
Submitted:
11 August 2023
Posted:
14 August 2023
You are already at the latest version
Abstract
Kluyveromyces marxianus is an attractive thermotolerant yeast species for ethanol production because of its ability to utilize various carbon sources as a fermentation substrate. The use of thermotolerant microbes enables performance of high-temperature ethanol fermentation, which has several advantages including reduction of cooling costs and minimization of contamination risk. To improve K. marxianus for ethanol fermentation under stress conditions, two strains, DMKU 3-1042 and DMKU 3-118, were adapted for heat resistance and toxic substances in pulp wastewater from a paper mill, respectively, resulting in the generation of KMR1042 and KMR118, respectively. Both adapted mutants exhibited clumpy clusters of cells as pseudohyphae and altered colony morphology, and their sedimentation speeds were much faster than those of the corresponding parent strains. The two mutants showed stronger tolerance to various stresses and higher performance for ethanol production than those of the corresponding parent strains at high temperatures or in the presence of toxic substances. Genome sequencing analysis revealed that both mutants had disruption of the same gene, SWI5, despite adaptation under different stress conditions, suggesting that the formation of pseudohyphae is a common strategy of K. marxianus for coping with stresses.
Keywords:
Kluyveromyces marxianus DMKU3-1042
; Kluyveromyces marxianus DMKU3-118
; stress-resistant mutant
; pseudohyphae formation
; SWI5
1. Introduction
Fossil energy sources such as oil and coal are being increasingly depleted, resulting in increases in oil prices and impacts on energy security and economic development. The use of fossil fuels is causing global warming due to CO2 emissions. Therefore, renewable, sustainable, and low-cost energy alternatives to fossil fuels are needed [1]. Bioethanol is one of the clean and sustainable alternatives. However, the supply of starch-based biomass, which is relatively easy to convert to ethanol, is approaching its limit. There is a need for the use of cellulosic biomass, but the high cost of pretreatment and saccharifying enzymes is a problem. High-temperature fermentation (HTF) to produce bioethanol is attracting attention because it can reduce the costs of cooling, saccharifying enzymes, and equipment [2]. Thermotolerant fermenting microbes are essential for HTF, but stable fermentation at high temperatures requires the use of microbes that are resistant not only to temperature but also to inhibitors present in the fermentation broth and have high ethanol productivity.
Kluyveromyces marxianus, which is one of the most thermotolerant yeasts, has several useful characteristics for industrial ethanol fermentation including efficient production of ethanol at high temperatures, high growth rate, short doubling times, weak glucose repression, and capability for assimilation of sugars present in various raw materials including glucose, xylose and sucrose [3,4,5,6,7]. K. marxianus DMKU 3-1042 has been extensively studied for elucidating its thermotolerance and developing HTF [8,9,10,11].
During the fermentation process, yeast survival and ethanol production are affected by various types of stress including high ethanol concentrations, high temperatures, high sugar concentrations, bacterial acids, and low pH. Yeast cells, therefore, need to deal with and overcome such multiple stresses simultaneously [6]. Control of cell proliferation and cellular stress responses are closely related. Cell cycle arrest may be necessary to remove damage that occurs during cell cycle phases (S and M) when cells are specifically vulnerable and to allow cells to adapt while in G1 (or G2) phase. It is well known that G1 cells, non-proliferating cells, have higher stress tolerance than do actively dividing cells [12].
In this study, in order to obtain a robust yeast strain for stress tolerance, we adapted K. marxianus DMKU 3-1042 and DMKU 3-118 to different conditions, exposure to a high temperature and exposure to pulp wastewater, respectively. A high temperature tends to generate reactive oxygen species (ROS) via an increase in mitochondrial membrane fluidity [13], and ROS cause damage to DNA, lipids, and proteins [14]. Pulp wastewater contains toxic chemicals including acids, furfural, and hydroxymethyl furfural in addition to hexose and pentose [15]. Surprisingly, both adapted mutants shared a similar phenotype, formation of pseudohyphae, suggesting that the formation of pseudohyphae is a common strategy for resistance to stresses. Genome sequencing analysis revealed a key gene responsible for the different stress tolerances. The results of this study may lead identification of targets for genetic engineering of K. marxianus to improve various stress tolerances. The mutant strains obtained were found to be resistant to various stresses and have higher ethanol productivity than those of the corresponding parent strains under stress conditions. Therefore, the mutant strains are expected to be used for more stable HTF to produce ethanol.
2. Materials and Methods
2.1. Yeast strains and media
The yeast strains used in this study were K. marxianus strains DMKU 3-1042 and DMKU 3-118 [3] and their derivatives, KMR1042 and KMR118. Cells were grown in YPD medium [1% (w/v) yeast extract, 2% (w/v) peptone and 2% (w/v) glucose] at 30 ˚C for pre-culture and at different temperatures for culture under a shaking condition at 160 rpm.
2.2. Screening of mutants
To obtain mutants that are resistant to stresses, two strategic approaches were used for two K. marxianus strains, DMKU 3-1042 and DMKU-118. For K. marxianus DMKU 3-1042, cells were spread on about 100 YPD agar plates and incubated at 45 ˚C for 12 days. One white colony found on the plates was selected for further experiments. The color of most colonies usually changes from white to brown after incubation for a long time at that temperature. For K. marxianus DMKU 3-118, cells were adapted in pulp wastewater (Nippon Paper Industries CO., Ltd) by serial subculture 52 times at 37 ˚C.
2.3. Preparation of genomic DNA, genome sequencing, and determination of mutations
The genomic DNA of mutated strains was isolated as described previously [16] from cultured cells and further purified using a Genomic-tip 20 kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. Genome library construction for KMR118 and KMR1042 was performed using a Twist 96-plex library preparation kit (Twist Bioscience, San Francisco, USA) according to the manufacturer’s instructions. Genome sequencing of the two strains, KMR118 and KMR1042, and that of DMKU 3-118 were performed using Illumina NextSeq 1000 and Illumina HiSeq 2500 platform (Hokkaido System Science Co., Ltd., Hokkaido, Japan), respectively. Genome sequencing was carried out by a massively parallel sequencer (MiSeq; Illumina KK, Tokyo, Japan) as reported previously [17]. The sequenced reads were screened by a quality score higher than the Phred score of 30 and were trimmed 12 bases from the 50 end and 20 bases from the 30 end. Truncated reads less than 150 bases or with ambiguous nucleotides were excluded from further analysis.
For genome mapping analysis, the reference genome sequence of K. marxianus DMKU 3-1042 (GenBank acc. No: AP012213–AP012221) was downloaded from NCBI ftp site, ftp://ftp.ncbi.nlm.nih.gov/ (accessed on March 28, 2020). The Illumina sequence reads of these strains were mapped to the reference sequences using BWA with default parameters [18]. All single nucleotide polymorphisms (SNPs) and indels were detected using the Genome Analysis Toolkit (GATK) [19,20]. Critical mutations including mutations of KLMA_50490 in KMR118 and KMR1042 were confirmed by the Sanger method [21]. Physiological functions of mutated genes were analyzed by a BLAST search at NCBI (https://www.ncbi.nlm.nih.gov (accessed on March 28, 2020)) or with the STRING database (https://string-db.org (accessed on March 28, 2020)).
