
Submitted:
22 August 2023
Posted:
24 August 2023
You are already at the latest version
Abstract
cGAS is a cytosolic DNA sensor that activates innate immune responses through production of the second messenger 2’3’-cGAMP, which activates the adaptor STING. cGAS senses dsDNA in a length-dependent but sequence-independent manner, and it cannot discriminate self-DNA from foreign DNA. In normal physiological conditions, cellular DNA is sequestered in the nucleus by nuclear envelope and in mitochondria by mitochondrial membrane. When self-DNA leaks into the cytosol during cellular stress, or during mitosis, the cGAS can be exposed to self-DNA and be activated. Recently, many studies have investigated how cGAS keeps inactive and avoids to be aberrantly activated by self-DNA. Thus, the aim of this narrative review is to summarize the mechanisms by which cGAS avoids to sense self-DNA under normal physiological conditions.
Keywords:
cGAS
; self-DNA
; mitosis
; autoimmune diseases
; DNases
1. Introduction
Innate immunity is the first immunological, non-specific mechanism for fighting against infections and other aggressions. The initial sensing of infection and injury is mediated by pattern recognition receptors (PRRs), which recognize pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs). Excessively accumulated cytosolic DNA can act as a PAMP or DAMP, and the presence of DNA in the cytoplasm is normally a sign of microbial infection or tissue damage [1]. The appearance of naked DNA in the cytoplasm of mammalian cells triggers a cellular response initiated by the DNA sensing pathway. Recently, cyclic GMP-AMP synthase (cGAS) was characterized as a primary cytosolic DNA sensor that triggers type I interferons (IFNs) and inflammatory cytokines upon binding dsDNA [2,3].
cGAS can be activated by dsDNA in a length-dependent but sequence-independent manner through binding the DNA phosphate backbone [4]. Thus, cGAS is activated not only by non-self DNA, such as DNA from DNA viruses or retroviruses, intracellular bacteria and protozoa, but also by self-DNA including mitochondrial and nuclear DNA that gain access to the cytoplasm. When self-DNA leaks into the cytoplasm during cellular stress (such as mitochondrial alteration, DNA damage, mitotic arrest, or senescence), or is present as cytosolic micronuclei, cGAS is activated leading to a state of sterile inflammation [5,6]. In the case of microbial infection, this is an important protective component of innate immunity, but in the case of exposure to self-DNA, it can result in autoimmune diseases [7].
In normal physiological conditions, cellular DNA is isolated in the nucleus and mitochondria through the nuclear envelope and the mitochondrial membrane, respectively [8]. However, the nuclear envelope is a highly dynamic structure that is disassembled and reassembled during mitosis in higher eukaryotes [9]. It is not surprising that the cGAS could be directly exposed to self-DNA during mitosis when the nuclear envelope is disassembled [9]. This has raised a question: When cGAS is exposed to self-DNA, how it keeps inactive? Thus, the aim of this narrative review is to summarize the mechanisms by which cGAS avoids to sense self-DNA and keeps inactive under normal physiological conditions.
2. Structural Domains and Modification Sites of cGAS
cGAS (also known as C6orf150, or male abnormal 21 domain containing 1 ) is located on chromosome 6q13 and encodes a protein with 522 amino acids in human [10]. The cGAS protein (human) is composed of an unstructured and not well-conserved N-terminus (1-160) and a highly conserved C terminus (161-522 ) that contains a nucleotidyltransferase (NTase) core domain (161-330) and the male abnormal 21 (Mab21) domain (213-513) [11]. The alignment of the amino acid sequence of cGAS within different mammalian species further suggested that the cGAS N-terminal domain (1-160) is highly diverse and disordered [12]. This less evolutionarily conserved sequence also plays a key role in determining nuclear, cytoplasmic distribution, sensing nuclear chromatin, binding to immune stimulatory DNA (ISD), plasma membrane, or centromere as well as assembly of lipid phase condensation [13]. Recent studies have shown that the serine (13, 37, 64, 129 and 143) residue in cGAS N-terminus are crucial for sensing genomic/chromatin DNA [13]. The C-terminal NTase domain contains several conserved amino acid residues within the NTase superfamily including G212, S213, E225, D227 and D319, which are critical for the enzyme activity of cGAS [14]. The Mab21 domain harbors the conserved Zn finger motif (H390-C405), which has been shown to be functionally important to scale the specificity of cGAS toward dsDNA [12].
The protein of cGAS contains many modification sites, which can be regulated by various post-translational modifications including phosphorylation, ubiquitination, sumoylation, and acetylation [15] (Figure 1). Recently, several studies have demonstrated that cGAS could be hyperphosphorylated in huamn at serine, threonine and tyrosine residues including Ser13, Ser37, Ser64, Ser116, Ser129, Ser143, Thr69, Thr91, Tyr215 and Tyr242 by phosphorylase kinases such as serine/threonine-protein kinase (AKT), cyclin-dependent kinase-1 (CDK1), and Aurora kinase B (AKB) and B lymphocyte kinase (BLK) respectively [12,16]. Ubiquitination and deubiquitination have additional control on cGAS activation [17]. Several ubiquitin ligases including RNF185, RINCK, TRIM56, ARIH1 and MARCH8 were shown to catalyze human cGAS at Lys-173, Lys335, Lys-347, Lys-384 and Lys414 respectively [18,19]. Sumoylation also plays a critical role in regulating the activity of cGAS [20]. TRIM38 maintains sumoylation of Lys231 and Lys479 in human cGAS, which prevents K48-linked cGAS polyubiquitination and degradation [20]. Sumoylation at Lys-347, Lys-384 and Lys-394 prevents DNA-binding, oligomerization and nucleotidyltransferase activity [15]. The acetylation of cGAS can regulate cGAS activity either positively or negatively depending upon the acetylation sites [21,22]. Human cGAS was reported to be acetylated at Lys47, Lys56, Lys62, Lys83, Lys198, Lys384, Lys394, and Lys414 [21].
3. How is the cGAS Activated
3.1. DNA-Induced Conformational Changes in cGAS Lead to Its Activation
cGAS activation requires direct binding to dsDNA to form cGAS-dsDNA complexes, within which the catalytic site of cGAS is structurally rearranged to activate its enzymatic activity to synthesize 2‘3′-cGAMP [23]. cGAS interacts with the dsDNA through extensive electrostatic interactions and hydrogen bonds, by employing a positively charged surface as well as the zinc thumb to interact with the DNA sugar-phosphate backbone. cGAS can be activated by dsDNA in a length-dependent but sequence-independent manner [24]. Structure of cGAS-DNA complex shows that their interaction interfaces cover about 10 bp on dsDNA [25]. Biochemical assays have shown that short DNAs of ~20 bp can bind to cGAS, but longer dsDNAs of >45 bp can form more stable ladder-like networks of cGAS dimers, leading to stronger enzymatic activity [25].
cGAS alone is monomeric and does not have catalytic activity as a result of local structure destabilization of the NTase domain. DNA binding induces conformational changes in cGAS and crosslinks two cGAS molecules to form a 2:2 dimer or higher-order complexes, resulting in the activation of cGAS. cGAS has three DNA binding sites (A, B, and C) that interact with the sugar-phosphate backbone of dsDNA or RNA–DNA hydrides in a sequence-independent pattern [2]. Site A is the primary site that mediates DNA-induced conformational change of the activation loop in cGAS. Sites A and B together mediate the formation of the 2:2 cGAS DNA complex. Site C provides an additional interaction between cGAS and DNA, contributing to the phase separation of the cGAS-DNA complex [26]. Interestingly, although ssDNA and dsRNA can bind cGAS, both fail to rearrange the cGAS catalytic pocket for cGAS activation [27].
3.2. Liquid-Liquid Phase Separation Can Enhance the Activation of cGAS
Liquid-liquid phase separation (LLPS) is a process by which bio-macromolecules, particularly proteins, condense into a dense phase that resembles liquid droplets [28]. LLPS is a concentration and environment dependent condensation process driven by solute–solute interactions that energetically overcome solute–solvent interactions [29]. In a liquid-like state, the condensed phase frequently exchanges materials with the dilute phase, and this liquid-like property has an important role in defining the composition and biochemical activity of molecules in the condensed phase [29].
