
Submitted:
17 January 2024
Posted:
17 January 2024
You are already at the latest version
Abstract
Physical exercise is the greatest stress on the cardiovascular system, not only in humans but also in all vertebrates. Consequently, the cardiovascular response cannot be thought of as having only one "anthropocentric" interpretation. The modulation of cardiac output (Q) in response to exercise in the five major groups of vertebrates exhibits considerable variability. Moreover, the response of the heart rate (HR) and the stroke volume (SV), the fundamental factors that cause the rise in Q, is also highly variable. The difference in the range of HR is difficult to determine in many vertebrates, both at rest and at maximal effort. Additionally, the increase in SV varies among vertebrates. For example, while rainbow trout and leopards increase their SV by increasing end diastolic volume (EDV), humans do so at the expense of both increasing EDV and reducing end systolic volume (ESV). The variation in arterial pressure that arises during exercise varies notably among vertebrates, such as amphibians, whose blood pressure values do not increase as significantly as in mammals. Such differences in cardiovascular response among vertebrates may be linked to their habitat or living environment. This review aims to analyze the cardiovascular response of vertebrates to physical exercise as the most common stressor.
Keywords:
vertebrates
; arterial pressure
; cardiac output
; exercise
; heart rate
; stroke volume
1. Introduction
Human interest in sport medicine and exercise physiology began in the 19th century. Some decades later, other athletic mammals received some attention, such as racing horses, dogs, and camels. In general terms, exercise is the greatest stress on the cardiovascular system, not only for humans but for all vertebrates. For this reason, the cardiovascular response cannot be considered to have only one “anthropocentric” meaning [1,2,3].
Athletic performance relies on aerobic capacity to generate energy for the exercising muscles, facilitated by an increase in HR and SV (the ability to increase Q), among other factors [4,5]. However, research on the cardiovascular response in vertebrates has serious limitations since its study is conditioned and limited by the methods needed for its assessment, such as stress tests, and the different models of locomotion (aquatic, terrestrial, aerial, and their combinations) in the different groups of vertebrates. Therefore, various types of ergometers are used, such as treadmills, flight tunnels, swimming tunnels, and running wheels. Other ways to measure cardiovascular response to physical activity include thermal stress due to cold or the straightening reflex [6,7].
Vertebrates that undertake enormous migrations, lasting from a few days to weeks, need to increase their metabolism eightfold in birds [8,9], and up to fifteenfold in salmon [10]. They must adjust their cardiovascular system correspondingly to sustain such considerable metabolic activity. Migratory birds maintain a high level of cardiovascular activity equivalent to the energy cost of a marathon competition in humans, ranging between 62 and 100% of their maximal rate of oxygen consumption (VO2max) [11].
The general characteristics of the cardiovascular system in vertebrates have previously been described [12]. Throughout the evolutionary process, all vertebrates have possessed a myogenic heart with evident intrinsic activity, although there are exceptions in fish that have more than one heart. This intrinsic activity of the heart has been demonstrated in vertebrate precursors, tunicates, and cephalochordates [13]. Blood flows from the heart to the head and the rest of the organism, and there are also one-way valves, which open and close, allowing only one direction of the blood to flow through them. The venous valves are considered a “passive” system [14,15]. The blood vessels exhibit vasomotor activity, such that changes in the degree of contraction of the smooth muscles alter the peripheral resistance, variations in blood pressure, and the storage of blood in the periphery [16,17]. Although with certain limitations, as occurs at the capillary level with the exchange of liquids, the cardiovascular system can be functionally considered a closed system [18].
Apart from the characteristics already mentioned, significant heterogeneity exists among the five principal groups of vertebrates, including fishes, amphibians, reptiles, birds, and mammals, in relation to their cardiovascular response to exercise [6,7,19,20].
In the closest living relatives of vertebrates, tunicates and cephalochordates, blood is transported in clear vessels to a range of vascular beds [21,22]. Differences are observed within the fish division; for instance, specific classes can breathe atmospheric air for a certain duration of time [23,24,25].
The amphibian heart consists of two anatomically separated atria, left and right, which receive blood from both the lungs and the systemic venous circulation, respectively. This blood then enters a ventricle that is highly trabeculated and undivided. The trabeculae, forming deep sacs, gather and retain blood during diastolic filling. This mechanism appears to enable a partial separation of the blood not only during this final phase but also while the blood is being expelled during systole [26,27,28]. There are also exceptions, such as tadpoles and some adult salamanders, which have pulmonocutaneous circulation. Nevertheless, notable differences exist between the two main classes of amphibians: Anura (frogs and toads) and Caudata (for example, the salamander) [15,29,30].
The differences among the classes of reptiles have been analyzed previously. The heart and central circulation in reptiles exhibit anatomical complexity, resulting in a parallel connection between the systemic and pulmonary systems. Furthermore, reptiles have a lower ratio of heart mass to body mass compared to mammals or birds. Extensive research has focused on the physiological implications of intermittent breathing patterns during both terrestrial and aquatic activities. Notably, reptiles demonstrate lower Q, which is a product of SV, mean blood pressure, and HR, when compared to mammals and birds [31,32,33,34].
The cardiovascular system of non-crocodilian reptiles is a paradigm of intracardiac shunting [12,35,36,37]. Cardiovascular shunts have been acquired as an adaptation associated with low metabolic activity and intermittent forms of apnea, especially in vertebrate ectotherms. The single ventricle of these vertebrates is partially divided by a muscular protrusion (homologous to the spiral valve of the amphibians heart) that partially separates flow and expels the blood from the ventricle according to the systemic and pulmonary vascular resistance Although there are two separate atria, the existence of a single ventricle with this valve device allows the pulmonary and systemic venous blood to be mixed [32,38,39,40].
Therefore, some reptiles with an undivided ventricle (except crocodilians, pythons, and monitor lizards) can perform right-left or left-right shunts. Possible theories regarding the functions of these derivations have been described by Burggren et al., giving them a holistic and integrative sense, and they may have several implications during exercise. In general, increasing exercise intensity results in cardiovascular bypass [38]. The right-left shunt allows the recirculation of venous blood to re-enter the systemic circulation and, consequently, a decrease in oxygen saturation, while the left-right shunt produces an increase in oxygen saturation and an improvement in tissue oxygenation in reptiles, aquatic anurans, and air-breathing fish. During exercise, amphibians and non-crocodilian reptiles have a left-right shunt [6,24,31,41,42,43,44,45,46,47,48,49,50,51,52].
Right-left shunts (Figure 1) would have a beneficial function in lowering metabolism, resulting in a decrease in blood flow at the pulmonary level and a decrease in the amount of oxygen in the body. The reduction of metabolism would reduce energy consumption when working conditions are more exigent, such as prolonged periods of apnea. On the other hand, the left-right derivations (Figure 2) minimize the possible inequalities of the ventilation/perfusion ratio, allowing more oxygen to reach the brain and upper body in amphibians and reptiles [13,18,32,38,53].
The heart of the crocodile is an example of extracardiac shunting, preventing the mixing of blood in the heart, which is common in other reptiles and amphibians, as cardiovascular shunting is produced outside the heart. The outflow tract is formed by the pulmonary and aortic components. The aortic component is double, with one aorta arising from the left ventricle and a second aorta taking its origin from the right ventricle, together with the pulmonary component [54,55].
Finally, the cardiovascular system of birds and mammals is completely divided into a pulmonary circuit, with low pressure, leaving the right ventricle to irrigate the lungs through the pulmonary artery, and a systemic circuit, with high pressure, originating from the left ventricle through the aorta to perfuse the rest of the body [56,57,58,59]. However, comparisons of the cardiovascular anatomy among genera and families, both of birds and mammals, reveal some differences [60,61,62]. Among them are the higher Q with the same body mass of birds compared to mammals, the more elongated heart of birds, and the greater cardiac mass compared to the body mass of birds in relation to mammals, finding significant differences between bird families. This may be due to the great need for aerobic power required to maintain flutter during flight [63].
HR in vertebrates is determined by the activity of the heart pacemaker and various factors, such us temperature, acetylcholine, adrenaline, or calcium, that modulate its electrical activity via nervous and hormonal pathways [64,65,66,67,68]. SV is determined by contractility and the characteristics of the ventricle, mainly regarding its filling capacity. Similarly, the control of the SV is accomplished by two different pathways, nervous and hormonal [7,17,69].
Although there are many common anatomic and functional characteristics in all vertebrates, the cardiovascular requirements in the five groups of vertebrates are highly dependent on the type of circulation and the environment. The domain of comparative physiology is inevitably linked to Laureate August Krogh (Nobel Prize) and adheres to the 'Krogh principle': "for many problems, there is an animal on which it can be most conveniently studied” [70]. By conducting comparative studies among various groups of vertebrates, research in exercise physiology in humans can be enriched.
The “amplitude” of the cardiovascular response is difficult to define because, except for vertebrates used in experimentation and some mammals, the values of rest and maximal exercise are not very precise. The knowledge and research on the cardiovascular response to physical exercise in various classes of vertebrates can assist in evaluating human cardiovascular responses during physical activity in diverse environmental conditions, including extreme temperatures, aquatic exercise, and high altitudes. Furthermore, this information may provide valuable insights into the interpretation of cardiovascular responses in humans engaging in physical exercise with disabilities or under pathological conditions. Previous reviews have focused on factors that limit physical exercise performance in vertebrates without emphasizing cardiovascular response [6,7]. A comparative analysis of cardiovascular parameters, together with an integrated description of the variables involved in the cardiovascular system's response to physical exercise in vertebrate groups, is essential. Focusing solely on the cardiovascular components is inadequate for understanding the adaptations in each vertebrate group, considering the diverse and individual variables attributed to each group.
The aim of this review is to examine the cardiovascular response in vertebrates based on hemodynamic parameters and the general hemodynamic equation, which establishes that the mean arterial pressure (MAP) equals Q divided by total peripheral resistance (TPR). Additionally, it aims to evaluate the integrated response of these cardiovascular variables, contributing to the adaptations to physical exercise across various vertebrate groups. The regulation of circulatory parameters during physical activity exhibits significant variation among vertebrates, attributable to their body structure and other determining factors affecting the regulation of Q and TPR. The parameters governing Q (the product of SV and HR) display substantial variability among the five groups of vertebrates, with some increasing Q via an increase in SV, while others achieve this solely through elevated HR.
2. Methodology
This paper constitutes a narrative review. The identification of research studies involved a comprehensive search across multiple databases, including PubMed, The Web of Science, Scopus, Google Scholar, and citation searches. The search terms included cardiovascular system, physical exercise, maximal effort, cardiovascular response, cardiovascular adaptations, exercise physiology, vertebrates, fish, amphibians, reptiles, birds, and mammals. These search terms were utilized in conjunction with the advanced tools provided by the search engines. The review did not set an arbitrary start date for the inclusion of studies. The searches were conducted up until September 2023.
3. Adjustment of the HR to exercise in vertebrates
All vertebrates have a pacemaker. The heart's activity is modulated by neural and hormonal factors. The intrinsic rate of the pacemaker shows important differences among the distinct groups of vertebrates [71,72,73]. Resting HR and maximal HR in ectotherm vertebrates are generally lower than those of endotherms, although there are exceptions such as tuna and small reptiles [74,75]. Despite all the problems related to determining resting HR (such as temperature, ventilatory arrhythmia, and the definition of the resting state), the differences are related to phylogeny, body mass, temperature, and, to a certain extent, the capacity to perform exercise [71,76].
Evidence indicates an allometric relationship with an exponent of -0.25 between body mass and resting HR, reaching its maximum in birds and small mammals [12]. The significant contrast in HR between the smallest birds and the largest mammals, as shown in Table 1, is reflected in this relationship. The considerable variations may be attributed, at least in part, to differences in sodium (Na+) conductance, which determines the slope of the potential of slow depolarization in the pacemaker cells [71,77].
Although the data are scarce, there is an allometric relationship with a similar exponent in snakes and frogs [12]. Resting HR in fit horses, for example, ranges from 30 to 40 bpm [78] and from 50 to 60 bpm in athletic humans under similar conditions [79,80]. Based on limited data, the maximum heart rate (HRmax) during exercise for the majority of the lower vertebrates is 120 bpm [81]. Tuna is the only known exception to this generalization, as a HRmax of more than 200 bpm has been found [71].
The upper limit of HR in lower vertebrates appears to be unrelated to body mass, suggesting a phylogenetic constraint associated with the pacemaker rate or modulating mechanisms [71,77]. Given the recognized scale of HR to body mass, it is probable that the highest HR values are found in early development when the animal is a neonate or still relatively young. Other recent studies have found that in alligators and turtles, HR did not scale with body mass after injecting adrenaline. The lack of a scaling pattern contradicts prior studies on reptiles demonstrating HR decreases with body mass [82,83].
