
Submitted:
26 July 2023
Posted:
31 July 2023
You are already at the latest version
Abstract
Fermented rice is known as a healthy food due to the presence of lactic acid bacteria as fermentative microorganisms. The study was carried out to discover and characterize the lactic acid bacteria (LAB) from white and red fermented rice (BG varieties). Fermentation was carried out naturally by soaking red, white, raw and cooked rice in water using an earthen pot. Species were confirmed based on 16S r DNA gene sequencing and studied for phenotypic characteristics including morphological, physiological (growth temperature, salt tolerance, milk coagulation) and biochemical characteristics (carbohydrate fermentation pattern) using API 50CH kits. Distinct clusters like cocci (48), diplo cocci (30) and rods (30) shaped bacteria were observed from fermented rice. Five species of lactic acid bacteria including Latilactobacillus curvatus GR Lb1,2 10 and 11 (predominant Bacillus species), Latilactobacillus graminis Lb8, Limosilactobacillus fermentum GRLb17, Weissella confuse GRLb4 and Pediococcus pentosaceus GRLc1 were identified. The base pair length of amplified DNA for the isolates was 1500 Bp. The beneficial physiological and biochemical properties of isolated Lactobacillus species from fermented rice revealed their potential applications in the food industry.
Keywords:
Fermentation
; 16SrDNA
; Lactic Acid Bacteria
; Phenotypical
; Genotypical.
1. Introduction
Fermentation is the most simple and economical way of improving the nutritional value, sensory properties and functional qualities of food which can also be applied in the production of functional food such as probiotic food [1]. Fermented foods are produced world-wide using various manufacturing techniques, raw materials and microorganisms [2]. Fermented foods offer beneficial health effects due to the antimicrobial effects of the Lactic acid bacteria present in them. Due to their interesting beneficial properties, LAB are widely being used as starter cultures, as probiotics, and as microbial cell factories for production of bioactive components including lactic acid which shows antimicrobial effect and interferes with the growth of pathogenic microorganisms [3]. Production of functional food prepared using non-dairy-based materials can satisfy the dietary requirements of consumers who have allergies with dairy-based products [4]. Microorganisms possess probiotic activity that promote the positive health image during fermentation [5]. Fermented foods can be consumed by vulnerable groups such as children, expectant, breastfeeding mothers, aged, sick and recovering persons for good health [6].
In the past, farmers and labourers from Asian countries kept the leftover rice for fermentation overnight and consumed it for breakfast which is known to offer several health benefits. This food gives sustained energy with all-natural nutritional supplements which arise from the fermenting bacteria in it. Consumption of fermented rice is believed to cause a soothing effect on the intestines because the rice is fermented and digested by bacteria. Bacteria add fibre to stool and also produce vitamin B complex (B12), vitamin D, and vitamin K. Fermented rice is locally called as ‘pazhaya sadham’, in India which is prepared by overnight soaking of cooked parboiled rice in water. Fermentation enhances the bioavailability of essential nutrients especially minerals through the enzymatic reduction of Phytate [7]. Fermented rice is one of the neglected foods in countries where rice is the staple food. The consumption of fermented rice as a contributor to a healthy lifestyle is either ignored or ridiculed as a poor man's lifestyle food. Many strains of probiotic Lactic acid bacteria have been isolated from different traditional fermented foods, insufficient research data are available on the study connected to identified lactic acid bacteria from fermented rice. Exploring the natural (wild) bacteria responsible for fermentation of rice is essential to confirm the health benefit of fermented food product. Pedicoccus pentosaceus and Lactobacillus plantarum are potentially probiotic bacteria isolated from fermented rice gruel prepared from Ponni variety rice (Oryza sativa) [8]. The parboiled rice and black grams are generally used in Sri Lanka for cooking and used for producing rice based- fermented product [9]. Fermented white and red Batalagoda (Bg) rice varieties are used for the preparation of fermented food like, hoppers, “Idli’ and “Dosai’ The study is attentive to distinguishing the responsible microorganism for the fermentation of red and white rice Bg varieties.
LAB constitutes a diverse group of industrially important and safe bacteria with interesting beneficial properties that are primarily being used as starter cultures, as probiotics, and as microbial cell factories for the production of highly valuable bioactive components [10]. Exploring LAB present in unknown niches may identify the unique species or strains with pertinent technological and probiotic properties. Autochthonous rather than allochthonous starter cultures are chosen in the contemporary industry of fermented food products, due to the better adaptation and performance of autochthonous strains to the matrix they originate from. The wide distribution and important nutritive value of cereals have focused attention on cereal as raw material for the development of new fermented functional foods. Most probiotic foods available worldwide are milk based and few attempts have been made to develop probiotics using cereal as the substrate. LAB, isolated from fermented rice has the potential to use as a starter culture in cereal-based fermented food products or probiotic food.
The identification of lactic acid bacteria based on phenotypic characterization is still being used as a starting point prior to molecular characterization of the key characteristics in bacterial taxonomy [11]. Seven predominant Lactobacillus species and one coccus were isolated from fermented white are red raw and cooked rice (Bg varieties) and identified at the genus level [12]. These novel Lactobacillus species should be confirmed by molecular study. Therefore, study was carried out to recognize the species of lactic acid bacteria isolated from fermented rice based on 16S r DNA gene sequencing with the phenotypic characterization.
2. Materials and Methods
2.1. Study location
Isolation and characterization of Lactobacillus sp. from fermented rice was carried out at the Department of Oral Medicine and Periodontology, Faculty of Dental Sciences, University of Peradeniya, Sri Lanka and molecular identification by 16Sr DNA sequencing has been carried out at the Department of Food Technology, University Putra, Malaysia (UPM).
2.2. Materials
Fermented red and white raw rice and cooked rice samples (Bg varieties -Red and white) were used as isolation sources. MRS media, MRS sorbitol (0.2%) and MRSL cysteine (0.25%) were used in order to improve the specificity of the medium for the isolation of diverse species of Lactic acid bacteria. The pH of the MRSagar, MRS sorbitol (0.2%) and MRS L cysteine (0.25%) were 6.2,6 and 5.8 respectively.
The survived distinguished predominant eight bacteria including seven rods shaped (Isolation code Lb-1, Lb-2, Lb-4, Lb-8, Lb-10, Lb-11, Lb-17 and Lc-1) and one spherical shaped bacteria were studied for phenotypic characterization and molecular identification.
2.3. Preparation of Rice samples
The red raw and white rice (Oryza sativa) were cleaned and washed twice, and cooked separately in water (rice: water 1:3) for 30 minutes up to get a soft consistency. The fermentation was carried out naturally by soaking raw or cooked rice(50g) in sterile distilled water (rice: water 1:3) overnight in earthen pots at 27 0C in an outside environment. Each rice variety of cooked and raw rice samples were kept in separate pots : sample were collected from each pot, introduced on the culture plates up to three dilution level and microorganisms were isolated from each plate.
2.4. Phenotypic characterization
Isolates were identified by their phenotypic characterization. Phenotypic characterization was carried out by studying the morphological, physiological, and biochemical characteristics based on Barrow, and Felthman [13] and these bacteria were identified at species level using phenotypic features and carbohydrate fermentation patterns in API 50CHL system.
2.4.1. Morphological characteristics of the isolates
The methods for Gram Staining, catalase activity, motility, and spore formation were followed according to Barrow and Felthman [13]. The morphological structure of isolates was observed with the light microscope using an oil immersion lens (10 X100) following Gram staining.
2.4.2. Physiological and biochemical characterization
- Culture preparation
Two to three colonies of the 8 isolates were inoculated in MRS-L cysteine (0.25%) broth (10 ml) and incubated at 37 °C for 16 h to obtain fresh culture (Figure A2). All experiments were carried out in replicates.
- Growth at different temperatures
The growth in different temperatures was evaluated with some modifications as described in Hatice [14]. MRS-L cysteine (0.25%) containing bromecresol purple (0.04 g/l) indicator was prepared as the test medium and 5 ml of the test medium was transferred into clear glass test tubes for observation. Fresh cultures (50 μL, 1%,18 h) were transferred into the tubes and incubated for 7 days at 10 °C, 37 °C, 40 °C, 45 °C and 55 °C in an incubator. The temperature ranges were selected based on the optimum growth temperatures of the lactobacillus species. During incubation, microbial growth was confirmed from the colour change of the cultures from purple to yellow. The MRS-L cysteine (0.25%) broth culture incubated at 37 °C and sterile broth (without culture) at the tested temperature were used as positive and negative controls respectively.
- Growth at different NaCl concentrations
Salt tolerance of bacteria was used to characterize the lactic acid bacteria for identification (Barrow and Felthman, 1995[13]). Isolated were tested for their tolerance against different NaCl concentrations based on the method by Hatice [14] with some modification. The NaCl concentrations of 2, 4 6.5 and 10% were selected. Test media containing bromocresol purple (5 ml) indicator was prepared according to the appropriate concentrations (0.04 g/l) and transferred into clear glass tubes which were inoculated with 1% cultures (18 h) and then incubated at 37 °C for 7 days. The cell growth was confirmed by the colour change from purple to yellow. The MRS-L cyteine (0.25%) broth culture without NaCl and broth without culture were used as a positive and negative control, respectively.