2.4. Cell morphology and sedimenting ability
For observation of cell morphology, cells were grown in 30 mL of YPD medium at 30 ˚C for 18 h under a shaking condition of 160 rpm, harvested by low-speed centrifugation, washed once with saline, suspended in saline, and subjected to observation under a microscope (E600, Nikon, Tokyo, Japan) at x400 magnification. For testing of sedimenting ability, cells were grown in 30 mL of YPD medium at 30 ˚C or 45 ˚C for 18 h under a shaking condition of 160 rpm, harvested by low-speed centrifugation, washed once with 0.1M EDTA, and suspended in 0.1M EDTA to adjust OD660 to 40 and 80 units for the mutant and wild-type strains, respectively. The ratio of OD660 of the mutant and wild-type strains was based on a standard curve of cell dry weight and OD660. The sedimenting test was started (0 min) after the cell suspension in a vial had been mixed by a vortex mixer. The sedimenting abilities were compared by taking photos at 5 min, 10 min, and 20 min. DMKU3-1042 was mainly used as a control because DMKU3-1042 and DMKU3-118 showed nearly the same sedimentation.
2.5. Analysis of stress tolerance
To prepare an inoculum, cells were grown at 30 ˚C and 160 rpm for 18 h in YPD medium, harvested, washed with distilled water, and resuspended in distilled water. The cell suspension was adjusted to OD660 of 1.0 and then ten-fold sequentially diluted and spotted onto YPD agar plates supplemented with (1) 0.1% (w/v) vanillin, (2) 10% (w/v) or 25% (w/v) glucose, (3) 6% (v/v) or 9% (v/v) ethanol, (4) 0.1% (v/v) or 0.2% (v/v) acetic acid, (5) 15 mM or 20 mM furfural, and (6) 4 mM, 8 mM or 16 mM hydrogen peroxide (H2O2). The plates were incubated at 30 ˚C for 48 h. In addition, the effect of temperature was examined on YPD agar plates. The plates were incubated at 30 ˚C, 48 ˚C, or 49 ˚C for 48 h. Each experiment was performed at least three times and similar results were obtained.
2.6. Analysis of ethanol fermentation ability
To investigate the ethanol fermentation ability of mutants and the corresponding parents, YP medium [1% (w/v) yeast extract and 2% (w/v) peptone] supplemented with 16% (w/v) of glucose was used. Cells were precultured in 30 mL of YPD medium in a 100-mL Erlenmeyer flask sealed with aluminum foil at 30 ˚C under a shaking condition at 160 rpm for 18 h. The preculture was inoculated to 30 mL of fresh YP medium supplemented with 16% (w/v) of glucose in a 100-mL Erlenmeyer flask at the initial OD660 of 0.1 and incubation was carried out at the appropriate temperature under a shaking condition at 100 rpm. Samples were taken at the times indicated until 36 h or 48 h of incubation. Yeast growth and concentrations of glucose and ethanol were determined.
Yeast growth was determined by measuring optical density at 660 nm on a UVmini-1240 spectrophotometer (Shimadzu, Japan). To determine glucose and ethanol concentrations in culture media, cultures were collected and centrifuged at 14,000 rpm for 5 min. The supernatant was passed through a 0.45-μm filter (Pall Corporation, Japan) and then subjected to quantitative analysis of glucose and ethanol on a high-performance liquid chromatography (HPLC) system (Hitachi, Japan) consisting of a Hitachi Model D-2000 Elite HPLC system Manager, column oven L-2130, pump L-2130, auto-sampler L-2200, and RI detector L-2490, equipped with a GL-C610-S gel pack column (Hitachi) in the mode of 0.5 mL/min eluent of deionized water at 60 ˚C.
2.7. Sequence data deposition
Illumina sequence reads of the KMR118 and KMR1042 strains and the DMKU 3-118 strain were deposited in the DDBJ Sequence Read Archive. The DRR accession numbers for KMR118, KMR1042, and DMKU 3-118 are DRR495714, DRR495716, and DRR495715, respectively.
3. Results
3.1. Stress-resistant mutants from K. marxianus strains DMKU 3-1042 and DMKU 3-118
To improve the performance of K. marxianus for ethanol fermentation under a multi-stress condition, two wild-type strains of K. marxianus, DMKU 3-1042 and DMKU 3-118, were exposed to different stresses: high temperature and drying on plates and toxic substances in pulp wastewater, respectively. Our previous observations revealed that the colony color of DMKU 3-1042 after incubation for several days at 45 ˚C on YPD agar plates changed from white to dark brown color, leading to cell death. We therefore spread K. marxianus DMKU 3-1042 cells on YPD agar plates (~100 colonies per plate) and incubated them at 45 ˚C for 12 days. After incubation, a white colony, named KMR1042, was obtained and was used for further experiments. On the other hand, the strain DMKU 3-118 was serially subcultured in pulp wastewater 52 times and an adapted strain, named KMR118, was obtained. Macroscopic observation revealed that the colony edges of both mutants on YPD plates were rough, and microscopic observation revealed that the two mutants formed pseudohyphae in YPD media (Figure 1).
3.2. Sedimenting ability of KMR1042 and KMR118
In the DMKU 3-1042 and DMKU 3-118 background, KMR1042 and KMR118 exhibited a defect in cell separation, resulting in clumpy clusters of cells and altered colony morphology. To quantify the cell separation defects, we tested the cell sedimenting ability of both mutants. Two different concentrations of cells that were grown at 30 ˚C and 45 ˚C were tested. As a result, both cells were rapidly sedimented compared to the corresponding parent cells, and the sedimentation of both mutant cells was nearly the same and almost completed within 10 min (Figure 2).
3.3. Stress tolerance of KMR1042 and KMR118
During a fermentation process, yeast cells are generally exposed to various stresses including heat stress, osmotic stress, acid stress, and ethanol stress, all of which have severe effects on cell viability and ethanol production [22,23]. In addition, cells during HTF tend to generate oxidative stress [24]. The degree of stress tolerance of the two mutants, KMR1042 and KMR118, was thus compared with that of the corresponding parents. Cells were spotted onto YPD agar plates and incubated at 30 ˚C, 48 ˚C or 49 ˚C (heat stress) for 48 h or onto YP agar plates supplemented with 10% or 25% glucose (osmotic stress), 6% or 9% ethanol (ethanol stress), 0.1% or 0.2% acetic acid (acid stress), 15 mM or 20 mM furfural, or 4 mM, 8 mM or 16 mM H2O2 (oxidative stress) and incubated at 30 ˚C for 48 h (Figure 3). At 48 ˚C and 49 ˚C, both mutants had much better growth than that of their parents. For osmotic stress, the two mutants and their parents showed similar levels of growth in the presence of 10% glucose, but the growth of both mutants was better than that of their parents in the presence of 25% glucose. For ethanol stress, KMR1042 and its parent showed similar levels of growth in the presence of 6% and 9% ethanol. However, the mutant KMR118 showed a level of growth similar to that of its parent in the presence of 6% ethanol but grew much better than its parent in the presence of 9% ethanol. For oxidative stress, the two mutants grew in the presence of 8 mM H2O2, but only KMR1042 could grow in the presence of 16 mM H2O2. It is known that toxic substances, including acetic acid, furfural and vanillin, that are generated in the process of hydrolysis of lignocellulosic materials prevent the growth or fermentation ability of microbes [25,26]. Cells were thus spotted onto YPD agar plates supplemented with 0.1% vanillin or 0.1% and 0.2% acetic acid or 15 mM and 20 mM furfural, and effects of the toxic substances on cell growth were evaluated after incubation at 30 ˚C for 48 h (Figure 3). The two mutants showed better growth than that of their parents. These findings suggest that KMR1042 and KMR118 are more suitable for ethanol fermentation under conditions of various stresses.
