
Submitted:
22 August 2023
Posted:
22 August 2023
You are already at the latest version
Abstract
Sepsis-associated encephalopathy (SAE) is a common brain dysfunction following sepsis that often results in severe cognitive and neurological sequelae and increases the mortality rate in patients with sepsis. Microglia are resident macrophages in the brain that play essential roles in the pathological and physiological processes of SAE. Depending on the nature of the stimulus, microglia can adopt two polarization states (M1/M2), which correspond to altered microglial morphology, gene expression, and function. Therefore, we systematically described the pathogenesis, morphology, function, and phenotype of microglial activation in SAE. We demonstrated that microglia are closely related to occurrence and development of SAE and concomitant cognitive impairement. Finally, we outlined some potential therapeutic approaches that can prime microglia and neuroinflammation towards the beneficial restorative microglial phenotype in SAE.
Keywords:
Sepsis-associated encephalopathy
; Microglial activation
; M1/M2 microglial polarization
; cognitive impairment
; therapeutic strategies
1. Introduction
Sepsis is a common cause of death among critically-ill patients in intensive care units. According to the latest global estimates of sepsis incidence and mortality, 49 million people suffer from sepsis annually, resulting in 11 million deaths, which is 20% of all deaths worldwide [1]. Sepsis is now defined as infection with organ dysfunction which codifies organ dysfunction using the Sequential Organ Failure Assessment (SOFA) score [2]. The World Health Assembly and World Health Organization (WHO) made sepsis a global health priority in 2017 and have adopted a resolution to improve the prevention, diagnosis, and management of sepsis [3]. A common complication of sepsis is diffuse brain dysfunction and cognitive impairment caused by infection outside the central nervous system (CNS), known as sepsis-associated encephalopathy (SAE) [4]. It frequently occurs in the absence of overt infection of the CNS, which manifest as mild disturbance of consciousness, disorientation, cognitive impairment, convulsion or deep coma [5].
The pathogenesis of SAE is likely multifactorial and has not been fully elucidated, treating and diagnosing SAE are also equally challenging tasks. Diagnosis of SAE in clinical practice occurs primarily through the detection of abnormalities in electroencephalogram recordings and abnormal mental status, along with clinical history, physical examination, laboratory tests, and neuroimaging evaluation [6]. The etiology and pathogenesis of SAE are complex, including microglial activation, blood-brain barrier (BBB) disruption, leukocyte infiltration, metabolic adaptations to systemic inflammation, bioenergetic shifts, cerebral coagulopathy or ischemia, oxidative stress due to inflammation, and mitochondrial dysfunction [7,8] . A localized and significant increase in CD68 positive microglia is observed in the brains of patients who die from septic shock due to severe systemic inflammation and increased microglial activity in the putamen, hippocampus, and cerebellum of the brain [9,10]. It was found that patients with sepsis had activation of pro-inflammatory microglia in their white matter, but not so much in their gray matter. In contrast to brain inflammatory or ischemic diseases, the anti-inflammatory microglia markers CD163 and CD206 were not expressed in acute sepsis [11]. Microglial activation can cause neuronal injury or apoptosis by releasing inflammatory mediators, reactive oxygen species, neurotransmitters, and other substances. Microglia also secrete cytokines and chemokines that protect the brain from inflammatory responses. However, because of the regulation of white blood cell migration and neuronal repair, the long-term activation of microglia has minimal protective effects on neurons and worsens inflammatory response in the brain [12,13]. In this review, we systematically investigated the critical role of microglia in SAE and summarized the prospects of therapies targeting microglial activation and neuroinflammation to alleviate cognitive impairment in SAE in recent years.
2. Microglia in homeostasis
Brain development and CNS homeostasis are normally regulated by microglia, which include programmed cell death, clearing apoptotic newborn neurons, and pruning axons and synapses that are developing. Throughout development and into adulthood, microglia processes are highly mobile and continually survey their local environment, contacting neurons, axons, and dendritic spines [14]. And microglia modulate synaptic transmissions and facilitate neural circuit formation by devouring eliminated synapses in a complement-dependent manner [15,16]. The human brain functions akin to a city that adheres to natural laws, wherein cell death and neogenesis are constant processes. Microglia, as the primary cleaners of the brain, engage in phagocytosis to eliminate dead neurons and minimize the accumulation of debris , which are derived from yolk-sac-derived progenitors, constitute about 10% of brain cells and approximately 20% of all glial cells [17]. They are the macrophage-like myeloid innate immune cells of the brain and spinal cord, act as the main immune defense in the CNS, and are rapidly activated in most neurological diseases, including traumatic brain injury, stroke, Alzheimer's disease, Parkinson's disease, multiple sclerosis, and schizophrenia et.al. [18,19].
Owing to blood-brain barrier tight junctions between endothelial cells, the basal lamina of these endothelial cells, and astrocytic end-feet processes, which significantly reduce the infiltration of macromolecules and immune cells to the parenchyma, the CNS has traditionally been considered immune-privileged. In addition, the brain lacks professional antigen-presenting cells and expresses low levels of major histocompatibility complex (MHC) class I and II [20]. Microglial activation can occur even with minimal disturbance, maintaining the homeostasis of the local brain parenchyma. Under physiological conditions, microglial processes are motile and exhibit a ramified morphology in the brain of healthy adults. However, microglia become activated and transform into a hypertrophic or ameboid shape in neurodegenerative diseases and upon neuronal injury [21,22]. Like macrophages, microglia respond to invading pathogens by sequestering and inoculating microbes and limiting the effects of cell damage and necrosis [23,24]. These acute responses include migration, proliferation, phagocytosis, antigen presentation, and the release of various effector substances, including superoxide, nitric oxide, proteases, anti-inflammatory (such as [interleukin] IL-10 and IL-4) and pro-inflammatory cytokines (such as IL-1β and IL-6) [25,26].
3. M1/M2 microglial polarization
Microglial activation is a dynamic process and may involve various temporally, physiologically, and/or spatially regulated events that govern the morphological and functional changes observed in the reactive microglia. The latest research found that microglia are most diverse in the developing, aged, and injured brain by mapping single cells of microglia in mice at various stages of development and after brain injury [27]. Depending on the milieu and factors that stimulate them, microglia can participate in classical activation, alternative activation, or acquired deactivation. Under physiological conditions, microglia were designated “resting” while the reactive morphology was termed “activated” (rounder cell body, with fewer and shorter processes, or amoeboid-shaped cell)[28]. To model this change, typical experiments involve the exposure of microglial cells in vitro to stimuli such as apoptotic cells, lipopolysaccharide, inflammatory cytokines, or aggregated proteins [29]. Microglia can be phenotypically polarized to develop either a classical (proinflammatory, M1) or an alternative (anti-inflammatory and pro-healing, M2) phenotype which in Figure 1. M1 microglia dominate the injury site at the end stage of the disease when M2 microglia's immune resolution and repair processes are impaired. M1 microglia produce cytokines and chemokines (IL-1β, IL-6, IL-12, tumor necrosis factor α [TNF-α], and chemokine (C-C motif) ligand 2), express nicotinamide adenine dinucleotide phosphate oxidase, and generate reactive oxygen and nitrogen species. Moreover, M1 microglia express major histocompatibility complex II, CD11b, CD11c integrins, CD36, CD45, and CD47 costimulatory molecules. M2 microglia are capable of releasing several anti-inflammatory cytokines (IL-10, transforming growth factor β [TGF-β]), growth factors (insulin-like growth factor, fibroblast growth factor, and colony-stimulating factor 1), neurotrophic growth factors (nerve-derived growth factor, brain-derived neurotrophic factor, neurotrophins, and glial cell–derived neurotrophic factor)[12].
However, some scholars have different opinions on dichotomic rigid categories of M1/M2, which is inconsistent with the wide repertoire of microglial states and functions in development, plasticity, aging, and diseases which were elucidated in recent years [30]. The advent of single-cell technologies has provided clear evidence that microglia in the living brain do not polarize to either of these categories, often co-expressing M1 and M2 markers [31]. At the molecular level, recent single-cell transcriptome analyses also revealed that human microglia show multiple clusters, indicating greater heterogeneity than in other mammalian species, such as mouse [32]. Therefore, this perspective assumes that the M1/M2 categories of macrophage activation are not essential and destructive for organizing our thinking about microglia. Implicit in that opinion is the mandate for a new microglial terminology, based on considerations including (but not limited to) transcriptomic and proteomic profiles, regional heterogeneity, sexual dimorphism, functions in the intact, healthy nervous system from fertilization to aging, and patterns of response to change, such as physical trauma, infection, systemic inflammation, tumor, ischemia, and neurodegeneration [30,33].
