
Submitted:
28 July 2023
Posted:
01 August 2023
You are already at the latest version
Abstract
Medicinal plants are being tested as options to prevent infectious diseases in many countries. Baccharis crispa Spreng. (Asteraceae), is a native medicinal species widely used throughout South America. Our objective was to evaluate the antiviral activity of four wild populations of B. crispa from the province of Córdoba (Argentina) and determine if there is variability between them. The cytotoxicity of the extracts (chlorofom, ethanol and aquose) was evaluated by the neutral red uptake method. Using the plaque-forming unit reduction assay, virucidal and antiviral activity against Herpes Simplex type 1 and Chikungunya viruses were tested. Phytochemical screening of each extract was also performed. Puesto Pedernera (PP) population showed the best antiviral and virucidal effect, standing out the ethanolic extract as the least toxic and with the best inhibitory activity against both viral models. Villa del Parque (VP) population was inactive, regardless of the extract and the virus tested. The results revealed that extracts bioactivities vary significantly depending on the geographic area of collection, with PP and VP populations showing the most significant variability, not only in activity but also in chemical composition. These findings support the potential use of extracts as a source of therapeutic agents against different viruses.
Keywords:
natural products
; population variability
; bioactivities
; Herpes Simplex virus
; Chikungunya virus
1. Introduction
Infectious viral diseases remain a significant worldwide problem [1]. In recent years, the growing emergence and re-emergence of viral infections represent one of the most worrying aspects of human health [2,3,4]. Within emerging virosis, infections by arthropod-borne viruses, called "arboviruses," are increasingly becoming an endemic medical problem affecting mainly developing countries or upcoming economies [5,6]. An example of a re-emerging arbovirus is the Chikungunya virus (CHIKV). CHIKV infection was first described in Tanzania in 1952 [7], and since then has caused significant epidemics of chikungunya fever. Epidemics crossed borders and seas, and the virus was introduced into at least 19 countries by travelers returning from affected areas. Because of the virus has been introduced into geographic areas where competent vectors are endemic, this disease has the potential to become established in new areas of Europe and the Americas [8,9,10,11]. Notably, in March 2016, the World Health Organization (WHO) notified the first outbreak of chikungunya fever in Argentina, and since then, the arbovirus has been under epidemiological surveillance. So far, no specific treatment or vaccine has been available to prevent infections by this virus [12].
Currently, only about 50 synthetic antivirals have been approved for human use, most of which are drugs for treating the Human Immunodeficiency Virus (HIV) [13]. To the limited number of antivirals available must be added the emergence of resistant strains to these drugs, such as strains of Herpes Simplex virus type 1 (HSV-I) resistant to acyclovir, which represent a serious concern, especially in immunosuppressed patients [14,15].
This lack of effective therapies has generated a growing search for new effective chemotherapeutic agents for treating these diseases [16]. Medicinal plants have been extremely valued for millennia as a rich source of therapeutic agents for managing viral disorders. Globally, a search for medicinal plants to treat viral diseases has been initiated, as they are relatively more accessible, inexpensive, and have less potential for toxicity [17]. There are no reports on plant extracts with antiviral activity against CHIKV; however, there are some novel reports on natural compounds with potential activity on this virus [18,19,20]. Baccharis crispa Spreng. (Asteraceae) is commonly known in Argentina as “carqueja” and is popularly used as an antiseptic, antirheumatic, cholagogue, diuretic, and hepatic agent [21,22]. Aqueous and organic extracts of several species of the genus Baccharis have demonstrated anti-herpetic activity [23,24,25,26,27,28,29,30]. Hydroalcoholic extracts from B. crispa have been reported to inhibit the replication of the Vesicular Stomatitis Virus and Poliovirus [31]. These results emphasize the need to continue with this line of research to improve the knowledge and validation of the antiviral properties of this vegetal species.
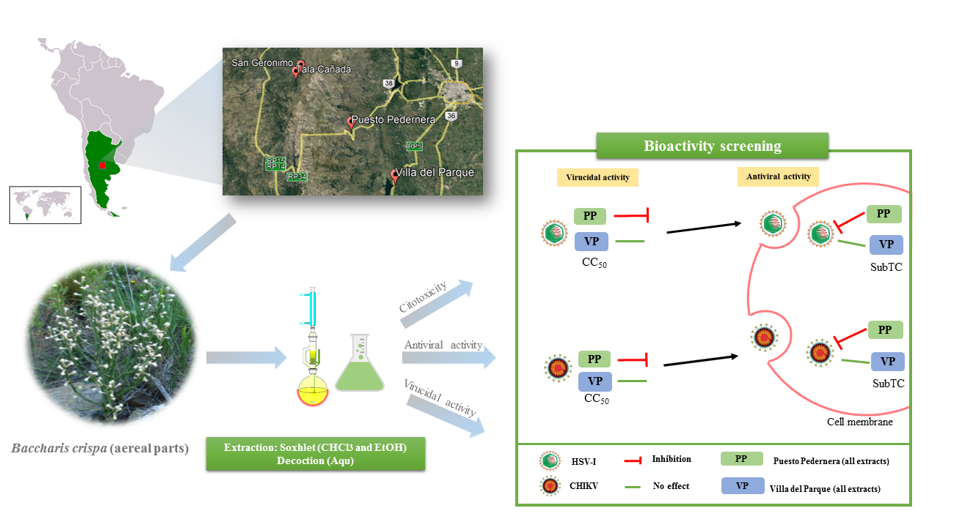
Plants rich in various secondary metabolites, belonging to different chemical classes such as tannins, terpenoids, alkaloids, and polyphenols are generally superior in their antimicrobial activities [32]. This suggests that the strength of the biological activities of a natural product depends on the diversity and quantity of such constituents [33]. It is also known that the content of these active principles in plants depend on the environmental factors that prevail in the place where the plants have developed [34,35] and on the adaptive response of different genotypes to their environment. Hence, in this work, extracts obtained from four wild populations of B. crispa from the province of Córdoba (Argentina) were evaluated in terms of variability in their in vitro bioactivities (cytotoxic and antiviral activity) between populations, especially their action against Herpes Simplex type 1 and Chikungunya viruses, assessing also their general chemical composition (groups of major secondary metabolites).
2. Materials and Methods
2.1. Plant material
Vegetal material (aerial parts) of Baccharis crispa Spreng. (Asteraceae) was collected in 2014 from four wild populations that grow in different areas of the natural distribution zone of this species in the province of Córdoba-Argentina: Villa del Parque, VP (31º49’14,7’’S, 64º30’17,8’’W, 827 m above sea level); Puesto Pedernera, PP (31º36’21,2’’S, 64º41’14,5’’W, 1515 m above sea level); Tala Cañada, TC (31º21’57,4’’S, 64º57’11,5’’W, 1342 m above sea level) and San Gerónimo, SG (31º20’02,3’’S, 64º55’50,7’’W, 1486 m above sea level). A voucher specimen of each population was herbarized and deposited in the ACOR herbarium (Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba). The specimens were incorporated into the Lucas M. Carbone (LMC) collection. The plant material was air-dried at room temperature, powdered, and kept in light amber containers until processing.