- Gas production with glucose
Carbon dioxide production from glucose was observed in order to determine the homofermentative and heterofermentative characteristics of the 7 isolates. MRS broth sans citrate and inverted Durham tubes were prepared and inoculated with 1% fresh cultures (18 h). The test tubes were incubated at 37 °C for 7 days. Gas accumulation in Durham tubes was observed during 7days which is evidence for CO2 production from glucose. The MRS-L cysteine (0.25%) broth without culture was used as the negative control [14].
- Milk coagulation assay
Milk coagulation assay in skim milk was performed as previously described by Marokki et al. [15] with some modifications. Isolates were introduced into MRS broth and incubated at 37 °C for 16-18 h for activation. The culture (1%) was transferred into culture tubes containing sterile skim milk (12.5%) and coagulation of milk was determined after 16 h of incubation at 37 °C. The probiotic mixed starter culture (ABT-3, Hansion Denmark, Lactobacillus acidophilus LA5 and Streptococcus thermophilus TH4 and Bifidobacterium bifidum Bb 12) inoculated with skim milk and the skim milk without culture were used as positive and negative controls, respectively.
2.5. Carbohydrate fermentation pattern of isolates on API 50 CHL system
The isolates were identified to species level using API 50 CH kits based on the carbohydrate’s fermentation profile of CHL media (BiomÈrieux, France) by following the manufacturer's instructions.
2.5.1. Preparation of the culture
Several identical colonies were picked up from MRS culture plates incubated at 37 °C for 2 days and introduced into the culture tubes containing sterile distilled water (2 ml) for the preparation of a heavy culture suspension. The suspension was added into the culture tubes containing sterile distilled water (5 ml) to prepare a suspension with turbidity equivalent to McFarland standard 2 and the number of the drops required were recorded (n). The API 50 CHL medium (10 ml) was inoculated by transferring twice the number of drops of prepared suspension (2n) into the API 50 CHL Medium ampule. This inoculated medium was mixed well and used immediately.
2.5.2. Preparation of incubation box
The references of the isolates were marked on the elongated flap of the tray and sterile distilled water (10 ml) into the honeycombed wells of the tray to create a humid atmosphere.
2.5.3. Preparation of the strips
Each API strip is made up of 5 smaller strips each containing 10 numbered tubes. The strips were cut into five smaller stripes (0-9, 10-19, 20-29, 30-39 and 40-49) and placed in the proper order in the incubation tray.
2.5.4. Inoculation of strip
The inoculated API 50 CHL medium was distributed using a sterile pipette among 50 tubes without the formation of bubbles and all the tubes were covered with mineral oil to maintain anaerobic conditions. The inoculated stripes were incubated at 37 °C for 48 h.
2.5.5. Reading and Interpretation
The strip was read for positive (+) and negative reactions (-) after 48 h incubation and the results were recorded on the result sheet. A positive test corresponds to acidification shown by the Bromcresol purple indicator contained in the API medium changing from purple to yellow except in the Esculin test (tube no. 25) where a change in color from purple to black was considered as positive. The biochemical profile obtained for the strain can be identified using the apiwebTM identification software with database (V5.1). The API profiles were analysed (Table 1 and Figues 1 and 2) using API Web TM identification software (Biom Èrieux, France). In cases where two or more significant taxa was indicated, reference was made to standard texts for identification by considering their physiological characteristics.
2.6. Identification of Lactobacillus sp. by 16SrDNA sequencing
2.6.1. Extraction of genomic DNA
The overnight MRS broth culture (maximum 2 x 109 cells) was harvested in a microcentrifuge tube by centrifuging for 10 min at 5000 x g and the supernatant was discarded. The Bacterial pellets were used for the extraction of DNA. DNA was extracted using the DNA extraction kit (QIAGene, Hilden, Germany) following the manufactures protocol with some modifications. Extracted DNA (3 μl of DNA) mixed with 2 μl of 6x loading dye and was visualized by agarose gel (1.5%) electrophoresis under UV light and documented with Gel Doc system. They were then taken to the PCR step.
2.6.2. Polymerase Chain Reaction for the amplification of 16S rDNA Region
The most common primer pair devised by Weisburg et al. [16] were forward primer 27F and reverse primer 1492R which are currently referred to as universal primers. Forward primer is complementary to the 5’end of 16SrDNA, and the reverse primer is complementary to the 3’ end of 16SrDNA region. Amplification of the 16S rDNA region was carried out using universal primers including forward primer 27F (5’-AGAGTTTGATCCTGG CTCAG-3’) and reverse primer 1492R (5’-GTTACCTTGTTACGACTT-3’) [17].
Amplification reactions were performed in a total reaction volume of 50 μl containing 2X Taq Master Mix (25 μl) (New England Biolabs Inc, Ipswich, UK), 0.25 mM forward primer (1μl), 0.25 mM (1μl) reverse primer (First base, Malaysia) and deionized water (22 μl)) and genomic DNA. The 1 μl of genomic DNA was mixed with 49μl of PCR mixture and the final reaction mixture was taken for PCR steps. A negative-DNA control was performed by adding 1 μl of nuclease-free water in the PCR mixture instead of genomic DNA. The preliminary experiments were carried for the confirmation of the amplification of the DNA. Gradient temperature PCR (Eppendorf, Homburg, Germany) was carried out with the initial heating of 95 °C for 3 min, followed by 40 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 55 s, ex-tension at 72 °C for 1 min, and terminating with a 10 min of final incubation of 72 °C.
2.6.3. Separation of amplified PCR products
Preparation of agarose gel: Agarose gel (Vivantis Biotechnology, Malaysia) was prepared by dissolving 0.8 g agarose in 100 ml of boiling TAE buffer(1X), cooled nearly to 45 °C and 1.5 % Red Safe nucleic acid staining solution (concentration 20000X, Intron Biotechnology) was added. The prepared agarose gel was poured into the gel casting stand and the combs were placed. The combs were taken out after the formation of rigid gel within the well. PCR products (5 μl) and 2 μl of loading dye (QIAGene, Hilden, Germany) were mixed and loaded into wells. A DNA size-marker (2 μl) (10 kb+100bp, Fermentas) was loaded into the first well to observe right amplified region.
Electrophoresis of the products: Extracted DNA and PCR products were electrophoresed at 180 V.80 mA for one hour in the gel electrophoresis system (Major Science, Taiwan). Amplified band pattern was visualized in a UV transilluminator (Major Science, Taiwan) and documented with Gel Doc system (UVIdoc System, model GAS9000/9010).
2.6.4. Sequencing and Phylogenic tree development.
The amplified 16S rDNA PCR products were sequenced by First BASE Laboratories, Malaysia (First BASE, Kuala Lumpur, Malaysia) using 27F and 1492R primers corresponding the positions of forward primer 27F (5’-AGAGTTTGATCCTGG CTCAG-3’) and reverse primer 1492R (5’-GTTACCTTGTTACGACTT-3’). The sequences of reverse primers1492R were converted into its reverse order in reverse complement. The sequences of the isolates were edited using the Bio edit -7.2 software. The sequences with the highest QV were selected and both reverse and forward sequence were combined. Sequence homologies of the isolates were examined by comparing the sequences obtained with 16S rDNA and sequences deposited in the nucleotide databases of the GenBank (NCBI), using the Basic Local Alignment Search Tool (BLAST) program and gene accession numbers were obtained.
The representative sequences of the (Lb-1, Lb-2, Lb-4, Lb-8, Lb-10, Lb-11, Lb-17 and LC-1) isolates were then aligned using ClustalW of MEGA 11package and the phylogenetic tree was built using the neighbor-joining method [18]. The stability of the tree was assessed by the bootstrap method using 1000 replications.
3. Results
3.1. Morphological characteristics of isolates
After Grams’ staining cell morphology of all isolates was observed using the oil immersion objective (10X100) of light microscope. Among studied colonies (Figure A1) the majority of the isolated microorganism from fermented rice appears like cluster of cocci and rods shaped bacillus.
Figure 1.
Gram positive, catalase negative bacteria and Candida sp. in the MRS and MRS modified media from fermented rice. WR-While Raw, WC-White cooked, RR-Red raw, and RC-Red cooked fermented rice.
Figure 1.
Gram positive, catalase negative bacteria and Candida sp. in the MRS and MRS modified media from fermented rice. WR-While Raw, WC-White cooked, RR-Red raw, and RC-Red cooked fermented rice.