4. Microglia as Central Actors in SAE
4.1. Experimental techniques
A better understanding of how microglial activation contributes to SAE may improve its treatment (An abridged diagram in Figure 2). Animal models of sepsis are typically divided into two types: intraperitoneal injection of lipopolysaccharide (LPS), cecal ligation perforation (CLP) and peritoneal contamination and infection (PCI). The CLP model has been widely adopted as a sepsis animal model, with a well-recognized reliability and clinical relevance. As well, we acknowledge that animal models of sepsis have limitations and may not reflect the high complexity of the sepsis condition in humans. As for cell model, Primary microglial cells are the best candidates for microglial research. Expression profiling has documented drastic differences between the microglia isolated immediately ex vivo and in vitro cultured primary microglia, and widely used cell lines such as BV-2[34]. Under different microenvironments, microglia undergo phenotypic changes, including M1/M2 polarization and other phenotypes such as pyroptosis, autophagy, and apoptosis. The SAE model established using CLP can also cause microglial overactivation and neuronal pyroptosis, aggravating brain tissue destruction and cognitive dysfunction [35]. Consistently, the body areas of microglia cell that represent the morphological alterations of activated microglia, was increased in septic mice [36].
Historically, microglial function has been studied primarily in the mouse and in mouse models of disease. In order to fully understand which mouse findings apply to humans, and whether microglia-targeted therapeutic approaches can be applied to human CNS disorders, it is essential that we invest in developing new innovative technologies including human induced pluripotent stem cells (iPSCs), organoids, two-photon imaging, whole-genome transcriptomic and epigenomic analyses with complementary bioinformatics, unbiased proteomics, cytometry by time-of-flight cytometry, and complex high-content experimental models such as slice culture and zebrafish [37,38].
4.2. Crosstalk between microglia, neurons, and astrocytes
Recent studies have shown that microglia interact with neurons astrocytes. Contacts between microglia and surrounding neurons are interesting: one microglial cell can touch several neurons, and several microglia cells can reach a neuron [39]. As soon as microglia are activated, they release inflammatory mediators, reactive oxygen species, neurotransmitters, and other substances that can injure or even cause apoptosis in neurons [40]. CLP model induced a gradual shift of 50% of surveillant microglia onto amoeboid hypertrophic-like and gitter cell-like reactive phenotypes with active phagocytosis and frequent contact with damaged neurons [41]. And complement-dependent synaptic pruning by microglia as a crucial pathomechanism for the development of neuronal defects during SAE, stereotactic intrahippocampal injection of anti-C1q complement antibody could prevent microglial engulfment of C1q-tagged synapses [42].
Microglia also communicate with astrocytes and coordinate their responses to neuronal damage. Liddelow et al. found that a subtype of reactive astrocytes, termed A1, is induced by classically activated neuroinflammatory microglia by secreting Il-1α, TNF, and C1q, and that these cytokines are necessary and sufficient to induce A1 astrocytes [43]. A microglia-astrocyte circuit mediated by the IL-33-ST2-AKT signaling axis supports microglial metabolic adaptation and phagocytic function during early development, with implications for neurodevelopmental and neuropsychiatric disorders [44]. It has been suggested in conventional view that debris from dead neurons triggers glia-mediated neuroinflammation, increasing neuronal death. But additionally, the expression of neurotoxic proteins in microglia alone is sufficient to trigger the death of naive neurons and propagate neuronal death through activation of naive astrocytes to the A1 state, and the propagation of injury is largely mediated by fragmented and dysfunctional microglial mitochondria [45]. Interferon gamma (IFNγ) remarkably increased the LPS-mediated release of TNFα and IL-1α in microglia and consequently induced the transformation of astrocytes to the A1 subtype, which ultimately resulted in neuronal damage. In addition, IFN-γ promotes cognitive impairment in endotoxemia by enhancing microglia-induced A1 astrocytes. Targeting IFN-γ is a novel strategy for preventing or treating cognitive dysfunction in patients with SAE [46]. The nucleotide-binding oligomerization domain, Leucine rich Repeat and Pyrin domain containing (NLRP3) inflammasome of microglia promotes the transformation of A1 astrocytes and consequently exacerbates neo-neuron decline, resulting in cognitive impairment after challenge with LPS [47]. Berberine targets HMGB1/RAGE signaling to inhibit microglia-stressed A1 astrocytes and neuronal decline, which consequently alleviates sepsis-induced cognitive impairment [48]. One and three days after LPS injection, ionized calcium binding adaptor molecule 1 (IBA1+) microglia in the periventricular white matter (PWM) damage were significantly increased in cells that generated excess amounts of IL-1α, TNF-α, and C1q. Melatonin effectively alleviates PWM damage in septic neonatal rats, which is most likely through the modulation of astrocyte phenotypic transformation from A1 to A2 via the MT1/JAK2/ STAT3 pathway [49].
4.3. Microglia activation in cognitive impairment
Microglial phenotypes change according to the stages and severity of the disease, which plays an essential role in SAE. Recent evidence indicates that many sepsis survivors develop long-term disabilities, including functional and cognitive impairments that affect their quality of life and return to activities of daily living. Cognitive impairment is a major post-sepsis sequela that affects 12.5‒21% of sepsis survivors [50] and can be progressive and permanent, although some patients may only present with transient problems. Memory, attention, and executive function are the cognitive domains most affected in sepsis survivors [51,52]. There is evidence that microglial activation is associated with delirium, and the expression of microglial markers CD68 and HLA-DR was significantly elevated in patients with delirium compared with that in controls in a postmortem case-control study [53]. Both humans and mice with SAE had large increase in chemokines for myeloid cell recruitment, and intravital imaging of the brains of septic mice revealed increased neutrophil and CCR2+ inflammatory monocyte recruitment (the latter being far more robust), accompanied by subtle microglial activation [54].
Mice subjected to CLP were lethargic, with reduced activity, slow movement, erect back hair, thick eyelid secretions, fecal adhesions at the anus, and cloudy urine. Dissection of the dead mice showed that the abdominal cavity was filled with turbid exudate, and the intestinal tract was adherent [55]. Activated microglia and reactive astrogliosis, which are the hallmarks of brain injury and may contribute to synaptic deficits, were observed in septic mice. IL-1β derived from activated microglia is the key molecule responsible for the hippocampal synaptic deficits observed in sepsis [51]. Most microglia are reportedly distributed around cerebral vessels 4 h after LPS injection. The extent of microglial activation was time-dependent, and the highest number of microglia was observed after 8 h in all brain regions [56]. Minocycline induced a downregulation, predominantly, of M1 markers [9], and high doses of minocycline prevented long-term potentiation impairment during sepsis [57]. Ketone body β-hydroxybutyrate (BHB) is produced in the liver and serves as an alternative energy source for the brain, heart, and skeletal muscles in mammals during states of energy deficits. Subcutaneous administration of BHB increased survival and body weight recovery in sepsis mice and improved learning and memory in sepsis surviving mice. The improvement of learning and memory in sepsis surviving mice was still detected even if BHB was administered at the late stage of sepsis [58].
5. Pharmacological intervention targeting microglia
In 2017, World Health Organization (WHO) declared sepsis a global health priority and the greatest unmet medical need of our time, and surviving sepsis campaign has been introduced since 2002. Sepsis survivorship, which is associated with substantial long-term morbidity and a high burden of cognitive dysfunction and disability, is a common and rapidly growing public health problem [59]. At present, there is a lack of specific and effective management methods for the diagnosis and treatment of SAE in modern medicine; Apart from early antimicrobial therapy, adequate tissue/organ perfusion, and timely source control at the early stage of sepsis, there is no specific method for preventing SAE or cognitive impairment following sepsis [60]. Therefore, there is an urgent need for novel anti-SAE therapeutic strategies. Microglial activation blocking or alleviating neurotoxic reactions after microglial activation is an important therapeutic target in anti-SAE therapy, and mastering stage-specific switching of M1/M2 phenotypes in appropriate time windows may produce therapeutic effects [61]. Summary of treatments in experimental models of SAE targeting microglia and neuroinflammation are found in Table 1.
5.1. Blockers of inflammatory factors and pyroptosis
The release of inflammatory factors and mediators is closely related to the pathophysiological process of SAE. Preemptive administration of IL-1 receptor antagonist (IL-1Ra) significantly reduced plasma cytokines and hippocampal microgliosis and ameliorated cognitive dysfunction, which suggested that blocking IL-1 signaling attenuated the inflammatory cascade in response to LPS, thereby reducing microglial activation and preventing behavioral abnormalities [62]. Metformin can partially reverse the severe prognosis caused by sepsis by blocking microglial proliferation and inhibiting the production of inflammatory factors [72]. Cortistatin-14 is a neuropeptide structurally resembling somastostatin, which relieves anxiety-related behaviors in CLP mice, decreases the levels of various inflammatory cytokines, reduces sepsis-induced BBB disruption, and inhibits microglial activation [70]. Dexmedetomidine is an α2 adrenergic agonist used as a sedative in critically ill patients, which can decrease the expression of TNF-α from microglia and alter the neuroinflammatory response of microglia [75].