3.4. Ethanol fermentation abilities of KMR1042 and KMR118 under stress conditions
To examine the fermentation abilities of KMR1042 and KMR118 at high temperatures, these cells were grown in a YP medium containing 16% glucose under a shaking condition at 45 ˚C, and cell growth and concentrations of glucose and ethanol in the medium were compared with those of the parents (Figure 4). The turbidity of all strains tested increased with decrease in glucose concentration, but the turbidity of the two mutants was much lower than that of their parents. The lag phase was found in the first 6 h in all strains. Ethanol was increased roughly in an anti-parallel manner with a decrease in glucose. Ethanol levels became nearly maximal at 24 h of incubation except for that in the case of DMKU 3-118, which was around 30 h. Both mutants showed higher ethanol concentrations with higher ethanol production rates than those of the parents. KMR1042 produced slightly less acetate than that produced by its parent, while KMR118 produced more acetate than that produced by its parent.
The fermentation abilities of KMR1042 and KMR118 were also examined in the presence of toxic substances, similar to previous study with other strains [27]. Cells were grown in a YP medium containing 2% or 10% glucose, 0.05% vanillin, 0.1% acetic acid, and 10 mM furfural under a shaking condition at 40 ˚C (Figure 5). In the case of 2% glucose with toxic substances, the growth of KMR1042 and KMR118 were similar to that of their parent counterparts, though their turbidity levels were lower than those of their parents. Ethanol accumulation of both mutants started earlier than that of their parents. On the other hand, in the case of 10% glucose with toxic substances, both mutants showed much faster growth and ethanol accumulation than those of their parents. These findings and those described above suggest that KMR1042 and KMR118 are robust strains against various stresses and may have a potential for ethanol fermentation at high temperatures or in the presence of toxic substances for industrial applications.
3.5. Analysis of mutation points of KMR1042 and KMR118
Genome sequencing was performed to identify the mutations of the two mutants (Table 1). As a result, KMR1042 and KMR118 were shown to have four and nine mutation points in protein-coding regions, respectively. Interestingly, both mutants were found to have mutations in the same gene, KLMA_50490, for metallothionein expression activator as an orthologue of SWI5 in S. cerevisiae. The KLMA_50490 mutations in KMR1042 and KMR118 caused a frame-shift mutation and a nonsense mutation, respectively, by the insertion of ten nucleotides and nucleotide substitution, respectively. Because there are no other overlapping gene mutations and disruption of the SWI5 homologue causes formation of pseudohyphae in S. cerevisiae (see Discussion section), the mutations of KLMA_50490 shared by the two mutants may be responsible for exhibiting a similar colony morphology, pseudohyphae formation, and tolerance phenotype against multi-stress. The shared disruption mutations may also contribute to the significant increase in ethanol fermentation at high temperatures. Given that the two mutants had nearly identical phenotypes, it is possible that additional mutations in KMR1042 and KMR118 contributed little to the phenotypes.
4. Discussion
In this study, we performed breeding of two K. marxianus strains under two different stress conditions to establish a much more stable HTF. The isolated mutants were resistant to multi-stress and achieved high ethanol production at high temperatures. Evolutionary adaptation has contributed to improved substrate utilization efficiency [28,29,30,31], functional implementation of alternative pathways [32,33], and increased tolerance to inhibitors [30,34] in various organisms. Our experimental results suggest that the mutants can be used to perform HTF even in the presence of toxic materials.
Notably, the two mutants acquired in this study have disrupted mutations in the same gene, KLMA_50490 as an orthologue of SWI5 in S. cerevisiae, which may be responsible for similar phenotypes described above. In S. cerevisiae, Ace2p, which shares 37% identity with Swi5p regulates the expression of several genes during the late M and early G1 stages of the mitotic cell cycle [35,36]. Targets of Ace2p are predominantly expressed in daughter cells and include CTS1 for chitinase and genes for proteins involved in separation from the mother cells [35,37,38]. Deletion of ACE2 results in increased pseudohyphal growth and invasion of agar [39,40]. On the other hand, SWI5 in S. cerevisiae encodes a transcription factor that activates the transcription of genes expressed at the M/G1 boundary and in the G1 phase of the cell cycle [41-43]. The DNA-binding domain of Swi5p is very similar to that of Ace2p [41], and both bind the same DNA sequences in vitro with similar affinities to regulate a shared set of genes in vivo [42,43]. However, the two regulators have distinct domains responsible for promoter-specific transcription activation and distinct promoter specificities in some genes; for example, transcription of the HO endonuclease gene is activated by Swi5p but not by Ace2p and CTS1 is activated by Ace2p and not by Swi5p.
Ace2p and Swi5p are conserved in species closely related to S. cerevisiae, including the opportunistic pathogenic yeast Candida glabrata [44]. The Candida albicans genome, however, contains only a single gene orthologue (C. albicans ACE2), the product of which is equidistant from both of the S. cerevisiae proteins [45]. Deletion of ACE2 in C. albicans results in gross attenuation of virulence of the fungus in a mouse model of disseminated infection [45]. In contrast, deletion of ACE2 in C. glabrata results in hypervirulence of the mutant strain in neutropenic mice [46] and in alterations in the expression of more than 60 proteins [47]. Mutants of C. albicans, C. glabrata, and S. cerevisiae with disruptions of ACE2 and of C. glabrata and S. cerevisiae with disruptions of SWI5 showed a clumping phenotype [48].
However, there has been no study on these genes in K. marxianus. A search of the NCBI database revealed that there is only one gene, KLMA_50490, for a protein that has a high identity with S. cerevisiae Swi5p and Ace2p, 34% and 30% identity, respectively, in K. marxianus and thus the KLMA_50490-encoded protein is an orthologue of S. cerevisiae Swi5p. A single orthologue of S. cerevisiae Swi5p or Ace2p may be enough in some yeasts including Kluyveromyces lactis [49] and Candida spp. [50]. The SWI5 disruption was found in both mutants, which were adapted to different stress conditions, suggesting that the SWI5 mutation is one of the strategies of K. marxianus to cope with various stresses.
Altering transcriptional programs is a major strategy for microorganisms to adapt to their surrounding environment. In S. cerevisiae, the environmental stress response involves about 900 genes for which expression is coordinately altered upon exposure to different types of stress [51]. Transcription analysis revealed that environmental stresses including heat, hyperosmolarity, and oxidative and starvation stress induce a very similar pattern of regulated genes in S. cerevisiae, C. albicans, Schizosaccharomyces pombe, and C. glabrata, which live in different environments [52,53,54]. As far as we know, there has been no report of SWI5 disruption causing multi-stress resistance in K. marxianus. Cellular stress responses protect cells from the harmful effects of stress and promote the repair of damage that occurs [55]. As shown in this study, exposure to one type of stress has been shown to increase tolerance to other types of stress [56]. This cross-protective phenomenon allows us to speculate that different stresses induce common cellular responses, such as modulating energy metabolism and expressing common protective proteins (such as heat shock proteins) and small protective molecules (compatible solutes such as glycerol or trehalose). Moreover, different stress responses may share certain means, possibly related to cross-protective phenomena. This important part of the stress response is referred to as the yeast general or environmental stress response [57].
We have almost no idea about the downstream from the SWI5 disruption to cause multi-stress resistance in K. marxianus. However, if, as S. cerevisiae Swi5p, K. marxianus Swi5p activates transcription of genes expressed at the M/G1 boundary and in the G1 phase of the cell cycle, it is possible that dysfunction of Swi5p causes cell cycle arrest at G1, which in turn provides time for the repair of cellular damage caused by stresses. Further studies downstream of Swi5p will allow us to understand mechanisms of multi-stress resistance, and that knowledge will be useful for the use of resulting mutants for stable HTF.
