
Submitted:
04 July 2023
Posted:
07 July 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
In post-COVID-19 syndrome, clinical presentation of the nerve fiber dysfunction plays an important role. The possibility of autoantigen cross-mimicry of coronavirus infection and the peripheral nervous system need to be investigated. The bioinformatic analysis was applied to search for possible common protein sequences located in the immunological epitopes. Among the autoantigens of the human nervous system, fibroblast growth factor receptor protein 3, myelin protein P0, myelin protein P2, sodium channel protein type 9, alpha protein subunit, plexin-D1 protein and ubiquitin-carboxyl-terminal hydrolase protein of the L1 isoenzyme were selected. The “Alignmentaj” program was created. The UniProt database, Protein Data Bank and AlphaFold databases were used. The analysis of protein sequence similarities of spike glycoproteins in human coronaviruses revealed common pentapeptides of the MERS-CoV-2 virus with the fibroblast growth factor receptor 3 and myelin protein P2. Among seasonal coronaviruses, common peptide sequences were identified in HCoV-HKU-1 virus with sodium channel protein type 9 subunit alpha and Plexin-D1, HCoV-OС43 with Plexin-D1, as well as HCoV-NL63 with Plexin-D1 and Ubiquitin carboxyl-terminal hydrolase isozyme L1. The data obtained make it possible to identify new potential targets for the development of autoimmune reactions that occur against the background of the activity of highly pathogenic and seasonal coronaviruses.
Keywords:
molecular mimicry
; post-COVID-19 syndrome
; SARS-CoV-2
; human coronaviruses
; autoimmunity
; autoantigens of nervous system
1. Introduction
Coronaviridae is a large family of RNA–containing pathogens infecting humans and other mammals [1]. Their virions have a complex immunologic composition, including three antigenically distinct subunits. Until the beginning of the 21st century, four seasonal coronaviruses (HCoV-229E, HCoV-OC43, HCoV-NL63 and HCoV-HKU-1) were described in the human population. [1]. In 2002 severe acute respiratory syndrome (caused by SARS-CoV-1), as well as in 2012 - Middle East respiratory syndrome (MERS, caused by MERS-CoV virus) were described, hence first couple of highly pathogenic members of the family appeared in the scope of medical attention, causing life-treating pneumonia and terminal respiratory failure.
In December 2019, the previously unknown coronavirus SARS-CoV-2 was evaluated [2], causing a pandemic of the novel highly pathogenic respiratory infection [3].
Novel coronavirus infection and autoimmunity
The respiratory system is the one to be the most impacted by the SARS-CoV-2 virus, which results in pulmonary complications such as alveolar damage, fibrosis in the interstitial tissue, and infiltration of the lung [4]. The ACE2 receptors and other molecules that allow the virus to enter cells are described in various human tissues, leading to a wide range of complications. The ACE2 enzyme can also exist in a soluble form, which may facilitate the binding of SARS-CoV-2 spike proteins and the formation of neoantigens [5].
SARS-CoV-2 is notorious for inducing an exaggerated response from the innate immune system and the production of pro-inflammatory substances, causing breaches in localized inflammation barriers and leading to the development of a cytokine storm and autoimmune complications [4,6]. A molecular mimicry can be the possible explanation for this hyperactive immune response, where similar sequences between foreign and self peptides can act as a trigger [7,8]. A study by Kanduc et al. identified a high degree of similarity between SARS-CoV-2 spike glycoprotein and human surfactant-related proteins, suggesting that this mechanism could explain the immunopathological processes in the lungs associated with coronavirus infection. Further investigation revealed extensive overlap between the spike glycoprotein of SARS-CoV-2 and the proteomes of humans and mice, both of which are known to experience prolonged health issues after acute SARS-CoV-2 infection [9]. Supporting this hypothesis, Kanduc identified more than 400 specific human peptides that match immunoreactive viral epitopes [10].
Small fiber neuropathy and the post-COVID-19 condition
Post-COVID syndrome is caused by a combination of immunologic, cardiovascular, and neurological issues. The dysfunction in both the central and peripheral nervous systems in these patients is evaluated [11]. While there have been cases of encephalitis or encephalomyelitis linked to the viral infection [12], it is still unclear, whether the coronavirus persists in the central nervous system without destructing the blood-brain barrier [13,14,15].