Microglia are the main cells where pyroptosis occurs in the CNS, and microglial pyroptosis-mediated neuroinflammation is a prominent pathogenesis of SAE [76]. NLRs recognize multiple stimuli and form complexes that cleave pro-Caspase-1 into activated Caspase-1. Activated Caspase-1 cleaves the pore-forming protein gasdermin D (GSDMD), pro-IL-1β, and pro-IL-18, leading to pyroptosis and secretion of IL-1β [77]. The caspase-1 inhibitor VX765 inhibited caspase-1, suppressed the expression of GSDMD and its cleavage form GSDMD-NT, and reduced pyroptosis in the brain at days 1 and 7 after sepsis. Meanwhile, the inhibition of caspase-1 mitigated the expression of IL1β, monocyte chemoattractant protein-1, and TNF-α in the serum and brain, diminished cognitive dysfunction and depressive-like behaviors and microglial activation in septic mice, and reduced sepsis-induced BBB disruption and ultrastructural damage in the brain as well [65]. As an NLRP3 inflammasome inhibitor, MCC950 inhibits SAE-induced NLRP3 expression, IL1β and IL18 cytokine release, neuronal apoptosis, and mitochondrial dysfunction. Moreover, hydrogen alleviates inflammation, neuronal apoptosis, and mitochondrial dysfunction by inhibiting the nuclear factor erythroid 2-related factor 2 (Nrf2)-mediated NLRP3 pathway [67]. Bromo- and extra-terminal protein inhibitor JQ1 protected the hippocampal BBB and neuronal damage by attenuating neuroinflammation via the inhibition of the inflammasome-dependent canonical pyroptosis pathway induced by LPS injection in mice, and also selectively suppressed the activation of hippocampal microglia [71]. The cannabinoid type 2 receptor (CB2R) -specific agonist HU308 has protective effects against SAE by inhibiting microglial activity and neuronal pyroptosis [35].
5.2. Signaling pathway inhibitors
Toll-like receptor 4 (TLR4) plays an essential role in promoting M1 polarization. Upon exposure to signals released at the site of inflammation, the transmembrane receptor TLR4 activates microglia and triggers the upregulation of pro-inflammatory genes [78]. Natural compounds targeting TLR4 may serve as important pharmacophores for developing potent drugs for treating SAE via the TLR4/MyD88/NF-κB signaling pathway in microglia [79]. Apart from the above, there is a possible association between microglial polarization and the mammalian target of rapamycin (mTOR)-autophagy signaling pathway [80]. mTOR and autophagy regulation can modulate microglial polarization and alleviate cerebral inflammation in neurodegenerative diseases and SAE [81]. Hydrogen gas can alleviate SAE by switching microglial polarization mediated by the mTOR-autophagy signaling pathway [82].
Interleukin-10 (IL-10) axis play a critical role in restoring murine microglia homeostasis, LPS-challenged mice harboring IL-10 receptor-deficient microglia displayed neuronal impairment and succumbed to fatal sickness [83]. Moreover, blocking IL-1β signaling ameliorates the inflammatory cascade in response to LPS and behavioral abnormalities were reduced by microglial activation [62]. A study indicated that chemokine receptor 5 (CXCR5) contributes to cognitive impairment in SAE mice by enhancing p38MAPK/NF-κB/STAT3 signaling, and CXCR5 knockout restores autophagy, polarizes microglia to the M2 phenotype, and inhibits p38MAPK and proinflammatory cytokine release in the hippocampus [84]. Indole-3-guanylhydrazone hydrochloride (LQM01) is an aminoguanidine derivative that has been described as an anti-inflammatory, antihypertensive, and antioxidant agent with potential applicability in inflammatory diseases. Heimfarth et al. found that LQM01 exposure during the neonatal period reduced anxiety-like behavior and cognitive impairment caused by LPS in adult life and attenuated inflammatory reactions and oxidative damage through the MAPK and NF-κB signaling pathways and microglial activation suppression [66]. Overall, this supports the critical role these signaling pathways can play in the progression of Microglial activation through SAE.
5.3. Mitochondrial targeting drugs
In recent years, mitochondrial targeting drugs have attracted much more attention. The mitochondrial-targeted antioxidant peptide SS-31 can reduce inflammation and oxidative stress through the inhibition of mitochondrial fission protein 1 ( Fis1) expression in LPS-stimulated microglia [85] . SS-31 improved the cognitive performance and survival rate of septic mice; alleviated hippocampal inflammation, reactive oxygen species production, and excessive mitochondrial fission; and effectively decreased the activation of NLRP3 inflammasome, mitochondrial translocation of dynamin-related protein 1 (Drp1), excessive mitochondrial fission, and mitochondrial membrane recruitment of GSDMD N-terminal in vivo microglial cell line BV-2 stimulated by LPS [73]. Recombinant human STC-1 (rhSTC1) suppressed pro-inflammatory cytokine production in LPS-stimulated microglia. In addition, rhSTC1 injected into the cisterna magna decreased acute hippocampal inflammation and oxidative stress and increased the activity of complex I and II of the mitochondrial respiratory chain and creatine kinase 24 h after sepsis; this was effective in preventing long-term cognitive impairment after CLP [69]. P110 as an inhibitor of Drp1-Fis1 interaction, which can eliminate BBB permeability and loss of tight-junctions after acute LPS injury [86].
Mitochondrial-targeted antioxidants may also serve as a novel strategy to overcome septic complications [87]. Drp1 inhibitor Mdivi-1 protected hippocampus from oxidative stresses and reduced TUNEL-positive cells in hippocampus [88] . Malvidin prevented cerebrum from LPS-induced mitochondrial dysfunction with enhanced JC-1 aggregates and Adenosine triphosphate (ATP) levels, and ROS accumulation with decreased lipid peroxidation and increased antioxidant enzymes [89].
5.4. Traditional Chinese medicine
Traditional Chinese medicine (TCM) focuses on the overall treatment of an individual, and Chinese herbal medicine is characterized by treatments with reduced side effects. The different herbs in the TCM formula are considered to have synergistic effects or reduce side effects, that is, the characteristic of “Jun Chen Zuo Shi” in TCM formulas [90]. Ginsenoside Rg1 (Rg1), a major component of ginseng, improved the postoperative survival rate and protected against learning and memory impairments in SAE (tested using the Morris water maze). In addition, Rg1 attenuated brain histopathological changes (hematoxylin and eosin staining), suppressed IBA1 activation, and decreased the expression of inflammatory cytokines [63]. Atractylon (Atr) is a major sesquiterpene compound belonging to the Asteraceae family, which attenuated LPS-induced cognitive impairment, neural apoptosis, inflammatory factors, and microglial activation, while Atr-induced silent information regulator 1 expression promoted BV2 from LPS-induced M1 to M2 phenotype in vitro [64].
Qiang Xin 1 (QX1) dramatically inhibited short- and long-term excessive pro-inflammatory cytokine production both peripherally and centrally and was accompanied by diminished microglial activation in septic mice. In addition, QX1 decreased the expression of M1 phenotype microglia gene markers such as CD32, Socs3, and CD68 and upregulated M2 phenotype marker genes, including Myc, Arg-1, and CD206 [36]. Fisetin is an effective ingredient of the TCM Cotinus coggygria. Its neuroprotective effects may be associated with the inhibition of neuroinflammation, represented by decreased expression of interleukin 1 receptor, pNF-κB, TNF-α, and inducible nitric oxide synthase in microglia. Additionally, fisetin-blocked NLRP3 inflammasome activation via the promotion of mitophagy may suppress the secretion of IL-1β into the CNS and contribute to the amelioration of cognitive impairment [74].
The above pharmacological intervention confers significant neuroprotection by inhibiting the inflammatory response in microglia and protecting against SAE, which may provide novel directions for reducing morbidity and ameliorating the neurological outcomes of SAE.
6. Conclusion
The brain is an important target organ during sepsis, microglial activation is a potentially harmful mechanism for brain damage. Microglia perform several critical functions in the brain, such as synaptogenesis, modification, plasticity changes, detection of local steady state, and immune functions, neuronal apoptosis regulation, myelin sheath regulation, and neurotrophic support. Microglial activation plays a critical role in the pathological and physiological processes of SAE, modulating microglial polarization to an anti-inflammatory phenotype also may be a potential therapeutic strategy for managing SAE, to reduce morbidity and ameliorate neurological outcomes. Although intensive studies were made by many researchers, the function of the microglial activation is still unclear which deserves further explorations to exact mechanisms and functional aspects of microglial activation to reach a clear conclusion.