2.2. Preparation of plant extracts
2.2.1. Organic Extracts
Forty grams of dried aerial parts from each wild population were initially subjected to maceration with hexane to extract fats and chlorophylls. Subsequently, they were extracted in a Soxhlet extractor with chloroform (CHCl3) and ethanol (EtOH). The plant material was in contact with each solvent until depletion. Each Extract was concentrated to dryness in a rotary evaporator (RV1 rotary evaporator, Figmay, Argentina) at moderate temperatures (30-40 °C). All organic solvents were previously distilled before use.
2.2.2. Aqueous Extract
Another fraction of dried and ground aerial parts (40 g) was extracted with enough water to cover the plant material at boiling temperature for one hour (decoction). Each aqueous Extract was lyophilized (Labconco LYPH LOCK 6, Lyophilizer, USA).
2.3. Phytochemical screening
Each dried extracts (2 mg) of of each wild population was dissolved with EtOH-H2O (1:1). Then, the alcoholic content was removed by evaporation at reduced pressure, and subsequently, a series of partitions were made to separate the chemical substances according to their polarity. At different stages of this analytical process, simple coloration reactions were carried out to characterize different families of chemical compounds [36]. The presence or absence of these secondary metabolites was determined by observing the development or not of a characteristic coloration indicating a positive or negative result for each metabolite investigated [37].
2.4. Cells and viruses
The host cells, African green monkey kidney cells (Cercophitecus aethiops, Vero cl 76 obtained from Laboratory of Cell Culture- Instituto de Virología “Dr. J. M. Vanella”, Argentina). Cells were grown and maintained in a humid atmosphere at 37 °C with 5% CO2. The growth medium (GM) consists of Eagle’s minimal essential medium (E-MEM, Gibco, USA) supplemented with 10% (v/v) Fetal Bovine Serum (FBS, Natocor, Argentina), L-glutamine (30 μg/mL, Sigma-Aldrich, USA), and gentamicin (50 μg/mL, Sigma-Aldrich); whereas the maintenance medium (MM) has the same formulation, but with 2% of FBS.
Herpes Simplex Virus Type-I (HSV-I) Kos strain (DNA virus) and Chikungunya virus (CHIKV) Asian-American genotype (RNA virus) were used. Virus stocks were propagated and titrated according to the methods described by [38]. The viral titration was expressed in plaque-forming units per milliliter (PFU/mL), which were 2 x 107 PFU/mL for HSV-I and 3 x 108 PFU/mL for CHIKV.
2.5. Sample solutions
Each extract was dissolved in dimethyl sulfoxide (DMSO) to obtain the stock solution (100 mg/mL), from which dilutions in MM were prepared to carry out the cytotoxicity and the antiviral activity assays. It is important to explain that although the DMSO has demonstrated an antiviral effect on HSV-I and cytotoxicity in different cell types [39], the final concentration used in our assays was always below 2%, a concentration that has no cytotoxic or antiviral effect [40,41,42].
2.6. In vitro Cytotoxicity assay
This assay determines the sample concentrations that do not affect the viability of the host cells to ensure that the observed cytopathic effect is due to the action of the virus and not to the toxicity of the extract. Employing the neutral red (NR) uptake assay, the cell viability (CV) was measured, following a procedure previously described by other authors with some minor modifications [43,44]. A confluent monolayer of Vero cells (1 x 105 cell/well ≈ 95% confluence), grown in 96-well culture plates, was exposed to 15 decreasing concentrations starting at 1 mg/ml of each extract, preparing three replicates of each concentration and including 3 wells of cells with MM as cell control (CC). After incubation for 72 h (37 °C in 5% CO2), the absorbance of the NR extracted from inside the cells was measured at 540 nm by using a microplate reader (BioTek ELx800). The percentage of CV (% CV) was calculated by comparing it with CC (100% CV). By using the Origin 8.6 software, % CV vs concentrations of each extract were plotted with non-linear regression analysis (Sigmoidal curve, R2>0.9).
From these graphs, the following toxicity parameters were estimated for each extract: the concentration that reduced the viable cells to 50% (CC50), and a subtoxic concentration (SubTC) defined as the concentration that causes 20% of cellular death and produces slight morphological changes observed by microscopy [45]. Obtaining these values allowed us to use non-toxic or sub-toxic concentrations in the antiviral activity assays.
2.7. Plaque reduction assay
2.7.1. Antiviral assays
The viral suspension (100 PFU/well) was inoculated into a confluent monolayer of Vero cells (4.8 x 107 cells/well) grown in 24-well culture plates. After 1 h of adsorption at 37 ºC, the residual inoculum was discarded, and the cells were washed with phosphate-buffered saline (PBS). Subsequently, the cells were covered with a semi-solid medium (1% agarose in a double concentration of E-MEM) and different extract concentrations (consecutive 2-fold serial dilutions from SubTC). Three replicates per concentration of each extract were assayed. Infected cells without extract, as viral control (VC), uninfected cells without extracts, as cellular control (CC), and positive control were also included in triplicate. The PFU were counted after 3-5 days of incubation at 37 °C with 5% CO2, according to the viral model, following the methodology described by [46]. Results were expressed as a percentage of inhibition (%I) in correlation with VC [47], and %I is plotted according to the concentration of the treatment to calculate the effective concentration 50 (EC50). Inhibition percentages below 50% were considered extracts without activity [48].
The selectivity index (SI) indicates how suitable a sample or extract is for decreasing viral replication without damaging host cells. SI was measured as the ratio between the concentration that produces toxicity in 50% of the host cells (CC50) and the concentration that inhibits viral replication by 50% (EC50).
2.7.2. Viral inactivation activity
The virucidal effect of each extract (capacity to inactivate the viral particles before they enter the host cell) was determined, using the PFU reduction test in Vero cells. For this, 100 μL of the viral stock (see subsection 2.4) was mixed with 100 μL of a concentration of each extract corresponding to its CC50 (1:1). Viruses in MM without extract were used as viral control (VC), the CC50 of each extract in MM was included as extract control and positive control was also tested. These mixtures were incubated for 1 h at 37 °C. Subsequently, serial dilutions (factor 10) were made from each mixture, and each dilution was added in duplicate to a confluent layer of Vero cells (4.8 x 107 cells/well). After 1 h of incubation (37 °C in a humid atmosphere with 5% CO2), the monolayer was washed with PBS and the cells were covered with a semi-solid medium (1% agarose in a double concentration of E-MEM). The PFU were counted after 3-5 days of incubation (37 °C with 5% CO2). The residual viral infectivity was determined by the PFU reduction assay [46]. Extracts that caused decreases ≤ 2 logarithms compared to the titer of the VC, were considered extracts without virucidal activity [25,49,50].
2.8. Positive controls
Acyclovir (Fada, Argentina) was used as a positive control of HSV-I at a concentration of 15 μM. In contrast, a positive antiviral control against CHIKV was not included because of the lack of effective antiviral drugs for this virus.
2.9. Statistical analysis
The CC50 and SubTC values were obtained from the dose-response curves generated from the non-linear regression analysis. Values were expressed as mean ± standard error ( ± SE) from 3 independent experiments. To evaluate variability in cytotoxicity and antiviral activity between populations, an analysis of variance (ANOVA) was performed. For this analysis, a bifactorial model was adjusted. To determine statistical significance, the posteriori Di Rienzo, Guzmán y Casanoves test (DGC) was applied [51] with a p≤0.05. The statistical software InfoStat [51] was used.
