Submitted:
31 July 2023
Posted:
02 August 2023
Read the latest preprint version here
Abstract
Memory and emotion, fundamental components of our mental existence, are highly vulnerable to psychiatric disorders like post-traumatic stress disorder (PTSD). This condition has been linked to serotonin (5-HT) metabolism disruptions. Over 95% of the 5-HT precursor tryptophan (Trp) is metabolized through the Trp-kynurenine (KYN) metabolic pathway, but little is known about its role in behavior. Kynurenine aminotransferases are responsible for the production of the Trp-KYN metabolite kynurenic acid. The gene aadat codes for mitochondrial kynurenine aminotransferase isotype 2. We generated CRISPR/Cas9-induced aadat knockout (kat2−/−) mice to examine the consequence of the gene deletion on negative valence in emotion, memory, and motor function in males 8 weeks of age and compared them to their wild-type counterparts. The forced swim test showed increased depression-like behaviors in transgenic mice. Anxiety and memory tests showed no significant differences. The transgenic mice had fewer center field and corner entries, shorter ambulation distances, and fewer jumping counts in the open field test. Overall, the transgenic mice exhibit depression-like behavior in a learned helplessness model, emotional indifference, and motor deficits. Here we present the first evidence that the deletion of the aadat gene triggers depression-like behaviors, uniquely associated with despair experience rather than fear memory. These findings have profound implications, opening avenues for further exploration into the main causes of experience-based depression linked to despair. This investigation has the potential to advance our understanding of these complex conditions and pave the way for improved therapeutic strategies by elucidating the relationship between Trp metabolism and the pathogenesis of PTSD.
Keywords:
posttraumatic stress disorder (PTSD)
; depression
; anxiety
; memory
; tryptophan
; serotonin (5-HT)
; kynurenine
; transaminases
; conversion disorder
; experimental animal models
1. Introduction
The interaction between memory and emotion involves a complex interplay of neural, cognitive, and physiological processes involving the amygdala, hippocampus, and prefrontal cortex [1,2,3,4,5]. Orderly function at multi-layered levels is essential to maintaining sound mental well-being [6,7,8,9]. The reciprocal interaction between cognitive function and affective states can significantly impact each other. Cognitive impairment may contribute to affective disturbances, leading to emotional responses like frustration, anxiety, and stress, especially when individuals perceive a loss of control over their cognitive abilities [10]. Similarly, emotional disturbances such as depression and anxiety can influence memory function, increasing vulnerability to cognitive challenges [11]. This complex bidirectional relationship between cognitive and affective domains can result in alterations in brain structure and function, modifications in behavior and lifestyle, and changes in neurotransmitter systems [12]. Memory impairment and emotional disturbance are associated with a wide range of systematic diseases and neuropsychiatric disorders such as Alzheimer’s disease (AD), Parkinson’s disease, traumatic brain injury, major depressive disorder (MDD), and post-traumatic stress disorder (PTSD) [13,14,15,16,17,18].
The serotonergic nervous system plays an important role in regulating mood, anxiety, and cognition [19]. Serotonin (5-HT) has been found to be involved in various aspects of cognitive processing, including attention, learning, and memory [20]. Studies have shown that 5-HT can enhance the consolidation of long-term memory and improve cognitive flexibility, which is the ability to switch between different cognitive tasks or mental sets [21,22,23,24,25,26,27,28,29,30]. 5-HT has been also implicated in the regulation of mood and anxiety, both of which can have significant effects on cognitive performance [31]. Dysregulation of 5-HT levels and function has been associated with various psychiatric conditions, including MDD, generalized anxiety disorder, obsessive-compulsive disorder, eating disorders, schizophrenia (SCZ), and PTSD [32]. Selective serotonin reuptake inhibitors (SSRIs) are commonly used for these conditions, targeting the serotonergic nervous system [33]. Furthermore, abnormality of the serotonergic nervous system involves other multiple factors, including norepinephrine and dopamine [34].
The intricate relationship between tryptophan (Trp)-kynurenine (KYN) metabolism and 5-HT metabolism holds significant implications for understanding the pathogenesis of mental illnesses [35,36]. The Trp-KYN metabolic system, closely associated with 5-HT metabolism, plays a pivotal role in the production of oxidants and antioxidants, regulation of the immune system, and the balance between neurotoxicity and neuroprotection [37,38]. Over 95% of Trp is catabolized through the KYN route, which safely to say that it governs Trp metabolism [39]. Various factors, including stress, inflammation, and the gut microbiome, influence this system [40,41,42]. Dysregulation of the KYN route has been linked to mental health conditions such as MDD, SCZ, and AD [43]. However, the understanding of the interplay between Trp-KYN and 5-HT metabolism in the pathogenesis of mental illnesses remains limited. Kynurenine aminotransferases (KATs) are members of the pyridoxal-5’-phosphate-dependent enzyme family involved in the KYN metabolic pathway. The KYN metabolism is responsible for the conversion of L-KYN to kynurenic acid (KYNA), a neuroactive metabolite with implications for various central nervous system (CNS) diseases [44,45,46]. Among the KAT isozymes, kynurenine/α-aminoadipate aminotransferase (KAT/AadAT, aka KATII) is a mitochondrial enzyme encoded in the gene aadat [47,48]. The KATII plays a prominent role in KYNA production in the human brain and is considered a crucial target for managing CNS disorders [49].
Preclinical research has significantly contributed to our understanding of mental illnesses by elucidating the underlying pathomechanisms and identifying potential therapeutic targets [50,51,52,53,54,55,56,57,58]. By utilizing preclinical models, researchers have studied the interplay of various factors involved in mental illnesses, explored the effects of genetic, environmental, and pharmacological manipulations on disease phenotypes, and gained a comprehensive understanding of the underlying pathology [59,60,61,62,63,64,65]. The use of in vitro models, such as cell cultures and organoids, has allowed for the examination of intricate molecular pathways associated with mental illnesses [66,67,68,69,70]. In vivo models, including animal models, have provided a platform to investigate the behavioral, cognitive, and physiological aspects of mental disorders [71,72,73,74,75,76,77,78]. These models allow researchers to simulate disease conditions, assess symptomatology, and evaluate the efficacy of potential interventions [79,80]. Transgenic animals serve as essential tools in biomedical research, allowing researchers to mimic specific human conditions by deleting ordinary genes or introducing foreign or altered genes into their genome [81]. These animals provide invaluable information on human diseases, allowing for the investigation of disease mechanisms, testing of potential treatments, and evaluation of therapeutic efficacy [82,83,84,85,86]. Moreover, they provide valuable insights regarding structural changes and imaging techniques for clinical cases [87,88,89,90,91,92,93,94,95,96,97,98,99,100,101]. Together, preclinical and clinical research contribute to the development of innovative therapeutics and personalized medicine [102,103,104,105,106,107].
This study involved manipulating the gene kat2 in mice to create a knockout (kat2−/−) model, allowing us to observe the behavioral consequences of KATII deficiency. By focusing on negative valence in emotional domain, memory acquisition, and motor function, we aimed to gain insights into the role of KATII in these specific behavioral domains in young adult kat2−/− mice. This research contributes to our understanding of the genetic factors influencing behaviors related to emotional valence, memory, and motor function.
2. Materials and Methods
CRISPR/Cas9 was applied on C57BL/6N and CD1 (ICR) mice to generate knockout kat2-/- mice, and Taqman allelic discrimination was used to prove that the gene had been deleted. The emotional domain, including depression-like and anxiety-like behaviors, was evaluated with the modified forced swim test (FST), tail suspension test (TST), elevated plus maze (EPM), open field (OF) test, light dark box (LDB) test, and marble burying (MB) test; the cognitive domain was evaluated with the passive avoidance test (PAT); and the motor domain was evaluated with the OF test.
2.1. Ethical approval
In accordance with the guidelines of the 8th Edition of the Guide for the Care and Use of Laboratory Animals, the Use of Animals in Research of the International Association for the Study of Pain, and the directive of the European Economic Community (86/609/ECC), the experiments conducted in this study received ethical approval from two committees. The Scientific Ethics Committee for Animal Research of the Protection of Animals Advisory Board (XI./95/2020) and the Committee of Animal Research at the University of Szeged (I-74-1/2022) both approved the experiments. Furthermore, Directive 2010/63/EU on the protection of animals used for scientific purposes provides guidance for the ethical evaluation of animal use proposals. The directive allows individual institutions to make determinations based on the recommendations of their ethical review committees. These ethical guidelines and regulations ensure that the experiments conducted on animals adhere to the highest standards of animal welfare and scientific integrity. The approval from the Scientific Ethics Committee for Animal Research of the Protection of Animals Advisory Board and the Committee of Animal Research at the University of Szeged demonstrates that the study was conducted in compliance with these ethical principles and regulations.
2.2. Animals
C57BL/6N and CD1 (ICR) mice were purchased from Japan SLC, Inc. (Hamamatsu, Japan) and Charles River Laboratories International, Inc. (Yokohama, Japan), respectively, in order to generate kat2−/− mice utilizing the CRISPR/Cas9 technique. After genetic modifications, breeding, and transport from Japan to Hungary, the animals were housed in groups of 4-5 in polycarbonate cages (530 cm2 floor space) under pathogen-free conditions in the Animal House of the Department of Neurology, University of Szeged, maintained at 24 ± 1°C and 45–55% relative humidity under a 12:12-h light:dark cycle. Throughout the duration of the investigation, mice had unrestricted access to standard rodent food and water. Animal experiments were conducted humanely in accordance with the Regulations for Animal Experiments of Kyushu University and the Fundamental Guidelines for Proper Conduct of Animal Experiments and Related Activities in Academic Research Institutions governed by the Ministry of Education, Culture, Sports, Science, and Technology of Japan, and with the approval of the Institutional Animal Experiment Committees of Kyushu University. After the transport of the animals, the studies complied with the Ethical Codex of Animal Experiments, were approved by the Committee of the Animal Research of the University of Szeged (I-74-49/2017), were authorized by the National Food Chain Safety Office with permission number XI/1101/2018, and complied with the guidelines for the Use of Animals in Research of the International Association for the Study of Pain and the directive of the European Parliament (2010/63/EU).
The deletion was introduced into the KATs gene using the CRISPR/Cas9 method. The single guide RNAs (sgRNA) were selected using the CRISPRdirect software. Artificially synthesized the sgRNA were purchased from FASMAC (Atsugi, Japan). The Female C57BL/6N mice were injected with pregnant mare serum gonadotropin (PMSG) and human chorionic gonadotropin (hCG) with a 48-h interval, and mated with male C57BL/6N mice. The fertilized one-cell embryos were collected from the oviducts. Then, 25 ng/µl of the sgRNA and 75 ng/µl Guide-it™ Recombinant Cas9 protein (TaKaRa, Japan) were injected into the cytoplasm of these one-cell-stage embryos. The injected two-cell embryos were then transferred into pseudopregnant ICR mice (Figure 1).
In the end result, the kat2−/− mouse line expresses a carboxy-terminal truncated polypeptide consisting of the first 47 amino acids of the intact KATII with a 2-nucleotide deletion (CCDS nucleotide sequence 32-33) in the mRNA.
2.3. DNA extraction and Sequencing
Genomic DNA of tails collected from mice was extracted using NucleoSpin Tissue (MACHEREY-NAGEL GmbH&Co, KG, Germany). Each targeted fragment around the sgRNA targeting site from the extracted genomic DNA as a part of the KATs genes was amplified with TAKARA Ex Taq (Takara Bio, Japan) and the 1st primers pair and subsequently with 2nd primers pair (Table 1). The polymerase chain reaction (PCR) product was purified with a Fast Gene Gel/PCR Extraction Kit (Nippon Genetics Co., Ltd., Tokyo, Japan), and the PCR products were purified by agarose gel electrophoresis and Monarch Gel Extraction Kit (NEW ENGLAND BioLabs Inc.). Then, the PCR products were sequenced with M-KAT II_2nd_R (Table 1).