Author Contributions
Jiyun H, Haisong Z, Xinren W, Binbin M, Shucai X, and Lina Z contributed to conception and design of the study. Jiyun H and Lina Z organized the database. Haisong Z and Shucai X assessed the quality of the study. Jiyun H wrote the first draft of the manuscript. Xinren W, Binbin M and Lina Z wrote sections of the manuscript. All authors contributed to manuscript revision, read, and approved the submitted version. and all authors commented on previous versions ofthe manuscript. All authors read and approved the final manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (82172145), the National Natural Science Foundation of China (81873956), China Postdoctoral Science Foundation(2022M7135).
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
SAE, Sepsis-associated encephalopathy; BBB, blood-brain barrier; CNS, central nervous system; MHC, major histocompatibility complex; IL, interleukin; TNF-α, tumor necrosis factor α; TGF-β, transforming growth factor β; TLR4, Toll-like receptor 4; LPS, lipopolysaccharide; CLP, cecal ligation perforation; GSDMD, gasdermin D; mTOR, mammalian target of rapamycin; IFNγ, interferon gamma; NLRP3, nucleotide-binding oligomerization domain, Leucine rich Repeat and Pyrin domain containing; IBA1+, ionized calcium binding adaptor molecule 1; PWM, periventricular white matter; CXCR5, chemokine receptor 5; rhSTC1, recombinant human STC-1; IL-1Ra, IL-1 receptor antagonist; Nrf2, nuclear factor erythroid 2-related factor 2; CB2R, cannabinoid type 2 receptor; TCM, Traditional Chinese Medicine; Atr, Atractylon; QX1, Qiang Xin 1; BHB, body β-hydroxybutyrate; LQM01, Indole-3-guanylhydrazone hydrochloride.
References
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.S.; Kissoon, N.; Finfer, S.; et al. Global, regional, and national sepsis incidence and mortality, 1990-2017: analysis for the Global Burden of Disease Study. Lancet (London, England) 2020, 395, 200–211. [Google Scholar] [CrossRef]
- Liu, V.; Escobar, G.J.; Greene, J.D.; Soule, J.; Whippy, A.; Angus, D.C.; Iwashyna, T.J. Hospital deaths in patients with sepsis from 2 independent cohorts. Jama 2014, 312, 90–92. [Google Scholar] [CrossRef]
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). Jama 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Reinhart, K.; Daniels, R.; Kissoon, N.; Machado, F.R.; Schachter, R.D.; Finfer, S. Recognizing Sepsis as a Global Health Priority - A WHO Resolution. The New England journal of medicine 2017, 377, 414–417. [Google Scholar] [CrossRef] [PubMed]
- Golzari, S.E.; Mahmoodpoor, A. Sepsis-associated encephalopathy versus sepsis-induced encephalopathy. Lancet Neurol 2014, 13, 967–968. [Google Scholar] [CrossRef] [PubMed]
- Sonneville, R.; de Montmollin, E.; Poujade, J.; Garrouste-Orgeas, M.; Souweine, B.; Darmon, M.; Mariotte, E.; Argaud, L.; Barbier, F.; Goldgran-Toledano, D.; et al. Potentially modifiable factors contributing to sepsis-associated encephalopathy. Intensive Care Med 2017, 43, 1075–1084. [Google Scholar] [CrossRef]
- Gofton, T.E.; Young, G.B. Sepsis-associated encephalopathy. Nature reviews. Neurology 2012, 8, 557–566. [Google Scholar] [CrossRef]
- Bozza, F.A.; D'Avila, J.C.; Ritter, C.; Sonneville, R.; Sharshar, T.; Dal-Pizzol, F. Bioenergetics, mitochondrial dysfunction, and oxidative stress in the pathophysiology of septic encephalopathy. Shock 2013, 39 Suppl 1, 10–16. [Google Scholar] [CrossRef]
- Bustamante, A.C.; Opron, K.; Ehlenbach, W.J.; Larson, E.B.; Crane, P.K.; Keene, C.D.; Standiford, T.J.; Singer, B.H. Transcriptomic Profiles of Sepsis in the Human Brain. Am J Respir Crit Care Med 2020, 201, 861–863. [Google Scholar] [CrossRef]
- Michels, M.; Abatti, M.R.; Ávila, P.; Vieira, A.; Borges, H.; Carvalho Junior, C.; Wendhausen, D.; Gasparotto, J.; Tiefensee Ribeiro, C.; Moreira, J.C.F.; et al. Characterization and modulation of microglial phenotypes in an animal model of severe sepsis. J Cell Mol Med 2020, 24, 88–97. [Google Scholar] [CrossRef]
- Lemstra, A.W.; Groen in't Woud, J.C.; Hoozemans, J.J.; van Haastert, E.S.; Rozemuller, A.J.; Eikelenboom, P.; van Gool, W.A. Microglia activation in sepsis: a case-control study. J Neuroinflammation 2007, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Zrzavy, T.; Höftberger, R.; Berger, T.; Rauschka, H.; Butovsky, O.; Weiner, H.; Lassmann, H. Pro-inflammatory activation of microglia in the brain of patients with sepsis. Neuropathology and applied neurobiology 2019, 45, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Colonna, M.; Butovsky, O. Microglia Function in the Central Nervous System During Health and Neurodegeneration. Annual review of immunology 2017, 35, 441–468. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, U.K. Microglia as a source and target of cytokines. Glia 2002, 40, 140–155. [Google Scholar] [CrossRef]
- Salter, M.W.; Stevens, B. Microglia emerge as central players in brain disease. Nat Med 2017, 23, 1018–1027. [Google Scholar] [CrossRef]
- Azam, S.; Haque, M.E.; Kim, I.S.; Choi, D.K. Microglial Turnover in Ageing-Related Neurodegeneration: Therapeutic Avenue to Intervene in Disease Progression. Cells 2021, 10. [Google Scholar] [CrossRef]
- Paolicelli, R.C.; Bolasco, G.; Pagani, F.; Maggi, L.; Scianni, M.; Panzanelli, P.; Giustetto, M.; Ferreira, T.A.; Guiducci, E.; Dumas, L.; et al. Synaptic pruning by microglia is necessary for normal brain development. Science (New York, N.Y.) 2011, 333, 1456–1458. [Google Scholar] [CrossRef]
- Lawson, L.J.; Perry, V.H.; Dri, P.; Gordon, S. Heterogeneity in the distribution and morphology of microglia in the normal adult mouse brain. Neuroscience 1990, 39, 151–170. [Google Scholar] [CrossRef]
- Fu, R.; Shen, Q.; Xu, P.; Luo, J.J.; Tang, Y. Phagocytosis of microglia in the central nervous system diseases. Mol Neurobiol 2014, 49, 1422–1434. [Google Scholar] [CrossRef]
- Barakat, R.; Redzic, Z. The Role of Activated Microglia and Resident Macrophages in the Neurovascular Unit during Cerebral Ischemia: Is the Jury Still Out? Medical principles and practice : international journal of the Kuwait University, Health Science Centre 2016, 25(Suppl 1), 3–14. [Google Scholar] [CrossRef]