Both in vitro and in vivo studies have shown that interactions between cGAS and DNA result in high-order oligomerization and formation of LLPS [30]. DNA binding to cGAS induces a robust phase transition to liquid like droplets, which could serve as a micro-reactor to accelerate 2′3′-cGAMP production by increasing local concentrations of proteins and reactants [30,31]. Additionally, cGAS forms liquid-like condensates with double-stranded DNA (dsDNA) to enhance the production of 2′3′-cyclic GMP–AMP (cGAMP) by protecting DNA from degradation by the exonuclease TREX1 [32]. This mechanism allows cGAS to detect the presence of DNA in the cytoplasm above a certain threshold to trigger a switch like response. Further studies also demonstrated that mutation and truncation of cGAS and short DNA (< 45 bp) attenuate the oligomerization and LLPS of cGAS-DNA, resulting in reduced or even eliminated cGAS activity [33,34].
3.3. Divalent Cations Substantially Promote the Activity of cGAS
Metal ions are essential for the functionality of a plethora of proteins [35]. In fact, about one-third of all known enzymes require particular metal co-factors, which serve catalytic, structural, or regulatory functions [35]. cGAS belongs to the NTase superfamily which catalyzes nucleophilic substitution reactions and is divalent cation-dependent [36,37]. The catalytic core structures of different NTases usually share a common structural fold and similar active sites harboring a highly conserved catalytic triad hG[GS], [DE]h[DE]h, h[DE]h (where h indicates a hydrophobic amino acid) for catalytic metal coordination [36]. The binding between cGAS and DNA involves extensive ionic interactions between the positively charged surfaces of cGAS and negatively charged DNA [30]. Such interactions are vulnerable to cytoplasmic salt concentrations. cGAS enzyme activity was much weaker in an assay with physiological buffer than in an assay with low-salt buffer [30]. Several studies have shown that divalent metal ions including zinc (Zn2+), manganese (Mn2+), magnesium (Mg2+) all play important roles in regulating the activation of cGAS [38,39].
Zn2+ could facilitate cGAS activation in cells by promoting cGAS phase transition in the presence of cytosolic DNA [38]. Measurements of free Zn2+ concentrations revealed that Zn2+ bound to cGAS but not to DNA. When L929 cells were depleted of zinc with the zinc-specific chelator TPEN, and the production of 2′3′-cGAMP was decreased in a TPEN concentration–dependent manner after transfecting with HT-DNA [30]. Furthermore, a study has indicated that Mn2+ could directly activate the cGAS, and that DNA is dispensable for Mn2+ induced cGAS activation [36]. Structural analysis reveals that Mn2+ activated cGAS undergoes globally similar conformational changes to DNA-activated cGAS, but forms a unique helix to widen the catalytic pocket, allowing substrate entry and 2′3′-cGAMP synthesis [36]. Further, Mn2+ improves the sensitivity of cGAS to intracellular DNA and its catalytic activity, which can promote cGAS to generate 2′3′-cGAMP under the stimulation of low concentration of DNA [39]. Mn-deficient mice produced decreased amounts of cytokines and were more vulnerable to DNA viruses [36]. Structural studies of ternary complexes of dsDNA-bound cGAS have shown that the triphosphate moieties of ATP and 5′-pppG(2′,5′)pG are coordinated to Mg2+ [36]. Incubation of human cGAS protein with ATP, GTP, Mg2+, and dsDNA could result in the induction of 2′3′-cGAMP [36].
4. How Does cGAS Avoid to Sense Self-DNA under Normal Conditions?
4.1. Self-DNA Is Cleared by the DNases
The deoxyribonucleases (DNases) has been suggested to degrade self-DNA under normal conditions to prevent aberrant activation of cGAS-mediated immune responses [40]. DNases maintain cytosolic DNA levels under the threshold of cGAS activation to retain immune silence. Three different DNases including DNase I, DNase II, and TREX1 (or DNase III) have been identified to degrade self double stranded DNA (dsDNA). DNase I is localized in serum where it degrades the chromatin released from dead cells and prevents autoimmune diseases [41]. DNase II is localized in the lysosomes and is largely responsible for the clearance of DNA from dead cells and expelled nuclei [42]. TREX1, localized in cytosol, is a major mammalian 3′-5′ DNA-exonuclease preventing endogenous DNA accumulation [43].
Dysfunction of DNases can lead to the accumulation of DNA in the cytoplasm, and therefore activation of cGAS-mediated immune responses (Figure 2). Loss-of-function mutations of TREX1 have been identified in human patients with autoimmune disorders such as Aicardi-Goutières syndrome (AGS) and lupus-like autoimmune disorders [44,45]. It has suggested that cGAS pathway activation is the key signaling for autoimmunity diseases caused by a TREX1 missense mutation [44]. A study has indicated that the deficiency of TREX1 could result in an aberrant accumulation of self-DNA in the cytosol, which could predominantly activate the innate immune response via the cGAS pathways [46]. Another in vivo study has suggested that genetic deletion of Trex1 or DNase II could lead to lethal autoimmune diseases in mice [47].
4.2. Plasma Membrane Localization of cGAS Prevents Recognition of self-DNA
cGAS is thought to diffuse throughout the cytosol for searching its DNA ligand. However, a study has indicated that cGAS is not a cytosolic protein but is rather a peripheral membrane protein that primarily resides on the plasma membrane [48]. Analysis of the distribution of cGAS in human THP1 monocyte has suggested that cGAS is a membrane protein that resides primarily at the surface of THP1 cells in its inactive state [48]. Within 30 min of DNA transfection, cGAS was no longer concentrated at the cell surface but was rather detected in various cytoplasmic puncta [48]. The unstructured N-terminus may play a role for cGAS plasma membrane attachment to restrain cGAS activation [48]. The N-terminus of cGAS in humans has a phosphoinositide-binding domain and interacts selectively with phosphatidylinositol 4,5-bisphosphate (PI(4,5)P2) in the plasma membrane [48]. The amino acids 64 to 75 of cGAS are important for its plasma membrane localization, and within this region two arginine residues are conserved or charge-conserved among humans, mice, and several other mammalian species [48].
Plasma membrane localization of cGAS is important for preventing recognition of self-DNA. In resting cells, its N terminus positions cGAS at the cell surface, where it is least likely to detect self-DNA and thus prevent aberrant activation [48]. Upon DNA detection, the N-terminus may release from PI(4,5)P2 and facilitate liquid droplet formation and signaling in the cytosol [48]. The N-terminal domain of cGAS is necessary and sufficient for cGAS localization to the plasma membrane and this interaction restrains cGAS activity from self-DNA [49]. Deleting the N-terminal localization domain from cGAS leads to heightened sensitivity to genotoxic stress [50]. Thus, localizing to the plasma membrane is an excellent strategy for cGAS to avoid inappropriate self-DNA detection (Figure 2).
4.3. Binding to Histones Prevents cGAS from Sensing Self-DNA during Mitosis
cGAS was first proposed as cytosolic DNA sensor, thereby, compartmentalization of self-DNA in mitochondria and nucleus is thought to be essential for cGAS to discriminate non-self and self-DNA. However, recent studies have indicated that cGAS is not only located in the cytoplasm and plasma membrane, but also found within the nucleus [4,13]. It been shown that cGAS is tightly associated with chromatin during mitosis, a phase of the cell cycle when the nuclear envelope breaks down [16]. The tight association of cGAS with chromatin during mitosis raises an important question: How cGAS activity is regulated during the cell cycle (Figure 3).