5. Conclusions
Thermotolerant and multi-stress-tolerant yeast strains are needed for stable HTF as low-cost fermentation. In this study, we carried out two different procedures and acquired one robust mutant from each of the procedures. Surprisingly, both mutants exhibited similar phenotypes including formation of pseudohyphae and multi-stress resistance, and they had disruption of the same gene, encoding an orthologue of S. cerevisiae Swi5p. It is therefore assumed that disruption of the gene is an effective mutation for a robust phenotype. Both mutants showed higher levels of ethanol production at high temperatures and under multi-stress conditions than those of their parents, indicating that they are candidates for stable HTF.
Author Contributions
N.L. and M.Y. organized the study and wrote the original manuscript. A.O. and K.H. performed adaptation to isolate mutants. S.P., S.N., A.O., and K.H. performed experiments and analyzed data. M.M. performed genome analysis. T.K. and S.L. gave advice and support for this project. M.Y. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Special Coordination Funds for Promoting Science and Technology, Ministry of Education, Culture, Sports, Science and Technology (MEXT), and the Advanced Low Carbon Technology Research and Development Program, Japan Science and Technology Agency (JST). This work was partially performed as collaborative research in the Asian Core Program and in the Core to Core Program, which was supported by the Scientific Cooperation Program agreed by the Japan Society for the Promotion of Science (JSPS), the National Research Council of Thailand (NRCT), the Ministry of Science and Technology in Vietnam, the National Univ. of Laos, the Univ. of Brawijaya, and Beuth Univ. of Applied Science Berlin. The research visit of N.L. to Japan was supported by Research Center for Thermotolerant Microbial Resources, Yamaguchi University.
Data Availability Statement
All data are reported in this manuscript.
Acknowledgments
We thank Kazunobu Matsushita, Toshiharu Yakushi, and Naoya Kataoka for their helpful discussion and the NODAI Genome Research Center, Tokyo University of Agriculture for the sequencing and its analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bai, F.W.; Anderson, W.A.; Moo-Young, M. Ethanol fermentation technologies from sugar and starch feedstocks. Biotechnol. Adv. 2008, 26, 89–105. [Google Scholar] [CrossRef] [PubMed]
- Lertwattanasakul, N.; Rodrussamee, N.; Kumakiri, I.; Pattanakittivorakul, S.; Yamada, M. (2023). Potential of thermo-tolerant microorganisms for production of cellulosic bioethanol. In the Handbook of Biorefinery Research and Technology, Eds.; Bisaria, V.; Springer Dordrecht, 2023; pp. 1–30.
- Limtong, S.; Sringiew, C.; Yongmanitchai, W. Production of fuel ethanol at high temperature from sugar cane juice by a newly isolated Kluyveromyces marxianus. Bioresour. Technol. 2007, 98, 3367–3374. [Google Scholar] [CrossRef] [PubMed]
- Goshima, T.; Tsuji, M.; Inoue, H.; Yano, S.; Hoshino, T.; Matsushika, A. Bioethanol production from lignocellulosic biomass by a novel Kluyveromyces marxianus strain. Biosci. Biotechnol. Biochem. 2013, 77, 1505–1510. [Google Scholar] [CrossRef] [PubMed]
- Nitiyon, S.; Keo-Oudone, C.; Murata, M.; Lertwattanasakul, N.; Limtong, S.; Kosaka, T.; Yamada, M. Efficient conversion of xylose to ethanol by stress-tolerant Kluyveromyces marxianus BUNL-21. Springerplus 2016, 27, 185. [Google Scholar] [CrossRef] [PubMed]
- Saini, P.; Beniwal, A.; Kokkiligadda, A.; Vij, S. Evolutionary adaptation of Kluyveromyces marxianus strain for efficient conversion of whey lactose to bioethanol. Process. Biochem. 2017, 62, 69–79. [Google Scholar] [CrossRef]
- Kosaka, T.; Nakajima, Y.; Ishii, A.; Yamashita, M.; Yoshida, S.; Murata, M.; Kato, K.; Shiromaru, Y.; Kato, S.; Kanasaki, Y.; Yoshikawa, H.; Matsutani, M.; Thanonkeo, P.; Yamada, M. Capacity for survival in global warming: adaptation of mesophiles to the temperature upper limit. PloS ONE 2019, 14, e0215614. [Google Scholar]
- Rodrussamee, N.; Lertwattanasakul, N.; Hirata, K.; Suprayogi; Limtong, S. ; Kosaka, T.; Yamada, M. Growth and ethanol fermentation ability on hexose and pentose sugars and glucose effect under various conditions in thermotolerant yeast Kluyveromyces marxianus. Appl. Microbiol. Biotechnol. 2011, 90, 1573–1586. [Google Scholar] [CrossRef]
- Lertwattanasakul, N.; Rodrussamee, N.; Suprayogi; Limtong, S. ; Thanonkeo, P.; Kosaka, T.; Yamada, M. Utilization capability of sucrose, raffinose and inulin and its less-sensitiveness to glucose repression in thermotolerant yeast Kluyveromyces marxianus DMKU 3-1042. AMB Expr. 2011, 1, 20. [Google Scholar] [CrossRef]
- Lertwattanasakul, N.; Suprayogi; Murata, M. ; Rodrussamee, N.; Limtong, S.; Kosaka, T.; Yamada, M. Essentiality of respiratory activity for pentose utilization in thermotolerant yeast Kluyveromyces marxianus DMKU 3-1042. Antonie van Leeuwenhoek 2013, 103, 933–945. [Google Scholar] [CrossRef]
- Nurcholis, M.; Murata, M.; Limtong, S.; Kosaka, T.; Yamada, M. MIG1 as a positive regulator for the histidine biosynthesis pathway and as a global regulator in thermotolerant yeast Kluyveromyces marxianus. Sci. Rep. 2019, 9, 9926. [Google Scholar] [CrossRef]
- Jiménez, J.; Bru, S.; Ribeiro, M.PC.; Clotet, J. Live fast, die soon: cell cycle progression and lifespan in yeast cells. Microb. Cell 2015, 2, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Tarrío, N.; García-Leiro, A.; Cerdán, M.E.; González-Siso, M.I. The role of glutathione reductase in the interplay between oxidative stress response and turnover of cytosolic NADPH in Kluyveromyces lactis. FEMS Yeast Res. 2008, 8, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Auesukaree, C. Molecular mechanisms of the yeast adaptive response and tolerance to stresses encountered during ethanol fermentation. J. Biosci. Bioeng. 2017, 124, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Jönsson, L.J. ; Alriksson, B.; Nilvebrant, N-O. Bioconversion of lignocellulose: inhibitors and detoxification. Biotechnol. Biofuels 2013, 6, 16. [CrossRef] [PubMed]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2008; Volume 1. [Google Scholar]
