
Submitted:
26 July 2023
Posted:
27 July 2023
You are already at the latest version
Abstract
Sclerotinia sclerotiorum is a devastating fungal pathogen that causes severe crop losses worldwide. It is of vital importance to understand its pathogenic mechanism for disease control. Through a forward genetic screen combined with next-generation sequencing, a putative protein kinase, SsCak1, was found to be involved in growth and pathogenicity of S. sclerotiorum. Knockout and complementation experiments confirmed that deletions in SsCAK1 caused defects in mycelium and sclerotia development, as well as appressoria formation and host penetration, leading to complete loss of virulence. These findings suggest that SsCak1 is essential for growth, development and pathogenicity of S. sclerotiorum. Therefore, SsCak1 could serve as a potential target for the control of S. sclerotiorum infection through host-induced gene silencing (HIGS), which could increase crop resistance to the pathogen.
Keywords:
Sclerotinia sclerotiorum
; SsCak1
; sclerotia
; appressoria
; pathogenesis
; HIGS
1. Introduction
Sclerotinia sclerotiorum is a phytopathogenic fungus which has a wide host range and can infect more than 600 species of plants, including rape, potato, cotton, tomato, soybean and other important crops [1,2,3]. S. sclerotiorum causes stem rot, resulting in the death and decay of host tissues. Its virulence involves the release of toxins (such as oxalic acid (OA) and cell wall-degrading enzymes (CWDEs) for infection initiation, followed by the extraction of nutrients from host cells [4]. As a necrotrophic pathogen, S. sclerotiorum, however, has a brief biotrophic phase that begins approximately 12-24 hours after infection [3,5]. During this stage, S. sclerotiorum establishes compatibility with the host by suppressing or disrupting its defense barriers [6]. Subsequently, the subcutaneous hyphae of S. sclerotiorum spread to multiple cell layers. With successful colonization by branched hyphae, S. sclerotiorum enters a necrotrophic phase, producing large amounts of reactive oxygen species, toxins, and CWDEs, leading to the development of host cell death and necrotic symptoms [5,7,8].
The complex appressorium is a key infection structure for establishing infection. It is formed by hyphae that undergo swelling, slow growth, and continuous branching. Previous studies have shown that these cells can enhance pathogen's adhesion to host surface [9], and helps penetrate the host epidermal barrier through mechanical pressure and/or enzymatic degradation [10]. The hyphae forming complex appressoria often become flattened and increase in diameter, from which narrow penetration pegs are formed to complete penetration [9,11]. Subsequently, subcutaneous infection hyphae are produced and differentiated. These hyphae grow horizontally beneath the epidermis, forming the colonization leading edge [3].
To date, host induced gene silencing (HIGS) has become a promising way to control fungal diseases, including sclerotinia stem rot. Andrade et al. [12] demonstrated for the first time that HIGS-mediated chitin synthase gene (CHS) enhanced T1 generation resistance to S. sclerotiorum in tobacco. In subsequent independent studies, by selecting Sscnd1, Ssoah1, ABHYRDOLASE-3 and SsTrxR1 as the target genes of HIGS, the resistance of plants to S. sclerotiorum was successfully improved [6,13,14,15]. Therefore, it is possible to achieve effective control of Sclerotinia stem rot (SSR) by conducting in-depth research on the pathogenic mechanisms of the S. sclerotiorum and screening their key genes, and enhancing the host's resistance to the pathogens through cross-kingdom RNA silencing.
Recently, Xu et al. developed a method combining forward genetic screening with high-throughput next-generation sequencing for rapid discovery of new genes involved in sclerotia development. Some of these genes can be used as HIGS target for disease control [16,17]. Here, we performed a genetic screen for virulence-related genes in a mutagenized population of S. sclerotiorum and isolated a mutant strain with defects in growth, sclerotia development and pathogenicity. Then we identified an important candidate gene, SsCak1, by NGS. Knockout and complementation experiments confirmed its involvement in the development of S. sclerotiorum mycelium, sclerotia, complex appressoria formation, and pathogenicity, making it a potential target for HIGS to enhance crop resistance against S. sclerotiorum.
2. Results
2.1. Identification of a Pathogenicity-Attenuated Mutant in S. sclerotiorum through Forward Genetic Screening
In order to study the pathogenic mechanism of S. sclerotiorum in depth, we carried out a forward genetics screen on a UV-mutagenized population of S. sclerotiorum [18]. Using leaves of lettuce (Figure S1), we aimed to identify mutants with virulence defects. Here we report on a mutant strain M14-9. On tobacco leaves, wild-type (WT) S. sclerotiorum caused severe leaf maceration 48 h after inoculation, whereas M14-9 did not (Figure 1A, B). Moreover, the M14-9 mutant exhibited markedly impaired growth (Figure 1C). The growth rate of M14-9 was much lower than that of WT (Figure 1D), and a large number of aerial hyphae were produced at the early stage of colony formation. In addition, the hyphae of the M14-9 appeared dense and short under the microscope (Figure 1E). M14-9 is deficient in sclerotia formation, with fewer sclerotia numbers per dish and less individual sclerotia weight than WT (Figure 1F-G). Thus, M14-9 shows dual defects in pathogenicity and growth.
2.2. Sscle_11g085070 Is the Candidate mutated Gene of M14-9
To identify the molecular lesion in M14-9, the whole genome of M14-9 was re-sequenced and analyzed by NGS. Gene sequences of two other mutants, 49-23 and A14-11, which were identified in the same screen and sequenced, were used as negative controls to exclude background mutations in M14-9. The NGS data were then analyzed with a modified NGS sequence analysis pipeline in S. sclerotiorum [19]. By removing background mutations, synonymous mutations, intron mutations, and intergenic mutations, three remaining SNP mutations became the main candidate mutations for subsequent analysis (Figure S2A).
In order to further narrow down the candidates, we analyzed the protein sequences encoded by these genes to examine the functions of their orthologs in closely related species. Literature research revealed that sscle_11g085070, whose ortholog, FgCak1, from Fusarium graminearum, is involved in hyphael development and pathogenicity [20]. Therefore, we first tested SsCak1 in S. sclerotiorum. Sanger sequencing of SsCak1 using DNA from M14-9 confirmed a single nucleotide mutation in the second exon of the gene, resulting in a premature stop codon, consistent with the NGS sequencing results (Figure S2B, C).
Furthermore, phylogenetic analysis results revealed that the homologous proteins of SsCak1 (protein accession number, APA13135.1 / XP001584936.1) are relatively conserved in fungi, and widely present in Botrytis cinerea and other pathogenic fungi (Figure 2A). All Cak1s have two conserved eukaryotic protein kinase domains, VIB and VIII (Figure 2B). According to the traditional classification method, SsCak1 belongs to the CMGG group of eukaryotic protein kinases. To gain preliminary insight into the role of SsCak1 in fungal development, RT-qPCR analysis was conducted to determine the abundance of SsCak1 mRNA in different growth stages of S. sclerotiorum. As shown in Figure 2C, SsCak1 was expressed constitutively in different developmental stages. However, the highest expression was observed in the mature sclerotium stage (7 dpi) (Figure 2C). When inoculated on leaves of Brassica napus, the expression of SsCak1 was significantly upregulated from 9 hpi to 24 hpi (Figure 2D), indicating that SsCak1 was strongly induced during S. sclerotiorum infection. These results suggest that SsCak1 may play an important role in growth, development and pathogenicity of S. sclerotiorum, and the mutated phenotype of M14-9 may be caused by the truncation of SsCak1.
2.3. SsCak1 Deletion Impairs Mycelial Growth and Sclerotia Development
To test the gene function of SsCak1, we generated a knockout mutant of Sscak1 in the background of WT strain by homologous recombination, and also obtained two independent complementation strains, Sscak1-C1 and Sscak1-C5, by introducing a WT copy of SsCak1 into the knockout mutant (Figure S3A, Figure 3A). PCR and RT-qPCR showed that SsCak1 was completely deleted in the knockout mutant, and its transcript was absent, while the two complementation strains restored the transcript levels (Figure S3B, Figure 3D). The mycelial morphology of Sscak1 was largely similar to that of M14-9, showing dense mycelial branching and slower growth rate (Figure 3B,E), while the complementation strains Sscak1-C1 and Sscak1-C5 regained the WT phenotypes. After 15 days of growth on PDA medium, the SsCak1 knockout mutant exhibited abnormal sclerotia development on the colony surface (Figure 3C). Although Sscak1 successfully formed sclerotia, the number of sclerotia produced per plate was significantly reduced (Figure S4A), and the average mass of each sclerotium was also lighter than that of WT (Figure S4B), indicating that loss of SsCak1 impairs growth of mycelium and sclerotia development in S. sclerotiorum.
2.4. Knockout of SsCak1 Results in Complete Loss of pathogenicity in S. sclerotiorum
For pathogenicity tests, we inoculated the WT, mutant M14-9, Sscak1, and complementation strains Sscak1-C1 and Sscak1-C5 onto intact and wounded leaves of Arabidopsis (Figure 4A) and N. benthamiana (Figure 4B). After 48 hours, on intact leaves, neither M14-9 nor Sscak1 was able to infect, whereas the complementation strains caused large areas of infection similar to that of WT (Figure 4C, D). On wounded leaves, M14-9 and Sscak1 caused mild infection damage compared to the WT and complementation strains (Figure 4C, D). These results confirmed that SsCak1 is essential for the pathogenicity of S. sclerotiorum, which likely involves both pre- and post-penetration regulations.
