
Submitted:
24 September 2023
Posted:
26 September 2023
You are already at the latest version
Abstract
Calicotome villosa is a eurioic legume with broad distribution in the south of Europe. It can grow in almost any type of soils as well as in humid and dry and nutrient deficient ecosystems. The broad distribution and eurioic nature of C. villosa may be attributed to its ability to establish symbiotic associations with plant growth promoting (PGP) bacteria housed in the nodules and rootless. This study examined legume-microbe symbiosis of C. villosa growing in two contrasted locations, hill top with high sun irradiance and drought; and a valley bottom, with low level or radiation and sufficient soil humidity for plant growth. Calicotome villosa adult plants established symbiotic in-teractions with bacteria in six genera, Pseudomonas, Stenotrophomonas, Bacillus, Paenibacillus, Brevicacterium and Rhizobium as well as one uncultured bacteria. Plants growing in the hill top had associated lower bacterial richness than those grown at the valley bottom. All strains were drought tolerant and produced siderophores, IAA, HCN and NH3 that stimulated plant performance in both, C. villosa plants and other four legumes commonly present in the understory of the shrubs communities The legumes’ capacity to selectively host symbiotic bacteria that enhance plant survival in harsh conditions partly accounts for the diverse partnerships between C. villosa plants and their symbionts, ultimately explaining the wide distribution of this plant species.
Keywords:
legumes
; siderophores
; IAA
; biomass
; bacterial richness
1. Introduction
Shrubs play a vital ecological role in the Iberian Peninsula due to their multifaceted contributions to the ecosystems [1]. These plants are particularly well adapted to the Mediterranean climate, characterized by hot and dry summers [2]. Shrubs provide habitat and food resources for a wide range of wildlife, including insects, birds, and mammals, contributing to biodiversity conservation [3]. Their intricate root systems help stabilize soil, preventing erosion and supporting water infiltration, which is essential in the face of frequent droughts [4]. Additionally, many shrub species have developed strategies to cope in nutrient-poor soils, promoting nutrient cycling and enriching the soil by establishing symbiotic interactions with soil borne microorganisms [5,6]. Their ability to thrive in challenging environmental conditions makes shrubs resilient components of the Iberian ecosystems, acting as pioneers in ecosystem restoration after disturbances such as fires [7]. .
The establishment of leguminous shrubs under Mediterranean type of climates faces several significant constraints that shape their distribution and abundance. One primary limitation is the nutrient-poor soils, which affects the growth and survival of leguminous species. These plants rely on a symbiotic relationship with nitrogen(N)-fixing bacteria to obtain essential nutrients, particularly nitrogen (N) and Phosphorus (P) [7,8]. Moreover, the Mediterranean climate can pose a challenge to water availability for these plants, especially during critical establishment phases [1]. Competition with other vegetation, including non-native species, further adds to the challenges leguminous shrubs face in securing their foothold in the Mediterranean ecosystems [9]. These constraints collectively influence the spatial distribution and success of leguminous shrubs, underscoring the intricate interplay between environmental factors and plant adaptability in the Mediterranean areas of the world [7].
In the afore-mentioned Mediterranean-type climates, light often functions as a critical limiting factor for plant establishment and growth. The intense sunlight characteristic in these regions can lead to high temperatures and increased evaporation rates, resulting in water stress for plants [6]. This stress, combined with limited water availability, can hinder photosynthesis and nutrient uptake, affecting the overall vitality and survival of plants. Additionally, the frequent occurrence of drought conditions further exacerbates the challenges posed by high light levels [2]. Plants in Mediterranean environments have evolved various strategies to cope with these conditions, such as developing thick, waxy leaves that reduce water loss through transpiration and employing mechanisms to optimize water use efficiency. These adaptations highlight the pivotal role of light availability in shaping plant distribution, community composition, and ecosystem dynamics in Mediterranean ecosystems, where the balance between water availability and light intensity plays a crucial role in determining the success of plant species [9].
Calicotome villosa (Poir.) Link, commonly known as hairy green weed or gergene, holds significant ecological importance in the southern part of Europe [10,39]. This shrub is well-adapted to the challenging Mediterranean climate . Its ability to thrive in such conditions makes it a valuable component of local ecosystems, contributing to soil stabilization, erosion control, and the prevention of desertification. Furthermore, its ability to establish symbiotic interactions with N-fixing bacteria enhances the nutrient availability in nutrient-poor soils, supporting the growth of other plants and fostering a healthier ecosystem [40,41]. In rural areas, this shrub has cultural significance and has been used historically for livestock forage and traditional practices and at the same time it plays a vital role in maintaining the ecological balance, resilience, and sustainability of the diverse ecosystems [10]. In the southern part of Spain, it can be observed growing both in shaded valleys and on the top of the hill, where high sunlight imposes increased stress coupled with intense drought.The wide geographic distribution of C. villosa suggests the influence of specific factors. This plant exhibits adaptability across diverse environmental conditions, thriving in regions characterized by high temperatures and intense radiation as well as in shaded valleys. Furthermore, it displays tolerance for a broad range of soil types. We posit that this extensive distribution is facilitated by the plant's capacity to selectively engage in symbiotic relationships with endophytic bacteria. Our objective in this study is to investigate the mechanisms by which C. villosa interacts with soil bacteria, aiming to gain insights into the key drivers behind its remarkable adaptability and widespread presence in varied ecological niches.
We propose that the resilience to high irradiance and the extreme conditions on top of the hills, roots in a greater microbial diversity associated to the plants in comparison with those in shaded environments. Microbial associations promote plant growth and maintenance by providing protective mechanisms absent in plants growing in milder environment. We also propose that the presence of bacteria associated to C. villosa induces qualitative changes in the understory species acting as modulators of plant growth triggered by the environmental conditions. We tested our hypothesis for the culturable bacteria associated with rootless and nodules of the shrubby legume C. villosa as these bacteria can be tested for the production of plant growth promoting bacteria and their effects on plant performance.
2. Materials and Methods
2.1. Plant material collection
Nodules and rootles of C. villosa were collected from wild plants growing in two populations of the Natural Park of Los Alcornocales in the Cadiz province (Spain). One of the populations was located at the top of a hill (36° 31´98´´N, 5° 45´59´´W and 1,796 m asl) and the other 158 m apart, at the bottom of the hill in a small valley (36° 31´85´´N, 5° 45´65´´W and 1,726 m asl). Thus, the two populations were 70m apart in altitude.
Root nodules and rootles were dissected from roots and subsequently transferred to Eppendorf tubes which were then stored in a portable refrigerator unit to maintain sample integrity during transport to the laboratory. Plant material was surface sterilized by immersion in 65% ethanol for 1 min followed by ten rinses with sterile distilled water. Nodules were them immersed in sodium hypochlorite (5% (v/v) for 4 minutes and then thoroughly rinsed for ten successive washes with sterile distilled water.