In the studied vertebrates, HR and its adjustment to physical exercise show notable differences and could be influenced by several intrinsic factors, such as age (as has already been evidenced in humans) [65,84,85,86,87,88,89]. As illustrated in Table 1, fish, amphibians, and reptiles show a limited range of HR. This may be because the environment in which they live do not require them to considerably alter their cardiovascular system, i.e., increase their Q. Another possible explanation could be that the adjustment of Q is produced by the Frank-Starling mechanism or contractility. Finally, the small increase in HR in response to exercise in these vertebrates may show the increase complexity of the control mechanisms that alter HR throughout evolution [7,56,74,90,91,92,93,94,95,96].

Table 1.
Examples of resting and exercise heart rates in different classes of vertebrates. HR differ even among different animals in the same group of vertebrates. Data: from various sources [56,74,92,93,94,95]. Abbreviations: HR-heart rate; bpm-beats per minute; HRmax-maximal heart rate.
| Vertebrate | Resting HR (bpm) | Exercise HRmax (bpm) |
|---|---|---|
| Human | 80 | 190 |
| Rat (mammals) | 320 | 600 |
| Dog (mammals) | 100 | 300 |
| Horse (mammals) | 38 | 207 |
| Golden-collared manakins (birds) | 250 | 1300 |
| Tuna (fishes) | 100 | 205 |
| Salmon (fishes) | 20 | 85 |
| Frog (amphibians) | 25 | 100 |
| Rhinella marina (amphibians) | 20 | 105 |
| Ophisaurus (lizard) (reptiles) | 20 | 60 |
| Iguana (reptiles) | 30 | 100 |
During strenuous physical exercise and VO2max conditions, Thoroughbred horses can reach an HRmax close to 220 bpm, and maximum values ranging from 180 to 200 bpm have been found in humans at a similar exercise intensity. These data show the ability to increase the HR from rest to maximal exercise intensity by an order of five to eightfold and three to fourfold in the equine and human species, respectively [97,98,99,100]. Surprisingly, ventricular filling does not appear to be compromised at maximal exercise, despite heart rates of 4 beats/s [101].These cardiovascular adjustments in response to exercise are one of the factors that may limit the VO2max in some vertebrates [3,6,102].
The control mechanisms of the HR (endocrine and nervous control) seem to show an evolutionary tendency. Catecholamines are present in all vertebrates; however, adrenergic cardiac innervation is lacking in Cyclostomata and Elasmobranchii. Thus, from an evolutionary perspective, tachycardia mediated by adrenaline seems to have acquired greater importance in birds and mammals. The muscarinic receptors mediate cholinergic inhibition of heart rate (HR), influenced by a cardiac branch of the vagus nerve, and are present in all vertebrates except certain fish [68,103,104,105,106,107].
The resting HR is influenced by both autonomic and non-autonomic stimuli. Horses and young people are predominantly under parasympathetic control, which tends to shift to a sympathetic predominance as aging progresses. This modification could be due to decreased parasympathetic or increased sympathetic nervous system input, as well as an adjustment in plasma volume and cardiac filling pressure, or a combination of all these factors [68,108,109,110,111,112].
4. Adjustment of the SV to exercise in vertebrates
SV is under the control of the Frank-Starling mechanism and contractility. The participation of each of the mechanisms in different vertebrates depends on factors such as the Frank-Starling mechanism. The Frank-Starling curves, or the curves of ventricular function, have been established in several groups of vertebrates [14,71,113,114,115]. Figure 3 shows variations in SV with respect to cardiac pressure.
The right ventricle of mammals is more sensitive to filling pressure than the left ventricle. It is not known if there are differences in the sensitivity of the right and left sides of the heart to filling pressure in amphibians and non-crocodilian reptiles, perhaps to help with intracardiac shunting [38,117].
Pulmonary arterial pressure in reptiles is several times higher than that in mammals when pulmonary blood flow increases, so there could be a different mechanism for adjusting SV during physical exercise [12].
Increases in SV of 10%-30% are observed in humans during exercise [116,118]. The relative increase in SV from rest to exercise for a racehorse and humans athletes has been measured and it can be doubled at high intensities [19,119]. Thoroughbred horses exhibit exceptional characteristics, doubling SV and tripling HR, resulting in an up to sixfold increase in Q. In these animals, it has been identified that there is an increase in SV from 1000 ml at rest up to 1700 ml at maximal exercise, consequent to an increase in blood volume, venous return, and filling pressures according to the Frank-Starling mechanism and it is highly enhanced by training [3,99,100,120]. The horse has evolved not only in accordance with the laws of nature like other vertebrates but also has been subjected to a thousand years of selective breeding based on athletic performance. This has created a disproportionate increase in the horse’s heart size and, consequently, the SV, which increases sharply at the onset of the exercise from 2-2,5 ml/kg until around the 40% VO2max. At maximal exercise, SV reaches 3-4 ml/kg and is determined principally by heart size [101]. This response is shown to a greater extent in the equine species than in humans, and it explains the enormous magnitude of the peak aerobic power in the horse compared to the elite human athlete [121,122]. Certainly, "athlete" horses are an exception among all vertebrates, so it is difficult to compare the results with other groups of vertebrates whose activity is limited to their ordinary lives.
In species where SV does not significantly rise during exercise, it is likely that the heart operates at the apex of the Frank-Starling curve under resting eupneic conditions. The Frank-Starling mechanism also maintains a long-term equilibrium between the Q of the right and left ventricles in mammalian hearts. Certain amphibians and reptiles exhibit slight reductions in SV during exercise, accompanied by increases in HR. The most notable alterations in SV in reptiles and amphibians occur as decreases associated with apnea. The greatest decrease in SV seen in these animals is associated with the apnea required during their physical effort in the water [7,17,53,66,69,86,123].
Unlike other vertebrates, certain fish species, including cyclostomata and teleosts, exhibit a twofold increase in SV during exercise [17,87,94]. The increase clearly underscores the functional importance of the Frank-Starling mechanism. In fact, fish appear to rely on HR modulation to achieve the augmented Q associated with aerobic exercise [65,67]. These differences in the SV response in fish may be a consequence of one of the following factors: greater sympathetic control of HR; the atria becoming less sensitive to filling pressure; a greater degree of sympathetic control of the tension in the vein walls; and a better shortening fraction [71,124].
Homeometric regulation is described as the capacity of the cardiac muscle to maintain blood flow independently of the development of pressure, in contrast to heterometric regulation (Starling’s Law). This factor is thoroughly documented in the hearts of various fish classes and might explain why alterations in vascular resistance do not affect SV despite the changes associated with cardiac functioning [71,125,126].
Concerning the factors mentioned above, it is noteworthy that the sensitivity of fish hearts to filling pressure is nearly ten times higher than that of mammals (approximately 1-2 mmHg compared to 10-20 mmHg for a maximal response). This heightened sensitivity may be linked to the fact that the atrial chamber in fish has thinner walls and is more distensible [12]. Moreover, intracardiac shunting may require stricter regulation of the dimensions of the ventricle, thus limiting SV but not HR [38].
Contractility is an easily defined cardiac function that is difficult to assess. Contractility is defined as the change undergone by the ventricle or atria when the parameters that can affect myocardial performance (HR, filling volume and pressure and arterial diastolic pressure) are kept constant. The most accurate assessment is through the peak derivative of left ventricular pressure in relation to time (LV dp/dt), which seems to rise progressively with exercise intensity but remains unchanged with training [12,127,128].
Studies have demonstrated that the values for mammals and tuna are five times greater than those of teleosts (370-480 mmHg/s), whereas the values for cyclostomata (22 mmHg/s) and sharks (30 mmHg/s) are ten times lower. Intermediate values have been recorded for lizards (95-180 mmHg/s) and anura, including Rhinella marina (toad) (60 mmHg/s and 110 mmHg/s) [124].
These differences may be due to different factors such as HR (the phenomenon known as the "negative staircase" refers to the inverse relationship between contraction rate and maximum isometric tension (Tmax); the increase in the frequency of electrical stimulation leads to a decrease in Tmax), temperature (Tmax decreases at high temperatures as a result of a shorter action potential duration), beta-adrenergic stimulation (in the cardiac strips of frogs and fish severely increases Tmax), extracellular calcium (an increase in extracellular [Ca2+] within the range of 1-9 mM induces an elevation in the maximum isometric tension (Tmax) of cardiac muscle), and inotropic agents (negative inotropic effects can be elicited by hypoxia, acidosis, and acetylcholine) [81,103,129,130,131,132,133,134,135,136,137].
Associated with contractile activity and the relationship between length and pressure, result of Starling’s Law and ventricular performance, SV increases due to of the increase in EDV and decrease in ESV, as SV is the difference between them (EDV-ESV). The ventricular ejection fraction in the rainbow trout is usually around 100% [138,139], while that of the leopard shark is 80% [140]. Hence, the significant increases in SV observed in fish arise from the corresponding increases in EDV. In amphibians and reptiles, the duration of the systole is 50% of the cardiac cycle, which potentially leading to a small ESV, which can be fundamental for intracardiac shunting [38,141]. At elevated heart rates, there is a decrease in filling time, which may account for the observed reduction in perfusion in the rainbow trout heart [142]. Similarly, in dogs, an increase in HR from 120 bpm to 180 bpm has minimal impact on Q due to the decrease in SV [143].
Unexpectedly, the maximum SV in various fish and mammals shows little variation (approximately 0.5-1.5 ml/kg of body mass). This narrow range might signify an anatomical limitation, as hearts with a larger volume require a disproportionately thicker wall to uphold the same tension, as per Laplace's Law [71]. A thicker cardiac wall presents a challenge to O2 diffusion. Therefore, the maximum SV is probably established as a compromise between the generation of wall tension and the adequate supply of myocardial O2. The notion that animals with a substantial SV exhibit low arterial pressure aligns with this concept. For instance, amphibians and non-crocodilian reptiles have a greater SV (3-5 ml/kg of body mass) compared to fish and mammals [71]. Moreover, the hearts of Antarctic fish demonstrate an exceptionally high SV (2-10 ml/kg of body mass) and a notably limited homeometric capacity [144].
Figure 4 shows the differences between a rainbow trout's cardiac cycle and that of human. While the rainbow trout seems to depend only on the Frank-Starling mechanism, the human improves its ejective capacity by increasing the final diastolic volume (Frank-Starling's law) and decreasing the ESV (contractility increase).
In most vertebrates, cardiac filling follows a vis-a-tergo (force from behind) pattern, where central venous blood pressure plays a crucial role in determining both cardiac filling and SV. In mammalian hearts, atria and ventricular filling occur simultaneously due to a favorable pressure gradient between the central veins and the heart chambers. As a result, filling pressure at the venous level serves as the primary determinant of both atrial and ventricular filling. Atria contraction alone contributes approximately 25% to ventricular filling, while a small portion results from the elastic recoil of the ventricle [17].
5. Adjustment of mean arterial pressure (MAP) to exercise in vertebrates
MAP is the product of Q and TPR. It is a parameter under the control of cardiovascular regulatory mechanisms, with the most important being pressure receptors (baroreceptors). In general, there is an evolutionary tendency towards an increase in pressure as progress is made from primitive circulatory systems to birds and endothermic mammals [12,145].
Species of fish adapted to fast swimming and high physiological performance have high levels of arterial pressure, exceeding 100 mm Hg in tuna and salmon [91]. The hearts of amphibians generally do not generate such high pressure. This could be linked to the single ventricle performing the function for both the pulmonary and systemic circuits [146].
Turtles have the lowest systemic arterial pressure, and terrestrial species have slightly higher pressure than the semi-aquatic species. Systemic arterial pressure at rest varies considerably among snakes, in part due to their diversified exposure to gravitational forces. There is an allometric relationship in snakes between arterial pressure and body mass (exponent = 0.15), probably because ventricular mass increases with the increase in body mass [147]. Resting systemic arterial pressure in crocodilians and lizards is similar to that of mammals, and is almost double in iguanas (finding mean blood pressure values in the iguana of 43.5 mmHg, in the varanus of 70 mmHg and in humans of 100 mmHg) [12,83].
The development of high arterial pressures is a feature of birds and mammals. The MAP is approximately 97 mmHg in the "average" mammal, whereas the "average" bird has a MAP of about 133 mmHg [148,149]. These variances appear to be associated with a higher Q (related to mass or metabolic rate) in birds rather than differences in peripheral resistance [150]. The literature describes a correlation between MAP and the vertical distance between the heart and brain in mammals; for example, giraffes would present a higher MAP to perfuse the brain [149]. However, compared to mammals, birds have larger hearts, a greater SV, and a lower HR. During exercise, MAP increases more than 50% above resting levels in these groups. As an example, MAP rises from 110 mmHg to 138 mmHg at rest to as high as 200 mmHg during maximal exercise [151].