- Akuzawa, S.; Nagaoka, J.; Kanekatsu, M.; Kanesaki, Y.; Suzuki, T. Draft genome sequence of Oceanobacillus picturae Heshi-B3, isolated from fermented rice bran in a traditional Japanese seafood dish. Genome Announc. 2016, 4, e01621–15. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics. 2010, 26, 589–595. [Google Scholar] [CrossRef]
- DePristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A. , del Angel, G.; Rivas, M.A.; Hanna, M.; McKenna, A.; Fennell, T.J.; Kernytsky, A.M.; Sivachenko, A.Y.; Cibulskis, K.; Gabriel, S.B.; Altshuler, D.; Daly, M.J. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef]
- Matsutani, M.; Matsumoto, N.; Hirakawa, H.; Shiwa, Y.; Yoshikawa, H.; Okamoto-Kainuma, A.; Ishikawa, M.; Kataoka, N.; Yakushi, T.; Matsushita, K. Comparative genomic analysis of closely related Acetobacter pasteurianus strains provides evidence of horizontal gene transfer and reveals factors necessary for thermotolerance. J Bacteriol. 2020, 202, e00553–19. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef]
- Gibson, B.R.; Lawrence, S.J.; Leclaire, J.P.; Powell, C.D.; Smart, K.A. Yeast responses to stresses associated with industrial brewery handling. FEMS Microbiol. Rev. 2007, 31, 535–569. [Google Scholar] [CrossRef]
- Puligundla, P.; Smogrovicova, D.; Obulam, V.S.R. , Ko, S. Very high gravity (VHG) ethanolic brewing and fermentation: a research update. J. Ind. Microbiol. Biotechnol. 2011, 38, 1133–1144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Shi, J.; Jiang, L. Modulation of mitochondrial membrane integrity and ROS formation by high temperature in Saccharomyces cerevisiae. Electron. J. Biotechnol. 2015, 18, 202–209. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Roberto, I.C. Alternatives for detoxification of diluted-acid lignocellulosic hydrolyzates for use in fermentative processes: a review. Bioresour. Technol. 2004, 93, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Behera, S.; Arora, R.; Nandhagopal, N.; Kumar, S. Importance of chemical pretreatment for bioconversion of lignocellulosic biomass. Renew. Sustain. Energy Rev. 2014, 36, 91–106. [Google Scholar] [CrossRef]
- Pattanakittivorakul, S.; Tsuzuno, T.; Kosaka, T.; Murata, M.; Kanesaki, Y.; Yoshikawa, H.; Limtong, S.; Yamada, M. Evolutionary adaptation by repetitive long-term cultivation with gradual increase in temperature for acquiring multi-stress tolerance and high ethanol productivity in Kluyveromyces marxianus DMKU 3-1042. Microorganisms 2022, 10, 798. [Google Scholar] [CrossRef]
- Wisselink, H.W.; Toirkens, M.J.; Wu, Q.; Pronk, J.T.; van Maris, A.J.A. Novel evolutionary engineering approach for accelerated utilization of glucose, xylose, and arabinose mixtures by engineered Saccharomyces cerevisiae strains. Appl. Environ. Microbiol. 2009, 75, 907–914. [Google Scholar] [CrossRef]
- Kuyper, M.; Toirkens, M.J.; Diderich, J.A.; Winkler, A.A.; van Dijken, J.P.; Pronk, J.T. Evolutionary engineering of mixed-sugar utilization by a xylose-fermenting Saccharomyces cerevisiae strain. FEMS Yeast Res. 2005, 5, 925–934. [Google Scholar] [CrossRef]
- Guadalupe-Medina, V.; Almering, M.J.H.; van Maris, A.J.A.; Pronk, J.T. Elimination of glycerol production in anaerobic cultures of a Saccharomyces cerevisiae strain engineered to use acetic acid as an electron acceptor. Appl. Environ. Microbiol. 2010, 76, 190–195. [Google Scholar] [CrossRef]
- Koppram, R.; Albers, E.; Olsson, L. Evolutionary engineering strategies to enhance tolerance of xylose utilizing recombinant yeast to inhibitors derived from spruce biomass. Biotechnol. Biofuels. 2012, 5, 32. [Google Scholar] [CrossRef]
- Zelle, R.M.; Harrison, J.C.; Pronk, J.T.; van Maris, A.J.A. Anaplerotic role for cytosolic malic enzyme in engineered Saccharomyces cerevisiae strains. Appl. Environ. Microbiol. 2011, 77, 732–738. [Google Scholar] [CrossRef]
- de Kok, S.; Nijkamp, J.F.; Oud, B.; Roque, F.C. , de Ridder, D.; Daran J-M., Pronk, J.T.; van Maris, A.J.A. Laboratory evolution of new lactate transporter genes in a jen1 delta mutant of Saccharomyces cerevisiae and their identification as ADY2 alleles by whole-genome resequencing and transcriptome analysis. FEMS Yeast Res. 2012, 12, 359–374. [Google Scholar] [PubMed]
- Adamo, G.M.; Brocca, S.; Passolunghi, S.; Salvato, B.; Lotti, M. Laboratory evolution of copper tolerant yeast strains. Microb. Cell. Fact. 2012, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Doolin, M.T.; Johnson, A.L.; Johnston, L.H.; Butler, G. Overlapping and distinct roles of the duplicated transcription factors Ace2p and Swi5p. Mol. Microbiol. 2001, 40, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Simon, I.; Barnett, J.; Hannett, N.; Harbison, C.T.; Rinaldi, N.J.; Volkert, T.L.; Wyrick, J.J.; Zeitlinger, J.; Gifford, D.K.; Jaakkola, T.S.; Young, R.A. Serial regulation of transcriptional regulators in the yeast cell cycle. Cell. 2001, 106, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Colman-Lerner, A.; Chin, T.E.; Brent, R. Yeast Cbk1 and Mob2 activate daughter-specific genetic programs to induce asymmetric cell fates. Cell. 2001, 107, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Weiss, E.L.; Kurischko, C.; Zhang, C.; Shokat, K.; Drubin, D.G.; Luca, F.C. The Saccharomyces cerevisiae Mob2p-Cbk1p kinase complex promotes polarized growth and acts with the mitotic exit network to facilitate daughter cell-specific localization of Ace2p transcription factor. J. Cell. Biol. 2002, 158, 885–900. [Google Scholar] [CrossRef]