Nodules and rootles were then crushed on sterile plates and streaked onto yeast-extract mannitol agar (YMA). Plates were incubated at 28 °C for 3–5 days [11]. The purity of the culture was validated by picking and re-streaking individual colonies on fresh plates. Bacteria with different colony morphology, among those isolated from nodules of plants growing in the same location, were selected for further identification.
Pure bacterial colonies were amplified through polymerase chain reaction (PCR) using 16S primers 63F (5’- CAGGCCTAACACATGCAAGTC -3’) and 1387R (5’- GGGCGGTGTGTACAA -3’) in a 100 µl volume containing 0.2 µl of template DNA extract, 20 pmol of each primer, 250 µM of deoxynucleoside triphosphate (Amersham Pharmacia Biotech, Piscataway, NJ, USA) and 2 U of PlatinumTaq Polimerase (Life Technologies/Invitrogen, Carlsbad, CA, USA). The reaction was carried out with a PCR System MyCycler Biorad (Germany) using the protocol usually followed in our laboratory: initial denaturation at 94 °C for 1 min; 30 cycles of 60 s at 94 °C, 60 s at 55 °C and 90 s at 72 °C; final extension at 72 °C for 7 min. The PCR products were sent to the Genetics laboratory at the University Complutense of Madrid, Spain for sequencing. The 16S rDNA sequences were submitted to the GenBank database to search for significant alignments.
A phylogenetic approach was used to determine the evolutionary relationship between bacterial nucleotide sequences. The nucleotide alignment was done using the MUSCLE tool in MEGA 11 and checked manually before constructing the phylogenetic tree using the neighbour-joining likelihood tree approach. Also, a bootstrap resampling was performed with 1000 replicates in accordance with the procedure by [12].
2.2. Authentication of the symbiotic interactions
The isolates were authenticated as root nodulating bacteria by re-inoculating 1 mL of three-day-old pure YEM broth culture of each of the isolates on individual host seedlings grown in slope YMA in sterile jars with four replicates per strain. Jars were covered with sterile hydrophobic cotton and maintained in an incubation chamber at 27 °C in a day -night regime of 14-10 hours. The negative control was set with non-inoculated seedlings. The isolates were also compared with the rhizobia strain from our collection (accession number AF461191) that has been proven for positive nodulation of Ornithopus compressus L. as reference strains [13]. After 35 days, a new extraction was conducted crushing roots and rootless. All bacterial material, from the first extraction and from the seedlings grown in slope YMA were molecularly identified.
2.3. Phenotypical characterization of the bacterial isolates
Bacteria were tested for their capacity to modify pH in the growing media, their tolerance to drought and to a salt gradient.
To test for the ability to modify the pH 1 μl inoculum of each target strain was plated onto Petri dishes containing selective Luria Bertani (LB) medium supplemented with 0.25 g/l of bromothymol blue indicator. Four replicates of each strain were prepared and allowed to grow at 27 °C for approximately 48 hours. The majority of strains that induced a change in medium color shifted it from green to blue, owing to the pH sensitivity range of bromothymol blue indicator, which operates actively within the pH range of 6.0 to 7.6. A medium pH below 6.5 is indicated by a yellow color, whereas a pH between 6.5 and 7.6 corresponds to a green. At pH values above 7.6, the indicator remains blue.
The strains were also grown in LB plates at increasing concentrations of NaCl (1%, 3%, 7% and 10%) for 3–5 days. Finally, to determine the resistance to drought, 1 mL of bacterial culture of each strain was grown in 5 mL of liquid LB with polyethylene glycol making the final osmotic pressure of the media -0.75. Preparations were incubated at 28 °C for approximately 24 hours. The presence of turbidity in the medium indicated strain growth, thus confirming its resistance to drought.
2.4. HCN production
All the isolates were screened for the production of hydrogen cyanide by adapting the method of Lorck [14]. Briefly, nutrient broth was amended with glycine 4.4 g/l and bacteria were streaked on modified agar plate. A Whatman filter paper no. 1 soaked in 2% sodium carbonate in 0.5% picric acid solution was placed in the top of the plate. Plates were sealed with parafilm and incubated at 28± 1ºC for 4 days. Development of orange to red color indicated HCN production.
2.5. Ammonia (NH3) production
Bacterial isolates were tested for the production of ammonia in peptone water. Freshly grown cultures were inoculated in 10 ml peptone water in each tube and incubated for 48–72 h at 28±1 °C. Nessler’s reagent (0.5 mL) was added in each tube. Confirmation of NH3 production was the development of yellow to brown color in the tube [15].
2.6. Assay for indoleacetic acid (IAA) production
IAA production was detected by the modified method as described by [16]. Quantitative analysis of IAA was performed using the method of [17] at different concentrations of tryptophan (0, 50, 150, 300, 400 and 500 mg/mL). Bacterial cultures were grown for 96 h (Rhizobium) 72 h (Paenibacillus and Brevibacterium) and 48 h (Pseudomonas, Stenotrophomonas and Bacillus) on their respective media at 28± 1 °C. Fully grown cultures were centrifuged at 3000 rpm for 30 min. The supernatant (2 mL) was mixed with two drops of orthophosphoric acid and 4ml of the Salkowski reagent (50 ml, 35% of perchloric acid, 1 mL 0.5 M FeCl3 solution). Development of pink color indicates IAA production. Optical density was measured at 536 nm with spectrophotometer Spectronic 20 D+. Concentration of IAA produced by cultures was measured with the help of standard graph of IAA obtained in the range of 10–100 mg/mL [16].
2.7. Siderophore production
Bacterial isolates were assayed for siderophores production on the Chrome azurol S agar medium (Sigma, Ltd.) described by [18]. Chrome azurol S agar plates were prepared and divided into equal sectors and spot inoculated with test organism (10 mL of 106 CFU/mL) and incubated at 28±1 °C for 48–72 h. Development of yellow–orange halo around the growth was considered as positive for siderophore production.
Quantitative estimation of siderophore production was performed by growing the isolates in a succinate medium at 28 ± 2 °C for 48 h [19]. The supernatant was centrifuged at 10,000 rpm for 15 min. Cell-free supernatant was assayed for the amount of siderophore according to CAS assay [18]. Percent siderophore unit (PSU) was calculated according to the following equation [20].
where
Ar= absorbance of reference (CAS solution and un-inoculated broth), and
As= absorbance of the sample (CAS solution and cell-free supernatant of the sample).
2.8. Effects of the rhizobacterial strains on C. villosa and understory accompanying species
All the rhizobacterial strains isolated and identified from the nodules and rootless of C. villosa plants were further screened for their effects on plants of C. villosa. Similarly, and at the same time, the strains were tested on Ornithopus compressus L., Coronilla junccea L., Vicia sativa L. and Trifolium repens L. All the species grow in the understory of C. villosa populations. The former two develop under high level of sun irradiance (open spaces among shrubs), whereas the latter two have better affinity for shaded areas (under the canopy of the shrubs).