2.4. Western blotting
For Western blotting, tissue extracts from the liver (20 mg) of the knockout and wild-type (WT) mice were prepared by the Total Protein Extraction Kit for Animal Cultured Cells and Tissues (Invent Biotechnologies, Minnesota, USA) according to the manufacturer’s instructions. Subsequently, the tissue extracts were passed through Protein G HP SpinTrapTM (Cytiva, Buckinghamshire, UK) to remove immunoglobulin G. 14 µl of each sample were mixed with 7 µl of 3X SDS Blue Loading Buffer (New England BioLabs) and separated on a 12% SDS-polyacrylamide gel. Subsequently, the protein was transferred to the membranes. The membranes were blocked and incubated with anti-human KAT II rabbit polyclonal antibody (1:500, Invitrogen, Thermo Fisher Scientific, Massachusetts, USA) at room temperature for 2 h, followed by combination with alkaline phosphatase-labeled secondary goat anti-rabbit IgG FC antibody (1:10000, Sigma-Aldrich) at room temperature for 2 h, followed by visualization of dystrophin and utrophin using Western Blue® Stabilized Substrate for Alkaline Phosphatase (Promega, Madison, WI, USA).
2.5. Behavioral tests
In order to make the results comparable, all behavioral experiments were performed between 8 a.m. and 12 p.m. The animals were transferred to the laboratory, where the measurements were made, one hour before the start of the experiment, thus they had time to acclimatize to the environmental conditions.
2.5.1. Modified forced swim test (FST)
The modified FST was performed as reported previously. The mice were placed individually in a glass cylinder of 12 cm in diameter and 30 cm in height. Water (25±1 °C) was filled to a height of 20 cm. Fresh water was used for each mouse. A 15-min pretest was carried out 24 hours before the 3-min test session. A time-sampling technique was conducted to count the duration of time spent with climbing, swimming, and immobility [108,109].
2.5.2. Tail suspension test (TST)
The mice were placed in a 28x28x23.5 cm wooden box with three side walls and a clip hanging from the top of the box. The animals were suspended by their tails from the base to the middle two-thirds using a clip and allowed to hang for six minutes. We measure the duration of immobility. A cotton swab was pre-attached to the clip’s interior to prevent the mice’s tails from injuring or severely restricting blood circulation. If the animal is able to climb or falls off the clip, it is removed from the experiment and its results are discarded [110,111].
2.5.3. Elevated plus maze (EPM) test
The animals were positioned in a plus-shaped apparatus with four arms measuring 35 x 10 cm. Two of the opposite arms are open, while the other two are closed, forming an angle of 90 degrees. The open arms have no side walls, while the closed arms have walls that are 20 centimeters tall. The entire apparatus is situated 50 cm off the ground. The device is surrounded by a screen that does not display any visual signals. The mouse was placed in the device’s center with its nose facing an open arm and allow it to explore for five minutes. We measure the time spent in each part (open arms, closed arms, and central part). The experiment was captured on video using a camera and software (EthoVision XT14). Between each animal, the apparatus was disinfected with 70% ethanol and left exposed to the air for 5 minutes [112,113].
2.5.4. Light Dark Box (LDB) test
The LDB apparatus is comprised of larger illuminated (2/3 of the box) and smaller dark (1/3 of the box) compartments that are connected by a 5x5 cm door. The length of time a mouse spent in the lighted compartment during the 5-minute session was determined five seconds after a mouse was placed in the bright area. After each session, the box was cleaned with 70% ethanol and allowed to air for five minutes [114,115,116].
2.5.5. Marble burying (MB) test
The animals were placed one by one in a 40 x 24 x 18-cm plastic box. The bottom of it was covered with fresh litter 5 cm thick, and on top we put a 40 x 24 cm, 1 cm thick, transparent plastic cover plate, which has 6 round holes with a diameter of 1 cm, which helped the ventilation. At one end of the box, on top of the litter, there were 16 marbles with a diameter of 1x1x1 cm3 arranged in a regular square shape (the edges are 3.5 cm from the sides of the box and 5 cm from each other). We placed the mice in the arena for 30 minutes, during which they were free to explore. The test was recorded using a video camera and software (EthoVision XT14). The marbles were classified into three different groups based on their condition after the 30-minute period. We considered immobile marbles those that had moved less than 50% from their original location and were not buried at all; the marbles that had moved at least 50% from their original position but were not buried were considered change of position marbles; and finally, the marbles on which the mice scratched the litter were considered buried marbles. We also distinguished between the buried marbles, classifying them into four groups based on the percentage of their total surface covered by litter (1-24%, 25-49%, 50-74%, and 75-100%) [117].
2.5.6. Passive avoidance test (PAT)
Each mouse was individually placed in a box containing two apparatuses with distinct lighting. The animals began in the bright compartment and had five minutes to pass through the 5 x 5-centimeter door into the dark, smaller portion of the box. As soon as the animals entered the dark compartment, they received a 0.3 mA electroshock through their paws, and the door shut. After 10 seconds, the animals were removed, and the experiment was repeated 24 hours later. Those animals that did not enter the dark area within five minutes during the pre-testing phase were omitted from the measurement. The box was cleaned with 70% ethanol and left to air for 5 minutes between mice [118].
2.5.7. Open field (OF) test
A standard table lamp illuminated the center of the 48 cm by 40 cm OF box, while the Conducta 1.0 system (Experimetria Ltd.) monitored the mouse’s movements. Each mouse was placed individually in the center of the box. Ambulation distance, time spent in the center zone, and number of entries to the center zone were measured for 10 minutes. After each session, the box was wiped down with 70% ethanol and allowed to for 5 minutes [119,120].
2.6. Statistical analysis
The Shapiro–Wilk test was used to determine the distribution of data. In addition, we used a Q-Q plot to find out if two sets of data come from the same distribution. Our data followed a normal distribution. One-way ANOVA test was used to evaluate the results of the TST, OF, FST, and EPM followed by the Tamhane post hoc test. For the Marble burying test, a mixed ANOVA model was used followed by the Tamhane post hoc test. Values p <0.05 were considered statistically significant. Our data are reported as means ± SEM for all parameters and groups.
3. Results
3.1. Generation of knockout mice by the CRIPR/Cas9 method
To generate knockout mice of KAT II gene, 25 ng/µl of sgRNA and 75ng/µl Cas9 protein were injected into the cytoplasm of the one-cell-stage embryos. Sequencing analyses with their founder mice showed that various deletions and/or insertions were introduced in the target sequence. One of the founders was selected and established the homozygous mouse line for further analyses. KAT II knockout mouse line expresses a carboxy-terminal truncated polypeptide consisting of the first 47 amino acids of the intact KAT II with 2 nucleotides deletion (CCDS nucleotide sequence 32-33) in the mRNA. Western blotting with antibodies against KAT II revealed that the band with approximately 50-kDa supposed to be KAT II was not detected in KAT II knockout mouse, while it was detected in the WT mouse (Figure 2).
3.2. Depression-like behavior
3.2.1. Forced swim test (FST)
3.2.2. Tail suspension test (TST)
There were no statistically significant distinctions observed between the transgenic mice and their WT counterparts (Table 2).
3.3. Aversive associative memory
The statistical analysis revealed no significant differences in PAT performance between kat2-/- mice and their WT counterparts (Table 2).
3.4. Anxiety-like behavior
3.4.1. Elevated plus maze (EPM) test, Light Dark Box (LDB) test, and Marble burying (MB) test
Comparing kat2-/- mice and WT mice using EPM, LDB, and MB tests, no statistically significant differences were observed (Table 2).
3.4.2. Open field (OF) test
3.5. Exploratory behavior and motor function
4. Discussion
Dysregulation of 5-HT metabolism plays a major role in the emergence of mental symptoms, and increasing attention has been paid to its imbalance with other neurotransmitters, including dopamine and norepinephrine, and biosystems such as substance P [121,122,123]. Alteration of 5-HT precursor Trp metabolism has also been observed in mental illnesses; however, little is known about its relationship with the Trp-KYN metabolic system [124]. KATs are cytosolic and mitochondrial aminotransferases that convert KYN to KYNA [125,126,127,128]. The isoform KATII is exclusively located in the mitochondria, which therefore may have a direct influence on cellular bioenergetics [129,130]. Using the CRISPR/Cas9, the gene kat2 was knocked out to generate kat2−/− mice to study the negative valence of the emotional domain and the impact of the aadat gene knockout in the young adult of 8 weeks to characterize change in behaviors.
Our study revealed that the aadat gene deletion has led to a distinguished behavioral phenotype in the negative emotional domain in a unique manner. The transgenic mice exhibit significantly more depression-like behaviors due to increased immobility time, which is predominantly caused by decreased swimming time in FST. This suggests that the kat2−/− mice showed more depressive-like behavior than their WT counterparts and that the depression-like behavior was more likely induced via the serotonergic nervous system. However, TST did not reveal any significant difference in immobility time between the two strains. This inconsistent finding in the tests for depression-like behavior may lie in the presence of conditioning in the FST. Nevertheless, the PA test failed to show acquisition of aversive associative memory in kat2−/− mice. It may be safe to state that the depression-like behavior was induced through despair experience but not fear memory. Tests for anxiety-like behaviors, including the EPM test, the LDB test, and the MB test, did not show any significant difference between the two strains. However, kat2−/− mice exhibited significantly fewer entry times into both center field and corners than their WT counterparts in the OF test. This behavior may suggest the presence of a la belle indifference-like trait in kat2−/− mice. Further, the OF test revealed significantly shorter ambulation distance and fewer jumping counts in kat2−/− mice. Thus, kat2−/− mice showed despair experience-based depression-like behavior without anxiety-like behaviors, with la belle indifference-like traits, and decreased motor functions. The results of this study demonstrate conclusively that the deletion of the aadat gene results in a phenotype characterized by motor deficits and a depression-like phenotype based on the experience of despair. The behavioral phenotype is comparable to human PTSD. In addition, the presence of a trait resembling la belle indifference may be indicative of complex PTSD, which is characterized by emotional dysregulation [131].
After receiving sensory input from the thalamus, fear memory is primarily encoded and stored in the amygdala, which also consolidates and retrieves fear memories from the original fear-inducing stimuli. Fear memory is thought to be mediated by the release of stress hormones such as adrenaline and cortisol, which activate the sympathetic nervous system and the hypothalamic-pituitary-adrenal axis [132]. This study does not show evidence of fear memory acquisition. In contrast, despair memory is primarily encoded and stored in the prefrontal cortex, which is also involved in regulating emotions playing a key role in the cognitive and emotional processing of negative experiences [133]. Despair memories can be also retrieved from long-term storage in response to cues that are associated with the original negative experience [134]. Despair memory involves dysregulation of the neuro-transmitters serotonin, norepinephrine, and dopamine [135]. Both fear memory and despair memory involve the encoding and retrieval of memories, but they are associated with different brain regions, neurotransmitters, and neural circuits.
Furthermore, despair memory and despair experience are distinct concepts. Despair experience refers to the immediate subjective experience of despair or hopelessness, which could occur in response to a current situation or trigger rather than a memory of a past event [136,137]. Long-term memories are consolidated and retrieved during despair memory, which is subject to influences from stress and emotion [138]. On the other hand, a despair experience involves the triggering of immediate emotional responses that are susceptible to a variety of influences, such as cognitive assessments, environmental cues, and physiological states [139]. Additionally, la belle indifference results from a discrepancy between the cognitive and emotional processing of symptoms, which involves altered emotional processing in the amygdala and insula, altered self-awareness in the medial prefrontal cortex, and altered activity in the somatosensory cortex via dopamine and serotonin [139]. Thus, kat2−/− mice show more despair-based depression-like behavior involving a change in 5-HT metabolism.
About 60% of individuals with depression on antidepressants including selective serotonin uptake inhibitors (SSRIs) for two months benefits from 50% reduction in their symptoms [141]. The observation makes prevail the monoamine hypothesis that the pathogenesis of depression lies in the low 5-HT level. Several transgenic models have generated to study behaviors in 5-HT dysmetabolism. The gene Tph produces tryptophan hydroxylase responsible for the rate-liming reaction of 5-HT synthesis [142]. Preclinical studies reported that a normal level of 5-HT and no change in behaviors in Tph1−/− mice, while the behaviors of Tph2−/− mice remain inconclusive [143,144]. The knock-in mice of the TPH2 variant (R439H) showed depression-like behavior in TST [145]. Intriguingly, the double knockout Tph1/Tph2−/− mice showed antidepression-like behavior in FST, but depression-like behavior in TST and anxiety-like behavior in MB test with low levels of 5-HT in the brain and the periphery [146]. 5-HT receptors may play an important role in the exhibition of PTSD-like phenotypes. 5-HT 1A receptor knockout 5-HT1AR−/− mice show increased fear memory to contextual cues [147]. 5-HT 2C receptor knockout 5-HT2CR−/− mice attenuates fear responses in contextual or cued but not compound context-cue fear conditioning [148]. 5-HT transporter gene knockout 5-HTT−/− mice show impaired stress response and impaired fear extinction with abnormal corticolimbic structure [149].