3. Results
3.1. Cytotoxicity assay
From the analysis of the plots % CV vs. concentration of each extract (Supplemental Material, Figure S1), the values of the cytotoxic parameters were estimated. The values of CC50 give an idea of the toxicity of the extracts: high CC50 values imply low toxicity, whereas low values, high toxicity. As shown in Table 1, aqueous extracts (Aqu) were found to be less cytotoxic than organic extracts (CHCl3 and EtOH), regardless of the population evaluated. Among the four populations evaluated, the EtOH extract of VP and the CHCl3 extract of PP proved to be the most cytotoxic, with values close to 100 μg/mL. Considering that some authors point out that CC50 values ≤ 20-30 μg/mL should be considered cytotoxic extracts [27,52,53], all tested extracts were found to be non-cytotoxic.
The interaction between the "extract" and "population" factors was significant (DGC, p≤0.05), which indicates that the factors do not act independently. Significant differences were found in the cytotoxicity between populations by type of extract for CC50 (DGC, p≤0.05). Considering SubTC no significant differences were found in the cytotoxic effect of the aqueous extract of VP and PP populations, as well as the CHCl3 extracts of TC and PP populations.
3.2. Plaque Reduction assay
3.2.1. Antiviral assays
As a cut-off point, those extracts showing an inhibition greater than 50% were selected. According to this criterion, seven extracts showed antiviral activity against CHIKV and five for HSV-I (Table 1). The antiviral activity of the extracts was also dose-dependent (Supplementary material, Figure S2). The inhibition percentage (%I) obtained at the highest concentration evaluated (SubTC) of each extract is shown in Table 1. The population of VP was inactive regardless of the extract and virus evaluated. PP was the wild population of B. crispa with better inhibitory activity on both viruses, being more active against CHIKV than HSV-I (Table 1). Specifically, the organic extracts (CHCl3 and EtOH, p >0.05) showed higher I% than the Aqu extract on CHIKV, and the EtOH extract was the most effective on HSV-I. However, the antiviral activity of the Aqu extract on CHIKV is interesting, considering that it is a preparation usually used by the population (herbal tea). The SG and TC populations showed an intermediate activity on both viruses, between 57 and 67 %I. In general, the EtOH extracts of the PP, TC, and SG populations turned out to be the most active against HSV-I, whereas CHCl3 extracts proved to be the most active against CHIKV (Table 1). These results could probably be related to different chemical profiles in the extracts. Assessment of the variability for antiviral activity determined that there are significant differences between populations for each type of extract tested for both HSV-I and CHIKV (DGC, p≤0.05). In general, SI values were greater than 2, except for Aqu extracts (Table 1).
3.2.2. Viral inactivation assay
Evaluation of the virucidal activity of the extracts against the two viral models showed that all populations had some activity. Only extracts that caused decreases in ≥2 logarithms relative to viral control (VC) were considered to have virucidal activity. As shown in Figure 1, the PP population had the best virucidal capacity with all extracts tested, highlighting the virucidal effect against HSV-I. The EtOH extract was the most active, reducing up to 5 logarithms of the formation of PFU compared to its control on both viruses. VP, TC, and SG showed intermediate and variable activity depending on the extract. Among the extracts of VP, only the EtOH had activity, this being exclusively against HSV-I; in the case of TC, only the Aqu extract was active for both viral models. Whereas for SG only the CHCl3 extract turned out to influence HSV-I, the EtOH and Aqu extracts on CHIKV.
Finally, and considering the variability in the viral inactivation for the active extracts, the results on HSV-I showed that there are significant differences between populations for the EtOH and Aqu extracts (DGC, p≤0.05). In both cases, the PP population stands out as different and with the greatest virucidal activity. In the case of CHIKV, the results obtained were similar, since there are also statistical differences between all populations for each extract evaluated (DGC, p≤0.05). The PP population always highlights, exhibiting the highest inhibition on this virus.
3.3. Phytochemical screening
The analyses indicated the presence of flavonoids, tannins, saponins, phytosterols, and carbohydrates. The population with the highest activity (PP) has these metabolites in different proportions (Table 2). In general, regardless of the polarity of the extracts, flavonoids are the major compounds, followed by tannins and saponins, and to a lesser extent phytosterols and carbohydrates. Populations with medium activities (SG and TC) contain lower amounts of saponins and phytosterols, the latter being absent in the VP population, which is the least active of all, with very low amounts of saponins, present only in the aqueous extract. Other secondary metabolites such as anthraquinones, cardiac glycosides, coumarins, and alkaloids were not detected (Table 2).
4. Discussion
Although there are some reports about the antiviral activity of several extracts from different Baccharis species on a wide variety of viruses (Junín virus, HIV, HSV-I, vesicular stomatitis virus (VSV), Poliovirus (PV) and Equid herpesvirus 1) [23,27,54,55,56,57,58], information on B. crispa is scarce. Thus, the in vitro and in vivo antiherpetic activity of some species of Baccharis [27,58] has already been reported. Until now, only hydroalcoholic extracts from B. crispa have been reported to inhibit the replication of VSV and PV [31,56]. Probably, due to the presence of apigenin (flavonoid, with broad spectrum antiviral activity) with reported antiviral activity against PV in other species of the genus [59,60].
Our work focuses on B. crispa, a species native to Argentina, whose medicinal and industrial use, mainly in the production of bitter beverages, has led to the decline of its wild populations that threatens the conservation of this plant species in some areas of our country. According to the Conservation Priority Index (CPI) for medicinal plants, B. crispa is in third place in the order of importance as a priority for conservation in the Paravachasca and Calamuchita Valley of the Province of Córdoba (Argentina) [61]. In this context, it is important to identify those wild populations with medicinal interest, to prioritize their conservation. Therefore, we have focused on the antiviral activity of 4 wild populations of this plant species, considering the problem of viral infections in the health of mankind.
The results obtained in this work show a significant virucidal and antiviral effect in vitro for all the extracts of the PP population against HSV-1 and CHIKV (Table 1 and Figure 1). In contrast, the rest of the populations showed an intermediate or low antiviral and virucidal effect. The results show that the PP population has a significant virucidal activity, especially on HSV-I reducing up to 5 logarithms, and with a good antiviral activity (51 to 79 %I, depending on extract polarity). Thus, we added another species of the Baccharis genus with in vitro antiherpetic activity and also demonstrated a high inhibitory effect of B. crispa on CHIKV (about 74 to 81 %I, depending on extract polarity), a pathogenic virus with no therapeutic or preventive treatment. These are important results, since they are not only new for this species and contribute to the knowledge of the bioactivities of the genus, but also reinforce previous data obtained about other Baccharis species on HSV-I [27,28,30].
In general, we found that EtOH and CHCl3 extracts from stems of B. crispa had better antiviral activity than Aqu extracts (decoction), regardless of the virus evaluated. It is even observed that EtOH extracts were the most active on HSV-I, whereas CHCl3 extracts exhibited greater action on CHIKV, regardless of the B. crispa population tested, except for the VP population which was inactive for the two viruses assayed.
The SI is a parameter that relates the concentrations of a sample (substance or extract) that generate antiviral effects to those that produce eventual toxicity in host cells. When the SI value is high, the cytotoxicity is low and the effect against the virus is high. As plant extracts are a complex mixture of chemical compounds, where each contributes differently to the biological effect of this matrix, their SI values are usually not high [62]. No value for this index defines whether a sample is a good antiviral agent; however, some authors consider that an SI ≥ 2 in natural products (extracts or compounds) would indicate a potential antiviral agent [63,64]. Considering this SI, it can be concluded that there is variability in the bioactivity among the wild populations evaluated, which is more accentuated between VP and PP populations (Table 1). The PP population was the most active, both in its antiviral and virucidal activity on both viruses, highlighting the EtOH extract as less toxic and with good inhibitory activity against both viral models. Then, with an intermediate activity is SG, followed by TC. The VP population, on the other hand, is discarded as it was inactive regardless of the extract and virus tested.