Autoimmune damage to the peripheral nervous system is also important in the post-COVID-19 complications development. A series of cases of Guillain-Barré syndrome and necrotizing autoimmune myositis as the post-viral sequel were described [11]. However, a less studied type of peripheral nervous system damage, known as small fiber neuropathy (SFN), can lead to numerous complications. SFN occurs when the smallest nerve fibers (A-delta and C-types) that penetrate almost all organs and tissues are damaged due to immunological inflammation. Standard electroneuromyography may not detect these changes, permitting the SFN to progress unrecognized.
The widespread presence of these nerve fibers in the body contributes to a diverse range of symptoms. Common manifestations include sensory dysfunction and dysautonomia, affecting the cardiovascular, urinary, gastrointestinal, skin, and mucous membrane systems. Due to the varying symptoms, the diagnosis of SFN can be challenging and may require comprehensive evaluation by specialists, such as neurologists. Given the importance of dysautonomia and polyneuropathy in the post-COVID-19 clinical evaluation, the presence of molecular mimicry between the antigens of small nerve fibers and different coronaviruses may lead to the new insights in this field. Moreover, our evaluation of small nerve fibers by confocal microscopy of the cornea shows the correctness of this approach (Figure 1).
The purpose of this article is to consider the possibility of implementing the phenomenon of molecular mimicry between autoantigens of the human nervous system and immunoreactive epitopes of human coronaviruses, which causes the emergence and development of immunological complications.
2. Materials and Methods
Human seasonal and highly pathogenic Coronaviruses SARS-CoV-2, SARS-CoV, MERS-CoV, HCoV-HKU-1, HCoV-OC43, HCoV-NL63, HCoV-229E were selected as the object of the study, in which spike proteins (S), membrane proteins (M) and nucleocapsid proteins (N) were evaluated. Among the autoantigens of small nerve fibers of the human peripheral nervous system, we selected the following ones mentioned in literature as potential targets in autoimmune neuropathies and candidates for mechanistic role in SFN: fibroblast growth factor receptor protein 3, myelin protein P0, myelin protein P2, sodium channel protein type 9 subunit alpha, plexin-D1 protein and ubiquitin-carboxyl-terminal hydrolase protein of the L1 isoenzyme. Amino acid sequences of proteins were obtained from the UniProt database [16] (https://www.uniprot.org/). The ID numbers of the amino acid sequences of these proteins from the UniProt database are shown in Table 1. Our original program “Alignmentaj” [https://github.com/muslimb/MyProekt1] (Registered by Russian Federal Agency for Intellectual Property: Certificate # 2023617186 of 6 April, 2023.) was created to conduct bioinformatic analysis. The primary sequences of S, M, N proteins were divided into pentapeptides shifted by one residue (i.e. MFVFL, FVFLV, VFLVL, FLVLL, etc.), after which the obtained viral pentapeptides were analyzed for the presence in human autoantigens. To show the location of similar pentapeptides in 3D structures of human coronavirus antigens and autoantigens of the nervous system, Protein Data Bank [17] (PDB, https://www.rcsb.org/) and AlphaFold [18] (https://alphafold.ebi.ac.uk/) databases were used, and the PyMol v.2.5.4 [19] (https://pymol.org/2/) program was used for structural visualization of pentapeptides.
To obtain information on the immunoreactivity of epitopes containing pentapeptides in human coronavirus antigens and human autoantigens, the IEDB database was used [20].
3. Results
The results obtained are presented in Table 1, Table 2 and Table 3 and clearly reflected in Figure 2, Figure 3 and Figure 4.
In addition, Table 1, Table 2 and Table 3 represent the number of immunoreactive epitopes containing pentapeptides in human coronavirus antigens and human autoantigens, where the IEDB database was used.
All ID numbers of immunoreactive epitopes in the IEDB database containing shared pentapeptides between human autoantigens and human coronaviruses were posted at this link (https://github.com/muslimb/Epitope).
Analysis of the data presented in Table 1 allows us to conclude that:
1. A Shared pentapeptide NLREF was found for Fibroblast growth factor receptor 3 and for 26 immunoreactive epitopes of the SARS-CoV-2 spike glycoprotein. This shared pentapeptide is located outside the immunoreactive epitopes of Fibroblast growth factor receptor 3.