2.5. SsCak1 is essential for appressorium formation and penetration
To further investigate the details of the pathogenicity defects in Sscak1 mutants, we examined its oxalate production, appressoria development and penetration ability. First, fresh mycelium blocks were inoculated on the PDA medium containing bromophenol blue as pH indicator. After 24 hours, all areas of the mycelium inoculated, including WT, M14-9, Sscak1, Sscak1-C1, and Sscak1-C5, had turned from blue to yellow (Figure S5), suggesting that SsCak1 does not affect oxalate production. However, M14-9 and Sscak1 failed to produce appressorium on glass slides, while the WT strain formed mature appressorium cell structures upon contact with the slide (Figure 5A, B). Onion epidermal penetration assays further revealed that WT strain formed numerous appressoria and invasive hyphae, whereas M14-9 and Sscak1 failed to produce them and the hyphae were unable to penetrate the onion epidermis (Figure 5C). Therefore, SsCak1 is an important factor in appressoria development, and the defect in appressorium development and permeability is likely the main reason for the loss of pathogenicity in Sscak1.
2.6. HIGS of SsCak1 in N. benthamiana Enhances Resistance to S. sclerotiorum
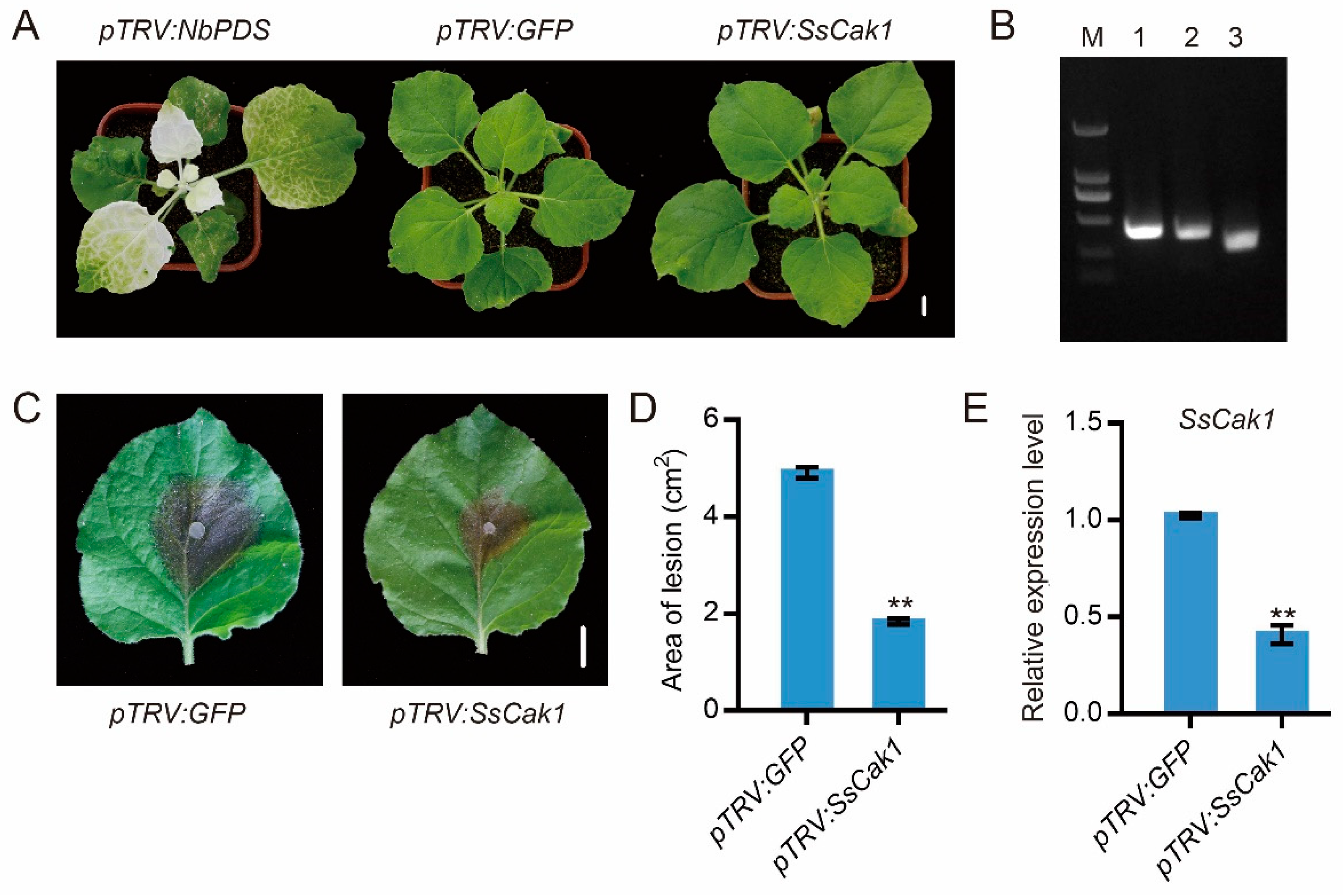
Conserved and functionally important genes can sometimes be used as target genes for the control of pathogenic microorganisms using HIGS. Since sequence alignment and phylogenetic tree analysis showed that there was no gene with similar sequence to SsCak1 in plants, SsCak1 may be a potential target for stem rot control using HIGS. Thus, we performed tobacco rattle virus (TRV)-mediated transient silencing of SsCak1 in N. benthamiana. Here, three agrobacterium constructs, including a positive control construct pTRV2::PDS, a negative control construct pTRV2::GFP and an experimental group pTRV2::SsCak1 construct, were agro-infiltrated into N. benthamiana leaves. After 7 days, the top leaves of N. benthamiana showed chlorosis with pTRV2::PDS infiltration, which gradually expanded to the entire plant (Figure 6A). At the same time, the expression of the target genes can be quantified by semi-quantitative (Figure 6B), demonstrating the feasibility of the gene silencing system. After 14 d TRV treatment, plants were inoculated with WT strain of S. sclerotiorum (Figure 6C). Compared with that in the control leaves (TRV::GFP), the lesion area on TRV::SsCak1 was reduced by 62% at 24 hpi (Figure 6D). Meanwhile, expression of SsCak1 in TRV::SsCak1 treated leaves was also reduced to 60% of control leaves (TRV::GFP) (Figure 6E). These results suggest that silencing of SsCak1 by TRV-HIGS does reduce virulence and enhance host resistance against S. sclerotiorum.
3. Discussion
Brassica napus is one of the world's most important oilseed crops. However, one of its common diseases, stem rot caused by S. sclerotiorum, causes significant damage, resulting in yield losses of up to 80% of the total disease damage and severely affecting B. napus production and quality [3,21]. In order to control this pathogen, various methods have been used, among which biological control has the advantages of safety and efficiency. Therefore, studying the growth and pathogenicity-related mechanisms of S. sclerotiorum can promote the development of new biological control methods. Previous studies have typically relied on homologous proteins identified from well-characterized proteins in other pathogenic fungi to study pathogenicity-related proteins in S. sclerotiorum. However, this research approach has limitations. With the development of sequencing technology and the revelation of the whole genome sequence of S. sclerotiorum, screening at the whole-genome level has become an efficient and innovative method for studying the growth, development, and pathogenicity mechanisms of S. sclerotiorum. In this study, we used forward genetic screening to obtain a stably inherited mutant, M14-9, which has dual defects in mycelial growth and pathogenicity. Then, we identified a putative CDK-activating kinase through an effective NGS-based method and conducted a series of experiments to explore its function.
Reversible phosphorylation is a major mechanism by which environmental and biochemical stimuli affect protein function and gene expression. Most eukaryotic protein kinases (ePKs) phosphorylate serine, threonine, or tyrosine and have a highly conserved catalytic domain consisting of 12 subdomains that make up the ATP-binding lobe (subdomains I-V) and peptide binding and phosphotransfer lobes (subdomains VI–XI). Subdomains VIB and VIII are involved in peptide substrate recognition, and conserved amino acids within them are used to classify ePKs into functional groups [22,23,24]. According to the multiple sequence alignment of homologues, Cak1 is relatively conserved in fungi and has a conserved eukaryotic kinase domain, which can be classified into CMGG groups according to their subdomains VIB and VIII. S. sclerotiorum ePKs CMGG group is widely produced during mycelium development, virulence formation, sclerotia development and host penetration. SMK1, another ePKs CMGG group by RNA-silencing showed impaired sclerotia formation in S. sclerotiorum, while in other pathogens inactivation of the SMK1 homologous gene resulted in loss of pathogenicity due to inability to form appressorium [25]. Disruption of the SMK3, an important ePKs of the CMGG group in S. sclerotiorum, results in an inability to aggregate and form appressorium leading to a severe reduction in virulence. The mutation also results in a loss of the ability to produce sclerotia, aerial bacteria, increased filaments and altered mycelial hydrophobicity [26]. In our study, SsCak1, which also belongs to a CMGG group, is a protein homologue of Cak1 in B. cinerea (Figure 2). Furthermore, by measuring the expression level of SsCak1 in different developmental stages and infection periods, the results showed that SsCak1 was expressed in all stages of mycelium development, especially at the time of sclerotia formation. And the expression level was the highest at 9-24 hours of infection, which is the critical period of infection. Thus, we speculated that SsCak1 may be involved in the fungal growth and cell cycle regulation of S.sclerotiorum. Indeed, Sscak1 knockout strains showed slowed down hyphal growth with a large number of aerial hyphae in the early stage and impaired sclerotia development, compared with WT strain (Figure 3).