These differences were also found in the screening of their main groups of phytoconstituents, being the population with the best activity (PP) the most chemically diverse. We infer that the differences found in biological activities (cytotoxicity and antiviral) among wild populations of B. crispa could be related to the variations in the chemical profiles obtained. The pattern of variation found may also reflect selective pressures in different ecological and geographical environments (ecotypes) [65,66,67,68,69,70]. Simões-Pires et al., [71] identified caffeoylquinic acid derivatives (quinic acid with various esterification with caffeic acid) as the major compounds in Aqu extracts of B. crispa, which exhibit recognized choleretic effects. On the other hand, in organic extracts, derivatives of flavonoids and neoclerodane diterpenes were the main isolated compounds [72,73,74]. Particularly, in EtOH extracts of aerial parts from B. crispa, flavonoids such as genkwanin and apigenin were found [72]. These flavonoids have shown in vitro antiviral activity against CHIKV, HSV-I, Hepatitis B virus, African Swine fever virus, Buffalopox virus, and Enterovirus-71 [18,75,76,77,78]. Therefore, these compounds could be involved in the antiviral activity observed in the organic extracts tested in this work.
The results obtained here motivate us to continue with a bio-guided fractionation as a next step, using the organic extracts of the PP population that proved to be the most active and with the highest SI (for both viruses), to determine if there is a one or several biologically active principle, or if it is the whole matrix of the extract that produces the observed activity. Continuing a posteriori with the study of the mechanism of viral inhibition against both viruses.
The variability observed among wild populations highlights the importance of considering the origin of the plant material used to carry out bioactivity evaluations. Furthermore, the study of different populations of a vegetal species allows choosing the population that has bioactivity for its future domestication. In another work of our group, it was found that different populations of the species Tagetes minuta L. (Asteraceae) have different chemical compositions and different antiviral activities [79]. Sartor et al., [80] agree that modifications in environmental conditions (biotic and abiotic), where different individuals of the same species grow, produce modifications in the chemical composition of the plant.
In summary, this study lays the basis for continuing the exhaustive study on the antiviral properties of the species B. crispa, deepens the background of bioactivity already reported on the genus and species, providing unpublished data on its antiviral activity, being the first record about bioactivities of wild populations of B. crispa from the province of Córdoba (Argentina). Besides, the results obtained contribute to the knowledge of the species to promote its conservation and sustainable use.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Experimental details relating to this paper are available online, alongside Figure S1 and Figure S2.
Author Contributions
Conceptualization, B.K. and S.Ñ.M.; methodology, G.L., B.K., and J.J.A.; validation, G.L.; formal analysis, G.L., and J.J.A.; resources, A.G.C, J.J.A, F.M, L.T., and E.L.; writing—original draft preparation, G.L., and B.K.; writing—review and editing, A.G.C., F.M., L.T., M.C., S.Ñ.M, and B.K.; supervision, A.G.C., and B.K.; project administration, B.K., and S.Ñ.M.; funding acquisition, S.Ñ.M, and B.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by SECyT-UNC (Type FORMAR s/Resol. 2020-233-E-UNC-SECYT#ACTIP and CONSOLIDAR N° 411/18; 203/14).
Acknowledgments
G. Lingua is a doctoral fellow of CONICET. B. S. Konigheim and S.C.Nuñez Montoya are members of the Research Career of CONICET.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Ben Sassi, A.B.; Harzallah-Skhiri, F.; Bourgougnon, N.; Aouni, M. Antiviral activity of some Tunisian medicinal plants against Herpes simplex virus type 1. Nat. Prod. Res 2008, 22, 53–65. [Google Scholar] [CrossRef]
- Hui, E.K. Reasons for the increase in emerging and re-emerging viral infectious diseases. Microbes Infect 2006, 8, 905–916. [Google Scholar] [CrossRef]
- Parvez, M.K.; Parveen, S. Evolution and Emergence of Pathogenic Viruses: Past, Present, and Future. Intervirology 2017, 60, 1–7. [Google Scholar] [CrossRef]
- WHO, 2018. Managing epidemics: key facts about major deadly diseases. World Health Organization, Geneva.
- Sautto, G.; Mancini, N.; Gorini, G.; Clementi, M.; Burioni, R. Possible future monoclonal antibody (mAb)-based therapy against arbovirus infections. Biomed Res Int. 2013, 2013, 838491. [Google Scholar] [CrossRef]
- Ketkar, H.; Herman, D.; Wang, P. Genetic Determinants of the Re-Emergence of Arboviral Diseases. Viruses 2019, 11, 150. [Google Scholar] [CrossRef]
- Ross, R.W. The Newala epidemic: III. The virus: isolation, pathogenic properties and relationship to the epidemic. J. Hyg (Lond) 1956, 54, 177–191. [Google Scholar] [CrossRef]
- Arankalle, V.A.; Shrivastava, S.; Cherian, S.; Gunjikar, R.S.; Walimbe, A.M.; Jadhav, S.M.; Sudeep, A.B.; Mishra, A.C. Genetic divergence of Chikungunya viruses in India (1963–2006) with special reference to the 2005–2006 explosive epidemic. J. Gen. Virol 2007, 88, 1967–1976. [Google Scholar] [CrossRef]
- Hochedez, P.; Hausfater, P.; Jaureguiberry, S.; Gay, F.; Datry, A.; Danis, M.; Bricaire, F.; Bossi, P. Cases of chikungunya fever imported from the islands of the South West Indian Ocean to Paris, France. Euro. Surveill 2007, 12, 13–14. [Google Scholar] [CrossRef]