One study indicated that cGAS is tethered to the chromatin through binding to histones that form the nucleosome core, and this interaction can inhibit the activation of cGAS [51]. Another study suggested that nucleosomes could interfere with cGAS activation by nucleosome-free regions, and nucleosomes have a higher affinity for cGAS than naked DNA [52]. The interaction between cGAS and the nucleosome prevents cGAS from further binding to dsDNA, and abolishes the dimerization of cGAS [53]. The structure of cGAS–nucleosome complexes has shown that cGAS interacts with the H2A–H2B heterodimer through the region around site B, and contacts the DNA from the adjacent nucleosome via residues within site C [53]. Thus, the dsDNA binding sites B and C are occupied by the bound nucleosomes. Although site A does not directly contact the nucleosome, it is not accessible to dsDNA due to steric hindrance. Therefore, cGAS is locked in its inactive state by the binding of nucleosomes, and this binding disrupts cGAS dimerization and prevents further dsDNA binding (Figure 3). Both the residues R236 and R255 were shown to be critical for binding of cGAS to the nucleosomal acidic patch formed by H2A-H2B, and play an important role in the inhibitory effect of nucleosomes on cGAS activity [53].
4.4. Binding to Barrier-to-Autointegration Factor 1 Restricts cGAS to Sense self-DNA during Mitosis
Excepting the H2A-H2B heterodimer, the barrier-to-autointegration factor (BAF) was also shown to compete with cGAS for access to DNA in the nucleus, thereby preventing sensing of host DNA [54]. BAF, a chromatin-binding protein that is essential for nuclear membrane reformation at the end of mitosis, is involved in multiple pathways including mitosis, nuclear assembly, viral infection, chromatin and gene regulation and the DNA damage response [55]. The unique DNA-binding properties of BAF are likely fundamental to its roles and BAF forms homodimers, each subunit of which binds double-stranded DNA in a sequence-independent manner [55]. Each BAF monomer has a helix-hairpin-helix DNA-binding domain, allowing BAF dimers to bridge two strands of DNA either intra-molecularly or inter-molecularly [56].
BAF exerts an important regulatory function over cGAS, and defects in BAF can trigger innate immune activation [54]. BAF-mediated suppression of the cGAS pathway is necessary for reactivation of Kaposi sarcoma-associated herpesvirus (KSHV) and Epstein-Barr virus (EBV) [54,57]. Inhibiting BAF expression in latently infected, reactivating, or uninfected cells leads to increased type I interferon-mediated antiviral responses and decreased viral replication [57]. Ablation of BAF by gene editing resulted in chromatin activation near host defense genes with associated increased expression of ISGs, including OAS2, Rsad2 (viperin), IFIT1, and ISG15 [57]. Down-regulation of BAF triggered a robust ISG response, whereas suppression of other relevant genes had no effect [54]. Rather than passively interfering with cGAS activity by blocking DNA binding sites, BAF dynamically displaces transiently bound cGAS monomers from dsDNA [54]. Dynamic competition by BAF at the nuclear periphery is a critical strategy used by the host to reconcile the advantages of maintaining a universal DNA recognition machinery with routine operations occurring within a living cell [54] (Figure 3).
4.5. The Activity of cGAS Is Suppressed via Phosphorylation during Mitosis
cGAS is hyperphosphorylated in the nucleus, and the hyperphosphorylation can suppress the activity of cGAS, with the cGAS hyperphosphorylation mediated by Aurora kinase B (AurB), CDK1-cyclin B complex and other kinases [16,58] (Figure 3). Human cGAS is hyperphosphorylated by AurB at the N-terminal serine and threonine residues including Ser13, Ser37, Ser64, Thr69, Thr91, Ser116, Ser129, and Ser143 [16]. Parallel reaction monitoring (PRM) showed that phosphorylation of the N-terminal serine and threonine residues in cGAS increased dramatically in mitotic cells compared with asynchronized cells [16]. Treating BJ-5ta cells with Aurora kinase inhibitors (AMG-900, MLN-8237, or AZD-1152) after cells exited the G2/M arrest, the hyperphosphorylation level of cGAS was reduced by aurora kinase inhibitors in a dose-dependent manner [16]. Like chemical inhibitors, siRNA-mediated knockdown of AurB also decreased cGAS hyperphosphorylation [16]. The hyperphosphorylation at the N-terminus could prevent cGAS phase separating into liquid droplets where cGAS can efficiently synthesize 2′3′-cGAMP.
Furthermore, human cGAS could be phosphorylated by the CDK1-cyclin B complex at a highly conserved site Ser305 during the mitotic phase of the cell cycle [58] (Figure 3). Phosphorylation at this site could inhibit the ability of cGAS to synthesize 2′3′-cGAMP upon mitotic entry, leading to unresponsiveness to DNA-triggered innate immunity in mitotic cells [58]. The type 1 phosphatase PP1 dephosphorylates cGAS upon mitotic exit to enable its DNA sensing ability. The human cGAS mutant (S305A) of S305 within the nuclear localization sequence (295DVIMKRKRGGS305), is localized to chromosomes in mitotic cells similarly as wild-type cGAS or cGAS (S305D); thus, phosphorylation of cGAS S305 is not required for its chromosomal localization, but preventing its activation and inhibiting the ability of cGAS to synthesize 2′3′-cGAMP during mitosis [58].
5. Consequences of Self-DNA Induced cGAS Activation
5.1. Activation of cGAS by Self-DNA Can Cause Autoimmune Diseases
Accumulated evidence has shown that imbalance of innate immune system was the main contributor to autoimmune diseases, and acute inflammation induced by tissue damage or microbial infection [59]. Self-DNA sensed by the immune system has emerged as a key contributing response in the pathogenesis of autoimmune diseases [60]. Recently, many researches have revealed that the regulatory role of cGAS pathway in autoimmune diseases including Aicardi Goutières syndrome (AGS), systemic lupus erythematosus (SLE) and Rheumatoid arthritis (RA) [57,61,62].
cGAS is required for lethal autoimmune disease in the Trex1-deficient mouse model of AGS [63]. Trex1-deficient mice die within a few months after birth because of multi-organ inflammation, especially myocarditis, while deletion of just one allele of the gene encoding cGAS in TREX1-deficient mice largely rescues them from the lethal autoimmunity phenotype [46]. Systemic lupus erythematosus (SLE) is an autoimmune disease in which pathogenic autoantibodies are produced against nucleic acids and their interacting proteins, resulting in inflammation and tissue damage [64]. Apoptosis-derived membrane vesicles (AdMVs) in SLE serum induce type I IFN (IFN-I) production through activation of the cGAS pathway [65]. The production of IFN-I could further activate immune responses that lead to tissue damage in various organs, resulting in more generation of AdMVs, triggering a positive-feedback loop of IFN-I production and further tissue damage [65]. Thus, blockade of the cGAS-STING axis represents a promising therapeutic target for AGS and SLE.
5.2. Activation of cGAS by Self-DNA Is a Double-Edged Sword in Cancer
The ectopic cytosolic dsDNA may exert both anti-tumorigenic and pro-tumorigenic effects dependent on the specific context as well as stage of tumor progression [66,67]. Cancer cells exhibit genomic instability and chromosomal abnormalities that commonly result in formation of cytosolic chromatin fragments and micronuclei [68]. Subsequently, cGAS recognizes the DNA source and responds quickly to activate downstream cascade reaction to eliminate tumor [69]. An increasing number of studies have indicated that activation of cGAS by self-DNA can inhibit tumors by enhancing tumor immune surveillance, accelerating cellular senescence, and promoting apoptosis [70,71]. The tumor cells need to evade this signaling pathway detection to survive in the harsh living environment; thus, the cGAS-STING axis was observed to be disrupted in tumors [72]. The cGAS-STING pathway suppression has been observed in colorectal carcinoma, melanoma, and cancer cells lacking telomerase [71].
However, a lot of other studies have revealed that cGAS activation by self-DNA could promote cancer development by inducing chronic and aberrant inflammation [1,69,73]. Chronic activation of cGAS signaling in cancer cells with high chromosomal instability promotes invasion and metastasis, attributed to a switch from IFN-I and canonical NF-κB signaling to non-canonical NF-κB cascades [74]. cGAS serves as a major factor in promoting hepatocellular carcinoma cell (HCC) tumor growth by suppressing ferroptosis in vivo [75]. Knocking down of cGAS significantly suppressed tumor growth of Hep3B cells in mouse xenograft model, which was completely rescued by the restored expression of cGAS, suggesting that the cGAS is important for tumor growth in HCC [75]. Another study indicated that tumor metastasis in mice brain was connected with the 2′3′-cGAMP transferred from tumor cells to astrocytes in an adjacent paracrine and endocytosis manner. In the process, the cGAS pathway in astrocytes was activated together with IFN-α and TNF-α activation, which contributed to a tumor growth advantage [76]. Thus, the activation of cGAS by self-DNA can carry out multiple functions in cancers.