- Galea, I.; Bechmann, I.; Perry, V.H. What is immune privilege (not)? Trends in immunology 2007, 28, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Roth, T.L.; Nayak, D.; Atanasijevic, T.; Koretsky, A.P.; Latour, L.L.; McGavern, D.B. Transcranial amelioration of inflammation and cell death after brain injury. Nature 2014, 505, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, U.K.; Kettenmann, H. Microglia: active sensor and versatile effector cells in the normal and pathologic brain. Nature neuroscience 2007, 10, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, A.; Wake, H.; Ishikawa, A.W.; Eto, K.; Shibata, K.; Murakoshi, H.; Koizumi, S.; Moorhouse, A.J.; Yoshimura, Y.; Nabekura, J. Microglia contact induces synapse formation in developing somatosensory cortex. Nature communications 2016, 7, 12540. [Google Scholar] [CrossRef]
- Michels, M.; Sonai, B.; Dal-Pizzol, F. Polarization of microglia and its role in bacterial sepsis. Journal of neuroimmunology 2017, 303, 90–98. [Google Scholar] [CrossRef]
- Ransohoff, R.M.; Perry, V.H. Microglial physiology: unique stimuli, specialized responses. Annual review of immunology 2009, 27, 119–145. [Google Scholar] [CrossRef]
- Masuda, T.; Amann, L.; Monaco, G.; Sankowski, R.; Staszewski, O.; Krueger, M.; Del Gaudio, F.; He, L.; Paterson, N.; Nent, E.; et al. Specification of CNS macrophage subsets occurs postnatally in defined niches. Nature 2022, 604, 740–748. [Google Scholar] [CrossRef]
- Hammond, T.R.; Dufort, C.; Dissing-Olesen, L.; Giera, S.; Young, A.; Wysoker, A.; Walker, A.J.; Gergits, F.; Segel, M.; Nemesh, J.; et al. Single-Cell RNA Sequencing of Microglia throughout the Mouse Lifespan and in the Injured Brain Reveals Complex Cell-State Changes. Immunity 2019, 50, 253–271.e256. [Google Scholar] [CrossRef]
- Madry, C.; Kyrargyri, V.; Arancibia-Cárcamo, I.L.; Jolivet, R.; Kohsaka, S.; Bryan, R.M.; Attwell, D. Microglial Ramification, Surveillance, and Interleukin-1β Release Are Regulated by the Two-Pore Domain K(+) Channel THIK-1. Neuron 2018, 97, 299–312.e296. [Google Scholar] [CrossRef]
- Nimmerjahn, A.; Kirchhoff, F.; Helmchen, F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science (New York, N.Y.) 2005, 308, 1314–1318. [Google Scholar] [CrossRef]
- Paolicelli, R.C.; Sierra, A.; Stevens, B.; Tremblay, M.E.; Aguzzi, A.; Ajami, B.; Amit, I.; Audinat, E.; Bechmann, I.; Bennett, M.; et al. Microglia states and nomenclature: A field at its crossroads. Neuron 2022, 110, 3458–3483. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M. A polarizing question: do M1 and M2 microglia exist? Nature neuroscience 2016, 19, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Masuda, T.; Sankowski, R.; Staszewski, O.; Böttcher, C.; Amann, L.; Sagar; Scheiwe, C. ; Nessler, S.; Kunz, P.; van Loo, G.; et al. Spatial and temporal heterogeneity of mouse and human microglia at single-cell resolution. Nature 2019, 566, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Borst, K.; Dumas, A.A.; Prinz, M. Microglia: Immune and non-immune functions. Immunity 2021, 54, 2194–2208. [Google Scholar] [CrossRef]
- Butovsky, O.; Jedrychowski, M.P.; Moore, C.S.; Cialic, R.; Lanser, A.J.; Gabriely, G.; Koeglsperger, T.; Dake, B.; Wu, P.M.; Doykan, C.E.; et al. Identification of a unique TGF-β-dependent molecular and functional signature in microglia. Nature neuroscience 2014, 17, 131–143. [Google Scholar] [CrossRef]
- Yang, L.; Li, Z.; Xu, Z.; Zhang, B.; Liu, A.; He, Q.; Zheng, F.; Zhan, J. Protective Effects of Cannabinoid Type 2 Receptor Activation Against Microglia Overactivation and Neuronal Pyroptosis in Sepsis-Associated Encephalopathy. Neuroscience 2022, 493, 99–108. [Google Scholar] [CrossRef]
- Wang, X.; Xu, X.; Guo, Y.; Huang, P.; Ha, Y.; Zhang, R.; Bai, Y.; Cui, X.; He, S.; Liu, Q. Qiang Xin 1 Formula Suppresses Excessive Pro-Inflammatory Cytokine Responses and Microglia Activation to Prevent Cognitive Impairment and Emotional Dysfunctions in Experimental Sepsis. Front Pharmacol 2020, 11, 579. [Google Scholar] [CrossRef]
- Gosselin, D.; Skola, D.; Coufal, N.G.; Holtman, I.R.; Schlachetzki, J.C.M.; Sajti, E.; Jaeger, B.N.; O'Connor, C.; Fitzpatrick, C.; Pasillas, M.P.; et al. An environment-dependent transcriptional network specifies human microglia identity. Science (New York, N.Y.) 2017, 356. [Google Scholar] [CrossRef]
- Abud, E.M.; Ramirez, R.N.; Martinez, E.S.; Healy, L.M.; Nguyen, C.H.H.; Newman, S.A.; Yeromin, A.V.; Scarfone, V.M.; Marsh, S.E.; Fimbres, C.; et al. iPSC-Derived Human Microglia-like Cells to Study Neurological Diseases. Neuron 2017, 94, 278–293.e279. [Google Scholar] [CrossRef]
- Mueller, K.L.; Hines, P.J.; Travis, J. Neuroimmunology. Science (New York, N.Y.) 2016, 353, 760–761. [Google Scholar] [CrossRef]
- Prinz, M.; Masuda, T.; Wheeler, M.A.; Quintana, F.J. Microglia and Central Nervous System-Associated Macrophages-From Origin to Disease Modulation. Annual review of immunology 2021, 39, 251–277. [Google Scholar] [CrossRef] [PubMed]
- Shulyatnikova, T.; Tumanskyi, V.; Hayden, M.R. Reactive Microgliosis in Sepsis-Associated and Acute Hepatic Encephalopathies: An Ultrastructural Study. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.Y.; Wickel, J.; Hahn, N.; Mein, N.; Schwarzbrunn, M.; Koch, P.; Ceanga, M.; Haselmann, H.; Baade-Büttner, C.; von Stackelberg, N.; et al. Microglia mediate neurocognitive deficits by eliminating C1q-tagged synapses in sepsis-associated encephalopathy. Science advances 2023, 9, eabq7806. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Xu, H.; Zhang, H.; Tang, R.; Lan, Y.; Xing, R.; Li, S.; Christian, E.; Hou, Y.; Lorello, P.; et al. Disruption of the IL-33-ST2-AKT signaling axis impairs neurodevelopment by inhibiting microglial metabolic adaptation and phagocytic function. Immunity 2022, 55, 159–173.e159. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.U.; Minhas, P.S.; Liddelow, S.A.; Haileselassie, B.; Andreasson, K.I.; Dorn, G.W., 2nd; Mochly-Rosen, D. Fragmented mitochondria released from microglia trigger A1 astrocytic response and propagate inflammatory neurodegeneration. Nature neuroscience 2019, 22, 1635–1648. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Yang, Y.; Peng, Z.; Xie, L.; Zhong, X.; Liang, F.; Yuan, C.; Lu, B. Silencing IFNγ inhibits A1 astrocytes and attenuates neurogenesis decline and cognitive impairment in endotoxemia. Biochem Biophys Res Commun 2020, 533, 1519–1526. [Google Scholar] [CrossRef]
- Xiao, T.; Ji, H.; Shangguan, X.; Qu, S.; Cui, Y.; Xu, J. NLRP3 inflammasome of microglia promotes A1 astrocyte transformation, neo-neuron decline and cognition impairment in endotoxemia. Biochem Biophys Res Commun 2022, 602, 1–7. [Google Scholar] [CrossRef]