- Alfaro-Toloza, P.; Clouet-Huerta, D.E.; Rodríguez-Morales, A.J. Chikungunya, the emerging migratory rheumatism. Lancet Infect. Dis. 2015, 15, 510–512. [Google Scholar] [CrossRef]
- Rodríguez-Morales, A.J.; Paniz-Mondolfi, A.E. Venezuela: far from the path to dengue and chikungunya control. J. Clin. Virol 2015, 66, 60–61. [Google Scholar] [CrossRef]
- Kantor, I.N. Dengue, Zika, Chikungunya y El Desarrollo De Vacunas. Medicina 2018, 78, 23–28. [Google Scholar] [PubMed]
- Antonelli, G.; Turriziani, O. Antiviral therapy: old and current Issues. Int. J. Antimicrob. Agents 2012, 40, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Chilukuri, S.; Rosen, T. Management of acyclovir-resistant herpes simplex virus. Dermatol. Clin. 2003, 21, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Chen, W.; Liang, H.; Wang, Z.; Chen, J.; Hong, H.; Xie, L.; Nie, H.; Xiong, S. Preparation of a monoPEGylated derivative of cyanovirin-N and its virucidal effect on acyclovir-resistant strains of herpes simplex virus type 1. Arch. Virol. 2019, 164, 1259–1269. [Google Scholar] [CrossRef]
- Kitazato, K.; Wang, Y.; Kobayashi, N. Viral infectious disease and natural products with antiviral activity. Drug Discov. Ther. 2007, 1, 14–22. [Google Scholar] [CrossRef]
- Akram, M.; Tahir, I.M.; Shah, S.M.A.; Mahmood, Z.; Altaf, A.; Ahmad, K.; Munir, N.; Daniyal, M.; Nasir, S.; Mehboob, H. Antiviral potential of medicinal plants against HIV, HSV, influenza, hepatitis, and coxsackievirus: A systematic review. Phytother. Res. 2018, 32, 811–822. [Google Scholar] [CrossRef]
- Murali, K.S.; Sivasubramanian, S.; Vincent, S.; Murugan., S.B.; Giridaran., B.; Dinesh., S.; Gunasekaran., P.; Krishnasamy, K.; Sathishkumar, R. Anti—chikungunya activity of luteolin and apigenin rich fraction from Cynodon dactylon. Asian Pac. J. Trop. Med. 2015, 8, 352–358. [Google Scholar] [CrossRef]
- Troost, B.; Mulder, L.M.; Diosa-Toro, M.; Van de Pol, D.; Rodenhuis-Zybert, I.A.; Smit, J.M. Tomatidine, a natural steroidal alkaloid shows antiviral activity towards chikungunya virus in vitro. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Patil, P.; Agrawal, M.; Almelkar, S.; Jeengar, M.K.; More, A.; Alagarasu, K.; Kumar, N.V.; Mainkar, P.S.; Parashar, D.; Cherian, S. In vitro and in vivo studies reveal α-Mangostin, a xanthonoid from Garcinia mangostana, as a promising natural antiviral compound against chikungunya virus. Virol. J. 2018, 18, 1–12. [Google Scholar] [CrossRef]
- Del Vitto, L.A.; Petenatti, E.M.; Petenatti, M.E. Recursos Herbolarios de San Luis (República Argentina) Primera Parte: Plantas Nativas. Multequina 1997, 6, 49–66. [Google Scholar]
- Abad, M.J.; Bermejo, P. Baccharis (Compositae): a review update. Arkivoc 2007, 7, 76–96. [Google Scholar] [CrossRef]
- Abad, M.J.; Bermejo, P.; Gonzales, E.; Iglesias, I.; Irurzun, A.; Carrasco, L. Antiviral activity of Bolivian plant extracts. Gen. Pharmacol 1999, 32, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Abad, M.J.; Bermejo, P.; Sanchez Palomino, S.; Chiriboga, X.; Carrasco, L. Antiviral activity of some South American Medicinal Plants. Phytother. Res 1999, 13, 142–146. [Google Scholar] [CrossRef]
- Zanon, S.M.; Ceriatti, F.S.; Rovera, M.; Sabini, L.J.; Ramos, B.A. Search for Antiviral activity of Certain Medicinal Plants from Córdoba, Argentina. Rev. Latinoam. Microbiol 1999, 41, 59–62. [Google Scholar] [PubMed]
- Montanha, J.A; Mentel, R.; Reiss, C.; Lindequist, U. Phytochemical screening and antiviral activity of some medicinal plants from the island Soqotra. Phytother. Res. 2006, 20, 298–302. [Google Scholar] [CrossRef]
- Torres, C.V.; Domínguez, M.J.; Carbonari, J.L.; Sabini, M.C.; Sabini, L.I.; Zanon, S.M. Study of Antiviral and Virucidal Activities of Aqueous Extract of Baccharis articulata against Herpes suis virus. Nat. Prod. Commun. 2011, 6, 994–994. [Google Scholar] [CrossRef]
- Visintini Jaime, M.F. Actividad antiviral de plantas medicinales argentinas de la familia Asteraceae. Identificación de compuestos bioactivos y caracterización del mecanismo de acción. Doctoral thesis. Universidad de Buenos Aires. Buenos Aires, Argentina. 2014.
- Simoni, I.C.; Aguiar, B.; Martineli de Araujo Navarro, A.; Martins Parreira, R.; Bittencourt Fernandes, M.J.; Frankland Sawaya, A.C.H.; Fávero, O.A. In vitro antiviral activity of propolis and Baccharis sp. extracts on animal herpesviruses. Arq. Inst. Biol. 2018, 85, 1–7. [Google Scholar] [CrossRef]
- Venturi, C.R.; De Loreto Bordignon, S.A.; Roehe, P.M.; Montanha, J.A.; Cibulski, S.P.; Gosman, G. Chemical analysis and antiviral activity evaluation of Baccharis anómala. Nat. Prod. Res. 2018, 32, 1960–1962. [Google Scholar] [CrossRef]
- Barboza, G.; Cantero, J.J.; Nuñez, C.; Ariza Espinar, L. Flora Medicinal de la provincia de Córdoba (Argentina). Pteridófitas y Antófitas silvestres o naturalizadas. Museo Botánico de Córdoba. Córdoba. 2006; pp: 323-329.
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbial. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef]
- Bouzada, M.L.; Fabri, R.L.; Nogueira, M.; Konno, T.U.; Duarte, G.G.; Scio, E. Antibacterial, cytotoxic and phytochemical screening of some traditional medicinal plants in Brazil. Pharm. Biol. 2009, 47, 44–52. [Google Scholar] [CrossRef]
- Ratera, E.L.; Ratera, M.O. Plantas de la flora argentina empleadas en medicina popular. Hemisferio Sur, Buenos Aires, Argentina. 1980.
- Del Vitto, L.A.; Petenatti, E.M.; Petenatti, M.E. Introducción a la Herboristería. Ser. Tec. Herbario UNSL 2002, 14, 10–61. [Google Scholar]
- Harborne, J.B. Phytochemical Methods – A Guide to Modern Techniques of Plant Analysis, 2nd ed; Chapman and Hall, London. 1984.