6. Conclusions and Perspectives
cGAS is a universal DNA sensor and cannot discriminate self from non-self DNA, thus both DNAs are capable of stimulating the cGAS pathway. Self-DNA sensing by the cGAS pathway can cause autoimmune diseases and regulate the progression of tumor. Under normal physiological conditions, cGAS avoids to sense self-DNA though the following mechanisms: DNases degrades self-DNA under normal conditions to prevent aberrant activation of cGAS-mediated immune responses. In the meantime, cGAS is localized to the plasma membrane during the stationary phase, which could hinder it from binding self-DNA in cytoplasm. During mitosis, cGAS is interacted with histone and BAF, which block cGAS to sense self-DNA. Further, cGAS is hyperphosphorylated by AurB and CDK1-cyclin B complex during mitosis, suppressing the activity of cGAS.
Not only the activity of cGAS was suppressed during mitosis, indeed, the phosphorylations of STING and IRF3 by TBK1 were also inhibited during mitosis by unknown mechanism [77]. Further, cGAS was shown as a shuttle protein transported among the plasma membrane, cytoplasm and nucleus [78]. The transportation of cGAS was dependent on several factors including pathogenic infection, cell cycle and DNA damage. However, the exact regulatory mechanism of cGAS transportation is still unclear. Thus, it is recommended that studies should be designed to explore these different aspects. Additionally, due to its indirect regulation and dual roles affecting diverse downstream regulatory factors, cGAS exerts a dichotomous effect on tumors after sensing self-DNA. Therefore, the activation status of cGAS and its function in a particular tumor should be considered when an intervention method targeting the cGAS pathway for cancer therapy is developed.
Author Contributions
J.Z. and W.Z. (Wanglong Zheng) proposed the idea and wrote the manuscript; W.Z. (Wangli Zheng) wrote the manuscript and drew the figures; N.C. and F.M. wrote the manuscript, made revisions, and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
The data presented in this study are available in insert article.
Acknowledgments
The work was partly supported by the National Natural Science Foundation of China (32202818; 32172867; 31872450), and A Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
References
- Ablasser, A.; Chen, Z.J.J. cGAS in action: Expanding roles in immunity and inflammation. Science 2019, 363, 1055. [Google Scholar] [CrossRef]
- Civril, F.; Deimling, T.; Mann, C.C.D.; Ablasser, A.; Moldt, M.; Witte, G.; Hornung, V.; Hopfner, K.P. Structural mechanism of cytosolic DNA sensing by cGAS. Nature 2013, 498, 332. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.L.; Liu, A.J.; Xia, N.W.; Chen, N.H.; Meurens, F.; Zhu, J.Z. How the Innate Immune DNA Sensing cGAS-STING Pathway Is Involved in Apoptosis. International journal of molecular sciences 2023, 24. [Google Scholar] [CrossRef]
- Song, J.X.; Villagomes, D.; Zhao, H.C.; Zhu, M. cGAS in nucleus: The link between immune response and DNA damage repair. Front Immunol 2022, 13. [Google Scholar] [CrossRef]
- Gong, T.; Liu, L.; Jiang, W.; Zhou, R.B. DAMP-sensing receptors in sterile inflammation and inflammatory diseases. Nature Reviews Immunology 2020, 20, 95–112. [Google Scholar] [CrossRef] [PubMed]
- Mata-Garrido, J.; Frizzi, L.; Nguyen, T.; He, X.Y.; Chang-Marchand, Y.; Xiang, Y.; Reisacher, C.; Casafont, I.; Arbibe, L. HP1 gamma Prevents Activation of the cGAS/STING Pathway by Preserving Nuclear Envelope and Genomic Integrity in Colon Adenocarcinoma Cells. International journal of molecular sciences 2023, 24. [Google Scholar] [CrossRef]
- Amadio, R.; Piperno, G.M.; Benvenuti, F. Self-DNA Sensing by cGAS-STING and TLR9 in Autoimmunity: Is the Cytoskeleton in Control? Front Immunol 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Hopfner, K.P.; Hornung, V. Molecular mechanisms and cellular functions of cGAS-STING signalling. Nat Rev Mol Cell Bio 2020, 21, 501–521. [Google Scholar] [CrossRef]
- Guttinger, S.; Laurell, E.; Kutay, U. Orchestrating nuclear envelope disassembly and reassembly during mitosis. Nat Rev Mol Cell Bio 2009, 10, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Rigby, R.E.; Leitch, A.; Jackson, A.P. Nucleic acid-mediated inflammatory diseases. Bioessays 2008, 30, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Liu, P.D. Cytosolic DNA sensing by cGAS: regulation, function, and human diseases. Signal transduction and targeted therapy 2021, 6. [Google Scholar] [CrossRef]
- Joshi, B.; Joshi, J.C.; Mehta, D. Regulation of cGAS Activity and Downstream Signaling. Cells 2022, 11. [Google Scholar] [CrossRef]
- Bai, J.L.; Liu, F. Nuclear cGAS: sequestration and beyond. Protein & cell 2022, 13, 90–101. [Google Scholar] [CrossRef]
- Wu, X.M.; Wu, F.H.; Wang, X.Q.; Wang, L.L.; Siedow, J.N.; Zhang, W.G.; Pei, Z.M. Molecular evolutionary and structural analysis of the cytosolic DNA sensor cGAS and STING. Nucleic Acids Res 2014, 42, 8243–8257. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.K.; Li, S.T. Role of Post-Translational Modifications of cGAS in Innate Immunity. International journal of molecular sciences 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Huang, T.Z.; Chen, Z.J.J. Phosphorylation and Chromatin Tethering Prevent cGAS activation During Mitosis. Journal of immunology 2021, 206. [Google Scholar] [CrossRef]
- Cui, Y.; Yu, H.S.; Zheng, X.; Peng, R.; Wang, Q.; Zhou, Y.; Wang, R.; Wang, J.H.; Qu, B.; Shen, N. , et al. SENP7 Potentiates cGAS Activation by Relieving SUMO-Mediated Inhibition of Cytosolic DNA Sensing. Plos Pathogens 2017, 13. [Google Scholar] [CrossRef]
- Wang, Q.; Huang, L.Y.; Hong, Z.; Lv, Z.S.; Mao, Z.M.; Tang, Y.J.; Kong, X.F.; Li, S.L.; Cui, Y.; Liu, H. , et al. The E3 ubiquitin ligase RNF185 facilitates the cGAS-mediated innate immune response. Plos Pathogens 2017, 13. [Google Scholar] [CrossRef]
- Ji, L.K.; Wang, Y.; Zhou, L.Y.; Lu, J.; Bao, S.W.; Shen, Q.; Wang, X.C.; Liu, Y.W.; Zhang, W. E3 Ubiquitin Ligases: The Operators of the Ubiquitin Code That Regulates the RLR and cGAS-STING Pathways. International journal of molecular sciences 2022, 23. [Google Scholar] [CrossRef]
- Hu, M.M.; Yang, Q.; Xie, X.Q.; Liao, C.Y.; Lin, H.; Liu, T.T.; Yin, L.; Shu, H.B. Sumoylation Promotes the Stability of the DNA Sensor cGAS and the Adaptor STING to Regulate the Kinetics of Response to DNA Virus. Immunity 2016, 45, 555–569. [Google Scholar] [CrossRef]