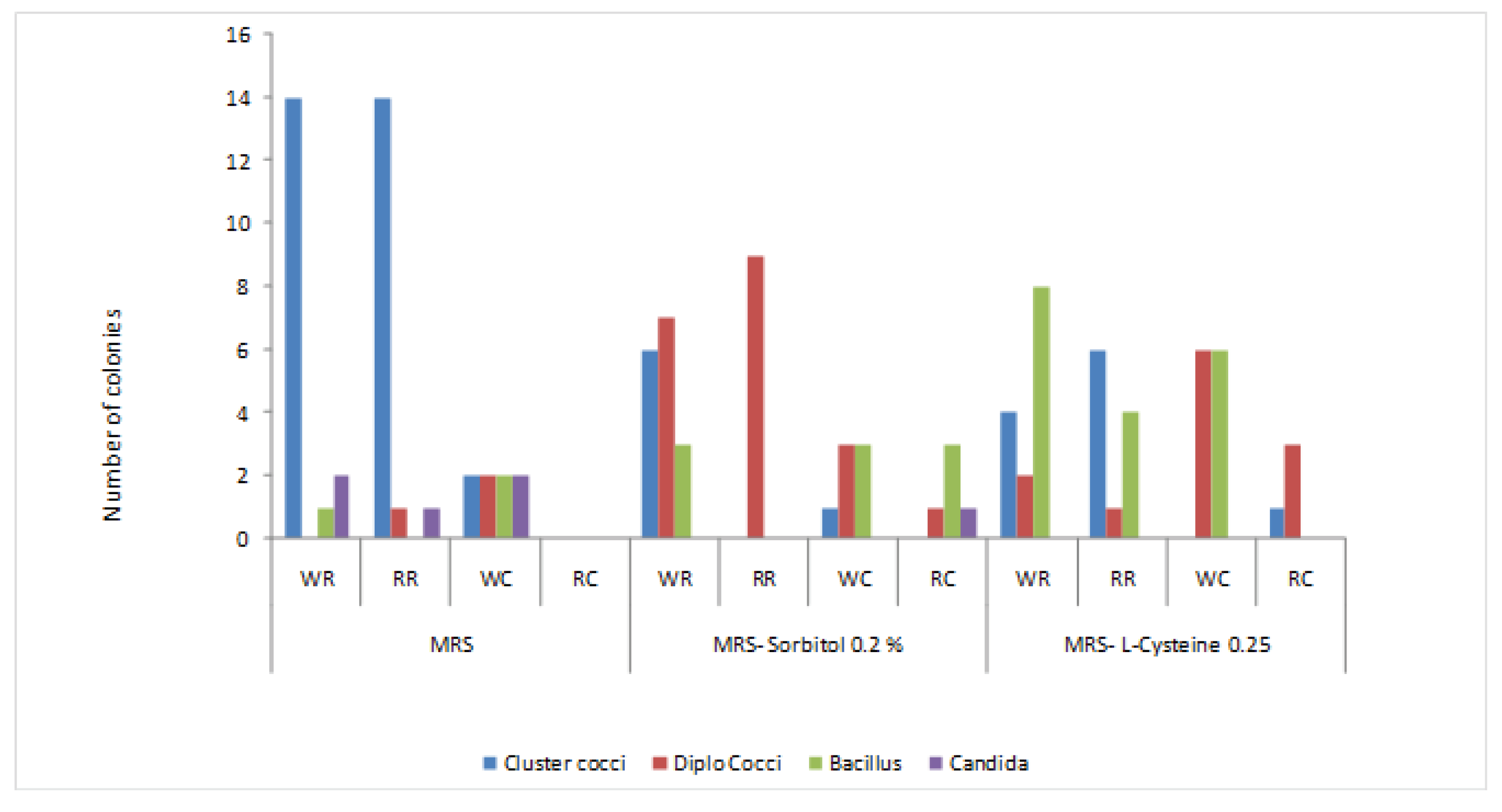
Three different morphotypes of the isolates including the cluster coccus (48), diplococcus (30) and rod (30) (Fiqure1,) were observed with Gram positive and catalase negative reactions in all three media. The number of clusters like coccus species are predominant in MRS media isolated from red and white raw rice and number of rod-shaped bacteria is high in MRSL cysteine (0.25%). The number of studied colonies shows that the majority of the isolated microorganism from fermented rice appears like cluster of cocci and rod-shaped bacillus. A subsequent (3-4 times) purification process was attempted for survived isolates, then rod-shaped bacteria and predominant cluster like coccus were tested for motility and endospore test. Seven rods and cluster like coccus were identified as lactic acid bacteria as they were Gram positive, catalase negative, non-motile and did not forming endospores which were preserved in MRS broth medium containing 20% (v/v) glycerol at -80 °C as frozen culture for long term storage.
Four morphologically distinguished types of lactobacilli were observed including short, medium, long and very long regular rods and appeared as single, pairs or V shaped short chains (Table 1) and coccus appeared as clusters in similar morphological appearance.
3.2. Physiological and biochemical characterization
Lactobacillus are mostly used as starter culture for industrial purpose, selected seven rod-shaped bacteria was studied for physiological characterization (Table 2).
3.2.1. Growth at different temperatures
Different temperatures (10 °C, 37 °C, 40 °C, 45 °C and 55 °C) were used to find the optimum growth temperature to use as incubation temperature for yoghurt production. The positive growth and survival of isolates was observed for all the isolates at temperatures 10°C, 37 °C, 40 °C and 45 °C with the color change from purple to yellow. (Table 2). A higher growth turbidity was observed visually in the isolates Lb-1. 2, 4, 10, 11and 17 (Latilactobacillus curvatus strain GRLb-1,2,4,11,17) at a temperature range of 10°- 40 °C compared to the turbidity at 45 °C. The growth temperature at 10 °C is a positive attribute to the probiotic Lactobacillus sp., there may be the possibility of maintaining the viable count of the probiotic at the therapeutic minimum count (107cfu/ml or g of carrier substrate) during refrigerated storage. The isolate Lb-8 (Latilactobacillus graminis strain GRLb8) grew dense at 37 °C and 40 °C. All isolates prefer the mesophilc temperature conditions than thermophilic temperatures for growth. The positive control at 37 °C shows the heavy growth with yellow colour while the negative control without culture appeared in purple color even after 7 days of incubation.
Generally, different species of the genus Lactobacillus can tolerate temperature ranging from 15 °C to 45 °C. Lactobacillus acidophilus, Lactobacillus delbruki and Lactobacillus salivarius can grow at 45 °C and unable to grow at 15 °C while Lactobacillus casei, Lactobacillus plantarum and Lactobacillus berevis can grow at 15°C and cannot grow at 45 °C [13].
3.2.2. Growth at different NaCl concentrations
It is observed that all the cultures showed positive heavy growth at 0, 2, 4 and 6.5% NaCl concentrations with the yellow color change of the media. However, no growth was observed for all cultures at 10% NaCl concentration (Table 2). The negative control (devoid of culture) did not show any growth and the colour of the medium remained purple. In a previous study, the Lactobacillus species isolated from Algerian goat milk has shown 2% and 4% of salt tolerance (Marroki, [15]). The tolerance to 6.5% NaCl centration is beneficial as a starter culture used in the fermentation process which should survive in fermentation environment with high salt concentration [19].
3.2.3. Gas production with glucose
In order to determine the homofermentative or heterofermentative nature of the cultures, all the isolates were studied for the gas production from glucose. The isolate Lb-8 (Latilactobacillus graminis GRLb8) produced gas during the fermentation of glucose showing the heterofermentative nature while others did not release gas from glucose and showed homofermentative nature (Table 2). Genus Lactobacillus can be either hetero or homofermentative (Barrow and Felthman, [13]).
Homofermentative organism ferment glucose to two moles of lactic acid, generating a net of 2 ATP per mole of glucose metabolized. Lactic acid is the major product of this fermentation. Heterofermentative lactic acid bacteria ferment 1 mole of glucose to 1 mole of lactic acid, 1 mole of ethanol, and 1 mole of CO2. One mole of ATP is generated per mole of glucose, resulting in less growth per mole of glucose metabolized. Because of the low energy yields, lactic acid bacteria often grow more slowly [13].
3.2.4. Milk coagulation and curd formation
Coagulation and curd formation of skim milk was shown by all isolates. LAB generally possesses the ability to coagulate milk due to the production of lactic acid leading to a decrease in pH and presence of a complete proteolytic system consisting of a cell envelope associated proteinase which allows the efficient degradation and utilization of casein [20]. The ability of bacteria to coagulate autoclaved milk within 16 h at 42 °C by 1% freshly coagulated inoculum in cheese production is defined as fast milk-coagulating (Fmc+) strain [21].
The isolates led to coagulation of milk within 16 h at 37 °C and can be considered as fast milk-coagulating (Fmc+) strains. Coagulation of milk by the isolates revealed their potential as starters or adjunct cultures in the production of fermented dairy food products.
3.2.5. Carbohydrate fermentation pattern of Lactobacillus sp.
Seven rods and one coccus-shaped bacteria were identified as Lactobacillus sp. as there were Gram positive, catalase negative, non-motile and without endospore formation. As per the carbohydrate fermentation pattern of isolated Lactobacillus species using classical and biochemical tests (using API 50 CHl BioMeriex, France), seven Lactobacillus species and one cluster like coccus were identified to species level (Table 3). There were five different Lactobacillus sp. and one Pedicoccus sp. identified among the tested organism. Isolates Lb-1, Lb-2 and Lb-11 were identified as Lactobacillus curvatus ssp. curvatus with 99% identity and Lc-1 was identified as Pediococcous pentosaceus with 99% identity.
The carbohydrate fermentation pattern of the isolates showed that all the isolates were not able to ferment most of the sugars and were able to ferment only some of the sugars (12-45%) tested using the API 50 kits (Figure 2 and Table S1). All the isolates were able to ferment maltose, D-glucose, D-frutose and D mannose. The isolate Lb-8 did not ferment the N-Acetyl glucosamine and D-galactose but all other isolates showed a positive reaction to them. The carbohydrate fermentation by isolates Lb-1, Lb-2 and Lb-11 showed a similar pattern.
