
Submitted:
20 July 2023
Posted:
21 July 2023
You are already at the latest version
Abstract
The intricacies of translational strategies make this stage of the implementation of genetic information one of the most difficult to understand and, at the same time, perhaps the most interesting. It also becomes clear that all this variety of strategies is not just a consequence of a long evolutionary history but is of paramount importance for fine-tuning gene expression and metabolic modulation. This is especially true for those organisms that demonstrate predominantly biochemical and physiological reactions with a lack of behavioural ones. Plants are just such a group of organisms. Overcoming unfavourable environmental conditions occupies a key place in plant physiology. This is especially true with the changing conditions of global warming and the irretrievable loss or depletion of natural ecosystems. Conceptually, the response of plants to abiotic stress is a set of complex and intricate strategies. This is dictated both by a variety of abiotic factors that provoke stressful conditions and by a variety of molecular genetic mechanisms that make it possible to fine-tune the metabolic pathways through which the plant organism overcomes non-standard and non-optimal conditions. In the presented review, we tried to focus on the state-of-the-art in the field of translational regulation in plants under abiotic stress conditions. Various regulatory elements and patterns are considered in relative chronological order. We also considered it necessary to pay attention to key high-performance methods for studying the dynamics of genetic information at the stage of translation.
Keywords:
abioitic stress
; plant translation
; IRES
; 5'-UTR
; uORF
; codon bias
; G-qudruplex
1. Introduction
Studying the response to stress in plants continues to be an urgent task in plant physiology. The fundamental component of this direction is complemented by the most important applied aspects: the preservation and increase in crop yields, the production of genetically modified plants, the fight against the consequences of environmental disasters, etc. The purpose of this review is to consider the main aspects of studying translation in plants under stressful conditions, namely, modulation of activity, changes in the profile of translated genes, patterns and mechanisms of regulation, as well as methods used in this area of research, both classical and developed specifically for this task. However, it is impossible to consider translation in isolation from all other stages of the implementation of genetic information. Stress affects all aspects of plant life, from the structure and functional activity of the genome to the metabolome and physiological reactions. The regulation of metabolism in all living organisms is largely confused due to evolutionary processes. In the presented review, we will focus on the sequential consideration of the process of plant response to the action of stress factors.
A plant organism’s recognition of stressful conditions and its response to them are formed due to a developed sensory system Figure 1. This set of sensors includes, first of all, cell membrane sensor proteins. Such proteins respond to changes in temperature, small signal molecules, changes in membrane fluidity, etc. Curiously, the sensory system of a plant cell is not limited to external sensors. For example, in recent years, researchers have attracted significant interest in sensors that perform an unfolded protein response. Proteins that perform this function are located in the endoplasmic reticulum and recognise misfolded or not completely folded protein molecules, signalling the development of stressful conditions inside the cell. In particular, the tomato model [1,2] and, of course, Arabidopsis [3,4] demonstrated the active use of this molecular mechanism in plants.
The initial mRNA population that enters the translation pathway does not come from nowhere. An array of molecular instructions is formed during transcription. However, this is not the very beginning. In addition to the nucleotide context (promoters and enhancers, etc.), equal actors are various epigenetic processes that operate at the stage of permanent storage of genetic information in the genome. Epigenetic processes change the availability of genes for translation. Abiotic stress has a wide impact on the expression regulation process and naturally affects the epigenetic aspect. Since a detailed consideration of epigenetic regulation in stressed plants is beyond the scope of this review, we will briefly review current works devoted only to chromatin modulation and DNA methylation. The genome, as a rule, is not directly accessible to the impact of stress factors. Using Arabidopsis thaliana as an example, it can be seen that stress factors, through signalling pathways (for example, the jasmonate pathway), trigger various types of epigenetic responses. It can be seen, for example, that different types of abiotic stress change the number of open chromatin regions (OCR) differently. So, compared with the control, drought and high temperatures increase the number of similar genome regions by 26 and 16 percent, respectively, and low-temperature stress, on the contrary, reduces the representation of these regions by 18 percent [5].
Using rice (Oryza sativa) as an example, global changes in chromatin structure have been demonstrated, including changes in the size of Topologically associating domains, loss of short-range interactions, and A/B compartment transition, in response to heat stress [6]. Heat stress also induces the activation of transposable elements (TEs), with the degree of activation being negatively correlated with the degree of chromatin interaction [7]. Methods of artificial epigenetic modification of the plant genome are of particular interest. This direction opens up additional possibilities in the regulation of expression, including under conditions of abiotic stress. A significant contribution to the development of these methods was made by a modification of the genomic editing technology (CRISPR:Cas9) [8]. To further explore the epigenetic regulation of gene expression in plants under stress, two comprehensive reviews can be recommended: [9,10].
An important role in plant acclimation under stress conditions is also played by the stages of posttranscriptional modifications: processing and splicing. It is known that, for example, under the influence of changing temperature, the mechanism of alternative splicing can be activated, which forms not only the temperature isoforms of various enzymes but also modifies the 5’-untranslated region (5’-UTR) [11]. Such changes in the composition of the 5’-UTR can affect its structure in various ways, changing the permeability of this mRNA region for ribosomes.
Billions of years of evolution have formed a very intricate and, at the same time, finely tuned mechanism of translation Figure 2. Translation as the final step in the transfer of genetic information is described in detail in terms of both chemical kinetics and as a complex multicomponent process [12,13]. Translation is a complex, multi-stage process. The efficiency of gene expression at the translation stage depends on a number of key participants: translational factors, uORFs (upstream open reading frames), IRES (intermal ribosome entry site), riboswitches (including RNA thermometers), codon composition, etc. Also, translation has its own staging, usually distinguished by initiation, elongation and termination. The key stage, as a rule, is considered to be the stage of translation initiation. The two main pathways by which translation initiation can occur include CEP-dependent and CEP-independent translation initiation. The CEP-dependent mechanism implies the recognition of 5’-CEP (7-methylguanosine) 43S by the pre-initiation complex. The 43S complex, in turn, consists of translation initiation factors: eIF3, eIF4, eIF2 (more precisely, eIF2:GTP), Met-tRNA, and, of course, the small subunit (40S) of the ribosome. The participation of the eIF2B factor is also required. Translation elongation begins when the entire complex completes its scan and is set at the start codon (AUG). Next, the large subunit of the ribosome is attached to the preinitiation complex, and the translation initiation factors are released. Cap-independent translation is best studied with IRES. In this case, the ribosome does not scan the mRNA in search of a start codon, but is recruited directly to a specific entry site (IRES) with the participation of IRES trans-acting factors. As a rule, for the implementation of elongation, two key factors are required: eEF1 (which delivers the necessary tRNAs) and eEF2 (which carries out the translocation of mRNA relative to the ribosome). Translation termination involves releasing factors that recognise stop codons. Details on the regulation of translation in plants under normal conditions can be found in [13].
Historically, the study of the influence of abiotic factors on the plant organism and the response of plants to these factors began with classical physiological articles. Further, a significant amount of data was obtained using biochemical approaches. However, the most active idea of responding to and overcoming the impact of abiotic environmental factors began to advance with the advent and development of molecular genetic methods and hybrid approaches that also involve classical biochemical techniques. Along with this important methodological branch in this area, the analysis of mutants that have lost the genes responsible for plant acclimation under stressful conditions has been and is still pending.
In the present review, we attempted to update the most modern ideas about the regulation of translation in plants under conditions of abiotic stress. Taking into account the fact that many regulatory elements can manifest themselves in all the main stages of translation, we decided to consider each type of regulatory code separately, without reference to the stages of translation.
2. Regulation of plant translation under stress conditions
2.1. Signaling pathways
A number of protein kinases are known to play a role in the modulation of translation in plants. In particular, common control kinase 2 (GCN2) phosphorylates eIF2 in plants. It was demonstrated [14,15] that signals generated by chloroplasts in response to cold and salinity stress, namely ROS or redox signals, promote GCN2 activation resulting in eIF2 phosphorylation. There have also been studies on the effect of osmoprotectants on GCN2 [16]. In the paper [17], it was shown that GCN2 from A.thaliana works under the control of reactive oxygen species (ROS) released from chloroplasts under various abiotic stresses, such as excess light. eIF2-phosphorylation under excessive light stress and attenuate GCN2 signalling activity. Another kinase that strictly coordinates translation is SnRK1, which is an evolutionarily conserved regulator of cellular energy stress signalling in plants. Stress conditions such as hypoxia impair photosynthesis and are one of the major energy threats to plants. Under such conditions, the plant must balance energy expenditure to maintain homeostasis [18]. showed that activation of SnRK1, a key sensor of energy stress signalling, leads to reprogramming of the transcriptome and metabolism to adapt to such cellular energy crises and survive them. SnRK1 phosphorylates the 4G1 isoform (eIFiso4G1) of the eukaryotic translation initiation factor (eIFiso17G4), which leads to a higher translational efficiency of specific transcripts, including those induced by hypoxia during flooding.
2.2. Translation factors are a necessary part of translation
The most well-studied element of translation control is initiation. It is currently believed that it is the initiation of translation that has the greatest influence on regulation, including during various stresses. However, it is initiation that has distinct components for different kingdoms, in contrast to elongation and termination, which are the most conservative. In this section, translation initiation factors will be discussed, but it is worth noting that within the plant kingdom there is great diversity in the translation apparatus, and most current ideas are based on data obtained from A.thaliana. According to modern concepts, the number of translation initiation factors for eukaryotes exceeds fifteen. In this section, we consider only the factors that have the greatest direct effect on translation in plants during stress.
Most of these factors, or factor subunits, are multidomain proteins, as two isoforms of eIF4F are present in plants. eIF4F, which consists of eIF4E and eIF4G, as well as the eIFiso4F isoform, which includes eIFiso4E and eIFiso4G [19]. The eukaryotic translation initiation factor eIF4F is an indispensable component of the translation mechanism and also plays a role in developmental processes and stress alleviation in plants and animals. Both eIF4F isoforms are associated with resistance to various stress factors. Mutations of both eIFIso4G have a negative impact on plant life by reducing the rate of translation. Cho et al. [20] found that eIF4F supports translation of mRNAs that have a structured 5’-UTR and lack a cap or contain multiple cistrons, while eIFiso4F prefers unstructured mRNAs or mRNAs with simple structures, which is also a feature of mRNAs that are translated during hypoxia. The study demonstrated that eIFiso4G1 is required for hypoxia tolerance and SnRK1-mediated translation of specific mRNAs, including well-known hypoxia-responsive genes. This indicates that in order to respond quickly to stress during hypoxia, SnRK1 phosphorylates eIFiso4G1, which enhances protein biosynthesis and supports energy production, enabling stress adaptation.
Translation initiation factors respond to an increase in temperature, so in plants, heat shock causes inhibition of cap-dependent translation; namely, strong heat stress can completely inhibit this mechanism of translation. This suggests that cap-related initiation factors may change after heat exposure, such as eIF-4B or eIF-4F. However, as we mentioned above, the translational factors of plants differ significantly from those of animals and yeast in that plants contain not only eIF-4F but also the eIF-iso4F isoform. Gallie et al. [21] it has been reported that while a number of translational factors (eIF-4E, eIF-4B and eIF-2) are modified in mammals after heat shock, in wheat only eIF-4A and eIF-4B are subject to thermally induced modifications. This is primarily due to the attached mode of existence in plants and their adaptation to environmental changes in order to minimise the harmful effects of stress [22]. One of the key steps in initiating translation is establishing the link between eIF4E and eIF4G. In most eukaryotes, this process is regulated by proteins that bind to eIF4E; in plants, CERES was discovered, which is an eIF4E-interacting protein [23]. Toribio and colleagues suggested that CERES, by reducing polysome load at ZT5 (zeitgeber time 5 (ZT5, 5 h after first light)), modulates glucose management in plants. CERES orthologues (homologues that also contain the canonical 4E-BS) were also analysed, resulting in no analogues being identified in other eukaryotes outside the plant kingdom. As a result, it was concluded that, unlike most eukaryotes, where the known translational regulators that bind to eIF4E are negative regulators of translation, CERES acts as a plant-specific translation initiator that enhances overall translation and regulates the translation of mRNA subsets at specific stages.
The group of eIF1 factors is involved in the stimulation and assembly of the 43S PIC (preinitiation complex) and includes eIF1 and eIF1A, both of which are rather conserved in all eukaryotes. The eIF1A factor functions as an mRNA scanner and AUG initiation codon locator. There are a number of works devoted to the relationship between the eIF1A factor and salt tolerance in plants [24,25]. In addition, it has been reported that eIF1A can regulate the expression of stress-related genes, including TOBLTP, GST, MnSOD, NtMPK1, poxN9, and CDPK1. The eukaryotic translation factor eIF5B in plants is of rather ancient origin and is a structural homologue of eubacterial IF2 [26]. Initiation factor 5B is a GTPase that acts late in translation initiation. Zhang et al. found [27] that the heat-sensitive mutant of A.thaliana (hot3-1) carries a mutation in the eIF5B1 gene. This mutant behaves like the wild type in the absence of stress but cannot acclimatise to high temperatures. A.thaliana is known to have three other eIF5B genes, but they do not replace eIF5B1. Polysome profiling and transcriptome analysis of hot3-1 plants show a delayed recovery of polysomes after heat stress and a decrease in the effectiveness of proteins that protect against stress factors, which demonstrates the important role of translational control in the early stages of heat acclimatisation.
eIF3 plays a key role in translation initiation through interaction with numerous factors as well as with the ribosome. In plants, it is represented by a complex consisting of 13 subunits [28]. The main role of eIF3 is to bind to the 40S subunit to participate in the formation of the 43S pre-initiation complex (PIC). Currently, there are no exact data to describe the structural composition of eIF3 in plants. Works on the effect of eIF3 overexpression on plant resistance to drought and other abiotic stresses have been published [29,30]. Eukaryotic translation initiation factor 4A (eIF4A) in the cell cytosol is present in various isoforms of eIF4A, namely eIF4A1, eIF4A2, and eIF4A3, and their expression is tightly regulated in cap-dependent translation. eIF4A is the prototype of a large family of RNA helicases; despite this, it has a minimal helicase core and does not have additional domains found in other helicases [31].
DEAD box helicases are known to play an important role in abiotic stress in flowering plants [32]. The role of eIF4A in plant tolerance to salinity and cold stress has been investigated, and novel engineering pathways have been proposed to maximise yield under suboptimal conditions [33]. It is known that eIF4A is phosphorylated in response to hypoxia and heat shock [21,34].
2.3. Protein thermosensors
In plants, one of the mechanisms of response to stress caused by an increase in temperature is the production of heat shock proteins (HSP). HSP is a family of molecular chaperones that ensures proper protein folding. It is important to note that these proteins, which support cell proteostasis, perform their functions both under stress and under normal conditions. Within the family of these proteins, several classes of HSPs are distinguished. Each class has members that are expressed constitutively and members whose expression is triggered by heat stress. Stress-induced triggering begins with an increase in membrane fluidity with increasing temperature. This leads to the activation of phospholipase C, whose products of activity are diacylglycerol and inositol-3-phosphate. The latter activates ER Ca channels, as a result of which the level of cytosolic Ca increases, activating calmodulin-dependent kinases. The targets of these kinases are transcription factors called HSFAs (Heat Shock transcription Factor A), which migrate into the nucleus and trigger the synthesis of HSP genes [35].
It has been shown that, under heat stress, translation initiation mechanisms are blocked and stress granules (SG) are formed, which sequester certain types of mRNA, preventing their degradation. Such mRNAs can be re-engaged into translation during the recovery period. Merret et al. showed that the mRNAs that are retained in the SG during heat stress are mRNAs of ribosomal proteins (RPs). The authors of the article also showed that the translation of these mRNAs actively begins after the end of heat stress and, leading to the accumulation of ribosomal proteins, provides post-stress recovery of the translational apparatus. The release of RB mRNA from SG occurs with the participation of HSP101. The problem of how the timely production of rRNA and RP is coordinated for the assembly of new ribosomes shortly after stress exposure has also been studied in detail. It has been shown that in tomato plants under heat stress, the production of rRNA transcripts continues while the process of their maturation does not occur, which leads to their accumulation in the nucleolus. Coordinated control of the formation of the basic molecules required for the generation of new ribosomes (storage of rRNA in the nucleolus and storage of RB-coding mRNA in SG) may be crucial for the formation of new ribosomes during the recovery period. In support of this, the authors showed that during the recovery period, the activity of RNA polymerases I and II is not required for the synthesis of new ribosomes. Thus, the rapid start of translation just a few minutes after the end of stress depends on the HSP101-dependent release of RB-encoding mRNAs from SG, which must be synchronised with the maturation of primary rRNAs in the nucleolus, which leads to the formation of new ribosomes [36].
A similar sequestration mechanism in SG has been shown not only for RB mRNA but also for translation initiation factors. In this case, small HSP and HSP101 play an important role. Small HSPs are involved in the formation of SH that capture eEF1B and eIF4A, and together with HSP101 are involved in the resolubilization of these proteins during the post-stress recovery phase [37].
Another class of chaperones, HSP70, together with its co-chaperones of the HSP40 class, coordinates the activity of chaperones of other classes, being a key component of a complex network of mutual regulation of these proteins (cited in [35]. Under normal conditions, HSP70 is associated with the polypeptide chain produced during translation and is involved in the folding of the peptide descending from the ribosome. In the absence of HSP70, translation stops around the first 50 ORF codons. Under heat stress, as the proportion of misfolded proteins increases, HSP70 is recruited to the region of protein aggregates, which leads to translation arrest [38]. Perhaps this is one of the reasons for the cessation of translation in the cell at the initial stages of heat stress.
During the recovery of translation after stress, the role of the translation factor eIF5B is important. Plants carrying a missense mutation in this protein did not show differences in translation efficiency under normal conditions compared to the wild type, but after stress exposure, the mutants were unable to acclimatise and restore translation [27].