- Shi, J.; Xu, H.; Cavagnaro, M.J.; Li, X.; Fang, J. Blocking HMGB1/RAGE Signaling by Berberine Alleviates A1 Astrocyte and Attenuates Sepsis-Associated Encephalopathy. Front Pharmacol 2021, 12, 760186. [Google Scholar] [CrossRef]
- Jiang, S.; Wang, H.; Zhou, Q.; Li, Q.; Liu, N.; Li, Z.; Chen, C.; Deng, Y. Melatonin ameliorates axonal hypomyelination of periventricular white matter by transforming A1 to A2 astrocyte via JAK2/STAT3 pathway in septic neonatal rats. Journal of Inflammation Research 2021, 14, 5919–5937. [Google Scholar] [CrossRef]
- Calsavara, A.J.C.; Nobre, V.; Barichello, T.; Teixeira, A.L. Post-sepsis cognitive impairment and associated risk factors: A systematic review. Australian critical care : official journal of the Confederation of Australian Critical Care Nurses 2018, 31, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Moraes, C.A.; Santos, G.; de Sampaio e Spohr, T.C.; D'Avila, J.C.; Lima, F.R.; Benjamim, C.F.; Bozza, F.A.; Gomes, F.C. Activated Microglia-Induced Deficits in Excitatory Synapses Through IL-1β: Implications for Cognitive Impairment in Sepsis. Mol Neurobiol 2015, 52, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Iwashyna, T.J.; Ely, E.W.; Smith, D.M.; Langa, K.M. Long-term cognitive impairment and functional disability among survivors of severe sepsis. Jama 2010, 304, 1787–1794. [Google Scholar] [CrossRef] [PubMed]
- Munster, B.C.; Aronica, E.; Zwinderman, A.H.; Eikelenboom, P.; Cunningham, C.; Rooij, S.E. Neuroinflammation in delirium: a postmortem case-control study. Rejuvenation research 2011, 14, 615–622. [Google Scholar] [CrossRef]
- Andonegui, G.; Zelinski, E.L.; Schubert, C.L.; Knight, D.; Craig, L.A.; Winston, B.W.; Spanswick, S.C.; Petri, B.; Jenne, C.N.; Sutherland, J.C.; et al. Targeting inflammatory monocytes in sepsis-associated encephalopathy and long-term cognitive impairment. JCI Insight 2018, 3. [Google Scholar] [CrossRef]
- Jing, G.; Zuo, J.; Fang, Q.; Yuan, M.; Xia, Y.; Jin, Q.; Liu, Y.; Wang, Y.; Zhang, Z.; Liu, W.; et al. Erbin protects against sepsis-associated encephalopathy by attenuating microglia pyroptosis via IRE1α/Xbp1s-Ca(2+) axis. J Neuroinflammation 2022, 19, 237. [Google Scholar] [CrossRef]
- Deng, Y.Y.; Fang, M.; Zhu, G.F.; Zhou, Y.; Zeng, H.K. Role of microglia in the pathogenesis of sepsis-associated encephalopathy. CNS Neurol Disord Drug Targets 2013, 12, 720–725. [Google Scholar] [CrossRef]
- Hoshino, K.; Hayakawa, M.; Morimoto, Y. Minocycline Prevents the Impairment of Hippocampal Long-Term Potentiation in the Septic Mouse. Shock 2017, 48, 209–214. [Google Scholar] [CrossRef]
- Wang, X.; Song, Y.; Chen, J.; Zhang, S.; Le, Y.; Xie, Z.; Ouyang, W.; Tong, J. Subcutaneous administration of β-hydroxybutyrate improves learning and memory of sepsis surviving mice. Neurotherapeutics : the journal of the American Society for Experimental NeuroTherapeutics 2020, 17, 616–626. [Google Scholar] [CrossRef]
- Iwashyna, T.J.; Cooke, C.R.; Wunsch, H.; Kahn, J.M. Population burden of long-term survivorship after severe sepsis in older Americans. Journal of the American Geriatrics Society 2012, 60, 1070–1077. [Google Scholar] [CrossRef]
- Armstrong, B.A.; Betzold, R.D.; May, A.K. Sepsis and Septic Shock Strategies. The Surgical clinics of North America 2017, 97, 1339–1379. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Yang, K.; Xiao, Q.; Hou, R.; Pan, X.; Zhu, X. Central role of microglia in sepsis-associated encephalopathy: From mechanism to therapy. Front Immunol 2022, 13, 929316. [Google Scholar] [CrossRef] [PubMed]
- Terrando, N.; Rei Fidalgo, A.; Vizcaychipi, M.; Cibelli, M.; Ma, D.; Monaco, C.; Feldmann, M.; Maze, M. The impact of IL-1 modulation on the development of lipopolysaccharide-induced cognitive dysfunction. Crit Care 2010, 14, R88. [Google Scholar] [CrossRef]
- Li, Y.; Wang, F.; Luo, Y. Ginsenoside Rg1 protects against sepsis-associated encephalopathy through beclin 1-independent autophagy in mice. J Surg Res 2017, 207, 181–189. [Google Scholar] [CrossRef]
- Tian, M.; Qingzhen, L.; Zhiyang, Y.; Chunlong, C.; Jiao, D.; Zhang, L.; Li, W. Attractylone attenuates sepsis-associated encephalopathy and cognitive dysfunction by inhibiting microglial activation and neuroinflammation. J Cell Biochem 2019, 120, 7101–7108. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.E.; Liu, L.; Wang, Y.C.; Wang, C.T.; Zheng, Q.; Liu, Q.X.; Li, Z.F.; Bai, X.J.; Liu, X.H. Caspase-1 inhibitor exerts brain-protective effects against sepsis-associated encephalopathy and cognitive impairments in a mouse model of sepsis. Brain Behav Immun 2019, 80, 859–870. [Google Scholar] [CrossRef]
- Heimfarth, L.; Carvalho, A.M.S.; Quintans, J.S.S.; Pereira, E.W.M.; Lima, N.T.; Bezerra Carvalho, M.T.; Barreto, R.S.S.; Moreira, J.C.F.; da Silva-Júnior, E.F.; Schmitt, M.; et al. Indole-3-guanylhydrazone hydrochloride mitigates long-term cognitive impairment in a neonatal sepsis model with involvement of MAPK and NFκB pathways. Neurochem Int 2020, 134, 104647. [Google Scholar] [CrossRef]
- Xie, K.; Zhang, Y.; Wang, Y.; Meng, X.; Wang, Y.; Yu, Y.; Chen, H. Hydrogen attenuates sepsis-associated encephalopathy by NRF2 mediated NLRP3 pathway inactivation. Inflamm Res 2020, 69, 697–710. [Google Scholar] [CrossRef]
- Rocha, M.; Vieira, A.; Michels, M.; Borges, H.; Goulart, A.; Fernandes, F.; Dominguini, D.; Ritter, C.; Dal-Pizzol, F. Effects of S100B neutralization on the long-term cognitive impairment and neuroinflammatory response in an animal model of sepsis. Neurochemistry International 2021, 142. [Google Scholar] [CrossRef]
- Bonfante, S.; Joaquim, L.; Fileti, M.E.; Giustina, A.D.; de Souza Goldim, M.P.; Danielski, L.G.; Cittadin, E.; De Carli, R.J.; de Farias, B.X.; Engel, N.A.; et al. Stanniocalcin 1 Inhibits the Inflammatory Response in Microglia and Protects Against Sepsis-Associated Encephalopathy. Neurotox Res 2021, 39, 119–132. [Google Scholar] [CrossRef]