- De Bessa, N.G.F. , Borges, J.C.M., Beserra, F. P., Carvalho, R H.A., Pereira, M.A.B., Fagundes, R., Alves, A. Prospecção fitoquímica preliminar de plantas nativas do cerrado de uso popular medicinal pela comunidade rural do assentamento vale verde-Tocantins. Revista Brasileira de Plantas Medicinais 2013, 15, 692–707. [Google Scholar] [CrossRef]
- Contigiani, M.; Sabattini, M. 1977. Virulencia diferencial de cepas de virus Junín por marcadores biológicos en ratones y cobayos. Medicina 1977, 37 (Suppl. 3), 244–251. [Google Scholar]
- Aguilar, J.S.; Roy, D.; Ghazal, P.; Wagner, E.K. Dimethyl sulfoxide blocks herpes simplex virus-1 productive infection in vitro acting at different stages with positive cooperativity. Application of micro-array analysis. BMC Infec. Dis. 2002, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Notman, R.; Noro, M.; O’Malley, B.; Anwar, J. Molecular basis for dimethylsulfoxide (DMSO) action on lipid membranes. J. Am. Chem. Soc. 2006, 128, 13982–13983. [Google Scholar] [CrossRef]
- De Ménorval, M.A.; Mir, L.M.; Fernández, M.L.; Reigada, R. Effects of dimethyl sulfoxide in cholesterol-containing lipid membranes: a comparative study of experiments in silico and with cells. PLoS One 2010, 7, e41733. [Google Scholar] [CrossRef]
- Galvao, J.; Davis, B.; Tilley, M.; Normando, E.; Duchen, M.R.; Cordeiro, M.F. Unexpected low-dose toxicity of the universal solvent DMSO. FASEB J. 2014, 28, 1317–1330. [Google Scholar] [CrossRef]
- Borenfreund, E.; Puerner, J.A. Toxicity determined in vitro by morphological alterations and neutral red absorption. Toxicol. Lett. 1985, 24, 119–124. [Google Scholar] [CrossRef]
- Ooi, L.S.; Wang, H.; Luk, C.W.; Ooi, V.E. Anticancer and antiviral activities of Youngia japonica (L.) DC (Asteraceae, Compositae). J. Ethnopharmacol. 2004, 94, 117–122. [Google Scholar] [CrossRef]
- Cholewa, M.; Legge, G.J.F.; Weigold, H.; Holan, G.; Birch, C.J. The use of a scanning proton microprobe to observe anti-HIV drugs within cells. Life Sci. 1994, 54, 1607–1612. [Google Scholar] [CrossRef]
- Cheng, H.Y.; Lin, L.T.; Huang, H.H.; Yang, C.M.; Lin, C.C. Yin Chen Hao Tang, a Chinese prescription, inhibits both herpes simplex virus type-1 and type-2 infections in vitro. Antiviral Res. 2008, 77, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Gescher, K.; Kühn, J.; Hafezi, W.; Louis, A.; Derksen, A.; Deters, A.; Lorentzen, E.; Hensel, A. Inhibition of viral adsorption and penetration by an aqueous extract from Rhododendron ferrugineum L. as antiviral principle against herpes simplex virus type-1. Fitoterapia. 2011, 82, 408–413. [Google Scholar] [CrossRef]
- Konigheim, B.S. Prospección de productos naturales con potencial actividad antiviral obtenidos a partir de especies nativas de género Larrea. Doctoral thesis. Universidad Nacional de Córdoba. Córdoba, Argentina. 2012.
- Andrei, G.M.; Coto, C.E.; De Torres, R.A. Ensayos de citotoxicidad y actividad antiviral de extractos crudos y semipurificados de hojas verdes de Melia azedarach L. Rev. Argent. Microbiol. 1985, 17, 187–194. [Google Scholar] [PubMed]
- Vanden Berghe, D.A.; Vlietinck, A.J.; Van Hoof, L. Plant products as potential antiviral agents. Bull lnst. Pasteur. 1986, 84, 101–147. [Google Scholar] [CrossRef]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. Centro de Transferencia InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. 2017.
- Geran, R.I.; Greenberg, N.H.; McDonald, M.M.; Schumacher, A.M.; Abbot, B.J. Protocol for screening chemical agents and natural products against animal tumors and other biological systems. Cancer Chemother. Rept. 1972, 3, 1–103. [Google Scholar]
- Cariddi, L.N.; Escobar, F.M.; Sabini, M.C.; Torres, C.V.; Zygadlo, J.L.; Sabini, L.I. First approaches in the study of cytotoxic and mutagenic damage induced by cold aqueous extract of Baccharis articulata on normal cells. Mol. Med. Chem. 2010, 21, 4–7. [Google Scholar]
- García Villalón, M.D. (Doctoral thesis). Inhibidores de la multiplicación del virus de la peste porcina africana (tesis doctoral). Universidad Complutense de Madrid. Madrid, España. 2002.
- Palomino, S.S.; Abad, M.J.; Bedoya, L.M.; Garcia, J.; Gonzales, E.; Chiriboga, X.; Bermejo, P.; Alcami, J. Screening of South American plants against human immunodeficiency vírus: preliminary fractionation of aqueous extract from Baccharis trinervis. Biol. Pharm. Bull. 2002, 25, 1147–1150. [Google Scholar] [CrossRef]
- Martinez, M.J.A.; Bessa, A.L.; Benito, P.B. Biologically active substances from the genus Baccharis L. (Compositae). Stud. Nat. Prod. Chem. 2005, 30, 703–759. [Google Scholar] [CrossRef]
- Montanha, J.A.; Moellerke, P.; Bordignon, S.A.; Schenkel, E.P.; Roehe, P.M. . Antiviral activity of Brazilian plant extracts. Acta Farm. Bonaer. 2004, 23, 183–186. [Google Scholar]
- Ferreira, C.G.T.; Campos, M.G.; Felix, D.M.; Santos, M.R.; de Carvalho, O.V.; Diaz, M.A.N.; de Almeida, M. R. Evaluation of the antiviral activities of Bacharis dracunculifolia and quercetin on Equid herpesvirus 1 in a murine model. Research in veterinary science 2018, 120, 70–77. [Google Scholar] [CrossRef]
- Visintini Jaime, M.F.; Redko, F.; Muschietti, L.V.; et al. In vitro antiviral activity of plant extracts from Asteraceae medicinal plants. Virol J 2013, 10, 245. [Google Scholar] [CrossRef]
- De Oliveira, C.B.; Comunello, L.N.; Maciel, É.S.; Giubel, S.R.; Bruno, A.N.; Chiela, E.C.; Gosmann, G. The inhibitory effects of phenolic and terpenoid compounds from Baccharis trimera in Siha cells: differences in their activity and mechanism of action. Molecules 2013, 18, 11022–11032. [Google Scholar] [CrossRef] [PubMed]
- Martínez, G.J.; Planchuelo, A.M.; Fuentes, E.; Ojeda, M. A numeric index to establish conservation priorities for medicinal plants in the Paravachasca Valley, Córdoba, Argentina. Biodivers. Conserv. 2005, 15, 2458–2475. [Google Scholar] [CrossRef]
- Andrighetti-Fröhner, C.R.; Sincero, T.C.M.; da Silva, A.C.; Savi, L.A.; Gaido, C.M.; Bettega, J.M.R.; Mancini, M.; de Almeida, M.T.R.; Barbosa, R.A.; Farias, M.R.; Barardi, C.R.M.; Simões, C.M.O. Antiviral evaluation of plants from Brazilian Atlantic Tropical Forest. Fitoterapia. 2005, 76, 374–378. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, E. Antiviral agents: characteristic activity spectrum depending on the molecular target with which they interact. Adv. Virus Res. 1993, 42, 1–55. [Google Scholar] [CrossRef] [PubMed]