Over 95% of 5-HT precursor Trp is catabolized in the Trp-KYN metabolic system, producing a variety of bioactive molecules including oxidants, antioxidants, inflammation suppressants, neurotoxins, neuroprotectants, and/or immunomodulators [150]. More and more data revealed the disturbance of KYN metabolism in MDD, bipolar disorder, and SCZ [151,152,153]. Previously, it was proposed that KYN metabolites are categorized into neuroprotective or neurotoxic molecules [154]. However, a growing number of evidence support that the action of KYN metabolites is versatile possibly depending on concentrations and microenvironment [155]. Previously, the cognitive domain and motor function of 129/SvEv kat2−/− mice were reported. The transgenic mice showed transitory hyperlocomotive activity, and abnormal motor coordination at postnatal day 21, but revealed significantly increased cognitive functions in object exploration and recognition task in PAT and T-maze test during postnatal day 17 to 26 [156,157].
Other biosystems play an important role in the pathogenesis of PTSD include dopaminergic and gamma-aminobutyric acid (GABA)ergic system and cannabinoids. Catechol-O-methyltransferase (COMT) degrades dopamine. COMT−/− mice exhibited an increased response to repeated stress exposures [158]. Glutamic acid decarboxylase (GAD) synthesizes GABA [159]. GAD6−/− mice shows increased generalized fear and impaired extinction of cued fear [160]. GABA receptor subunit B1a knockout GABAB1a−/− mice showed a generalization of conditioned fear to nonconditioned stimuli [161]. Cannabinoid 1 receptor (CBIR) knockout CB1R−/− mice showed an increased response to repeated stress exposures [162].
The potential of this study is to characterize the negative valence of emotional domain in context with fear memory and despair experience in the young adult (8 week) of kat2−/− mice. The findings complement the previous studies of kat2−/− mice in the early adolescence (2 and 1/2 to 4 week) to reveal that, toward the adulthood, there is a dynamic change in emotional susceptibility and motor function derived from despair experience in adjunct to Trp metabolism. This study may open a new avenue toward understanding the deletion of the gene aadat as a boundary between fear and despair, a distinction between memory and experience, their mechanism in the formation of intrusive memories, and the pathogenesis leading to PTSD. The ultimate goal is to probe a potential interventionable stage in age where the progression of PTSD is preventable and to identify targets which drugs or psychotherapy can relieve symptoms of PTSD. The greatest challenge lies in preclinical animal models that are difficult to simulate and interpolate to mental illnesses to achieve high model validity.
The results of this study suggest that behavioral sampling in rodents may be able to distinguish between fear-, memory-, and despair-based depression-like behavior, which may result from the deletion of the Trp metabolism gene. Further studies may reinforce this finding by incorporating neurochemical, neurogenetic, and electrophysiological biomarkers such as c-fos, brain-derived neurotrophic factor, and electroencephalogram signature. Preclinical research is a driving force of advance in clinical application to precision medicine, novel drug discovery, and building personalized treatment plan. The authors acknowledge the presence of the weakness and the limitation in this study. The Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition pays more attention to the presence of four symptom clusters in diagnosis of PTSD [163]. The transgenic mice did not show signs referring to negative cognitions and mood, and this study did not investigate arousal state and reactivity of the transgenic mice.
5. Conclusions
Psychiatric disorders, including PTSD, have a significant impact on memory and emotion, and disruptions in 5-HT metabolism have been associated with these disorders. The Trp-KYN metabolic pathway plays a crucial role in metabolizing over 95% of the serotonin precursor Trp. To investigate the effects of gene deletion on negative valence in emotion, memory, and motor function, transgenic kat2−/− mice were created and compared to WT mice. The kat2−/− mice exhibited depression-like behavior characterized by despair experiences, diminished motor functions, and la belle indifference-like characteristics without anxiety-like behavior. This study provides insights into the negative valence of the emotional domain in the context of fear memory and despair experiences in 8-week-old kat2−/− mice. It has the potential to identify a stage in aging where the progression of PTSD can be prevented and targets for drug treatments or psychotherapy to alleviate PTSD symptoms can be identified. Understanding the complex interplay between memory, emotion, and genetic factors is crucial for advancing our knowledge of psychiatric disorders. By elucidating the specific effects of gene deletion on negative valence and related behaviors, this research contributes to our understanding of the underlying mechanisms and potential interventions.
Author Contributions
Conceptualization, Á.S., E.O., L.V., and M.T.; methodology, Á.S., E.S., M.S., K.T., K.O., H.I., S.Y., and M.T.; software, Á.S., E.S., M.S., K.O.; validation, Á.S., E.S., M.S., K.T., K.O., and M.T.; formal analysis, Á.S., E.S., M.S., K.O., and M.T.; investigation, Á.S., E.S., K.T., K.O., and M.T.; resources, Á.S. and E.O.; data curation, Á.S., E.S., K.O.; writing—original draft preparation, Á.S., M.T.; writing—review and editing, Á.S., E.S., J.T., E.O., L.V., and M.T.; visualization, Á.S., K.O.; supervision, J.T., E.O., L.V., and M.T.; project administration, J.T., E.O., and L.V; funding acquisition, J.T., E.O., L.V., and M.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by OTKA-138125-K, TUDFO/47138-1/2019-ITM, ELKH-SZTE Eötvös Loránd Research Network, the University of Szeged, and JSPS Joint Research Projects under Bilateral Programs Grant Number JPJSBP120203803.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Ethics Committee of the Scientific Ethics Committee for Animal Research of the Protection of Animals Advisory Board (XI./95/2020) and the Committee of Animal Research at the University of Szeged (I-74-1/2022).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The figures are created with biorender.com.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| 5-HT | serotonin |
| AD | Alzheimer’s disease |
| CBIR | cannabinoid 1 receptor |
| CNS | central nervous system |
| COMT | catechol-O-methyltransferase |
| EPM | elevated plus maze |
| FST | forced swim test |
| GABA | gamma-aminobutyric acid |
| GAD | glutamic acid decarboxylase |
| hCG | human chorionic gonadotropin |
| KATII | α-aminoadipate aminotransferase/kynurenine aminotransferase II |
| KATs | kynurenine aminotransferases |
| KYN | kynurenine |
| KYNA | kynurenic acid |
| LDB | light dark box |
| MB | marble burying |
| MDD | major depressive disorder |
| OF | open field |
| PAT | passive avoidance test |
| PCR | polymerase chain reaction |
| PMSG | pregnant mare serum gonadotropin |
| PTSD | posttraumatic stress disorder |
| SCZ | schizophrenia |
| sgRNA | single guide RNA |
| SSRI | selective serotonin reuptake inhibitors |
| Trp | tryptophan |
| TST | tail suspension test |
| WT | wild-type |
References
- Tyng, C.M.; Amin, H.U.; Saad, M.N.M.; Malik, A.S. The Influences of Emotion on Learning and Memory. Front. Psychol. 2017, 8, 1454. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, S.; Garofalo, S.; di Pellegrino, G.; Starita, F. Revaluing the Role of vmPFC in the Acquisition of Pavlovian Threat Conditioning in Humans. J. Neurosci. 2020, 40, 8491–8500. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, S.; Harrison, B.J.; Falana, M.A. Does the human ventromedial prefrontal cortex support fear learning, fear extinction or both? A commentary on subregional contributions. Mol. Psychiatry 2022, 27, 784–786. [Google Scholar] [CrossRef] [PubMed]
- Sumsuzzman, D.M.; Choi, J.; Jin, Y.; Hong, Y. Neurocognitive effects of melatonin treatment in healthy adults and individuals with Alzheimer’s disease and insomnia: A systematic review and meta-analysis of randomized controlled trials. Neurosci. Biobehav. Rev. 2021, 127, 459–473. [Google Scholar] [CrossRef] [PubMed]
- Borgomaneri, S.; Battaglia, S.; Sciamanna, G.; Tortora, F.; Laricchiuta, D. Memories are not written in stone: Re-writing fear memories by means of non-invasive brain stimulation and optogenetic manipulations. Neurosci. Biobehav. Rev. 2021, 127, 334–352. [Google Scholar] [CrossRef] [PubMed]
- Hasson, U.; Chen, J.; Honey, C.J. Hierarchical process memory: memory as an integral component of information processing. Trends Cogn. Sci. 2015, 19, 304–13. [Google Scholar] [CrossRef] [PubMed]
- Clewett, D.; Sakaki, M.; Nielsen, S.; Petzinger, G.; Mather, M. Noradrenergic mechanisms of arousal’s bidirectional effects on episodic memory. Neurobiol. Learn. Mem. 2017, 137, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, S.; Nazzi, C.; Thayer, J.F. Fear-induced bradycardia in mental disorders: Foundations, current advances, future perspectives. Neurosci. Biobehav. Rev. 2023, 149, 105163. [Google Scholar] [CrossRef]
- Battaglia, S.; Di Fazio, C.; Vicario, C.M.; Avenanti, A. Neuropharmacological Modulation of N-methyl-D-aspartate, Noradrenaline and Endocannabinoid Receptors in Fear Extinction Learning: Synaptic Transmission and Plasticity. Int. J. Mol. Sci. 2023, 24, 5926. [Google Scholar] [CrossRef]
- Hayes, J.P.; Vanelzakker, M.B.; Shin, L.M. Emotion and cognition interactions in PTSD: a review of neurocognitive and neuroimaging studies. Front. Integr. Neurosci. 2012, 6, 89. [Google Scholar] [CrossRef]
- Dillon, D.G.; Pizzagalli, D.A. Mechanisms of Memory Disruption in Depression. Trends Neurosci. 2018, 41, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Hasebe, K.; Kendig, M.D.; Morris, M.J. Mechanisms Underlying the Cognitive and Behavioural Effects of Maternal Obesity. Nutrients 2021, 13, 240. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Vélez, E.; Feinstein, J.S.; Tranel, D. Feelings without memory in Alzheimer disease. Cogn. Behav. Neurol. 2014, 27, 117–29. [Google Scholar] [CrossRef] [PubMed]
- Mack, J.; Marsh, L. Parkinson’s Disease: Cognitive Impairment. Focus (Am. Psychiatr. Publ). 2017, 15, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Mckee, A.C.; Daneshvar, D.H. The neuropathology of traumatic brain injury. Handb. Clin. Neurol. 2015, 127, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Feng, L.; Liu, X.; Zhang, M.; Fu, B.; Wang G, Lu, S. ; Zhong, N.; Hu, B. Emotional working memory in patients with major depressive disorder. J. Int. Med. Res. 2018, 46, 1734–1746. [Google Scholar] [CrossRef]
- Samuelson, K.W. Post-traumatic stress disorder and declarative memory functioning: a review. Dialogues Clin. Neurosci. 2011, 13, 346–51. [Google Scholar] [CrossRef] [PubMed]
- Dere, E.; Pause, B.M.; Pietrowsky, R. Emotion and episodic memory in neuropsychiatric disorders. Behav. Brain Res. 2010, 215, 162–71. [Google Scholar] [CrossRef]
- Berger, M.; Gray, J.A.; Roth, B.L. The expanded biology of serotonin. Annu. Rev. Med. 2009, 60, 355–66. [Google Scholar] [CrossRef]
- Švob Štrac, D.; Pivac, N.; Mück-Šeler, D. The serotonergic system and cognitive function. Transl. Neurosci. 2016, 7, 35–49. [Google Scholar] [CrossRef]
- Battaglia, S.; Cardellicchio, P.; Di Fazio, C.; Nazzi, C.; Fracasso, A.; Borgomaneri, S. Stopping in (e)motion: Reactive action inhibition when facing valence-independent emotional stimuli. Front. Behav. Neurosci. 2022, 16, 998714. [Google Scholar] [CrossRef] [PubMed]
- Cowen, P.; Sherwood, A.C. The role of serotonin in cognitive function: evidence from recent studies and implications for understanding depression. J. Psychopharmacol. 2013, 27, 575–83. [Google Scholar] [CrossRef]
- Battaglia, S.; Cardellicchio, P.; Di Fazio, C.; Nazzi, C.; Fracasso, A.; Borgomaneri, S. The Influence of Vicarious Fear-Learning in “Infecting” Reactive Action Inhibition. Front. Behav. Neurosci. 2022, 16, 946263. [Google Scholar] [CrossRef]
- Battaglia, S.; Thayer, J.F. Functional interplay between central and autonomic nervous systems in human fear conditioning. Trends Neurosci. 2022, 45, 504–506. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, S.; Orsolini, S.; Borgomaneri, S.; Barbieri, R.; Diciotti, S.; di Pellegrino, G. Characterizing cardiac autonomic dynamics of fear learning in humans. Psychophysiology 2022, 59, e14122. [Google Scholar] [CrossRef] [PubMed]
- Di Gregorio, F.; La Porta, F.; Petrone, V.; Battaglia, S.; Orlandi, S.; Ippolito, G.; Romei, V.; Piperno, R.; Lullini, G. Accuracy of EEG Biomarkers in the Detection of Clinical Outcome in Disorders of Consciousness after Severe Acquired Brain Injury: Preliminary Results of a Pilot Study Using a Machine Learning Approach. Biomedicines 2022, 10, 1897. [Google Scholar] [CrossRef] [PubMed]
- Borgomaneri, S.; Battaglia, S.; Avenanti, A.; Pellegrino, G.D. Don’t Hurt Me No More: State-dependent Transcranial Magnetic Stimulation for the treatment of specific phobia. J. Affect. Disord. 2021, 286, 78–79. [Google Scholar] [CrossRef]
- Khalil, R.; Godde, B.; Karim, A.A. The Link Between Creativity, Cognition, and Creative Drives and Underlying Neural Mechanisms. Front. Neural. Circuits. 2019, 13, 18. [Google Scholar] [CrossRef]
- Borgomaneri, S.; Battaglia, S.; Garofalo, S.; Tortora, F.; Avenanti, A.; di Pellegrino, G. State-Dependent TMS over Prefrontal Cortex Disrupts Fear-Memory Reconsolidation and Prevents the Return of Fear. Curr. Biol. 2020, 30, 3672–3679. [Google Scholar] [CrossRef]
- Battaglia, S.; Garofalo, S.; di Pellegrino, G. Context-dependent extinction of threat memories: influences of healthy aging. Sci. Rep. 2018, 8, 12592. [Google Scholar] [CrossRef]
- Albert, P.R.; Vahid-Ansari, F.; Luckhart, C. Serotonin-prefrontal cortical circuitry in anxiety and depression phenotypes: pivotal role of pre- and post-synaptic 5-HT1A receptor expression. Front. Behav. Neurosci. 2014, 8, 199. [Google Scholar] [CrossRef] [PubMed]
- Brewerton, T.D. Toward a unified theory of serotonin dysregulation in eating and related disorders. Psychoneuroendocrinology 1995, 20, 561–90. [Google Scholar] [CrossRef] [PubMed]
- Chu, A.; Wadhwa, R. Selective Serotonin Reuptake Inhibitors. [Updated 2023 May 1]. In: StatPearls [https://www.ncbi.nlm.nih.gov/books/NBK554406/]. Treasure Island (FL): StatPearls Publishing; 2023 Jan-. (accessed on 20 July 2023).