2.4. RNA binding proteins
Post-transcriptional control of gene expression is carried out through the regulation of RNA metabolism (stability, splicing, polyadenylation, and transport). RNA-binding proteins (RBPs) play an important role in this process. They can directly make modifications or recruit other regulatory factors. The A.thaliana AtRBP45b protein contains three RNA-recognition domains (RRM) and has a Glu-rich region at the C-terminus involved in establishing protein-protein interactions. R. Mahalingam and colleagues showed that AtRBP45b interacts with the EDC4 protein (enhancer of decapping protein), which is involved in translational repression [39]. EDC4 interacts with decapping enzymes, which leads to the suppression of translation and promotes RNA degradation. Other studies have shown that EDC4 interacts with RNA but has no RRM motifs. Possibly, AtRBP45b is the platform on which EDC4 interacts with RNA. It is known that EDC4 is localised in the cytoplasm in P-bodies, the formation of which is associated with stress. In these areas, mRNA sequestration and translation suppression occur. The researchers obtained results demonstrating that AtRBP45b interacts with EDC4, as well as with the cap-binding protein CBP20 and the heat shock protein HSP40. During the study, co-immunoprecipitation methods, two-hybrid analysis, and FRET analysis (Förster resonance method) were used. However, it is worth mentioning that the object used was not a wild-type plant but an AtRBP45b mutant line with restored expression. Another example of the involvement of RBP in the regulation of responses to stress is AtRBP-DR1, which is a positive regulator of the salicylate-mediated immune response in plants. It is possible that the effect of AtRBP-DR1 is mediated through an increase in the stability of the transcript of the SID2 (isochorismate synthase) gene involved in the synthesis of salicylic acid [40].
Genes encoding PR proteins (pathogenesis-related proteins) are represented by 17 families, among which there is a family containing the CAP (cysteine-rich secretory protein, antigen 5 and pathogenesis-related-1) domain. Aydin Akbudak et al. showed that members of this family show an increased level of expression in tomato in response to drought stress [41]. According to Chien et al. [42] in A.thaliana, short peptides (CARE peptides) obtained by cleavage of PR proteins containing the CAP domain (PROAtCAPE) play a regulatory role in response to various types of abiotic stresses. Thus, in the roots under salinity, the level of representation of a number of transcripts (PROAtCAPE1, PROAtCAPE3, and PROAtCAPE4) decreased under salinity, the content of the transcript of the PROAtCAPE3 and PROAtCAPE6 genes also decreased under drought and cold stress, while the representation of the PROAtCAPE7 transcript under cold stress increased. The content of the PROAtCAPE1 transcript was regulated mainly by salt, while the transcripts of the remaining PROAtCAPEs were regulated by more than two abiotic stresses. Under normal conditions, CAPE1 (a cleavage product of the PROAtCAPE1 gene) suppresses the expression of genes responsible for plant tolerance to salinity. According to the authors of the article, CARE1 reduces the expression of genes involved in the synthesis of osmolytes (P5CS1 and GolS2). The content of endogenous CAPE1 in shoots was 25% of the corresponding content in roots. Since the expression of PROAtCAPE1 in the shoots was not determined, the CARE1 found in them may have a root origin . Alternative splicing (AS), which generates various transcript variants, is a mechanism for the formation of resistance to abiotic stresses. AS is carried out with the participation of the spliceosome, a complex of nuclear ribonucleoproteins (snRNPs). Serine/arginine-rich proteins (SR proteins) play an important role in splicing site recognition; they are associated with the spliceosome, but snRNPs are not [43]. These proteins are mainly involved in pre-mRNA splicing but may also perform other functions such as mRNA export, stability control, and some others. Characterised by the presence of one or two N-terminal RNA recognition motifs (RRMs) and a C-terminal arginine or serine-containing domain that can be phosphorylated. Phosphorylation status controls subcellular distribution. The role of SR proteins in plant responses to abiotic stress is important [44]. For example, loss of function of the SR34b gene, whose upregulation is induced by cadmium (Cd), causes increased accumulation of this metal ion and increased cytotoxicity associated with it. It is important that the IRT1 gene, which encodes the putative Cd transporter, is absent in the sr34b mutant, which suggests the presence of mechanisms for controlling Cd tolerance using the SR protein [45] . In another study, A.thaliana RS40 and RS41 were found to interact with the HOS5 protein (osmotic stress response protein). RS40 and RS41 knockout mutants showed hypersensitivity to salt and ABA, as well as intron retention in many stress-related genes [46].
Six gene families (small heat shock proteins) have been identified depending on localization and sequence similarity. The expression of these proteins is induced by exposure to high temperatures as well as other types of abiotic stresses. The accumulation of these proteins in response to adverse conditions correlates with stress resistance. For example, in the study of chloroplast sHsps in Agrostis stolonifera, several variants of these proteins were isolated: sHsp26.2 in the heat-resistant plant variant and identical sHsp26.2m in the heat-sensitive variant, which contained a point mutation that generated a premature stop codon [47]. Another example is the presence of additional Hsp25 polypeptides in heat-resistant variants of Agrostis palustris (cited at [48]). It is known that sHsps act as molecular chaperones by binding to unstable conformations of various stress-forming proteins. sHsps, together with other classes of chaperones, ensure the formation of the correct conformation, facilitating folding. They can also promote transport to a specific subcellular compartment or participate in the degradation of misfolded proteins.
2.5. MicroRNAs
MicroRNAs, small non-coding RNA molecules 18–25 nucleotides long, are involved in the transcriptional and translational regulation of gene expression through RNA interference. The genesis of miRNAs can be conditionally divided into three stages. The first step is the transcription of the miRNA precursor by RNA polymerase II (or, in some cases, RNA polymerase III), which is about 1000 bp long. At the second stage, this precursor molecule is cleaved by RNase III DCL-1 (Dicer-Like 1), resulting in the formation of pre-miRNA, which is a miRNA:miRNA duplex. Pre-miRNA is exported from the nucleus to the cytoplasm with the participation of the transport protein HASTY. At the third stage in the cytoplasm, the pre-miRNA binds to the Argonaute (AGO) protein, which is part of the RNA-induced gene shutdown complex (RNA-induced silencing complex (RISC)). Gene shutdown can be accomplished by degrading the mRNA or preventing its translation. If the miRNA is fully complementary to the target mRNA, then AGO can cut the mRNA and lead to its direct degradation. If there is no complete complementarity, then shutdown is achieved by preventing translation [49]. Using an in vitro model, Japanese scientists Hiro-oki Iwakawa and Yukihide Tomari showed that the AGO1-RISC complex can be involved in both target mRNA cleavage and translational repression. At the same time, several different mechanisms are reported for translational repression, depending on the location of target sites for miRNAs. Thus, if the AGO1-RISC complex is localised in the 3’-UTR region, translation is inhibited at the stage of formation of the 48S complex. This mechanism is similar to that used in animal cells. It is assumed that repression is mediated through eIF4E, eIF4G, and eIF4A factors. If the binding site of the AGO1-RISC complex is located in the ORF or 5’-UTR region, then repression is achieved by sterically blocking the advancement of the ribosome or its landing on mRNA, respectively [50] . In general, the results of recent studies suggest that plant miRNAs have sites of complementarity to the 5’-UTRs of target transcripts, and most likely, translational repression is more likely to be mediated by AGO1-RISC binding to the 5’-regions, while in animals the preferred mechanism is 3’-mediated repression [51].
In plants, microRNAs are actively involved in the regulation of gene expression at the level of translation under various types of abiotic stresses. At the same time, the targets of known miRNAs are very often transcription factors or proteins involved in auxin signalling, as a result of which the effect is systemic. In some cases, the targets are proteins that are specifically involved in the regulation of a particular process (under stress caused by high temperatures, heat shock proteins; under salt stress, membrane ion transporters). Next, we will consider a number of the most studied examples of translational regulation by miRNAs under abiotic stress in plants.
The predicted target of mir393 is the mRNA of the TIR1 protein. TIR1 is a component of the ubiquitin ligase complex, the activity of which is directed to Aux/IAA repressor proteins associated with promoters of auxin-regulated genes. Thus, an increased level of mir393 leads to a decrease in the amplitude of the auxin response and slows down plant growth under unfavourable conditions. The expression of this microRNA increases in response to cold stress as well as to moisture deficiency (cited in [52]).
In a group led by Youxin Jin, microRNAs of the miR169 family were found. It was found that the formation of miR169g is induced by drought, while that of miR169n is induced by salt stress. In the promoters of these microRNA genes, sequences were found that bind transcription factors associated with drought and salinity, respectively, which is another proof of their inducible expression. Also, it was shown that the target of miR169 is the transcription factor of the NF-Y family [53]. Previously, this transcription factor was shown to be involved in the response to drought stress (cited in [54]).
The miR395 family is an important regulator of sulphate assimilation in A.thaliana. The targets of these miRNAs are high-affinity sulphate transporters as well as three enzymes of the ATP-sulfurylase class involved in the process of uptake [55]. The level of miR395 varies depending on exposure to various metabolites that regulate sulphate assimilation [56].
Chiou et al. showed that the regulation of phosphorus assimilation is regulated by miR399 through downregulation of E2-ubiquitin ligase. The authors propose an elegant scheme for the regulation of phosphorus fluxes in wild-type and transgenic plants expressing miR399, which is not expressed at all in intact plants in the absence of phosphorus deficiency [57]. More recent studies report that miR399 has a so-called pseudo-target that can complementarily bind miRNAs but does not contain a cleavage site. It has been shown that in textitA.thaliana deficiency of inorganic phosphate, the expression of IPS (induced by phosphate starvation) genes increases, which, being pseudotargets of miR399, regulate the expression of the true target of miR399, E2-ubiquitin ligase (PHO2), involved in the renewal of the phosphate transporter [58].
Another microRNA representative involved in the response to mineral starvation is miR398, whose target is Cu-dependent superoxide desmutase (SOD). SOD is involved in the elimination of reactive oxygen species; therefore, under oxidative stress, the expression level of miR398 decreases, allowing the accumulation of SOD transcripts. However, SOD, being a metal-dependent enzyme, is involved in the homeostasis of ions, and, when this mineral nutrient is starved, the amount of miR398 increases [59].
The availability of nitrogen affects the architecture of the root system through mir393, which was discussed earlier in the context of drought. The target of this miRNA can be auxin receptors (TIR1, AFB1, AFB2, and AFB3), as well as the transcription factor bHLH (basic helix-loop-helix) [60]. It is reported that mir393 is expressed under osmotic stress and, by cleaving its TIR1 and AFB2 targets, leads to blocking of the auxin signal and lateral root initiation [61]. Another miRNA whose expression is controlled by nitrogen availability is miR167. Its target is one of the auxin receptors, ARF8. The presence of transport forms of nitrogen (glutamate) is a signal for the suppression of miR167 expression, which removes the blocking of the auxin signal and leads to the initiation of the formation of lateral roots. miR167 expression is also downregulated under drought and high temperatures, while miR167 levels increase under cold stress. A complex regulatory network, including miR167, mir393, and ARFs, has been reported that regulates auxin-mediated signalling during osmotic stress (cited at [62]).
2.6. 5’-Untranslated regions’ unstructured features
The 5’ untranslated regions are extremely important for both the canonical and alternative translation processes in both prokaryotes and eukaryotes. Plants are no exception in this matter. The 5’-UTR is involved in translation control in two separate but related ways: 1) it serves as a container for specific regulatory elements [63], and 2) it acts on its own, even without distinguishable patterns, due to its structure and nucleotide composition. If everything is clear with the first type of determination, we will further consider in detail various regulatory elements and patterns localised in the 5’-UTR. However, there is no specific consensus on the role of the 5’-UTR per se. In this section, we will attempt to discuss what is currently known about the role of 5’-UTR in shaping the response to stress at the translation stage.
It is known that heat stress suppresses the translation of almost all genes. However, this rule does not apply to heat shock proteins, and their translation, contrary to expectations, increases. Among the factors determining translation under conditions of a radical increase in temperature is 5’-UTR. For the 5’-UTR of the Hsp81-3 protein gene, it was shown [64] that this untranslated region provides a scanning mechanism for translation initiation (to prove this, the authors integrated uAUG) for both capped and non-capped mRNAs; the level of reporter gene translation under the control of this 5’-UTR was also shown to be indifferent to temperature changes. Unfortunately, the authors do not provide a detailed mechanism for the described process.
A likely answer to the above question is a recent study [65], whose authors found (in HeLa cell culture) that -methyladenosine () in the proximal part of the 5’-UTR is able to directly bind eIF3, which, in turn, recruits the 43S complex. In fact, the initiation mechanism presented is an alternative to both Cap-dependent and IRES-dependent translation initiation mechanisms. In this work, it was found that this initiation mechanism is activated in response to stress factors, in particular heat stress, when the Cap-dependent translation mechanism is suppressed due to inhibition of the eIF4E factor. Despite the fact that the authors of this study demonstrated the pattern they discovered in HeLa cell culture, there is no reason to believe that such a mechanism will not work for plants. For A.thaliana, mRNA isoforms are shown that differ in the 5’-UTR and are characteristic of different temperature conditions. This, according to the authors of the study [11], may be due to the different stability of the 5’-UTR of such transcripts. A fairly striking example is the expression of the gene encoding the PSY protein (phytoene synthase), which is involved in the metabolism of carotenoids. For the mRNA of this protein, the presence of two isoforms of the 5’-UTR was demonstrated: long and short. The long one contains a stem-loop structure, which, according to the researchers, could potentially be a riboswitch that reacts to carotenoids or their metabolites. The short one does not contain such a secondary structure and therefore can quickly increase the PSY level, which is necessary in the case of the synthesis of abscisic acid under, for example, salinity [66].
The context of the proximal parts of the 5’-UTR may also affect the efficiency of translation under conditions of heat stress [67] and low temperatures [68] .
While it can be difficult to isolate specific regulatory regions, the inherent intrinsic characteristics of the 5’-UTR are length and GC content [69]. The authors of this study showed that the translateate (i.e., the predominant fraction of translated mRNAs) of rice (Oryza sativa) can be characterised by the predominance of short and GC-rich 5’-UTR sequences. In our opinion, it seems necessary to consider in detail the role of the length and nucleotide composition of the 5’-UTR in plants under the action of stress factors and without. At the same time, it is worth paying special attention to the development of methods that make it possible to assess the contribution of these factors (length and composition) and exclude previously unidentified ones, for example, areas with a developed secondary and tertiary structure. Going back also, we find it extremely interesting to be able to demonstrate the presence in plants of an alternative scanning mechanism for translation initiation involving -methyladenosine.
2.7. lncRNA
Natural antisense transcripts NATs (natural antisense transcripts) are a type of long non-coding RNA. NATs are transcribed from the non-coding strand of protein-coding regions of the genome and are involved in the regulation of transcription and translation of the complementary strand product (sense). In this case, they are called cis-NATs. If the target of NAT is mRNA translated from another gene segment, such molecules are called trans-NATs. Julia Bailey-Serres et al. showed the presence in the structure of some cis-NATs of open reading frames encoding small peptides. Translation from these frames, the so-called sORFs (small open reading frames), correlated with increased translation of the corresponding sense mRNAs. The authors suggest that translation of sORFs may stabilise or promote targeting of cis-NATs. The authors conducted studies on A.thaliana under conditions of phosphorus deficiency, showing that the genes subject to the above regulation include two representatives of the ATP-binding transporters of the ABC family (ABCG2 and ABCG20), as well as a representative of the receptor kinase family (POLLEN-SPECIFIC RECEPTOR-LIKE KINASE 7). These genes are involved in the regulation of the absorption of minerals as well as in the formation of lateral roots, which is one of the conditions for adaptation to phosphorus starvation [58].
A group of scientists led by Jinhui Chen, using third-generation sequencing (Iso-Seq) and high-throughput RNA sequencing, showed an increase in the content of transcripts of five genes of eIF2Ds translation initiation factors, as well as a number of long non-coding RNAs, including lncPs3, against the background of heat stress in Populus simonii (Chinese poplar). Using bioinformatic methods, the interaction of lncPs3 with eIF2D proteins was predicted. Thus, the authors suggest the involvement of long noncoding RNAs in the regulation of translation under heat stress in poplar [70].
An interesting study [71] demonstrated the involvement of an lncRNA called CRIR1 (cold- responsive intergenic lncRNA 1) in the response to cold stress in cassava (Manihot esculenta Crantz). Regulation by this lncRNA bypasses C-repeat-bindingin gene expression regulation via multiple ways, including transcriptional factors (CBFs), but due to the close interaction of this lncRNA with MeCSP5 (a cold shock domain-containing protein), the connection between this process of regulation and the process of translation is not entirely clear.
2.8. IRES
Most eukaryotic mRNAs have a cap structure at the 5’ end. The cap structure plays a key role in binding to ribosomes via the mRNA translation initiation factor eIF4F [72]. However, under conditions where cap-dependent initiation is impaired, such as by viral infection [73], apoptosis [74] or mitosis [75]; [76], an alternative translation initiation mechanism is enabled. Such mechanisms include ribosome access to mRNA through binding to an internal ribosome entry site (IRES) located at the 5’-UTR. IRES-driven translation initiation was first described for picornavirus RNA [77]. In cells infected with picornavirus, cap-dependent translation initiation is inhibited by proteolytic cleavage of the translation initiation factor eIF4G [78]. The sequences of the various IRESs are not conserved. For example, the IRES sequence of picornavirus and the IRES sequence of foot-and-mouth disease virus match by 50%. Heat stress is a condition in which overall cap-dependent translation is severely retarded while the translation of mRNAs encoding heat shock proteins (HSPs) functions at its maximum efficiency. Evidence suggests that stress activates internal initiation of gene translation (HSP) [79]. Some of the maize germ mRNAs are capable of cap-independent translation initiation during germination [80]. Hsp101 mRNA, encoding the heat shock protein, activates cell thermal stability under stress conditions and is present among maize germline mRNA transcripts.
The 5’-UTR sequence of Hsp101 maize cell mRNA functions as an IRES-like element responsible for cap-independent translation during heat stress. Removal of the 5’ untranslated region (UTR) from Hsp101 mRNA disables cap-independent translation. The study [80] used bicistronic constructs containing the 5´UTR of Hsp101 mRNA in the sense and antisense directions between two reporter genes. A similar bicistronic construct containing an element of the viral ribosome entry site (IRES) between the reporter genes was used as a control. Translation of the second reporter gene occurred when the 5’-UTR of Hsp101 was only in the sense orientation of the bicistronic construct. The study found that the 5´UTR mRNA of Hsp101 has the characteristics of a functional IRES element and is suitable for the regulation of translation not only in maize but possibly in other plants as well [80]. The study [81] also used a bicistronic system containing an internal ribosome entry site (IRES) sequence from the 5’-UTR of the tobacco heat shock protein NtHSF-1 to study the salt tolerance of tobacco plants. In [82], the element used in the bicistronic IRES construct, involving two reporter genes, gfp and uidA, showed functional activity in tobacco protoplasts and human HeLa cells.