Surface sterilized scarified seeds of the five plant species were germinated for 2 days on 1% (w/v) water agar. Ten seedlings per species were planted into a 180 mm pot containing sterile river sand. Seedlings were inoculated with 1ml of one-day old cultures of each strain, grown at 28ºC on glucose minimal salt medium. A 1 cm layer of sterilized plastic beads was placed on the surface of each pot to prevent contamination and to maintain humidity in the pot. Plants were grown in a glasshouse at 25°C located at the University Pablo de Olavide, Seville (Spain). Plants were watered every second day through a PVC tube with nutrient solution containing 3% (w/v) KNO3. The experimental design was completely randomized with five replicates per strain. After 75 days of growth, plants were harvested, separated into shoot and root and oven dried at 72 °C until constant dry weight.
2.9. Data analysis
Data from laboratory and glasshouse experiments were analyzed separately for each experiment using software SPSS 19 for Windows 18th edition developed by VSN International Ltd., UK [21]. Normality and homoscedasticity of data were tested by means of tests given by [22]. A linear mixed model was fitted to each measurement. Data from C. villosa, O. compressus, C. juncea, V. sativa and T. repens growth after treatment with the 12 bacterial strains, were analyzed separately. The 12 strains used were those obtained from the roots and rootless of C. villosa identified in the present study. All measurements of mass were log transformed prior to analysis as residual variance increased with mass. All measurements of mass were log transformed prior to analysis as residual variance increased with mass.
3. Results
3.1. Bacterial identification
All the strains were retrieved from the in vitro inoculated seedlings, thus proving their ability be endophytic bacteria and maybe, to form symbiosis with C. villosa and excluding the hypothesis of some being surface contaminants.
Sequence comparison of 16S ribosomal RNA partial sequences revealed the presence of multiple bacterial species in the nodules and rootles of C. villosa plants (Figure 1). A total of 12 strains were identified with the greatest bacterial richness observed in plants originating from a valley, where plants were well irrigated and at low level of sun irradiance. Strains from these plants reached a maximum of 7 strains in the 4 genera, Bacillus, Brevibacterium, Paenibacillus, and Rhizobium. Bacterial richness was lower in plants growing at the top of a hill, with high sun irradiance and water scarcity. Five strains were identified in three genera, Pseudomonas, Stenotrophomonas and Rhizobium. Two strains were identified in nodules from either C. villosa population. . The remaining strains came from the rootless.
The phylogenetic tree reveals two main clusters of bacteria. Strains on cluster 1 come from plants growing at the top of a hill with high sun irradiance and water scarcity. Isolates in cluster 2 come from plants growing at the bottom of a valley with low sun irradiance and sufficient soil humidity.
3.2. Phenotypical characterization of the bacterial isolates
All the analyzed strains demonstrated the capacity to modify the pH of the growth medium (Table 1). The strains of P. panacis and P. baetica derived from plants growing at the top of the hill induced a pH shift below 6.5, discernible by a yellow hue on the Petri dishes. Similarly, B. megaterium, B. frigotolerans, and P. taichungensis exhibited analogous pH-modifying effects. The remaining strains generated a pH range between 6.5 and 7.6, visualized by a green halo encircling the colonies.
All the strains showed tolerance to salinity, being able to form colonies across all the four tested NaCl concentrations. Pertaining to drought tolerance, the five strains isolated from the plants growing at the hill top formed clear and well-developed colonies. In contrast, only P. Polymyxa and P. taichungensis demonstrated such attributes among the strains isolated from plants growing at the bottom of the valley.
3.3. HCN and NH3 production
Six of the strains were capable of producing cyanic acid, as indicated by the development of an orange to red color on the filter paper placed over the colonies. Among the strains from plants grown at the top of the hill, all but R. cellulositycum produced HCN. Among the strains from plants growing at the bottom of the valley, only two (P. polymyxa and P. taichungensis) exhibited this capability. Regardless of their provenance, all strains were able to produce NH3.
3.4. Assay for indoleacetic acid (IAA) production
Varying levels of IAA production were recorded depending on the bacterial strain (Figure 2). The range of IAA production ranked between 2.26 µg/L in Brevibacterium frigoritolerans and 29.24 µg/L in Stenotrophomonas maltophilia. It is noteworthy that bacterial strains from plants growing on top of the hill were the ones producing the highest concentrations of IAA whereas those concentrations in bacteria collected from plants growing at the bottom of the valley, produced low concentrations of IAA.
3.5. Siderophore production
In CAS assay showed a positive reaction (orange color zone) for siderophore production in all strains but two, Penibacillus taichungensis and one of three strains of Paenibacillus polymyxa (Figure 3). The estimated siderophore production ranged from 298 µg/mL in Pseudomonas panacis to 7 µg/mL in one of the three strains of Paenibacillus polymyxa. Similar to the case of IAA production, the highest concentrations of siderophores were produced by bacteria collected from plants growing at the top of the hill, while considerably lower amounts were measured in bacteria from plants grown at the bottom of the valley.
3.6. Effects of the rhizobacterial strains on C. villosa and understory accompanying species
As a general rule, all bacterial isolates exerted a growth-promoting effect on the five studied plant species (Table 2). However, we also found some significantly deleterious effects at the root level. These deleterious effects were observed in C. villosa following the application of Pseudomonas baetica, P. panacis, B. frigoritolerans, P. taichungensis and the three isolates in the P. polymyxa strains: in O. compressus, C. juncea, and V. sativa following the application of P. panacis. The remaining results from the application of bacterial isolates were positive. Particularly noteworthy for their positive effects are those derived from Rhizobium cellulosilyticum and R. leguminosarum, as well as those from Stenotrophomonas maltophilia, Bacillus megaterium and Brevibacterium frigoritolerans (Table 2). Although increases in plant length were observed, none of the results were statistically significant.
Interestingly, T. repens is the plant species in which significantly higher biomass values were obtained in both roots and shoots compared to the control when treated with all other bacterial isolates, except for Pseudomonas baetica. As a general trend, the bacterial isolates from plants of C. villosa grown at the hilltop exhibited a significantly greater growth enhancement capacity on all studied plant species compared to isolates originating from plants grown at the bottom of the valley (Table 2).