2. A Shared pentapeptide NLNES was found for Plexin-D1 and for 32 immunoreactive epitopes of the SARS-CoV-2 spike glycoprotein. This Shared pentapeptide is located outside the immunoreactive epitopes of Plexin D1.
3. A Shared pentapeptide NLNES was found for Plexin-D1 and for two immunoreactive epitopes of the SARS-CoV spike glycoprotein. This Shared pentapeptide is located outside the immunoreactive epitopes of Plexin D1.
4. Shared pentapeptides ALALC, LALCV, LDVLE were discovered for Fibroblast growth factor receptor 3 and spike glycoprotein of the MERS-CoV. Shared pentapeptides ALALC, LALCV are located outside the immunoreactive epitopes in both spike glycoprotein of the MERS-CoV and Fibroblast growth factor receptor 3. The Shared pentapeptide LDVLE is located within the 14 of immunoreactive epitopes of the MERS-CoV spike glycoprotein and within immunoreactive epitope of Fibroblast growth factor receptor 3.
5. A shared pentapeptide EYSLY was discovered for myelin protein P2 and immunoreactive epitope of the MERS-CoV spike glycoprotein. The shared pentapeptide EYSLY is located within immunoreactive epitope of myelin protein P2.
6. Shared pentapeptides KLSDV, IILFT, ILFTG, KLSDF were discovered for sodium channel protein type 9 subunit alpha and HCoV-HKU-1 spike glycoprotein. Shared pentapeptides KLSDV and ILFTG are located within immunoreactive epitopes of HCoV-HKU-1 spike glycoprotein and outside the immunoreactive epitopes of sodium channel protein type 9 subunit alpha. The common pentapeptide IILFT is located outside the immunoreactive epitopes in both spike glycoprotein of the HCoV-HKU-1 and sodium channel protein type 9 subunit alpha. The common pentapeptide KLSDF is located within the 15 of immunoreactive epitopes of the HCoV-HKU-1 spike glycoprotein and within one immunoreactive epitope of sodium channel protein type 9 subunit alpha.
7. Common pentapeptides YVLPL, SDLQL, DLQLG were discovered for autoantigens of Plexin-D1 and immunoreactive epitopes of HCoV-HKU-1 spike glycoprotein. The common pentapeptide YVLPL is located within one immunoreactive epitope of HCoV-HKU-1 spike glycoprotein and within 7 immunoreactive epitopes of Plexin-D1. Common pentapeptides SDLQL and DLQLG are located within immunoreactive epitopes of HCoV-HKU-1 spike glycoprotein, but outside immunoreactive epitopes of Plexin-D1.
8. A Shared pentapeptide KLSDV was discovered for autoantigen of sodium channel protein type 9 subunit alpha and immunoreactive epitopes of HCoV-ОС43 spike glycoprotein. A Shared pentapeptide is located within 10 immunoreactive epitopes of HCoV-ОС43 spike glycoprotein, but outside of immunoreactive epitopes of sodium channel protein type 9 subunit alpha.
9. Shared pentapeptides AEAEA, GTCPA, DLQLG were discovered for autoantigen Plexin-D1 and immunoreactive epitopes of HCoV-ОС43 spike glycoprotein. Shared pentapeptides AEAEA, GTCPA are located within immunoreactive epitopes of both HCoV-ОС43 spike glycoprotein (6 - AEAEA and 11 - GTCPA) and Plexin-D1. A common pentapeptide DLQLG is located outside immunoreactive epitopes of Plexin-D1.
10. A Shared pentapeptide YSDNG was discovered for Myelin protein P0 and HCoV-NL63 spike glycoprotein. A Shared pentapeptide YSDNG is located outside immunoreactive epitopes in both HCoV-NL63 spike glycoprotein and Myelin protein P0.
11. Shared pentapeptides TIVGA, IVGAL, GRSAL were discovered for sodium channel protein type 9 subunit alpha and immunoreactive epitopes HCoV-NL63 spike glycoprotein. Shared pentapeptides are located outside and immunoreactive epitopes of sodium channel protein type 9 subunit alpha.
12. Shared pentapeptides LSNAS, VTATF were discovered for Plexin-D1 and immunoreactive epitopes HCoV-NL63 spike glycoprotein. Shared pentapeptide VTATF is located within immunoreactive epitope in both HCoV-NL63 spike glycoprotein (one epitope) and Plexin-D1 (one epitope).