The authors of [81] used constructs in which the wheat genes encoding vacuolar ion carriers, antiporter (TNHXS1), and -pyrophosphatase (TVP1) are linked through the IRES sequence [81]. The main goal of the cloning strategy was to physically link the TNHXS1 and TVP1 genes and position them side by side on DNA that would be passed as a single unit into the tobacco genome. Molecular analysis of transgenic tobacco plants revealed the integration of the TNHXS1-IRES-TVP1 construct into the tobacco genome and the synthesis of full-length bicistronic mRNA under the control of the CaMV 35S promoter. It was noted that transgenic lines showed better growth than wild-type plants under normal conditions. The results confirm the data that plant vacuolar -pyrophosphatase plays an important role in plant organogenesis by controlling the transport and distribution of auxin [83]. Overexpression of plant vacuolar -pyrophosphatase affects the development of shoots and roots in transgenic plants of tomato [84], cotton [85], corn [86], and A.thaliana[87]. A strong root system facilitates water uptake under abiotic stresses such as drought and salinity. Ectopic expression of TVP1 and TNHXS1 potentially activates proton pumps, -Pases, and V-ATPases. The activation of V-ATPase can further enhance the salt tolerance of plants. The resulting transgenic plants simultaneously expressing the TNHXS1 and TVP1 genes showed salt tolerance in comparison with control plants, both in vitro and in greenhouse conditions. The transgenic plants had a high content of chlorophyll and a high biomass under saline conditions in comparison with the control. Probably due to greater leaf turgor as a result of a balanced osmotic potential. Thus, the obtained results demonstrate that the IRES sequence can coordinate the expression of genes involved in resistance to abiotic stresses [81].
Totipotent, or undifferentiated, plant cells are hypersensitive to environmental fluctuations throughout their life cycle. The mechanism by which plants protect totipotent cell homeostasis in response to environmental hazards has not been fully established. The WUSCHEL (WUS) transcription factor homeodomain maintains a pool of totipotent cells in the shoot apical meristem of A.thaliana. It was found that the translation of WUS mRNA is controlled by IRES in the 5’-untranslated region [88]. The AtLa1 protein is an RNA-binding factor that interacts with the 5’ UTR and initiates IRES-dependent translation of WUS mRNA. Knockdown of AtLa1 expression suppresses WUS IRES-dependent translation and stops plant growth and development. The AtLa1 protein is mainly located in the nuclear nucleoplasm. However, exposure to a stressful situation can determine the nuclear-cytoplasmic translocation of the AtLa1 protein, which further enhances the IRES-dependent translation of WUS mRNA. Genetic data indicate that the WUS protein increases the resistance of the shoot apical meristem to abiotic fluctuations in environmental conditions. Based on the obtained results, it was concluded that undifferentiated A.thaliana cells, in response to negative environmental influences, activate IRES-dependent translation of WUS mRNA under the control of the AtLa1 protein [88].
In the study [89], transgenic tomato plants overexpressing – antiporter genes (TNHXS1) were obtained either alone or with -pyrophosphatase (-PPiase) using the IRES element in a bicistronic design. Transgenic tomato lines co-expressing both genes (LNV) showed higher salt tolerance than wild-type lines or lines expressing only the TNHXS1 (LN) gene. Under conditions of salt stress, transgenic plants increased biomass more efficiently and retained high levels of chlorophyll and catalase (CAT). Also, the translation of antiporters and pyrophosphatase coupled in the transformation vector determined the early flowering of tomato plants and the production of more fruits in comparison with control plants.
Intracellular NHX proteins are and antiporters involved in homeostasis and pH regulation. Proteins also play a major role in salinity tolerance [90]. The study overexpressed the NHX TNHXS1 gene alone and in combination with the TVP1 pyrophosphatase (-PPiase) gene using the IRES element in a bicistronic vector. Both genes were isolated from common wheat (Triticum aestivum L.). The work achieved an improvement in salt tolerance and an increase in the content of in the leaves and fruits of transgenic tomato plants. An important feature is the formation of lateral roots, which allow plants to adapt to abiotic stresses such as drought and salinity. Plant vacuolar -pyrophosphatase has previously been shown to play an important role in regulating root formation by controlling auxin transport and distribution [83]. As a result of the work, it was found that the coordinated expression of the NHX and VP1 genes and their translation using the IRES element determines an effective strategy not only for increasing the salt tolerance of tomatoes but also for saturating this vegetable crop with potassium [89].
2.9. Leaderless mRNAs
Of particular interest are leaderless, i.e., devoid of 5’-UTR, mRNAs. Originally discovered in eubacteria [91,92], they were later found in archaea and eukaryotes. Given the need for a 5’-UTR for the main mechanism of translation initiation in both prokaryotes and eukaryotes, it would seem that such a model would not work. But it was shown that in, for example, E. coli, in cases of cold stress, leaderless mRNAs gain an advantage in translation (apparently due to the lack of a developed secondary and tertiary structure before the starting AUG) [91]. For mammalian mitochondria (which are known to be of prokaryotic origin), translation of leaderless mRNAs has also been found [93,94]. The need for participation in this process by only the translation initiation factors IF-2mt and IF-3mt was shown; other factors are not necessary. Nuclear-derived leaderless mRNAs can bind to the 80S ribosome-Met-tRNA complex in mammalian cells in the absence of any translation initiation factors [95,96])
If we talk about representatives of the plant kingdom (Viridiplantae), similar data were obtained only for mitochondria of Chlamydomonas reinhardtii [97]. In this organism, eight protein-coding genes are transcribed as two polycistronic transcripts, which are subsequently hydrolyzed in such a way that transcripts are formed that begin with a start codon and have an extended 3’-UTR. The authors of this study showed that such mRNA molecules are looped, forming a kind of pseudo-5’-UTR. The study of leaderless RNAs still requires additional large-scale experiments, forming the following open questions: Are the patterns found in mammalian and bacterial cells applicable to plants? How common are leaderless mRNAs in higher plants? Do they gain an advantage when broadcasting under stressful conditions? What is the situation with the rate of degradation of leaderless mRNAs? Are there mechanisms that protect leaderless mRNAs from rapid degradation?
2.10. uORFs
uORFs (upstream open reading frames) located within the 5’-untranslated region (5’-UTR) of some plant mRNAs can regulate the translation of the main open reading frame (mORF), which encodes the main protein product of the transcript [98,99,100,101]. uORFs regulate the translation of the underlying core ORF through small protein signalling molecules. In plants, uORFs were found in 24–30% of the 5’-UTR region of mRNA [63,102]. The uORF prevalence approximation is based on the use of AUG as the start codon, but ribosome profiling studies show that non-canonical codons (eg, ACG) in uORF can also serve as start codons [103].
In response to abiotic stresses such as drought, salt stress, or low temperatures, gene expression can change in plants. uORF may play an important role in this process by controlling the translation of mORF (main ORF). Typically, uORFs inhibit translation initiation at the underlying CDS coding sequence. The inhibitory mechanism of uORF is explained by the fact that in eukaryotes, the 43S pre-initiation complex binds to the 5’-cap structure of mRNA, processively scans in the 5’-3’ direction, and initiates translation at the first encountered initiation codon, which is in the optimal environment [104]. The 43S preinitiation complex consists of several factors, including eukaryotic initiation factor (eIF)-3, eIF1, eIF1A, the eIF2/GTP/Met-tRNAiMet ternary complex, and the 40S ribosomal small subunit [105]. Dissociation of the eIF2 ternary complex and other initiation factors during uORF translation is hypothesised to cause repression of subsequent translational re-initiation in downstream CDSs [106]. uORF-mediated inhibition of ORF translation can occur either passively, in the event of ribosome dissociation after uORF translation, or actively, in the case of ribosome arrest caused by uORF translation [100,101]. The arrest may isolate translating ribosomes, blocking their access to downstream ORFs, or may be recognised as an abnormal translational termination triggering transcript destruction via the nonsense-mediated decay (NMD) mRNA mechanism [107,108]. In each of these cases, translation of the downstream mORF is inhibited.
The use of various approaches has revealed a significant number of genes with uORF-encoded peptides that are highly conserved among different representatives of eukaryotes, including plants. These uORFs have been independently termed upstream conserved coding regions (uCCs) or conserved peptide uORFs (CPuORFs). The uCC nomenclature was first used to describe elements in AZIN1 and ODC mRNA in many organisms, from fungi to mammals [109]. uCCs encode conserved polypeptides; they generally do not have canonical AUG start codons and are initiated by a related start codon. The term CPuORF is more commonly used for plants [110]. CPuORFs are usually initiated by the AUG start codon. Relatively few of the plant CPuORFs have a known biological function; those characterised are involved in the translational regulation of mORFs in a largely metabolite-dependent manner. Below are examples of plant transcription factors such as bZIP (basic leucine zipper) and bHLH (basic helix-loop-helix) and organic compounds such as ascorbate (vitamin C) and polyamines (PA), the induction of gene expression of which is somehow accompanied by abiotic stresses and whose translation is controlled in a CPuORF-dependent manner. The main and best studied example of metabolite-controlled conserved peptide-dependent regulation of uORF translation in plants is the bZIP transcription factor of the A.thaliana C/S1 group (bZIP1, bZIP2/GBP5, ATB2/bZIP11, bZIP44, and bZIP53).
The bZIP transcription factors control overall metabolism as well as amino acid and sugar metabolism. Translation of their mRNA is regulated by sucrose concentration [111]. It was sucrose that was found to cause ribosome arrest during uORF translation [112]. Ribosome arrest is specific to sucrose, while other sugars, including d-fructose, d-glucose, UDP-d-glucose, and sucrose-6-phosphate, do not cause ribosome arrest. S1-bZIP plays an important role in plant adaptation to adverse conditions [113,114]. It has been confirmed that S1-bZIP plays an important role in plant innate immunity, especially against attacks by various pathogens [113,115], as well as in response to abiotic stresses such as low temperatures [116,117], drought [118,119] and salinity [119]. It has been demonstrated that the C-/S1-bZIP-SnRK1 complex is involved in the reprogramming of primary metabolism associated with carbohydrates and amino acids and induces tolerance to salt stress through ABA-independent signalling in A.thaliana roots [119].
In plants, polyamines play an important role in adaptation to adverse conditions and stress resistance in general. The influence of polyamines on the adaptation of various plant species to various abiotic stresses, such as drought [120,121], salt stress [122], high temperature, and cold stress [123]. Polyamine homeostasis is tightly regulated in plants. A key target of polyamine-regulated translational control in many eukaryotic systems is AdoMetDC (AMD1) [124]. The 5’ leader of plant AdoMetDC mRNA contains two conserved overlapping uORFs, tiny and small. The tiny uORF consists of only three codons. The small uORF is in the range of 40–65 codons and shows greater conservation at the amino acid level. The C-terminus of a small uORF always ends with PS [125]. Overexpression of AdoMetDC mRNA using a construct in which uORFs are eliminated by mutation of start codons leads to an increase in the enzymatic activity of AdoMetDC, disruption of polyamine homeostasis, and abnormal plant growth and development [126].
Thermospermine is a structural isomer of spermine. In A.thaliana, the production of thermospermine is catalyzed by the enzyme thermospermine synthase (ACL5), which converts spermidine to thermospermine. ACL5 loss-of-function mutants cause severe stem elongation defects due to excessive proliferation of xylem vessels [127]. SAC51 encodes the basic transcription factor helix-loop-helix (bHLH). Sac51-d is the dominant mutation in SAC51 [128]. The sac51-d mutation is a premature termination codon in one of several uORFs in SAC51 mRNA. Mutated uORF in sac51-d is highly conserved at the amino acid level in plants [129]. In the sac51-d mutant, mORF translation is increased relative to wild type, as determined in reporter experiments, indicating that uORF inhibits downstream CDS translation [128]. Plant studies have shown that thermospermin suppresses the expression of reporter genes containing the wild-type SAC51 leader [130]. Whether thermospermin exerts its action directly through uORF (during its translation) or works indirectly remains to be explored.
Ascorbate (ASC) plays a critical role in the removal of reactive oxygen species by plants under abiotic stress conditions such as intense light, salinity, and drought. These stresses can cause ROS accumulation in plants, changing the cellular composition and disrupting growth and development [131,132]. As an antioxidant, ASC can effectively remove accumulated ROS and is thus critical for repairing oxidative damage and increasing plant resistance to abiotic stress [133,134]. The amount of ASC in a cell is regulated by GDP-1-galactose phosphorylase (GGP); part of the regulation of GGP is at the transcriptional level; GGP mRNA is also translationally regulated by ascorbate in the autoregulatory loop, with high levels of ascorbate resulting in suppression of translation of GGP mRNA. This translational regulation is mediated by uORF, which is initiated by the ACG start codon. The uORF-encoded peptide is conserved in all plants and eukaryotic algae.
Experiments with reporter genes and transformation of A.thaliana cells show that uORF represses mORF GGP expression and that this repression is enhanced in the presence of ascorbate. The same experiments also identified a conserved histidine residue located 20 amino acids upstream of the AGGG motif, which is required for both repression and regulation of ascorbate. Mutation of the ACG start codon of uORF to the AUG codon results in constitutive repression of mORF expression [135]. Inactivation of uORF GGP in A.thaliana by homozygous modification by CRISPR results in plants with over 70% higher levels of ascorbate, confirming the involvement of uORF in vitamin C homeostasis [136].
2.11. Riboswitches
Thiamine pyrophosphate (TPP) riboswitches are the only type of riboswitches described for eukaryotes [137,138]. TPP is a substance derived from vitamin B1, which is part of the most important enzymes, including oxidative decarboxylation. Since TPP is part of the enzymes involved in perception and response to abiotic influences, the regulation of TPP production in plants during stress is carried out through the key enzymes of its biosynthesis. It is assumed that a decrease in TPP, which is actively involved in the synthesis of functional enzymes under stress, can enhance its production in the cell through a feedback mechanism using a riboswitch. Plant TPP riboswitches are located in the 3’UTR mRNA of one of the key genes for the synthesis of TPP phosphomethylpyrimidine synthase AtTHIC (THIAMINE C SYNTHASE). This distinguishes them from the corresponding bacterial and fungal riboswitches located in the 5’-UTR of this gene. In the absence of TPP, the riboswitch is in a conformation that shields the alternative splicing site at the 3’ end of the mRNA, resulting in the preservation of the polyadenylation signal (position U370) in the mature mRNA. When the concentration of TPP begins to exceed the threshold value, its excess binds to the riboswitch, which causes a change in its conformation and entails the exposure of an alternative 3’ splicing site. The availability of this area for the spliceosome leads to the removal of the standard polyadenylation signal (U370). This produces a longer mRNA with polyA tails in several positions (between U531 and C945), which makes it unstable, and AtTHIC translation does not occur, thereby leading to a decrease in TPP synthesis [138].
2.12. RNA thermometers
RNA thermometers are sections of mRNA that can change their secondary and tertiary structures when the temperature changes in the physiological range. These regulatory elements have been studied in most detail in prokaryotes. In particular, for bacteria, the fundamental possibility of using almost any biological molecules as temperature sensors has been shown: DNA, RNA, proteins, and lipids [139,140]. Bacterial RNA thermometers perform a wide range of functions, including: induction of genes responsible for invasion (in the case of warm-blooded pathogens); sense of quorum; adaptation to temperature increase; adaptation to lower temperatures; formation of biofilms; transition from lysogenic to lytic cycle in bacteriophage [141].
For animals, the situation with the knowledge of RNA thermometers looks much more modest [142]. Of the regulatory elements acting directly as thermosensors, we can mention the 5’-UTR mRNA encoding the heat shock protein 90 (HSP90) in Drosophila, as well as the non-coding RNA Hsr1 (which will be discussed below). Along with this, in Drosophila, the dependence of the intensity and site-specificity of RNA editing on temperature was demonstrated [142].
The study of plant RNA thermometers is at an early stage. Review papers [138] and [139] demonstrate an approach based on the suggestion of similarity of the molecular mechanisms in plants, prokaryotes, and non-plant eukaryotes (mammals, protists, and insects). One of the first discovered RNA thermometers in plants is the hairpin located in the 5’-UTR of the PIF7 transcription factor. PIF7 is involved in the activation of genes responsible for the response to heat stress in A.thaliana. PIF7 also directly interacts with PIF4 (PHYTOCHROME INTERACTING FACTOR 4), which, in turn, is involved in the control of auxin synthesis. A similar hairpin has also been described for the transcript encoding the transcription factor WRKY22 and for HSFA2, which in turn controls the expression of heat shock proteins [143]. Hsr1 is a short, conserved RNA that controls the expression of heat shock proteins. The papers [144] and [142] found in a wide variety of organisms. At present, it seems to be a consequence of horizontal gene transfer, or contamination. Since the Hsr1 gene has been shown to have a sequence identical to the bacterial voltage-gated chloride channel protein.
At first glance, two different mechanisms underlie the functioning of RNA thermometers in prokaryotes and eukaryotes. The former are characterised, as a rule, by the binding of the Shine-Dalgarno sequence (SD) and the start codon (AUG) as part of the stem-loop structure; in eukaryotes, hairpins prevent the implementation of the scanning mechanism of translation initiation; the hairpin mechanically prevents the movement of the complex of the small subunit of the ribosome and translation initiation factors from the 5’-Cap to the start codon surrounded by the Kozak consensus sequence. In an experiment to study the structure of rice transcripts, only a few candidate sequences for the role of RNA thermometers were found. And none of them melted at 42C [145].
Information on the role of RNA thermometers localised in the 3’-untranslated region in the regulation of translation is extremely scarce. So for Leishmania, the presence of a structured region in the 3’-UTR was demonstrated, which undergoes partial melting with increasing temperature in the range of 26-37C. Presumably, this may facilitate the cyclization of the mRNA molecule and the subsequent reinitiation of translation at elevated temperatures [146].
At the present stage, the topic of RNA thermometers in the context of plants raises more questions than it answers. In particular, the question of the degree of representation of RNA thermometers in higher plant mRNAs is of great interest. If the answer to this question is negative, then it is necessary, in our opinion, to clarify the mechanisms that prevent such a scheme of translation regulation. If the answer is yes, then what are the algorithms for finding such patterns? Can there be specific RNA sensors sensitive to temperature decreases? Potentially, two ways are possible in this situation: first, such thermometers change their structure with decreasing temperature to one that is more passable for ribosomes but with higher potential energy (for example, two low-molecular hairpins instead of one high-molecular hairpin, similar to the tryptophan operator); or as mRNA structures that block the expression of genes that themselves act as suppressors when the temperature is lowered. A separate fact that needs clarification is whether plant RNA thermometers are integrated into larger regulatory regions (for example, IRES). Can RNA thermometers be involved in the process of RNA interference?