- Wen, Q.; Ding, Q.; Wang, J.; Yin, Y.; Xu, S.; Ju, Y.; Ji, H.; Liu, B. Cortistatin-14 Exerts Neuroprotective Effect Against Microglial Activation, Blood-brain Barrier Disruption, and Cognitive Impairment in Sepsis-associated Encephalopathy. Journal of immunology research 2022, 2022, 3334145. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Chen, Z.; Wang, Y.; Mao, M.; Deng, Y.; Shi, M.; Xu, Y.; Chen, L.; Cao, W. JQ1 attenuates neuroinflammation by inhibiting the inflammasome-dependent canonical pyroptosis pathway in SAE. Brain Res Bull 2022, 189, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Liang, H.; Song, H.; Ding, X.; Wang, D.; Zhang, X.; Sun, T. Metformin Improves the Prognosis of Adult Mice with Sepsis-Associated Encephalopathy Better than That of Aged Mice. Journal of immunology research 2022, 2022, 3218452. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Ren, X.; Ai, Y.; Liu, Z. SS-31 Improves Cognitive Function in Sepsis-Associated Encephalopathy by Inhibiting the Drp1-NLRP3 Inflammasome Activation. Neuromolecular medicine 2022, 10.1007/s12017-022-08730-1. [Google Scholar] [CrossRef]
- Ding, H.; Li, Y.; Chen, S.; Wen, Y.; Zhang, S.; Luo, E.; Li, X.; Zhong, W.; Zeng, H. Fisetin ameliorates cognitive impairment by activating mitophagy and suppressing neuroinflammation in rats with sepsis-associated encephalopathy. CNS Neurosci Ther 2022, 28, 247–258. [Google Scholar] [CrossRef]
- Scott, M.C.; Haase, C.M.; Olson, S.D.; Cox, C.S., Jr. Dexmedetomidine Alters the Inflammatory Profile of Rat Microglia In Vitro. Neurocrit Care 2023, 38, 688–697. [Google Scholar] [CrossRef]
- Walsh, J.G.; Muruve, D.A.; Power, C. Inflammasomes in the CNS. Nature reviews. Neuroscience 2014, 15, 84–97. [Google Scholar] [CrossRef]
- Voet, S.; Srinivasan, S.; Lamkanfi, M.; van Loo, G. Inflammasomes in neuroinflammatory and neurodegenerative diseases. EMBO molecular medicine 2019, 11. [Google Scholar] [CrossRef]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: in vivo veritas. The Journal of clinical investigation 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Rahimifard, M.; Maqbool, F.; Moeini-Nodeh, S.; Niaz, K.; Abdollahi, M.; Braidy, N.; Nabavi, S.M.; Nabavi, S.F. Targeting the TLR4 signaling pathway by polyphenols: A novel therapeutic strategy for neuroinflammation. Ageing Res Rev 2017, 36, 11–19. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, H.; Tang, X.; Bai, W.; Wang, G.; Tian, X. Repetitive transcranial magnetic stimulation regulates L-type Ca(2+) channel activity inhibited by early sevoflurane exposure. Brain Res 2016, 1646, 207–218. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Q.; Lou, Y.; Xu, J.; Feng, Z.; Chen, Y.; Tang, Q.; Zheng, G.; Zhang, Z.; Wu, Y.; et al. Salidroside attenuates neuroinflammation and improves functional recovery after spinal cord injury through microglia polarization regulation. J Cell Mol Med 2018, 22, 1148–1166. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Yu, Y.; Jiang, Y.; Zhao, S.; Wang, Y.; Su, L.; Xie, K.; Yu, Y.; Lu, Y.; Lv, G. Molecular hydrogen attenuates sepsis-induced neuroinflammation through regulation of microglia polarization through an mTOR-autophagy-dependent pathway. Int Immunopharmacol 2020, 81, 106287. [Google Scholar] [CrossRef] [PubMed]
- Shemer, A.; Scheyltjens, I.; Frumer, G.R.; Kim, J.S.; Grozovski, J.; Ayanaw, S.; Dassa, B.; Van Hove, H.; Chappell-Maor, L.; Boura-Halfon, S.; et al. Interleukin-10 Prevents Pathological Microglia Hyperactivation following Peripheral Endotoxin Challenge. Immunity 2020, 53, 1033–1049.e1037. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, Y.; Du, J.; Jiang, B.; Shan, T.; Li, H.; Bao, H.; Si, Y. CXCR5 down-regulation alleviates cognitive dysfunction in a mouse model of sepsis-associated encephalopathy: potential role of microglial autophagy and the p38MAPK/NF-κB/STAT3 signaling pathway. J Neuroinflammation 2021, 18, 246. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.; Deng, S.; Zhang, L.; Huang, Y.; Li, W.; Peng, Q.; Liu, Z.; Ai, Y. SS-31 reduces inflammation and oxidative stress through the inhibition of Fis1 expression in lipopolysaccharide-stimulated microglia. Biochem Biophys Res Commun 2019, 520, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Haileselassie, B.; Joshi, A.U.; Minhas, P.S.; Mukherjee, R.; Andreasson, K.I.; Mochly-Rosen, D. Mitochondrial dysfunction mediated through dynamin-related protein 1 (Drp1) propagates impairment in blood brain barrier in septic encephalopathy. J Neuroinflammation 2020, 17, 36. [Google Scholar] [CrossRef]
- Galley, H.F. Oxidative stress and mitochondrial dysfunction in sepsis. Br J Anaesth 2011, 107, 57–64. [Google Scholar] [CrossRef]
- Deng, S.; Zhang, L.; Mo, Y.; Huang, Y.; Li, W.; Peng, Q.; Huang, L.; Ai, Y. Mdivi-1 attenuates lipopolysaccharide-induced acute lung injury by inhibiting MAPKs, oxidative stress and apoptosis. Pulm Pharmacol Ther 2020, 62, 101918. [Google Scholar] [CrossRef]
- Zhao, P.; Li, X.; Yang, Q.; Lu, Y.; Wang, G.; Yang, H.; Dong, J.; Zhang, H. Malvidin alleviates mitochondrial dysfunction and ROS accumulation through activating AMPK-α/UCP2 axis, thereby resisting inflammation and apoptosis in SAE mice. Front Pharmacol 2022, 13, 1038802. [Google Scholar] [CrossRef]
- Gao, X.; Liu, Y.; An, Z.; Ni, J. Active Components and Pharmacological Effects of Cornus officinalis: Literature Review. Front Pharmacol 2021, 12, 633447. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Uncontrolled or overactivated microglia are detrimental for the pathological conditions. Resting microglia can be polarized to the pro-inflammatory phenotype M1 (amoeboid-shaped cell) or alternatively to the anti-inflammatory phenotype M2 by different stimulators. The original figure was created using figdraw (www.figdraw.com).
Figure 1.
Uncontrolled or overactivated microglia are detrimental for the pathological conditions. Resting microglia can be polarized to the pro-inflammatory phenotype M1 (amoeboid-shaped cell) or alternatively to the anti-inflammatory phenotype M2 by different stimulators. The original figure was created using figdraw (www.figdraw.com).
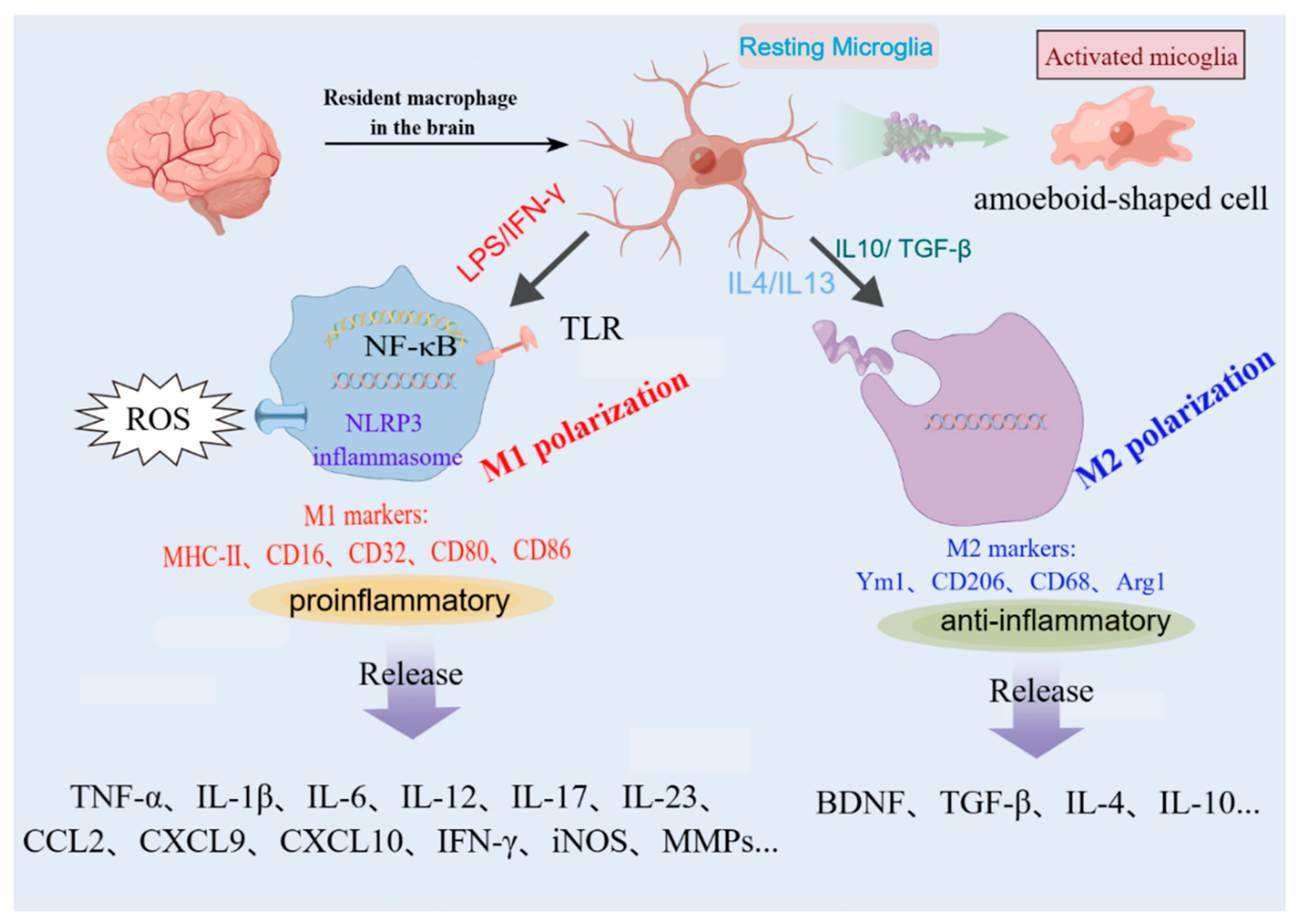
Figure 2.
Schematic representation of functions and mechanisms of microglia-mediated neurotoxicity in SAE. Activated microglia mediate neural interactions and cell death in the CNS, and play a crucial role in cognitive impairment of SAE. The original figure was created using figdraw (www.figdraw.com).
Figure 2.
Schematic representation of functions and mechanisms of microglia-mediated neurotoxicity in SAE. Activated microglia mediate neural interactions and cell death in the CNS, and play a crucial role in cognitive impairment of SAE. The original figure was created using figdraw (www.figdraw.com).