Figure 2.
Carbohydrate fermentation pattern of Lb-10(a) and Lb-1 (b) from fermented rice tested using API 50 CHL system. The sugar fermentation was identified by the colour change of Bromcresol purple indicator in the API medium to yellow except in the Esculin test (tube no. 25) where a change in color from purple to black was considered as positive. The medium remined in purple indicated negative result for sugar fermentation.
Figure 2.
Carbohydrate fermentation pattern of Lb-10(a) and Lb-1 (b) from fermented rice tested using API 50 CHL system. The sugar fermentation was identified by the colour change of Bromcresol purple indicator in the API medium to yellow except in the Esculin test (tube no. 25) where a change in color from purple to black was considered as positive. The medium remined in purple indicated negative result for sugar fermentation.

The phenotypic characteristics of microorganisms are useful tools for presumptive classification. Identification of species, especially within the genus Lactobacillus based on the information on the biochemical and metabolic traits of LAB is not confirmative due to an increasing number of lactic acid bacteria species which vary on a small number of biochemical traits [22]. The fermentation ability of some carbohydrates is plasmid-encoded. The losing and acquiring plasmids lead to metabolite inconsistencies and affect the fermentation pattern [23]. The combination of commercially available API systems with conventional phenotypic methods and genotypic techniques ensures the identification of species with its known phenotypic properties.
3.3. Molecular identification and genotypic characteristics of Lactobacillus sp.
3.3.1. Genomic DNA Isolation
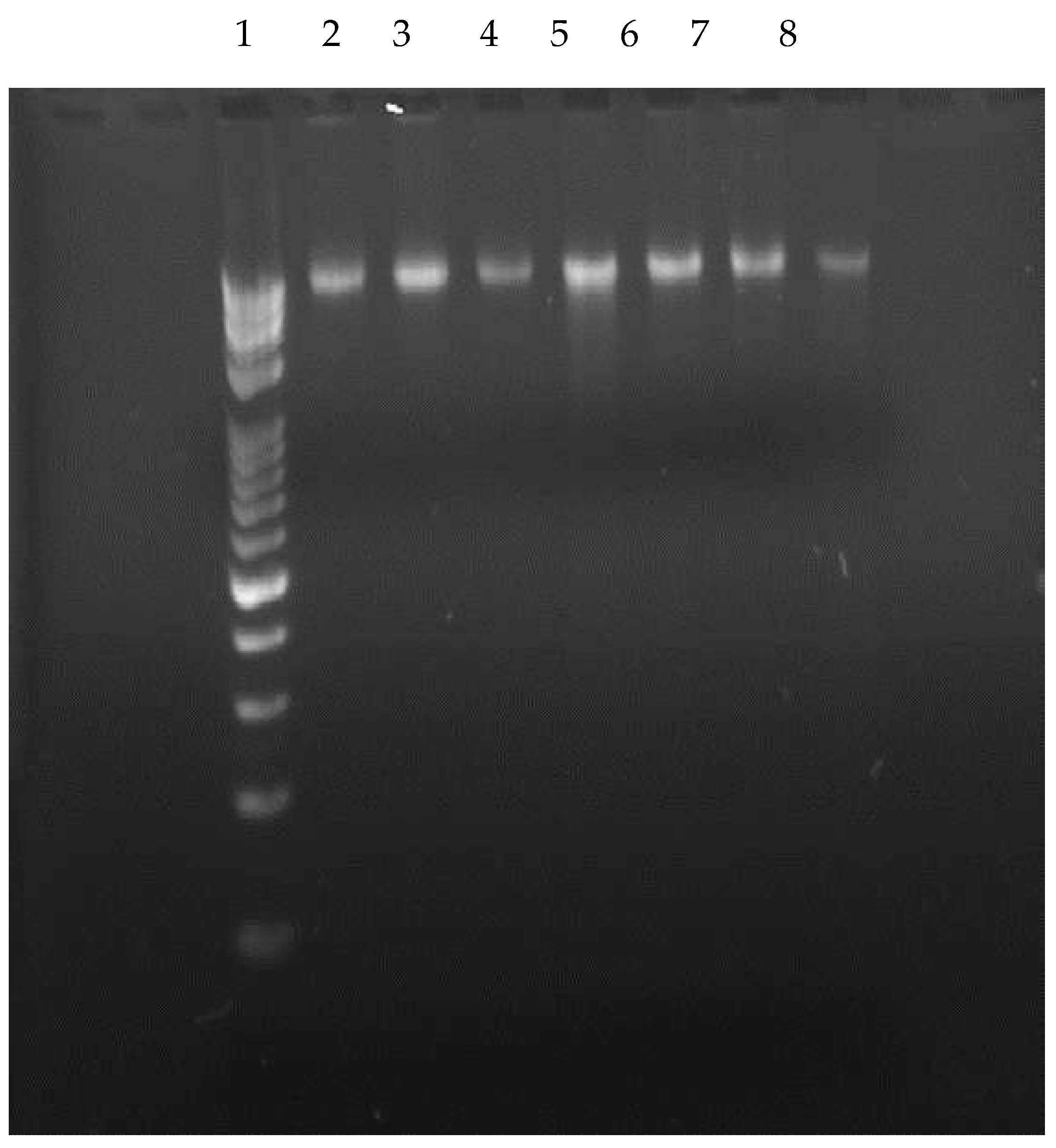
Extraction of genomic DNAs of isolates were carried out according the manufacture instruction and used for amplification of 16SrDNA Region by PCR protocol. Figure 3 shows the extracted DNA from isolated Lactobacillus species which confirm the present of the DNA after the extraction procedure.
Figure 3.
Visualization of extracted DNA by gel electrophoresis. 1. 10kb+100bp DNA Ladder, 2. Lb-1, 3. Lb-2, 4. Lb-4, 5. Lb-8, 6. Lb-10,7. Lb-11, 8. Lb-17. Band pattern in the gel confirmed the presence of extracted DNA.
Figure 3.
Visualization of extracted DNA by gel electrophoresis. 1. 10kb+100bp DNA Ladder, 2. Lb-1, 3. Lb-2, 4. Lb-4, 5. Lb-8, 6. Lb-10,7. Lb-11, 8. Lb-17. Band pattern in the gel confirmed the presence of extracted DNA.
3.3.2. Amplification of 16S rDNA region
After the confirmation of DNA extraction, extracted DNA was taken up for PCR for the amplification of 16S rDNA region and the amplification of the PCR products was visualized by agarose gel electrophoresis under UV light. The amplification of the PCR product bands was observed in gel documentation system and the length of amplification products was nearly 1500 bp (Figure 4).
Figure 4.
Amplified Products of 16SrDNAregion of the isolates. 1. Lb-1, 2. Lb-2, 3. Lb-4, 4. Lb-8, 5. Lb-10, 6. Lb-11, 7. Lb-17, 8.10kb+100bp DNA Ladder.
Figure 4.
Amplified Products of 16SrDNAregion of the isolates. 1. Lb-1, 2. Lb-2, 3. Lb-4, 4. Lb-8, 5. Lb-10, 6. Lb-11, 7. Lb-17, 8.10kb+100bp DNA Ladder.
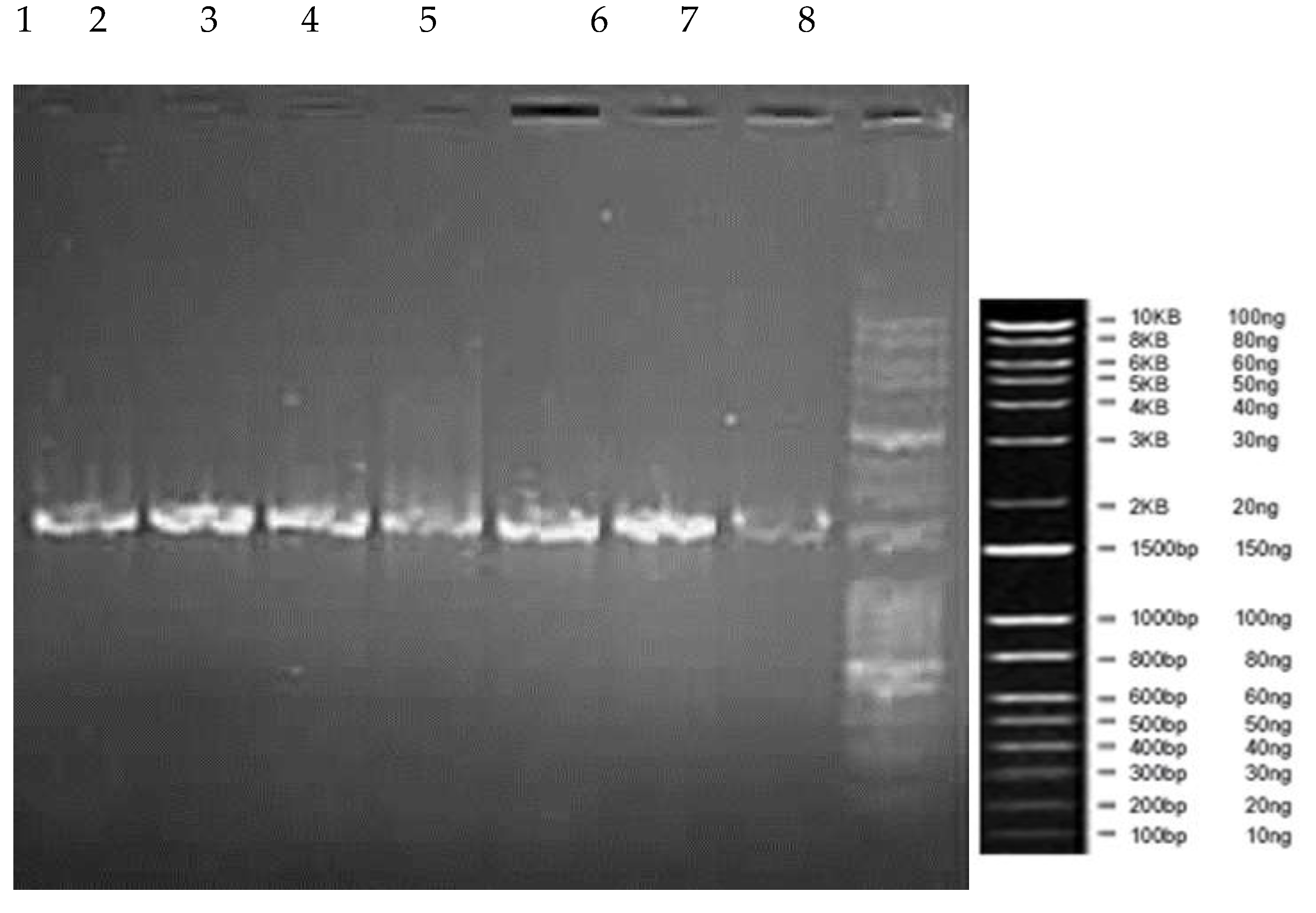
The base pair length of amplified DNA for all the isolates was in the similar level which stay at the level of 1500 Bp of DNA ladder.