2.13. Plant ribosomes also act as an independent sensor
The authors of a recent study showed that a decrease in the rate of translation under the influence of cold (and cycloheximide) leads to an increase in the concentration of calcium ions in the cytoplasm and nucleus. An increase in the concentration of , in turn, induces the expression of the CAMTA complex, which already triggers the expression of the CBF gene responsible for overcoming cold shock. The participation of ribosome proteins in this signalling pathway has been demonstrated, which suggests that ribosomes can act as independent thermosensors. Ribosome-inactivating proteins are able to release adenine from RNA and DNA molecules [147]. Models of Hura crepitans, Phytolaeca americana [148] and Mesembryanthemum crystallinum [149] showed that the content of these proteins increases in ageing leaves and leaves subjected to heat or osmotic stress. Changes in the population of ribosomal proteins during acclimation after cold stress (from days 1 to 7) were shown in an A.thaliana model using mutants for the REIL (Rei-like) protein gene [150].
2.14. G-quadruplexes
G-quadruplexes are secondary and tertiary structures within mRNA (and DNA) based on non-Watson-Crick interactions between successive guanines. This area of research is currently developing very dynamically [151]. Initially, G-quadruplexes are considered regulatory elements active at the transcription stage. However, over time, data are accumulating about the important role of G-quadruplexes in the regulation of translation, too. Moreover, like many regulatory elements, GQS (G-quadruplex sequence) modulates translation in two ways. Typically, GQS suppresses 5’-UTR ribosome scanning. An alternative way is to additionally stabilise the GQS of various regulatory elements with a developed secondary and tertiary structure (IRES, ribiswitches, etc.). GQs can also serve as binding sites for various proteins and small molecules. Such proteins can both (fragile X mental retardation protein (FMRP)) stabilise the structure of G-quadruplexes (suppressing translation) and act as specific helicases (RHAU/DHX36), on the contrary, they can increase the intensity of translation by reducing the complexity of the mRNA structure [152]. An example [153] for plants is the regulation of the gene encoding the SUPPRESSOR OF MAX2 1-LIKE4/5 (SMXL4/5) protein. This protein is responsible for the development of the phloem in vascular plants. The authors of this study, using Populus tremula and A.thaliana as examples, showed that SMXL4/5 contains a 5’-UTR sequence capable of forming GQS. Another zinc-finger protein, JULGI (JUL), binds to this region as part of the 5’-UTR and induces the formation of a G-quadruplex (i.e., in the native version, without JUL, it is not formed), which inhibits the translation of SMXL4/5. In silico analysis (using BLASTp and FIMO) identified over 400 proteins potentially capable of binding to GQS [154].
The situation with the interaction of small molecules and G-quadruplexes is similar to the situation with proteins. A number of compounds can additionally stabilise the structure of GQSs; other substances, on the contrary, cause the breakdown of these mRNA structures and increase their permeability for ribosomes. In this case, G-quadruplexes can act as riboswitches. In addition, synthetic aptamers can be constructed based on sequences containing GQS [155]. The stability of GQSs is also determined by their dehydration, i.e., its increase can be facilitated by an increase in the concentration of ions or osmolytes in the cytosol. An equally intriguing hypothesis was put forward by the authors of the study [156]. In their work, they showed the ability of ultraviolet radiation to destabilise GQS under in vitro conditions. If such a mechanism occurs in living plant cells, it can act as a potential sensor responsible for the response to increasing solar radiation and related stress conditions (drought and temperature stress).
GQS have also been noted in the context of liquid-liquid phase separation in A.thaliana SHORT ROOT (SHR) [157].An active involvement of SHR liquid-liquid phase separation mRNA was demostrated. Also, for the mRNA of this gene, the formation of a G-quadruplex was demonstrated under certain conditions (with a change in the concentration of potassium ions). Using normal and mutant variants of the mRNA sequence, the authors of this work showed the involvement of GQS in the process of phase separation. Given that phase separation can occur during stress response (in the form of stress granule formation, for example), this study opens up another new avenue of research—it is unlikely that this G-quadruplex alone is involved in phase separation. A new stage in the study of G-quadruplexes has been the use of genome-wide and whole-transcriptome approaches based on the sequencing of all transcripts and subsequent screening of potential GQSs (and sometimes verification of individual candidate sequences). Full translational analysis revealed the putative involvement of G-qualruplexes in the regulation of translation under heat stress in A.thaliana [158].
Genome-wide analysis (on the A.thaliana model) using Markov models in combination with GO-enrichment analysis showed significant changes in some gene groups: Overrepresented: catalytic activity, protein amino acid phosphorylation, transferase activity, etc. Underrepresented: translation, gene expression, RNA binding, etc. [159]. Potential GQSs associated with responses to various types of abiotic stress (including hypoxia and nutrient deficiency) have also been found in the maize genome. Quite curious is the presence of GQS in the antisense strand in the 5’-UTR and at the 5’ end of some genes responsible for acclimation under these stress conditions (a similar pattern is also characteristic of the wheat genome [160]). However, GQSs that directly regulate broadcasting have not been identified [161]. A genome-wide search for potential GQSs using barley as an example showed an increased representation of these motifs within 1Kb of the start codons of genes (300 per 1e6 versus 100 per 1e6 for the entire genome) [162] . These data are also confirmed by experimental work carried out on the wheat genome [160].
The widespread use of in silico methods, as can be seen from the above works, can be explained by the availability of these methods for direct use. Nevertheless, we see it as necessary to develop evidence-based methods using molecular cloning tools and ome-wide techniques to detect both GQS directly in transcripts and protein-GQS interactions. An interesting approach was proposed in a study [163]. It is based on the obstruction of the area containing GQS for RNA-dependent DNA polymerase. Thus, using specific primers for reverse transcription, it is possible to detect the presence of a formed G-quadruplex in mRNA. Modulation of the GQ structure is based on the selectivity of this motif for potassium ions, i.e., in the presence of a G-quadruplex is formed, but not in the presence of [163]. This approach has been further developed and improved for full transcriptome search and GQS analysis [164,165]. Along with this, there is a method based on the precipitation of RNA containing GQS and further sequencing of these molecules [166].
Considering that most of the studies on the contribution of G-quadruplexes to the regulation of translation have been carried out in animals, it is reasonable to consider the role of GQS in plants by analogy. However, additional avenues for future research can be identified: (i) the study of GQS-binding proteins in plants (ii) Distribution of GQS among differentially expressed genes under stress that provokes the accumulation of osmolytes (cold, drought) (iii) Prospects for using GQS as riboswitches in plants, in particular as aptamers for specific low molecular weight ligands and as RNA thermometers (i.e., the stability dynamics of these elements under in vivo conditions at varying temperatures is interesting).
2.15. Kozak consensus sequence
One of the key steps in the control of gene expression (as mentioned above) is the mechanism of regulation of translation initiation, in which the sequence context around the start codon AUG, which forms the translation initiation site (TIS), plays an important role. To date, the main model of translation initiation is considered to be the model described by Marilyn Kozak, according to which the 40S ribosomal subunit associated with the 5’-terminal region of mRNA, together with a number of translation initiation factors, begins a linear scan along the mRNA until it reaches the start codon, most often AUG [167]. It is generally accepted that successful recognition of the AUG start codon by eukaryotic ribosomes depends on its nucleotide environment. Thus, in the 1980s, Kozak conducted research in which 699 vertebrate mRNAs were analysed, resulting in the optimal TIS consensus sequence, known as the Kozak motif gccgcC(A/G)ccAUGG, where AUG represents the start codon. In this case, the position of the purine at -3 (97% mRNA) and G (61%) at position +4 (where A in AUG has position +1) forms the optimal sequence for TIS recognition and subsequent translation initiation [168].
In addition, sequences surrounding the start codon were classified based on the presence of two conserved nucleotides at positions -3 and +4. So, the Kozak motif NNN(A/G)NNAUGG (where N is any base), which contains both important nucleotides, is called "strong", while the sequence GCC(A/G)CCAUGG is usually called "optimal", In turn, NNN (A/G)NNAUG(A/C/U) or NNN(C/U)NNAUGG is "adequate" because it contains only one of these nucleotides; "weak" is the sequence NNN(C/U)NNAUG(A/C /U) that does not contain any of the key nucleotides [169].
Further studies of other taxonomic groups, including plants, expanded the understanding of the consensus sequence surrounding TIS [170]; however, significant differences were observed between taxa, which differed from the initial beliefs about the universality of the consensus Kozak for all eukaryotes. In studies by Gupta et al., a comparative genome-wide analysis of 14 plant species (seven monocots and seven dicots) was conducted, in which 500 genes with the highest protein content and 500 genes with the lowest protein content were taken. The results obtained demonstrated a direct relationship between highly conserved nucleotide positions at positions -3A/G and +4G and high protein levels in monocots and dicots. In addition, the C at position +5 also showed an increase in translation efficiency, in contrast to Kozak’s early data, where the nucleotide at position +5 did not enhance translation initiation [171]. Also, a comparative analysis revealed a significant predisposition of the A/C nucleotides in position -2 to highly expressed genes of both monocots and dicots. Interestingly, the -2A/C position is found in the TIS consensus sequences of chordates, invertebrates, unicellular fungi, and protists, which makes it rather conserved [172]. As a result, the sequences GCNAUGGC, AANAUGGC, and GCNAUGGC were established as the archetypal TIS signals in monocots, dicots, and plants in general, respectively [173].
The regulation of gene expression at the translational level enables cells to elicit rapid and reversible responses to sudden environmental changes. At the same time, the use of Kozak sequences capable of increasing the level of gene expression opens up a wide range of possibilities for practical application. Thus, in studies aimed at increasing the resistance of Oryza sativa L. to dehydration stress, Zhou et al. produced transgenic rice plants expressing the fungal gene for glutamate dehydrogenase, which can act as a stress-responsive enzyme to detoxify high intracellular ammonia and produce glutamate for proline synthesis under stress. In order to increase the level of translation of eukaryotic genes in transgenic rice plants, the Kozak sequence (GCCACC) was added before the MgGDH start codon. As the authors note, as a result of a high level of expression of the MgGDH gene, transgenic rice plants demonstrated an increase in tolerance to dehydration stress [174]. The data obtained are consistent with previous results demonstrating a high level of expression of genes containing strong Kozak sequences [173].
However, there are studies demonstrating that site-directed mutagenesis experiments with the Kozak sequence can tune the physiological response to stress. Thus, a recent study showed that, in response to stress, Saccharomyces cerevisiae produce ribosomes lacking certain proteins (for example, Δ Rps26), which initiate the activation of certain signalling pathways (Hog1 and Rim101) that respond to osmotic stress and pH stress, respectively, as a result of which cells are more resistant to high salt and high pH conditions. At the same time, Rps26 preferentially initiates translation of mRNAs containing A in positions -2 and -4, while mRNAs containing G in positions -2/-4 were translated using ΔRps26 [175]. It is also noted that ΔRps26 does not activate the cell wall integrity (CWI) pathway because the Rho1 gene of the CWI pathway contains an A at position -2/-4. As a result of site-directed mutagenesis, a mutant Rho1 containing -2/-4G was obtained. At the same time, cells containing modified Rho1 showed increased resistance to zymolysis with depletion of ΔRps26, in contrast to cells containing native Rho1. Also, this site-directed mutagenesis method was applied to the Rnr2 and Ras2 genes of the DNA damage response and filamentation pathways, respectively. As before, the modified genes turned out to be sensitive to the accumulation of ΔRps26 in the cell, in contrast to the native ones. Thus, by introducing point mutations in the Kozak sequence of individual genes, the authors managed to reprogram the cellular response to severe salt stress, including the pathways of filamentation, activation of DNA repair, and strengthening of the cell wall [176].
The relationship between the environment of the start codon and the regulation of translation under stressful conditions in plants is not obvious. It is difficult at the moment to give an unambiguous answer as to whether this connection can take place in principle. Given that the environment of the start codon is rarely modified. Nevertheless, several principal ways can be outlined for further study of this issue: the first is an analysis of the representation of different types of Kozak consensus sequences at all stages of the realisation of genetic information, and enrichment (enrichment) and bias (bias) in the frequencies of these types under stress. No less interesting is the presence of alternative start codons and the extent to which they are used under stress. The third way is the study of epigenetic modifications of nucleotides in the environment of the start codon in plants.
2.16. Codon usage
Codon bias is a well-known phenomenon that consists of the preference for some synonymous triplets over others in protein coding sequences. The connection between this phenomenon and the control of translation is quite obvious due to the different concentrations of transfer RNAs and the different frequencies of the synonymous codons themselves. Some kind of independent, non-competing channel of translation is formed. Under conditions of heat stress in A.thaliana plants, optimal and conservative codons change the rate of translation elongation, thereby affecting the global translation of host genes. Based on this, it can be concluded that functionally important codons/ sites, or genes are usually involved in the control of translation in response to heat shock [177]. It is most likely that this interaction of factors was formed by natural selection in plants during the process of evolution.
The study [178] established the location of salinity-sensitive genes in rice, which are located on chromosomes 1–6. Preference for codon usage has been suggested to affect expression levels in response to environmental stimuli [179]. Most of the analysed genes involved in the mechanism of rice salt tolerance showed a high degree of homology (> 50%) in the use of CUB (codon usage bias) [179]. Another study of codon usage patterns found that different rice abiotic stress sensitivity genes widely use different synonymous codons. They also have a balanced ratio of AT-GC in the third nucleotide position of the codon [180].
Mohasses et al. studied the correlation between the use of synonymous codons (SCU) in drought conditions in rice plants [181]. An evaluation of 60 genes with different expression efficiencies associated with stress was carried out. Data selected from a meta-analysis of the use of various rice codons. The following indices were taken into account, the effective number of codons used, the effective number of codon usage (ENC), the codon adaptation index (CAI), and the relative synonymous codon usage (RSCU). It was found that for highly expressed genes under drought conditions, the GC content was higher, the ENC value was lower, changes in the preferred codons of individual amino acids were determined, and the ratio of RSCU of GC-terminal codons relative to AT-terminal codons for 18 amino acids increased significantly compared to other genes. To denote this phenomenon, the abbreviation The average ratio of RSCUs ARSCU was introduced as the average ratio of RSCU GC-terminal codons to AT-terminal codons in each gene, which can be divided according to the degree of expression during drought. To predict the expression level of rice genes during drought, the ARSCU-Calculator software was developed. An ARSCU index above the cut-offs indicates that the gene may belong to the "highly expressed gene group under drought conditions". The data obtained can be used to optimise codons in the genetic engineering of rice. To confirm the findings, another 60 genes were used (randomly selected from 43,233 genes studied for their response to drought). In this evaluation, the ARSCU value was able to predict the level of expression in 88.33% of cases. When comparing a third set of 60 highly expressed non-drought genes, only 31.65% of the genes showed an ARSCU value above the set threshold. The obtained results indicate that the revealed pattern may be unique for genes associated with drought. As a rationale for the correlation between CUB and high gene expression under drought conditions, a possible role of post-transcriptional modification of tRNA and tRFs (transfer RNA-derived fragments) underlying the biological mechanism has been hypothesised [181].
Another plant study [182] aims to investigate heat shock proteins in the tomato genome by analysing the codon abundance of various amino acids. The focus of this work is on the selected HsfA sequence (Accession No. NM 001247342.2 cultivar Heinz 1706 NCBI) with 3534 base pairs. The HsfA sequence has a transcription activation domain for resistance to temperature rise and other abiotic stresses. The codon adaptation index (CAI) value corresponded to 0.80 in the sequence NM 001247342.2 based on the analysis of the mRNA of tomato heat shock proteins. The amino acids asparagine (AAT, AAS), aspartic acid (GAT, GAC), phenylalanine (TTT, TTC), and tyrosine (TAT, TAC) have relative synonymous codon usage (RSCU) values above 0.5. In modified relative codon bias (MRCBS), high expression of amino acid genes under heat stress corresponded to histidine, tryptophan, asparagine, aspartic acid, lysine, phenylalanine, isoleucine, cysteine, and threonine. RSCU values corresponding to less than 0.5 were considered rare codons affecting the rate of translation. As a result of the analysis, it was shown that the RSCU, RCBS, and MRCBS indices can be used to predict gene expression at various stages of tomato growth [182].
Wu et al. established the use of optimal codons and the frequency of occurrence of amino acids in the cell that affect the rate of protein synthesis [183]. It was found that the use of certain codons largely determined protein expression at various temperature manifestations of heat stress, from 30 to 40C. Codon usage may play an important role in the active translation of proteins, especially for the codon (AAG) of the amino acid threonine [183].
In [177], an extensive assessment of the effect of heat shock on the functioning of translation in A.thaliana plants was carried out. The following indices were evaluated: CUB, (tRNA adaptation index) tAI, nitrogen cost, length identity of CDS sequences, contents of CDS GC sequences, and (minimum free energy) MFE per 1 kilobase of sequence. CUB, tAI, nitrogen cost, and CDS identity are evolutionary features of an organism, while length, GC content, and MFE are structural features. CUB is a deviation in the use of a synonymous codon. The tAI index assesses the availability of tRNA synonymous codes. The nitrogen cost index is the cost of biosynthesis for a synonymous codon. The identity index represents the similarity between the sequences of orthologous proteins and indicates the level of conservation. Index MFE values are negatively correlated with the stability of secondary structures. The study carried out a paired correlation analysis between the mentioned indices. To investigate the contribution of each function to the (translation efficiency) TE and eliminate potential errors among the studied functions, multiple regression analysis was carried out. Based on the analysis, it was determined that CUB partially determines the distribution of the source (similar to tRNA) during the decoding process and also determines the level of gene preservation. Genes with high CUB content are translated prior to heat stress. It has been found that genes with high MFE are less folded in the secondary structure and more sensitive to high temperatures. With an increase in temperature, the efficiency of translation of such genes sharply decreases. Genes whose products have a high cost of biosynthesis cease to be translated under stress. Thus, nitrogen cost is negatively correlated with ΔTE under heat shock. Uncertainty in the field of codon usage studies lies (in our opinion only) in (i) the identification of more formalised patterns in codon preference patterns. (ii) Intriguing and not fully understood is the existence of stable translation channels formed by preferred codons. (iii) If such channels exist, what is their capacity?