- Teleanu, R.I.; Niculescu, A.G.; Roza, E.; Vladâcenco, O.; Grumezescu, A.M.; Teleanu, D.M. Neurotransmitters-Key Factors in Neurological and Neurodegenerative Disorders of the Central Nervous System. Int. J. Mol. Sci. 2022, 23, 5954. [Google Scholar] [CrossRef] [PubMed]
- Muneer, A. Kynurenine Pathway of Tryptophan Metabolism in Neuropsychiatric Disorders: Pathophysiologic and Therapeutic Considerations. Clin. Psychopharmacol. Neurosci. 2020, 18, 507–526. [Google Scholar] [CrossRef]
- Tanaka, M.; Török, N.; Tóth, F.; Szabó, Á.; Vécsei, L. Co-Players in Chronic Pain: Neuroinflammation and the Tryptophan-Kynurenine Metabolic Pathway. Biomedicines 2021, 9, 897. [Google Scholar] [CrossRef]
- Huang, Y.; Zhao, M.; Chen, X.; Zhang, R.; Le, A.; Hong, M.; Zhang, Y.; Jia, L.; Zang, W.; Jiang, C.; Wang, J.; Fan, X.; Wang, J. Tryptophan Metabolism in Central Nervous System Diseases: Pathophysiology and Potential Therapeutic Strategies. Aging Dis. 2023, 14, 858–878. [Google Scholar] [CrossRef]
- Polyák, H.; Galla, Z.; Nánási, N.; Cseh, E.K.; Rajda, C.; Veres, G.; Spekker, E.; Szabó, Á.; Klivényi, P.; Tanaka, M.; Vécsei, L. The Tryptophan-Kynurenine Metabolic System Is Suppressed in Cuprizone-Induced Model of Demyelination Simulating Progressive Multiple Sclerosis. Biomedicines 2023, 11, 945. [Google Scholar] [CrossRef]
- Hubková, B.; Valko-Rokytovská, M.; Čižmárová, B.; Zábavníková, M.; Mareková, M.; Birková, A. Tryptophan: Its Metabolism along the Kynurenine, Serotonin, and Indole Pathway in Malignant Melanoma. Int. J. Mol. Sci. 2022, 23, 9160. [Google Scholar] [CrossRef]
- Mor, A.; Tankiewicz-Kwedlo, A.; Krupa, A.; Pawlak, D. Role of Kynurenine Pathway in Oxidative Stress during Neurodegenerative Disorders. Cells 2021, 10, 1603. [Google Scholar] [CrossRef]
- Bosi, A.; Banfi, D.; Bistoletti, M.; Giaroni, C.; Baj, A. Tryptophan Metabolites Along the Microbiota-Gut-Brain Axis: An Interkingdom Communication System Influencing the Gut in Health and Disease. Int. J. Tryptophan. Res. 2020, 13, 1178646920928984. [Google Scholar] [CrossRef]
- Spekker, E.; Tanaka, M.; Szabó, Á.; Vécsei, L. Neurogenic Inflammation: The Participant in Migraine and Recent Advancements in Translational Research. Biomedicines 2022, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Réus, G.Z.; Jansen, K.; Titus, S.; Carvalho, A.F.; Gabbay, V.; Quevedo, J. Kynurenine pathway dysfunction in the pathophysiology and treatment of depression: Evidences from animal and human studies. J. Psychiatr. Res. 2015, 68, 316–28. [Google Scholar] [CrossRef]
- Tanaka, M.; Tóth, F.; Polyák, H.; Szabó, Á.; Mándi, Y.; Vécsei, L. Immune Influencers in Action: Metabolites and Enzymes of the Tryptophan-Kynurenine Metabolic Pathway. Biomedicines 2021, 9, 734. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Bohár, Z.; Martos, D.; Telegdy, G.; Vécsei, L. Antidepressant-like effects of kynurenic acid in a modified forced swim test. Pharmacol. Rep. 2020, 72, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Martos, D.; Tuka, B.; Tanaka, M.; Vécsei, L.; Telegdy, G. Memory Enhancement with Kynurenic Acid and Its Mechanisms in Neurotransmission. Biomedicines 2022, 10, 849. [Google Scholar] [CrossRef]
- National Library of Medicine. Available online: https://www.ncbi.nlm.nih.gov/gene/51166.
- Goh, D.L.; Patel, A.; Thomas, G.H.; Salomons, G.S.; Schor, D.S.; Jakobs, C.; Geraghty, M.T. Characterization of the human gene encoding alpha-aminoadipate aminotransferase (AADAT). Mol. Genet. Metab. 2002, 76, 172–80. [Google Scholar] [CrossRef] [PubMed]
- Modoux, M.; Rolhion, N.; Mani, S.; Sokol, H. Tryptophan Metabolism as a Pharmacological Target. Trends Pharmacol. Sci. 2021, 42, 60–73. [Google Scholar] [CrossRef]
- Palotai, M.; Telegdy, G.; Tanaka, M.; Bagosi, Z.; Jászberényi, M. Neuropeptide AF induces anxiety-like and antidepressant-like behavior in mice. Behav. Brain Res. 2014, 274, 264–9. [Google Scholar] [CrossRef]
- Tanaka, M.; Telegdy, G. Involvement of adrenergic and serotonergic receptors in antidepressant-like effect of urocortin 3 in a modified forced swimming test in mice. Brain Res. Bull. 2008, 77, 301–5. [Google Scholar] [CrossRef]
- Tanaka, M.; Schally, A.V.; Telegdy, G. Neurotransmission of the antidepressant-like effects of the growth hormone-releasing hormone antagonist MZ-4-71. Behav. Brain Res. 2012, 228, 388–91. [Google Scholar] [CrossRef]
- Tanaka, M.; Telegdy, G. Neurotransmissions of antidepressant-like effects of neuromedin U-23 in mice. Behav. Brain Res. 2014, 259, 196–9. [Google Scholar] [CrossRef] [PubMed]
- Telegdy, G.; Tanaka, M.; Schally, A.V. Effects of the growth hormone-releasing hormone (GH-RH) antagonist on brain functions in mice. Behav. Brain Res. 2011, 224, 155–8. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Csabafi, K.; Telegdy, G. Neurotransmissions of antidepressant-like effects of kisspeptin-13. Regul. Pept. 2013, 180, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Rákosi, K.; Masaru, T.; Zarándia, M.; Telegdy, G.; Tóth, G.K. Short analogs and mimetics of human urocortin 3 display antidepressant effects in vivo. Peptides 2014, 62, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Vécsei, L. Editorial of Special Issue ‘Dissecting Neurological and Neuropsychiatric Diseases: Neurodegeneration and Neuroprotection’. Int. J. Mol. Sci. 2022, 23, 6991. [Google Scholar] [CrossRef] [PubMed]
- Tran, K.N.; Nguyen, N.P.K.; Nguyen, L.T.H.; Shin, H.-M.; Yang, I.-J. Screening for Neuroprotective and Rapid Antidepressant-like Effects of 20 Essential Oils. Biomedicines 2023, 11, 1248. [Google Scholar] [CrossRef]
- Tanaka, M.; Szabó, Á.; Vécsei, L. Integrating Armchair, Bench, and Bedside Research for Behavioral Neurology and Neuropsychiatry: Editorial. Biomedicines 2022, 10, 2999. [Google Scholar] [CrossRef]
- Tanaka, M.; Kádár, K.; Tóth, G.; Telegdy, G. Antidepressant-like effects of urocortin 3 fragments. Brain Res. Bull. 2011, 84, 414–8. [Google Scholar] [CrossRef]
- Telegdy, G.; Adamik, A.; Tanaka, M.; Schally, A.V. Effects of the LHRH antagonist Cetrorelix on affective and cognitive functions in rats. Regul. Pept. 2010, 159, 142–147. [Google Scholar] [CrossRef]
- Baliellas, D.E.M.; Barros, M.P.; Vardaris, C.V.; Guariroba, M.; Poppe, S.C.; Martins, M.F.; Pereira, Á.A.F.; Bondan, E.F. Propentofylline Improves Thiol-Based Antioxidant Defenses and Limits Lipid Peroxidation following Gliotoxic Injury in the Rat Brainstem. Biomedicines 2023, 11, 1652. [Google Scholar] [CrossRef]
- Montanari, M.; Imbriani, P.; Bonsi, P.; Martella, G.; Peppe, A. Beyond the Microbiota: Understanding the Role of the Enteric Nervous System in Parkinson’s Disease from Mice to Human. Biomedicines 2023, 11, 1560. [Google Scholar] [CrossRef] [PubMed]
- Garifulin, R.; Davleeva, M.; Izmailov, A.; Fadeev, F.; Markosyan, V.; Shevchenko, R.; Minyazeva, I.; Minekayev, T.; Lavrov, I.; Islamov, R. Evaluation of the Autologous Genetically Enriched Leucoconcentrate on the Lumbar Spinal Cord Morpho-Functional Recovery in a Mini Pig with Thoracic Spine Contusion Injury. Biomedicines 2023, 11, 1331. [Google Scholar] [CrossRef]