The length of amplification of the 16SrDNA region depends on the nucleotide sequence. Generally, the length of 16S rDNA of fragments obtained from different lactobacilli species was approximately 1500 bp [24]. The PCR of the 16S rRNA genes of Lactobacillus salivarus from the blood culture and gall bladder pus showed bands at 1,515 bp [25] and the length of amplified product for Lactobacillus helvelictus and Lactobacillus acidophilus isolated from breast milk varied from 1500-2000 bp [14].
3.3.3. Identification of LAB based on phylogenetic analyses of 16S rDNA sequences
The seven rod and one coccus presumptively identified as LAB were subjected to 16S rDNA sequence analyses. The microorganism was deposited in Gene Bank (NCBI) and gene accession number were obtained (Table 5).
Based on the closely related strain from BLAST, isolated LAB were identified to species level (Table 4) including the four main genera of lactic acid bacteria from fermented rice such as Latilactobacillus sp, Limosilactobacillus , Weissella confusa and Pediococcus pentsaceus (Table 4). The four isolates Lb-1, Lb-2, Lb-10 and Lb-11 showed similar sequences to Latilactobacillus curvatus which was the dominant Lactobacillus sp. among the studied isolates. Other Lactobacillus sp. such as Latilactobacillus graminis and Limosilactobacillus fermentum were also identified from fermented rice.
3.3.4. Phytogenic tree development
The phylogenetic tree shows a relationship among the LAB species (Figure 5 and Figure 6) isolated from fermented rice. The isolates Latilactobacillus curvatus strains GRLb2, GRLb10 and GRLb11 shows 98% homology among them. The Pediococcus pentosaceus strain GRLc1 shows 74 % identity with Latilactobacillus graminis GRLb8. Weissella confusa strain GRLb4 shows 99% phylogenetic relationship with Latilactobacillus curvatus strains GRLb1 and Limosilactobacillus fermentum GRLb17. Latilactobacillus curvatus strains GRLb1 shows 55% identity with Limosilactobacillus fermentum GRLb17. Microbial community in the phylogenic tree is varied for different isolation sources, The phylogenic tree of isolated microorganisms from fermented teff dough showed that Lactobacillus and Enterococcus sp were the predominant microorganisms involved in the fermentation of teff dough [26]. Current study proved that genus Limosilactobacillus, Latilactobacillus, Weissella and Pedicoccus are the predominant microorganisms of fermented rice.
Figure 5.
The phylogenetic trees (a, b, c, d, e) for each isolate (Latilactobacillus curvatus strains GRLb1 GRLb2, GRLb10 and GRLb11, Latilactobacillus graminis GRLb8, Limosilactobacillus fermentum GRLb17, Weissella confusa strain GRLb4, Pediococcus pentosaceus strain GRLc1) which were constructed using the neighbour-joining method in MEGA 11. The gene sequences of mostly related stains from Gene Bank (NCBI) were used to construct the phylogenetic trees.
Figure 5.
The phylogenetic trees (a, b, c, d, e) for each isolate (Latilactobacillus curvatus strains GRLb1 GRLb2, GRLb10 and GRLb11, Latilactobacillus graminis GRLb8, Limosilactobacillus fermentum GRLb17, Weissella confusa strain GRLb4, Pediococcus pentosaceus strain GRLc1) which were constructed using the neighbour-joining method in MEGA 11. The gene sequences of mostly related stains from Gene Bank (NCBI) were used to construct the phylogenetic trees.

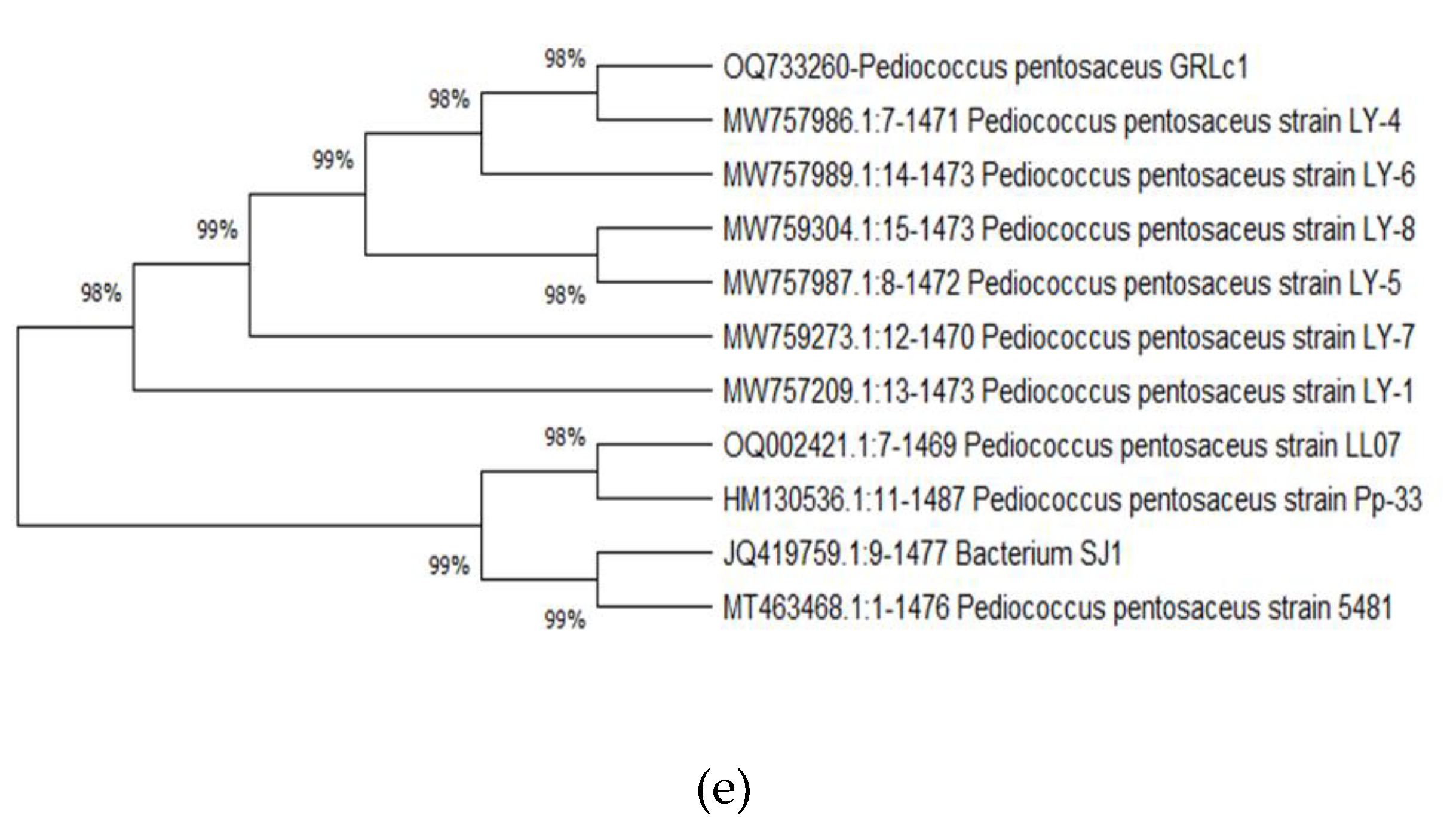
Figure 6.