Traditionally, books on physiology deal with mechanisms for regulating arterial pressure in the short, medium, and long term. In short-term regulation, the innervation of the heart is very varied [152]. In teleosts vagal innervation shows a higher tone at rest. The heart of amphibians has sympathetic and parasympathetic innervation, both in the caudata and anura. The activity of anura leads to an increase in both HR and arterial pressure. Initially, this rise is due to adrenergic effluents and is sustained by circulating catecholamines. In reptiles, there is considerable variation in blood pressure levels, influenced by differences among species, environmental conditions, and non-steady states resulting from various causes. In spite of the high HR in relation to many other vertebrates, the heart of birds is subject to an important cholinergic (vagal) and adrenergic tone. Walking increases blood pressure in several types of birds, particularly at high intensities, due to the increase in sympathetic tone and, to a lesser extent, to vagal inhibition [153].
All the mentioned data suggest that the baroreflex function in the immediate control of arterial pressure is very important in amphibians (anura), reptiles, birds, and mammals, with the latter being the group that has provided the most evidence about this issue. Moreover, the central nervous system (CNS) plays a role in mediating diverse cardiovascular responses across all vertebrates, including those associated with excitement, emotions, and cognitive functions. Several of these responses are immediate, such as the defense reactions involving tachycardia and elevated blood pressure. Mammalian central nervous mechanisms can inhibit the baroreceptor reflex responses during exercise, and there is evidence suggesting that the brain can modify the reference point around which arterial pressure is controlled or stabilized over the long term. The CNS is crucial in integrating both the heart and circulation and controlling sustained sympathetic activity, which is essential for maintaining peripheral vascular tone and Q. Without sympathetic vascular tone, the reflex controls cannot effectively stabilize pressure at any moment. Skeletal muscle arterial and arteriolar vasodilation allow a fall in TPR in exercising mammals, facilitating an increase in Q related to a modest rise in MAP. The control of these events involves a complex array of mechanical, humoral, and neural mechanisms [154].
Blood volume in vertebrates hardly exceeds 10% of body mass. It is supposed that all mechanisms controlling blood volume reflect steady-state conditions, depending on factors such as hydration, activity, hibernation, and other factors. The regulatory mechanisms of vertebrates are summarized below:
The elevation in efferent nervous activity can induce renal vasoconstriction, significantly decreasing renal blood flow in turn. Renal regulation may be particularly important in amphibians, some fish, and reptiles [155].
Endocrine factors, such as arginine vasopressin, the renin-angiotensin system, and atrial natriuretic peptide, play a role in fluid balance. The retention of fluid at the kidney level is promoted by arginine vasopressin (AVP), and its primary physiological significance lies in its indirect effects on blood pressure. AVP is found in the neurohypophysis of all mammalian vertebrates. The neurohypophyseal peptides in reptiles and birds closely resemble those found in mammals [156]. Except in cyclostomata and elasmobranchii, the renin-angiotensin-aldosterone system (RAAS) is present in all vertebrates. It stands out as one of the most crucial hormonal systems for the long-term modulation of renal function and hemodynamics in mammals [157,158]. The involvement of angiotensin II in regulating blood pressure in fish becomes evident when pressure declines following the administration of angiotensin-converting enzyme inhibitors. Finally, the atrial natriuretic factor comprises a family of peptides synthesized in the atrial myocytes in response to the tension in the local wall (increase in intra auricular volume), and it is present in all vertebrates [159].
Factors derived from the endothelium. Numerous peptides produce vasoactive effects and are present in all classes of vertebrates [160].
Other factors. Various molecules with autocrine or paracrine effects (such us histamine, bradykinin, adenosine, or nitric oxide) are present in the cells and affect the cardiovascular system of vertebrates [161].
As observed, an integrated response to changes in blood volume is controlled by all the afore-mentioned factors. The most prevalent natural disturbance in terrestrial environments, involving dehydration through evaporation, convection, and irradiation, as well as in marine environments marked by dehydration through osmotic imbalances, is the reduction in volumes of blood and body fluids in most vertebrates. Generally, mammals are less tolerant to loss of blood than other vertebrates such as fish [162], amphibians [163], reptiles [164,165] and birds [166,167], which can tolerate a greater degree of loss of plasma volume and mobilize compensatory fluid more quickly than mammals.
6. Integrated response to exercise in vertebrates
Metabolic demand in all vertebrates during exercise requires some cardiovascular system adjustments that are not quantitatively proportional. While VO2 can increase from ten to twentyfold, the changes in Q are smaller. The increased needs from muscle metabolism during exercise impose control by matching the delivery of VO2 and VO2max. This is due to two factors. First, the increase in oxygen extraction allows a greater supply of O2 to musculoskeletal muscles without a necessary increase in blood flow. Secondly, the supply of VO2 to tissues depends not only on the cardiovascular system but also on the respiratory apparatus [19,20].
Di Prampero [168] calculated the contribution of each element that contributes to an increase in VO2 to its maximum value. In humans, the central component (respiratory apparatus and cardiovascular system) is responsible for 75%, minimizing the participation of the peripheral route (mitochondrial oxidation and use of energetic stores). Ventricular size is a limiting factor for the central contribution as it determines SV, Q, and hence aerobic capacity and exercise performance. In mammals, the heart constitutes the same fraction of body mass according to the size of the mammals, from mice to cows [81]. In horses, the heart size ranges between 0.9 and 2% of the body mass in elite individuals, which is greater than in other species [3].
Thus, the cardiovascular system of vertebrates during exercise requires an increase in Q and a variation in TPR. In general, the adjustment of these variables differs among the five major groups of vertebrates. The way in which Q is augmented during exercise varies across vertebrates, with the majority increasing Q to a level between 1.3 and 3.3 times higher than resting values [169,170]. Exceptions to this pattern include trained human athletes and Thoroughbred horses. The contribution of each parameter that determines Q depends on the class of vertebrate [69].
While maximum HR is crucial in the increase of Q during exercise as an allometric function of body mass, SV may be an extremely important determinant in vertebrates with a certain body weight. Flying pigeons can increase HR sixfold without changing SV [169]. The majority of amphibians, reptiles, birds, and mammals depend to a large extent on tachycardia, although the range varies considerably [91] (see Table 1) as SV changes relatively little [171]. For example, in the rainbow trout, the considerable increase in EDV during exercise rises from about 0.4 ml/kg to 1.1 ml/kg, that is, almost threefold. In the leopard shark, the rise of the EDV during exercise is only 1.5 times, bearing in mind that it weighs 50 times more than the rainbow trout. In contrast to fish, humans increase EDV to a lesser degree, but ESV decreases [12]. However, training in mammals increases the role of SV. In mammals, the maximum HR in goats and dogs, ponies, calves, and horses is almost identical for animals of the same size, despite a difference of 1.4 to 2.5 times in their VO2max [172]. Tachycardia typically involves the withdrawal of vagal influence and, whenever feasible, adrenergic stimulation.
Central arterial pressure rises during exercise. Nevertheless, species with respiratory and systemic circulations in series, such as fish and mammals, generally exhibit slightly greater increases in systemic arterial pressure compared to species with two parallel circulations, such as amphibians and non-crocodilian reptiles [173]. The amplitude of the increase in arterial pressure is reduced because of the decrease in total vascular resistance. The most active mammals appear to have the ability to achieve a more significant reduction in vascular resistance. The overall outcome of the rise in Q and arterial pressure is an elevation in myocardial energy expenditure, which naturally necessitates a simultaneous increase in myocardial oxygen consumption. Another response involves directing blood flow preferentially to the actively engaged skeletal muscles. The regulation of these varied responses is not fully comprehended, particularly in the case of lower vertebrates. In mammals during exercise, the vasomotor center coordinates afferent information from active muscles (proprioceptors), arterial and cardiac mechanoreceptors, chemoreceptors, the cortex, and the hypothalamus to ensure appropriate autonomic output [12].
The change in the regional distribution of blood flow with exercise has been measured in several types of fish. In the rainbow trout and the big-mouthed trout, blood flow in red muscle constitutes 9% and 0.57% of cardiac output (Q) at rest, respectively. During exercise, these values increase to 42% and 13.2%, respectively [174,175]. Simultaneously, while blood flow to the skeletal muscles increases, it diminishes in the abdominal viscera [176]. The reduction in blood flow in the celiac, mesenteric, and intestinal arteries can be particularly significant, reaching approximately 30% in Atlantic cod [144] and 70% in Chinook salmon [177]. As systemic resistance diminishes, the impact of vasodilation in the skeletal muscles must surpass that of vasoconstriction in the visceral region. The regulation of these vascular responses remains poorly understood. The potential modulation of systemic alpha-adrenergic tone is considered, and the roles of locally produced metabolites [178], adrenergic and non-cholinergic fibers, particularly in the intestinal circulation, have yet to be thoroughly investigated [179].
Nevertheless, our knowledge regarding the regional distribution of Q during activity in amphibians and reptiles is extremely limited [180,181,182]. One of the scarce studies of this kind assessed the blood flows in the left pulmonary and aortic regions during both rest and swimming in the green sea turtle Chelonia mydas [183]. During exercise, it was hypothesized that Q would rise, primarily attributed to an increase in HR from 24 bpm to 40 bpm (at 28ºC). Both pulmonary and aortic blood flows showed significant increases during exercise, but there was limited evidence of a redistribution of Q, except for what is typically associated with intermittent pulmonary ventilation and breath-holding periods. Another more recent study shows that alligators, which during development have lower oxygen levels, also show long-term changes in cardiovascular response due to hypoxia during the embryonic period. Many of the changes were masked during exercise, probably due to differences in adrenergic activity. The authors conclude that it seems likely that changes in cardiovascular response have an evolutionary origin in vertebrates [86].
7. Limitations
One of the main methodological limitations in comparative studies of the cardiovascular response to physical exercise in vertebrates is the use of different models of locomotion. This leads to varying approaches for assessing the response to physical exercise. Whereas, treadmills have been used in mammals and reptiles, more sophisticated ergometers such as aerodynamic tunnels have been used in birds and aquatic tunnels in fish. It is important to note that this type of evaluation may not be applicable to all vertebrates [6,7]. For instance, in reptiles, the cardiovascular response to posture changes, made manually or by mechanical devices, has been studied. These posture changes that are achieved through the development of 'straightening reflexes' [38,42]. Additionally, evaluations of ectothermic animals include adjustments in their cardiovascular system in response to heat stress [184].
Other limitations include the study of vertebrates in their natural habitat, which poses methodological challenges, including the inability to control the timing of the highest demands on the cardiovascular system [6,7,19,20,69].
Therefore, it is evident that the limitations described pose significant obstacles to fully comprehending the cardiovascular response to exercise, which involves the threshold of demand among varying vertebrate groups.
8. Conclusions and Future Directions
In conclusion, the cardiovascular adjustments needed during exercise in all vertebrates, including changes in Q, TPR, and pressure gradient, are quantitatively smaller than the increase in oxygen consumption. Different types of circulation in vertebrates determine differences in cardiovascular response and regulation. The mechanism by which Q (either the SV or the HR) is enhanced during exertion varies significantly among vertebrates. The diverse physical exercise responses observed in arterial pressure and TPR can be attributed to the existing differences in the pulmonary and systemic circulation systems between vertebrates. "Athlete" vertebrates (e.g. horses) or those that have been genetically manipulated (e.g. rats with high exercise capacity) may represent models of maximum adaptation of the cardiovascular system. However, they do not reflect the typical response in vertebrates.
Exploring the examination of an integrated cardiovascular response and the elements that influence the diverse cardiovascular adaptations to physical exercise in varied vertebrate groups may be of significance for understanding the cardiovascular response of athletes in different conditions, including extreme temperatures, immersions, or high altitudes, as well as in individuals participating in physical exercise with different physical and functional disabilities. It is recommended to further research comparative cardiovascular physiology across vertebrate groups to comprehend previously described situations and develop mechanisms to support or improve the athletic population.
Supplementary Materials
Table S1. Example of resting and exercise heart rate in different classes of vertebrates. Figure S1. Right-left shunt and functionality in adult non-crocodilian reptiles. Figure S2. Left-right shunt and functionality in adult non-crocodilian reptiles. Figure S3. Importance of Frank-Starling's law in regulating stroke volume. Figure S4. Comparative cardiac cycle between trout and human.