As a cell cycle kinase, CDK-activating kinase (Cak) primarily exerts its activation mechanism through phosphorylation of a conserved threonine residue located in the T-loop region of CDK [27,28]. In the realm of yeasts, Saccharomyces cerevisiae harbors a vital Cak gene, denoted as Cak1, while Schizosaccharomyces pombe employs two partially redundant Cak systems, namely the MCS6-MCS2 complex and Csk1, to facilitate the activation of Cdc2 during cellular division [28,29,30]. Intriguingly, within S. sclerotiorum, the elimination of SsCak1 did not yield lethal consequences, as the resulting SsCak1 knockout mutants exhibited comparable deficiencies in growth and infection levels to the previously reported SsCDC28 mutants [31]. Consequently, this observation proposes that Cak1 might actively engage in the cell cycle progression of S. sclerotiorum by effectuating the phosphorylation of CDC28. Nonetheless, the veracity of this proposition necessitates further validation through subsequent experimental investigations. The association of cell cycle-regulated kinases with appressorium formation has been demonstrated in many important pathogenic fungi [32,33]. In some cases, specific cell cycle phases must be completed prior to attachment formation, as has been described in M. oryzae, where completion of the S and M phases is mandatory for attachment formation and function [33]. In other cases, however, the cell cycle must be stopped at a specific cell cycle stage to allow appressorium to form. In U. maydis, infectious filaments (where the attachment will differentiate) must stop in G2 phase [34]. In M. oryzae, lack of cyclin-dependent kinase CDC14 hinders appressorium generation [35]. Considering that the main role of the appressorium is to facilitate the penetration of the fungal hyphae within its host to proliferate after invading plant tissue, the formation of appressorium is subordinate to the regulation of the cell cycle. This affiliation seems reasonable to ensure that normal genetic information is loaded into the invading hyphae during this process. As expected, appressoria are completely absent in Sscak1 mutants, therefore we hypothesis that the loss of SsCak1 results in disordered cell cycle regulation, thereby affecting the formation of appressorium and leads to the completely loss of pathogenicity in S. sclerotiorum. Nevertheless, its function in other aspects of pathogenicity besides facilitating penetration cannot be excluded. As when inoculated on wounded A. thaliana and N. benthamiana leaves (with a dissecting needle), SsCak1 deletion mutants produced lesion spots, but the area was much smaller than that of the WT strain (Figure 4).
At present, the prevention and control of sclerotinia is almost entirely dependent on chemical fungicides, and the perennial use of medicaments has made sclerotinia produce obvious drug resistance [36]. Thus, breeding of disease-resistant varieties becomes more and more important for environmentally friendly control strategy. However, strong host monogenic resistance to S. sclerotiorum has not yet been found [2]. RNAi-based approach HIGS offers a flexible and environmentally friendly solution for crop protection [37,38,39]. Hence, HIGS technology provides a new generation of species-specific and environmentally safe method for the control of S. sclerotiorum. Here, we selected SsCak1 as the target gene and transiently expressed the RNAi construct of SsCak1 in N. benthamiana using HIGS, and the results showed that HIGS-mediated silencing of the SsCak1 significantly increased the resistance of plants to S. sclerotiorum. Although, a limitation of RNAi technology lies in potential off-target effects [40]. However, the expression of SsCak1 decreased by 60% in WT strain after infection, which further verified the correctness of HIGS (Figure 6). Therefore, in the genome of S. sclerotiorum, genes similar to SsCak1 that are conserved in pathogens without homology in host plants are expected to be target genes of HIGS.
In summary, we identified a putative SsCak1 (sscle_11g085070) by a combination of forward genetic screening and NGS, and demonstrated that the gene was persistently expressed during hyphal development and highly expressed during S. sclerotiorum infection. A knockout strain of SsCak1 was subsequently generated by split-tagging and then transformed with a WT copy to generate two complementary strains. Deletion of SsCak1 led to defects in mycelium development, sclerotia development, appressorium formation, and penetration, resulting in complete loss of virulence, suggesting that SsCak1 is essential for both growth and pathogenicity regulation, and thus can serve as a potential target for enhancing crop resistance to S. sclerotiorum through HIGS.
4. Materials and Methods
4.1. Fungal Strains, Plants and Culture Conditions
The WT S. sclerotiorum 1980 was cultured and maintained on potato dextrose agar (PDA). The deletion mutant strain and complementary strain was grown on PDA containing 150 μg/mL hygromycin B (Roche) in a 20°C culture room as previously described [41]. WT A. thaliana (Col-0), N. benthamiana for virulence test were grown in a growth room at 22°C with a 16 h light/8 h dark cycle.
4.2. Inoculation and Virulence Assessment
For inoculation of S. sclerotiorum with mycelial suspension, 6 fresh mycelium agar plugs with a diameter of 5 mm were put into a 250 mL flask containing 150 mL PDB and incubated at 22°C and 150 rpm for 24 hours. The resulting mycelial spheres were collected by filtering the medium with filter paper and washed 3 times with ddH2O and PDB, respectively. The mycelium balls were ground on ice to homogenize. The resulting liquid mycelial suspension was then adjusted to OD600 = 1.0 with PDB solution, and inoculation was referred to previous standard techniques [42,43]. The expanded detached or un-detached leaves were inoculated with actively growing mycelial agar plugs (1 mm diameter for A. thaliana leaves and 6 mm diameter for N. benthamiana leaves). Ten leaves of A. thaliana and N. benthamiana were inoculated respectively in each replication. The experiments were replicated at least three times. The inoculated leaves were incubated at 22°C with 95% - 100% relative humidity. The lesion size was photographed at 24 hpi.
4.3. Screening for mutants defective in virulence
Ascospores were collected from the ascus of the WT strain S. sclerotiorum 1980. Ascospores were subjected to UV mutagenesis as previously described [18]. Screening of mutagenized populations using lettuce leaves and multiple replicates to confirm putative mutants.
4.4. Genomic DNA Extraction and NGS
The agar block with a diameter of 5 mm and fresh hyphae was inoculated on PDA covered with cellophane. After 2 days, the mycelium was scraped off with a sterile tip, quick-frozen in liquid nitrogen, ground into powder, and genomic DNA was extracted using cetyltrime-thylammonium bromide method [44]. The crude extract was further purified for NGS with a commercial service (Novogene, Bioinformatics Technology Co., Ltd., Beijing, China). DNA degradation and contamination were evaluated on a 1% agarose gel. Paired-end library was built by Novogene, using Illumina sequencer NovaSeq 6000, and clean reads were aligned to the S. sclerotiorum genome (ASM185786v1).
4.5. Candidate Genes Identification
The sequence reads from NGS were mapped to the reference genome of WT strain S. sclerotiorum 1980. Mutations were identified by SAMtools with default parameters [45]. Annotation of the mutations was performed with germline short variant discovery (SNPs + INDELs) based on GATK best practices [46]. False mutations in repetitive sequences were manually removed as previously described [19].
4.6. Target Gene Knockout and Transgene Complementation
Using the genomic DNA of WT S. sclerotiorum as a template, the flanking sequences of 1032 bp upstream and 998 bp downstream of SsCak1 were amplified by PCR, and fused with the left and right parts of the hygromycin expression cassette to obtain the split marker fragments by overlapping PCR. The clone primer sequences are shown in Table S1. The resulting split marker fragments were transformed into WT S. sclerotiorum by PEG-mediated protoplast transformation, as previously described [47]. Transformants were selected three times on PDA medium containing 150 mg/L hygromycin. And the transformants were purified by transferring at least three times with mycelial tip. The deletion of SsCak1 gene was verified by amplifying the sequence of SsCak1 from cDNA of transformants.
For the genetic complementation of the SsCak1 deletion mutant, a genomic region containing the full-length fragment of SsCak1, including upstream and downstream of the coding sequence, was cloned from WT S. sclerotiorum gDNA. This fragment was then cloned into the modified pCH-NEO1 vector [48]. Complementary transformants were selected on PDA medium containing 100 μg/ml G418 and then verified by PCR.
4.7. RNA Extraction and cDNA Synthesis
Total RNA from S. sclerotiorum or N. benthamiana leaves with S. sclerotiorum inoculated was extracted using the EastepTM Super Total RNA Extraction Kit (Promega, Madison, WI, USA). According to the manufacturer's instructions, first-strand cDNA was synthesized by the GoScript™ Reverse Transcription System Kit (Promega, Madison, WI, USA).