- Del Barrio Alonso, G.; Álvarez Rodríguez, Á.L.; Valdés García, S.M.; Parra Fernández, F. Metodología de pesquisa preclínica de actividad anti-herpesvirus a partir de productos naturales. Rev. Cubana Farm. 2008, 42, 10–13. [Google Scholar]
- Loayza, I.; Abujder, D.; Aranda, R.; Jakupovic, J.; Collin, G.; Deslauriers, H.; Jean, F.I. Essential oils of Baccharis salicifolia, B. latifolia and B. dracunculifolia. Phytochemistry. 1995, 38, 381–389. [Google Scholar] [CrossRef]
- He, K.; Montenegro, G.; Hoffmann, J.J.; Timmermann, B.N. Diterpenoids from Baccharis linearis. Phytochemistry 1996, 41, 1123–1127. [Google Scholar] [CrossRef]
- Silva, F.G.; Oliveira, C.; Pinto, J.E.B.; Nascimento, V.E.; Santos, S.C.; Seraphin, J.C.; Ferri, P.H. Seasonal variability in the essential oils of wild and cultivated Baccharis trimera. J. Braz. Chem. Soc. 2007, 18, 990–997. [Google Scholar] [CrossRef]
- Frizzo, C.D.; Atti-Serafini, L.; Laguna, S.E.; Cassel, E.; Lorenzo, D.; Dellacassa, E. Essential oil variability in Baccharis uncinella DC and Baccharis dracunculifolia DC growing wild in southern Brazil, Bolivia and Uruguay. Flavour. Fragr. J. 2008, 23, 99–106. [Google Scholar] [CrossRef]
- Chaves, A.G.; Bruentti, P.C.; Massuh, Y.; Ocaño, S.F.; Torres, L.E.; Ojeda, M.S. Variabilidad entre poblaciones silvestres de Baccharis crispa Spreng. de la Provincia de Córdoba, Argentina. Phyton. 2014, 83, 145–153. [Google Scholar]
- Moreira, X.; Abdala-Roberts, L.; Nell, C.S.; Vázquez-González, C.; Pratt, J.D.; Keefover-Ring, K.; Mooney, K.A. Sexual and genotypic variation in terpene quantitative and qualitative profiles in the dioecious shrub Baccharis salicifolia. Sci. Rep. 2019, 9, 14655. [Google Scholar] [CrossRef] [PubMed]
- Simões-Pires, C.A.; Queiroz, E.F.; Henriques, A.T.; Hostettmann, K. Isolation and on-line identification of anti-oxidant compounds from three Baccharis species by HPLC-UV-MS/MS with post-column derivatisation. Phytochem. Anal. 2005, 16, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Palacios, P.; Gutkind, G.O.; Rondina, R.V.D.; Torres, R.A.D.; Coussio, J.D. Actividad antimicrobiana de Baccharis crispa Sprengel ("Carqueja", FA) y Baccharis notosergila Gris. Acta Farm. Bonaer. 1983, 2. [Google Scholar]
- Ceñal, J.P.; Giordano, O.S.; Rossomando, P.C.; Tonn, C.E. Neoclerodane diterpenes from Baccharis crispa. J. Nat. Prod. 1997, 60, 490–492. [Google Scholar] [CrossRef]
- Velázquez, A.M.; Mallorquín, Z.E.; Montalbetti, Y.; Campuzano-Bublitz, M.A.; Hellión-Ibarrola, M.C.; Kennedy, M.L.; Ibarrola, D.A. Assessment of General effects and gastrointestinal prokinetic activity of Baccharis crispa in mice. J. App. Biol. Biotech. 2019, 7, 30–34. [Google Scholar] [CrossRef]
- Li, H.; Zhou, C.; Pan, Y.; Gao, X.; Wu, X.; Bai, H.; Zhou, L.; Chen, Z.; Zhang, S.; Shi, S.; Luo, J.; Xu, J.; Chen, L.; Zheng, X.; Zhao, Y. Evaluation of antiviral activity of compounds isolated from Ranunculus sieboldii and Ranunculus sceleratus. Planta Med. 2005, 71, 1128–33. [Google Scholar] [CrossRef]
- Özçelik, B.; Kartal, M.; Orhan, I. Cytotoxicity, antiviral and antimicrobial activities of alkaloids, flavonoids, and phenolic acids. Pharm. Biol. 2011, 49, 396–402. [Google Scholar] [CrossRef]
- Zhang, W.; Qiao, H.; Lv, Y.; Wang, J.; Chen, X.; Hou, Y.; Tan, R.; Li, E. Apigenin Inhibits Enterovirus-71 Infection by Disrupting Viral RNA Association with trans-Acting Factors. PLoS ONE 2014, 9, e110429. [Google Scholar] [CrossRef]
- Hakobyan, A.; Arabyan, E.; Kotsinyan, A.; Karalyan, Z.; Sahakyan, H.; Arakelov, V.; Nazaryan, K.; Ferreira, F.; Zakaryan, H. Inhibition of African swine fever virus infection by genkwanin. Antiviral Res. 2019, 167, 78–82. [Google Scholar] [CrossRef]
- Martinez, F.; Massuh, Y.; Aguilar, J.J.; Ojeda, M.S.; Contigiani, M.S.; Núñez Montoya, S.C.; Konigheim, B.S. Cultivars of Tagetes minuta L. (Asteraceae) as a source of potential natural products with antiviral activity. J. Herb. Med 2020, 24, 100397. [Google Scholar] [CrossRef]
- Sartor, B.T.; Xavier, V.B.; Falcão, M.A.; Mondin, C.A.; dos Santos, M.A.; Cassel, E.; Astarita, L.V.; Santarém, E.R. Seasonal changes in phenolic compounds and in the biological activities of Baccharis dentata (Vell.) G.M. Barroso. Ind. Crop. Prod. 2013, 51, 355–359. [Google Scholar] [CrossRef]
Figure 1.
Effect of extracts from four wild populations of B. crispa against HSV-I and CHIKV. Viral inactivation of aqueous (Aqu) and organic extracts (CHCl3 and EtOH) from four wild populations of B. crispa, against HSV-I and CHIKV. Data represented virus yield (log PFU/mL) compared to viral control (infected and untreated cells), expressed as ± SE (n = 3) of three independent experiments. The dashed line represents the cut-off point for the virucidal activity, extracts below it are positive. *Significant difference between wild populations for the CHCl3 extracts p ≤0.05 (ANOVA DGC test). **Significant difference between wild populations for the EtOH extracts p ≤0.05 (ANOVA DGC test). ***Significant difference between wild populations for the Aqu extracts p ≤0.05 (ANOVA DGC test). CHIKV: Chikungunya virus; HSV-I: Herpes Simplex Virus Type 1; CHCl3: chloroform extract; EtOH: ethanolic extract; Aqu: aqueous extract; VP: Villa del Parque population; TC: Tala Cañada population; SG: San Gerónimo population; PP: Puesto Pedernera population.
Figure 1.
Effect of extracts from four wild populations of B. crispa against HSV-I and CHIKV. Viral inactivation of aqueous (Aqu) and organic extracts (CHCl3 and EtOH) from four wild populations of B. crispa, against HSV-I and CHIKV. Data represented virus yield (log PFU/mL) compared to viral control (infected and untreated cells), expressed as ± SE (n = 3) of three independent experiments. The dashed line represents the cut-off point for the virucidal activity, extracts below it are positive. *Significant difference between wild populations for the CHCl3 extracts p ≤0.05 (ANOVA DGC test). **Significant difference between wild populations for the EtOH extracts p ≤0.05 (ANOVA DGC test). ***Significant difference between wild populations for the Aqu extracts p ≤0.05 (ANOVA DGC test). CHIKV: Chikungunya virus; HSV-I: Herpes Simplex Virus Type 1; CHCl3: chloroform extract; EtOH: ethanolic extract; Aqu: aqueous extract; VP: Villa del Parque population; TC: Tala Cañada population; SG: San Gerónimo population; PP: Puesto Pedernera population.