Author Contributions
RGL contributed to the investigation, data curation, formal analysis, and writing—original draft; EJH contributed to the data curation, supervision, and writing—original draft; ALR contributed to funding acquisition, visualization, resources, and writing; JJR and JCM contributed to supervision, conceptualization, methodology, project administration, validation, and writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Universidad Alfonso X El Sabio.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hunter, P. The evolution of human endurance: Research on the biology of extreme endurance gives insights into its evolution in humans and animals. EMBO Rep 2019, 20, e49396. [Google Scholar] [CrossRef]
- Lavin, K.M.; Coen, P.M.; Baptista, L.C.; Bell, M.B.; Drummer, D.; Harper, S.A.; Lixandrao, M.E.; McAdam, J.S.; O'Bryan, S.M.; Ramos, S.; et al. State of Knowledge on Molecular Adaptations to Exercise in Humans: Historical Perspectives and Future Directions. Compr Physiol 2022, 12, 3193–3279. [Google Scholar] [CrossRef]
- Poole, D.C.; Erickson, H.H. Highly athletic terrestrial mammals: horses and dogs. Compr Physiol 2011, 1, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Lavie, C.J.; Arena, R.; Swift, D.L.; Johannsen, N.M.; Sui, X.; Lee, D.C.; Earnest, C.P.; Church, T.S.; O'Keefe, J.H.; Milani, R.V.; et al. Exercise and the cardiovascular system: clinical science and cardiovascular outcomes. Circ Res 2015, 117, 207–219. [Google Scholar] [CrossRef]
- Lavie, C.J.; Ozemek, C.; Carbone, S.; Katzmarzyk, P.T.; Blair, S.N. Sedentary Behavior, Exercise, and Cardiovascular Health. Circ Res 2019, 124, 799–815. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, M.S.; Hancock, T.V.; Hillman, S.S. Metabolism at the Max: How Vertebrate Organisms Respond to Physical Activity. Compr Physiol 2015, 5, 1677–1703. [Google Scholar] [CrossRef]
- Wang, T.; Joyce, W.; Hicks, J.W. Similitude in the cardiorespiratory responses to exercise across vertebrates. Curr Opin Physiol 2019, 10, 137–145. [Google Scholar] [CrossRef]
- Bowlin, M.S.; Bisson, I.A.; Shamoun-Baranes, J.; Reichard, J.D.; Sapir, N.; Marra, P.P.; Kunz, T.H.; Wilcove, D.S.; Hedenstrom, A.; Guglielmo, C.G.; et al. Grand challenges in migration biology. Integr Comp Biol 2010, 50, 261–279. [Google Scholar] [CrossRef] [PubMed]
- Piersma, T. Why marathon migrants get away with high metabolic ceilings: towards an ecology of physiological restraint. J Exp Biol 2011, 214, 295–302. [Google Scholar] [CrossRef]
- Morash, A.J.; Vanderveken, M.; McClelland, G.B. Muscle metabolic remodeling in response to endurance exercise in salmonids. Front Physiol 2014, 5, 452. [Google Scholar] [CrossRef]
- McWilliams, S.; Pierce, B.; Wittenzellner, A.; Langlois, L.; Engel, S.; Speakman, J.R.; Fatica, O.; DeMoranville, K.; Goymann, W.; Trost, L.; et al. The energy savings-oxidative cost trade-off for migratory birds during endurance flight. Elife 2020, 9. [Google Scholar] [CrossRef]
- Burggren, W.W.; Farrell, A.P.; Lillywhite, H.B. Vertebrate cardiovascular systems. In Handbook of Physiology. Comparative physiology; Am. Physiol.Soc.: Bethesda MD, 1997; Volume 1, pp. 215–308. [Google Scholar]
- Katano, W.; Moriyama, Y.; Takeuchi, J.K.; Koshiba-Takeuchi, K. Cardiac septation in heart development and evolution. Dev Growth Differ 2019, 61, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Crystal, G.J.; Pagel, P.S. The Physiology of Oxygen Transport by the Cardiovascular System: Evolution of Knowledge. J Cardiothorac Vasc Anesth 2020, 34, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Monahan-Earley, R.; Dvorak, A.M.; Aird, W.C. Evolutionary origins of the blood vascular system and endothelium. J Thromb Haemost 2013, 11 Suppl 1, 46–66. [Google Scholar] [CrossRef]
- Cowburn, A.S.; Macias, D.; Summers, C.; Chilvers, E.R.; Johnson, R.S. Cardiovascular adaptation to hypoxia and the role of peripheral resistance. Elife 2017, 6. [Google Scholar] [CrossRef]
- Joyce, W.; Wang, T. What determines systemic blood flow in vertebrates? J Exp Biol 2020, 223. [Google Scholar] [CrossRef] [PubMed]
- Aslanger, E. The Evolution of the Cardiovascular System: A Hemodynamic Perspective. Turk Kardiyol Dern Ars 2022, 50, 518–526. [Google Scholar] [CrossRef]
- Ramsook, A.H.; Dominelli, P.B.; Angus, S.A.; Senefeld, J.W.; Wiggins, C.C.; Joyner, M.J. The oxygen transport cascade and exercise: Lessons from comparative physiology. Comp Biochem Physiol A Mol Integr Physiol 2023, 282, 111442. [Google Scholar] [CrossRef] [PubMed]
- Hicks, J.W.; Wang, T. Arterial blood gases during maximum metabolic demands: Patterns across the vertebrate spectrum. Comp Biochem Physiol A Mol Integr Physiol 2021, 254, 110888. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, D.; Nourizadeh, S.; De Tomaso, A.W. The biology of the extracorporeal vasculature of Botryllus schlosseri. Dev Biol 2019, 448, 309–319. [Google Scholar] [CrossRef]
- Rodriguez, D.; Taketa, D.A.; Madhu, R.; Kassmer, S.; Loerke, D.; Valentine, M.T.; Tomaso, A.W. Vascular Aging in the Invertebrate Chordate, Botryllus schlosseri. Front Mol Biosci 2021, 8, 626827. [Google Scholar] [CrossRef]
- Lefevre, S.; Damsgaard, C.; Pascale, D.R.; Nilsson, G.E.; Stecyk, J.A. Air breathing in the Arctic: influence of temperature, hypoxia, activity and restricted air access on respiratory physiology of the Alaska blackfish Dallia pectoralis. J Exp Biol 2014, 217, 4387–4398. [Google Scholar] [CrossRef] [PubMed]
- Damsgaard, C.; Baliga, V.B.; Bates, E.; Burggren, W.; McKenzie, D.J.; Taylor, E.; Wright, P.A. Evolutionary and cardio-respiratory physiology of air-breathing and amphibious fishes. Acta Physiol (Oxf) 2020, 228, e13406. [Google Scholar] [CrossRef] [PubMed]
- Turko, A.J.; Rossi, G.S.; Wright, P.A. More than Breathing Air: Evolutionary Drivers and Physiological Implications of an Amphibious Lifestyle in Fishes. Physiology (Bethesda) 2021, 36, 307–314. [Google Scholar] [CrossRef]
- Hillman, S.S. Cardiac scope in amphibians: transition to terrestrial life. Canadian journal of zoology 1991, 69, 2010–2013. [Google Scholar] [CrossRef]
- Meyer, S.; Lauridsen, H.; Pedersen, K.; Andersson, S.A.; van Ooij, P.; Willems, T.; Berger, R.M.F.; Ebels, T.; Jensen, B. Opportunities and short-comings of the axolotl salamander heart as a model system of human single ventricle and excessive trabeculation. Sci Rep 2022, 12, 20491. [Google Scholar] [CrossRef]
- Schmidt-Ukaj, S.; Gumpenberger, M.; Posautz, A.; Strauss, V. The Amphibian Heart. Vet Clin North Am Exot Anim Pract 2022, 25, 367–382. [Google Scholar] [CrossRef]
- Lewis, Z.R.; Hanken, J. Convergent evolutionary reduction of atrial septation in lungless salamanders. J Anat 2017, 230, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Guangming, G.; Zhe, Y.; Mei, Z.; Chenchen, Z.; Jiawei, D.; Dongyu, Z. Comparative Morphology of the Lungs and Skin of two Anura, Pelophylax nigromaculatus and Bufo gargarizans. Sci Rep 2020, 10, 11420. [Google Scholar] [CrossRef]
- Hicks, J.W. The physiological and evolutionary significance of cardiovascular shunting patterns in reptiles. News Physiol Sci 2002, 17, 241–245. [Google Scholar] [CrossRef]
- Jensen, B.; Christoffels, V.M. Reptiles as a Model System to Study Heart Development. Cold Spring Harbor Perspectives in Biology 2020, 12, a037226. [Google Scholar] [CrossRef] [PubMed]
- Jensen, B.; Lauridsen, H.; Webb, G.J.W.; Wang, T. Anatomy of the heart of the leatherback turtle. J Anat 2022, 241, 535–544. [Google Scholar] [CrossRef]
- Jensen, B.; Moorman, A.F.; Wang, T. Structure and function of the hearts of lizards and snakes. Biol Rev Camb Philos Soc 2014, 89, 302–336. [Google Scholar] [CrossRef] [PubMed]
- Jensen, B.; Joyce, W.; Gregorovicova, M.; Sedmera, D.; Wang, T.; Christoffels, V.M. Low incidence of atrial septal defects in nonmammalian vertebrates. Evol Dev 2020, 22, 241–256. [Google Scholar] [CrossRef]
- Mitova, E.; Wittnich, C. Cardiac structures in marine animals provide insight on potential directions for interventions for pediatric congenital heart defects. Am J Physiol Heart Circ Physiol 2022, 322, H1–h7. [Google Scholar] [CrossRef] [PubMed]
- Rocha, L.I.Q.; Oliveira, M.; Dias, L.C.; Franco de Oliveira, M.; de Moura, C.E.B.; Magalhães, M.D.S. Heart morphology during the embryonic development of Podocnemis unifilis Trosquel 1948 (Testudines: Podocnemididae). Anat Rec (Hoboken) 2023, 306, 193–212. [Google Scholar] [CrossRef] [PubMed]
- Burggren, W.; Filogonio, R.; Wang, T. Cardiovascular shunting in vertebrates: a practical integration of competing hypotheses. Biol Rev Camb Philos Soc 2020, 95, 449–471. [Google Scholar] [CrossRef] [PubMed]
- Gregorovicova, M.; Bartos, M.; Jensen, B.; Janacek, J.; Minne, B.; Moravec, J.; Sedmera, D. Anguimorpha as a model group for studying the comparative heart morphology among Lepidosauria: Evolutionary window on the ventricular septation. Ecol Evol 2022, 12, e9476. [Google Scholar] [CrossRef]
- Jensen, B.; Van Den Berg, G.; Van Den Doel, R.; Oostra, R.-J.; Wang, T.; Moorman, A.F.M. Development of the Hearts of Lizards and Snakes and Perspectives to Cardiac Evolution. PLoS ONE 2013, 8, e63651. [Google Scholar] [CrossRef]
- Gleeson, T.T.; Mitchell, G.S.; Bennett, A.F. Cardiovascular responses to graded activity in the lizards Varanus and Iguana. Am J Physiol 1980, 239, R174–179. [Google Scholar] [CrossRef]
- Leite, C.A.C.; Taylor, E.W.; Wang, T.; Abe, A.S.; De Andrade, D.O.V. Ablation of the ability to control the right-to-left cardiac shunt does not affect oxygen consumption, specific dynamic action or growth in rattlesnakes,<i>Crotalus durissus</i>. Journal of Experimental Biology 2013, 216, 1881–1889. [Google Scholar] [CrossRef] [PubMed]
- Ishimatsu, A.; Hicks, J.W.; Heisler, N. Analysis of Cardiac Shunting in the Turtle <i>Trachemys</i> (<i>Pseudemys</i>) <i>Scripta</i>: Application of the Three Outflow Vessel Model. Journal of Experimental Biology 1996, 199, 2667–2677. [Google Scholar] [CrossRef] [PubMed]
- Boutilier, R.G.; Shelton, G. Gas exchange, storage and transport in voluntarily diving Xenopus laevis. J Exp Biol 1986, 126, 133–155. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Hedrick, M.S.; Ihmied, Y.M.; Taylor, E.W. Control and interaction of the cardiovascular and respiratory systems in anuran amphibians. Comp Biochem Physiol A Mol Integr Physiol 1999, 124, 393–406. [Google Scholar] [CrossRef]
- Burggren, W.W.; Johansen, K. Circulation and Respiration in Lungfishes (Dipnoi). J Morphol 1986, 217–236. [Google Scholar] [CrossRef]
- Graham, J.B.; Lee, H.J. Breathing air in air: In what ways might extant amphibious fish biology relate to prevailing concepts about early tetrapods, the evolution of vertebrate air breathing, and the vertebrate land transition? Physiol Biochem Zool 2004, 77, 720–731. [Google Scholar] [CrossRef] [PubMed]
- Randall, D.J. Cardio-respiratory modeling in fishes and the consequences of the evolution of airbreathing. Cardioscience 1994, 5, 167–171. [Google Scholar]
- Filogonio, R.; Wang, T.; Taylor, E.W.; Abe, A.S.; Leite, C.A. Vagal tone regulates cardiac shunts during activity and at low temperatures in the South American rattlesnake, Crotalus durissus. J Comp Physiol B 2016, 186, 1059–1066. [Google Scholar] [CrossRef]