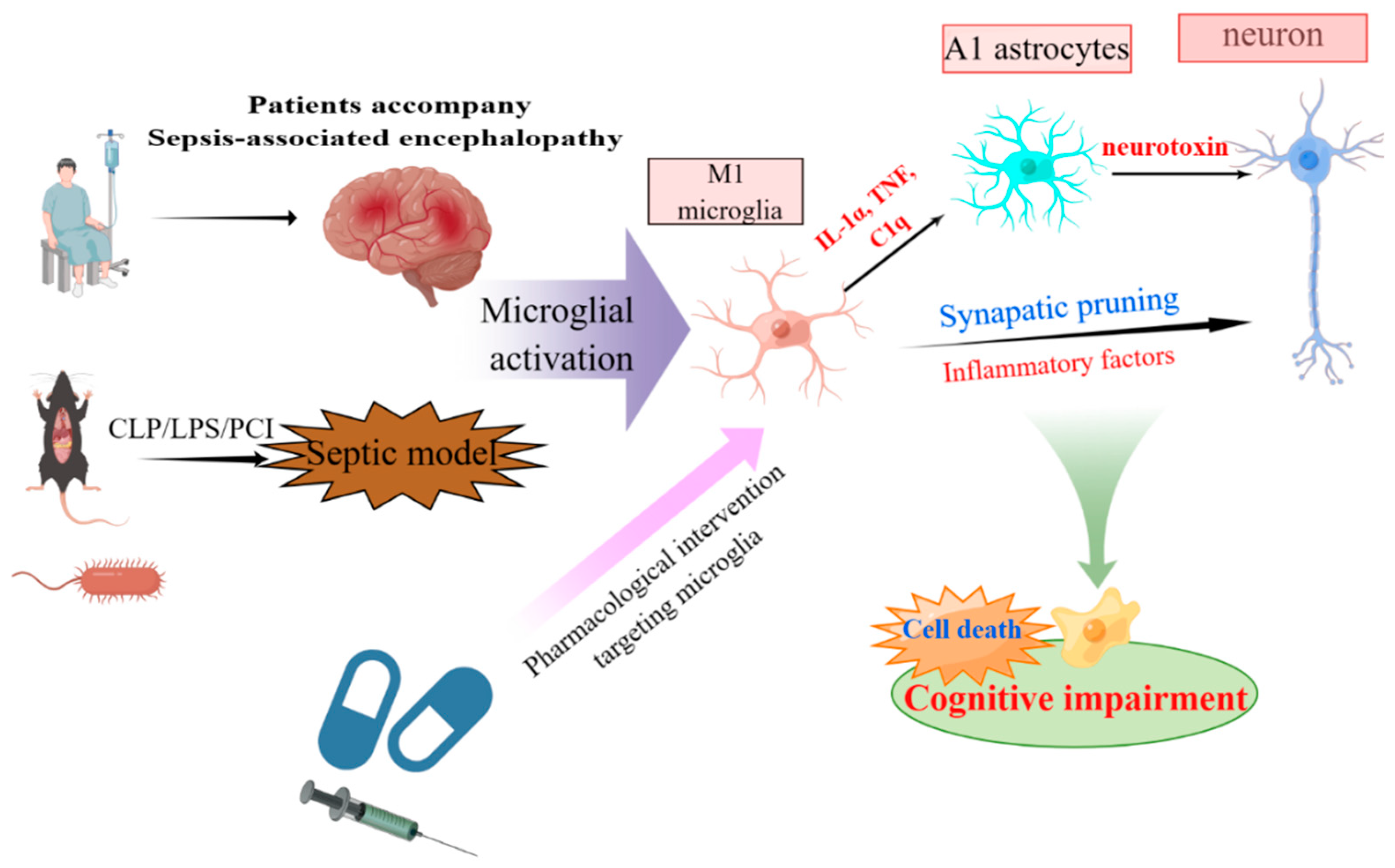

Table 1.
Treatments in experimental models of SAE targeting microglia and neuroinflammation.
| References | Species, strain, sex |
Model | Treatment and drug dose | Mode of administration and duration | Simplified treatment outcomes |
|---|---|---|---|---|---|
| Terrando 2010[62] | Mouse, WT C57BL/6 and IL-1R-/-, ♂ | LPS | IL-1 receptor antagonist (IL-1Ra), 100 mg/kg | Subcutaneous, immediately before LPS administration | Reduced plasma cytokines and hippocampal microgliosis and ameliorated cognitive dysfunction |
| Li 2017[63] | Mouse, C57 BL/6, ♂ | CLP | Ginsenoside Rg1, 40 and 200 mg/kg | I.p., 1 h before the CLP operation | Improved the survival rate; suppressed IBA1 activation and learning and memory impairments |
| Hoshino 2017[57] | Mouse, NA | CLP | Minocycline, 60 mg/kg | I.p., 3 consecutive days | Prevented impaired long-term potentiation in the hippocampus |
| Tian 2019 [64] | Mouse, C57 BL/6, ♂ | LPS | Attractylon, 25 mg/kg | I.p., with LPS injection | Attenuated LPS-induced cognitive impairment, neural apoptosis, inflammatory factors, and microglial activation |
| Xu 2019[65] | Mouse, BALB/c, ♂ | CLP | Caspase-1 inhibitor VX765, 0.2 mg per mouse | Intragastric administration, twice daily (10 a.m. and 4 p.m.) until mice were sacrificed | Reversed cognitive dysfunction and depressive-like behaviors; reduced microglia activation and BBB disruption and ultrastructure damages in the brain |
| Michels 2019[9] | Rat, Wistar, ♂ | CLP | Minocycline, 100 ug/kg | I.c.v, immediately after CLP operation | Induced a down-regulation of M1 markers |
| Wang 2020[58] | Mouse, C57 BL/6, ♂ | CLP | β-hydroxybutyrate, 250 mg/kg | Subcutaneous administration/i.c.v., every 6 hours from the fourth day to the seventh day after CLP/twice daily for 7 days | Increased survival and body weight recovery of sepsis mice and improved learning and memory; limited neuroinflammation and neuroplasticity damage |
| Heimfarth 2020[66] | Mouse, albino Swiss, ♂/♀ | LPS | Indole-3-guanylhydrazone hydrochloride, 50 mg/kg | I.p., after LPS administration and for 5 consecutive days | Attenuated inflammatory reactions through the MAPK and NFκB signaling pathways, and microglia activation suppression reduces anxiety-like behavior and cognitive impairment |
| Xie 2020[67] | Mouse, WT and Nrf2 KO, ♂ | CLP | MCC950/ Hydrogen-rich saline solution, 50 mg/kg/ 5 mL/kg | I.p., before operation/1 h and 6 h after CLP | Alleviated inflammation, neuronal apoptosis, and mitochondrial dysfunction via inhibiting Nrf2-mediated NLRP3 pathway. |
| Rocha 2021[68] | Rat, Wistar, ♂ | CLP | Anti-S100B monoclonal antibody, 10 μg/kg | I.c.v, 15 days after CLP | Increased the time of grooming; alleviated microglia activation |
| Bonfante 2021[69] | Rat, Wistar, ♂ | CLP | Stanniocalcin-1; 20/50/100 ng/kg | I.c.v, immediately after the CLP procedure | Improved hippocampal mitochondrial function and creatine kinase activity, reduced oxidative stress, neuroinflammation, and long-term memory impairment. |
| Wang 2022[36] | Mouse, C57 BL/6, ♂ | CLP | Qiang Xin 1, 0.5 /1/2 g/kg | Oral, 2 h after CLP | Attenuated cognitive deficits, emotional dysfunction, and reduces neuroinflammatory responses to improve survival. |
| Wen 2022[70] | Mouse, C57 BL/6 J, ♂ | CLP | Cortistatin-14, 200 ug/kg | I.p., 30 min after CLP | Relieved anxiety-related behaviors and the levels of various inflammatory cytokines; reduced BBB disruption and microglial activation |
| Zhong 2022[71] | Mouse, C57 BL/6, ♂ | LPS | JQ-1, 50 mg/kg | I.p., 1 h before LPS | Protected the hippocampal BBB and neuronal damage and microglia activation through the attenuation of neuroinflammation |
| Song 2022[72] | Mouse, C57 BL/6, ♂ | LPS | Metformin, 25 mg/kg | I.p., 1 h after LPS | Blocked microglial proliferation and production of inflammatory factors |
| Zhong 2022[73] | Mouse, C57 BL/J, ♂ | CLP | SS-31, 5 mg/kg | I.p., once daily for 1 week | Improved the survival rate and cognitive and memory dysfunctions in CLP mice |
| Yang 2022[35] | Mouse, C57 BL/6, ♂ | CLP | CB2R agonist HU308, 2.5 mg/kg | I.p., three consecutive days after CLP | Inhibited microglia activity and neuronal pyroptosis |
| Ding 2022[74] | Rat, NA, ♂ | CLP | Fisetin, 20 mg/kg | Intragastrical administration, once a day for three consecutive days before CLP | Blocked NLRP3 inflammasome activation by promoting mitophagy and ameliorating cognitive impairment |
CLP: Cecal ligation and puncture; LPS: lipopolysaccharide; BBB: blood-brain barrier; NA: not announced; WT: Wild type; KO: knockout; kg: kilogram; g: gram; h: hour; d: day; i.c.v.: intracerebroventricular injection; i.p.: intraperitoneal injection; male: ♂; female: ♀.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