- King, L.; Butler, G. Ace2p, a regulator of Cts1 (chitinase) expression, affects pseudohyphal production in Saccharomyces cerevisiae. Curr. Genet. 1998, 34, 183–191. [Google Scholar] [CrossRef]
- O’Conallain, C.; Doolin, M.T.; Taggart, C.; Thornton, F.; Butler, G. Regulated nuclear localization of the yeast transcription factor Ace2p controls expression of chitinase (CTS1) in Saccharomyces cerevisiae. Mol. Gen. Genet. 1999, 262, 275–282. [Google Scholar] [CrossRef]
- Dohrmann, P.R.; Butler, G.; Tamai, K.; Dorland, S.; Greene, J.R.; Thiele, D.J.; Stillman, D.J. Parallel pathways of gene regulation: homologous regulators SWI5 and ACE2 differentially control transcription of HO and chitinase. Genes. Dev. 1992, 6, 93–104. [Google Scholar] [CrossRef]
- Dohrmann, P.R.; Voth, W.P.; Stillman, D.J. Role of negative regulation in promoter specificity of the homologous transcriptional activators Ace2p and Swi5p. Mol. Cell. Biol. 1996, 16, 1746–1758. [Google Scholar] [CrossRef]
- McBride, H.J.; Yu, Y.; Stillman, D.J. Distinct regions of the Swi5 and Ace2 transcription factors are required for specific gene activation. J. Biol. Chem. 1999, 274, 21029–21036. [Google Scholar] [CrossRef] [PubMed]
- Dujon, B.; Sherman, D.; Fischer, G.; Durrens, P.; Casaregola, S.; Lafontaine, I.; De Montigny, J.; Marck, C.; Neuveglise, C.; Talla, E.; Goffard, N.; Frangeul, L.; Aigle, M.; Anthouard, V.; Babour, A.; Barbe, V.; Barnay, S.; Blanchin, S.; Beckerich, J.M.; Beyne, E.; Bleykasten, C.; Boisrame, A.; Boyer, J.; Cattolico, L.; Confanioleri, F.; De Daruvar, A.; Despons, L.; Fabre, E.; Fairhead, C.; Ferry-Dumazet, H.; Groppi, A.; Hantraye, F.; Hennequin, C.; Jauniaux, N.; Joyet, P.; Kachouri, R.; Kerrest, A.; Koszul, R.; Lemaire, M.; Lesur, I.; Ma, L. , Muller, H.; Nicaud, J.M., Nikolski, M., Oztas, S., Ozier-Kalogeropoulos, O.; Pellenz, S., Potier, S.; Richard, G.F.; Straub, M.L.; Suleau, A.; Swennen, D.; Tekaia, F.; Wesolowski-Louvel, M.; Westhof, E.; Wirth, B.; Zeniou-Meyer, M.; Zivanovic, I.; Bolotin-Fukuhara, M.; Thierry, A.; Bouchier, C.; Caudron, B.; Scarpelli, C.; Gaillardin, C.; Weissenbach, J.; Wincker, P.; Souciet, J.L. Genome evolution in yeasts. Nature 2004, 430, 35–44. [Google Scholar] [PubMed]
- Kelly, M.T.; MacCallum, D.M. , Clancy, S.D.; Odds, F.C.; Brown, A.J.P.; Butler, G. The Candida albicans CaACE2 gene affects morphogenesis, adherence and virulence. Mol. Microbiol. 2004, 53, 969–983. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.; Calcagno, A.M.; Findon, H.; Bignell, E.; Jones, M.D.; Warn, P.; Hopkins, P.; Denning, D.W.; Butler,G. ; Rogers, T.; Muhlschlegel, F.A.; Haynes, K. Inactivation of transcription factor gene ACE2 in the fungal pathogen Candida glabrata results in hypervirulence. Eukaryot. Cell 2004, 3, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Stead, D.; Findon, H.; Yin, Z.K.; Walker, J.; Selway, L.; Cash, P.; Dujon, B.A.; Hennequin, C.; Brown, A.J.P.; Haynes, K. Proteomic changes associated with inactivation of the Candida glabrata ACE2 virulence-moderating gene. Proteomics 2005, 5, 1838–1848. [Google Scholar] [CrossRef] [PubMed]
- MacCallum, D.M.; Findon, H.; Kenny, C.C.; Butler, G.; Haynes, K.; Odds, F.C. Different consequences of ACE2 and SWI5 gene disruptions for virulence of pathogenic and nonpathogenic yeasts. Infect Immun. 2006, 74, 5244–5248. [Google Scholar] [CrossRef]
- Bussereau, F.; Casaregola, S.; Lafay, J.F.; Bolotin-Fukuhara, M. The Kluyveromyces lactis repertoire of transcriptional regulators. FEMS Yeast Research 2006, 6, 325–335. [Google Scholar] [CrossRef]
- Mulhern, S.M.; Logue, M.E.; Butler, G. Candida albicans transcription factor Ace2 regulates metabolism and Is Required for filamentation in hypoxic conditions. Eukaryot. Cell 2006, 5, 2001–2013. [Google Scholar] [CrossRef]
- Gasch, A.P.; Spellman, P.T.; Kao, C.M.; Carmel-Harel, O.; Eisen, M.B.; Storz, G.; Botstein, D.; Brown, P.O. Genomic expression programs in the response of yeast cells to environmental changes. Mol. Biol. Cell. 2000, 11, 4241–4257. [Google Scholar] [CrossRef]
- Chen, D.; Toone, W.M.; Mata, J.; Lyne, R.; Burns, G.; Kivinen, K.; Brazma, A.; Jones, N.; Bähler, J. Global transcriptional responses of fission yeast to environmental stress. Mol. Biol. Cell. 2003, 14, 214–229. [Google Scholar] [CrossRef]
- Enjalbert, B.; Nantel, A.; Whiteway, M. Stress-induced gene expression in Candida albicans: absence of a general stress response. Mol. Biol. Cell. 2003, 14, 1460–1467. [Google Scholar] [CrossRef] [PubMed]
- Gasch, A.P. Comparative genomics of the environmental stress response in ascomycete fungi. Yeast. 2007, 24, 961–976. [Google Scholar] [CrossRef] [PubMed]
- Eardley, J.; Timson, D.J. Yeast cellular stress: impacts on bioethanol production. Fermentation 2020, 6, 109. [Google Scholar] [CrossRef]
- Święciło, A. Cross-stress resistance in Saccharomyces cerevisiae yeast—new insight into an old phenomenon. Cell Stress Chaperones. 2016, 21, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Gasch, A.P. The environmental stress response: a common yeast response to diverse environmental stresses. In Yeast Stress Responses. Eds.; Hohmann, S., Mager, W.H., Springer-Verlag Berlin Heidelberg, New York. 2003; Volume 1, pp. 11–70.
Figure 1.
Morphological observation of adapted mutants and their parents. After growing in YPD medium at 30 ˚C for 18 h under a shaking condition of 160 rpm, the cells were subjected to morphological observation at x400 magnification. Cell preparation and other details are described in Materials and Methods.
Figure 1.
Morphological observation of adapted mutants and their parents. After growing in YPD medium at 30 ˚C for 18 h under a shaking condition of 160 rpm, the cells were subjected to morphological observation at x400 magnification. Cell preparation and other details are described in Materials and Methods.
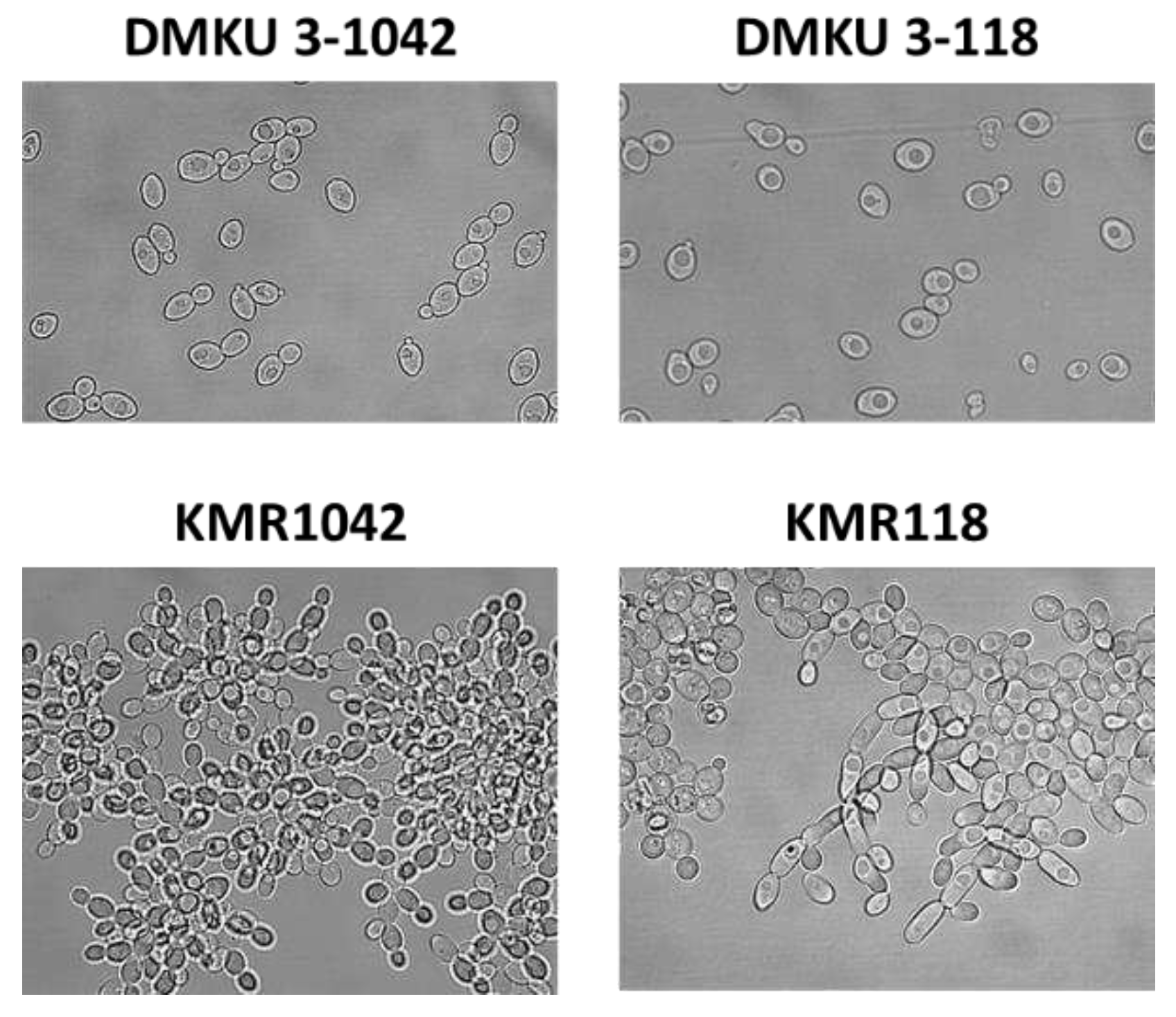
Figure 2.