13. A Shared pentapeptide FVDVL was discovered for autoantigen of Ubiquitin carboxyl-terminal hydrolase isozyme L1 and immunoreactive epitopes of HCoV-NL63 spike glycoprotein. Shared pentapeptide FVDVL is located within immunoreactive epitope in both HCoV-NL63 spike glycoprotein (2 epitopes) and autoantigen of Ubiquitin carboxyl-terminal hydrolase isozyme L1 (3 epitopes).
14. Shared pentapeptides TLANV, LANVS were discovered for Fibroblast growth factor receptor 3 and immunoreactive epitopes of HCoV-229E spike glycoprotein. Shared pentapeptides are located within immunoreactive epitope of HCoV-229E spike glycoprotein, but outside immunoreactive epitopes in autoantigen of Fibroblast growth factor receptor 3.
15. Shared pentapeptides TFIVL, SYDSV were discovered for Sodium channel protein type 9 subunit alpha and immunoreactive epitopes of HCoV-229E spike glycoprotein. Shared pentapeptides are located outside immunoreactive epitopes in autoantigen of Fibroblast growth factor receptor 3.
Analysis of the data presented in Table 2 allows us to conclude that:
1. A Shared pentapeptide TVEEL was discovered for autoantigen of sodium channel protein type 9 subunit alpha and immunoreactive epitopes of membrane protein of SARS- CoV-2. Shared pentapeptide TVEEL is located within immunoreactive epitopes (25 epitopes) of SARS-CoV-2, but outside immunoreactive epitopes of sodium channel protein type 9 subunit alpha.
2. A Shared pentapeptide TVEEL was discovered for autoantigen of sodium channel protein type 9 subunit alpha and immunoreactive epitopes of membrane protein of SARS- CoV. Shared pentapeptide TVEEL is located within immunoreactive epitopes (4 epitopes) of SARS-CoV, but outside immunoreactive epitopes of sodium channel protein type 9 subunit alpha.
3. A Shared pentapeptide IISIV was discovered for sodium channel protein type 9 subunit alpha and membrane protein of HCoV-HKU-1. A shared pentapeptide IISIV is located outside immunoreactive epitopes in both membrane protein of HCoV-HKU-1 and sodium channel protein type 9 subunit alpha.
4. A Shared pentapeptide ILTVF was discovered for sodium channel protein type 9 subunit alpha and immunoreactive epitope of membrane protein of HCoV-NL63. A shared pentapeptide ILTVF is located outside immunoreactive epitopes of sodium channel protein type 9 subunit alpha.
5. A Shared pentapeptide VGQLP was discovered for Plexin-D1 and two immunoreactive epitopes of membrane protein of HCoV-NL63. A shared pentapeptide VGQLP is located outside immunoreactive epitopes of Plexin-D1.
Analysis of the data presented in Table 3 allows us to conclude that:
1. Shared pentapeptides YFYYL, LALLL were discovered for autoantigen of sodium channel protein type 9 subunit alpha and immunoreactive epitopes of nucleocapsid protein of SARS-CoV-2. Shared pentapeptides YFYYL, LALLL are located within immunoreactive epitopes of SARS-CoV-2 (YFYYL – 10 epitopes, LALLL – 4 epitopes), but outside immunoreactive epitopes of sodium channel protein type 9 subunit alpha.
2. A shared pentapeptide ALLLL was discovered for autoantigen of Ubiquitin carboxyl-terminal hydrolase isozyme L1 and 12 immunoreactive epitopes of SARS-CoV-2. Shared pentapeptide outside immunoreactive epitopes of Ubiquitin carboxyl-terminal hydrolase isozyme L1.
3. Shared pentapeptides YFYYL, LALLL were discovered for autoantigen of sodium channel protein type 9 subunit alpha and immunoreactive epitopes of nucleocapsid protein of SARS-CoV. Shared pentapeptides YFYYL, LALLL are located within immunoreactive epitopes of SARS-CoV (YFYYL – 1 epitope, LALLL – 5 epitopes), but outside immunoreactive epitopes of autoantigen sodium channel protein type 9 subunit alpha.
4. A shared pentapeptide ALLLL was discovered for autoantigen of Ubiquitin carboxyl-terminal hydrolase isozyme L1 and 5 immunoreactive epitopes of SARS-CoV. Shared pentapeptide ALLLL is located outside immunoreactive epitopes of Ubiquitin carboxyl-terminal hydrolase isozyme L1.
