molecular orientation of the superimposed models showed in (a).
Figure 7.
Structural features of eryngitin 3 and 4 by 3D-molecular model. (a), superimposed 3D molecular model of eryngitin 4 (cyan ribbon) and eryngitin 3 (orange ribbon) obtained by AlphaFold software. Stick representation highlights the catalytic triad (Asp62, Asp64 and His75, numbering referred to eryngitin 4 amino acid sequence). (b) and (c), electrostatics surfaces of eryngitin 4 and 3, respectively. Blue and red show positive and negative surface regions, respectively. Yellow and green arrows highlight the N- and C-terminal region of the two proteins, respectively. The disposition of 3D models in (b) and (c) is in the same molecular orientation of the superimposed models showed in (a).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