- Kirby, A.R.; Smith, B.; Crossley, D.A. Regulation of blood flow in the pulmonary and systemic circuits during submerged swimming in common snapping turtle (Chelydra serpentina). J Exp Biol 2019, 222. [Google Scholar] [CrossRef]
- Corno, A.F.; Zhou, Z.; Uppu, S.C.; Huang, S.; Marino, B.; Milewicz, D.M.; Salazar, J.D. The Secrets of the Frogs Heart. Pediatr Cardiol 2022, 43, 1471–1480. [Google Scholar] [CrossRef]
- Nie, S. Use of Frogs as a Model to Study the Etiology of HLHS. J Cardiovasc Dev Dis 2023, 10. [Google Scholar] [CrossRef] [PubMed]
- Joyce, W.; White, D.W.; Raven, P.B.; Wang, T. Weighing the evidence for using vascular conductance, not resistance, in comparative cardiovascular physiology. J Exp Biol 2019, 222. [Google Scholar] [CrossRef] [PubMed]
- Cook, A.C.; Tran, V.H.; Spicer, D.E.; Rob, J.M.H.; Sridharan, S.; Taylor, A.; Anderson, R.H.; Jensen, B. Sequential segmental analysis of the crocodilian heart. J Anat 2017, 231, 484–499. [Google Scholar] [CrossRef] [PubMed]
- Poelmann, R.E.; Gittenberger-de Groot, A.C.; Goerdajal, C.; Grewal, N.; De Bakker, M.A.G.; Richardson, M.K. Ventricular Septation and Outflow Tract Development in Crocodilians Result in Two Aortas with Bicuspid Semilunar Valves. J Cardiovasc Dev Dis 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Barske, J.; Eghbali, M.; Kosarussavadi, S.; Choi, E.; Schlinger, B.A. The heart of an acrobatic bird. Comp Biochem Physiol A Mol Integr Physiol 2019, 228, 9–17. [Google Scholar] [CrossRef]
- Chang, Y.H.; Sheftel, B.I.; Jensen, B. Anatomy of the heart with the highest heart rate. J Anat 2022, 241, 173–190. [Google Scholar] [CrossRef] [PubMed]
- Jensen, B.; Wang, T.; Moorman, A.F.M. Evolution and Development of the Atrial Septum. Anat Rec (Hoboken) 2019, 302, 32–48. [Google Scholar] [CrossRef]
- Mori, S.; Tretter, J.T.; Spicer, D.E.; Bolender, D.L.; Anderson, R.H. What is the real cardiac anatomy? Clin Anat 2019, 32, 288–309. [Google Scholar] [CrossRef]
- Rowlatt, U. Comparative anatomy of the heart of mammals. Zoological Journal of the Linnean Society 1990, 98, 73–110. [Google Scholar] [CrossRef]
- Franziska Hein, R.; Kiefer, I.; Pees, M. A Spectral Computed Tomography Contrast Study: Demonstration of the Avian Cardiovascular Anatomy and Function. Vet Clin North Am Exot Anim Pract 2022, 25, 435–451. [Google Scholar] [CrossRef]
- Stephenson, A.; Adams, J.W.; Vaccarezza, M. The vertebrate heart: an evolutionary perspective. J Anat 2017, 231, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Lansford, R.; Rugonyi, S. Follow Me! A Tale of Avian Heart Development with Comparisons to Mammal Heart Development. Journal of Cardiovascular Development and Disease 2020, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.; Wheen, P.; Brandon, L.; Maree, A.; Kenny, R.A. Heart rate: control mechanisms, pathophysiology and assessment of the neurocardiac system in health and disease. Qjm 2022, 115, 806–812. [Google Scholar] [CrossRef] [PubMed]
- Joyce, W.; Pan, Y.K.; Garvey, K.; Saxena, V.; Perry, S.F. Regulation of heart rate following genetic deletion of the ß1 adrenergic receptor in larval zebrafish. Acta Physiol (Oxf) 2022, 235, e13849. [Google Scholar] [CrossRef]
- Joyce, W.; Wang, T. Regulation of heart rate in vertebrates during hypoxia: A comparative overview. Acta Physiol (Oxf) 2022, 234, e13779. [Google Scholar] [CrossRef] [PubMed]
- Stoyek, M.R.; MacDonald, E.A.; Mantifel, M.; Baillie, J.S.; Selig, B.M.; Croll, R.P.; Smith, F.M.; Quinn, T.A. Drivers of Sinoatrial Node Automaticity in Zebrafish: Comparison With Mechanisms of Mammalian Pacemaker Function. Front Physiol 2022, 13, 818122. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.W.; Wang, T.; Leite, C.A.C. An overview of the phylogeny of cardiorespiratory control in vertebrates with some reflections on the 'Polyvagal Theory'. Biol Psychol 2022, 172, 108382. [Google Scholar] [CrossRef]
- Joyce, W.; Wang, T. How cardiac output is regulated: August Krogh's proto-Guytonian understanding of the importance of venous return. Comp Biochem Physiol A Mol Integr Physiol 2021, 253, 110861. [Google Scholar] [CrossRef]
- Krogh, A. The Progress of Physiology. Science 1929, 70, 200–204. [Google Scholar] [CrossRef]
- Farrell, A.P. From Hagfish to Tuna: A Perspective on Cardiac Function in Fish. Physiological Zoology 1991, 64, 1137–1164. [Google Scholar] [CrossRef]
- Boukens, B.J.D.; Kristensen, D.L.; Filogonio, R.; Carreira, L.B.T.; Sartori, M.R.; Abe, A.S.; Currie, S.; Joyce, W.; Conner, J.; Opthof, T.; et al. The electrocardiogram of vertebrates: Evolutionary changes from ectothermy to endothermy. Prog Biophys Mol Biol 2019, 144, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Burkhard, S.; van Eif, V.; Garric, L.; Christoffels, V.M.; Bakkers, J. On the Evolution of the Cardiac Pacemaker. J Cardiovasc Dev Dis 2017, 4. [Google Scholar] [CrossRef] [PubMed]
- Lillywhite, H.B.; Zippel, K.C.; Farrell, A.P. Resting and maximal heart rates in ectothermic vertebrates. Comp Biochem Physiol A Mol Integr Physiol 1999, 124, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Parlin, A.F.; Schaeffer, P.J. Cardiovascular contributions and energetic costs of thermoregulation in ectothermic vertebrates. J Exp Biol 2022, 225. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, J.; Shiozawa, M.; Shiode, D. Heart rate and cardiac response to exercise during voluntary dives in captive sea turtles (Cheloniidae). Biol Open 2020, 9. [Google Scholar] [CrossRef]
- Farrell, A.P. Cardiac scope in lower vertebrates. Canadian Journal of Zoology 1991, 69, 1981–1984. [Google Scholar] [CrossRef]
- Lindner, A.; Esser, M.; López, R.; Boffi, F. Relationship between Resting and Recovery Heart Rate in Horses. Animals 2020, 10, 120. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.; van Mourik, R.A.; Martin, K.J.; Eijsvogels, T.M.H.; Longoria, K.A. 100 Long-Distance Triathlons in 100 Days: A Case Study on Ultraendurance, Biomarkers, and Physiological Outcomes. Int J Sports Physiol Perform 2023, 18, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, T.I.; Jeon, J.Y.; Lindsay, T.; Westgate, K.; Perez-Pozuelo, I.; Hollidge, S.; Wijndaele, K.; Rennie, K.; Forouhi, N.; Griffin, S.; et al. Resting heart rate is a population-level biomarker of cardiorespiratory fitness: The Fenland Study. PLoS One 2023, 18, e0285272. [Google Scholar] [CrossRef]
- Burggren, W.; Farrell, A.; Lillywhite, H. Vertebrate cardiovascular systems. Handbook of Physiology. Comprehensive physiology 2010, 215–308. [Google Scholar]
- Filogonio, R.; Dubansky, B.D.; Dubansky, B.H.; Leite, C.A.C.; Crossley, D.A. , 2nd. Prenatal hypoxia affects scaling of blood pressure and arterial wall mechanics in the common snapping turtle, Chelydra serpentina. Comp Biochem Physiol A Mol Integr Physiol 2021, 260, 111023. [Google Scholar] [CrossRef]
- Filogonio, R.; Dubansky, B.D.; Dubansky, B.H.; Wang, T.; Elsey, R.M.; Leite, C.A.C.; Crossley, D.A. , 2nd. Arterial wall thickening normalizes arterial wall tension with growth in American alligators, Alligator mississippiensis. J Comp Physiol B 2021, 191, 553–562. [Google Scholar] [CrossRef]
- Nyerges-Bohák, Z.; Nagy, K.; Rózsa, L.; Póti, P.; Kovács, L. Heart rate variability before and after 14 weeks of training in Thoroughbred horses and Standardbred trotters with different training experience. PLoS One 2021, 16, e0259933. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.; Stieglitz, J.D.; Cox, G.K.; Heuer, R.M.; Benetti, D.D.; Grosell, M.; Crossley, D.A. , 2nd. Cardio-respiratory function during exercise in the cobia, Rachycentron canadum: The impact of crude oil exposure. Comp Biochem Physiol C Toxicol Pharmacol 2017, 201, 58–65. [Google Scholar] [CrossRef]
- Joyce, W.; Miller, T.E.; Elsey, R.M.; Wang, T.; Crossley, D.A. , 2nd. The effects of embryonic hypoxic programming on cardiovascular function and autonomic regulation in the American alligator (Alligator mississippiensis) at rest and during swimming. J Comp Physiol B 2018, 188, 967–976. [Google Scholar] [CrossRef]
- Joyce, W.; Egginton, S.; Farrell, A.P.; Crockett, E.L.; O'Brien, K.M.; Axelsson, M. Exploring nature's natural knockouts: in vivo cardiorespiratory performance of Antarctic fishes during acute warming. J Exp Biol 2018, 221. [Google Scholar] [CrossRef]
- Alves, J.C.; Santos, A.M.; Jorge, P.I.; Lafuente, P. A protocol for the determination of the maximal lactate steady state in working dogs. J Sports Med Phys Fitness 2020, 60, 942–946. [Google Scholar] [CrossRef] [PubMed]
- Farinatti, P.; Monteiro, W.; Oliveira, R.; Crisafulli, A. Cardiorespiratory responses and myocardial function within incremental exercise in healthy unmedicated older vs. young men and women. Aging Clin Exp Res 2018, 30, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Hagberg, J.M.; Allen, W.K.; Seals, D.R.; Hurley, B.F.; Ehsani, A.A.; Holloszy, J.O. A hemodynamic comparison of young and older endurance athletes during exercise. J Appl Physiol (1985) 1985, 58, 2041–2046. [Google Scholar] [CrossRef]
- Farrell, A.P.; Davie, P.S.; Franklin, C.E.; Johansen, J.A.; Brill, R.W. Cardiac physiology in tunas. I. In vitro perfused heart preparations from yellowfin and skipjack tunas. Canadian Journal of Zoology 1992, 70, 1200–1210. [Google Scholar] [CrossRef]
- Wang, T.; Taylor, E.W.; Andrade, D.; Abe, A.S. Autonomic control of heart rate during forced activity and digestion in the snake Boa constrictor. J Exp Biol 2001, 204, 3553–3560. [Google Scholar] [CrossRef] [PubMed]
- Hornicke, H.; von Engelhardt, W.; Ehrlein, H.J. Effect of exercise on systemic blood pressure and heart rate in horses. Pflugers Arch 1977, 372, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Farrell, A.P. A review of cardiac performance in the teleost heart: intrinsic and humoral regulation. Canadian Journal of Zoology 1984, 62, 523–536. [Google Scholar] [CrossRef]
- Barske, J.; Fusani, L.; Wikelski, M.; Feng, N.Y.; Santos, M.; Schlinger, B.A. Energetics of the acrobatic courtship in male golden-collared manakins (Manacus vitellinus). Proc Biol Sci 2014, 281, 20132482. [Google Scholar] [CrossRef] [PubMed]
- Grubb, B.; Jorgensen, D.D.; Conner, M. Cardiovascular changes in the exercising emu. J Exp Biol 1983, 104, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Noakes, T.D. Lord of running; Oxford University Press: Cape Town, 1992. [Google Scholar]
- Physick-Sheard, P.W. Cardiovascular response to exercise and training in the horse. Vet Clin North Am Equine Pract 1985, 1, 383–417. [Google Scholar] [CrossRef] [PubMed]
- Navas de Solis, C. Cardiovascular Response to Exercise and Training, Exercise Testing in Horses. Vet Clin North Am Equine Pract 2019, 35, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Greco-Otto, P.; Bond, S.; Sides, R.; Bayly, W.; Leguillette, R. Conditioning equine athletes on water treadmills significantly improves peak oxygen consumption. Vet Rec 2020, 186, 250. [Google Scholar] [CrossRef] [PubMed]
- Poole, D.C.; Erickson, H.H. 31- Heart and vessels: Function during exercise and training adaptations. In Equine Sports Medicine and Surgery (Second Edition), Hinchcliff, K.W., Kaneps, A.J., Geor, R.J., Eds.; W.B. Saunders: 2014; pp. 667-694.