4.8. RT-qPCR
Quantitative real-time RCR was performed on a StepOneTM Real-time PCR Instrument Thermal Cycling Block using SYBR® Green Premix Pro Taq HS qPCR Kit II (AG11702, Accurate Biotechnology (Hunan) Co., Ltd, Changsha, China). Use the following PCR program: 40 cycles of 2 min at 94°C, 15 s at 94°C and 1 min at 58°C. The internal reference gene for S. sclerotiorum is Tubulin1. Relative gene expression levels were analyzed using the 2-ΔΔCT method [49].
4.9. Compound Appressoria Observation
S. sclerotiorum mycelium plugs of 5 mm were placed on glass slides and cultured for 16 h to observe the formation and number of appressorium. After 16 hours of inoculation with S. sclerotiorum, onion epidermis were soaked in a 0.5% trypan blue solution for 30 min and then destained using a bleach solution (ethanol:acetic acid:glycerol = 3:1:1). Samples were examined and photographed under an optical microscope (Axio Imager 2, ZEISS, Oberkochen, Germany).
4.10. Construction of TRV-HIGS Vectors and Agro-infiltration in N. benthamiana
Short cDNA fragments of SsCak1 in S. sclerotiorum were amplified by PCR using gene specific primers with EcoRI and BamHI linkers. The resultant PCR product was digested with EcoRI and BamHI and cloned into a linearized pTRV2 vector. A TRV2-based construct harbouring GFP was used as a negative control of VIGS and HIGS [50,51]. A TRV2-based construct harbouring phytoene desaturase (PDS) from N. benthamiana (pTRV2::PDS) was used as a positive control for VIGS efficiency (Senthil-Kumar and Mysore, 2014). Finally, pTRV1 and these pTRV2-based constructs were separately transformed into Agrobacterium tumefaciens GV3101 by electroporation.
The mixed Agrobacterium solution was then infiltrated into the first and second leaves of 2-week-old N. benthamiana using a needle-free syringe as previously described [36]. N. benthamiana plants were then grown in a growth chamber for at least 14 days.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Identification of a pathogenicity-deficient mutant M14-9 by screening on lettuce leaves; Figure S2: Identification of the molecular lesions of the M14-9 mutants; Figure S3: Identification of Sscak1 by PCR; Figure S4: Sclerotia number and weight of SsCak1 knockout and complementation strains; Figure S5: WT, M14-9, Sscak1, Sscak1-C1 and Sscak1-C5 grown on PDA medium containing bromophenol blue. Experiments were conducted three times with similar results.
Author Contributions
L.Q. X.L. and S.X. planned and designed the research. L.Q., J.N., X.G., and Y.X. performed research. Y.X. and Y. Q. contributed to the mutagenized population of S. sclerotiorum. X.T. conducted bioinformatics analysis. L.Q. and K.C. analyzed and validated the data. L.Q., J.N., X.L. and S.X. wrote the original draft of the manuscript. All authors discussed the data, edited, and approved the manuscript.
Funding
This research was funded by National Natural Science Foundation of China, grant number 31971836 and Natural Science Foundation of Changsha City, grant number kq2208141.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We cordially thank Dr. Daohong Jiang (Huazhong Agricultural University) for sharing pCH-EF-1 plasmid, and Jeffrey Rollins (University of Florida) for sharing WT S. sclerotiorum strain 1980.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Boland, G.J.; Hall, R. Index of plant hosts of sclerotinia sclerotiorum. Canadian journal of plant pathology 1994, 2, 93–108. [Google Scholar] [CrossRef]
- Bolton, M.D.; Thomma, B.P.; Nelson, B.D. Sclerotinia sclerotiorum (Lib.) de Bary: Biology and molecular traits of a cosmopolitan pathogen. Molecular plant pathology 2006, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Rollins, J.A. Mechanisms of broad host range necrotrophic pathogenesis in Sclerotinia sclerotiorum. Phytopathology 2018, 108, 1128–1140. [Google Scholar] [CrossRef] [PubMed]
- Rollins, J.A.; Dickman, M.B. pH signaling in Sclerotinia sclerotiorum: Identification of a pacC/RIM1 homolog. Applied and environmental microbiology 2001, 67, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Seifbarghi, S.; Borhan, M.H.; Wei, Y.; Coutu, C.; Robinson, S.J.; Hegedus, D.D. Changes in the Sclerotinia sclerotiorum transcriptome during infection of Brassica napus. BMC genomics 2017, 18, 266. [Google Scholar] [CrossRef]
- Ding, Y.; Chen, Y.; Yan, B.; Liao, H.; Dong, M.; Meng, X.; Wan, H.; Qian, W. Host-induced gene silencing of a multifunction gene Sscnd1 enhances plant resistance against Sclerotinia sclerotiorum. Frontiers in microbiology 2021, 12, 693334. [Google Scholar] [CrossRef]
- Williams, B.; Kabbage, M.; Kim, H.J.; Britt, R.; Dickman, M.B. Tipping the balance: Sclerotinia sclerotiorum secreted oxalic acid suppresses host defenses by manipulating the host redox environment. PLoS pathogens 2011, 7, e1002107. [Google Scholar] [CrossRef]
- Chittem, K.; Yajima, W.R.; Goswami, R.S.; Del Río Mendoza, L.E. Transcriptome analysis of the plant pathogen Sclerotinia sclerotiorum interaction with resistant and susceptible canola (Brassica napus) lines. PLoS ONE 2020, 15, e0229844. [Google Scholar] [CrossRef]
- Huang, L.; Buchenauer, H.; Han, Q.; Zhang, X.; Kang, Z. Ultrastructural and cytochemical studies on the infection process of Sclerotinia sclerotiorum in oilseed rape. Journal of Plant Diseases and Protection 2008, 115, 9–16. [Google Scholar] [CrossRef]
- Tariq, V.N.; Jeffries, P. Ultrastructure of penetration of Phaseolus spp. by Sclerotinia sclerotiorum. Canadian Journal of Botany 1986, 64, 2909–2915. [Google Scholar] [CrossRef]
- Garg, H.; Li, H.; Sivasithamparam, K.; Kuo, J.; Barbetti, M.J. The infection processes of Sclerotinia sclerotiorum in cotyledon tissue of a resistant and a susceptible genotype of Brassica napus. Annals of botany 2010, 106, 897–908. [Google Scholar] [CrossRef] [PubMed]
- Andradeab, C.M.; Tinocoa, M.L.P.; Rietha, A.F.; Maiaa, F.C.O.; Araga~Oa, F.J.L. Host-induced gene silencing in the necrotrophic fungal pathogen Sclerotinia sclerotiorum. Plant Pathology 2016, 65. [Google Scholar] [CrossRef]
- Rana, K.; Yuan, J.; Liao, H.; Banga, S.S.; Kumar, R.; Qian, W.; Ding, Y. Host-induced gene silencing reveals the role of Sclerotinia sclerotiorum oxaloacetate acetylhydrolase gene in fungal oxalic acid accumulation and virulence. Microbiol Res. 2022, 258, 126981. [Google Scholar] [CrossRef]
- Wytinck, N.; Ziegler, D.J.; Walker, P.L.; Sullivan, D.S.; Biggar, K.T.; Khan, D.; Sakariyahu, S.K.; Wilkins, O.; Whyard, S.; Belmonte, M.F. Host induced gene silencing of the Sclerotinia sclerotiorum ABHYDROLASE-3 gene reduces disease severity in Brassica napus. PLoS ONE 2022, 17, e0261102. [Google Scholar] [CrossRef]
- Rana, K.; Ding, Y.; Banga, S.S.; Liao, H.; Zhao, S.; Yu, Y.; Qian, W. Sclerotinia sclerotiorum Thioredoxin1 (SsTrx1) is required for pathogenicity and oxidative stress tolerance. Molecular plant pathology 2021, 22, 1413–1426. [Google Scholar] [CrossRef]
- Xu, Y.; Qiu, Y.; Zhang, Y.; Li, X. A cAMP phosphodiesterase is essential for sclerotia formation and virulence in Sclerotinia sclerotiorum. Front Plant Sci 2023, 14, 1175552. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Josh, L.; Yan, X.; Yilan, Q.; Xin, L. A MAP kinase cascade broadly regulates development and virulence of Sclerotinia sclerotiorum and can be targeted by HIGS for disease control. bioRxiv 2003, 2023.2003.2001.530680. [Google Scholar] [CrossRef]
- Xu, Y.; Ao, K.; Tian, L.; Qiu, Y.; Huang, X.; Liu, X.; Hoy, R.; Zhang, Y.; Rashid, K.Y.; Xia, S.; et al. A forward genetic screen in Sclerotinia sclerotiorum revealed the transcriptional regulation of its sclerotial melanization Pathway. Molecular plant-microbe interactions: MPMI 2022, 35, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Sun, Y.; Orduna, A.R.; Jetter, R.; Li, X. The Mediator kinase module serves as a positive regulator of salicylic acid accumulation and systemic acquired resistance. The Plant journal 2019, 98, 842–852. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, S.; Ma, J.; Dai, Y.; Li, C.; Lyu, X.; Wang, C.; Xu, J.R. Two Cdc2 Kinase Genes with distinct functions in vegetative and infectious hyphae in Fusarium graminearum. PLoS pathogens 2015, 11, e1004913. [Google Scholar] [CrossRef]