4. Discussion
Leguminous plant species have the ability to accept endophytic bacteria and to establish symbiotic interactions with soil-borne bacteria, and these associations can vary significantly depending on the environmental conditions they encounter [7,23]. Our results evidenced that depending on where plants of C. villosa grow, the associated bacteria to the rootless and nodules vary (Figure 1) [24]. Thus, plants exposed to high irradiance and low water availability had lower bacterial richness associated and have preference for bacteria in the genus Pseudomonas and Strenotrophomonas whereas plants growing at the bottom of a valley, where sun irradiance was lower and water was not limited showed higher bacterial diversity and preferred strains in the genus Bacillus, Brevibacterium and Paenibacillus. The only genus that was present in plants from both provenances was Rhizobium (Figure 1). This sanctions that plants impose in favor of certain bacterial genera can be explained by the Filter Theory [25,26], which suggests that certain microbial communities are filtered out based on specific environmental factors. When these leguminous plants grow in a well-irrigated and shaded environment, the availability of water and reduced sunlight can favor the establishment of bacterial species that are well-suited to such conditions as is has been demonstrated in P. sativum L. [27]. These bacteria may provide benefits such as enhanced nutrient uptake and disease resistance to the plants, helping them thrive in this specific environment. In fact, we have observed that bacteria from plants from the hilltop were able to produce significantly higher amounts of IAA (Figure 2) and siderophores (Figure 3), what are plant growth promoting (PGP) compounds. In addition, and unlikely bacteria isolated from plants at the valley bottom, all of the bacteria isolated at the hilltop were drought tolerant and produce HCN (Table 1). Previous studies in legumes, such as alfalfa, common bean, pea, fava bean, and Medicago sp has demonstrated that the presence of endophytic bacteria producing Plant Growth-Promoting Bacteria (PGPB) contributes to enhanced biomass production, increased survival rates, and improved drought tolerance[28,29,30,31,32].
Endophytic bacteria isolated from plants thriving in the well-irrigated and low sun irradiance environment from the valley bottom exhibit a higher richness compared to their counterparts growing under water stress (hilltop). This observation can be attributed to the availability of abundant water and low sun irradiance, which provide favorable conditions for bacterial growth and proliferation. The increased abundance of endophytic bacteria in these plants can result from the presence of more nutrient resources, promoting a conducive environment for microbial colonization and interaction [33,34] Conversely, plants experiencing water stress undergo physiological changes that impact their symbiotic associations. In response to water scarcity, plants may allocate resources towards stress adaptation mechanisms, leading to a potential reduction in bacterial colonization [30,31,32]. Simultaneously, the water-stressed plants may exhibit an elevated production of hydrogen cyanide (HCN), a compound known for its defensive properties. The heightened HCN production in water-stressed plants serves as an adaptive response to potential challenges posed by herbivores and pathogens in environments where resources are limited. The allocation of resources towards HCN production in water-stressed plants underscores the trade-off between symbiotic investment and stress tolerance.
The intricate interplay between environmental factors and bacterial-plant symbiotic interactions leads to variations in the production of indole-3-acetic acid (IAA) and siderophores. In the context of plants growing in the shaded environment of a valley bottom with sufficient water availability, the low sun irradiation may contribute to reduced photosynthetic activity. Consequently, the availability of photosynthates, which are essential precursors for IAA and siderophore biosynthesis, may be limited. This subdued metabolic activity can be the reason why bacteria isolated from plants growing at the valley bottom show a decrease in IAA and siderophore production.
Conversely, when plants face water stress and high sun irradiance, they often activate stress-responsive pathways that can influence bacterial symbionts [35]. In such conditions, plants may allocate more resources to stress adaptation, including enhanced production of IAA and siderophores. The elevated production of IAA in water-stressed plants may serve as a growth-promoting hormone, aiding in root development and nutrient uptake. Additionally, heightened siderophore production can contribute to enhanced iron uptake under water stress conditions, mitigating potential nutrient limitations what concurs with our results. For example, in legumes such as alfalfa, common bean, pea, fava bean, and Medicago sp., endophytic bacteria have been shown to produce PGP compounds such as siderophores, auxins, and cytokinins, which can enhance plant performance under water stress conditions [28,29,30,31,32]
In addition to that, we have seen that the five leguminous species tested in the present study show similar responses to all the bacterial strains, with positive responses in terms of plant biomass production. Even thought, no significant differences were detected for shoot and root lengths, in most cases bacterial strains induced plant elongation. However, and to certain extent, some of the bacteria exerted a deleterious effect of the herbaceous species. This intriguing phenomenon reflects the intricate arms race between microorganisms and plants, where some bacteria deploy toxins or inhibitory molecules as a survival strategy. While seemingly counter intuitive, such antagonistic interactions play a role in shaping plant communities by exerting selective pressures on plant-microbe associations. In our case, the deleterious effects on plant performance was profusely observed in bacteria coming from the valley bottom. Due to the milder environmental condition for plant growth there is more competition for space and nutrients and bacteria have evolved mechanisms to survive under strong competition [36,37,38].
The distinct responses observed in IAA and siderophore production between plants in the valley bottom and those under water stress highlight the dynamic nature of plant-bacteria interactions. These responses underscore how environmental conditions, particularly sunlight availability and water stress, influence the allocation of resources and metabolic pathways in both plants and associated bacteria. As a result, the varying levels of IAA and siderophore production underscore the adaptability of symbiotic partnerships to different ecological contexts.
The empirical findings derived from our experimental study have substantiated and reinforced the aforementioned hypotheses. Our investigation substantiates the notion that the plants' robustness in the face of heightened irradiance and drought can be, at least partlyattributed to the enriched microbial diversity that the plants host, distinguishing them from their counterparts thriving in shaded ecosystems. This microbial synergy actively fosters plant growth and sustenance by eliciting protective mechanisms that are notably absent in plants inhabiting more temperate settings. Moreover, we propose that the presence of bacteria intricately associated with C. villosa serves as a catalyst for qualitative transformations within the understory species, functioning as potent regulators of plant growth responses triggered by the prevailing environmental conditions.
In summary, the variance in endophyti bacteria abundance between plants of C. villosa at the bottom of the valley and those under water stress underscores the intricate interplay between environmental conditions, plant-bacteria interactions, and stress-related responses. This dynamic relationship highlights the adaptability of both plants and their symbiotic partners to varying ecological contexts, where resource availability and stress tolerance influence symbiont prevalence and secondary metabolite production. Ultimately, the symbiotic relationships formed between leguminous plants and soil-borne bacteria contribute to the plants' ability to thrive in a range of environmental contexts by providing them with valuable support and resources tailored to their specific needs what can be one of the explanations for the broad distribution of C. villosa.
Author Contributions
The three authors have similarly contributed to the manuscript, from its design, laboratory experimentation, data treatment and writing up
Funding
This research has been self-funded with no external support.
Institutional Review Board Statement
Not apply
Data Availability Statement
Data will be made available on request.