- Hillman, S.S.; Hancock, T.V.; Hedrick, M.S. A comparative meta-analysis of maximal aerobic metabolism of vertebrates: implications for respiratory and cardiovascular limits to gas exchange. J Comp Physiol B 2013, 183, 167–179. [Google Scholar] [CrossRef]
- Ask, J.A. Comparative aspects of adrenergic receptors in the hearts of lower vertebrates. Comp Biochem Physiol A Comp Physiol 1983, 76, 543–552. [Google Scholar] [CrossRef]
- Taylor, E.W.; Leite, C.A.; Sartori, M.R.; Wang, T.; Abe, A.S.; Crossley, D.A. , 2nd. The phylogeny and ontogeny of autonomic control of the heart and cardiorespiratory interactions in vertebrates. J Exp Biol 2014, 217, 690–703. [Google Scholar] [CrossRef] [PubMed]
- Mueller, P.J.; Clifford, P.S.; Crandall, C.G.; Smith, S.A.; Fadel, P.J. Integration of Central and Peripheral Regulation of the Circulation during Exercise: Acute and Chronic Adaptations. Compr Physiol 2017, 8, 103–151. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, P.S.; Hadaya, J.; Khalsa, S.S.; Yu, C.; Chang, R.; Shivkumar, K. The vagus nerve in cardiovascular physiology and pathophysiology: From evolutionary insights to clinical medicine. Semin Cell Dev Biol 2024, 156, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Filogonio, R.; Joyce, W. Chapter 1 - Evolution of the cardiovascular autonomic nervous system in vertebrates. In Primer on the Autonomic Nervous System (Fourth Edition), Biaggioni, I., Browning, K., Fink, G., Jordan, J., Low, P.A., Paton, J.F.R., Eds.; Academic Press: 2023; pp. 3-9.
- de Marneffe, M.; Jacobs, P.; Haardt, R.; Englert, M. Variations of normal sinus node function in relation to age: role of autonomic influence. Eur Heart J 1986, 7, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Sugano, S. Relation of intrinsic heart rate and autonomic nervous tone to resting heart rate in the young and the adult of various domestic animals. Nihon Juigaku Zasshi 1989, 51, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, M.A.; Weinberg, C.R.; Cook, D.; Best, J.D.; Reenan, A.; Halter, J.B. Differential changes of autonomic nervous system function with age in man. Am J Med 1983, 75, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Zavorsky, G.S. Evidence and possible mechanisms of altered maximum heart rate with endurance training and tapering. Sports Med 2000, 29, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Porges, S.W. The vagal paradox: A polyvagal solution. Compr Psychoneuroendocrinol 2023, 16, 100200. [Google Scholar] [CrossRef]
- Han, J.C.; Taberner, A.J.; Loiselle, D.S.; Tran, K. Cardiac efficiency and Starling's Law of the Heart. J Physiol 2022, 600, 4265–4285. [Google Scholar] [CrossRef]
- Sequeira, V.; van der Velden, J. Historical perspective on heart function: the Frank-Starling Law. Biophys Rev 2015, 7, 421–447. [Google Scholar] [CrossRef]
- Shiels, H.A.; White, E. The Frank-Starling mechanism in vertebrate cardiac myocytes. J Exp Biol 2008, 211, 2005–2013. [Google Scholar] [CrossRef]
- Farrell, A.P. Circulation of body fluids. In Environmental and metabolic animal physiology, Prosser, C.L., Ed.; Wiley/Liss: New York, 1991; pp. 509–558. [Google Scholar]
- Tello, K.; Naeije, R.; de Man, F.; Guazzi, M. Pathophysiology of the right ventricle in health and disease: an update. Cardiovasc Res 2023, 119, 1891–1904. [Google Scholar] [CrossRef] [PubMed]
- Furrer, R.; Hawley, J.A.; Handschin, C. The molecular athlete: exercise physiology from mechanisms to medals. Physiol Rev 2023, 103, 1693–1787. [Google Scholar] [CrossRef]
- Stephenson, R.B. Cardiovascular physiology. In Textbook of veterinary physiology, 4th ed.; Saunders/Elsevier St. Louis, Mo.: St. Louis, Mo, 2007. [Google Scholar]
- Thomas, D.P.; Fregin, G.F. Cardiorespiratory and metabolic responses to treadmill exercise in the horse. J Appl Physiol Respir Environ Exerc Physiol 1981, 50, 864–868. [Google Scholar] [CrossRef] [PubMed]
- Derman, K.D.; Noakes, T.D. Comparative Aspects of Exercise Physiology. In The athletic horse, Hodgson, D.R., Rose, R.J., Eds.; WB Saunders: Philadelphia, 1984; pp. 13–25. [Google Scholar]
- Mazan, M. Equine exercise physiology-challenges to the respiratory system. Anim Front 2022, 12, 15–24. [Google Scholar] [CrossRef]
- Joyce, W.; Elsey, R.M.; Wang, T.; Crossley, D.A. , 2nd. Maximum heart rate does not limit cardiac output at rest or during exercise in the American alligator ( Alligator mississippiensis). Am J Physiol Regul Integr Comp Physiol 2018, 315, R296–r302. [Google Scholar] [CrossRef]
- Farrell, A.P.; Jones, D.R. The Heart. In The Cardiovascular System, Hoar, W.S., Randall, D.J., Farrell, A.P., Eds.; Academic Press: San Diego, CA, 1992; pp. 1–88. [Google Scholar]
- Mazza, R.; Pasqua, T.; Gattuso, A. Cardiac heterometric response: the interplay between Catestatin and nitric oxide deciphered by the frog heart. Nitric Oxide 2012, 27, 40–49. [Google Scholar] [CrossRef]
- Reil, J.C.; Reil, G.H.; Kovács, Á.; Sequeira, V.; Waddingham, M.T.; Lodi, M.; Herwig, M.; Ghaderi, S.; Kreusser, M.M.; Papp, Z.; et al. CaMKII activity contributes to homeometric autoregulation of the heart: A novel mechanism for the Anrep effect. J Physiol 2020, 598, 3129–3153. [Google Scholar] [CrossRef]
- Ahmadian, M.; Williams, A.M.; Mannozzi, J.; Konecny, F.; Hoiland, R.L.; Wainman, L.; Erskine, E.; Duffy, J.; Manouchehri, N.; So, K.; et al. A cross-species validation of single-beat metrics of cardiac contractility. J Physiol 2022, 600, 4779–4806. [Google Scholar] [CrossRef] [PubMed]
- Muir, W.W.; Hamlin, R.L. Myocardial Contractility: Historical and Contemporary Considerations. Front Physiol 2020, 11, 222. [Google Scholar] [CrossRef] [PubMed]
- Vornanen, M. Regulation of contractility of the fish (Carassius carassius L.) heart ventricle. Comparative Biochemistry and Physiology Part C: Comparative Pharmacology 1989, 94, 477–483. [Google Scholar] [CrossRef]
- Driedzic, W.R.; Gesser, H. Ca2+ protection from the negative inotropic effect of contraction frequency on teleost hearts. Journal of Comparative Physiology B 1985, 156, 135–142. [Google Scholar] [CrossRef]
- Driedzic, W.R.; Gesser, H. Differences in force-frequency relationships and calcium dependency between elasmobranch and teleost hearts. Journal of Experimental Biology 1988, 140, 227–241. [Google Scholar] [CrossRef]
- Gesser, H.; Poupa, O. Acidosis and cardiac muscle contractility: comparative aspects. Comp Biochem Physiol A Comp Physiol 1983, 76, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Lo, M.; Shahriari, A.; Roa, J.N.; Tresguerres, M.; Farrell, A.P. Differential effects of bicarbonate on severe hypoxia- and hypercapnia-induced cardiac malfunctions in diverse fish species. J Comp Physiol B 2021, 191, 113–125. [Google Scholar] [CrossRef]
- Aguilar-Sanchez, Y.; Rodriguez de Yurre, A.; Argenziano, M.; Escobar, A.L.; Ramos-Franco, J. Transmural Autonomic Regulation of Cardiac Contractility at the Intact Heart Level. Front Physiol 2019, 10, 773. [Google Scholar] [CrossRef] [PubMed]
- López Alarcón, M.M.; Rodríguez de Yurre, A.; Felice, J.I.; Medei, E.; Escobar, A.L. Phase 1 repolarization rate defines Ca(2+) dynamics and contractility on intact mouse hearts. J Gen Physiol 2019, 151, 771–785. [Google Scholar] [CrossRef]
- Bazmi, M.; Escobar, A.L. Autonomic Regulation of the Goldfish Intact Heart. Front Physiol 2022, 13, 793305. [Google Scholar] [CrossRef]
- Kharin, S.; Shmakov, D. A comparative study of contractility of the heart ventricle in some ectothermic vertebrates. Acta Herpetologica, 2009, 4, 57–71. [Google Scholar] [CrossRef]
- Franklin, C.E.; Davie, P.S. Myocardial power output of an isolated eel (Anguilla dieffenbachii) heart preparation in response to adrenaline. Comparative Biochemistry and Physiology Part C: Comparative Pharmacology 1992, 101, 293–298. [Google Scholar] [CrossRef]
- Ma, K.G.L.; Gamperl, A.K.; Syme, D.A.; Weber, L.P.; Rodnick, K.J. Echocardiography and electrocardiography reveal differences in cardiac hemodynamics, electrical characteristics, and thermal sensitivity between northern pike, rainbow trout, and white sturgeon. J Exp Zool A Ecol Integr Physiol 2019, 331, 427–442. [Google Scholar] [CrossRef] [PubMed]
- Lai, N.C.; Shabetai, R.; Graham, J.B.; Hoit, B.D.; Sunnerhagen, K.S.; Bhargava, V. Cardiac function of the leopard shark, Triakis semifasciata. Journal of Comparative Physiology B 1990, 160, 259–268. [Google Scholar] [CrossRef]
- Johansen, K.; Burggren, W.W. Venous return and cardiac filling in varanid lizards. J. Exp. Biol 1984, 113, 389–399. [Google Scholar] [CrossRef]
- Brijs, J.; Sandblom, E.; Axelsson, M.; Sundell, K.; Sundh, H.; Kiessling, A.; Berg, C.; Grans, A. Remote physiological monitoring provides unique insights on the cardiovascular performance and stress responses of freely swimming rainbow trout in aquaculture. Sci Rep 2019, 9, 9090. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.R.; Fennell, W.H.; Young, J.B.; Palomo, A.R.; Quinones, M.A. Differential systemic arterial and venous actions and consequent cardiac effects of vasodilator drugs. Prog Cardiovasc Dis 1982, 24, 353–374. [Google Scholar] [CrossRef]
- Tota, B.; Acierno, R.; Agnisola, C. Mechanical Performance of the Isolated and Perfused Heart of the Haemoglobinless Antarctic Icefish Chionodraco hamatus (Lonnberg): Effects of Loading Conditions and Temperature. 1991. [CrossRef]
- Nishiyama, A.; Kitada, K.; Suzuki, M. Blood pressure adaptation in vertebrates: comparative biology. Kidney Int 2022, 102, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Burggren, W.W. Central Cardiovascular Function in Amphibians: Qualitative Influences of Phylogeny, Ontogeny, and Season. In Mechanisms of Systemic Regulation. Advances in Comparative and Environmental Physiology, Heisler, N., Ed.; Springer-Verlag: Berlin Heidelberg, 1995; Volume vol 21, pp. 175-197. [Google Scholar]
- Enok, S.; Slay, C.; Abe, A.S.; Hicks, J.W.; Wang, T. Intraspecific scaling of arterial blood pressure in the Burmese python. J Exp Biol 2014, 217, 2232–2234. [Google Scholar] [CrossRef]
- Goetz, R.H.; Warren, J.V.; Gauer, O.H.; Patterson, J.L., Jr.; Doyle, J.T.; Keen, E.N.; Mc, G.M. Circulation of the giraffe. Circ Res 1960, 8, 1049–1058. [Google Scholar] [CrossRef]