- Bueno, C.R.d.S.; Tonin, M.C.C.; Buchaim, D.V.; Barraviera, B.; Ferreira Junior, R.S.; Santos, P.S.d.S.; Reis, C.H.B.; Pastori, C.M.; Pereira, E.d.S.B.M.; Nogueira, D.M.B.; Cini, M.A.; Rosa Junior, G.M.; Buchaim, R.L. Morphofunctional Improvement of the Facial Nerve and Muscles with Repair Using Heterologous Fibrin Biopolymer and Photobiomodulation. Pharmaceuticals 2023, 16, 653. [Google Scholar] [CrossRef] [PubMed]
- Scalise, S.; Zannino, C.; Lucchino, V.; Lo Conte, M.; Scaramuzzino, L.; Cifelli, P.; D’Andrea, T.; Martinello, K.; Fucile, S.; Palma, E.; Gambardella, A.; Ruffolo, G.; Cuda, G.; Parrotta, E.I. Human iPSC Modeling of Genetic Febrile Seizure Reveals Aberrant Molecular and Physiological Features Underlying an Impaired Neuronal Activity. Biomedicines 2022, 10, 1075. [Google Scholar] [CrossRef] [PubMed]
- Datki, Z.; Sinka, R. Translational biomedicine-oriented exploratory research on bioactive rotifer-specific biopolymers. Adv. Clin. Exp. Med. 2022, 31, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Kwon, K.-M.; Lee, M.-J.; Chung, H.-S.; Pak, J.-H.; Jeon, C.-J. The Organization of Somatostatin-Immunoreactive Cells in the Visual Cortex of the Gerbil. Biomedicines 2022, 10, 92. [Google Scholar] [CrossRef]
- Simon, C.; Soga, T.; Ahemad, N.; Bhuvanendran, S.; Parhar, I. Kisspeptin-10 Rescues Cholinergic Differentiated SHSY-5Y Cells from α-Synuclein-Induced Toxicity In Vitro. Int. J. Mol. Sci. 2022, 23, 5193. [Google Scholar] [CrossRef]
- Chen, B.; Hasan, M.M.; Zhang, H.; Zhai, Q.; Waliullah, A.S.M.; Ping, Y.; Zhang, C.; Oyama, S.; Mimi, M.A.; Tomochika, Y.; Nagashima, Y.; Nakamura, T.; Kahyo, T.; Ogawa, K.; Kaneda, D.; Yoshida, M.; Setou, M. UBL3 Interacts with Alpha-Synuclein in Cells and the Interaction Is Downregulated by the EGFR Pathway Inhibitor Osimertinib. Biomedicines 2023, 11, 1685. [Google Scholar] [CrossRef] [PubMed]
- Song, A.; Cho, G.W.; Vijayakumar, K.A.; Moon, C.; Ang, M.J.; Kim, J.; Park, I.; Jang, C.H. Neuroprotective Effect of Valproic Acid on Salicylate-Induced Tinnitus. Int. J. Mol. Sci. 2021, 23, 23. [Google Scholar] [CrossRef]
- Ibos, K.E.; Bodnár, É.; Bagosi, Z.; Bozsó, Z.; Tóth, G.; Szabó, G.; Csabafi, K. Kisspeptin-8 Induces Anxiety-Like Behavior and Hypolocomotion by Activating the HPA Axis and Increasing GABA Release in the Nucleus Accumbens in Rats. Biomedicines 2021, 9, 112. [Google Scholar] [CrossRef]
- Uliana, D.L.; Zhu, X.; Gomes, F.V.; Grace, A.A. Using animal models for the studies of schizophrenia and depression: The value of translational models for treatment and prevention. Front. Behav. Neurosci. 2022, 24, 16–935320. [Google Scholar] [CrossRef] [PubMed]
- Puri, S.; Kenyon, B.M.; Hamrah, P. Immunomodulatory Role of Neuropeptides in the Cornea. Biomedicines 2022, 10, 1985. [Google Scholar] [CrossRef]
- Mirchandani-Duque, M.; Barbancho, M.A.; López-Salas, A.; Alvarez-Contino, J.E.; García-Casares, N.; Fuxe, K.; Borroto-Escuela, D.O.; Narváez, M. Galanin and Neuropeptide Y Interaction Enhances Proliferation of Granule Precursor Cells and Expression of Neuroprotective Factors in the Rat Hippocampus with Consequent Augmented Spatial Memory. Biomedicines 2022, 10, 1297. [Google Scholar] [CrossRef] [PubMed]
- Taschereau-Dumouchel, V.; Michel, M.; Lau, H.; Hofmann, S.G.; LeDoux, J.E. Putting the “mental” back in “mental disorders”: a perspective from research on fear and anxiety. Mol. Psychiatry 2022, 27, 1322–1330. [Google Scholar] [CrossRef]
- Li, J.; Li, C.; Subedi, P.; Tian, X.; Lu, X.; Miriyala, S.; Panchatcharam, M.; Sun, H. Light Alcohol Consumption Promotes Early Neurogenesis Following Ischemic Stroke in Adult C57BL/6J Mice. Biomedicines 2023, 11, 1074. [Google Scholar] [CrossRef] [PubMed]
- Petković, A.; Chaudhury, D. Encore: Behavioural animal models of stress, depression and mood disorders. Front. Behav. Neurosci. 2022, 16, 931964. [Google Scholar] [CrossRef] [PubMed]
- Bahor, Z.; Nunes-Fonseca, C.; Thomson, L.D.; Sena, E.S.; Macleod, M.R. Improving our understanding of the in vivo modelling of psychotic disorders: A protocol for a systematic review and meta-analysis. Evid. Based Preclin. Med. 2016, 3, e00022. [Google Scholar] [CrossRef] [PubMed]
- Nasini, S.; Tidei, S.; Shkodra, A.; De Gregorio, D.; Cambiaghi, M.; Comai, S. Age-Related Effects of Exogenous Melatonin on Anxiety-like Behavior in C57/B6J Mice. Biomedicines 2023, 11, 1705. [Google Scholar] [CrossRef]
- Tanaka, M.; Spekker, E.; Szabó, Á.; Polyák, H.; Vécsei, L. Modelling the neurodevelopmental pathogenesis in neuropsychiatric disorders. Bioactive kynurenines and their analogues as neuroprotective agents-in celebration of 80th birthday of Professor Peter Riederer. J. Neural. Transm. (Vienna) 2022, 129, 627–642. [Google Scholar] [CrossRef]
- Sobolewska-Nowak, J.; Wachowska, K.; Nowak, A.; Orzechowska, A.; Szulc, A.; Płaza, O.; Gałecki, P. Exploring the Heart–Mind Connection: Unraveling the Shared Pathways between Depression and Cardiovascular Diseases. Biomedicines 2023, 11, 1903. [Google Scholar] [CrossRef]
- Tug, E.; Fidan, I.; Bozdayi, G.; Yildirim, F.; Tunccan, O.G.; Lale, Z.; Akdogan, D. The relationship between the clinical course of SARS-CoV-2 infections and ACE2 and TMPRSS2 expression and polymorphisms. Adv. Clin. Exp. Med. 2023, in press. [Google Scholar] [CrossRef] [PubMed]
- Fan, P.; Miranda, O.; Qi, X.; Kofler, J.; Sweet, R.A.; Wang, L. Unveiling the Enigma: Exploring Risk Factors and Mechanisms for Psychotic Symptoms in Alzheimer’s Disease through Electronic Medical Records with Deep Learning Models. Pharmaceuticals 2023, 16, 911. [Google Scholar] [CrossRef]
- Festa, F.; Medori, S.; Macrì, M. Move Your Body, Boost Your Brain: The Positive Impact of Physical Activity on Cognition across All Age Groups. Biomedicines 2023, 11, 1765. [Google Scholar] [CrossRef]
- Alhaddad, A.; Radwan, A.; Mohamed, N.A.; Mehanna, E.T.; Mostafa, Y.M.; El-Sayed, N.M.; Fattah, S.A. Rosiglitazone Mitigates Dexamethasone-Induced Depression in Mice via Modulating Brain Glucose Metabolism and AMPK/mTOR Signaling Pathway. Biomedicines 2023, 11, 860. [Google Scholar] [CrossRef] [PubMed]
- Statsenko, Y.; Habuza, T.; Smetanina, D.; Simiyu, G.L.; Meribout, S.; King, F.C.; Gelovani, J.G.; Das, K.M.; Gorkom, K.N.-V.; Zareba, K.; Almansoori, T.M.; Szolics, M.; Ismail, F.; Ljubisavljevic, M. Unraveling Lifelong Brain Morphometric Dynamics: A Protocol for Systematic Review and Meta-Analysis in Healthy Neurodevelopment and Ageing. Biomedicines 2023, 11, 1999. [Google Scholar] [CrossRef] [PubMed]
- Dang, J.; Tao, Q.; Niu, X.; Zhang, M.; Gao, X.; Yang, Z.; Yu, M.; Wang, W.; Han, S.; Cheng, J.; Zhang, Y. Meta-Analysis of Structural and Functional Brain Abnormalities in Cocaine Addiction. Front. Psychiatry 2022, 13, 927075. [Google Scholar] [CrossRef] [PubMed]
- Okanda Nyatega, C.; Qiang, L.; Jajere Adamu, M.; Bello Kawuwa, H. Altered striatal functional connectivity and structural dysconnectivity in individuals with bipolar disorder: A resting state magnetic resonance imaging study. Front. Psychiatry 2022, 13, 1054380. [Google Scholar] [CrossRef]
- Battaglia, S.; Schmidt, A, Hassel, S.; Tanaka, M. Case Reports in Neuroimaging and Stimulation. Front. Psychiatry (submitted).