- Wu, J.; Cai, G.; Tu, J.; Li, L.; Liu, S.; Luo, X.; Zhou, L.; Fan, C.; Zhou, Y. Identification of QTLs for resistance to sclerotinia stem rot and BnaC.IGMT5.a as a candidate gene of the major resistant QTL SRC6 in Brassica napus. PLoS ONE 2013, 8, e67740. [Google Scholar] [CrossRef]
- Hanks, S.K.; Hunter, T. Protein kinases 6. The eukaryotic protein kinase superfamily: Kinase (catalytic) domain structure and classification. FASEB journal 1995, 9, 576–596. [Google Scholar] [CrossRef] [PubMed]
- Manning, G.; Plowman, G.D.; Hunter, T.; Sudarsanam, S. Evolution of protein kinase signaling from yeast to man. Trends in biochemical sciences 2002, 27, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Miranda-Saavedra, D.; Barton, G.J. Classification and functional annotation of eukaryotic protein kinases. Proteins 2007, 68, 893–914. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Harel, A.; Gorovoits, R.; Yarden, O.; Dickman, M.B. MAPK regulation of sclerotial development in Sclerotinia sclerotiorum is linked with pH and cAMP sensing. Molecular plant-microbe interactions: MPMI 2004, 17, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Bashi, Z.D.; Gyawali, S.; Bekkaoui, D.; Coutu, C.; Lee, L.; Poon, J.; Rimmer, S.R.; Khachatourians, G.G.; Hegedus, D.D. The Sclerotinia sclerotiorum Slt2 mitogen-activated protein kinase ortholog, SMK3, is required for infection initiation but not lesion expansion. Canadian journal of microbiology 2016, 62, 836–850. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Kipreos, E.T. Evolution of cyclin-dependent kinases (CDKs) and CDK-activating kinases (CAKs): Differential conservation of CAKs in yeast and metazoa. Mol Biol Evol 2000, 17, 1061–1074. [Google Scholar] [CrossRef]
- Morgan, D.O. Cyclin-dependent kinases: Engines, clocks, and microprocessors. Annu Rev Cell Dev Biol 1997, 13, 261–291. [Google Scholar] [CrossRef]
- Espinoza, F.H.; Farrell, A.; Erdjument-Bromage, H.; Tempst, P.; Morgan, D.O. (1996). A cyclin-dependent kinase-activating kinase (CAK) in budding yeast unrelated to vertebrate CAK. Science 1997, 273, 1714–1717. [Google Scholar] [CrossRef]
- Saiz, J.E.; Fisher, R.P. A CDK-activating kinase network is required in cell cycle control and transcription in fission yeast. Curr Biol 2000, 12, 1100–1105. [Google Scholar] [CrossRef]
- Zhang, B.W. The Function of Cell Division Cycle Protein SsCdc28 in Sclerotinia sclerotiorum. Master, Jilin University, Jilin, 2019. [Google Scholar]
- Castanheira, S.; Pérez-Martín, J. Appressorium formation in the corn smut fungus Ustilago maydis requires a G2 cell cycle arrest. Plant signaling & behavior 2015, 10, e1001227. [Google Scholar] [CrossRef]
- Osés-Ruiz, M.; Talbot, N.J. Cell cycle-dependent regulation of plant infection by the rice blast fungus Magnaporthe oryzae. Communicative & integrative biology 2017, 10, e1372067. [Google Scholar] [CrossRef]
- de la Torre, A.; Castanheira, S.; Pérez-Martín, J. Incompatibility between proliferation and plant invasion is mediated by a regulator of appressorium formation in the corn smut fungus Ustilago maydis. Proceedings of the National Academy of Sciences of the United States of America 2020, 117, 30599–30609. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Cao, S.; Zhang, C.; Zhang, Y.; Zhang, Q.; Xu, J.R.; Wang, C. MoCDC14 is important for septation during conidiation and appressorium formation in Magnaporthe oryzae. Molecular plant pathology 2018, 19, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Hawkins, N.J.; Sanglard, D.; Gurr, S.J. Worldwide emergence of resistance to antifungal drugs challenges human health and food security. Science (New York, NY) 2018, 360, 739–742. [Google Scholar] [CrossRef] [PubMed]
- Nowara, D.; Schweizer, P.; Gay, A.; Lacomme, C.; Shaw, J.; Ridout, C.; et al. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell 2010, 22, 3130–3141. [Google Scholar] [CrossRef]
- Zhang, T.; Jin, Y.; Zhao, J.H.; Gao, F.; Zhou, B.J.; Fang, Y.Y.; et al. (2016). Host-induced gene silencing of the target gene in fungal cells confers effective resistance to the cotton wilt disease pathogen Verticillium dahliae. Mol. Plant 2016, 9, 939–942. [Google Scholar] [CrossRef] [PubMed]
- Spada, M.; Pugliesi, C.; Fambrini, M.; Pecchia, S. (2021). Silencing of the Slt2-type MAP kinase Bmp3 in Botrytis cinerea by application of exogenous dsRNA affects fungal growth and virulence on Lactuca sativa. Int. J. Mol. Sci 2021, 22, 5362. [Google Scholar] [CrossRef]
- Lundgren, J.G.; Duan, J.J. RNAi-based insecticidal crops: Potential effects on nontarget species. Bioscience 2013, 63, 657–665. [Google Scholar] [CrossRef]
- Yang, C.; Li, W.; Huang, X.; Tang, X.; Qin, L.; Liu, Y.; Xia, Y.; Peng, Z.; Xia, S. SsNEP2 contributes to the virulence of Sclerotinia sclerotiorum. Pathogens (Basel, Switzerland) 2022, 11, 446. [Google Scholar] [CrossRef] [PubMed]
- Mei, J.; Liu, Y.; Wei, D.; Wittkop, B.; Ding, Y.; Li, Q.; Li, J.; Wan, H.; Li, Z.; Ge, X.; et al. Transfer of sclerotinia resistance from wild relative of Brassica oleracea into Brassica napus using a hexaploidy step. Theor Appl Genet 2015, 128, 639–644. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, C.; Wan, J.; Li, Z.; Liu, D.; Yamamoto, N.; Zhou, E.; Shu, C. Functional validation of pathogenicity genes in rice sheath blight pathogen Rhizoctonia solani by a novel host-induced gene silencing system. Molecular plant pathology 2021, 22, 1587–1598. [Google Scholar] [CrossRef] [PubMed]
- Allen, G.C.; Flores-Vergara, M.A.; Krasynanski, S.; Kumar, S.; Thompson, W.F. A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethylammonium bromide. Nature protocols 2006, 1, 2320–2325. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics (Oxford, England) 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Van der Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; Del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. From FastQ data to high confidence variant calls: The Genome Analysis Toolkit best practices pipeline. Current protocols in bioinformatics 2013, 43, 11.10.11–11.10.33. [Google Scholar] [CrossRef] [PubMed]
- Catlett, N.L.; Lee, B.N.; Yoder, O.C.; Turgeon, B.G. Split-marker recombination for efficient targeted deletion of fungal genes. Fungal Genetics Newsletter 2003, 50, 9–11. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Y.; Liu, J.; Zhang, Y.; Pan, H. The Sclerotinia sclerotiorum FoxE2 gene is required for apothecial development. Phytopathology 2016, 106, 484–490. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Senthil-Kumar, M.; Mysore, K.S. Tobacco rattle virus-based virus-induced gene silencing in Nicotiana benthamiana. Nat Protoc 2014, 9, 1549–1562. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.; Page, J.E. Optimized cDNA libraries for virus-induced gene silencing (VIGS) using tobacco rattle virus. Plant Methods 2008, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Pathogenicity and mycelial growth phenotypes of M14-9 mutant. (A) Necrotic areas caused by wild type (WT) and M14-9 on tobacco leaves at 48 hpi; The experiment was repeated at least three times. Bar = 1 cm. (B) Quantification of lesion size of WT and M14-9 on tobacco leaves. Image J was used to quantify the lesion size. **p < 0.01, one-way ANOVA test. (C) Mycelial morphology of WT and M14-9 strains after 2 d or 14 d on potato dextrose agar (PDA) media (D) Colony diameter of M14-9 and WT strains cultured on PDA plates. (E) Morphology of mycelium under light microscope of M14-9 and WT strains. Bar = 200 μm. (F-G) Sclerotia number (F) or weight (G) per plate of M14-9 and WT strains. **p < 0.01, one-way ANOVA test.
Figure 1.
Pathogenicity and mycelial growth phenotypes of M14-9 mutant. (A) Necrotic areas caused by wild type (WT) and M14-9 on tobacco leaves at 48 hpi; The experiment was repeated at least three times. Bar = 1 cm. (B) Quantification of lesion size of WT and M14-9 on tobacco leaves. Image J was used to quantify the lesion size. **p < 0.01, one-way ANOVA test. (C) Mycelial morphology of WT and M14-9 strains after 2 d or 14 d on potato dextrose agar (PDA) media (D) Colony diameter of M14-9 and WT strains cultured on PDA plates. (E) Morphology of mycelium under light microscope of M14-9 and WT strains. Bar = 200 μm. (F-G) Sclerotia number (F) or weight (G) per plate of M14-9 and WT strains. **p < 0.01, one-way ANOVA test.