- Poulsen, C.B.; Wang, T.; Assersen, K.; Iversen, N.K.; Damkjaer, M. Does mean arterial blood pressure scale with body mass in mammals? Effects of measurement of blood pressure. Acta Physiol (Oxf) 2018, 222, e13010. [Google Scholar] [CrossRef]
- Bishop, C.M. Heart mass and the maximum cardiac output of birds and mammals: implications for estimating the maximum aerobic power input of flying animals. Philos Trans R Soc Lond B Biol Sci 1997, 29, 3447–3456. [Google Scholar] [CrossRef]
- Evans, D.L. The cardiovascular system: anatomy, physiology, and adaptations to exercise and training. In The athletic horse, Hodgson, D.R., Rose, R.J., Eds.; WB Saunders: Philadelphia, 1994; pp. 129–144. [Google Scholar]
- Hirsch, E.F. The innervation of the human heart. IV. (1) The fiber connections of the nerves with the perimysial plexus (Gerlach-Hofmann); (2) The role of nerve tissues in the repair of infarcts. Arch Pathol 1963, 75, 378–401. [Google Scholar] [CrossRef] [PubMed]
- Sturkie, P.D.; Poorvin, D.; Ossorio, N. Levels of epinephrine and norepinephrine in blood and tissues of duck, pigeon, turkey, and chicken. Proc Soc Exp Biol Med 1970, 135, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Segal, S.S. Integration of blood flow control to skeletal muscle: key role of feed arteries. Acta Physiol Scand 2000, 168, 511–518. [Google Scholar] [CrossRef]
- Dantzler, W.H. Comparative Physiology of the Vertebrate Kidney; Springer-Verlag: New York, 2016; p. 292. [Google Scholar]
- Pang, P.K.; Furspan, P.B.; Sawyer, W.H. Evolution of neurohypophyseal hormone actions in vertebrates. American Zoologist 1983, 23, 655–662. [Google Scholar] [CrossRef]
- Fournier, D.; Luft, F.C.; Bader, M.; Ganten, D.; Andrade-Navarro, M.A. Emergence and evolution of the renin-angiotensin-aldosterone system. J Mol Med (Berl) 2012, 90, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, H. Renin-angiotensin system in vertebrates: phylogenetic view of structure and function. Anat Sci Int 2017, 92, 215–247. [Google Scholar] [CrossRef]
- Cerra, M.C. Cardiac distribution of the binding sites for natriuretic peptides in vertebrates. Cardioscience 1994, 5, 215–224. [Google Scholar] [PubMed]
- Kipryushina, Y.O.; Yakovlev, K.V.; Odintsova, N.A. Vascular endothelial growth factors: A comparison between invertebrates and vertebrates. Cytokine Growth Factor Rev 2015, 26, 687–695. [Google Scholar] [CrossRef]
- Donald, J.A.; Forgan, L.G.; Cameron, M.S. The evolution of nitric oxide signalling in vertebrate blood vessels. J Comp Physiol B 2015, 185, 153–171. [Google Scholar] [CrossRef]
- Duff, D.W.; Olson, K.R. Response of rainbow trout to constant-pressure and constant-volume hemorrhage. Am J Physiol 1989, 257, R1307–1314. [Google Scholar] [CrossRef]
- Hillman, S.S.; Withers, P.C. The hemodynamic consequences of hemorrhage and hypernatremia in two amphibians. J Comp Physiol B 1988, 157, 807–812. [Google Scholar] [CrossRef]
- Smits, A.W.; Kozubowski, M.M. Partitioning of body fluids and cardiovascular responses to circulatory hypovolaemia in the turtle, Pseudemys scripta elegans. Journal of experimental biology 1985, 116, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Smits, A.W.; Lillywhite, H.B. Maintenance of blood volume in snakes: transcapillary shifts of extravascular fluids during acute hemorrhage. Journal of Comparative Physiology B 1985, 155, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Djojosugito, A.M.; Folkow, B.; Kovach, A.G. The mechanisms behind the rapid blood volume restoration after hemorrhage in birds. Acta Physiol Scand 1968, 74, 114–122. [Google Scholar] [CrossRef]
- Schindler, S.L.; Gildersleeve, R.P. Comparison of recovery from hemorrhage in birds and mammals. Comp Biochem Physiol A Comp Physiol 1987, 87, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Di Prampero, P.E. Metabolic and circulatory limitations to VO2 max at the whole animal level. Journal of Experimental Biology 1985, 115, 319–331. [Google Scholar] [CrossRef]
- Butler, P.J.; West, N.H.; Jones, D.R. Respiratory and cardiovascular responses of the pigeon to sustained, level flight in a wind-tunnel. Journal of Experimental Biology 1977, 71, 7–26. [Google Scholar] [CrossRef]
- Korner, P.I. Integrative neural cardiovascular control. Physiol Rev 1971, 51, 312–367. [Google Scholar] [CrossRef]
- Axelsson, M.; Davison, W.; Forster, M.E.; Farrell, A.P. Cardiovascular responses of the red-blooded Antarctic fishes Pagothenia bernacchii and P. borchgrevinki. Journal of Experimental Biology 1992, 167, 179–201. [Google Scholar] [CrossRef]
- Jones, J.H.; Longworth, K.E.; Lindholm, A.; Conley, K.E.; Karas, R.H.; Kayar, S.R.; Taylor, C.R. Oxygen transport during exercise in large mammals. I. Adaptive variation in oxygen demand. J Appl Physiol (1985) 1989, 67, 862–870. [Google Scholar] [CrossRef]
- Brill, R.W.; Bushnell, P.G. Metabolic and cardiac scope of high energy demand teleosts, the tunas. Canadian Journal of Zoology 1991, 69, 2002–2009. [Google Scholar] [CrossRef]
- Kolok, A.S.; Spooner, M.R.; Farrell, A.P. The effect of exercise on the cardiac output and blood flow distribution of the largescale sucker Catostomus macrocheilus. Journal of Experimental Biology 1993, 183, 301–321. [Google Scholar] [CrossRef]
- Randall, D.J.; Daxboeck, C. Cardiovascular changes in the rainbow trout (Salmo gairdneri Richardson) during exercise. Canadian Journal of Zoology 1982, 60, 1135–1140. [Google Scholar] [CrossRef]
- Thorarensen, H.; McLean, E.; Donaldson, E.M.; Farrell, A.P. The blood vasculature of the gastrointestinal tract in chinook, Oncorhynchus tshawytscha (Walbaum), and coho, O. kisutch (Walbaum), salmon. Journal of fish biology 1991, 38, 525–531. [Google Scholar] [CrossRef]
- Axelsson, M.; Fritsche, R. Effects of exercise, hypoxia and feeding on the gastrointestinal blood flow in the Atlantic cod Gadus morhua. Journal of Experimental Biology 1991, 158, 181–198. [Google Scholar] [CrossRef] [PubMed]
- Canty, A.A.; Farrell, A.P. Intrinsic regulation of flow in an isolated tail preparation of the ocean pout (Macrozoarces americanus). Canadian journal of zoology, 1985, 63, 2013–2020. [Google Scholar] [CrossRef]
- Kimmel, P.B. Adrenergic receptors and the regulation of vascular resistance in bullfrog tadpoles (Rana catesbeiana). J Comp Physiol B 1992, 162, 455–462. [Google Scholar] [CrossRef]
- Butler, P.J.; Milsom, W.K.; Woakes, A.J. Respiratory, cardiovascular and metabolic adjustments during steady state swimming in the green turtle,Chelonia mydas. Journal of Comparative Physiology B 1984, 154, 167–174. [Google Scholar] [CrossRef]
- Hillman, S.S.; Withers, P.C.; Palioca, W.B.; Ruben, J.A. Cardiovascular consequences of hypercalcemia during activity in two species of amphibian. Journal of Experimental Zoology 1987, 242, 303–308. [Google Scholar] [CrossRef]
- Withers, P.C.; Hillman, S.S.; Simmons, L.A.; Zygmunt, A.C. Cardiovascular adjustments to enforced activity in the anuran amphibian, Bufo marinus. Comparative Biochemistry and Physiology Part A: Physiology 1988, 89, 45–49. [Google Scholar] [CrossRef]
- West, N.H.; Butler, P.J.; Bevan, R.M. Pulmonary blood flow at rest and during swimming in the green turtle, Chelonia mydas. Physiological zoology 1992, 65, 287–310. [Google Scholar] [CrossRef]
- Filatova, T.S.; Abramochkin, D.V.; Shiels, H.A. Thermal acclimation and seasonal acclimatization: a comparative study of cardiac response to prolonged temperature change in shorthorn sculpin. J Exp Biol 2019, 222. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Right-left shunt and functionality in adult non-crocodilian reptiles. A right-to-left shunt occurs when oxygen-poor venous blood (right atrium) is diverted to the systemic circulation without being reoxygenated in the lungs. Cardiovascular shunts should be viewed as part of integrated systems with multiple functions. High vagal tone (typical of rest) promotes R-L shunting, whereas adrenaline reduces the R-L shunt. The reductions in R-L provide for an additional means to elevate systemic oxygen delivery during exercise. Colours: red for oxygenated blood, blue for deoxygenated blood, purple for mixed blood. Data: from different authors [31,32,38,41,42,43]
Figure 1.
Right-left shunt and functionality in adult non-crocodilian reptiles. A right-to-left shunt occurs when oxygen-poor venous blood (right atrium) is diverted to the systemic circulation without being reoxygenated in the lungs. Cardiovascular shunts should be viewed as part of integrated systems with multiple functions. High vagal tone (typical of rest) promotes R-L shunting, whereas adrenaline reduces the R-L shunt. The reductions in R-L provide for an additional means to elevate systemic oxygen delivery during exercise. Colours: red for oxygenated blood, blue for deoxygenated blood, purple for mixed blood. Data: from different authors [31,32,38,41,42,43]
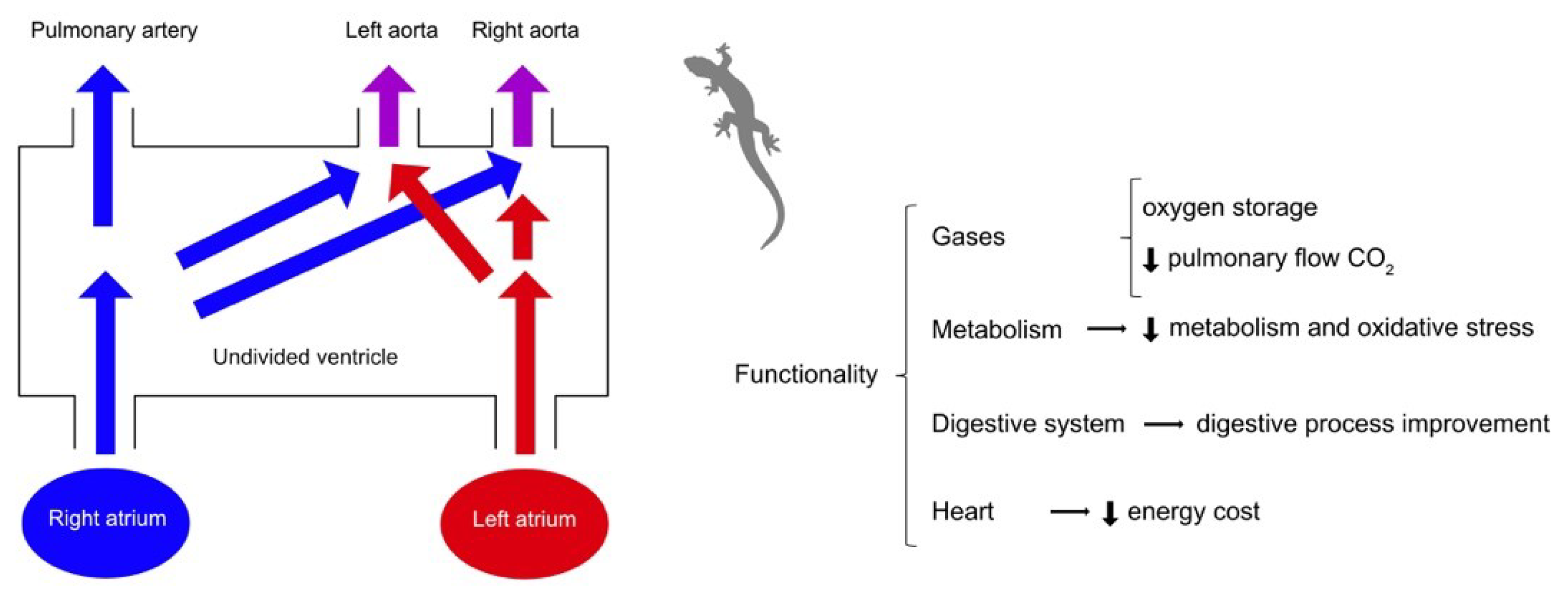
Figure 2.
Left to right shunt and functionality in adult non-crocodilian reptiles. Left-to-right shunts occur when oxygenated blood (left atrium) is directly recirculated to the lungs and represent the recirculation into the pulmonary arterial circulation. Cardiovascular shunts are adaptions with multiple functions, selected for because of their favourable cost/benefit ratio. The increase in adrenaline during exercise may invoke a left-to-right shunt and a recirculation of pulmonary venous blood back into the lung. Colours: red for oxygenated blood, blue for deoxygenated blood, purple for mixed blood. Data: from different authors [31,32,38,41,42,43].
Figure 2.
Left to right shunt and functionality in adult non-crocodilian reptiles. Left-to-right shunts occur when oxygenated blood (left atrium) is directly recirculated to the lungs and represent the recirculation into the pulmonary arterial circulation. Cardiovascular shunts are adaptions with multiple functions, selected for because of their favourable cost/benefit ratio. The increase in adrenaline during exercise may invoke a left-to-right shunt and a recirculation of pulmonary venous blood back into the lung. Colours: red for oxygenated blood, blue for deoxygenated blood, purple for mixed blood. Data: from different authors [31,32,38,41,42,43].