Acknowledgments
We thank Dr. Francisco Javier Gallego from the1Department of Genetics, Physiology and Microbiology, Faculty of Biological Sciences, University Complutense, Madrid, Spain for assistance with sequencing of the bacterial strainsI.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Segarra-Moragues, J. G.; Carrión-Marco, Y.; Castellanos, M.C; Molina, M. J.; García-Fayos, P. Ecological and historical determinants of population genetic structure and diversity in the Mediterranean shrub Rosmarinus officinalis (Lamiaceae). Botanical Journal of the Linnean Society 2016, 180, 50–63. [Google Scholar] [CrossRef]
- Turco, M.; Von Hardenberg, J.; AghaKouchak, A.; Llasat, M. C.; Provenzale, A.; Trigo, R. M. On the Key Role of Droughts in the Dynamics of Summer Fires in Mediterranean Europe. Sci Rep 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- García, D.; Martínez, D. Species Richness Matters for the Quality of Ecosystem Services: A Test Using Seed Dispersal by Frugivorous Birds. Proc. R. Soc. B. 2012, 279, 3106–3113. [Google Scholar] [CrossRef] [PubMed]
- Haase, P.; Pugnaire, F. I.; Fernández, E. M.; Puigdefábregas, J.; Clark, S. C.; Incoll, L. D. An Investigation of Rooting Depth of the Semiarid Shrub Retama Sphaerocarpa (L.) Boiss. by Labelling of Ground Water with a Chemical Tracer. Journal of Hydrology 1996, 177, (1–2). [Google Scholar] [CrossRef]
- Thamer, S.; Schädler, M.; Bonte, D.; Ballhorn, D. J. Dual Benefit from a Belowground Symbiosis: Nitrogen Fixing Rhizobia Promote Growth and Defense against a Specialist Herbivore in a Cyanogenic Plant. Plant Soil 2011, 341, (1–2). [Google Scholar] [CrossRef]
- De Lara-Del Rey, I. A.; Pérez-Fernández, M. A. Regulatory Effect of Light and Rhizobial Inoculation on the Root Architecture and Plant Performance of Pasture Legumes. Agronomy 2023, 13. [Google Scholar] [CrossRef]
- Pérez-Fernández, M. A.; Calvo-Magro, E.; Valentine, A. Benefits of the Symbiotic Association of Shrubby Legumes for the Rehabilitation of Degraded Soils under Mediterranean Climatic Conditions: Inoculated Legumes Recover Contaminated Soil. Land Degrad. Develop. 2016, 27, 395–405. [Google Scholar] [CrossRef]
- Thrall, P. H.; Millsom, D. A.; Jeavons, A. C.; Waayers, M.; Harvey, G. R.; Bagnall, D. J.; Brockwell, J. Seed Inoculation with Effective Root-Nodule Bacteria Enhances Revegetation Success: Seed Inoculation with Root-Nodule Bacteria. Journal of Applied Ecology 2005, 42, 740–751. [Google Scholar] [CrossRef]
- Berger, J. D.; Shrestha, D.; Ludwig, C. Reproductive Strategies in Mediterranean Legumes: Trade-Offs between Phenology, Seed Size and Vigor within and between Wild and Domesticated Lupinus Species Collected along Aridity Gradients. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Alonso-Vargas, M.A.; Crespo-Villalva, M.B.; Martínez-Azorín. M.; Pérez-Botella, J. Calicotome villosa (Poir.) Link (Fabaceae), novedad para la flora de la Comunidad Valenciana. Flora Montiberica 2020, 77, 104–107. [Google Scholar]
- Vincent, J. M. A Manual for the Practical Study of Root-Nodule Bacteria; IBP handbook no. 15; [Published for the] International Biological Programme [by] Blackwell Scientific, 1970.
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Molecular Biology and Evolution 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Pérez-Fernández, M. A.; Hill, Y. J.; Calvo-Magro, E.; Valentine, A. Competing Bradyrhizobia Strains Determine Niche Occupancy by Two Native Legumes in the Iberian Peninsula. Plant Ecol 2015, 216, 1537–1549. [Google Scholar] [CrossRef]
- Lorck, H. Production of Hydrocyanic Acid by Bacteria. Physiol. Plant. 1948, 1, 142–146. [Google Scholar] [CrossRef]
- Cappuccino, J.C.; Sherman, N. (1992). A Laboratory Manual, In: Microbiology third ed. Benjamin/cummings Pub. Co., New York, pp. 125–179.
- Bric, J. M.; Bostock, R. M.; Silverstone, S. E. Rapid In Situ Assay for Indoleacetic Acid Production by Bacteria Immobilized on a Nitrocellulose Membrane. Appl Environ Microbiol 1991, 57, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Loper, J.E.; Scroth, M. N. Influence of bacterial sources on indole-3 acetic acid on root elongation of sugarbeet. Phytopathology 1986, 76, 386–389. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J. B. Universal Chemical Assay for the Detection and Determination of Siderophores. Analytical Biochemistry 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Meyer, J. M.; Abdallah, M. A. The Fluorescent Pigment of Pseudomonas fluorescens: Biosynthesis, Purification and Physicochemical Properties. Journal of General Microbiology 1978, 107, 319–328. [Google Scholar] [CrossRef]
- Payne, S. M. [25] Detection, Isolation, and Characterization of Siderophores. In Methods in Enzymology; Elsevier, 1994; Vol. 235, pp 329–344. [CrossRef]
- IBM SPSS Software. Available online: https://www.ibm.com/analytics/spss-statistics-software.
- Cochran, W.G. The distribution of the largest of a set of estimated variances as a fraction of their total. Ann Eugen 1941, 11, 47–61. [Google Scholar] [CrossRef]