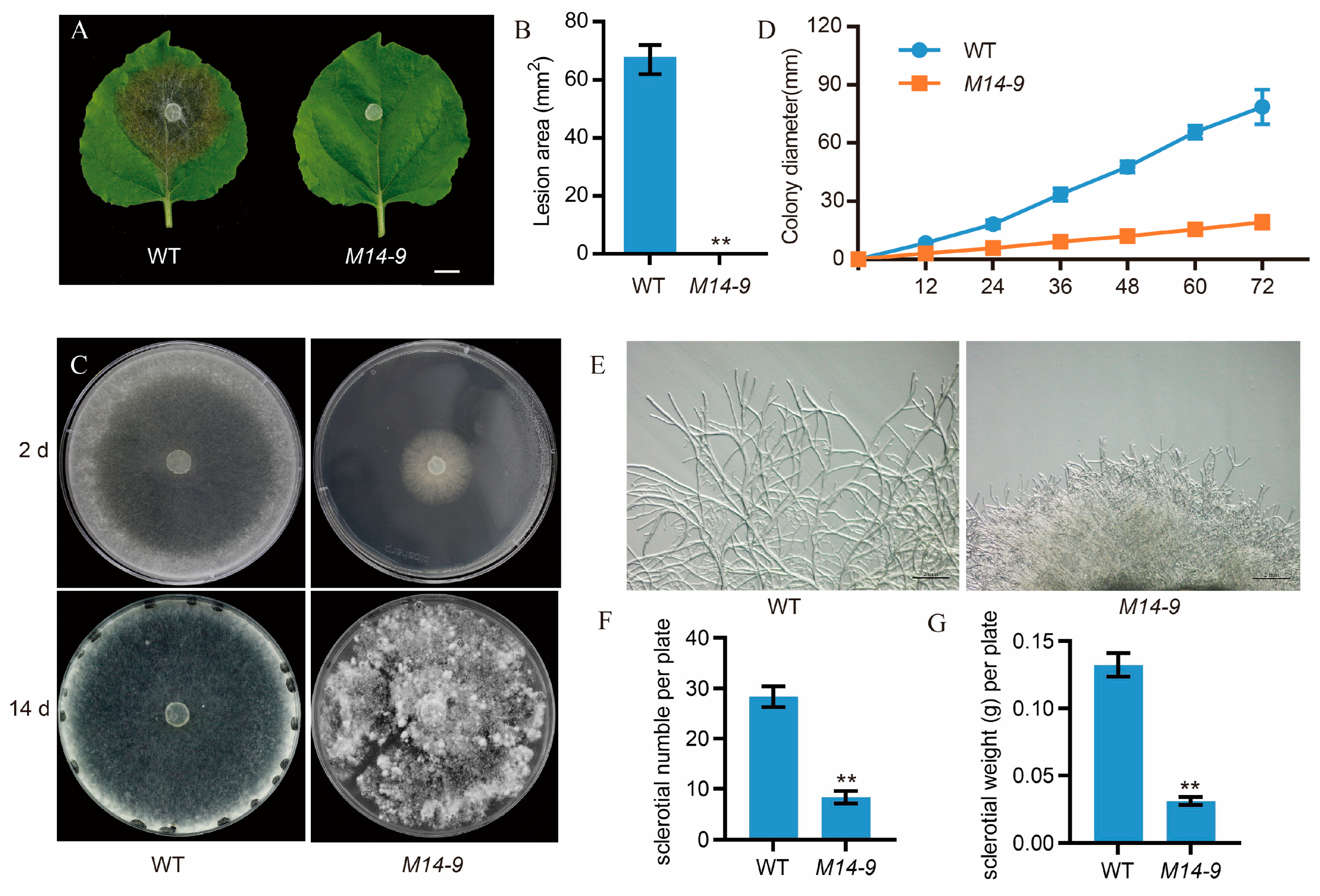
Figure 2.
Sequence and expression analysis of the candidate gene in M14-9. (A) Phylogenetic analysis of S. sclerotiorum sscle_11g085070 and other homologous Cak1 from Botrytis cinerea, Monilinia laxa, Rutstroemla sp. NJR-2017a WRK4, Lachnellula suecica, Hyaloscypha variabilis F, Cadophora malorum, Fusarium graminearum and Saccharomyces cerevisiae. Phylogenetic analysis was performed using MEGA 6.0 software with the maximum likelihood method. SsCAK1 was marked with ▲. (B) Multiple alignment of sscle_11g085070 with homologous sequences of B. cinerea, M. laxa, R. sp. NJR-2017a WRK4, L. suecica, H. variabilis F, C. malorum, F.graminearum and S. cerevisiae; (C) Quantitative real-time reverse transcription-polymerase chain reaction (RT-qPCR) analysis of SsCak1 expression during different developmental stages of S. sclerotiorum grown on potato dextrose agar (PDA) plates. The quantity of tubulin1 (Sstub1) was used to normalize the expression levels of SsCak1 in different samples. Error bars represent ±SD (n = 3). **p < 0.01, one-way ANOVA test. (D) Expression analysis of SsCak1 in S. sclerotiorum after being inoculated on N. benthamiana leaves. The quantity of tubulin1 (Sstub1) was used to normalize the expression levels of SsCak1 in different samples. Error bars represent ±SD (n = 3). **p < 0.01, one-way ANOVA test.
Figure 2.
Sequence and expression analysis of the candidate gene in M14-9. (A) Phylogenetic analysis of S. sclerotiorum sscle_11g085070 and other homologous Cak1 from Botrytis cinerea, Monilinia laxa, Rutstroemla sp. NJR-2017a WRK4, Lachnellula suecica, Hyaloscypha variabilis F, Cadophora malorum, Fusarium graminearum and Saccharomyces cerevisiae. Phylogenetic analysis was performed using MEGA 6.0 software with the maximum likelihood method. SsCAK1 was marked with ▲. (B) Multiple alignment of sscle_11g085070 with homologous sequences of B. cinerea, M. laxa, R. sp. NJR-2017a WRK4, L. suecica, H. variabilis F, C. malorum, F.graminearum and S. cerevisiae; (C) Quantitative real-time reverse transcription-polymerase chain reaction (RT-qPCR) analysis of SsCak1 expression during different developmental stages of S. sclerotiorum grown on potato dextrose agar (PDA) plates. The quantity of tubulin1 (Sstub1) was used to normalize the expression levels of SsCak1 in different samples. Error bars represent ±SD (n = 3). **p < 0.01, one-way ANOVA test. (D) Expression analysis of SsCak1 in S. sclerotiorum after being inoculated on N. benthamiana leaves. The quantity of tubulin1 (Sstub1) was used to normalize the expression levels of SsCak1 in different samples. Error bars represent ±SD (n = 3). **p < 0.01, one-way ANOVA test.
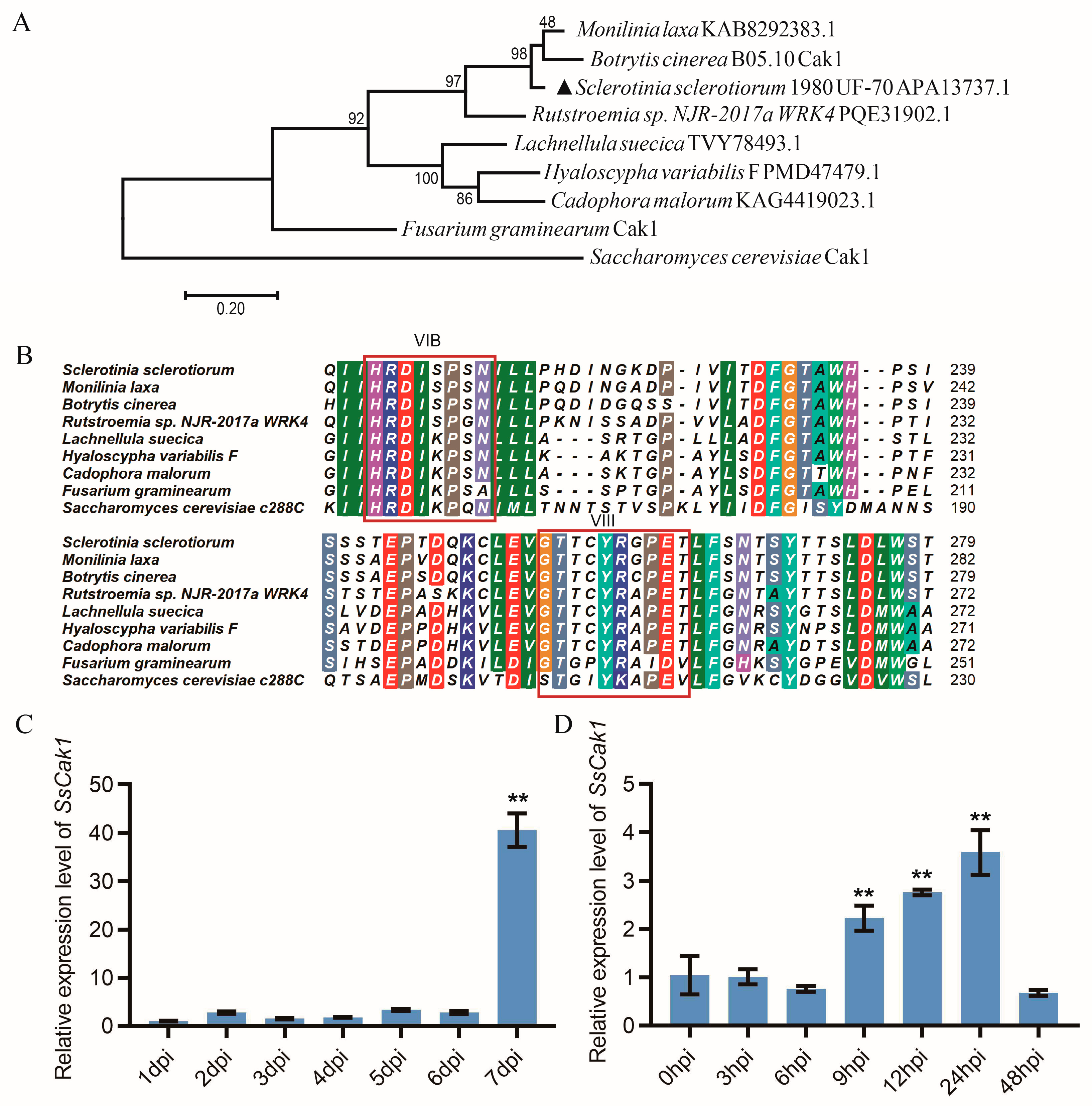
Figure 3.