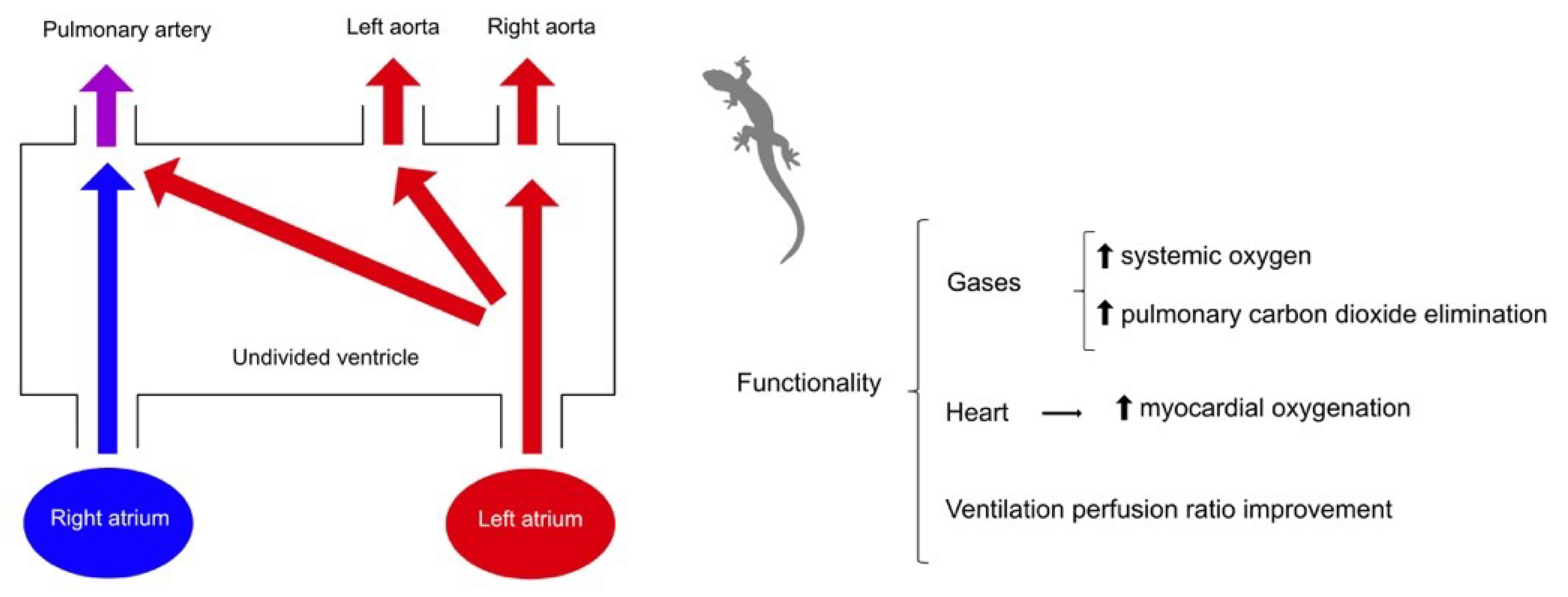
Figure 3.
Importance of Frank-Starling's law in regulating stroke volume. An elevation in cardiac filling pressure leads to an augmented stroke volume through the Frank-Starling mechanism. Exercise is an extrinsic mechanism of cardiac stimulation that increase cardiac filling pressure and stroke volume. Tuna has less chance of improving ejection through the Frank-Starling mechanism since, at zero ventricular pressure, it reaches maximum stroke volume, unlike trout and sea raven, which maintain a lower stroke volume at higher cardiac pressures. Abbreviations: ml-milliliter, kg-kilogram, mmHg-millimeters of mercury. Data: from various authors [7,12,91,94,115,116].
Figure 3.
Importance of Frank-Starling's law in regulating stroke volume. An elevation in cardiac filling pressure leads to an augmented stroke volume through the Frank-Starling mechanism. Exercise is an extrinsic mechanism of cardiac stimulation that increase cardiac filling pressure and stroke volume. Tuna has less chance of improving ejection through the Frank-Starling mechanism since, at zero ventricular pressure, it reaches maximum stroke volume, unlike trout and sea raven, which maintain a lower stroke volume at higher cardiac pressures. Abbreviations: ml-milliliter, kg-kilogram, mmHg-millimeters of mercury. Data: from various authors [7,12,91,94,115,116].
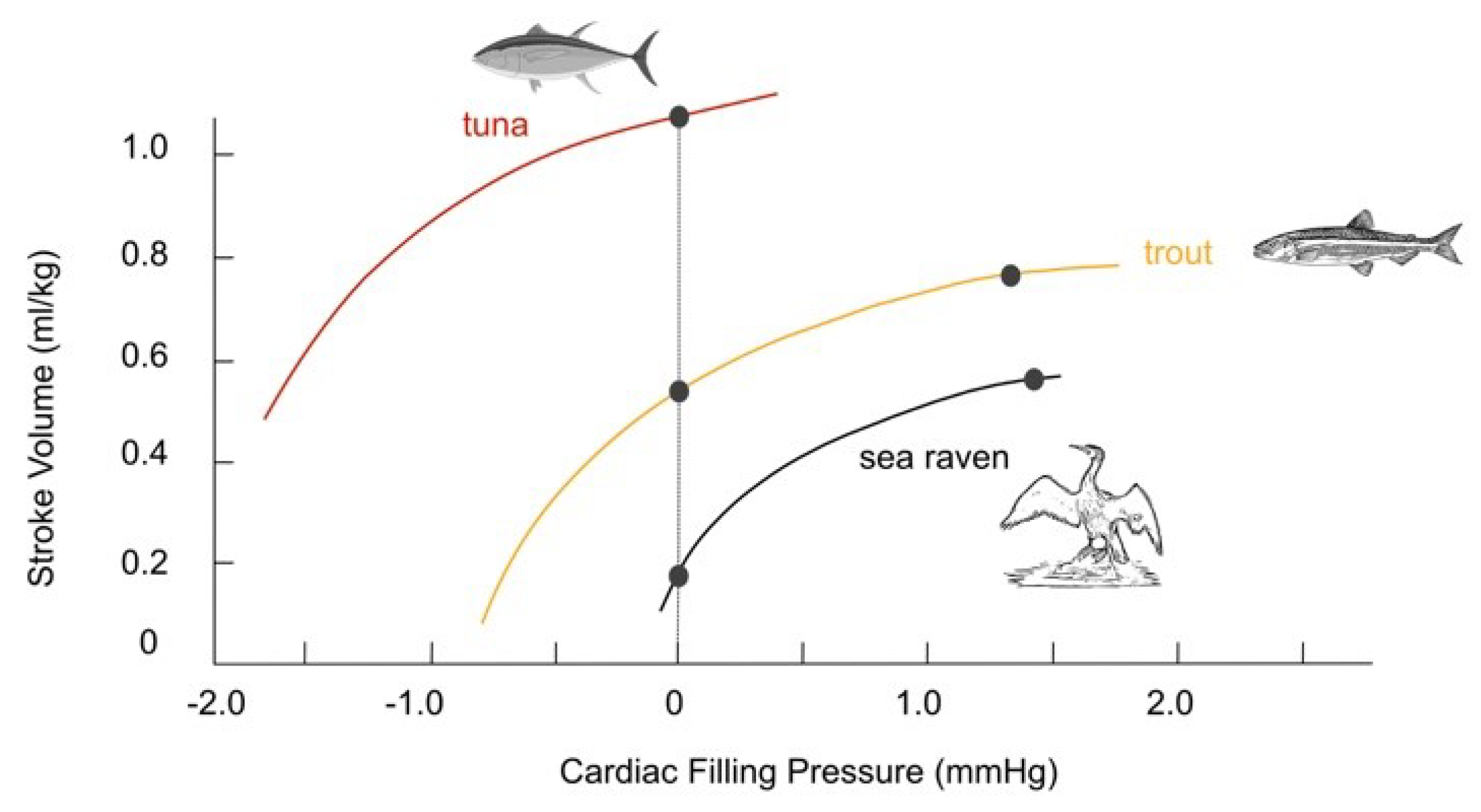
Figure 4.
Comparative cardiac cycle between trout and human. The ventricular pressure-volume relationship can be described by a loop diagram with volume depicted on the x-axis and left ventricular pressure on the y-axis. Rainbow trout increase their stroke volume by increasing end diastolic volume, whereas human do so at the expense of both increasing end diastolic volume and reducing end systolic volume. Abbreviations: SV-stroke volume, EDV-end diastolic volume, ESV-end systolic volume, ml-milliliter, kg- kilogram, mmHg-millimeters of mercury. Data: from various authors [6,7,12,71,91,94,116,124].
Figure 4.
Comparative cardiac cycle between trout and human. The ventricular pressure-volume relationship can be described by a loop diagram with volume depicted on the x-axis and left ventricular pressure on the y-axis. Rainbow trout increase their stroke volume by increasing end diastolic volume, whereas human do so at the expense of both increasing end diastolic volume and reducing end systolic volume. Abbreviations: SV-stroke volume, EDV-end diastolic volume, ESV-end systolic volume, ml-milliliter, kg- kilogram, mmHg-millimeters of mercury. Data: from various authors [6,7,12,71,91,94,116,124].
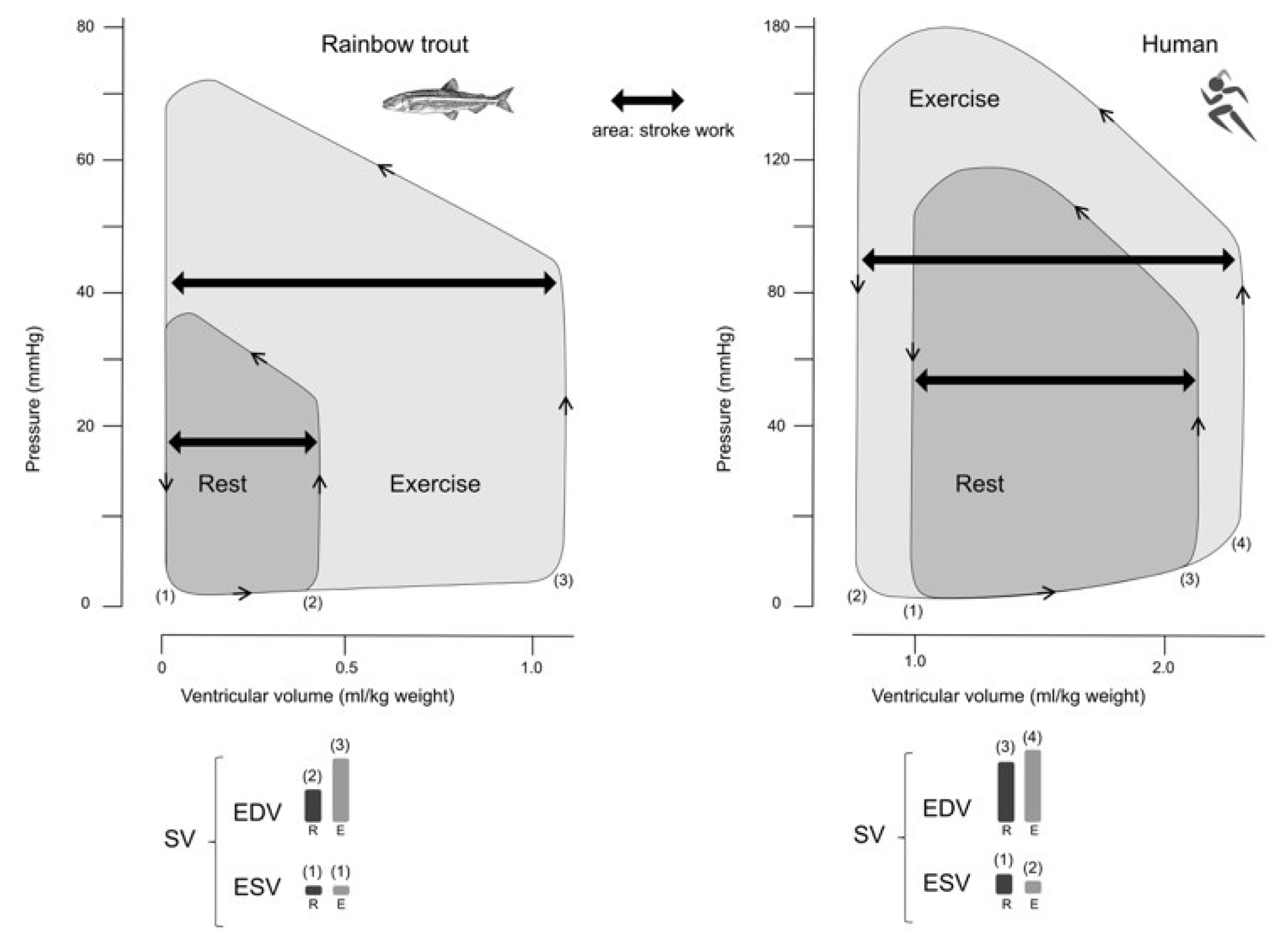
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