- Bever, J. D.; Platt, T. G.; Morton, E. R. Microbial Population and Community Dynamics on Plant Roots and Their Feedbacks on Plant Communities. Annu. Rev. Microbiol. 2012, 66, 265–283. [Google Scholar] [CrossRef]
- Bhattacharyya, P. N.; Jha, D. K. Plant Growth-Promoting Rhizobacteria (PGPR): Emergence in Agriculture. World J Microbiol Biotechnol 2012, 28, 1327–1350. [Google Scholar] [CrossRef]
- Lawton, J.H. A Direct Test of the Theory of Habitat Selection. Ecology 1970, 51, 704–707. [Google Scholar] [CrossRef]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The Importance of the Microbiome of the Plant Holobiont. New Phytol 2015, 206, 1196–1206. [Google Scholar] [CrossRef]
- Prudent, M.; Dequiedt, S.; Sorin, C.; Girodet, S.; Nowak, V.; Duc, G.; Salon, C.; Maron, P. The Diversity of Soil Microbial Communities Matters When Legumes Face Drought. Plant Cell Environ 2020, 43, 1023–1035. [Google Scholar] [CrossRef]
- Amine-Khodja, I. R.; Boscari, A.; Riah, N.; Kechid, M.; Maougal, R. T.; Belbekri, N.; Djekoun, A. Impact of Two Strains of Rhizobium leguminosarum on the Adaptation to Terminal Water Deficit of Two Cultivars Vicia Faba. Plants 2022, 11. [Google Scholar] [CrossRef]
- Antolín, M. C.; Yoller, J.; Sánchez-Díaz, M. Effects of Temporary Drought on Nitrate-Fed and Nitrogen-Fixing Alfalfa Plants. Plant science (Limerick) 1995, 107, 159–165. [Google Scholar] [CrossRef]
- Lodeiro, A. R.; González, P.; Hernández, A.; Balagué, L. J.; Favelukes, G. Comparison of Drought Tolerance in Nitrogen-Fixing and Inorganic Nitrogen-Grown Common Beans. Plant Science 2000, 154, 31–41. [Google Scholar] [CrossRef]
- Staudinger, C.; Mehmeti-Tershani, V.; Gil-Quintana, E.; Gonzalez, E. M.; Hofhansl, F.; Bachmann, G.; Wienkoop, S. Evidence for a Rhizobia-Induced Drought Stress Response Strategy in Medicago truncatula. Journal of Proteomics 2016, 136, 202–213. [Google Scholar] [CrossRef]
- Turetschek, R.; Staudinger, C.; Wienkoop, S. Rhizobial Symbiosis Influences Response to Early Salt and Drought Stress of the Medicago truncatula Root Proteome. In The Model Legume Medicago truncatula; 2020; pp 253–260. [CrossRef]
- Frindte, K.; Pape, R.; Werner, K.; Löffler, J.; Knief, C. Temperature and Soil Moisture Control Microbial Community Composition in an Arctic–Alpine Ecosystem along Elevational and Micro-Topographic Gradients. ISME J 2019, 13, 2031–2043. [Google Scholar] [CrossRef]
- Griffiths, B. S.; Ritz, K.; Wheatley, R.; Kuan, H. L.; Boag, B.; Christensen, S.; Ekelund, F.; Sørensen, S. J.; Muller, S.; Bloem, J. An Examination of the Biodiversity–Ecosystem Function Relationship in Arable Soil Microbial Communities. Soil Biology and Biochemistry 2001, 33, 1713–1722. [Google Scholar] [CrossRef]
- Maire, V.; Gross, N.; Da Silveira Pontes, L.; Picon-Cochard, C.; Soussana, J.-F. Trade-off between Root Nitrogen Acquisition and Shoot Nitrogen Utilization across 13 Co-Occurring Pasture Grass Species. Functional Ecology 2009, 23, 668–679. [Google Scholar] [CrossRef]
- Berg, G.; Smalla, K. Plant Species and Soil Type Cooperatively Shape the Structure and Function of Microbial Communities in the Rhizosphere: Plant Species, Soil Type and Rhizosphere Communities. FEMS Microbiology Ecology 2009, 68, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lebeis, S. L.; Paredes, S. H.; Lundberg, D. S.; Breakfield, N.; Gehring, J.; McDonald, M.; Malfatti, S.; Glavina Del Rio, T.; Jones, C. D.; Tringe, S. G.; Dangl, J. L. Salicylic Acid Modulates Colonization of the Root Microbiome by Specific Bacterial Taxa. Science 2015, 349, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Panke-Buisse, K.; Poole, A. C.; Goodrich, J. K.; Ley, R. E.; Kao-Kniffin, J. Selection on Soil Microbiomes Reveals Reproducible Impacts on Plant Function. ISME J 2015, 9, 980–989. [Google Scholar] [CrossRef] [PubMed]
- Lattanzi, E. The distribution of three species of the genus Calicotome in Italy. Flora Mediterr. 2008, 18, 123–125. [Google Scholar]
- Zahran, H. H. Rhizobia from wild legumes: Diversity, taxonomy, ecology, nitrogen fixation and biotechnology. Journal of Biotechnology, 2001, 91:143–153. [CrossRef]
- Zakhia, F. 2004. Diversity of bacteria hosts of mediterranean legumes in Tunisia and Lebanon. PhD thesis, Universite´ de Montpellier II.
Figure 1.
Phylogenetic tree of the bacterial strains identified in root nodules and rootless of mature plants of Calicotome villosa collected in two populations in the Natural Park of Los Alconocales (Cadiz, Spain). Strains on cluster 1 come from plants growing at the top of a hill with high sun irradiance and water scarcity. Isolates in cluster 2 come from plants growing at the bottom of a valley with low sun irradiance and sufficient irrigation.
Figure 1.
Phylogenetic tree of the bacterial strains identified in root nodules and rootless of mature plants of Calicotome villosa collected in two populations in the Natural Park of Los Alconocales (Cadiz, Spain). Strains on cluster 1 come from plants growing at the top of a hill with high sun irradiance and water scarcity. Isolates in cluster 2 come from plants growing at the bottom of a valley with low sun irradiance and sufficient irrigation.
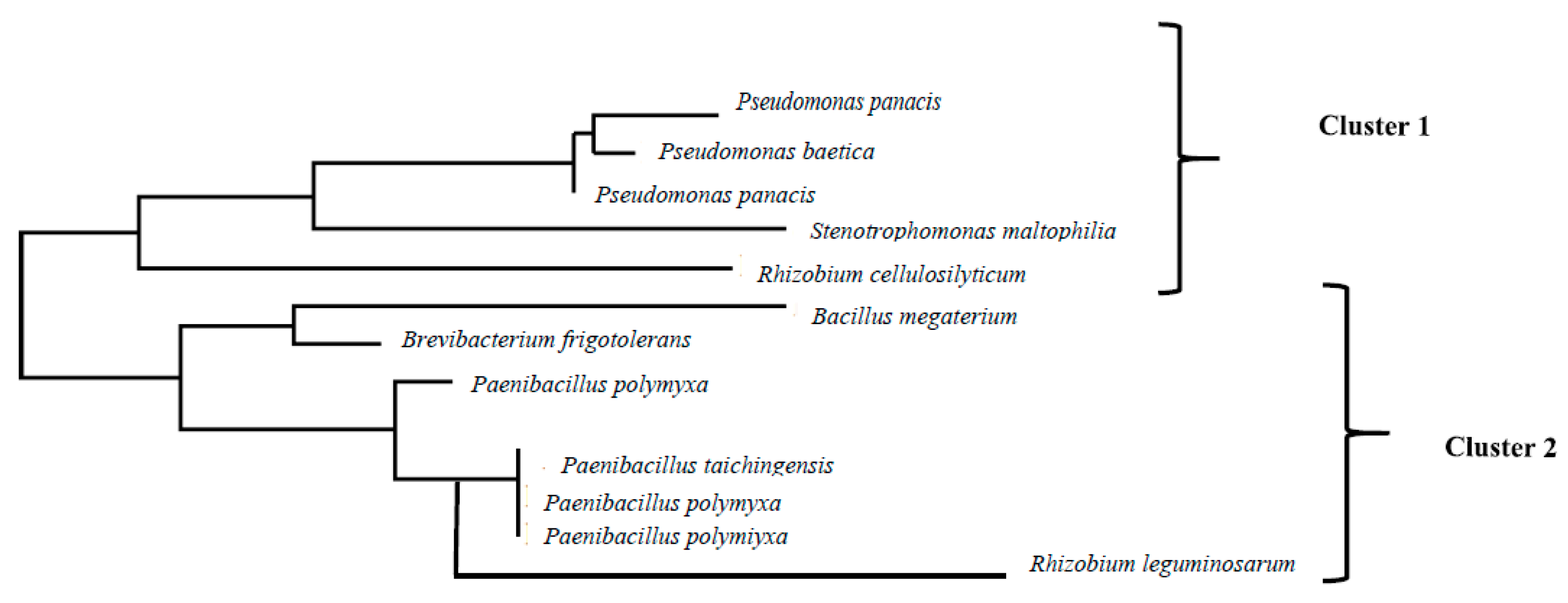
Figure 2.