- Du, H.; Yang, B.; Wang, H.; Zeng, Y.; Xin, J.; Li, X. The non-linear correlation between the volume of cerebral white matter lesions and incidence of bipolar disorder: A secondary analysis of data from a cross-sectional study. Front. Psychiatry 2023, 14, 1149663. [Google Scholar] [CrossRef]
- Chen, Y.; Yu, R.; DeSouza, J.F.X.; Shen, Y.; Zhang, H.; Zhu, C.; Huang, P.; Wang, C. Differential responses from the left postcentral gyrus, right middle frontal gyrus, and precuneus to meal ingestion in patients with functional dyspepsia. Front. Psychiatry 2023, 14, 1184797. [Google Scholar] [CrossRef]
- Adamu, M.J.; Qiang, L.; Nyatega, C.O.; Younis, A.; Kawuwa, H.B.; Jabire, A.H.; Saminu, S. Unraveling the pathophysiology of schizophrenia: insights from structural magnetic resonance imaging studies. Front. Psychiatry 2023, 14, 1188603. [Google Scholar] [CrossRef]
- Chang, C.H.; Wang, W.L.; Shieh, Y.H.; Peng, H.Y.; Ho, C.S.; Tsai, H.C. Case Report: Low-Frequency Repetitive Transcranial Magnetic Stimulation to Dorsolateral Prefrontal Cortex and Auditory Cortex in a Patient With Tinnitus and Depression. Front. Psychiatry 2022, 13, 847618. [Google Scholar] [CrossRef] [PubMed]
- Zakia, H.; Iskandar, S. Case report: Depressive disorder with peripartum onset camouflages suspected intracranial tuberculoma. Front. Psychiatry 2022, 13, 932635. [Google Scholar] [CrossRef] [PubMed]
- Nyatega, C.O.; Qiang, L.; Adamu, M.J.; Kawuwa, H.B. Gray matter, white matter and cerebrospinal fluid abnormalities in Parkinson’s disease: A voxel-based morphometry study. Front. Psychiatry 2022, 13, 1027907. [Google Scholar] [CrossRef] [PubMed]
- Rymaszewska, J.; Wieczorek, T.; Fila-Witecka, K.; Smarzewska, K.; Weiser, A.; Piotrowski, P.; Tabakow, P. Various neuromodulation methods including Deep Brain Stimulation of the medial forebrain bundle combined with psychopharmacotherapy of treatment-resistant depression-Case report. Front. Psychiatry 2023, 13, 1068054. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Xie, X.; Xie, J.; Tian, S.; Du, X.; Feng, H.; Zhang, H. Early-onset Alzheimer’s disease with depression as the first symptom: a case report with literature review. Front. Psychiatry 2023, 14, 1192562. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.H.; Kim, S.H.; Han, C.; Jeong, H.G.; Lee, M.S.; Kim, J. Antidepressant-induced mania in panic disorder: a single-case study of clinical and functional connectivity characteristics. Front. Psychiatry 2023, 14, 1205126. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Cao, Y.; Deng, G.; Fang, J.; Qiu, C. Transient splenial lesion syndrome in bipolar-II disorder: a case report highlighting reversible brain changes during hypomanic episodes. Front. Psychiatry. 2023, 14, 1219592. [Google Scholar] [CrossRef]
- Veldema, J. Non-Invasive Brain Stimulation and Sex/Polypeptide Hormones in Reciprocal Interactions: A Systematic Review. Biomedicines 2023, 11, 1981. [Google Scholar] [CrossRef]
- Balogh, L.; Tanaka, M.; Török, N.; Vécsei, L.; Taguchi, S. Crosstalk between Existential Phenomenological Psychotherapy and Neurological Sciences in Mood and Anxiety Disorders. Biomedicines 2021, 9, 340. [Google Scholar] [CrossRef]
- Di Gregorio, F.; Battaglia, S. Advances in EEG-based functional connectivity approaches to the study of the central nervous system in health and disease. Adv. Clin. Exp. Med. 2023, 32, 607–612. [Google Scholar] [CrossRef]
- Tanaka, M.; Diano, M.; Battaglia, S. Editorial: Insights into structural and functional organization of the brain: evidence from neuroimaging and non-invasive brain stimulation techniques. Front. Psychiatry 2023, 14, 1225755. [Google Scholar] [CrossRef] [PubMed]
- Hakamata, Y.; Hori, H.; Mizukami, S.; Izawa, S.; Yoshida, F.; Moriguchi, Y.; Hanakawa, T.; Inoue, Y.; Tagaya, H. Blunted diurnal interleukin-6 rhythm is associated with amygdala emotional hyporeactivity and depression: a modulating role of gene-stressor interactions. Front. Psychiatry 2023, 14, 1196235. [Google Scholar] [CrossRef] [PubMed]
- Rassler, B.; Blinowska, K.; Kaminski, M.; Pfurtscheller, G. Analysis of Respiratory Sinus Arrhythmia and Directed Information Flow between Brain and Body Indicate Different Management Strategies of fMRI-Related Anxiety. Biomedicines 2023, 11, 1028. [Google Scholar] [CrossRef] [PubMed]
- Vasiliu, O. Efficacy, Tolerability, and Safety of Toludesvenlafaxine for the Treatment of Major Depressive Disorder—A Narrative Review. Pharmaceuticals 2023, 16, 411. [Google Scholar] [CrossRef] [PubMed]
- Detke, M.J.; Rickels, M.; Lucki, I. Active behaviors in the rat forced swimming test differentially produced by serotonergic and noradrenergic antidepressants. Psychopharmacology (Berl.) 1995, 121, 66–72. [Google Scholar] [CrossRef]
- Khisti, R.T.; Chopde, C.T.; Jain, S.P. Antidepressant-like effect of the neurosteroid 3alpha-hydroxy-5alpha-pregnan-20-one in mice forced swim test. Pharmacol. Biochem. Behav. 2000, 67, 137–43. [Google Scholar] [CrossRef]
- Steru, L.; Chermat, R.; Thierry, B.; Simon, P. The tail suspension test: a new method for screening antidepressants in mice. Psychopharmacology (Berl.) 1985, 85, 367–70. [Google Scholar] [CrossRef]
- Cryan, J.F.; Mombereau, C.; Vassout, A. The tail suspension test as a model for assessing antidepressant activity: review of pharmacological and genetic studies in mice. Neurosci. Biobehav. Rev. 2005, 29, 571–625. [Google Scholar] [CrossRef]
- Lister, R.G. The use of a plus-maze to measure anxiety in the mouse. Psychopharmacology (Berl.) 1987, 92, 180–5. [Google Scholar] [CrossRef]
- Pellow, S.; Chopin, P.; File, SE.; Briley, M. Validation of open: closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J. Neurosci. Meth. 1985, 14, 149–167. [Google Scholar] [CrossRef]
- Misslin, R.; Belzung, C.; Vogel, E. Behavioural validation of a light/dark choice procedure for testing anti-anxiety agents. Behav. Processes 1989, 18, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Costall, B.; Coughlan, J.; Horovitz, Z.P.; Kelly, M.E.; Naylor, R.J.; Tomkins, D.M. The effects of ACE inhibitors captopril and SQ29,852 in rodent tests of cognition. Pharmacol. Biochem. Behav. 1989, 33, 573–9. [Google Scholar] [CrossRef] [PubMed]
- Onaivi, E.S.; Martin, B.R. Neuropharmacological and physiological validation of a computer-controlled two-compartment black and white box for the assessment of anxiety. Prog. Neuropsychopharmacol. Biol. Psychiatry 1989, 13, 963–76. [Google Scholar] [CrossRef] [PubMed]
- Broekkamp, C.L.; Rijk, H.W.; Joly-Gelouin, D.; Lloyd, K.L. Major tranquillizers can be distinguished from minor tranquillizers on the basis of effects on marble burying and swim-induced grooming in mice. Eur. J. Pharmacol. 1986, 126, 223–9. [Google Scholar] [CrossRef] [PubMed]
- van der Poel, A.M. Ethological study of the behaviour of the albino rat in a passive-avoidance test. Acta Physiol. Pharmacol. Neerl. 1967, 14, 503–5. [Google Scholar]
- Hall, C.; Ballachey, E.L. A study of the rat’s behavior in a field: a contribution to method in comparative psychology. University of California Publications in Psychology 1932, 6, 1–12. [Google Scholar]
- Gould, T.D.; Dao, D.T.; Kovacsics, C.E. The Open Field Test. In: Gould, T. Mood and Anxiety Related Phenotypes in Mice. Neuromethods, vol 42. Humana Press, Totowa, NJ, 2009. [CrossRef]
- ScienceDirect. Available online: https://www.sciencedirect.com/topics/psychology/neurotransmitter (accessed on 20 July 2023).
- Schwarz, M.J.; Ackenheil, M. The role of substance P in depression: therapeutic implications. Dialogues Clin. Neurosci. 2002, 4, 21–9. [Google Scholar] [CrossRef]
- Tanaka, M.; Török, N.; Vécsei, L. Are 5-HT1 receptor agonists effective anti-migraine drugs? Expert Opin. Pharmacother. 2021, 22, 1221–1225. [Google Scholar] [CrossRef]
- Kindler, J.; Lim, C.K.; Weickert, C.S.; Boerrigter, D.; Galletly, C.; Liu, D.; Jacobs, K.R.; Balzan, R.; Bruggemann, J.; O’Donnell, M.; Lenroot, R.; Guillemin, G.J.; Weickert, T.W. Dysregulation of kynurenine metabolism is related to proinflammatory cytokines, attention, and prefrontal cortex volume in schizophrenia. Mol. Psychiatry 2020, 25, 2860–2872. [Google Scholar] [CrossRef]
- Han, Q.; Cai, T.; Tagle, D.A.; Li, J. Structure, expression, and function of kynurenine aminotransferases in human and rodent brains. Cell Mol. Life Sci. 2010, 67, 353–68. [Google Scholar] [CrossRef]
- Okada, K.; Angkawidjaja, C.; Koga, Y.; Kanaya, S. Structural and mechanistic insights into the kynurenine aminotransferase-mediated excretion of kynurenic acid. J. Struct. Biol. 2014, 185, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Kucukkarapinar, M.; Yay-Pence, A.; Yildiz, Y.; Buyukkoruk, M.; Yaz-Aydin, G.; Deveci-Bulut, T.S.; Gulbahar, O.; Senol, E.; Candansayar, S. Psychological outcomes of COVID-19 survivors at sixth months after diagnose: the role of kynurenine pathway metabolites in depression, anxiety, and stress. J. Neural. Transm. (Vienna) 2022, 129, 1077–1089. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Török, N.; Vécsei, L. Novel Pharmaceutical Approaches in Dementia. In: Riederer, P., Laux, G., Nagatsu, T., Le, W., Riederer, C. (eds) NeuroPsychopharmacotherapy. Springer, Cham. 2021. [CrossRef]
- Tanaka, M.; Szabó, Á.; Spekker, E.; Polyák, H.; Tóth, F.; Vécsei, L. Mitochondrial Impairment: A Common Motif in Neuropsychiatric Presentation? The Link to the Tryptophan–Kynurenine Metabolic System. Cells 2022, 11, 2607. [Google Scholar] [CrossRef]
- Tanaka, M.; Bohár, Z.; Vécsei, L. Are Kynurenines Accomplices or Principal Villains in Dementia? Maintenance of Kynurenine Metabolism. Molecules 2020, 25, 564. [Google Scholar] [CrossRef]
- Ford, J.D.; Courtois, C.A. Complex PTSD and borderline personality disorder. Borderline Personal Disord. Emot. Dysregul. 2021, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Barsy, B.; Kocsis, K.; Magyar, A.; Babiczky, Á.; Szabó, M.; Veres, J.M.; Hillier, D.; Ulbert, I.; Yizhar, O.; Mátyás, F. Associative and plastic thalamic signaling to the lateral amygdala controls fear behavior. Nat. Neurosci. 2020, 23, 625–637. [Google Scholar] [CrossRef] [PubMed]
- Spielberg, J.M.; Stewart, J.L.; Levin, R.L.; Miller, G.A.; Heller, W. Prefrontal Cortex, Emotion, and Approach/Withdrawal Motivation. Soc. Personal. Psychol. Compass 2008, 2, 135–153. [Google Scholar] [CrossRef] [PubMed]
- Frankland, P.W.; Josselyn, S.A.; Köhler, S. The neurobiological foundation of memory retrieval. Nat. Neurosci. 2019, 22, 1576–1585. [Google Scholar] [CrossRef]
- Grace, A.A. Dysregulation of the dopamine system in the pathophysiology of schizophrenia and depression. Nat. Rev. Neurosci. 2016, 17, 524–32. [Google Scholar] [CrossRef]
- TenHouten, W. The Emotions of Hope: From Optimism to Sanguinity, from Pessimism to Despair. Am. Soc. 2023, 54, 76–100. [Google Scholar] [CrossRef]
- Westerhof, G.J.; Bohlmeijer, E.T.; McAdams, D.P. The Relation of Ego Integrity and Despair to Personality Traits and Mental Health. J. Gerontol. B. Psychol. Sci. Soc. Sci. 2017, 72, 400–407. [Google Scholar] [CrossRef]
- Jing, L.; Duan, T.T.; Tian, M.; Yuan, Q.; Tan, J.W.; Zhu, Y.Y.; Ding, Z.Y.; Cao, J.; Yang, Y.X.; Zhang, X.; Mao, R.R.; Richter-Levin, G.; Zhou, Q.X.; Xu, L. Despair-associated memory requires a slow-onset CA1 long-term potentiation with unique underlying mechanisms. Sci. Rep. 2015, 5, 15000. [Google Scholar] [CrossRef] [PubMed]
- Kwong, J. Despair and Hopelessness. Journal of the American Philosophical Association 2023, 1–18. [Google Scholar] [CrossRef]
- van Meerkerk-Aanen, P.J.; de Vroege, L.; Khasho, D.; Foruz, A.; van Asseldonk, J.T.; van der Feltz-Cornelis, C.M. La belle indifférence revisited: a case report on progressive supranuclear palsy misdiagnosed as conversion disorder. Neuropsychiatr. Dis. Treat. 2017, 13, 2057–2067. [Google Scholar] [CrossRef] [PubMed]
- Cologne, Germany: Institute for Quality and Efficiency in Health Care (IQWiG); 2006-. Depression: How effective are antidepressants? [Updated 2020 Jun 18]. Available online: https://www.ncbi.nlm.nih.gov/books/NBK361016/ (accessed on 20 July 2023).