2.17. Global changes in plant mRNA structure under abiotic stress
Global changes in mRNA structure are thematically closely related to the general characteristics of 5’-untranslated regions. Quite curious is the approach based on the full transcriptomic analysis of double-stranded elements in the structure of transcripts [145]. The essence of the approach proposed by the authors, Structure-seq2 - is to methylate in vivo adenine and cytosine as part of single-stranded mRNA regions. Such methylated bases cause the synthesis reaction to stop during sequencing. The researchers also took into account the fact that methylations will be avoided by bases not only in the composition of double-stranded RNA regions, but also protected by proteins associated with nucleic acids. PIP-seq was used to clarify this issue. In the present study, it was found that mRNAs are actively degraded under heat stress, and this process is closely related to the melting of the secondary structure of these molecules. Also, it was not found (which in itself does not mean the absence) of mechanisms for detecting an increase in temperature similar to prokaryotic ones.
Analysis of the textitA.thaliana translateatome under conditions of prolonged heat stress revealed the predominance of genes translated under these conditions whose mRNAs have a weakly structured or unstructured 5’-UTR, especially in the vicinity of the start codon [158]. The same study showed that a significant proportion of upregulated genes have consensus sequences in the 5’- and 3’-UTRs. A small number of works in the declared area do not allow for drawing any far-reaching conclusions. Apparently, we have yet to see systematic studies on global changes in the structure of mRNA.
3. Methods for studying translation in plants
The study of translation in plants is a rather diverse process. First of all, as in the study of transcription, it is necessary to determine the frequencies of genes in the composition of the transcriptome and translatome. Depending on the objectives of the study, it may be necessary to determine the secondary and tertiary structure of RNA, protein binding sites, and the structure of certain already known regulatory elements in the transcriptome.
3.1. Analysis of the involvement of mRNA in the translation process
Ribosomal profiling [184,185,186]) makes it possible to identify a subset of polysome-bound mRNAs as well as sites of messenger RNA molecules preferred by ribosomes (regions with rare codons, developed secondary structures, landing sites for specific proteins, etc.) [187]. Polysome profiling is an earlier and simpler version of ribosomal profiling. This approach makes it possible to fractionate polysomes according to the number of ribosomes in their composition. The disadvantage of this method is the lack of information about the ribosome-preferred sites mentioned. At the same time, high-resolution polysomal profiling, both in terms of separation of polysom fractions and with sufficient sequencing depth, provides detailed information on the dynamics of translation of specific transcripts. TRAP-seq [188,189] should be considered as an alternative to polysomic profiling, which, however, does not reduce its effectiveness and also allows analysis of changes in gene expression on the scale of the entire translatome [190]. RNC-seq makes it possible to obtain information about all translated transcripts but does not provide quantitative information about the dynamics of translation itself or about specific nucleotide contexts.
As can be expected, the combination of these approaches increases both the overall resolution of the experiment and the range of information obtained. However, the main information that these methods provide is the transcript frequencies in the translation involvement gradient [191].
3.2. Examination of mRNA secondary structure
Given the variety of methods for analysing the secondary structure of mRNA, here we only note the progressive full transcriptome approaches. A detailed review of all modern approaches is given in two review papers devoted to the study of plant structure [166] and RNA structure in general [192].
Most modern methods of structural analysis are based on the identification of nucleotides that make up single-stranded regions of target molecules. As a rule, the nucleotides in these regions are chemically modified or subjected to enzymatic degradation. However, it should be noted that protection against chemical modification or degradation by means of enzymes can be provided both by binding nucleotides in secondary structures (hairpins, etc.) and by shielding protein molecules bound to mRNA. Another factor that introduces uncertainty is the conditions of the experiment itself: in vivo vs. in vitro. It will be no secret that conditions in a living cell can be very different from those in a test tube. To eliminate all these sources of uncertainty, a number of experiments were developed that are not universal in themselves but, when used together, make it possible to understand the true state of affairs with the structure of mRNA in its natural environment. Modern methods for analysing the structure of RNA molecules are based on a number of general provisions:
- For analysis under in vivo conditions, chemical modification of bases should be preferred since large molecules of RNases and proteases may have significant problems crossing the plasmalemma and, in the case of plants, the cell wall.
-
Reagents used to modify unbound bases must not be toxic.
- −
- For example, dimethyl sulphate (DMS) [193], selectively methylating adenines and cytosines as part of single-stranded RNA regions; or N-cyclohexyl- N’-(2-morpholinoethyl)carbodiimide metho-p-toluenesulfonate (CMCT), which forms adducts with N1 and N3 of pseudouridine, N3 of uridine, and N1 of guanosine and inosine, as well as a combination of these two methods [194].
- −
- A separate method is icSHAPE [195], the essence of which is to modify free (single-stranded) RNA bases of all 4 types.
- −
- A similar approach is SHAPE-MaP [196]
- When analysing the temperature modulation of translation, one should take into account the increase in reactivity of modifying reagents and carry out the appropriate normalisation of the obtained data.
-
To eliminate the ambiguity arising from the protection of bases by RNA-bound proteins, it is necessary to apply approaches based on selective combinatorial degradation:
- −
- double-stranded RNA
- −
- single-stranded RNA
- −
- proteins that protect the backbone of the RNA molecule
The second group of methods for studying the structurome are approaches based on the formation of crosslinks or ligation [192]. These techniques are based on the chemical combination of macromolecules that are closely spaced or actively interacting. The main example for this group of methods for analysing both the structure of RNA and RNA regions shielded by protein molecules is PIP-seq [197]. In fact, it is a development of the previously known approach of RNase-mediated protein footprint sequencing [198]. In short, the sample is cross-linked (due to formaldehyde treatment). Then the sample is divided into several. Further, some of the samples undergo deproteinization, while others are immediately processed by either duplex-specific or single-stranded RNA-specific nucleases. Deproteinized samples are also hydrolyzed in the same way. A strand-specific library preparation and sequencing complete the analysis. Analysis of multiple fragments (after, of course, alignment) allows you to get an idea about the structure of RNA and the proteins associated with it. The obvious disadvantage of this approach is that most of the manipulations are carried out already in conditions in vitro, and not in vivo. But this difficulty is easily overcome by combining PIP-seq with Structure-seq or similar methods, as was done, for example, in [145].
3.3. In silico mRNA structure prediction
Speaking about approaches to studying the structure of mRNA, one cannot fail to mention the prediction of this structure by in silico methods. A distinction is made between similarity-based (or comparative) methods and folding calculation methods. Comparative approaches (as the name implies) are based on the search and comparison of conserved elements in mRNA sequences. With high accuracy, the application of this type of analysis is hampered by the lack of large databases and the high diversity of RNA molecules. Folding prediction, as a rule, involves various systems for calculating scoring parameters, which can be obtained empirically, set arbitrarily, or based on probabilistic models (in particular, using machine learning). However, the most modern approaches are based on the use of various machine learning methods and neural networks [199,200]).
4. Concluding remarks
We were unable to cover the entire process of translational regulation in plants under abiotic stress in a single review. For example, stop codon readthrough (SCR) has been shown in many organisms. This phenomenon consists in overcoming the canonical stop codon by the ribosome. In plants, this mechanism is not well understood. However, [201] demonstrated on the A.thaliana SCR model for 144 genes. Based on GO analysis, these genes can be divided into three main subgroups: translation, photosynthesis, and response to abiotic stress. The peptides themselves, synthesised at the C-terminus during this process, can be signals of localization in peroxisomes, the nucleus, or the membrane, as well as unstructured peptides without a clearly defined function. Another interesting regulatory mechanism, however, needs further experimental verification, for example, when analysing the stress response of plants. Another interesting study [202] demonstrated an increase in the length of the poly-A tail of some Arabidopsis genes (including heat shock protein genes) in response to heat stress. The mechanism, however, is not clear. The circadian rhythm also has a significant effect on the set of translated mRNAs [203]. The specific mechanism remains unclear, and it is not entirely clear how to distinguish between the contributions of transcription and translation in this particular case. Similar examples can be continued. The study of translation in plants is the study of a complex system. As can be seen from the above, more than a dozen mechanisms may be involved in the control of translation. These processes differ both in speed and stability. In our opinion, such diversity makes it possible, first, to fine-tune and quickly (from an evolutionary point of view) control the translation of specific genes. Second, a large set of alternative disjoint possibilities organises several independent translation channels, which in turn expands the range of metabolic pathways for which co-orchestration becomes available. However, multithreading creates additional difficulties in the analysis of data, especially those obtained as a result of *ome-wide experiments. In our opinion, the next step in the development of the study of translation in general (and in plants in particular) is the creation of modern, updated databases that allow the possibility of generalisation and prediction based on the data obtained. Since the analysis by the efforts of the researchers themselves is already very difficult at the present stage. In conclusion, we would like to note that the gaps in the model of translation in plants (including those under the influence of abiotic stress conditions, as an example of modifying factors) are rather an invitation to further work. If some mechanism is discovered in other taxa, by analogy, it is possible to show the presence of the same regulatory circuit for plants (since studies on this group of organisms are traditionally delayed). If there are difficulties with this approach, it will be no less intriguing to find out why this mechanism does not work in plants and the nature of these restrictions.
Author Contributions
All authors contributed equally to this work. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Russian Science Foundation, RSF grant number 22-74-10064; and the research was carried out within the state assignment of Ministry of Science and Higher Education of the Russian Federation (theme No. 122042700043-9).
Institutional Review Board Statement
Not applicable.
Acknowledgments
We apologize to authors whose work has not been cited here, because of the limitations associated with the review format.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Czékus, Z.; Csíkos, O.; Ördög, A.; Tari, I.; Poór, P. Effects of Jasmonic Acid in ER Stress and Unfolded Protein Response in Tomato Plants. Biomolecules 2020, 10, 1031. [Google Scholar] [CrossRef] [PubMed]
- Czékus, Z.; Szalai, G.; Tari, I.; Khan, M.I.R.; Poór, P. Role of ethylene in ER stress and the unfolded protein response in tomato (Solanum lycopersicum L.) plants. Plant Physiology and Biochemistry 2022, 181, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Je, S.; Lee, Y.; Yamaoka, Y. Effect of Common ER Stress–Inducing Drugs on the Growth and Lipid Phenotypes of Chlamydomonas and Arabidopsis. Plant and Cell Physiology 2023, 64, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.Y.; Kanehara, K. The Unfolded Protein Response Modulates a Phosphoinositide-Binding Protein through the IRE1-bZIP60 Pathway. Plant Physiology 2020, 183, 221–235. [Google Scholar] [CrossRef]
- Raxwal, V.K.; Ghosh, S.; Singh, S.; Katiyar-Agarwal, S.; Goel, S.; Jagannath, A.; Kumar, A.; Scaria, V.; Agarwal, M. Abiotic stress-mediated modulation of the chromatin landscape in Arabidopsis thaliana. Journal of Experimental Botany 2020, 71, 5280–5293. [Google Scholar] [CrossRef]
- Liang, Z.; Zhang, Q.; Ji, C.; Hu, G.; Zhang, P.; Wang, Y.; Yang, L.; Gu, X. Reorganization of the 3D chromatin architecture of rice genomes during heat stress. BMC Biology 2021, 19, 53. [Google Scholar] [CrossRef]
- Sun, L.; Jing, Y.; Liu, X.; Li, Q.; Xue, Z.; Cheng, Z.; Wang, D.; He, H.; Qian, W. Heat stress-induced transposon activation correlates with 3D chromatin organization rearrangement in Arabidopsis. Nature Communications 2020, 11, 1886. [Google Scholar] [CrossRef]
- Selma, S.; Orzáez, D. Perspectives for epigenetic editing in crops. Transgenic Research 2021, 30, 381–400. [Google Scholar] [CrossRef]
- Kumar, S.; Kaur, S.; Seem, K.; Kumar, S.; Mohapatra, T. Understanding 3D Genome Organization and Its Effect on Transcriptional Gene Regulation Under Environmental Stress in Plant: A Chromatin Perspective. Frontiers in Cell and Developmental Biology 2021, 9. [Google Scholar] [CrossRef]
- Nunez-Vazquez, R.; Desvoyes, B.; Gutierrez, C. Histone variants and modifications during abiotic stress response. Frontiers in Plant Science 2022, 13. [Google Scholar] [CrossRef]
- James, A.B.; Sullivan, S.; Nimmo, H.G. Global spatial analysis of Arabidopsis natural variants implicates 5’UTR splicing of LATE ELONGATED HYPOCOTYL in responses to temperature. Plant, Cell and Environment 2018, 41, 1524–1538. [Google Scholar] [CrossRef] [PubMed]
- Gorban, A.N.; Harel-Bellan, A.; Morozova, N.; Zinovyev, A. Basic, simple and extendable kinetic model of protein synthesis. Mathematical Biosciences and Engineering 2019, 16, 6602–6622. [Google Scholar] [CrossRef] [PubMed]
- Muench, D.G.; Zhang, C.; Dahodwala, M. Control of cytoplasmic translation in plants. Wiley Interdisciplinary Reviews: RNA 2012, 3, 178–194. [Google Scholar] [CrossRef] [PubMed]