Phylogenic tree of the isolates from fermented rice. The tree was constructed based on 16S rDNA gene sequences by using the neighbour-joining method in MEGA 11 software.
Figure 6.
Phylogenic tree of the isolates from fermented rice. The tree was constructed based on 16S rDNA gene sequences by using the neighbour-joining method in MEGA 11 software.

4. Discussion
Lactic acid bacteria (LAB) are well known for their ability for synthesizing a wide range of metabolites that beneficially affect the nutritional, sensorial, and technological properties of fermented food products. Among the studied isolates obtained from fermented rice, seven isolates appeared as rod, and one as spherical in shape found as cluster. All isolates were Gram positive and catalase negative and seven rod-shaped isolates were non-motile without spore forming. Lactobacillus appeared as rod-shaped cells in short chains while Pediococcus has spherical cells in pairs or tetrads. The morphological appearances of Lactobacillus and Pediococcus were rod and spherical shape, respectively. Seven rod shaped isolates and coccus could be belonged to the genus Lactobacillus and Pediococcus respectively as described by [27].
The phenotypic identification of microorganisms is important in the isolation and screening of lactic acid bacteria (LAB) with potential probiotic properties from new sources. Lactobacillus sp. are frequently found in vegetation, dairy products and the human intestine where it forms part of the normal flora of the gut [13]. Phenotypic characteristics of LAB can be used for identification at the genus level. The genus Lactobacillus can be confirmed with certain biochemical tests and Gram staining. Lactobacillus species are Gram-positive, catalase negative, non-spore-forming and non-motile rods (ranging from coccobacilli to long, slender bacilli) [28]. The physiological characteristics and fermentation of different type of sugars are used in the classification of Lactobacillus into groups, however, identification at species level is time-consuming, laborious and difficult to interpret by the conventional phenotypic methods [29]. Identification of Lactobacillus to species level has been unreliable only based on physiological biochemical and sugar fermentation tests [15]. The use of molecular-based techniques offers a rapid and species or strain specific alternative for identification [29].
Identification of Lactobacillus to the species level based on carbohydrate fermentation patterns using API50 CHL kit is not a reliable method due to the similar nutrition requirement of lactobacilli. In a previous study the carbohydrate fermentation pattern analysis, of the two strains, L.amylovorus DSM20531 and L. sobrius DSM16698 were identified as L. crispatus while Lactobacillus reuteri DSM 20016 was identified as L. fermentum1[30]. PCR based molecular techniques are precise and operative to identify the species of lactic acid bacteria [31]. Species-specific PCR primers that target the 16S-23S rRNA spacer region are available for a limited number of Lactobacillus species. The 16S rRNA gene is used for phylogenetic studies as it is highly conserved between different species of bacteria and archaea [16]. The accurate identification of Lactobacillus species can be accomplished by reference to 16S rRNA gene sequences and has been successful in the identification of different species of lactobacilli including L. plantarum, L.pentosus, and L fermentum from different sources such as the honey stomach of honey bee [32]. Furthermore, Lactobacillus species including L. gastricus, L.santri, L.kalixensis and L.ultunensis isolated from human stomach mucosa [33], Lactobacillus plantarum, L. rhamnosus, L. fermentum. L. pentosus from goat milk [15], L. nodensis from rice bran [34], L. plantarum and L. fermentum from fermented mustard (Susilini et al., [17]) L saerimneri from pig faeces [35] and L. casei NRC AM2, L.s rhamnosus NRC AM6, P. pentosaceus NRC AM4 and P. acidilactici NRC AM8 from fermented dairy product [31] have been successfully identified at molecular level using 16S rRNA gene sequences. The molecular identification method is more accurate and confirmative in identification of Lactobacillus sp. compared with other biochemical identification methods [36]. Therefore, the isolated Lactobacillus sp. were confirmed by16S r DNA gene sequencing.
In the current study isolates Lb-4, Lb-8, Lb-10 and Lb-17 were not identified as similar species by the biochemical identification using API 50 Kits and 16SrDNA method. Isolates Lb-1,2,11 and Lc-1 were identified as similar species as Lactobailus curvatus and Pediococcus pentosaceus by both identification methods. Lacti acid bacteria belongs to genus Lactobacillus were identified as L. curvats, L. delbrueckii, L. helveticus, L. pentosus and L. plantarum and Pediococcus pentoceous using the API 50CHL system. While, molecular identification by 16SrDNS confirmed the five lactic acid bacteria as Weissella confusa, Pediococcus pentosaceus, Latilactobacillus curvatus Latilactobacillus graminis and Limosilactobacillus fermentum from fermented rice, Latilactobacillus curvatus was the dominant Lactobacillus sp from fermented rice (Table 3 and Table 4).
Previous studies show that similar species isolated from fermented food have probiotic characteristics. Twenty-five strains of lactic acid bacteria (LAB) have been isolated from South Indian traditional fermented foods including “Kallappam” batter, “Koozh” and “Mor Kuzhambu”. Six strains were identified with higher antimicrobial activity among the twenty-five strains [37]. Lactobacillus fermentum which was isolated from Thailand traditional fermented food products (fermented pork, fermented fish, fermented tea leaves, and pickled garlic) showed extremely high survival rates in acid or bile salts, inhibited pathogenic bacteria and were sensitive to antibiotics [38]. Lactobacillus species isolated from fermented rice showed antimicrobial activity against pathogenic bacteria including Salmonella thyphi, S. enteritidis, S. thyphimurium, Escherichia coli (ATCC 259222), Shigella sonnei, S. flexneri and Candida spp. [39]. Lactobacillus curvatus A61 inhibited the growth of Listeria monocytogenes and Bacillus cereus strains and has shown antifungal activity against Cladosporium and Fusarium sp. [40]. Weissella confusa C3-7 shows antifungal activity against Penicillium roqueforti, Aspergillus niger and Endomyces fibuliger in contaminated bakery products [41]. Pediococcus pentoceus was the dominant isolate found in fermented cooked white rice and cococonut milk [42]. The strains of Pediococcus pentosaceus (MY-800, NS75, KC007, SH 740) isolated from Indian fermented food possessed probiotic characteristics [43]. L. fermentum FTL2311 and L. fermentum FTL10BR isolated from fermented tea leaves were probiotics with satisfactory acid and bile tolerance and antimicrobial activity [38]. Lactobacillus curvatus HY7602 added Fermented Antler (FA) is a functional food which effectively controls muscle atrophy caused by aging and considered as novel alternative treatment for sarcopenia [44]. The review study by Ying Chen et. al. [45] shows that Latilactobacillus curvatus is a probiotic with excellent fermentation properties and offer health benefits. LAB isolated from ‘idli’ batter [46] and Lactobacillus species isolated from fermented buffalo milk gel demonstrated promising probiotic potential with antibiotic sensitivity, antimicrobial properties, bile acid tolerance and acid tolerance [47]. As per the above review, the similar species of studied isolates were identified as probiotic and used as stater culture for the production of fermented food. Thus, the microbial community from fermented rice may possess probiotic properties and fermented food may considered as functional food. Probiotic characterization of the species identified in the present study warrants further studies.
5. Conclusions
Five species of lactic acid bacteria including L. curvatus, L. graminis and L. fermentum, W. confusa and P. pentosaceus were identified by 16SrDNA molecular technology. L. curvatus was the dominant Lactobacillus sp. among the studied isolates from fermented rice. Most of the isolates showed homofermentative characteristics except the isolate Lb-8 which showed hetero-fermentation. Isolates are able to grow at temperatures ranging between 10 °C and 45 °C, tolerate NaCl concentrations from 2%, to 6.5, and coagulate the milk. Fermented rice may offer health benefits due to the presence of identified LAB. The favorable physiological characteristics of these isolates confirm their potential application in the food industry. However, further studies are required to confirm the probiotic properties of these isolates.
Author Contributions
Conceptualization, N.J., A.G., T.M. and O.M.; Data curation, N.J. and T.M; Formal analysis, N.J. and A.G.; Investigation, N.J., and C.S.R.; Software, N.J.A.G., and O.M.; Methodology, N.J. A.G., T.M. and O.M.; Validation, N.J., O.M., and T.M.; Writing - original draft preparation. N.J.; Writing Review and editing C.S.R., M.Y.M., A.G., O.M. and T.M.; Supervision, C.S.R.., M.Y.M., and T.M.; Resources, C.S.R., M.Y.M., A.G. and T.M.
Funding
“This research was funded by HETC (Higher Education for twentieth Century projects) grant number ref: JFN/Vav-C/N1” to conduct the research for Doctor of Philosophy which is allocated for staff higher education for the University.
Data Availability Statement
The data presented in this study are available upon request.
Conflicts of Interest
The authors declare that the research was conducted in the absence of anycommercial or financial relationships that could be construed as a potential conflict of interest.
References
- Salovaara, H.; Simson, L. Fermented Cereal Based Functional Foods. In Hand Book of Food and Beverage Fermentation Technology; Hui, Y. H., Meunier-Goddik, L., Josephsen, J., Nip, W., Stanfield, P.S., Eds.; CRC press: Marcel Dekker, New York, 2004; p. 831. [Google Scholar]
- Blandinob, A.; Al-Aseeria, M.E.; Pandiellaa, S.S.; Canterob, D.; Webba, C. Review- Cereal-based fermented foods and beverages. Food Res Intern 2003, 36, 27–543. https://comenius.susqu.edu/biol/312/cerealbasedfermentedfoodsandbeverages.pdf.