- Raison, S.; Weissmann, D.; Rousset, C.; Pujol, J.F.; Descarries, L. Changes in steady-state levels of tryptophan hydroxylase protein in adult rat brain after neonatal 6-hydroxydopamine lesion. Neuroscience 1995, 67, 463–75. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, J.P.; Medvedev, I.O.; Caron, M.G. The 5-HT deficiency theory of depression: perspectives from a naturalistic 5-HT deficiency model, the tryptophan hydroxylase 2Arg439His knockin mouse. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2012, 367, 2444–59. [Google Scholar] [CrossRef]
- Xu, C.J.; Wang, J.L.; Jing-Pan, Min-Liao. Tph2 Genetic Ablation Contributes to Senile Plaque Load and Astrogliosis in APP/PS1 Mice. Curr. Alzheimer Res. 2019, 16, 219–232. [Google Scholar] [CrossRef]
- Angoa-Pérez, M.; Kane, M.J.; Briggs, D.I.; Herrera-Mundo, N.; Sykes, C.E.; Francescutti, D.M.; Kuhn, D.M. Mice genetically depleted of brain serotonin do not display a depression-like behavioral phenotype. ACS Chem. Neurosci. 2014, 5, 908–19. [Google Scholar] [CrossRef]
- Sbrini, G.; Hanswijk, S.I.; Brivio, P.; Middelman, A.; Bader, M.; Fumagalli, F.; Alenina, N.; Homberg, J.R.; Calabrese, F. Peripheral Serotonin Deficiency Affects Anxiety-like Behavior and the Molecular Response to an Acute Challenge in Rats. Int. J. Mol. Sci. 2022, 23, 4941. [Google Scholar] [CrossRef] [PubMed]
- Klemenhagen, K.C.; Gordon, J.A.; David, D.J.; Hen, R.; Gross, C.T. Increased fear response to contextual cues in mice lacking the 5-HT1A receptor. Neuropsychopharmacology 2006, 31, 101–11. [Google Scholar] [CrossRef]
- Bouchekioua, Y.; Nebuka, M.; Sasamori, H.; Nishitani, N.; Sugiura, C.; Sato, M.; Yoshioka, M.; Ohmura, Y. Serotonin 5-HT2C receptor knockout in mice attenuates fear responses in contextual or cued but not compound context-cue fear conditioning. Trans. Psychiatry 2022, 12, 58. [Google Scholar] [CrossRef] [PubMed]
- Wellman, C.L.; Izquierdo, A.; Garrett, J.E.; Martin, K.P.; Carroll, J.; Millstein, R.; Lesch, K.P.; Murphy, D.L.; Holmes, A. Impaired stress-coping and fear extinction and abnormal corticolimbic morphology in serotonin transporter knock-out mice. J. Neurosci. 2007, 27, 684–91. [Google Scholar] [CrossRef] [PubMed]
- Sorgdrager, F.J.H.; Naudé, P.J.W.; Kema, I.P.; Nollen, E.A.; Deyn, P.P. Tryptophan Metabolism in Inflammaging: From Biomarker to Therapeutic Target. Front. Immunol. 2019, 10, 2565. [Google Scholar] [CrossRef]
- Myint, A.M. Kynurenines: from the perspective of major psychiatric disorders. FEBS J. 2012, 279, 1375–85. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, M.R.; Di Fazio, C.; Battaglia, S. Activated Tryptophan-Kynurenine Metabolic System in the Human Brain is Associated with Learned Fear. Front. Mol. Neurosci. 2023, 16. [Google Scholar] [CrossRef] [PubMed]
- Skorobogatov, K.; Autier, V.; Foiselle, M.; Richard, J.R.; Boukouaci, W.; Wu, C.L.; Raynal, S.; Carbonne, C.; Laukens, K.; Meysman, P. , et al. Kynurenine pathway abnormalities are state-specific but not diagnosis-specific in schizophrenia and bipolar disorder. Brain Behav. Immun. Health 2023, 27, 100584. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Xie, S.; He, Y.; Xu, M.; Qiao, X.; Zhu, Y.; Wu, W. Kynurenine Pathway Metabolites as Biomarkers in Alzheimer’s Disease. Dis. Markers 2022, 2022, 9484217. [Google Scholar] [CrossRef]
- Marszalek-Grabska, M.; Walczak, K.; Gawel, K.; Wicha-Komsta, K.; Wnorowska, S.; Wnorowski, A.; Turski, W.A. Kynurenine emerges from the shadows—Current knowledge on its fate and function. Pharmacol. Ther. 2021, 225, 107845. [Google Scholar] [CrossRef]
- Yu, P.; Di Prospero, N.A.; Sapko, M.T.; Cai, T.; Chen, A.; Melendez-Ferro, M.; Du, F.; Whetsell, W.O. Jr.; Guidetti, P.; Schwarcz, R.; Tagle, D.A. Biochemical and phenotypic abnormalities in kynurenine aminotransferase II-deficient mice. Mol. Cell. Biol. 2004, 24, 6919–30. [Google Scholar] [CrossRef]
- Potter, M.C.; Elmer, G.I.; Bergeron, R.; Albuquerque, E.X.; Guidetti, P.; Wu, H.Q.; Schwarcz, R. Reduction of endogenous kynurenic acid formation enhances extracellular glutamate, hippocampal plasticity, and cognitive behavior. Neuropsychopharmacology 2010, 35, 1734–42. [Google Scholar] [CrossRef]
- Desbonnet, L.; Tighe, O.; Karayiorgou, M.; Gogos, J.A.; Waddington, J.L.; O’Tuathaigh, C.M. Physiological and behavioural responsivity to stress and anxiogenic stimuli in COMT-deficient mice. Behav. Brain Res. 2012, 228, 351–8. [Google Scholar] [CrossRef] [PubMed]
- Kash, S.F.; Tecott, L.H.; Hodge, C.; Baekkeskov, S. Increased anxiety and altered responses to anxiolytics in mice deficient in the 65-kDa isoform of glutamic acid decarboxylase. Proc. Natl. Acad. Sci. U. S. A. 1999, 96, 1698–703. [Google Scholar] [CrossRef] [PubMed]
- Sangha, S.; Narayanan, R.T.; Bergado-Acosta, J.R.; Stork, O.; Seidenbecher, T.; Pape, H.C. Deficiency of the 65 kDa isoform of glutamic acid decarboxylase impairs extinction of cued but not contextual fear memory. J. Neurosci. 2009, 29, 15713–20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.H.; Zhou, J.; Pan, H.Q.; Wang, X.Y.; Liu, W.Z.; Zhang, J.Y.; Yin, X.P.; Pan, B.X. δ Subunit-containing GABAA receptor prevents overgeneralization of fear in adult mice. Learn. Mem. 2017, 24, 381–384. [Google Scholar] [CrossRef] [PubMed]
- Sideris, A.; Piskoun, B.; Russo, L.; Norcini, M.; Blanck, T.; Recio-Pinto, E. Cannabinoid 1 receptor knockout mice display cold allodynia, but enhanced recovery from spared-nerve injury-induced mechanical hypersensitivity. Mol. Pain. 2016, 12, 1744806916649191. [Google Scholar] [CrossRef] [PubMed]
- Clinical SynthesisFull Access Highlights of Changes from DSM-IV to DSM-5: Posttraumatic Stress Disorder American Psychiatric Association Division of Research Published Online:1 Jul 2013. Available online: https://doi.org/10.1176/appi.focus.11.3.358 (accessed on 20 July 2023). [CrossRef]
Figure 1.
Generation of the knockout kat2-/- mice. Female C57BL/6N mice were treated with pregnant mare serum gonadotropin (PMSG) and human chorionic gonadotropin (hCG) with a 48-hour interval between administrations, then mated with male C57BL/6N mice. From the oviducts, fertilized one-cell embryos were collected and injected with single guide RNA (sgRNA) and Guide-itTM Recombinant Cas9 protein. At the two-cell stage, the embryos were transferred into pseudopregnant ICR mice. PMSG: pregnant mare serum gonadotropin; hCG: human chorionic gonadotropin; sgRNA: single guide RNA.
Figure 1.
Generation of the knockout kat2-/- mice. Female C57BL/6N mice were treated with pregnant mare serum gonadotropin (PMSG) and human chorionic gonadotropin (hCG) with a 48-hour interval between administrations, then mated with male C57BL/6N mice. From the oviducts, fertilized one-cell embryos were collected and injected with single guide RNA (sgRNA) and Guide-itTM Recombinant Cas9 protein. At the two-cell stage, the embryos were transferred into pseudopregnant ICR mice. PMSG: pregnant mare serum gonadotropin; hCG: human chorionic gonadotropin; sgRNA: single guide RNA.
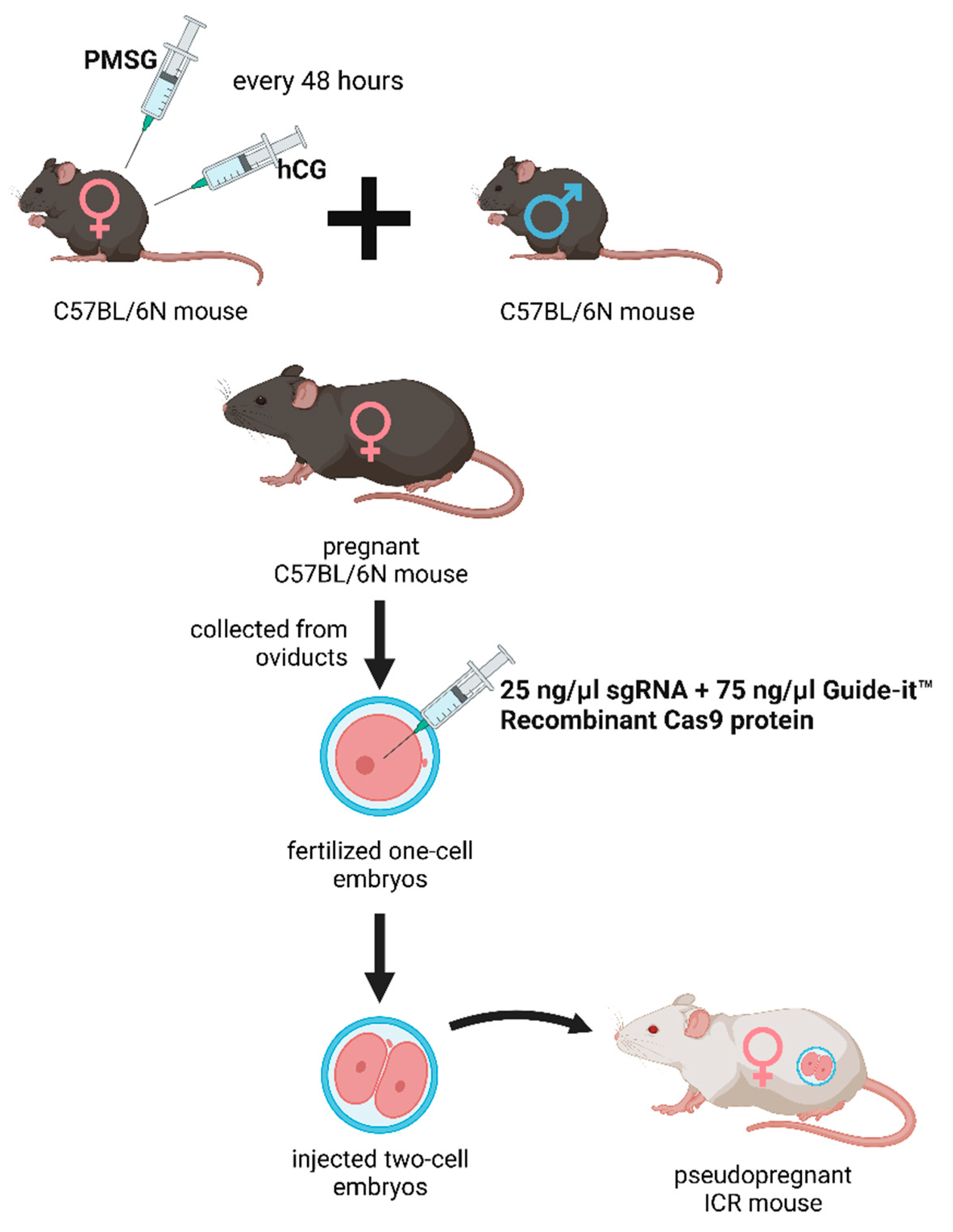
Figure 2.