Concentration of IAA produced by each of the 12 strains isolated from plants of Calicotome villosa growing in two different populations. Green columns represent strains from the population located at the bottom of a valley, characterized by low sun irradiance and sufficient soil humidity. Orange columns represent strains from the population located at the top of a hill, characterized by high sun irradiance and water scarcity.
Figure 2.
Concentration of IAA produced by each of the 12 strains isolated from plants of Calicotome villosa growing in two different populations. Green columns represent strains from the population located at the bottom of a valley, characterized by low sun irradiance and sufficient soil humidity. Orange columns represent strains from the population located at the top of a hill, characterized by high sun irradiance and water scarcity.
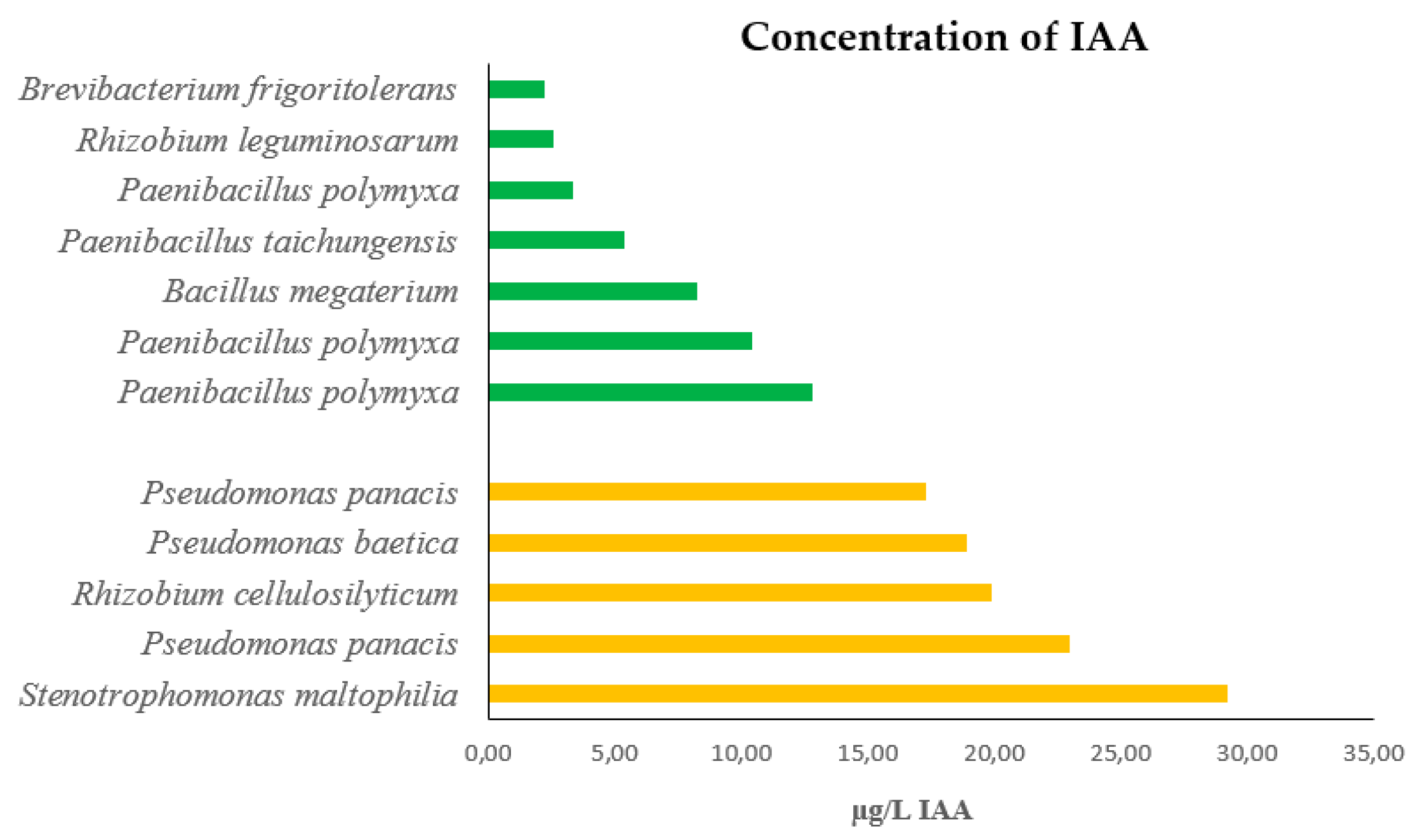
Figure 3.
Concentration of siderophores produced by each of the 12 strains isolated from plants of Calicotome villosa growing in two different populations. Clear columns represent strains from the population located at the bottom of a valley, characterized by low sun irradiance and sufficient soil humidity. Dark columns represent strains from the population located at the top of a hill, characterized by high sun irradiance and water scarcity.
Figure 3.
Concentration of siderophores produced by each of the 12 strains isolated from plants of Calicotome villosa growing in two different populations. Clear columns represent strains from the population located at the bottom of a valley, characterized by low sun irradiance and sufficient soil humidity. Dark columns represent strains from the population located at the top of a hill, characterized by high sun irradiance and water scarcity.

Table 1.
Phenotypical characterization of the 12 bacterial isolates isolated from nodules and rootless from Calicotome villosa plants growing in two natural populations in the Natural Park of Los Alcornocales (Cádiz). The upper part of the table covers isolates from plants growing at the top of the hill with high irradiance and water scarcity. The lower part of the table covers isolates from plants growing at the bottom of a valley with low irradiance and well irrigated.
Table 1.
Phenotypical characterization of the 12 bacterial isolates isolated from nodules and rootless from Calicotome villosa plants growing in two natural populations in the Natural Park of Los Alcornocales (Cádiz). The upper part of the table covers isolates from plants growing at the top of the hill with high irradiance and water scarcity. The lower part of the table covers isolates from plants growing at the bottom of a valley with low irradiance and well irrigated.
| pH | Tolerance to salinity | Resistance to Drought | HCN | NH3 | ||||
|---|---|---|---|---|---|---|---|---|
| Bacterial strain | 1% | 3% | 7% | 10% | ||||
| Pseudomonas panacis | Yellow | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Pseudomonas baetica | Yellow | Yes | Yes | Yes | Yes | Yes | Yes | |
| Pseudomonas panacis | Yellow | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Stenotrophomonas maltophilia | Green | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Rhizobium cellulosilyticum | Green | Yes | Yes | Yes | Yes | Yes | No | Yes |
| Bacillus megaterium | Yellow | Yes | Yes | Yes | No | Yes | Yes | Yes |
| Brevibacterium frigoritolerans | Yellow | Yes | Yes | Yes | Yes | No | No | Yes |
| Paenibacillus polymyxa | Green | Yes | Yes | Yes | Yes | No | No | Yes |
| Paenibacillus taichungensis | Yellow | Yes | Yes | Yes | No | No | No | Yes |
| Paenibacillus polymyxa | Green | Yes | Yes | No | No | No | No | Yes |
| Paenibacillus polymyxa | Green | Yes | Yes | No | No | Yes | No | Yes |
| Rhizobium leguminosarum | Green | Yes | Yes | Yes | Yes | No | Yes | Yes |
Table 2.