- Dai, J.; Huang, Y.J.; He, X.H.; Zhao, M.; Wang, X.Z.; Liu, Z.S.; Xue, W.; Cai, H.; Zhan, X.Y.; Huang, S.Y. , et al. Acetylation Blocks cGAS Activity and Inhibits Self-DNA-Induced Autoimmunity. Cell 2019, 176, 1447. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.M.; Lin, H.; Yi, X.M.; Guo, W.; Hu, M.M.; Shu, H.B. KAT5 acetylates cGAS to promote innate immune response to DNA virus. Proceedings of the National Academy of Sciences of the United States of America 2020, 117, 21568–21575. [Google Scholar] [CrossRef] [PubMed]
- Dowling, Q.M.; Volkman, H.E.; Gray, E.E.; Ovchinnikov, S.; Cambier, S.; Bera, A.K.; Sankaran, B.; Johnson, M.R.; Bick, M.J.; Kang, A.L. , et al. Computational design of constitutively active cGAS. Nat Struct Mol Biol 2023, 30, 72. [Google Scholar] [CrossRef] [PubMed]
- Herzner, A.M.; Schlee, M.; Bartok, E. The many faces of cGAS: how cGAS activation is controlled in the cytosol, the nucleus, and during mitosis. Signal transduction and targeted therapy 2021, 6. [Google Scholar] [CrossRef]
- Mann, C.D.C.; Hornung, V. Molecular mechanisms of nonself nucleic acid recognition by the innate immune system. European journal of immunology 2021, 51, 1897–1910. [Google Scholar] [CrossRef]
- Zhang, X.W.; Bai, X.C.; Chen, Z.J.J. Structures and Mechanisms in the cGAS-STING Innate Immunity Pathway. Immunity 2020, 53, 43–53. [Google Scholar] [CrossRef]
- Zheng, J.Y.; Mo, J.L.; Zhu, T.; Zhuo, W.; Yi, Y.N.; Hu, S.; Yin, J.Y.; Zhang, W.; Zhou, H.H.; Liu, Z.Q. Comprehensive elaboration of the cGAS-STING signaling axis in cancer development and immunotherapy. Molecular cancer 2020, 19. [Google Scholar] [CrossRef]
- Robb, C.G.; Dao, T.P.; Ujma, J.; Castaneda, C.; Beveridge, R. Ion Mobility Mass Spectrometry Unveils Global Protein Conformations in Response to Conditions that Promote and Reverse Liquid-Liquid Phase Separation. J Am Chem Soc 2023, 145, 12541–12549. [Google Scholar] [CrossRef]
- Xiao, Q.; McAtee, C.K.; Su, X.L. Phase separation in immune signalling. Nature Reviews Immunology 2022, 22, 188–199. [Google Scholar] [CrossRef]
- Du, M.J.; Chen, Z.J.J. DNA-induced liquid phase condensation of cGAS activates innate immune signaling. Science 2018, 361, 704. [Google Scholar] [CrossRef]
- Hu, M.M.; Shu, H.B. Innate Immune Response to Cytoplasmic DNA: Mechanisms and Diseases. Annu Rev Immunol 2020, 38, 79–98. [Google Scholar] [CrossRef]
- Zhou, W.; Mohr, L.; Maciejowski, J.; Kranzusch, P.J. cGAS phase separation inhibits TREX1-mediated DNA degradation and enhances cytosolic DNA sensing. Molecular cell 2021, 81, 739. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Shu, C.; Yi, G.H.; Chaton, C.T.; Shelton, C.L.; Diao, J.S.; Zuo, X.B.; Kao, C.C.; Herr, A.B.; Li, P.W. Cyclic GMP-AMP Synthase Is Activated by Double-Stranded DNA-Induced Oligomerization. Immunity 2013, 39, 1019–1031. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Chen, Z.J. DNA-induced liquid phase condensation of cGAS activates innate immune signaling. Science 2018, 361, 704–709. [Google Scholar] [CrossRef]
- Mitic, N.; Miraula, M.; Selleck, C.; Hadler, K.S.; Uribe, E.; Pedroso, M.M.; Schenk, G. Catalytic Mechanisms of Metallohydrolases Containing Two Metal Ions. Adv Protein Chem Str 2014, 97, 49–81. [Google Scholar] [CrossRef]
- Zhao, Z.; Ma, Z.X.; Wang, B.; Guan, Y.K.; Su, X.D.; Jiang, Z.F. Mn2+ Directly Activates cGAS and Structural Analysis Suggests Mn2+ Induces a Noncanonical Catalytic Synthesis of 2 ‘ 3 ‘-cGAMP. Cell Rep 2020, 32. [Google Scholar] [CrossRef]
- Liu, H.S.; Yan, Z.Z.; Zhu, D.Y.; Xu, H.Y.; Liu, F.; Chen, T.; Zhang, H.H.; Zheng, Y.; Liu, B.Y.; Zhang, L.; et al. CD-NTase family member MB21D2 promotes cGAS-mediated antiviral and antitumor immunity. Cell death and differentiation 2023. [Google Scholar] [CrossRef] [PubMed]
- Rozenberg, J.M.; Kamynina, M.; Sorokin, M.; Zolotovskaia, M.; Koroleva, E.; Kremenchutckaya, K.; Gudkov, A.; Buzdin, A.; Borisov, N. The Role of the Metabolism of Zinc and Manganese Ions in Human Cancerogenesis. Biomedicines 2022, 10. [Google Scholar] [CrossRef]
- Wang, C.G.; Guan, Y.K.; Lv, M.Z.; Zhang, R.; Guo, Z.Y.; Wei, X.M.; Du, X.X.; Yang, J.; Li, T.; Wan, Y. , et al. Manganese Increases the Sensitivity of the cGAS-STING Pathway for Double-Stranded DNA and Is Required for the Host Defense against DNA Viruses. Immunity 2018, 48, 675. [Google Scholar] [CrossRef]
- Ma, R.H.; Serrano, T.P.O.; Davis, J.; Prigge, A.D.; Ridge, K.M. The cGAS-STING pathway: The role of self-DNA sensing in inflammatory lung disease. Faseb Journal 2020, 34, 13156–13170. [Google Scholar] [CrossRef]
- Koyama, R.; Arai, T.; Kijima, M.; Sato, S.; Miura, S.; Yuasa, M.; Kitamura, D.; Mizuta, R. DNase, DNase I and caspase-activated DNase cooperate to degrade dead cells. Genes Cells 2016, 21, 1150–1163. [Google Scholar] [CrossRef]
- Anindya, R. Cytoplasmic DNA in cancer cells: Several pathways that potentially limit DNase2 and TREX1 activities. Bba-Mol Cell Res 2022, 1869. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.R.; Hemphill, W.O.; Hudson, T.; Perrino, F.W. TREX1-Apex predator of cytosolic DNA metabolism. DNA Repair 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Xiao, N.Y.; Wei, J.J.; Xu, S.; Du, H.K.; Huang, M.H.; Zhang, S.T.; Ye, W.W.; Sun, L.J.; Chen, Q. cGAS activation causes lupus-like autoimmune disorders in a TREX1 mutant mouse model. J Autoimmun 2019, 100, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Giordano, A.M.S.; Luciani, M.; Gatto, F.; Abou Alezz, M.; Beghe, C.; Della Volpe, L.; Migliara, A.; Valsoni, S.; Genua, M.; Dzieciatkowska, M. , et al. DNA damage contributes to neurotoxic inflammation in Aicardi-Goutieres syndrome astrocytes. Journal of Experimental Medicine 2022, 219. [Google Scholar] [CrossRef]
- Fang, L.W.; Ying, S.C.; Xu, X.; Wu, D. TREX1 cytosolic DNA degradation correlates with autoimmune disease and cancer immunity. Clin Exp Immunol 2023, 211, 193–207. [Google Scholar] [CrossRef]
- Gao, D.X.; Li, T.; Li, X.D.; Chen, X.; Li, Q.Z.; Wight-Carter, M.; Chen, Z.J. Activation of cyclic GMP-AMP synthase by self-DNA causes autoimmune diseases. Proceedings of the National Academy of Sciences of the United States of America 2015, 112, E5699–E5705. [Google Scholar] [CrossRef]
- Barnett, K.C.; Coronas-Serna, J.M.; Zhou, W.; Ernandes, M.J.; Cao, A.; Kranzusch, P.J.; Kagan, J.C. Phosphoinositide Interactions Position cGAS at the Plasma Membrane to Ensure Efficient Distinction between Self- and Viral DNA. Cell 2019, 176, 1432. [Google Scholar] [CrossRef]