DNA sequence analysis of knockout kat2-/- mouse line. a) Genomic sequences around the mutation site of knockout kat2-/- mouse line. b) Western blot analysis of knockout kat2-/- mouse line.
Figure 2.
DNA sequence analysis of knockout kat2-/- mouse line. a) Genomic sequences around the mutation site of knockout kat2-/- mouse line. b) Western blot analysis of knockout kat2-/- mouse line.
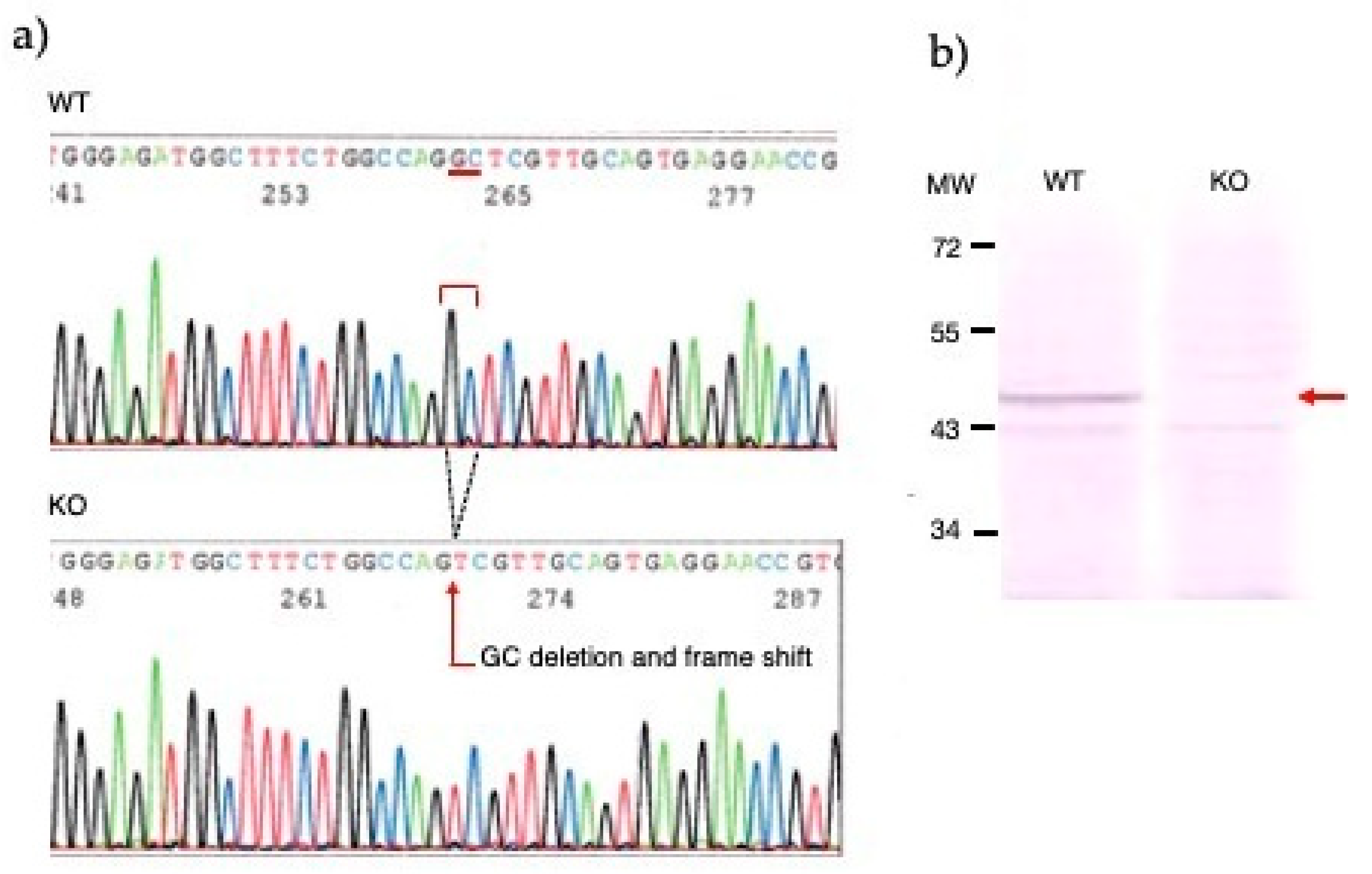
Figure 3.
Behavioral tests. a) Time spent immobile in the modified forced swim test (FST); b) Time spent swimming in the modified FST; c) Number of entries into the center and corner zones in the open-field (OF) test; d) Ambulation distance in the OF test; and e) Number of jumps in the OF test. Wild-type mice (light green); kat2-/- mice (dark green). WT: wild-type; kat2-/-: kynurenine aminotransferase II knockout mice; •: outliner. Mean ± SEM. *: p ≤ 0.05; **: p ≤ 0.01; ***: p ≤ 0.001.
Figure 3.
Behavioral tests. a) Time spent immobile in the modified forced swim test (FST); b) Time spent swimming in the modified FST; c) Number of entries into the center and corner zones in the open-field (OF) test; d) Ambulation distance in the OF test; and e) Number of jumps in the OF test. Wild-type mice (light green); kat2-/- mice (dark green). WT: wild-type; kat2-/-: kynurenine aminotransferase II knockout mice; •: outliner. Mean ± SEM. *: p ≤ 0.05; **: p ≤ 0.01; ***: p ≤ 0.001.
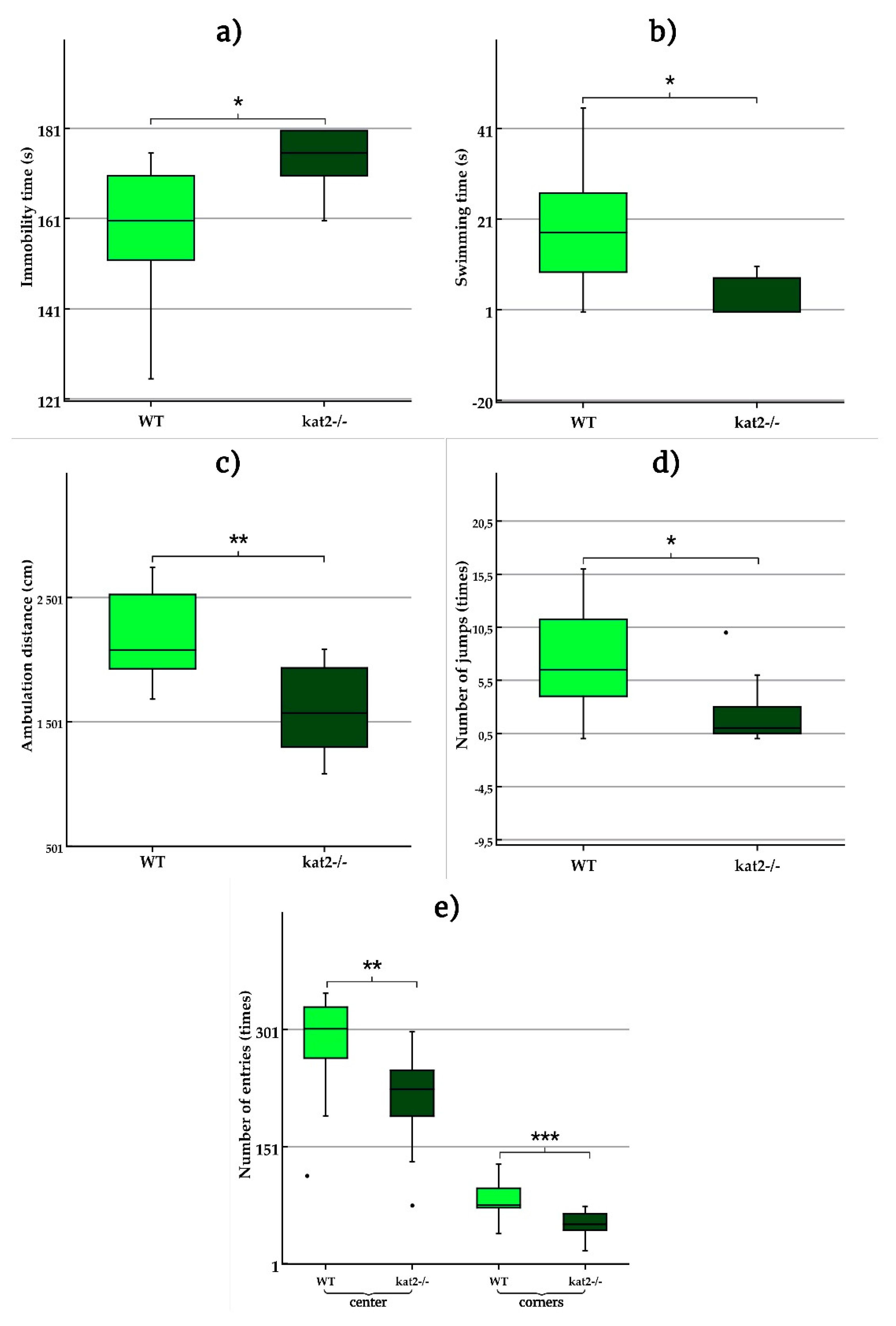

Table 1.
Properties of sgRNA, primers and KAT gene. sgRNA: single guide RNA; KAT: kynurenine aminotransferase; KAT II: aminoadipate aminotransferase; CCDS: Consensus Coding Sequence.
Table 1.
Properties of sgRNA, primers and KAT gene. sgRNA: single guide RNA; KAT: kynurenine aminotransferase; KAT II: aminoadipate aminotransferase; CCDS: Consensus Coding Sequence.
| Name of sgRNA | Sequence | ||
|---|---|---|---|
| M-KAT II-2 | GTTCCTCACTGCAACGAGCCguuuuagagcuagaaauagcaaguuaaaaaaggcuaguccguuaucaacuugaaaaaguggcacggacucggugcuuuu | ||
| Name of primer | Sequence | ||
| M-KAT II_1st_F | CCCTCTGTGGATGGACTTTG | ||
| M-KAT II_1st _R | TTGAAAGATGTGCCTCATGC | ||
| M-KAT II_2nd_F | GGATGGACTTTGTCCCTTCT | ||
| M-KAT II_2nd_R | ATGTGCCTCATGCTTGGCCC | ||
| Name of KAT gene | Transcript ID | CCDS | CCDS Nucleotide Sequence |
| Aadat-201 | ENSMUST00000079472.4 | CCDS22320 | 32-33 (2 nucleotide deletion) |
Table 2.
Behaviors of kat2−/− mice and the wild-type counterparts.
| Test type | Perspectives | p-value |
|---|---|---|
| Modified forced swim test (FST) | Immobility time (s) | 0,022 |
| Swimming time (s) | 0,014 | |
| Climbing time (s) | 0,681 | |
| Tail suspension test (TST) | Immobility time (s) | 0,625 |
| Passive avoidance (PA) test | Time spent in the lit box on the training day (s) | 0,979 |
| Time spent in the lit box on the test day (s) | 0,822 | |
| Elevated plus maze (EPM) test | Time spent in the open arms (s) | 0,500 |
| Light-dark box (LDB) test | Time spent in the lit box (s) | 0,957 |
| Marble burying (MB) test | Number of immobile marbles (times) | 0,824 |
| Number of marbles that changed position (times) | 0,568 | |
| Number of 0-25% burried marbles (times) | 0,577 | |
| Number of 25-50% burried marbles (times) | 0,926 | |
| Number of 50-75% burried marbles (times) | 0,949 | |
| Number of 75-100% burried marbles (times) | 0,909 | |
| Open-field test (OFT) | Number of entries to the center zones (times) | 0,011 |
| Number of entries to the corner zones (times) | 0,001 | |
| Ambulation distance (cm) | 0,002 | |
| Number of jumps (times) | 0,034 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