Sedimenting ability of adapted mutants and their parents. After growing in YPD medium at 30 ˚C or 45 ˚C for 18 h under a shaking condition of 160 rpm, the cells were subjected to a sedimentation test. Cell preparation and other details are described in Materials and Methods. Two different cell suspensions for each strain (WT, wild type: OD660 of 20 and 80; mutants: OD660 of 10 and 40) were used. DMKU 3-1042 was used as a control because the sedimentation abilities of DMKU 3-1042 and DMKU 3-118 were almost the same. The sedimenting test was started (0 min) after the cell suspension in a vial had been mixed by a vortex mixer. Photos were taken at 5 min, 10 min, and 20 min.
Figure 2.
Sedimenting ability of adapted mutants and their parents. After growing in YPD medium at 30 ˚C or 45 ˚C for 18 h under a shaking condition of 160 rpm, the cells were subjected to a sedimentation test. Cell preparation and other details are described in Materials and Methods. Two different cell suspensions for each strain (WT, wild type: OD660 of 20 and 80; mutants: OD660 of 10 and 40) were used. DMKU 3-1042 was used as a control because the sedimentation abilities of DMKU 3-1042 and DMKU 3-118 were almost the same. The sedimenting test was started (0 min) after the cell suspension in a vial had been mixed by a vortex mixer. Photos were taken at 5 min, 10 min, and 20 min.

Figure 3.
Tolerance of adapted mutants and their parents to various stresses. After growing in YPD medium at 30 ˚C for 18 h under a shaking condition of 160 rpm, the cells were subjected to a stress tolerance test. Cell preparation and other details are described in Materials and Methods. Ten-fold sequentially diluted cells were spotted onto YPD agar plates supplemented with various substances as stress. The plates were incubated at 30 ˚C for 48 h. To see temperature tolerance, YPD agar plates without supplements were incubated at 30 ˚C or 45 ˚C for 48 h.
Figure 3.
Tolerance of adapted mutants and their parents to various stresses. After growing in YPD medium at 30 ˚C for 18 h under a shaking condition of 160 rpm, the cells were subjected to a stress tolerance test. Cell preparation and other details are described in Materials and Methods. Ten-fold sequentially diluted cells were spotted onto YPD agar plates supplemented with various substances as stress. The plates were incubated at 30 ˚C for 48 h. To see temperature tolerance, YPD agar plates without supplements were incubated at 30 ˚C or 45 ˚C for 48 h.

Figure 4.
Growth and ethanol fermentation of adapted mutants and their parents at 45 ˚C. Cells precultured in YPD medium at 30 ˚C for 18 h were inoculated into YP medium containing 16% glucose and cultured under a shaking condition of 160 rpm at 45 ˚C. Samples were taken at the times indicated, and glucose, ethanol and acetate concentrations were determined as described in Materials and Methods. Error bars represent ±SD of values from experiments performed in triplicate.
Figure 4.
Growth and ethanol fermentation of adapted mutants and their parents at 45 ˚C. Cells precultured in YPD medium at 30 ˚C for 18 h were inoculated into YP medium containing 16% glucose and cultured under a shaking condition of 160 rpm at 45 ˚C. Samples were taken at the times indicated, and glucose, ethanol and acetate concentrations were determined as described in Materials and Methods. Error bars represent ±SD of values from experiments performed in triplicate.
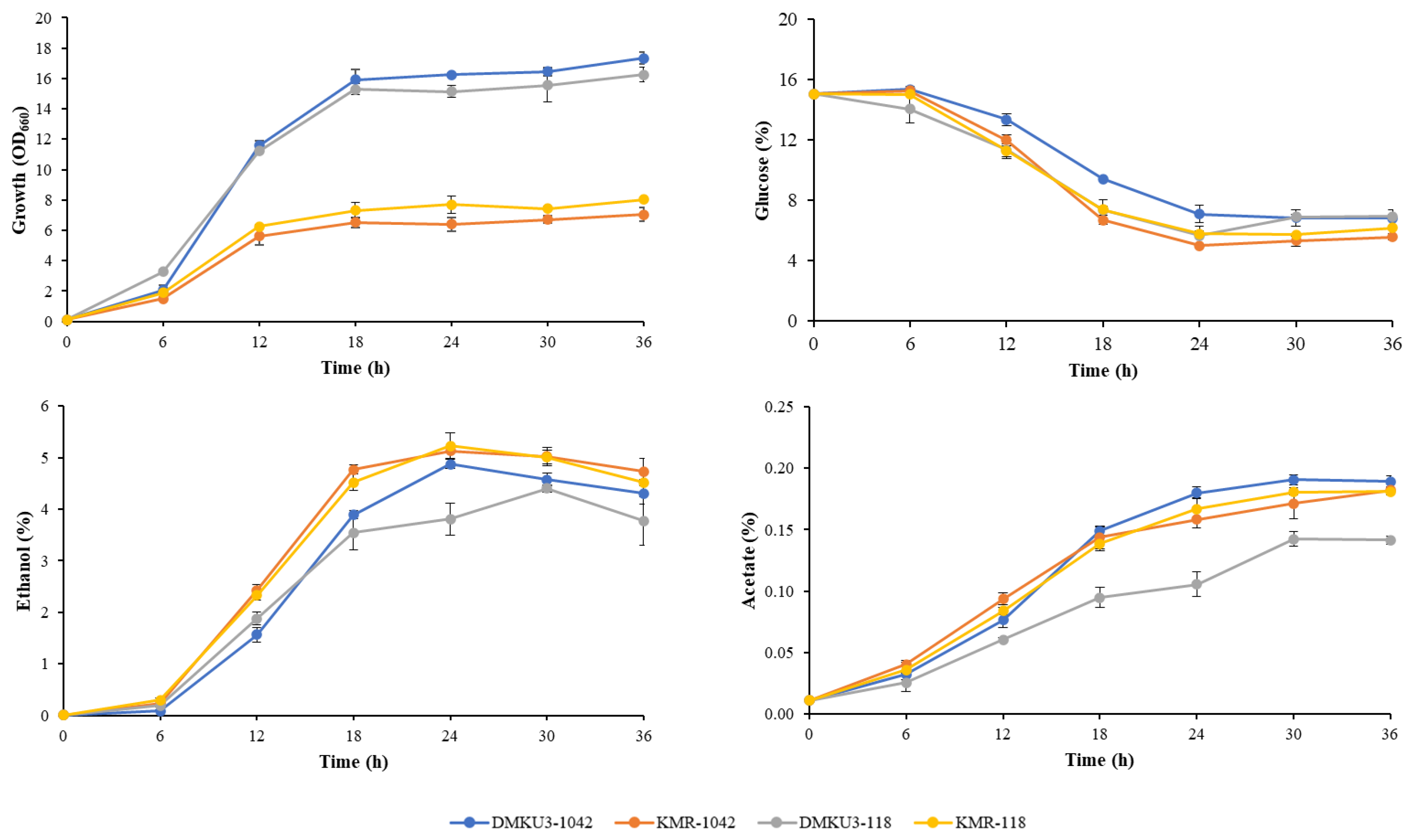
Figure 5.