Promoting and deleterious effects of 12 bacterial isolates isolated from nodules and rootless from Calicotome villosa plants growing in two natural populations in the Natural Park of Los Alcornocales (Cádiz) on plant performance. The upper part of the table covers isolates from plants growing at the hill top with high irradiance and water scarcity. The lower part of the table covers isolates from plants growing at the bottom of a valley with low irradiance and well irrigated. The results in the table have been calculated by subtracting the value obtained after treatment with each isolate for each variable from the control value. Positive values (green) indicate a significantly beneficial effect of the bacterium while negative values (orange) indicate a significantly detrimental effect of the bacterium.
Table 2.
Promoting and deleterious effects of 12 bacterial isolates isolated from nodules and rootless from Calicotome villosa plants growing in two natural populations in the Natural Park of Los Alcornocales (Cádiz) on plant performance. The upper part of the table covers isolates from plants growing at the hill top with high irradiance and water scarcity. The lower part of the table covers isolates from plants growing at the bottom of a valley with low irradiance and well irrigated. The results in the table have been calculated by subtracting the value obtained after treatment with each isolate for each variable from the control value. Positive values (green) indicate a significantly beneficial effect of the bacterium while negative values (orange) indicate a significantly detrimental effect of the bacterium.
| C. villosa | O. compressus | V. sativa | ||||||||||
| Shoot length (cm) | Shoot length (cm) | Shoot mass (mg) | Root mass (mg) | Shoot length (cm) | Root length (cm) | Shoot mass (mg) | Root mass (mg) | Shoot length (cm) | Root length (cm) | Shoot mass (mg) | Root mass (mg) | |
| Pseudomonas panacis | 2.95 | 2.93 | 54.09 | 64.09 | 3.07 | 5.90 | 110.17 | -227.45 | 2.65 | 2.38 | 156.45 | -75.81 |
| Pseudomonas baetica | -1.40 | 0.40 | 80.09 | -87.40 | -2.05 | -1.80 | 148.90 | 80.00 | -6.22 | -2.39 | 211.45 | 26.67 |
| Pseudomonas panacis | 0.02 | -2.08 | 132.54 | -89.42 | -4.20 | -1.20 | 226.93 | 67.25 | -3.86 | 0.31 | 322.25 | 22.42 |
| Rhizobium cellulosilyticum | 7.60 | 4.90 | 1977.23 | 331.75 | 8.35 | 14.06 | 1483.67 | 835.23 | 7.02 | 13.43 | 2106.82 | 278.41 |
| Stenotrophomonas maltophilia | 8.72 | 4.29 | 911.82 | 439.61 | 10.11 | 21.89 | 493.67 | 251.23 | 20.60 | 14.53 | 701.02 | 83.75 |
| Bacillus megaterium | 6.98 | 3.27 | 613.85 | 199.73 | 2.28 | 11.44 | 943.03 | 632.26 | 15.80 | 10.13 | 1339.11 | 210.76 |
| Brevibacterium frigoritolerans | 0.30 | 0.51 | 280.24 | -52.77 | 0.48 | 0.53 | 446.70 | 299.49 | 0.45 | 0.45 | 634.32 | 99.83 |
| Paenibacillus polymyxa | 0.53 | 0.55 | 20.33 | -102.96 | 0.30 | 0.89 | 59.99 | -18.57 | -0.34 | 0.19 | 85.19 | -6.19 |
| Paenibacillus taichungensis | 0.02 | -0.20 | 42.53 | -94.28 | -0.22 | 0.09 | 93.01 | 36.44 | -0.20 | -0.61 | 132.08 | 12.15 |
| Paenibacillus polymyxa | -0.14 | 0.58 | 11.52 | -101.79 | -0.50 | -0.31 | 46.88 | -11.17 | -0.42 | 1.72 | 66.58 | -3.71 |
| Paenibacillus polymyxa | -0.97 | -0.29 | 33.04 | -103.73 | -0.82 | 0.33 | 78.90 | -23.45 | -0.28 | 0.14 | 112.04 | -7.81 |
| Rhizobium leguminosarum | 6.70 | 3.80 | 1597.17 | 797.10 | 9.42 | 6.28 | 918.21 | 615.62 | 15.65 | 12.34 | 1303.88 | 205.21 |
| C. juncea | T. repens | |||||||||||
| Shoot length (cm) | Shoot length (cm) | Shoot mass (mg) | Root mass (mg) | Shoot length (cm) | Root length (cm) | Shoot mass (mg) | Root mass (mg) | |||||
| Pseudomonas panacis | 2.45 | 2.18 | 151.27 | -33.06 | 4.65 | 2.83 | 10.28 | 10.12 | ||||
| Pseudomonas baetica | -6.42 | -2.59 | 185.64 | 30.99 | 3.07 | 5.90 | -2.91 | -2.03 | ||||
| Pseudomonas panacis | -4.06 | 0.11 | 254.89 | 28.34 | -2.05 | -1.80 | 350.60 | 184.72 | ||||
| Rhizobium cellulosilyticum | 6.82 | 13.23 | 1870.25 | 1188.33 | -4.20 | -1.20 | 406.33 | 175.61 | ||||
| Stenotrophomonas maltophilia | 20.40 | 14.33 | 491.63 | 66.67 | 8.35 | 14.06 | 1404.01 | 824.17 | ||||
| 10.11 | 21.89 | 696.86 | 407.02 | |||||||||
| Bacillus megaterium | 15.60 | 9.93 | 1290.43 | 146.05 | -4.65 | -2.83 | -10.28 | -10.12 | ||||
| Brevibacterium frigoritolerans | 0.25 | 0.25 | 481.94 | 76.72 | 2.28 | 11.44 | 1017.83 | 679.19 | ||||
| Paenibacillus polymyxa | -0.54 | -0.01 | 106.73 | 10.46 | 0.48 | 0.53 | 663.31 | 441.50 | ||||
| Paenibacillus taichungensis | -0.40 | -0.81 | 136.04 | 21.92 | 0.30 | 0.89 | 387.09 | 214.31 | ||||
| Paenibacillus polymyxa | -0.62 | 1.52 | 95.10 | 12.01 | -0.22 | 0.09 | 410.68 | 253.60 | ||||
| Paenibacillus polymyxa | -0.48 | -0.06 | 123.51 | 9.44 | -0.50 | -0.31 | 377.73 | 219.59 | ||||
| Rhizobium leguminosarum | 15.45 | 12.14 | 2668.41 | 1142.58 | -0.82 | 0.33 | 400.60 | 210.82 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