- Ablasser, A.; Hur, S. Regulation of cGAS- and RLR-mediated immunity to nucleic acids. Nature immunology 2020, 21, 17–29. [Google Scholar] [CrossRef]
- Wu, Y.K.; Song, K.; Hao, W.Z.; Li, J.; Wang, L.Y.; Li, S.T. Nuclear soluble cGAS senses double-stranded DNA virus infection. Communications biology 2022, 5. [Google Scholar] [CrossRef]
- Wischnewski, M.; Ablasser, A. Interplay of cGAS with chromatin. Trends Biochem Sci 2021, 46, 822–831. [Google Scholar] [CrossRef]
- Zierhut, C.; Yamaguchi, N.; Paredes, M.; Luo, J.D.; Carroll, T.; Funabiki, H. The Cytoplasmic DNA Sensor cGAS Promotes Mitotic Cell Death. Cell 2019, 178, 302. [Google Scholar] [CrossRef]
- Cao, D.F.; Han, X.A.; Fan, X.Y.; Xu, R.M.; Zhang, X.Z. Structural basis for nucleosome-mediated inhibition of cGAS activity. Cell research 2020, 30, 1088–1097. [Google Scholar] [CrossRef]
- Guey, B.; Wischnewski, M.; Decout, A.; Makasheva, K.; Kaynak, M.; Sakar, M.S.; Fierz, B.; Ablasser, A. BAF restricts cGAS on nuclear DNA to prevent innate immune activation. Science 2020, 369, 823. [Google Scholar] [CrossRef]
- Jamin, A.; Wiebe, M.S. Barrier to Autointegration Factor (BANF1): interwoven roles in nuclear structure, genome integrity, innate immunity, stress responses and progeria. Current opinion in cell biology 2015, 34, 61–68. [Google Scholar] [CrossRef]
- Bradley, C.M.; Ronning, D.R.; Ghirlando, R.; Craigie, R.; Dyda, F. Structural basis for DNA bridging by barrier-to-autointegration factor. Nat Struct Mol Biol 2005, 12, 935–936. [Google Scholar] [CrossRef]
- Broussard, G.; Ni, G.X.; Zhang, Z.G.; Li, Q.; Cano, P.; Dittmer, D.P.; Damania, B. Barrier-to-autointegration factor 1 promotes gammaherpesvirus reactivation from latency. Nature communications 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Hu, M.M.; Bian, L.J.; Liu, Y.; Chen, Q.; Shu, H.B. Phosphorylation of cGAS by CDK1 impairs self-DNA sensing in mitosis. Cell Discov 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- Navegantes, K.C.; Gomes, R.D.; Pereira, P.A.T.; Czaikoski, P.G.; Azevedo, C.H.M.; Monteiro, M.C. Immune modulation of some autoimmune diseases: the critical role of macrophages and neutrophils in the innate and adaptive immunity. J Transl Med 2017, 15. [Google Scholar] [CrossRef] [PubMed]
- Benmerzoug, S.; Ryffel, B.; Togbe, D.; Quesniaux, V.F.J. Self-DNA Sensing in Lung Inflammatory Diseases. Trends in immunology 2019, 40, 719–734. [Google Scholar] [CrossRef]
- Tonduti, D.; Fazzi, E.; Badolato, R.; Orcesi, S. Novel and emerging treatments for Aicardi-Goutieres syndrome. Expert Rev Clin Immu 2020, 16, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Pokatayev, V.; Hasin, N.; Chon, H.; Cerritelli, S.M.; Sakhuja, K.; Ward, J.M.; Morris, H.D.; Yan, N.; Crouch, R.J. RNase H2 catalytic core Aicardi-Goutieres syndrome-related mutant invokes cGAS-STING innate immune-sensing pathway in mice. Journal of Experimental Medicine 2016, 213, 329–336. [Google Scholar] [CrossRef]
- Gray, E.E.; Treuting, P.M.; Woodward, J.J.; Stetson, D.B. Cutting Edge: cGAS Is Required for Lethal Autoimmune Disease in the Trex1-Deficient Mouse Model of Aicardi-Goutieres Syndrome. Journal of immunology 2015, 195, 1939–1943. [Google Scholar] [CrossRef]
- Crow, M.K. Pathogenesis of systemic lupus erythematosus: risks, mechanisms and therapeutic targets. Annals of the rheumatic diseases 2023. [Google Scholar] [CrossRef]
- Kato, Y.; Park, J.; Takamatsu, H.; Konaka, H.; Aoki, W.; Aburaya, S.; Ueda, M.; Nishide, M.; Koyama, S.; Hayama, Y. , et al. Apoptosis-derived membrane vesicles drive the cGAS-STING pathway and enhance type I IFN production in systemic lupus erythematosus. Annals of the rheumatic diseases 2018, 77, 1507–1515. [Google Scholar] [CrossRef]
- Duan, D.D.; Shang, M.J.; Han, Y.X.; Liu, J.Y.; Liu, J.W.; Kong, S.H.; Hou, J.Y.; Huang, B.Q.; Lu, J.; Zhang, Y. EZH2-CCF-cGAS Axis Promotes Breast Cancer Metastasis. International journal of molecular sciences 2022, 23. [Google Scholar] [CrossRef]
- Huang, J.L.; Chang, Y.T.; Hong, Z.Y.; Lin, C.S. Targeting DNA Damage Response and Immune Checkpoint for Anticancer Therapy. International journal of molecular sciences 2022, 23. [Google Scholar] [CrossRef]
- Guscott, M.; Saha, A.; Maharaj, J.; McClelland, S.E. The multifaceted role of micronuclei in tumour progression: A whole organism perspective. Int J Biochem Cell B 2022, 152. [Google Scholar] [CrossRef]
- Shen, R.; Liu, D.S.; Wang, X.N.; Guo, Z.; Sun, H.A.; Song, Y.F.; Wang, D.G. DNA Damage and Activation of cGAS/ STING Pathway Induce Tumor Microenvironment Remodeling. Frontiers in cell and developmental biology 2022, 9. [Google Scholar] [CrossRef]
- Yum, S.; Li, M.H.; Chen, Z.J.J. Old dogs, new trick: classic cancer therapies activate cGAS. Cell research 2020, 30, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Khoo, L.T.; Chen, L.Y. Role of the cGAS-STING pathway in cancer development and oncotherapeutic approaches. Embo Rep 2018, 19. [Google Scholar] [CrossRef]
- Decout, A.; Katz, J.D.; Venkatraman, S.; Ablasser, A. The cGAS-STING pathway as a therapeutic target in inflammatory diseases. Nature Reviews Immunology 2021, 21, 548–569. [Google Scholar] [CrossRef] [PubMed]
- Bakhoum, S.F.; Ngo, B.; Laughney, A.M.; Cavallo, J.A.; Murphy, C.J.; Ly, P.; Shah, P.; Sriram, R.K.; Watkins, T.B.K.; Taunk, N.K. , et al. Chromosomal instability drives metastasis through a cytosolic DNA response. Nature 2018, 553, 467. [Google Scholar] [CrossRef]
- Bakhoum, S.; Ngo, B.; Bakhoum, A.; Cavallo-Fleming, J.A.; Murphy, C.W.; Powell, S.N.; Cantley, L. Chromosomal Instability Drives Metastasis Through a Cytosolic DNA Response. Int J Radiat Oncol 2018, 102, S118–S118. [Google Scholar] [CrossRef]
- Qiu, S.Q.; Zhong, X.Y.; Meng, X.; Li, S.T.; Qian, X.Y.; Lu, H.; Cai, J.; Zhang, Y.; Wang, M.J.; Ye, Z.J. , et al. Mitochondria-localized cGAS suppresses ferroptosis to promote cancer progression. Cell research 2023, 33, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Boire, A.; Jin, X.; Valiente, M.; Er, E.E.; Lopez-Soto, A.; Jacob, L.S.; Patwa, R.; Shah, H.; Xu, K. , et al. Carcinoma-astrocyte gap junctions promote brain metastasis by cGAMP transfer (vol 533, pg 493, 2016). Nature 2017, 544, 124–124. [Google Scholar] [CrossRef] [PubMed]
- Uhlorn, B.L.; Gamez, E.R.; Li, S.Z.; Campos, S.K. Attenuation of cGAS/STING activity during mitosis. Life Sci Alliance 2020, 3. [Google Scholar] [CrossRef]
- Liu, H.P.; Wang, F.; Cao, Y.J.; Dang, Y.F.; Ge, B.X. The multifaceted functions of cGAS. J Mol Cell Biol 2022, 14. [Google Scholar] [CrossRef]
Figure 1.