- Jagadeeswari, S.; Vidya, P.; MukeshKumar, D.J.; Balakumaran, M.D. Isolation and Characterization of Bacteriocin producing Lactobacillus sp from Traditional Fermented Foods. Elect J Environ Agri Food 2010, 9, 575–581. https://www.cabdirect.org/cabdirect/abstract/20103323984.
- Dahiya, D.; Nigam, P.S. Use of Characterized Microorganisms in Fermentation of Non-Dairy-Based Substrates to Produce Probiotic Food for Gut-Health and Nutrition. Fermentation. 2023, 9, 1–14. [Google Scholar] [CrossRef]
- Heller, K.J. Probiotic bacteria in fermented foods: product characteristics and starter organisms. Am J Clin Nutr 2001, 73, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Kauli, C.M.; Mathara, J.M.; Kutima, P.M. Probiotic potential of spontaneously fermented cereal-based foods – A review. Afr J Biotechnol 2010, 9, 2490–2498. Available online: https://www.ajol.info/index.php/ajb/article/view/79704.
- Kumar, P.; Begum, V.H.; Kumaravel, S. Mineral nutrients of ‘pazhaya sadham’: A traditional fermented food of Tamil Nadu, India. Intern J Nutr Metab 2012, 4, 151–152. [Google Scholar] [CrossRef]
- Kowsalya, M.; Sudha, K.G.; Ali1, S.; Velmurugan, T.; Karunakaran, G.; Rajeshkumar, M.P. In-vitro assessment of probiotic properties of lactic acid bacteria isolated from naturally fermented rice gruel of south India. J Microbiol Biotechnol Food Sci 2022, 12, 1–9. [Google Scholar] [CrossRef]
- Wikeramanayake, T.W. Legumes. In Food and Nutrition. 2nd edition. Hector Kobbekaduwa Agrarian Training Institute: Colombo, Sri Lanka, 2002; ISBN: 955-565-000-4,168.
- Hatti-Kaul, R.; Chen, L.; Dishisha, T.; Enshasy, H.E. Lactic acid bacteria: from starter cultures to producers of chemicals. FEMS Microbiol Lett. 2018, 365(I20), 1–20. [Google Scholar] [CrossRef]
- Axelsson, L. Lactic Acid Bacteria Classification and Physiology in Lactic Acid Bacteria Microbiological and Functional Aspects, 3rded.; Salminen, S., Wright, A.V., Ouwehand, A., Eds.; Marcel Dekker: New York, 2004; pp. 1–67. [Google Scholar] [CrossRef]
- Jeyagowri, N.; Parahitiyawa, N.B.; Jeyatilake, J.A.M.S.; Ranadheera, C.S.; Madhujith, M.M.T. Study on Isolation of potentially probiotic Lactobacillus spp from fermented rice. Tropic Agri Res 2015, 26, 428–440. [Google Scholar] [CrossRef]
- Barrow, G.H.; Feltham, R.K.A. Cowan and Steel’s Manual for Identification of Medical Bacteria, 3rd ed.; Cambridge University Press: Cambridge, 1993; p. 331. [Google Scholar] [CrossRef]
- Hatice, Y.; Isolation, Characterization and determination of probiotic properties of lactic acid bacteria from human milk. Thesis for Master of Science. Graduate School of Engineering and Sciences of Izmir Institute of Technology, Turkey. 2007. Available online: https://www.academia.edu/3341345/ (accessed on 16 October 2011).
- Marrokil, A.; Zuniga, M.; Kihal, M.; Martinez, G. Characterization of Lactobacillus from Algerian goats’s milk based on phenotypic, 16SrDNA Sequencing and their technological properties. Braz J Microbiol 2011, 42, 58–171. [Google Scholar]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 1. 16S ribosomal DNA amplification for phylogenetic study. J Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Sulistiani,; Abinawanto,;Sukara, E.; Salamah, A.;Dinoto, A.; Mangunwardoyo, W. Identification of lactic acid bacteria in sayurasin from Central Java (Indonesia) based on 16S rDNA sequence. Intern Food Res J 2014, 21, 527–532.
- Hall, B.G. Building Phylogenetic Trees from Molecular Data with MEGA. Mol Biol Evol 2013, 30, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Vaccalluzzo, A.; Pino., A.; Angelis., M.D.; Bautista-Gallego, J.; Romeo, F.V.; Foti, P.; Caggia, C.; Randazzo, C.L. Effects of Different Stress Parameters on Growth and on Oleuropein-Degrading Abilities of Lactobacillus plantarum Strains Selected as Tailored Starter Cultures for Naturally Table Olives. Microorganisms 2020, 8, 1607. [Google Scholar] [CrossRef] [PubMed]
- Kunji, E.R.S.; Mierau, I.A.; Poolman, B.; Konings, W.N. The proteolytic systems of lactic acid bacteria. Antonie Leeuwenhoek 1996, 70, 187–221. [Google Scholar] [CrossRef]
- Wang, H.; Yu, W.; Coolbear, T.; Sullivan, D.; Mckay, L.L. A deficiency in aspartate biosynthesis in Lactococcus lactis subsp. lactis C2 causes slow milk coagulation. Appl Environ Microbiol 1998, 64, 1673. [Google Scholar] [PubMed]
- Quere, F.; Deschamps, A.; Urdaci, M.C. DNA probe and PCR specific reaction for Lactobacillus plantarum. J Appl Microbiol 1997, 82, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Ahrne, S.; Molin, G.; Stahl, S. Plasmids in Lactobacillus strains isolated from meat and meat products. System Appl Microbiol 1989, 11, 320–325. [Google Scholar] [CrossRef]
- Hamid, M.; Abulfazl. Isolation and Molecular Study of Potentially Probiotic Lactobacilli in Traditional White Cheese of Tabriz in Iran. Annals of Biological Research 2012, 3, 2019–2022. [PubMed]
- Patrick, C.Y.; Woo, A.M.Y.; Fung.; Susanna, K.P.L.; Yuen, K.Y. Identification by 16S rRNA Gene Sequencing of Lactobacillus salivarius Bacteremic cholecystitis. J Clin Microbiol 2002, 40, 265–267.
- Tilahun, B.; Testaye, A.; Muleta, D; Baihru, A.; Terefework, Z.; Wessel G. Isolation and Molecular Identification of Lactic acid Bacteria using 16srRNA from fermented Teff (Eragrostis tef (Zucc.)) Dough. Intern J Food Sci 2018. [PubMed]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A.; Staley, J.T.; Williams, S.T. Bergey's Manual of Determinative Bacteriology, 9th ed.; Williams and Wilkins: Baltimore, 1994; pp. 505–542. [Google Scholar]
- Hoque, M.Z.; Akter, F.; Hossain, K.M.; Rahman, M.S.M.; Billah, M.M.; Islam, K.M.D. Isolation, Identification and Analysis of Probiotic Properties of Lactobacillus Sp. From Selective Regional Yoghurts. World J Dairy Food Sci 2010, 5, 39–46. [Google Scholar]
- Mohammed, M.; Abd El-Aziz, H.; Omran, N.; Anwar, S.; Awad, S.; El-Soda, M. Rep-PCR characterization and biochemical selection of lactic acid bacteria isolated from the Delta area of Egypt. Intern J Food Microbiol 2009, 128, 417–423. [Google Scholar] [CrossRef]
- Khunajakr, N.; Wongwicharn, W.; Moonmangmee, D. Screening and identification of lactic acid bacteria producing antimicrobial compounds from Pig gastrointestinal Tracts. KMITL Sci Technol J 2008, 8, 10. [Google Scholar]
- Hassan, Z.; Effat, B.; Magdoub, M.; Tawfik, N.; Sadek, Z.; Mabrouk, A. Molecular Identification of lactic bacteria isolated from fermented dairy product. Intern J Biol Pharm Allied Health Sci 2016, 5, 3221–3230. [Google Scholar]
- Tajabadi, N.; Mardan, M.; Nazamid, S.; Shuhaimi, M.; Bahreini, R.; Manap, M.Y.A. Identification of Lactobacillus plantarum, Lactobacillus pentosus and Lactobacillus fermentum from honey stomach of honeybee. Braz J Microbiol 2013, 44, 717–722. [Google Scholar] [CrossRef]
- Roos, S.; Engstrand, L.; Jonsson, H. Lactobacillus gastricus sp. nov., Lactobacillus antri sp. nov., Lactobacillus kalixensis sp. nov. and Lactobacillus ultunensis sp. nov., isolated from human stomach mucosa. Intern J Syst Evol Microbiol 2005, 55, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Kashiwagi, T.; Suzuki, T.; Kamakura, T. Lactobacillus nodensis sp. nov., isolated from rice bran. Intern J Syst Evol Microbiol 2009, 59, 83–86. [Google Scholar] [CrossRef]
- Pedersen, C.; Roos, S. Lactobacillus saerimneri sp. nov isolated from pig faces. Intern J Syst Evol Microbiol 2004, 54, 1365–1368. [Google Scholar] [CrossRef] [PubMed]
- Brolazo, E.M.; Leite, D.S.; Tiba, M.R.; Villarroel, M.; Marconi, C.; Simoes, J.A. Correlation between API 50CH and Multiple Polymerase Chain Reaction for the identification of Vaginal Lactobacillus in Isolates. Braz J Microbiol 2011, 42, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Satishkumar, R.; Ragu-Varman, R.; Kanmani, P.N.; Yuvaraj, N.; Paari, K.A.; Pattukumar, V.; Arul, V. Isolation, Characterization and Identification of a Potential Probiont from South Indian Fermented Foods (Kallappam, Koozh and MorKuzhambu) and Its Use as Biopreservative. Prob Antimicrob Protein 2010, 2, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Srikanjana, K.; Siriporn, O. Antibacterial and Antioxidant actives of Acid and bile resistant strains of Lactobacillus fermentum isolated from Miang. Braz J Microbio 2009, 40, 757–766. [Google Scholar]
- Jeyagowri, N.; Parahitiyawa, N.B.; Jeyatilake, J.A.M.S.; Ranadheera, C.S.; Madhujith, W.M.T. Antimicrobial activity of potentially probiotic Lactobacillus spp from fermented rice. Proceedings Peradeniya University International Research Sessions (iPURSE 2014) Peradeniya. Sri Lanka, 4–5 July 2014; University of Peradeniya, Sri lanka, 18, p. 182. https://www.pdn.ac.lk/ipurse/2014/proceeding_book/FL/210.pdf.