- Lokdarshi, A.; Morgan, P.W.; Franks, M.; Emert, Z.; Emanuel, C.; von Arnim, A.G. Light-Dependent Activation of the GCN2 Kinase Under Cold and Salt Stress Is Mediated by the Photosynthetic Status of the Chloroplast. Frontiers in Plant Science 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Kørner, C.; Du, X.; Vollmer, M.; Pajerowska-Mukhtar, K. Endoplasmic Reticulum Stress Signaling in Plant Immunity—At the Crossroad of Life and Death. International Journal of Molecular Sciences 2015, 16, 26582–26598. [Google Scholar] [CrossRef]
- Liu, X.; Afrin, T.; Pajerowska-Mukhtar, K.M. Arabidopsis GCN2 kinase contributes to ABA homeostasis and stomatal immunity. Communications Biology 2019, 2, 302. [Google Scholar] [CrossRef]
- Lokdarshi, A.; von Arnim, A.G.; Akuoko, T.K. Modulation of GCN2 activity under excess light stress by osmoprotectants and amino acids. Plant Signaling and Behavior 2022, 17. [Google Scholar] [CrossRef]
- Son, S.; Im, J.H.; Song, G.; Park, S.R. SNF1-Related Protein Kinase 1 Activity Represses the Canonical Translational Machinery. Plants 2022, 11, 1359. [Google Scholar] [CrossRef]
- Patrick, R.M.; Browning, K.S. The eIF4F and eIFiso4F Complexes of Plants: An Evolutionary Perspective. Comparative and Functional Genomics 2012, 2012, 1–12. [Google Scholar] [CrossRef]
- Cho, H.; Lu, M.J.; Shih, M. The Sn RK 1- eIF iso4G1 signaling relay regulates the translation of specific mRNA s in Arabidopsis under submergence. New Phytologist 2019, 222, 366–381. [Google Scholar] [CrossRef]
- Gallie, D.R.; Le, H.; Caldwell, C.; Tanguay, R.L.; Hoang, N.X.; Browning, K.S. The Phosphorylation State of Translation Initiation Factors Is Regulated Developmentally and following Heat Shock in Wheat. Journal of Biological Chemistry 1997, 272, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- Tuteja, N.; Vashisht, A.A.; Tuteja, R. Translation initiation factor 4A: a prototype member of dead-box protein family. Physiology and Molecular Biology of Plants 2008, 14, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Toribio, R.; Muñoz, A.; Castro-Sanz, A.B.; Merchante, C.; Castellano, M.M. A novel eIF4E-interacting protein that forms non-canonical translation initiation complexes. Nature Plants 2019, 5, 1283–1296. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.L.; Hong, S.K. Sensitivity of Translation Initiation Factor eIF1 as a Molecular Target of Salt Toxicity to Sodic-Alkaline Stress in the Halophytic Grass Leymus chinensis. Biochemical Genetics 2013, 51, 101–118. [Google Scholar] [CrossRef]
- Yang, G.; Yu, L.; Wang, Y.; Wang, C.; Gao, C. The Translation Initiation Factor 1A (TheIF1A) from Tamarix hispida Is Regulated by a Dof Transcription Factor and Increased Abiotic Stress Tolerance. Frontiers in Plant Science 2017, 8. [Google Scholar] [CrossRef]
- Hashem, Y.; des Georges, A.; Dhote, V.; Langlois, R.; Liao, H.Y.; Grassucci, R.A.; Hellen, C.U.; Pestova, T.V.; Frank, J. Structure of the Mammalian Ribosomal 43S Preinitiation Complex Bound to the Scanning Factor DHX29. Cell 2013, 153, 1108–1119. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, X.; Gaikwad, K.; Kou, X.; Wang, F.; Tian, X.; Xin, M.; Ni, Z.; Sun, Q.; Peng, H.; Vierling, E. Mutations in eIF5B Confer Thermosensitive and Pleiotropic Phenotypes via Translation Defects in Arabidopsis thaliana. The Plant Cell 2017, 29, 1952–1969. [Google Scholar] [CrossRef]
- Lorsch, J.R.; Dever, T.E. Molecular View of 43 S Complex Formation and Start Site Selection in Eukaryotic Translation Initiation. Journal of Biological Chemistry 2010, 285, 21203–21207. [Google Scholar] [CrossRef]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought — from genes to the whole plant. Functional Plant Biology 2003, 30, 239. [Google Scholar] [CrossRef]
- Singh, G.; Jain, M.; Kulshreshtha, R.; Khurana, J.P.; Kumar, S.; Singh, P. Expression analysis of genes encoding translation initiation factor 3 subunit g (TaeIF3g) and vesicle-associated membrane protein-associated protein (TaVAP) in drought tolerant and susceptible cultivars of wheat. Plant Science 2007, 173, 660–669. [Google Scholar] [CrossRef]
- Putnam, A.A.; Jankowsky, E. DEAD-box helicases as integrators of RNA, nucleotide and protein binding. Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms 2013, 1829, 884–893. [Google Scholar] [CrossRef]
- Tyagi, V.; Parihar, V.; Malik, G.; Kalra, V.; Kapoor, S.; Kapoor, M. The DEAD-box RNA helicase eIF4A regulates plant development and interacts with the hnRNP LIF2L1 in Physcomitrella patens. Molecular Genetics and Genomics 2020, 295, 373–389. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Mudgil, Y.; Pandey, S.; Fartyal, D.; Reddy, M.K. Structural modelling and phylogenetic analyses of PgeIF4A2 (Eukaryotic translation initiation factor) from Pennisetum glaucum reveal signature motifs with a role in stress tolerance and development. Bioinformation 2016, 12, 416–419. [Google Scholar] [CrossRef] [PubMed]
- Boex-Fontvieille, E.; Daventure, M.; Jossier, M.; Zivy, M.; Hodges, M.; Tcherkez, G. Photosynthetic Control of Arabidopsis Leaf Cytoplasmic Translation Initiation by Protein Phosphorylation. PLoS ONE 2013, 8, e70692. [Google Scholar] [CrossRef] [PubMed]
- Guihur, A.; Rebeaud, M.E.; Goloubinoff, P. How do plants feel the heat and survive? Trends in Biochemical Sciences 2022, 47, 824–838. [Google Scholar] [CrossRef]
- Merret, R.; Carpentier, M.C.; Favory, J.J.; Picart, C.; Descombin, J.; Bousquet-Antonelli, C.; Tillard, P.; Lejay, L.; Deragon, J.M.; yung Charng, Y. Heat Shock Protein HSP101 Affects the Release of Ribosomal Protein mRNAs for Recovery after Heat Shock. Plant Physiology 2017, 174, 1216–1225. [Google Scholar] [CrossRef]
- McLoughlin, F.; Basha, E.; Fowler, M.E.; Kim, M.; Bordowitz, J.; Katiyar-Agarwal, S.; Vierling, E. Class I and II small heat-shock proteins protect protein translation factors during heat stress. Plant Physiology, 2016. [Google Scholar] [CrossRef]
- Walters, R.W.; Parker, R. Coupling of Ribostasis and Proteostasis: Hsp70 Proteins in mRNA Metabolism. Trends in Biochemical Sciences 2015, 40, 552–559. [Google Scholar] [CrossRef]
- Muthuramalingam, M.; Wang, Y.; Li, Y.; Mahalingam, R. Interacting protein partners of Arabidopsis RNA-binding protein AtRBP45b. Plant Biology 2017, 19, 327–334. [Google Scholar] [CrossRef]
- Qi, Y.; Tsuda, K.; Joe, A.; Sato, M.; Nguyen, L.V.; Glazebrook, J.; Alfano, J.R.; Cohen, J.D.; Katagiri, F. A Putative RNA-Binding Protein Positively Regulates Salicylic Acid–Mediated Immunity in Arabidopsis. Molecular Plant-Microbe Interactions® 2010, 23, 1573–1583. [Google Scholar] [CrossRef]
- Akbudak, M.A.; Yildiz, S.; Filiz, E. Pathogenesis related protein-1 (PR-1) genes in tomato (Solanum lycopersicum L.): Bioinformatics analyses and expression profiles in response to drought stress. Genomics 2020, 112, 4089–4099. [Google Scholar] [CrossRef]
- Chien, P.S.; Nam, H.G.; Chen, Y.R. A salt-regulated peptide derived from the CAP superfamily protein negatively regulates salt-stress tolerance in Arabidopsis. Journal of Experimental Botany 2015, 66, 5301–5313. [Google Scholar] [CrossRef] [PubMed]
- Duque, P. A role for SR proteins in plant stress responses. Plant Signaling and Behavior 2011, 6, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Laloum, T.; Martín, G.; Duque, P. Alternative Splicing Control of Abiotic Stress Responses. Trends in Plant Science 2018, 23, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Du, B.; Liu, D.; Qi, X. Splicing factor SR34b mutation reduces cadmium tolerance in Arabidopsis by regulating iron-regulated transporter 1 gene. Biochemical and Biophysical Research Communications 2014, 455, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Cui, P.; Chen, H.; Ali, S.; Zhang, S.; Xiong, L. A KH-Domain RNA-Binding Protein Interacts with FIERY2/CTD Phosphatase-Like 1 and Splicing Factors and Is Important for Pre-mRNA Splicing in Arabidopsis. PLoS Genetics 2013, 9, e1003875. [Google Scholar] [CrossRef]
- Wang, D.; Luthe, D.S. Heat Sensitivity in a Bentgrass Variant. Failure to Accumulate a Chloroplast Heat Shock Protein Isoform Implicated in Heat Tolerance. Plant Physiology 2003, 133, 319–327. [Google Scholar] [CrossRef]
- Sun, W.; Montagu, M.V.; Verbruggen, N. Small heat shock proteins and stress tolerance in plants. Biochimica et Biophysica Acta (BBA) - Gene Structure and Expression 2002, 1577, 1–9. [Google Scholar] [CrossRef]
- Gao, Z.; Nie, J.; Wang, H. MicroRNA biogenesis in plant. Plant Growth Regulation 2021, 93, 1–12. [Google Scholar] [CrossRef]
- oki Iwakawa, H.; Tomari, Y. Molecular Insights into microRNA-Mediated Translational Repression in Plants. Molecular Cell 2013, 52, 591–601. [Google Scholar] [CrossRef]
- Sunkar, R.; Zhu, J.K. Novel and Stress-Regulated MicroRNAs and Other Small RNAs from Arabidopsis[W]. The Plant Cell 2004, 16, 2001–2019. [Google Scholar] [CrossRef]
- Lee, M.S. The role of the activated macrophage in clearing Listeria monocytogenes infection. Frontiers in Bioscience 2007, 12, 2683. [Google Scholar] [CrossRef]
- Zhao, B.; Ge, L.; Liang, R.; Li, W.; Ruan, K.; Lin, H.; Jin, Y. Members of miR-169 family are induced by high salinity and transiently inhibit the NF-YA transcription factor. BMC Molecular Biology 2009, 10, 29. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.C.; Liu, P.P.; Goloviznina, N.A.; Nonogaki, H. microRNA, seeds, and Darwin?: diverse function of miRNA in seed biology and plant responses to stress. Journal of Experimental Botany 2010, 61, 2229–2234. [Google Scholar] [CrossRef] [PubMed]
- Yuan, N.; Yuan, S.; Li, Z.; Li, D.; Hu, Q.; Luo, H. Heterologous expression of a rice miR395 gene in Nicotiana tabacum impairs sulfate homeostasis. Scientific Reports 2016, 6, 28791. [Google Scholar] [CrossRef] [PubMed]
- Matthewman, C.A.; Kawashima, C.G.; Húska, D.; Csorba, T.; Dalmay, T.; Kopriva, S. miR395 is a general component of the sulfate assimilation regulatory network in Arabidopsis. FEBS Letters 2012, 586, 3242–3248. [Google Scholar] [CrossRef] [PubMed]
- Chiou, T.J.; Aung, K.; Lin, S.I.; Wu, C.C.; Chiang, S.F.; lin Su, C. Regulation of Phosphate Homeostasis by MicroRNA in Arabidopsis. The Plant Cell 2006, 18, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Bazin, J.; Baerenfaller, K.; Gosai, S.J.; Gregory, B.D.; Crespi, M.; Bailey-Serres, J. Global analysis of ribosome-associated noncoding RNAs unveils new modes of translational regulation. Proceedings of the National Academy of Sciences 2017, 114. [Google Scholar] [CrossRef]
- Kumar, R. Role of MicroRNAs in Biotic and Abiotic Stress Responses in Crop Plants. Applied Biochemistry and Biotechnology 2014, 174, 93–115. [Google Scholar] [CrossRef]
- Vidal, E.A.; Araus, V.; Lu, C.; Parry, G.; Green, P.J.; Coruzzi, G.M.; Gutiérrez, R.A. Nitrate-responsive miR393/ AFB3 regulatory module controls root system architecture in Arabidopsis thaliana. Proceedings of the National Academy of Sciences 2010, 107, 4477–4482. [Google Scholar] [CrossRef]
- Long, R.; Li, M.; Li, X.; Gao, Y.; Zhang, T.; Sun, Y.; Kang, J.; Wang, T.; Cong, L.; Yang, Q. A Novel miRNA Sponge Form Efficiently Inhibits the Activity of miR393 and Enhances the Salt Tolerance and ABA Insensitivity in Arabidopsis thaliana. Plant Molecular Biology Reporter 2017, 35, 409–415. [Google Scholar] [CrossRef]
- Liu, X.; Huang, S.; Xie, H. Advances in the regulation of plant development and stress response by miR167. Frontiers in Bioscience-Landmark 2021, 26, 655. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Lu, Y.; Zinta, G.; Lang, Z.; Zhu, J.K. UTR-Dependent Control of Gene Expression in Plants. Trends in Plant Science 2018, 23, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, H.; Shinmyo, A.; Kato, K. Preferential translation mediated by Hsp81-3 5’-UTR during heat shock involves ribosome entry at the 5’-end rather than an internal site in Arabidopsis suspension cells. Journal of Bioscience and Bioengineering 2008, 105, 39–47. [Google Scholar] [CrossRef]
- Meyer, K.D.; Patil, D.P.; Zhou, J.; Zinoviev, A.; Skabkin, M.A.; Elemento, O.; Pestova, T.V.; Qian, S.B.; Jaffrey, S.R. 5’ UTR m6A Promotes Cap-Independent Translation. Cell 2015, 163, 999–1010. [Google Scholar] [CrossRef]
- Álvarez, D.; Voß, B.; Maass, D.; Wüst, F.; Schaub, P.; Beyer, P.; Welsch, R. Carotenogenesis Is Regulated by 5’UTR-Mediated Translation of Phytoene Synthase Splice Variants. Plant Physiology 2016, 172, 2314–2326. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, H.; Takenami, S.; Kubo, Y.; Ueda, K.; Ueda, A.; Yamaguchi, M.; Hirata, K.; Demura, T.; Kanaya, S.; Kato, K. A Computational and Experimental Approach Reveals that the 5’-Proximal Region of the 5’-UTR has a Cis-Regulatory Signature Responsible for Heat Stress-Regulated mRNA Translation in Arabidopsis. Plant and Cell Physiology 2013, 54, 474–483. [Google Scholar] [CrossRef]
- Yamasaki, S.; Sanada, Y.; Imase, R.; Matsuura, H.; Ueno, D.; Demura, T.; Kato, K. Arabidopsis thaliana cold-regulated 47 gene 5’-untranslated region enables stable high-level expression of transgenes. Journal of Bioscience and Bioengineering 2018, 125, 124–130. [Google Scholar] [CrossRef]
- Zhao, D.; Hamilton, J.P.; Hardigan, M.; Yin, D.; He, T.; Vaillancourt, B.; Reynoso, M.; Pauluzzi, G.; Funkhouser, S.; Cui, Y.; Bailey-Serres, J.; Jiang, J.; Buell, C.R.; Jiang, N. Analysis of Ribosome-Associated mRNAs in Rice Reveals the Importance of Transcript Size and GC Content in Translation. Genes|Genomes|Genetics 2017, 7, 203–219. [Google Scholar] [CrossRef]
- Xu, J.; Du, R.; Meng, X.; Zhao, W.; Kong, L.; Chen, J. Third-Generation Sequencing Indicated that LncRNA Could Regulate eIF2D to Enhance Protein Translation Under Heat Stress in Populus simonii. Plant Molecular Biology Reporter 2021, 39, 240–250. [Google Scholar] [CrossRef]
- Li, S.; Cheng, Z.; Dong, S.; Li, Z.; Zou, L.; Zhao, P.; Guo, X.; Bao, Y.; Wang, W.; Peng, M. Global identification of full-length cassava lncRNAs unveils the role of cold-responsive intergenic lncRNA 1 in cold stress response. Plant, Cell and Environment 2022, 45, 412–426. [Google Scholar] [CrossRef]
- Dever, T.E. Translation initiation: adept at adapting. Trends in Biochemical Sciences 1999, 24, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Terenin, I.M.; Dmitriev, S.E.; Andreev, D.E.; Royall, E.; Belsham, G.J.; Roberts, L.O.; Shatsky, I.N. A Cross-Kingdom Internal Ribosome Entry Site Reveals a Simplified Mode of Internal Ribosome Entry. Molecular and Cellular Biology 2005, 25, 7879–7888. [Google Scholar] [CrossRef] [PubMed]
- Holcik, M.; Sonenberg, N.; Korneluk, R.G. Internal ribosome initiation of translation and the control of cell death. Trends in Genetics 2000, 16, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, S.; Bruynooghe, Y.; Denecker, G.; Huffel, S.V.; Tinton, S.; Beyaert, R. Identification and Characterization of a Novel Cell Cycle–Regulated Internal Ribosome Entry Site. Molecular Cell 2000, 5, 597–605. [Google Scholar] [CrossRef]
- Pyronnet, S.; Pradayrol, L.; Sonenberg, N. A Cell Cycle–Dependent Internal Ribosome Entry Site. Molecular Cell 2000, 5, 607–616. [Google Scholar] [CrossRef]
- Merrick, W.C. Cap-dependent and cap-independent translation in eukaryotic systems. Gene 2004, 332, 1–11. [Google Scholar] [CrossRef]
- Gingras, A.C.; Raught, B.; Sonenberg, N. eIF4 Initiation Factors: Effectors of mRNA Recruitment to Ribosomes and Regulators of Translation. Annual Review of Biochemistry 1999, 68, 913–963. [Google Scholar] [CrossRef]