Sequence structure and post-translational modifications of cGAS. (1) Human cGAS is a protein consisting of 522 amino acids, and it is composed of an N-terminus (1-160) and a C terminus (161-522) that contains a nucleotidyltransferase (NTase) core domain (161-330) and the male abnormal 21 (Mab21) domain (213-513). The Mab21 domain harbors the conserved Zn finger motif (390-405). (2) cGAS is regulated by various posttranslational modifications, and the phosphorylation, ubiquitination, sumoylation and acetylation are major post-translational modifications in the celluar cGAS. The common post-translational modification sites of cGAS was illustrated.
Figure 1.
Sequence structure and post-translational modifications of cGAS. (1) Human cGAS is a protein consisting of 522 amino acids, and it is composed of an N-terminus (1-160) and a C terminus (161-522) that contains a nucleotidyltransferase (NTase) core domain (161-330) and the male abnormal 21 (Mab21) domain (213-513). The Mab21 domain harbors the conserved Zn finger motif (390-405). (2) cGAS is regulated by various posttranslational modifications, and the phosphorylation, ubiquitination, sumoylation and acetylation are major post-translational modifications in the celluar cGAS. The common post-translational modification sites of cGAS was illustrated.
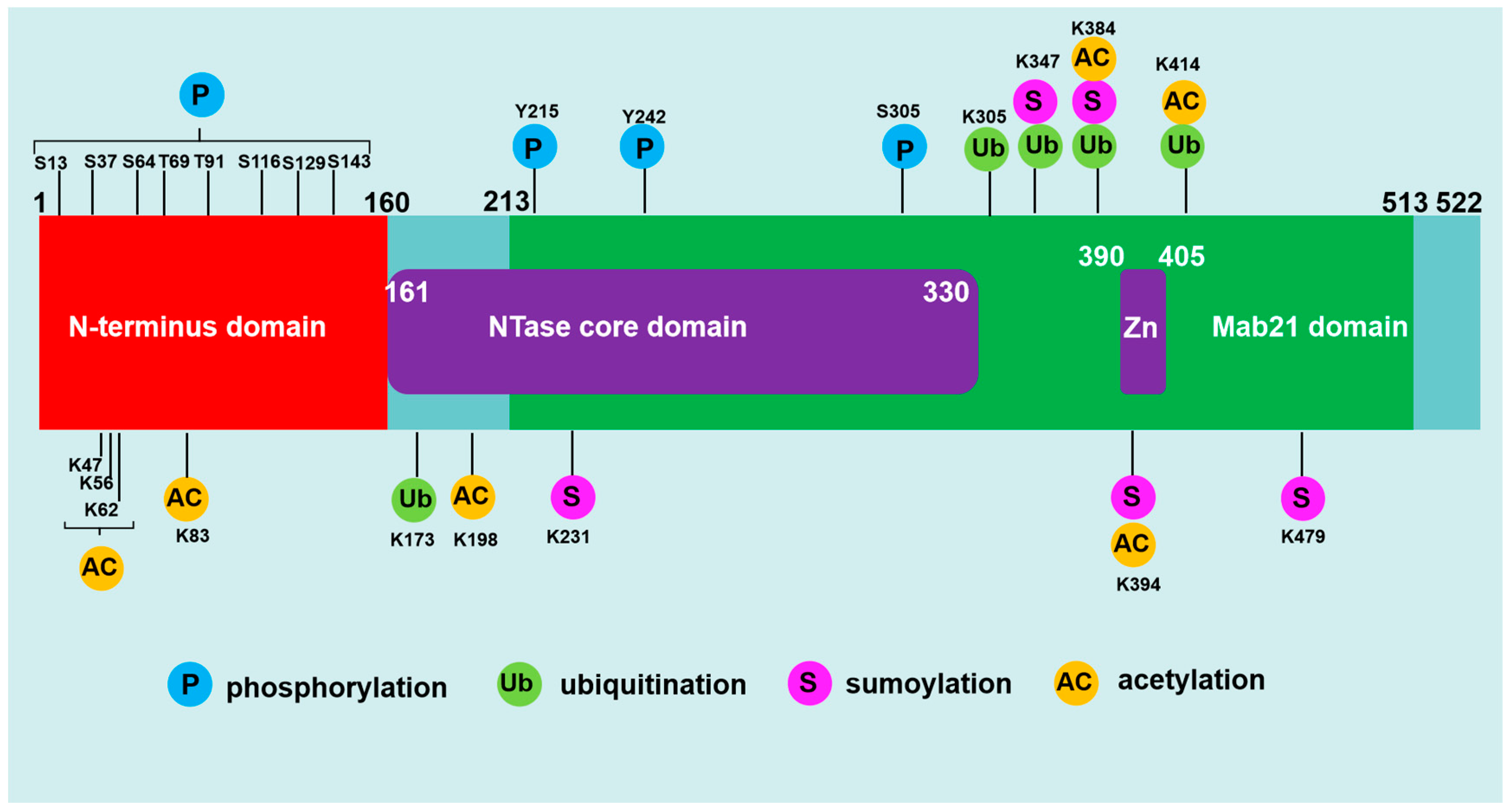
Figure 2.
The mechanisms by cGAS to avoid sensing self-DNA during interphase. ① Self-DNA in serum or on cell surface from dead cell is degraded by DNase I. ② Self-DNA in endosome or lysosome is degraded by DNase II. ③ Self-DNA in cytoplasm is degraded by TREX1. ④ cGAS is located in the plasma membrane through interacting with PI(4,5)P2, in which it is least likely to detect self-DNA and prevent aberrant activation.
Figure 2.
The mechanisms by cGAS to avoid sensing self-DNA during interphase. ① Self-DNA in serum or on cell surface from dead cell is degraded by DNase I. ② Self-DNA in endosome or lysosome is degraded by DNase II. ③ Self-DNA in cytoplasm is degraded by TREX1. ④ cGAS is located in the plasma membrane through interacting with PI(4,5)P2, in which it is least likely to detect self-DNA and prevent aberrant activation.