Morphology of the SsCak1 knockout and complementation strains. (A) Colony morphology of WT, Sscak1, Sscak1-C1 and Sscak1-C5 strains after 2 d on potato dextrose agar (PDA) media. (B) Morphology of mycelium under light microscope of WT, Sscak1, Sscak1-C1 and Sscak1-C5 strains. Bar = 200 μm. (C) Sclerotia morphology at 15 d on potato dextrose agar (PDA) media. (D) RT-qPCR was used to monitor the expression levels of SsCak1 in WT, Sscak1, Sscak1-C1 and Sscak1-C5 strains. The data were normalized to Sstub1 transcript level of WT strain. Error bars represent ±SD (n = 3). **p < 0.01, one-way ANOVA test. (E) Colony diameter of the WT, Sscak1, Sscak1-C1 and Sscak1-C5 cultured on PDA plates.
Figure 3.
Morphology of the SsCak1 knockout and complementation strains. (A) Colony morphology of WT, Sscak1, Sscak1-C1 and Sscak1-C5 strains after 2 d on potato dextrose agar (PDA) media. (B) Morphology of mycelium under light microscope of WT, Sscak1, Sscak1-C1 and Sscak1-C5 strains. Bar = 200 μm. (C) Sclerotia morphology at 15 d on potato dextrose agar (PDA) media. (D) RT-qPCR was used to monitor the expression levels of SsCak1 in WT, Sscak1, Sscak1-C1 and Sscak1-C5 strains. The data were normalized to Sstub1 transcript level of WT strain. Error bars represent ±SD (n = 3). **p < 0.01, one-way ANOVA test. (E) Colony diameter of the WT, Sscak1, Sscak1-C1 and Sscak1-C5 cultured on PDA plates.
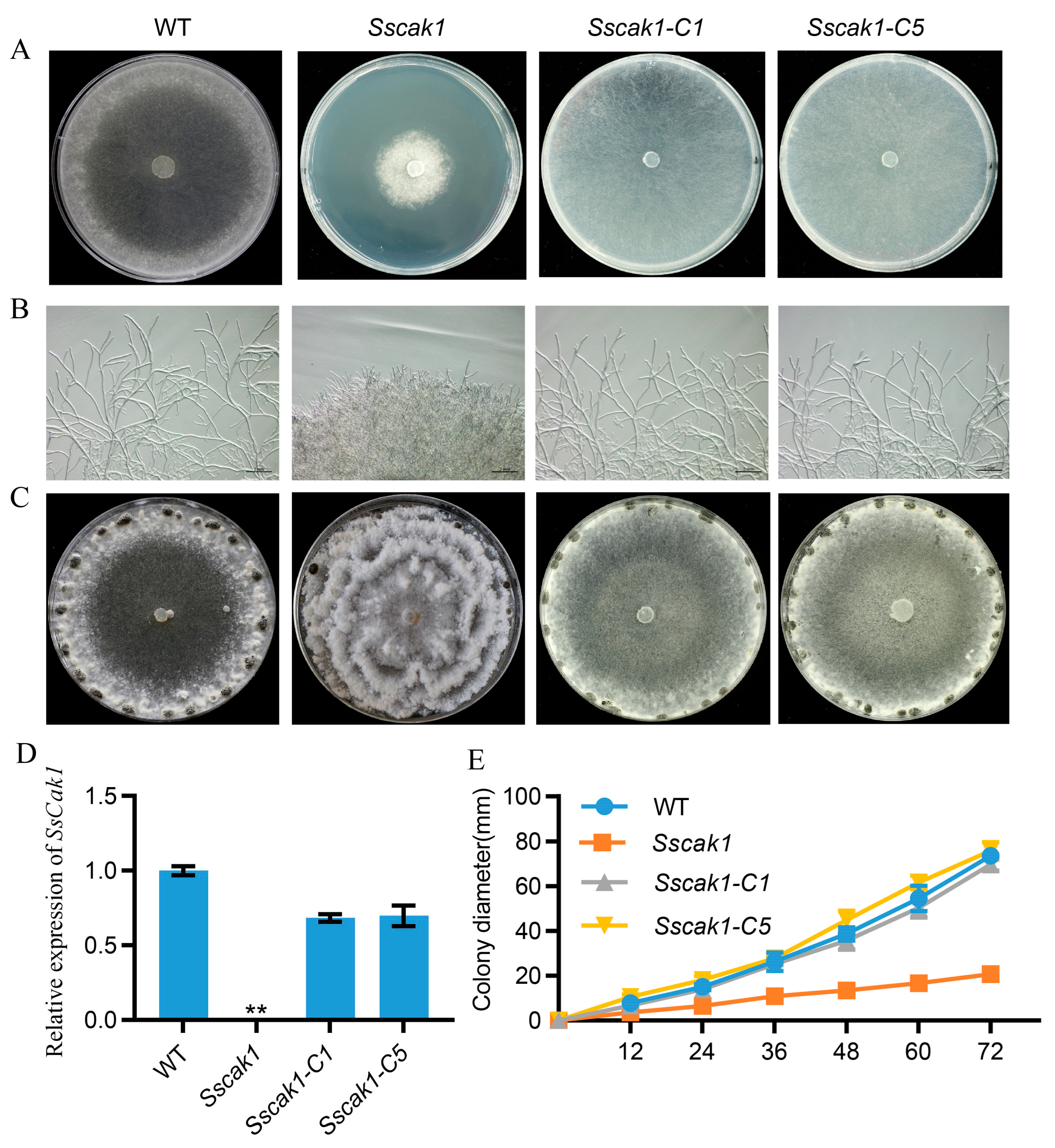
Figure 4.
Deletion of SsCak1 leads to loss of pathogenicity in S. sclerotiorum. (A-B) Lesion area caused by each strain on intact and injured A. thaliana (A) and N. benthamiana (B) leaves. The experiment was repeated at least three times. Bar = 1 cm. (C-D) Statistical analysis of the lesion area in panels above (C, A. thaliana; D, N. benthamiana). Error bars represent ±SD (n = 3). **p < 0.01, one-way ANOVA test.
Figure 4.
Deletion of SsCak1 leads to loss of pathogenicity in S. sclerotiorum. (A-B) Lesion area caused by each strain on intact and injured A. thaliana (A) and N. benthamiana (B) leaves. The experiment was repeated at least three times. Bar = 1 cm. (C-D) Statistical analysis of the lesion area in panels above (C, A. thaliana; D, N. benthamiana). Error bars represent ±SD (n = 3). **p < 0.01, one-way ANOVA test.