- Rubtsova, M.P.; Sizova, D.V.; Dmitriev, S.E.; Ivanov, D.S.; Prassolov, V.S.; Shatsky, I.N. Distinctive Properties of the 5’-Untranslated Region of Human Hsp70 mRNA. Journal of Biological Chemistry 2003, 278, 22350–22356. [Google Scholar] [CrossRef]
- Dinkova, T.D.; Zepeda, H.; Martínez-Salas, E.; Martínez, L.M.; Nieto-Sotelo, J.; Jiménez, E.S. Cap-independent translation of maize Hsp101. The Plant Journal 2005, 41, 722–731. [Google Scholar] [CrossRef]
- Gouiaa, S.; Khoudi, H.; Leidi, E.O.; Pardo, J.M.; Masmoudi, K. Expression of wheat Na+/H+ antiporter TNHXS1 and H+- pyrophosphatase TVP1 genes in tobacco from a bicistronic transcriptional unit improves salt tolerance. Plant Molecular Biology 2012, 79, 137–155. [Google Scholar] [CrossRef]
- Dorokhov, Y.L.; Skulachev, M.V.; Ivanov, P.A.; Zvereva, S.D.; Tjulkina, L.G.; Merits, A.; Gleba, Y.Y.; Hohn, T.; Atabekov, J.G. Polypurine (A)-rich sequences promote cross-kingdom conservation of internal ribosome entry. Proceedings of the National Academy of Sciences 2002, 99, 5301–5306. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, H.; Peer, W.A.; Richter, G.; Blakeslee, J.; Bandyopadhyay, A.; Titapiwantakun, B.; Undurraga, S.; Khodakovskaya, M.; Richards, E.L.; Krizek, B.; Murphy, A.S.; Gilroy, S.; Gaxiola, R. Arabidopsis H+ -PPase AVP1 Regulates Auxin-Mediated Organ Development. Science 2005, 310, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Habib, K.; Aida, N.K.; S. ; ra, G.; Khaled, M. Optimization of regeneration and transformation parameters in tomato and improvement of its salinity and drought tolerance. African Journal of Biotechnology 2009, 8, 6068–6076. [Google Scholar] [CrossRef]
- Asad, S.; Mukhtar, Z.; Nazir, F.; Hashmi, J.A.; Mansoor, S.; Zafar, Y.; Arshad, M. Silicon Carbide Whisker-Mediated Embryogenic Callus Transformation of Cotton (Gossypium hirsutum L.) and Regeneration of Salt Tolerant Plants. Molecular Biotechnology 2008, 40, 161–169. [Google Scholar] [CrossRef]
- LIU, H.; WANG, Q.; YU, M.; ZHANG, Y.; WU, Y.; ZHANG, H. Transgenic salt-tolerant sugar beet ( Beta vulgaris L.) constitutively expressing an Arabidopsis thaliana vacuolar Na+ / H+ antiporter gene, AtNHX3 , accumulates more soluble sugar but less salt in storage roots. Plant, Cell and Environment 2008, 31, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- LI, Z.; BALDWIN, C.M.; HU, Q.; LIU, H.; LUO, H. Heterologous expression of Arabidopsis H+ -pyrophosphatase enhances salt tolerance in transgenic creeping bentgrass ( Agrostis stolonifera L.). Plant, Cell and Environment 2010, 33, 272–289. [Google Scholar] [CrossRef]
- Cui, Y.; Rao, S.; Chang, B.; Wang, X.; Zhang, K.; Hou, X.; Zhu, X.; Wu, H.; Tian, Z.; Zhao, Z.; Yang, C.; Huang, T. AtLa1 protein initiates IRES-dependent translation of WUSCHEL mRNA and regulates the stem cell homeostasis of A rabidopsis in response to environmental hazards. Plant, Cell and Environment 2015, 38, 2098–2114. [Google Scholar] [CrossRef]
- Gouiaa, S.; Khoudi, H. Co-expression of vacuolar Na+/ H+ antiporter and H+-pyrophosphatase with an IRES-mediated dicistronic vector improves salinity tolerance and enhances potassium biofortification of tomato. Phytochemistry 2015, 117, 537–546. [Google Scholar] [CrossRef]
- Barragán, V.; Leidi, E.O.; Andrés, Z.; Rubio, L.; Luca, A.D.; Fernández, J.A.; Cubero, B.; Pardo, J.M. Ion Exchangers NHX1 and NHX2 Mediate Active Potassium Uptake into Vacuoles to Regulate Cell Turgor and Stomatal Function in Arabidopsis. The Plant Cell 2012, 24, 1127–1142. [Google Scholar] [CrossRef]
- Moll, I.; Grill, S.; Gualerzi, C.O.; Blasi, U. Leaderless mRNAs in bacteria: surprises in ribosomal recruitment and translational control. Molecular Microbiology 2002, 43, 239–246. [Google Scholar] [CrossRef]
- Moll, I. Translation initiation with 70S ribosomes: an alternative pathway for leaderless mRNAs. Nucleic Acids Research 2004, 32, 3354–3363. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Matsunaga, N.; Akabane, S.; Yasuda, I.; Ueda, T.; Takeuchi-Tomita, N. Reconstitution of mammalian mitochondrial translation system capable of correct initiation and long polypeptide synthesis from leaderless mRNA. Nucleic Acids Research 2021, 49, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Remes, C.; Khawaja, A.; Pearce, S.F.; Dinan, A.M.; Gopalakrishna, S.; Cipullo, M.; Kyriakidis, V.; Zhang, J.; Dopico, X.C.; Yukhnovets, O.; Atanassov, I.; Firth, A.E.; Cooperman, B.; Rorbach, J. Translation initiation of leaderless and polycistronic transcripts in mammalian mitochondria. Nucleic Acids Research 2023, 51, 891–907. [Google Scholar] [CrossRef] [PubMed]
- Andreev, D.E.; Terenin, I.M.; Dunaevsky, Y.E.; Dmitriev, S.E.; Shatsky, I.N. A Leaderless mRNA Can Bind to Mammalian 80S Ribosomes and Direct Polypeptide Synthesis in the Absence of Translation Initiation Factors. Molecular and Cellular Biology 2006, 26, 3164–3169. [Google Scholar] [CrossRef]
- Akulich, K.A.; Andreev, D.E.; Terenin, I.M.; Smirnova, V.V.; Anisimova, A.S.; Makeeva, D.S.; Arkhipova, V.I.; Stolboushkina, E.A.; Garber, M.B.; Prokofjeva, M.M.; Spirin, P.V.; Prassolov, V.S.; Shatsky, I.N.; Dmitriev, S.E. Four translation initiation pathways employed by the leaderless mRNA in eukaryotes. Scientific Reports 2016, 6, 37905. [Google Scholar] [CrossRef]
- Cahoon, A.B.; Qureshi, A.A. Leaderless mRNAs are circularized in Chlamydomonas reinhardtii mitochondria. Current Genetics 2018, 64, 1321–1333. [Google Scholar] [CrossRef]
- Barbosa, C.; Peixeiro, I.; Romão, L. Gene Expression Regulation by Upstream Open Reading Frames and Human Disease. PLoS Genetics 2013, 9, e1003529. [Google Scholar] [CrossRef]
- Johnstone, T.G.; Bazzini, A.A.; Giraldez, A.J. Upstream ORF s are prevalent translational repressors in vertebrates. The EMBO Journal 2016, 35, 706–723. [Google Scholar] [CrossRef]
- Kurihara, Y. uORF Shuffling Fine-Tunes Gene Expression at a Deep Level of the Process. Plants 2020, 9, 608. [Google Scholar] [CrossRef]
- von Arnim, A.G.; Jia, Q.; Vaughn, J.N. Regulation of plant translation by upstream open reading frames. Plant Science 2014, 214, 1–12. [Google Scholar] [CrossRef]
- Shashikanth, M.; Krishna, A.R.; Ramya, G.; Devi, G.; Ulaganathan, K. Genome-wide comparative analysis of Oryza sativa (japonica) and Arabidopsis thaliana 5’-UTR sequences for translational regulatory signals. Plant Biotechnology 2008, 25, 553–563. [Google Scholar] [CrossRef]
- de Arce, A.J.D.; Noderer, W.L.; Wang, C.L. Complete motif analysis of sequence requirements for translation initiation at non-AUG start codons. Nucleic Acids Research 2018, 46, 985–994. [Google Scholar] [CrossRef]
- Kozak, M. The scanning model for translation: an update. Journal of Cell Biology 1989, 108, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.J.; Hellen, C.U.T.; Pestova, T.V. The mechanism of eukaryotic translation initiation and principles of its regulation. Nature Reviews Molecular Cell Biology 2010, 11, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Young, S.K.; Wek, R.C. Upstream Open Reading Frames Differentially Regulate Gene-specific Translation in the Integrated Stress Response. Journal of Biological Chemistry 2016, 291, 16927–16935. [Google Scholar] [CrossRef]
- Lloyd, J.P.B. The evolution and diversity of the nonsense-mediated mRNA decay pathway. F1000Research 2018, 7, 1299. [Google Scholar] [CrossRef]
- Yamashita, A. Role of SMG-1-mediated Upf1 phosphorylation in mammalian nonsense-mediated mRNA decay. Genes to Cells 2013, 18, 161–175. [Google Scholar] [CrossRef]
- Ivanov, I.P.; Loughran, G.; Atkins, J.F. uORFs with unusual translational start codons autoregulate expression of eukaryotic ornithine decarboxylase homologs. Proceedings of the National Academy of Sciences 2008, 105, 10079–10084. [Google Scholar] [CrossRef]
- van der Horst, S.; Filipovska, T.; Hanson, J.; Smeekens, S. Metabolite Control of Translation by Conserved Peptide uORFs: The Ribosome as a Metabolite Multisensor. Plant Physiology 2020, 182, 110–122. [Google Scholar] [CrossRef]
- Wiese, A.; Elzinga, N.; Wobbes, B.; Smeekens, S. A Conserved Upstream Open Reading Frame Mediates Sucrose-Induced Repression of Translation[W]. The Plant Cell 2004, 16, 1717–1729. [Google Scholar] [CrossRef]
- Alves, M.; Dadalto, S.; Gonçalves, A.; Souza, G.D.; Barros, V.; Fietto, L. Plant bZIP Transcription Factors Responsive to Pathogens: A Review. International Journal of Molecular Sciences 2013, 14, 7815–7828. [Google Scholar] [CrossRef] [PubMed]
- Sornaraj, P.; Luang, S.; Lopato, S.; Hrmova, M. Basic leucine zipper (bZIP) transcription factors involved in abiotic stresses: A molecular model of a wheat bZIP factor and implications of its structure in function. Biochimica et Biophysica Acta (BBA) - General Subjects 2016, 1860, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Noman, A.; Liu, Z.; Aqeel, M.; Zainab, M.; Khan, M.I.; Hussain, A.; Ashraf, M.F.; Li, X.; Weng, Y.; He, S. Basic leucine zipper domain transcription factors: the vanguards in plant immunity. Biotechnology Letters 2017, 39, 1779–1791. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; Sato, K.; Berberich, T.; Miyazaki, A.; Ozaki, R.; Imai, R.; Kusano, T. LIP19, a Basic Region Leucine Zipper Protein, is a Fos-like Molecular Switch in the Cold Signaling of Rice Plants. Plant and Cell Physiology 2005, 46, 1623–1634. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, F.; Maeta, E.; Terashima, A.; Kawaura, K.; Ogihara, Y.; Takumi, S. Development of abiotic stress tolerance via bZIP-type transcription factor LIP19 in common wheat. Journal of Experimental Botany 2008, 59, 891–905. [Google Scholar] [CrossRef]
- Ditzer, A.; Bartels, D. Identification of a dehydration and ABA-responsive promoter regulon and isolation of corresponding DNA binding proteins for the group 4 LEA gene CpC2 from C. plantagineum. Plant Molecular Biology 2006, 61, 643–663. [Google Scholar] [CrossRef]
- Shekhawat, U.K.S.; Ganapathi, T.R. Transgenic banana plants overexpressing MusabZIP53 display severe growth retardation with enhanced sucrose and polyphenol oxidase activity. Plant Cell, Tissue and Organ Culture (PCTOC) 2014, 116, 387–402. [Google Scholar] [CrossRef]
- Hartmann, L.; Pedrotti, L.; Weiste, C.; Fekete, A.; Schierstaedt, J.; Göttler, J.; Kempa, S.; Krischke, M.; Dietrich, K.; Mueller, M.J.; Vicente-Carbajosa, J.; Hanson, J.; Dröge-Laser, W. Crosstalk between Two bZIP Signaling Pathways Orchestrates Salt-Induced Metabolic Reprogramming in Arabidopsis Roots. The Plant Cell 2015, 27, 2244–2260. [Google Scholar] [CrossRef]
- Liu, Y.; Liang, H.; Lv, X.; Liu, D.; Wen, X.; Liao, Y. Effect of polyamines on the grain filling of wheat under drought stress. Plant Physiology and Biochemistry 2016, 100, 113–129. [Google Scholar] [CrossRef]
- Krishnan, S.; Merewitz, E.B. Polyamine Application Effects on Gibberellic Acid Content in Creeping Bentgrass during Drought Stress. Journal of the American Society for Horticultural Science 2017, 142, 135–142. [Google Scholar] [CrossRef]
- Li, S.; Jin, H.; Zhang, Q. The Effect of Exogenous Spermidine Concentration on Polyamine Metabolism and Salt Tolerance in Zoysiagrass (Zoysia japonica Steud) Subjected to Short-Term Salinity Stress. Frontiers in Plant Science 2016, 7. [Google Scholar] [CrossRef]
- Peynevandi, K.M.; Razavi, S.M.; Zahri, S. The ameliorating effects of polyamine supplement on physiological and biochemical parameters of Stevia rebaudiana Bertoni under cold stress. Plant Production Science 2018, 21, 123–131. [Google Scholar] [CrossRef]
- Dever, T.E.; Ivanov, I.P. Roles of polyamines in translation. Journal of Biological Chemistry 2018, 293, 18719–18729. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, I.P.; Atkins, J.F.; Michael, A.J. A profusion of upstream open reading frame mechanisms in polyamine-responsive translational regulation. Nucleic Acids Research 2010, 38, 353–359. [Google Scholar] [CrossRef]
- Hanfrey, C.; Elliott, K.A.; Franceschetti, M.; Mayer, M.J.; Illingworth, C.; Michael, A.J. A Dual Upstream Open Reading Frame-based Autoregulatory Circuit Controlling Polyamine-responsive Translation. Journal of Biological Chemistry 2005, 280, 39229–39237. [Google Scholar] [CrossRef]
- Imai, A.; Akiyama, T.; Kato, T.; Sato, S.; Tabata, S.; Yamamoto, K.T.; Takahashi, T. Spermine is not essential for survival of Arabidopsis. FEBS Letters 2004, 556, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Imai, A.; Hanzawa, Y.; Komura, M.; Yamamoto, K.T.; Komeda, Y.; Takahashi, T. The dwarf phenotype of the Arabidopsis acl5 mutant is suppressed by a mutation in an upstream ORF of a bHLH gene. Development 2006, 133, 3575–3585. [Google Scholar] [CrossRef]
- Hayden, C.A.; Jorgensen, R.A. Identification of novel conserved peptide uORF homology groups in Arabidopsis and rice reveals ancient eukaryotic origin of select groups and preferential association with transcription factor-encoding genes. BMC Biology 2007, 5, 32. [Google Scholar] [CrossRef]
- Ishitsuka, S.; Yamamoto, M.; Miyamoto, M.; Kuwashiro, Y.; Imai, A.; Motose, H.; Takahashi, T. Complexity and Conservation of Thermospermine-Responsive uORFs of SAC51 Family Genes in Angiosperms. Frontiers in Plant Science 2019, 10. [Google Scholar] [CrossRef]
- You, J.; Chan, Z. ROS Regulation During Abiotic Stress Responses in Crop Plants. Frontiers in Plant Science 2015, 6. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. The Plant Journal 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic Acid-A Potential Oxidant Scavenger and Its Role in Plant Development and Abiotic Stress Tolerance. Frontiers in Plant Science 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Broad, R.C.; Bonneau, J.P.; Beasley, J.T.; Roden, S.; Sadowski, P.; Jewell, N.; Brien, C.; Berger, B.; Tako, E.; Glahn, R.P.; Hellens, R.P.; Johnson, A.A.T. Effect of Rice GDP-L-Galactose Phosphorylase Constitutive Overexpression on Ascorbate Concentration, Stress Tolerance, and Iron Bioavailability in Rice. Frontiers in Plant Science 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Laing, W.A.; Martínez-Sánchez, M.; Wright, M.A.; Bulley, S.M.; Brewster, D.; Dare, A.P.; Rassam, M.; Wang, D.; Storey, R.; Macknight, R.C.; Hellens, R.P. An Upstream Open Reading Frame Is Essential for Feedback Regulation of Ascorbate Biosynthesis in Arabidopsis. The Plant Cell 2015, 27, 772–786. [Google Scholar] [CrossRef]
- Zhang, H.; Si, X.; Ji, X.; Fan, R.; Liu, J.; Chen, K.; Wang, D.; Gao, C. Genome editing of upstream open reading frames enables translational control in plants. Nature Biotechnology 2018, 36, 894–898. [Google Scholar] [CrossRef]
- Serganov, A.; Nudler, E. A Decade of Riboswitches. Cell 2013, 152, 17–24. [Google Scholar] [CrossRef]
- Bocobza, S.; Adato, A.; Mandel, T.; Shapira, M.; Nudler, E.; Aharoni, A. Riboswitch-dependent gene regulation and its evolution in the plant kingdom. Genes and Development 2007, 21, 2874–2879. [Google Scholar] [CrossRef]
- Klinkert, B.; Narberhaus, F. Microbial thermosensors. Cellular and Molecular Life Sciences 2009, 66, 2661–2676. [Google Scholar] [CrossRef]
- Kortmann, J.; Narberhaus, F. Bacterial RNA thermometers: molecular zippers and switches. Nature Reviews Microbiology 2012, 10, 255–265. [Google Scholar] [CrossRef]
- Samtani, H.; Unni, G.; Khurana, P. Microbial Mechanisms of Heat Sensing. Indian Journal of Microbiology 2022, 62, 175–186. [Google Scholar] [CrossRef]