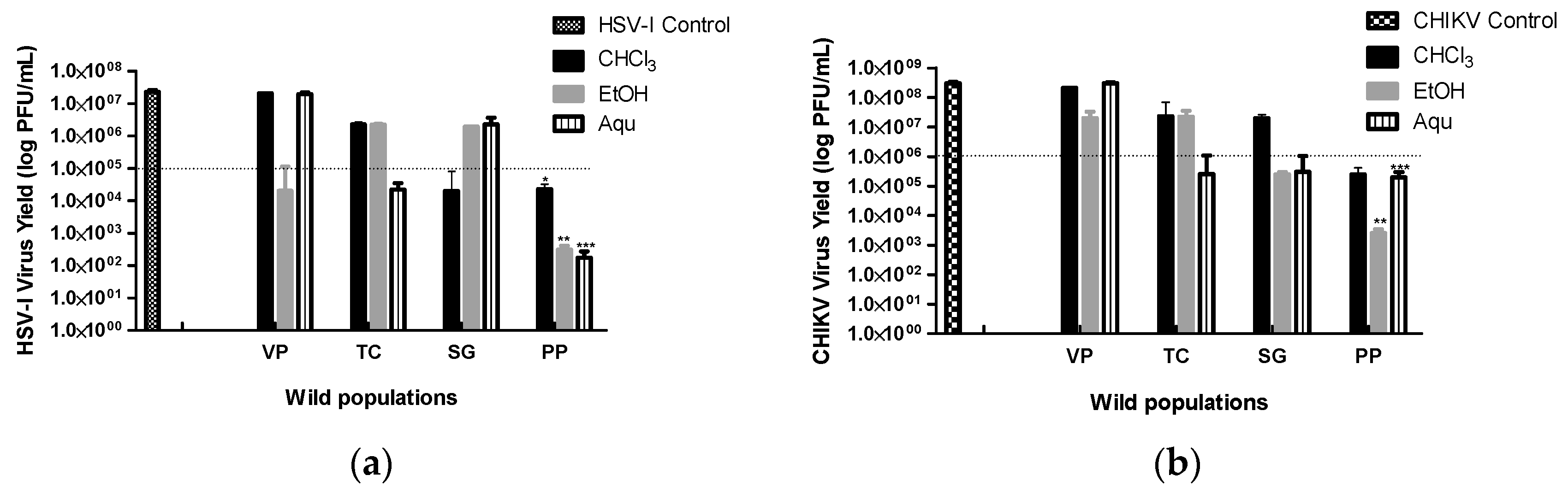

Table 1.
Cytotoxic and antiviral activities of aqueous and organic extracts from four wild populations of B. crispa.
Table 1.
Cytotoxic and antiviral activities of aqueous and organic extracts from four wild populations of B. crispa.
| Extracts/ positive control |
Population | CC50 (µg/mL)1 | SubTC (µg/mL)2 | CHIKV | HSV-I | ||||
|---|---|---|---|---|---|---|---|---|---|
| I (%)3 | EC504 (µg/mL) |
SI5 | I (%) | EC50 (µg/mL) |
SI | ||||
| CHCl3 | VP | 326.2 ± 6.7d | 134.9 ± 5.6c | 45.7 ± 0.8 | - | - | 17.6 ± 0.6 | - | - |
| TC | 109.3 ± 4.6b | 52.0 ± 2.6a | 59.4 ± 0.6c | 44.2 ± 0.2 | 2.5 | 41.3 ± 0.6 | - | - | |
| SG | 169.1 ± 5.7c | 105.6 ± 10.1b | 67.0 ± 1.2b | 78.8 ± 0.5 | 2.1 | 48.3± 0.1 | - | - | |
| PP | 94.8 ± 1.6a | 45.7 ± 0.4a | 81.6 ± 0.9a | 28.2 ± 0.1 | 3.4 | 55.2 ± 0.4 | 41.4 ± 0.1 | 2.3 | |
| EtOH | VP | 92.9 ± 3.5ª | 20.2 ± 1.1a | 35.9 ± 0.6 | - | - | 32.5 ± 0.6 | - | - |
| TC | 413.6 ± 9,5c | 219.8 ± 5.4c | 43.0 ± 0.9 | - | - | 57.6 ± 0.4b | 190.8 ± 0.3 | 2.2 | |
| SG | 159.2. ± 1.5b | 103.6 ± 2.8b | 63.5 ± 0.9b | 81.6 ± 0.4 | 2 | 61.6 ± 0.5b | 84.1 ± 0.6 | 1.9 | |
| PP | 571.9 ± 0.3d | 326.8 ± 9.8d | 79.0 ± 0.8a | 188.7 ± 0.1 | 3 | 78.9 ± 0.2a | 207.1 ± 0.1 | 2.8 | |
| Aqu | VP | 700.4 ± 17.3d | 482.0 ± 11.4b | 27.8 ± 0.1 | - | - | 19.4 ± 0.2 | - | - |
| TC | 611.9 ± 9.1c | 507.0 ± 0.7c | 35.8 ± 0.2 | - | - | 30.2 ± 0.2 | - | - | |
| SG | 422.3 ± 9.6a | 342.8 ± 9.3a | 57.9 ± 1.1b | 296.0 ± 0.2 | 1.4 | 38.9 ± 0.1 | - | - | |
| PP | 573.4 ± 2.3b | 491.9 ± 0.7b | 74.0 ± 0.6a | 332.4 ± 0.5 | 1.7 | 51.2 ± 0.7 | 480.4 ± 0.1 | 1.2 | |
| Acyclovir | - | >200 | - | - | - | - | 100 | >1000 | >20.000 |
Data are mean values from three independent experiments ± SE performed in triplicate. Values in bold font indicate positive extracts. CHCl3: chloroform extract; EtOH: ethanol extract; Aqu: aqueous extract; VP: Villa del Parque population; TC: Tala Cañada population; SG: San Gerónimo population; PP: Puesto Pedernera population; CHIKV: Chikungunya virus; HSV-I: Herpes Simplex virus type 1. 1. CC50: concentration that reduced the viable cells to 50%. 2. SubTC: concentration that ensures 80% of the viable cells. 3. Percentages of viral inhibition at SubTC of extracts of B.crispa on CHIKV or HSV-I. The results are expressed as a percentage of inhibition compared to viral controls (100%). 4. EC50: concentration of the extract that produces a 50% inhibition of the virus-induced cytopathic effect. 5. SI: selectivity index was determined by the ratio of CC50 /EC50. a-d Different letters indicate significant differences between wild populations for the same type of extract: p ≤0.05 (ANOVA DGC test).
Table 2.
Phytochemical screening of organic and aqueous extracts of wild populations of B. crispa.
| Compound groups | Extracts | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CHCl3 | EtOH | Aqu | ||||||||||
| VP | TC | SG | PP | VP | TC | SG | PP | VP | TC | SG | PP | |
| Tanins | + | ++ | ++ | ++ | ++ | ++ | ++ | ++ | + | + | +++ | +++ |
| Antocianins | - | - | - | - | - | - | - | - | - | - | - | - |
| Cardiac glycoside | - | - | - | - | - | - | - | - | - | - | - | - |
| Carbohydrate | - | - | - | + | - | - | + | ++ | - | - | - | ++ |
| Saponins | - | - | - | + | - | - | + | ++ | + | ++ | ++ | +++ |
| Flavonoids | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | - | + | + | ++ |
| Anthraquinone | - | - | - | - | - | - | - | - | - | - | - | - |
| Phytosterols | - | + | + | ++ | - | - | - | ++ | - | - | - | - |
| Alkaloids | - | - | - | - | - | - | - | - | - | - | - | - |
+++ (Very abundant); ++ (Abundant); + (Not abundant); - (Not detected); VP (Villa del Parque); PP (Puesto Pedernera); CHCl3 (Chloroform); EtOH (ethanolic); Aqu (Aqueous).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