- Ahmadova, A.; Todorov, S.D.; Hadji-Sfaxi, I.; Choiset, Y.; Rabesona, H.; Messaoudi, S.; Kuliyev, A.; Franco, B.D.; Chobert, J.M.; Haertle, T. Antimicrobial and antifungal activities of Lactobacillus curvatus strain isolated from homemade Azerbaijani cheese. Anaerobe. 2013, 20, 42–49. [Google Scholar] [CrossRef]
- Valerio, F.; Favilla, M.; De Bellis, P.; Sisto, A.; Valerio, F. Antifungal activity of strains of lactic acid bacteria isolated from a semolina ecosystem against Penicillium roqueforti, Aspergillus niger and Endomyces fibuliger contaminating bakery products. Systemic Appl Microbiol 2013, 32, 438–448. [Google Scholar] [CrossRef]
- Divisekera, D.M.W.D.; Gooneratne, J.; Jayawardana, D. Isolation and characterization and identification of Lactic acid bacteria and yeast from fermented organically grown rice and coconut milk. In: Proceedings Peradeniya University International Research Sessions (iPURSE 2014) Peradeniya. Sri Lanka, 4–5 July 2014; University of peradeniya, Sri lanka,.18, pp. 210. https://www.pdn.ac.lk/ipurse/2014/proceeding_book/FL/210.pdf.
- Sukumar, G.; Ghosh, A.R. Pediococcus spp. – A potential probiotic isolated from Khadi (an Indian fermented food) and identified by 16SrDNA sequence analysis. Afr J Food Sci 2010, 4, 597–602. [Google Scholar]
- Jeon, H.; Kim, Y.-T.; Jang, W.Y.; Kim, J.-Y.; Heo, K.; Shim, J.-J.; Lee, J.-L.; Yang, D.-C.; Kang, S.C. Effects of Lactobacillus curvatus HY7602-Fermented Antlers in Dexamethasone-Induced Muscle Atrophy. Fermentation 2022, 8, 454. [Google Scholar] [CrossRef]
- Chen, Y.; Yu, L.; Qiao,; Xiao,; Tian,; Zhao, J.; Zhang,H.; Chen,W.; Zhai.O. Latilactobacillus curvatus: A Candidate Probiotic with Excellent Fermentation Properties and Health Benefits. Foods 2020, 9, 1336. Available online: https://pubmed.ncbi.nlm.nih.gov/32993033/.
- Bernard, D.; Jeyagowri, N.; Madhujith, T. Characterization of Lactic Acid Bacteria isolated from Idli Batter and their susceptibility to Antibiotics. Tropical Agricultural Research 2021, 32, 453–461. [Google Scholar] [CrossRef]
- Adikari, A.M.M.U.; Priyashantha, H.; Disanayaka, J.N.K.; Jayatileka, D.V.; Kodithuwakku, S.P.; Jayatilake, J.A.M.S.; Vidanarachchi, J.K. Isolation, identification and characterization of Lactobacillus species diversity from Meekiri: traditional fermented buffalo milk gels in Sri Lanka. Heliyon 2021, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]

Table 1.
Morphological characteristics of isolated Lactic Acid Bacteria from fermented rice.
| Isolates | Morphological description | Appearance under light microscope (x1000 magnification) |
|---|---|---|
| Latilactobacillus curvatus strain GRLb1 | Medium size, Rod (regular)shaped bacteria, arranged as single /pair or group in ‘V’ arrangement | |
| Latilactobacillus curvatus strain GRLb2 | Medium size, Rod (regular)shaped bacteria, arranged as single /pair or group in ‘V’ arrangement | |
| Weissella confusa strain GRLb4 | Very long size, Rod (regular)shaped bacteria, arranged as single /pair or group in ‘V’ arrangement | |
| Latilactobacillusgraminis strain GRLb8 | Small size, Rod (regular)shaped bacteria, arranged as single /pair or group in ‘V’ arrangement | |
| Latilactobacillus curvatus strain GRLb10 | Medium size, Rod (regular)shaped bacteria, arranged as single /pair or group in ‘V’ arrangement | |
| Latilactobacillus curvatus strain GRLb11 | Medium size, Rod (regular)shaped bacteria, arranged as single /pair or group in V arrangement | |
| Limosilactobacillusfermentum strain GRLb17 | Long size, Rod (regular)shaped bacteria, arranged as single /pair or group in V arrangement | |
| Pediococcus pentosaceus strain GRLc1 | Coccus shaped bacteria, arranged as single, tetrad, group/ cluster |
Table 2.
Physiological and biochemical characterization of the isolates.
| Physiological and biochemical characteristics | Lb-1 | Lb-2 | Lb-4 | Lb-8 | Lb-10 | Lb-11 | Lb-17 | Positive | Negative |
|---|---|---|---|---|---|---|---|---|---|
| Gram test | + | + | + | + | + | + | + | NA | NA |
| Catalase test | - | - | - | - | - | - | - | NA | NA |
| Motility | - | - | - | - | - | - | - | NA | NA |
| Spore formation | - | - | - | - | - | - | - | NA | NA |
| Gas from glucose | - | - | - | + | - | - | - | ND | - |
| Growth at 10°C | ++ | ++ | ++ | + | ++ | ++ | ++ | NA | - |
| Growth at 37°C | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | - |
| Growth at 40°C | ++ | ++ | ++ | ++ | ++ | ++ | ++ | NA | - |
| Growth at 45°C | + | + | + | + | + | + | + | NA | - |
| Growth at 55°C | - | - | - | - | - | - | - | NA | - |
| 0% Nacl | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | |
| 2% NaCl | ++ | ++ | ++ | ++ | ++ | ++ | ++ | NA | - |
| 4% NaCl | ++ | ++ | ++ | ++ | ++ | ++ | ++ | NA | - |
| 6.5% NaCl | ++ | ++ | ++ | ++ | ++ | ++ | ++ | NA | - |
| 10% NaCl | - | - | - | - | - | - | - | NA | - |
| Milk coagulation | + | + | + | + | + | + | + | + | - |
NA- Not applicable, ND-Not determined, ++: heavy growth.
Table 3.
Identification of Lactic acid bacteria using API 50 Kits.
| Isolate code | Species of Lactobacillus | *Identity (%) |
|---|---|---|
| Lb-1 | Lactobacillus curvatus ssp.curvatus | 98.9 |
| Lb-2 | Lactobacillus curvatus ssp.curvatus | 99.3 |
| Lb-4 | Lactobacillus helveticus | 86.3 |
| Lb-8 | Lactobacillus delbrueckii ssp. delbrueckii | 95.5 |
| Lb-10 | Lactobacillus pentosus | 63.3 |
| Lb-11 | Lactobacillus curvatus ssp.curvatus | 99.3 |
| Lb-17 | Lactobacillus plantrum | 91.3 |
| Lc-1 | Pediococcus pentosaceus | 99.9 |
* Identity percentage of bacteria using api web system.
Table 4.
Identification of isolated LAB using16S rDNA sequences.
| Sequence ID | Microorganism | Accession Number | Sequence length |
|---|---|---|---|
| Lb1 | Latilactobacillus curvatus GRLb1 | OQ733261 | 1475 bp |
| Lb2 | Latilactobacillus curvatus GRLb2 | OQ733262 | 1465 bp |
| Lb4 | Weissella confusa strain GRLb4 | OQ733263 | 1486 bp |
| Lb8 | Latilactobacillus graminis GRLb8 | OQ861079 | 1097 bp |
| Lb10 | Latilactobacillus curvatus GRLb10 | OQ733264 | 1469 bp |
| Lb11 | Latilactobacillus curvatus GRLb11 | OQ733265 | 1456 bp |
| Lb17 | Limosilactobacillus fermentum GRLb17 | OQ861078 | 819 bp |
| Lc1 | Pediococcus pentosaceus GRLc1 | OQ733260 | 1480 bp |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