- Somero, G.N. RNA thermosensors: how might animals exploit their regulatory potential? Journal of Experimental Biology 2018, 221. [Google Scholar] [CrossRef]
- Chung, B.Y.W.; Balcerowicz, M.; Antonio, M.D.; Jaeger, K.E.; Geng, F.; Franaszek, K.; Marriott, P.; Brierley, I.; Firth, A.E.; Wigge, P.A. An RNA thermoswitch regulates daytime growth in Arabidopsis. Nature Plants 2020, 6, 522–532. [Google Scholar] [CrossRef]
- Choi, D.; Oh, H.J.; Goh, C.J.; Lee, K.; Hahn, Y. Heat Shock RNA 1, Known as a Eukaryotic Temperature-Sensing Noncoding RNA, Is of Bacterial Origin. Journal of Microbiology and Biotechnology 2015, 25, 1234–1240. [Google Scholar] [CrossRef]
- Su, Z.; Tang, Y.; Ritchey, L.E.; Tack, D.C.; Zhu, M.; Bevilacqua, P.C.; Assmann, S.M. Genome-wide RNA structurome reprogramming by acute heat shock globally regulates mRNA abundance. Proceedings of the National Academy of Sciences 2018, 115, 12170–12175. [Google Scholar] [CrossRef]
- Thomas, S.E.; Balcerowicz, M.; Chung, B.Y.W. RNA structure mediated thermoregulation: What can we learn from plants? Frontiers in Plant Science 2022, 13. [Google Scholar] [CrossRef]
- Guillaume-Schöpfer, D.; Jaeger, K.E.; Geng, F.; Doccula, F.G.; Costa, A.; Webb, A.A.R.; Wigge, P.A. Ribosomes act as cryosensors in plants. bioRxiv, 2020. [Google Scholar] [CrossRef]
- Stirpe, F.; Barbieri, L.; Gorini, P.; Valbonesi, P.; Bolognesi, A.; Polito, L. Activities associated with the presence of ribosome-inactivating proteins increase in senescent and stressed leaves. FEBS Letters 1996, 382, 309–312. [Google Scholar] [CrossRef]
- Rippmann, J.F.; Michalowski, C.B.; Nelson, D.E.; Bohnert, H.J. Induction of a ribosome-inactivating protein upon environmental stress. Plant Molecular Biology 1997, 35, 701–709. [Google Scholar] [CrossRef]
- Martinez-Seidel, F.; Beine-Golovchuk, O.; Hsieh, Y.C.; Eshraky, K.E.; Gorka, M.; Cheong, B.E.; Jimenez-Posada, E.V.; Walther, D.; Skirycz, A.; Roessner, U.; Kopka, J.; Firmino, A.A.P. Spatially Enriched Paralog Rearrangements Argue Functionally Diverse Ribosomes Arise during Cold Acclimation in Arabidopsis. International Journal of Molecular Sciences 2021, 22, 6160. [Google Scholar] [CrossRef]
- Stefos, G.C.; Theodorou, G.; Politis, I. DNA G-quadruplexes: functional significance in plant and farm animal science. Animal Biotechnology 2021, 32, 262–271. [Google Scholar] [CrossRef]
- Bugaut, A.; Balasubramanian, S. 5’-UTR RNA G-quadruplexes: translation regulation and targeting. Nucleic Acids Research 2012, 40, 4727–4741. [Google Scholar] [CrossRef]
- Cho, H.; Cho, H.S.; Nam, H.; Jo, H.; Yoon, J.; Park, C.; Dang, T.V.T.; Kim, E.; Jeong, J.; Park, S.; Wallner, E.S.; Youn, H.; Park, J.; Jeon, J.; Ryu, H.; Greb, T.; Choi, K.; Lee, Y.; Jang, S.K.; Ban, C.; Hwang, I. Translational control of phloem development by RNA G-quadruplex–JULGI determines plant sink strength. Nature Plants 2018, 4, 376–390. [Google Scholar] [CrossRef] [PubMed]
- Volná, A.; Bartas, M.; Nezval, J.; Špunda, V.; Pečinka, P.; Červeň, J. Searching for G-Quadruplex-Binding Proteins in Plants: New Insight into Possible G-Quadruplex Regulation. BioTech 2021, 10, 20. [Google Scholar] [CrossRef] [PubMed]
- Tucker, W.O.; Shum, K.T.; Tanner, J.A. G-quadruplex DNA Aptamers and their Ligands: Structure, Function and Application. Current Pharmaceutical Design 2012, 18, 2014–2026. [Google Scholar] [CrossRef] [PubMed]
- Volná, A.; Bartas, M.; Karlický, V.; Nezval, J.; Kundrátová, K.; Pečinka, P.; Špunda, V.; Červeň, J. G-Quadruplex in Gene Encoding Large Subunit of Plant RNA Polymerase II: A Billion-Year-Old Story. International Journal of Molecular Sciences 2021, 22, 7381. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, M.; Duncan, S.; Yang, X.; Abdelhamid, M.A.S.; Huang, L.; Zhang, H.; Benfey, P.N.; Waller, Z.A.E.; Ding, Y. G-quadruplex structures trigger RNA phase separation. Nucleic Acids Research 2019. [Google Scholar] [CrossRef]
- Lukoszek, R.; Feist, P.; Ignatova, Z. Insights into the adaptive response of Arabidopsis thaliana to prolonged thermal stress by ribosomal profiling and RNA-Seq. BMC Plant Biology 2016, 16, 221. [Google Scholar] [CrossRef]
- Mullen, M.A.; Olson, K.J.; Dallaire, P.; Major, F.; Assmann, S.M.; Bevilacqua, P.C. RNA G-Quadruplexes in the model plant species Arabidopsis thaliana: prevalence and possible functional roles. Nucleic Acids Research 2010, 38, 8149–8163. [Google Scholar] [CrossRef]
- Cagirici, H.B.; Sen, T.Z. Genome-Wide Discovery of G-Quadruplexes in Wheat: Distribution and Putative Functional Roles. G3 Genes|Genomes|Genetics 2020, 10, 2021–2032. [Google Scholar] [CrossRef]
- Andorf, C.M.; Kopylov, M.; Dobbs, D.; Koch, K.E.; Stroupe, M.E.; Lawrence, C.J.; Bass, H.W. G-Quadruplex (G4) Motifs in the Maize (Zea mays L.) Genome Are Enriched at Specific Locations in Thousands of Genes Coupled to Energy Status, Hypoxia, Low Sugar, and Nutrient Deprivation. Journal of Genetics and Genomics 2014, 41, 627–647. [Google Scholar] [CrossRef]
- Cagirici, H.B.; Budak, H.; Sen, T.Z. Genome-wide discovery of G-quadruplexes in barley. Scientific Reports 2021, 11, 7876. [Google Scholar] [CrossRef]
- Kwok, C.K.; Balasubramanian, S. Targeted Detection of G-Quadruplexes in Cellular RNAs. Angewandte Chemie International Edition 2015, 54, 6751–6754. [Google Scholar] [CrossRef]
- Kwok, C.K.; Marsico, G.; Balasubramanian, S. Detecting RNA G-Quadruplexes (rG4s) in the Transcriptome. Cold Spring Harbor Perspectives in Biology 2018, 10, a032284. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Chow, E.Y.C.; Yeung, P.Y.; Zhang, Q.C.; Chan, T.F.; Kwok, C.K. Enhanced transcriptome-wide RNA G-quadruplex sequencing for low RNA input samples with rG4-seq 2.0. BMC Biology 2022, 20, 257. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yang, M.; Deng, H.; Ding, Y. New Era of Studying RNA Secondary Structure and Its Influence on Gene Regulation in Plants. Frontiers in Plant Science 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Kozak, M. How do eucaryotic ribosomes select initiation regions in messenger RNA? Cell 1978, 15, 1109–1123. [Google Scholar] [CrossRef]
- Kozak, M. An analysis of 5’-noncoding sequences from 699 vertebrate messenger RNAs. Nucleic Acids Research 1987, 15, 8125–8148. [Google Scholar] [CrossRef]
- MEIJER, H.A.; THOMAS, A.A. Control of eukaryotic protein synthesis by upstream open reading frames in the 5’-untranslated region of an mRNA. Biochemical Journal 2002, 367, 1–11. [Google Scholar] [CrossRef]
- Cavener, D.R.; Ray, S.C. Eukaryotic start and stop translation sites. Nucleic Acids Research 1991, 19, 3185–3192. [Google Scholar] [CrossRef]
- Kozak, M. Recognition of AUG and alternative initiator codons is augmented by G in position +4 but is not generally affected by the nucleotides in positions +5 and +6. The EMBO Journal 1997, 16, 2482–2492. [Google Scholar] [CrossRef]
- Hernández, G.; Osnaya, V.G.; Pérez-Martínez, X. Conservation and Variability of the AUG Initiation Codon Context in Eukaryotes. Trends in Biochemical Sciences 2019, 44, 1009–1021. [Google Scholar] [CrossRef]
- Gupta, P.; Rangan, L.; Ramesh, T.V.; Gupta, M. Comparative analysis of contextual bias around the translation initiation sites in plant genomes. Journal of Theoretical Biology 2016, 404, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, C.; Lin, J.; Yang, Y.; Peng, Y.; Tang, D.; Zhao, X.; Zhu, Y.; Liu, X. Over-expression of a glutamate dehydrogenase gene, MgGDH, from Magnaporthe grisea confers tolerance to dehydration stress in transgenic rice. Planta 2015, 241, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, M.B.; Ghalei, H.; Ward, E.A.; Potts, E.L.; Karbstein, K. Rps26 directs mRNA-specific translation by recognition of Kozak sequence elements. Nature Structural and Molecular Biology 2017, 24, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, M.B.; Barre, J.L.; Karbstein, K. Translational Reprogramming Provides a Blueprint for Cellular Adaptation. Cell Chemical Biology 2018, 25, 1372–1379.e3. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.; Wei, L. Characterizing the heat response of Arabidopsis thaliana from the perspective of codon usage bias and translational regulation. Journal of Plant Physiology 2019, 240, 153012. [Google Scholar] [CrossRef]
- Ganie, S.A.; Molla, K.A.; Henry, R.J.; Bhat, K.V.; Mondal, T.K. Advances in understanding salt tolerance in rice. Theoretical and Applied Genetics 2019, 132, 851–870. [Google Scholar] [CrossRef]
- Younesi-Melerdi, E.; Nematzadeh, G.A.; Shokri, E. Isolation and Gene Expression Investigation in Photosynthetic Isoform of Phosphoenolpyruvate Carboxylase Gene in Halophytic Grass Aeluropus Littoralis under Salinity Stress. Journal of Crop Breeding 2018, 9, 69–78. [Google Scholar] [CrossRef]
- Singha, H.S.; Chakraborty, S.; Deka, H. Stress induced MAPK genes show distinct pattern of codon usage in Arabidopsis thaliana, Glycine max and Oryza sativa. Bioinformation 2014, 10, 436–442. [Google Scholar] [CrossRef]
- Mohasses, F.C.; Solouki, M.; Ghareyazie, B.; Fahmideh, L.; Mohsenpour, M. Correlation between gene expression levels under drought stress and synonymous codon usage in rice plant by in-silico study. PLOS ONE 2020, 15, e0237334. [Google Scholar] [CrossRef]
- Almutairi, M.M.; Almotairy, H.M. Analysis of Heat Shock Proteins Based on Amino Acids for the Tomato Genome. Genes 2022, 13, 2014. [Google Scholar] [CrossRef]
- Wu, B.; Qiao, J.; Wang, X.; Liu, M.; Xu, S.; Sun, D. Factors affecting the rapid changes of protein under short-term heat stress. BMC Genomics 2021, 22, 263. [Google Scholar] [CrossRef] [PubMed]
- Kage, U.; Powell, J.J.; Gardiner, D.M.; Kazan, K. Ribosome profiling in plants: what is not lost in translation? Journal of Experimental Botany 2020, 71, 5323–5332. [Google Scholar] [CrossRef] [PubMed]
- Ingolia, N.T. Ribosome profiling: new views of translation, from single codons to genome scale. Nature Reviews Genetics 2014, 15, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Shi, J.; Chen, J.; Zhang, M.; Sun, S.; Xie, S.; Li, X.; Zeng, B.; Peng, L.; Hauck, A.; Zhao, H.; Song, W.; Fan, Z.; Lai, J. Ribosome profiling reveals dynamic translational landscape in maize seedlings under drought stress. The Plant Journal 2015, 84, 1206–1218. [Google Scholar] [CrossRef]
- Wu, H.Y.L.; Song, G.; Walley, J.W.; Hsu, P.Y. The Tomato Translational Landscape Revealed by Transcriptome Assembly and Ribosome Profiling. Plant Physiology 2019, 181, 367–380. [Google Scholar] [CrossRef]
- Juntawong, P.; Girke, T.; Bazin, J.; Bailey-Serres, J. Translational dynamics revealed by genome-wide profiling of ribosome footprints in Arabidopsis. Proceedings of the National Academy of Sciences 2014, 111. [Google Scholar] [CrossRef]
- Zanetti, M.E.; Chang, I.F.; Gong, F.; Galbraith, D.W.; Bailey-Serres, J. Immunopurification of Polyribosomal Complexes of Arabidopsis for Global Analysis of Gene Expression. Plant Physiology 2005, 138, 624–635. [Google Scholar] [CrossRef]
- Mustroph, A.; Zanetti, M.E.; Jang, C.J.H.; Holtan, H.E.; Repetti, P.P.; Galbraith, D.W.; Girke, T.; Bailey-Serres, J. Profiling translatomes of discrete cell populations resolves altered cellular priorities during hypoxia in Arabidopsis. Proceedings of the National Academy of Sciences 2009, 106, 18843–18848. [Google Scholar] [CrossRef]
- Sablok, G.; Powell, J.J.; Kazan, K. Emerging Roles and Landscape of Translating mRNAs in Plants. Frontiers in Plant Science 2017, 8. [Google Scholar] [CrossRef]
- Wang, X.W.; Liu, C.X.; Chen, L.L.; Zhang, Q.C. RNA structure probing uncovers RNA structure-dependent biological functions. Nature Chemical Biology 2021, 17, 755–766. [Google Scholar] [CrossRef]
- Talkish, J.; May, G.; Lin, Y.; Woolford, J.L.; McManus, C.J. Mod-seq: high-throughput sequencing for chemical probing of RNA structure. RNA 2014, 20, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Incarnato, D.; Neri, F.; Anselmi, F.; Oliviero, S. Genome-wide profiling of mouse RNA secondary structures reveals key features of the mammalian transcriptome. Genome Biology 2014, 15, 491. [Google Scholar] [CrossRef] [PubMed]
- Spitale, R.C.; Flynn, R.A.; Zhang, Q.C.; Crisalli, P.; Lee, B.; Jung, J.W.; Kuchelmeister, H.Y.; Batista, P.J.; Torre, E.A.; Kool, E.T.; Chang, H.Y. Structural imprints in vivo decode RNA regulatory mechanisms. Nature 2015, 519, 486–490. [Google Scholar] [CrossRef]
- Smola, M.J.; Weeks, K.M. In-cell RNA structure probing with SHAPE-MaP. Nature Protocols 2018, 13, 1181–1195. [Google Scholar] [CrossRef] [PubMed]
- Silverman, I.M.; Gregory, B.D. Transcriptome-wide ribonuclease-mediated protein footprinting to identify RNA–protein interaction sites. Methods 2015, 72, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Silverman, I.M.; Li, F.; Alexander, A.; Goff, L.; Trapnell, C.; Rinn, J.L.; Gregory, B.D. RNase-mediated protein footprint sequencing reveals protein-binding sites throughout the human transcriptome. Genome Biology 2014, 15, R3. [Google Scholar] [CrossRef]
- Singh, J.; Hanson, J.; Paliwal, K.; Zhou, Y. RNA secondary structure prediction using an ensemble of two-dimensional deep neural networks and transfer learning. Nature Communications 2019, 10, 5407. [Google Scholar] [CrossRef]
- Sato, K.; Akiyama, M.; Sakakibara, Y. RNA secondary structure prediction using deep learning with thermodynamic integration. Nature Communications 2021, 12, 941. [Google Scholar] [CrossRef]
- Sahoo, S.; Singh, D.; Singh, A.; Eswarappa, S.M. Identification of mRNAs that undergo stop codon readthrough in Arabidopsis thaliana. bioRxiv, 2021. [Google Scholar] [CrossRef]
- Wu, X.; Wang, J.; Wu, X.; Hong, Y.; Li, Q.Q. Heat Shock Responsive Gene Expression Modulated by mRNA Poly(A) Tail Length. Frontiers in Plant Science 2020, 11. [Google Scholar] [CrossRef]
- Bonnot, T.; Nagel, D.H. Time of the day prioritizes the pool of translating mRNAs in response to heat stress. The Plant Cell 2021, 33, 2164–2182. [Google Scholar] [CrossRef]
Figure 1.
A general scheme of detection and coping with abiotic factors by plants. ER - endoplasmic reticulum.
Figure 1.
A general scheme of detection and coping with abiotic factors by plants. ER - endoplasmic reticulum.
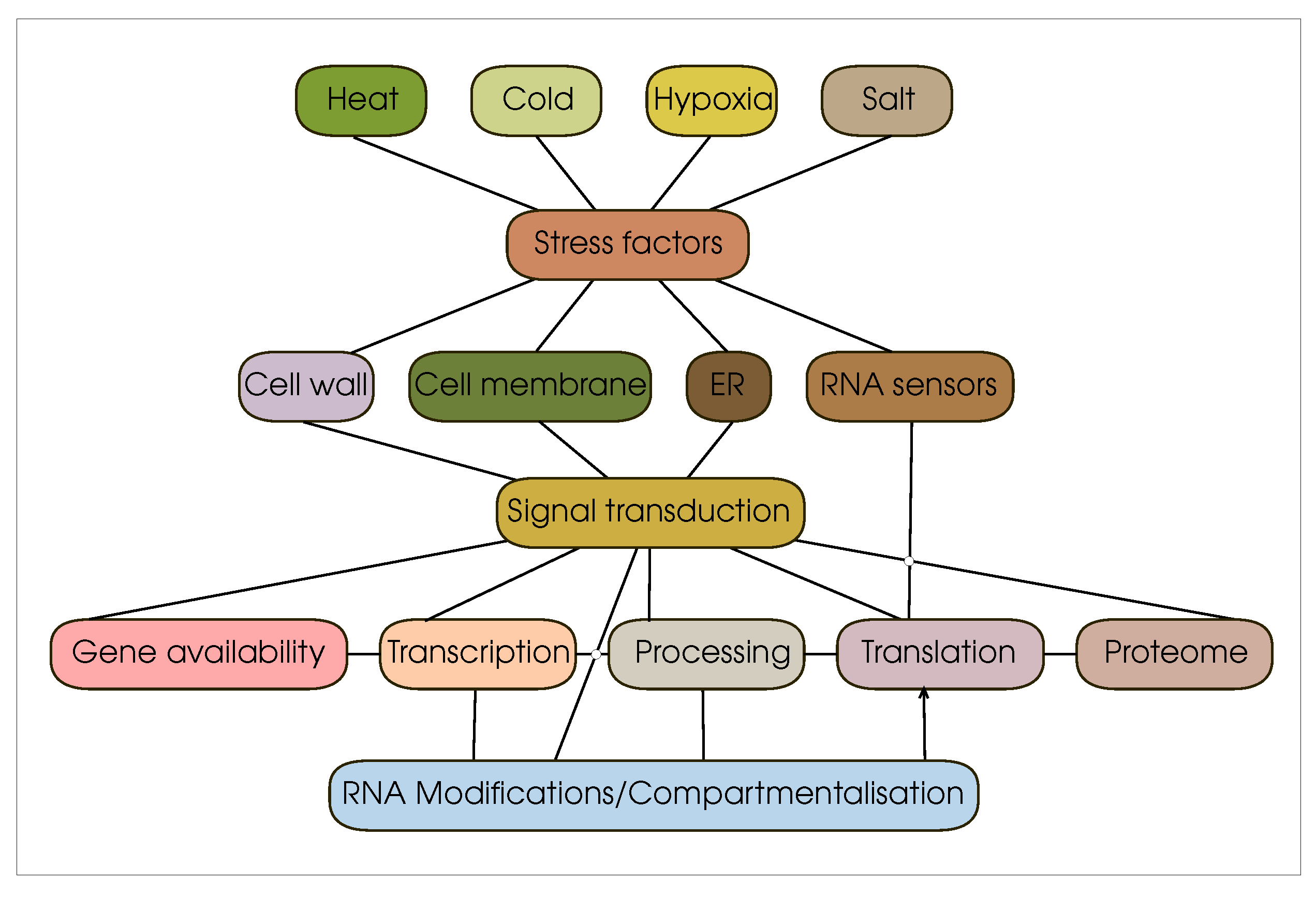
Figure 2.
The journey of mRNA from translation initiation to the nascent peptide chain. Key regulatory elements and patterns: Cap – five-prime cap, 43S – 43S preinitiation complex, IRES – internal ribosome entry site, ITAFs – IRES-transacting factors , – -methyladenosine, G4 – G-quadruplexes, uORF – upstream open reading frames, mAUG – main AUG, KOZAK – Kozak consensus sequences, aatRNA – aminoacetilated tRNA, eIF* – eukariotic initiation factors, eEF* – eukariotic elongation factors. Edges corresponding to the direct course of the translation process are marked in black; cycles within the translation are marked in red.
Figure 2.
The journey of mRNA from translation initiation to the nascent peptide chain. Key regulatory elements and patterns: Cap – five-prime cap, 43S – 43S preinitiation complex, IRES – internal ribosome entry site, ITAFs – IRES-transacting factors , – -methyladenosine, G4 – G-quadruplexes, uORF – upstream open reading frames, mAUG – main AUG, KOZAK – Kozak consensus sequences, aatRNA – aminoacetilated tRNA, eIF* – eukariotic initiation factors, eEF* – eukariotic elongation factors. Edges corresponding to the direct course of the translation process are marked in black; cycles within the translation are marked in red.
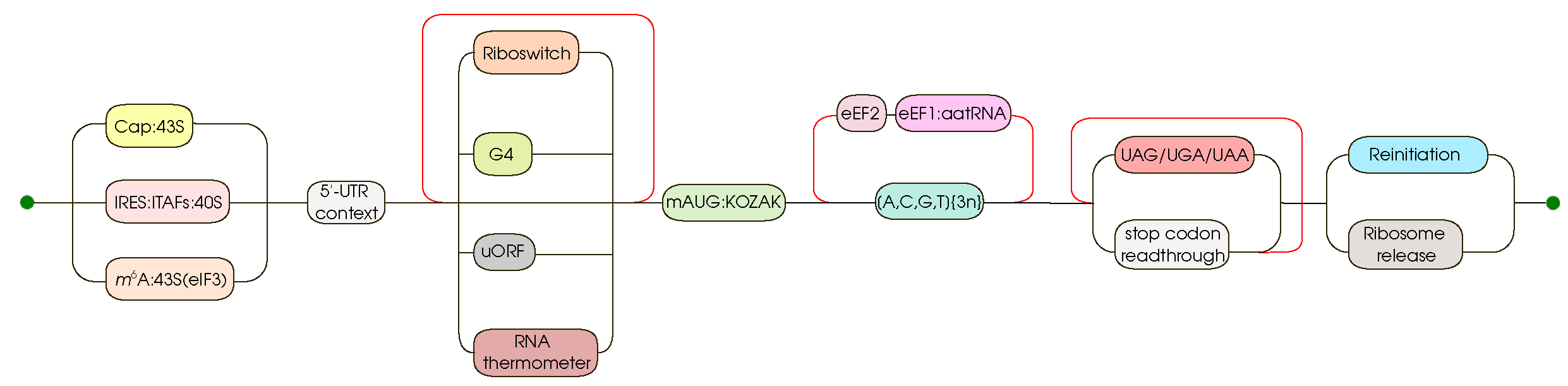
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