Growth and ethanol fermentation of adapted mutants and their parents under multi-stress conditions. Cells precultured in YPD medium at 30 ˚C for 18 h were inoculated into YP medium containing 2% or 10% glucose supplemented with 0.05% (v/v) vanillin, 0.1% (v/v) acetic acid, and 10 mM furfural and cultured under a shaking condition of 160 rpm at 40 ˚C. Samples were taken at the times indicated, and glucose and ethanol concentrations were determined as described in Materials and Methods. Error bars represent ± SD of values from experiments performed in triplicate.
Figure 5.
Growth and ethanol fermentation of adapted mutants and their parents under multi-stress conditions. Cells precultured in YPD medium at 30 ˚C for 18 h were inoculated into YP medium containing 2% or 10% glucose supplemented with 0.05% (v/v) vanillin, 0.1% (v/v) acetic acid, and 10 mM furfural and cultured under a shaking condition of 160 rpm at 40 ˚C. Samples were taken at the times indicated, and glucose and ethanol concentrations were determined as described in Materials and Methods. Error bars represent ± SD of values from experiments performed in triplicate.
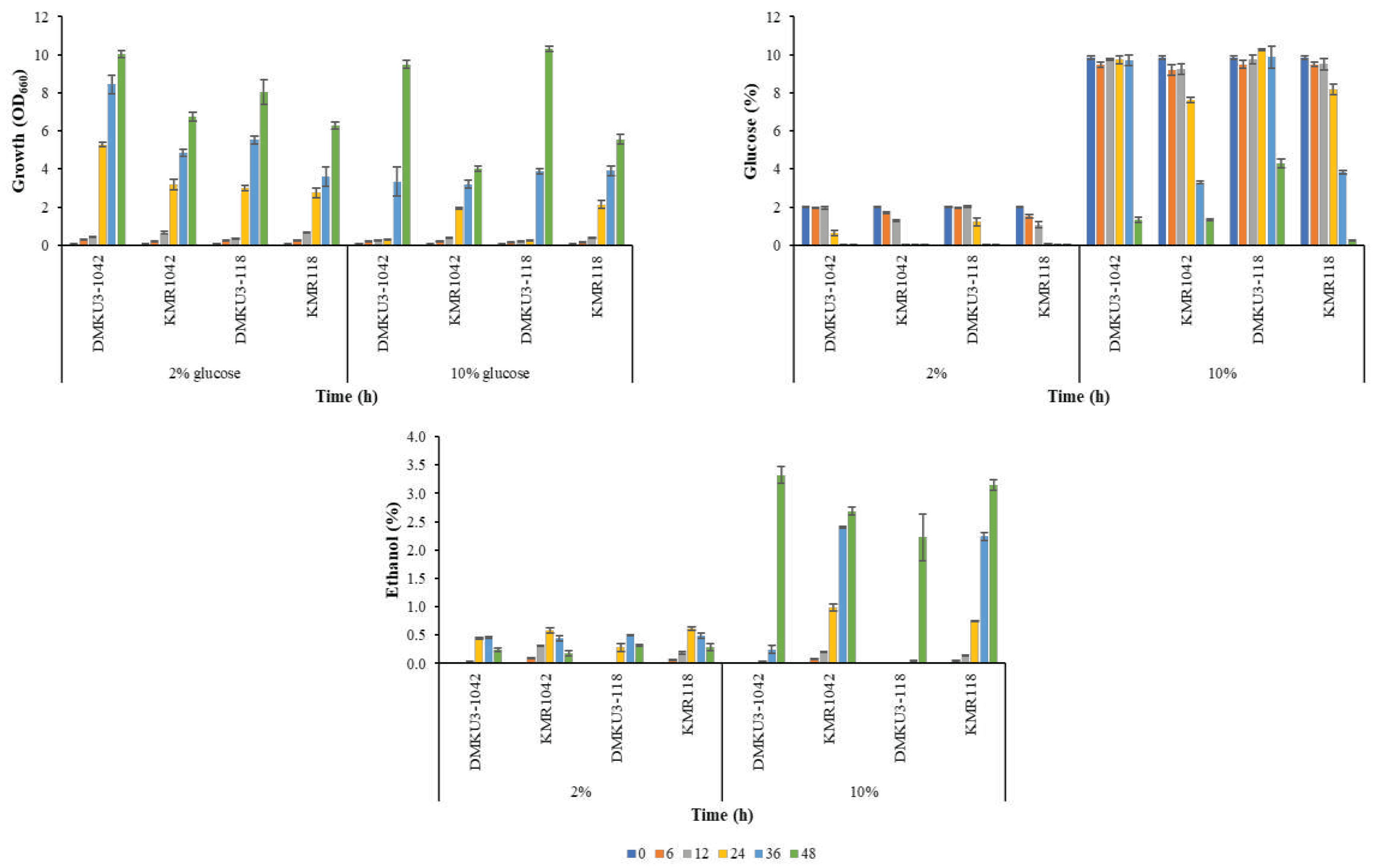

Table 1.
List of mutation sites in K. marxianus mutant strains, KMR1042 and KMR118.
| KMR1042 | |||||||
| locus_tag | gene | product | reference genome (NCBI ref seq acc. no.) | position | REF | ALT | AA sub |
| KLMA_10406 | TVP38 | Golgi apparatus membrane protein TVP38 | AP012213.1 | 858831 | C | T | Val158Val |
| KLMA_50490 | SWI5 | metallothionein expression activator | AP012217.1 | 1055034 | TACCACCATTA | T | Asn320fs |
| KLMA_50562 | UPS2 | protein MSF1 | AP012217.1 | 1204779 | C | T | Asp37Asn |
| KLMA_80154 | - | putative zinc metalloproteinase YIL108W | AP012220.1 | 339946 | C | CG | Ala660fs |
| KMR118 | |||||||
| locus_tag | gene | product | reference genome (NCBI refseq acc. no.) | position | REF | ALT | |
| KLMA_10572 | SRV2 | adenylyl cyclase-associated protein | AP012213.1 | 1202978 | G | A | Pro65Pro |
| KLMA_10608 | - | hypothetical protein | AP012213.1 | 1272508 | G | T | Thr373Thr |
| KLMA_20191 | HPR1 | THO complex subunit HPR1 | AP012214.1 | 431928 | C | A | Ile202Ile |
| KLMA_30667 | HKR1 | herpes_gp2 | AP012215.1 | 1429635 | C | G | Pro158Ala |
| KLMA_30667 | HKR1 | herpes_gp2 | AP012215.1 | 1429636 | C | A | Pro158His |
| KLMA_30667 | HKR1 | herpes_gp2 | AP012215.1 | 1429661 | A | AC | Asp168fs |
| KLMA_50490 | SWI5 | metallothionein expression activator | AP012217.1 | 1053571 | G | T | Tyr810stop |
| KLMA_60329 | IXR1 | HMGB-UBF_HMG-box containing protein | AP012218.1 | 691365 | A | AGCTTGG | Ala173_Gln174ins |
| KLMA_70080 | - | conserved hypothetical protein | AP012219.1 | 159037 | G | A | Ala417Thr |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