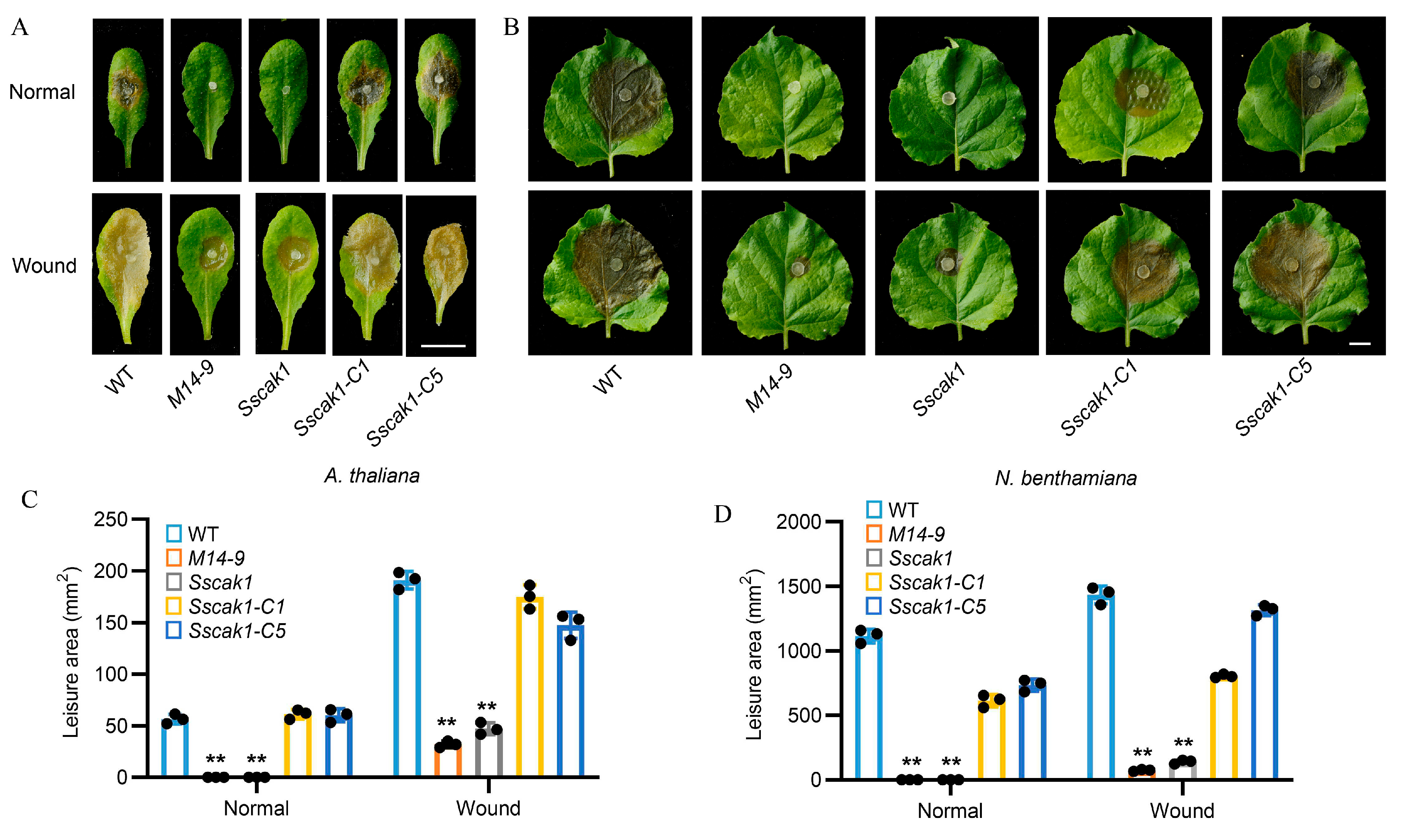
Figure 5.
Knockout of SsCak1 results in defects in appressorium formation and penetration. (A-B) WT, M14-9, Sscak1, Sscak1-C1 and Sscak1-C5 strains placed on glass slides and cultured for 16 h to observe the number of appressoria formed. Bar = 200 μm. (C) Penetration assay of WT, M14-9, Sscak1, Sscak1-C1 and Sscak1-C5 on onion epidermis cells. Invasion hyphae were stained with trypan blue. Bar = 100 μm.
Figure 5.
Knockout of SsCak1 results in defects in appressorium formation and penetration. (A-B) WT, M14-9, Sscak1, Sscak1-C1 and Sscak1-C5 strains placed on glass slides and cultured for 16 h to observe the number of appressoria formed. Bar = 200 μm. (C) Penetration assay of WT, M14-9, Sscak1, Sscak1-C1 and Sscak1-C5 on onion epidermis cells. Invasion hyphae were stained with trypan blue. Bar = 100 μm.
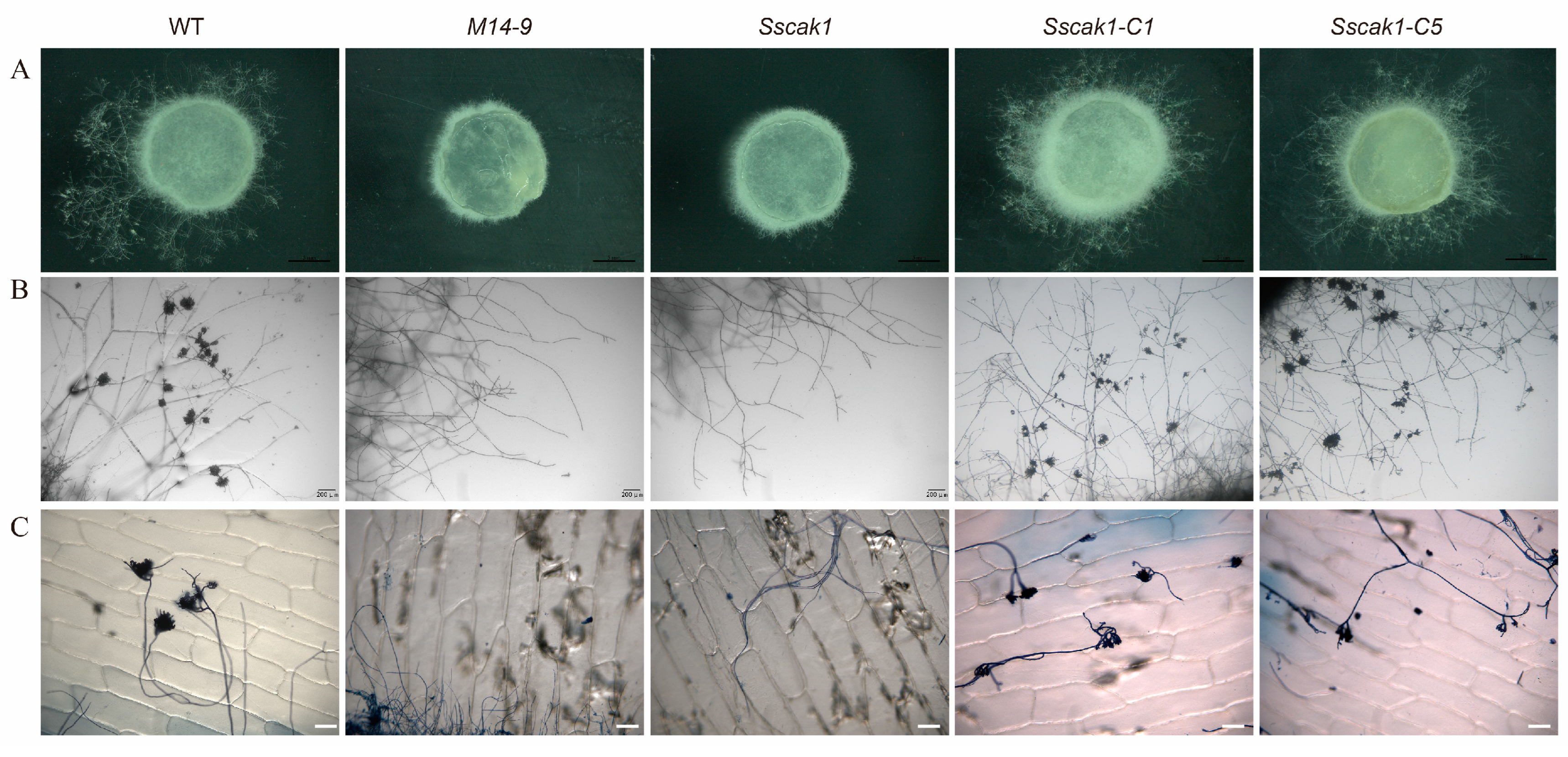
Figure 6.
Effects of host-induced gene silencing (HIGS) of SsCak1 in N. benthamiana. (A) The phenotype of pTRV2::SsCak1-infiltrated plants at 14 days after infiltration. Bar =1 cm. (B) Reverse transcription PCR was used to monitor expression levels of target genes in infiltrated plants. M, 2-kb DNA ladder; 1, pTRV2::GFP; 2, pTRV2::PDS; 3, pTRV2::SsCak1. (C) Necrotic symptoms on leaves inoculated with WT strain at 2 d post-inoculation (dpi). One of the representative biological replicates is shown. Bar =1 cm. (D) Statistical analysis of the lesion area. Error bars represent ±SD (n = 10). **p < 0.01, one-way ANOVA test. (E) Gene expression levels of SsCak1. The data were normalized to the Sstub1 transcript levels of WT strain. Error bars represent ±SD (n = 3). **p < 0.01, one-way ANOVA test.
Figure 6.
Effects of host-induced gene silencing (HIGS) of SsCak1 in N. benthamiana. (A) The phenotype of pTRV2::SsCak1-infiltrated plants at 14 days after infiltration. Bar =1 cm. (B) Reverse transcription PCR was used to monitor expression levels of target genes in infiltrated plants. M, 2-kb DNA ladder; 1, pTRV2::GFP; 2, pTRV2::PDS; 3, pTRV2::SsCak1. (C) Necrotic symptoms on leaves inoculated with WT strain at 2 d post-inoculation (dpi). One of the representative biological replicates is shown. Bar =1 cm. (D) Statistical analysis of the lesion area. Error bars represent ±SD (n = 10). **p < 0.01, one-way ANOVA test. (E) Gene expression levels of SsCak1. The data were normalized to the Sstub1 transcript levels of WT strain. Error bars represent ±SD (n = 3). **p < 0.01, one-way ANOVA test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