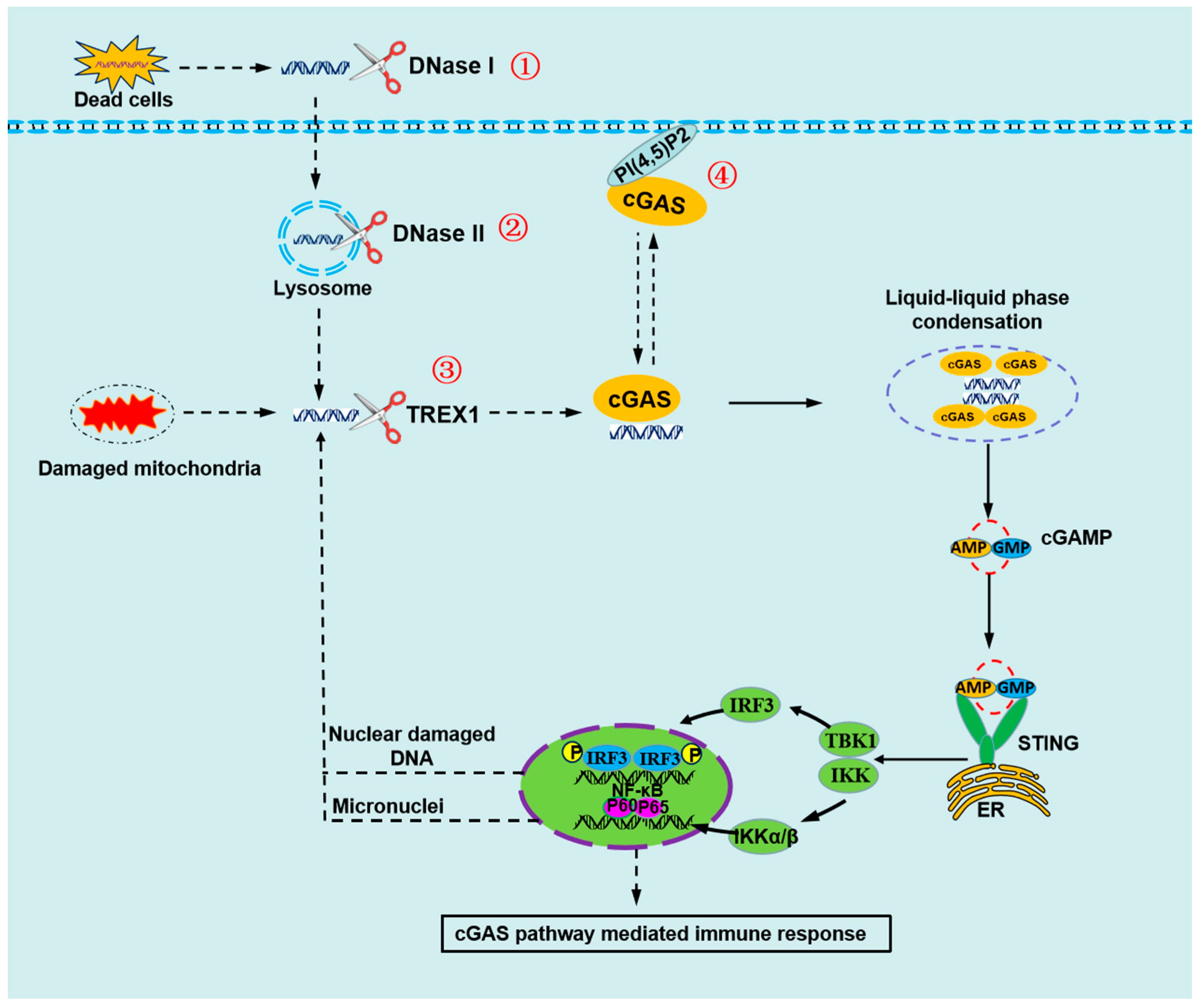
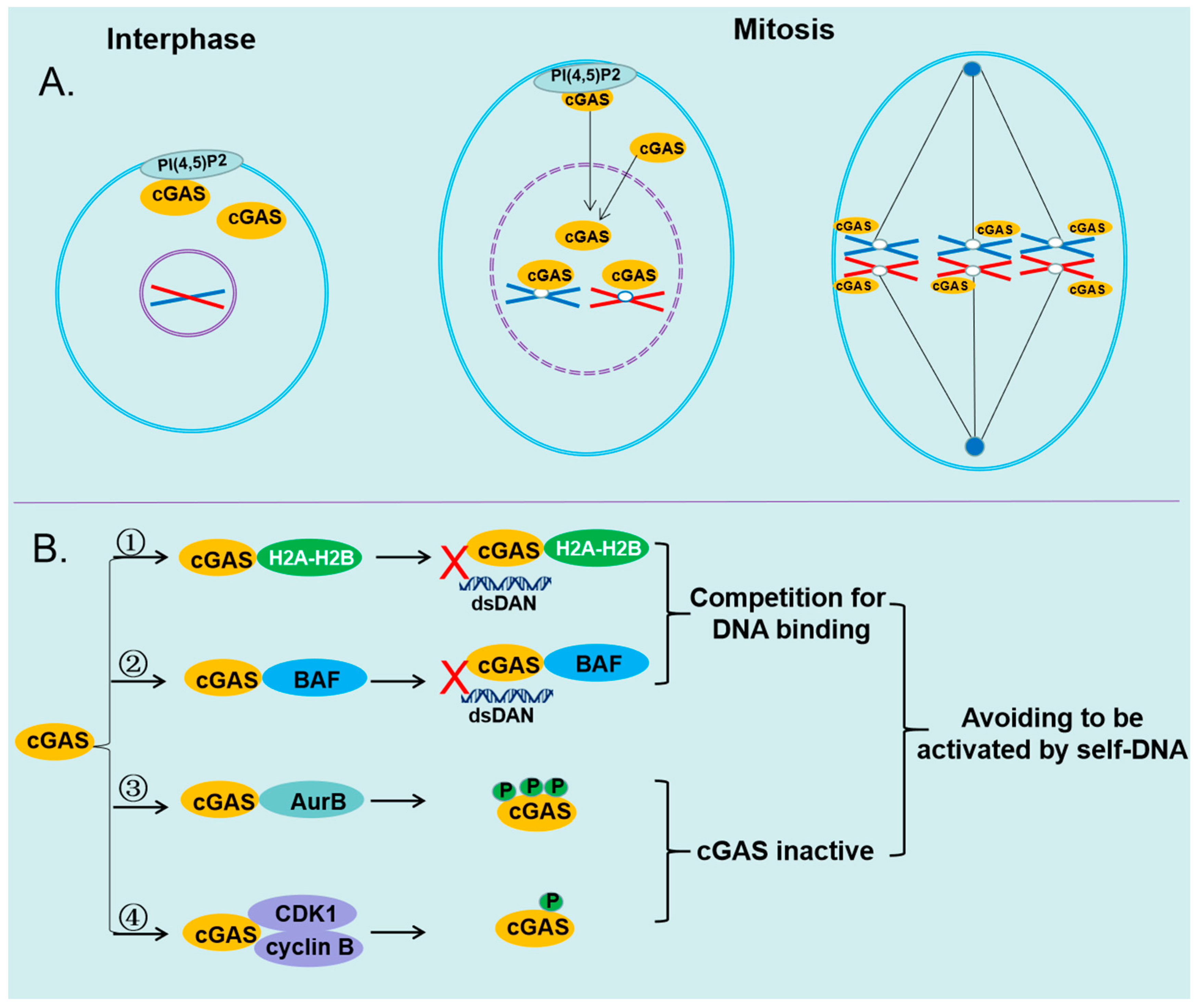
Figure 3.
The mechanisms of cGAS avoiding to be activated by self-DNA.(A)cGAS is transported to nucleus and interacts with chromatin during mitosis, when the nuclear envelope is disassembled. (B) ①cGAS interacts with the H2A–H2B heterodimer, which prevents it further binding to dsDNA.②cGAS interacts with BAF, which dynamically displaces transiently bound cGAS monomer from dsDNA. ③ cGAS is hyperphosphorylated by AurB at the N-terminal serine and threonine residues, which inhibits the ability of cGAS to form Liquid-liquid phase condensation. ④ cGAS is phosphorylated by the CDK1-cyclin B complex at a highly conserved site Ser305, which impairs the ability of cGAS to synthesize 2′3′-cGAMP.
Figure 3.
The mechanisms of cGAS avoiding to be activated by self-DNA.(A)cGAS is transported to nucleus and interacts with chromatin during mitosis, when the nuclear envelope is disassembled. (B) ①cGAS interacts with the H2A–H2B heterodimer, which prevents it further binding to dsDNA.②cGAS interacts with BAF, which dynamically displaces transiently bound cGAS monomer from dsDNA. ③ cGAS is hyperphosphorylated by AurB at the N-terminal serine and threonine residues, which inhibits the ability of cGAS to form Liquid-liquid phase condensation. ④ cGAS is phosphorylated by the CDK1-cyclin B complex at a highly conserved site Ser305, which impairs the ability of cGAS to synthesize 2′3′-cGAMP.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