5. A shared pentapeptide TDAPS was discovered for autoantigen of Fibroblast growth factor receptor 3 and 7 immunoreactive epitopes of MERS-CoV. Shared pentapeptide TDAPS is located outside immunoreactive epitopes of autoantigen of Fibroblast growth factor receptor 3.
6. Shared pentapeptides YFYYL, GKDSK were discovered for autoantigen of sodium channel protein type 9 subunit alpha and immunoreactive epitopes (two and one, respectively) of HCoV-HKU-1. Shared pentapeptides YFYYL, GKDSK are located outside immunoreactive epitopes of autoantigen of sodium channel protein type 9 subunit alpha.
7. A shared pentapeptide LGKDS was discovered for autoantigen of Plexin-D1 and two immunoreactive epitopes of HCoV-HKU-1. Shared pentapeptide LGKDS is located outside immunoreactive epitopes of autoantigen of Plexin-D1.
8. Shared pentapeptides YFYYL, PIAPG were discovered for autoantigen of sodium channel protein type 9 subunit alpha and immunoreactive epitopes (two and 6, respectively) of HCoV-OC43. Shared pentapeptides YFYYL, PIAPG are located outside immunoreactive epitopes of autoantigen of sodium channel protein type 9 subunit alpha.
9. Shared pentapeptides VLNAS, VAAVT, AAVTL were discovered for autoantigen of Fibroblast growth factor receptor 3 and immunoreactive epitopes (4, 4 and 4, respectively) of HCoV-NL63. Shared pentapeptides VLNAS, VAAVT, AAVTL are located outside immunoreactive epitopes of autoantigen of Fibroblast growth factor receptor 3.
Based on the results, summarized in Table 3, an assumption can be made, that the most important targets for the molecular similarities are the sodium channel subunits and fibroblast growth factor receptor 3, both for seasonal and highly pathogenic coronaviruses.
As a result, we found 50 common shared pentapeptides between human autoantigens of the small nerve fibers and antigens of human coronaviruses. We found 30 mimicking pentapeptides in spike proteins of human coronaviruses, 5 pentapeptides in membrane proteins of human coronaviruses and 15 shared pentapeptides in nucleocapsid proteins of human coronaviruses.
4. Discussion
The aim of the study was to tackle the possibility of the molecular mimicry between human coronaviruses and autoantigens of the human peripheral nervous system. Both seasonal and highly pathogenic coronaviruses were analyzed with an assessment of possible pentapeptide candidates among their spike, membrane and nucleocapsid proteins. The literature search was initialized to find out potential protein candidates from the peripheral nervous system, that may cross-react with coronaviruses with the development of autoimmune reactions. 50 pentapeptides similarities between human autoantigens of small nerve fibers and antigens of human coronaviruses were evaluated, where 60% of the matches were with spike proteins, and 20% each are membrane and nucleocapsid proteins of coronaviruses.
Anticipating the main points of the discussion of the results obtained, attention should be paid to the fact that shared pentapeptides can be located both within immunogenic epitopes and outside them.
Three variants of shared pentapeptides arrangements are possible. The first variant: the TVIIS pentapeptide, shared for two autoantigens of the myelin P2 protein and 12 immunoreactive epitopes of the MERS-CoV virus, is located in the immunoreactive epitopes of both the virus and the autoantigen of nervous system. The second variant: the LALCV pentapeptide, shared for the autoantigen of the fibroblast growth factor receptor 3 and MERS-CoV virus, is located outside the immunoreactive epitopes in both the virus and the autoantigen of nervous system. The third variant: the NLNES pentapeptide, shared for the autoantigen of nervous system plexin-D1 and 32 immunoreactive epitopes of SARS-CoV-2, is located within the immunoreactive epitopes of the virus, but outside the immunoreactive epitopes of the autoantigen of the nervous system. Theoretically possible forth variant, when the shared pentapeptide is located within immunoreactive epitopes of the nervous system, but outside of the immunoreactive epitopes of the virus, we have not encountered in this paper. The considered statements are clearly visible on the example of the search for shared pentapeptides for autoantigens of the nervous system and for the spike protein of human coronaviruses (Table 1).
Analysis of the search for shared pentapeptides for autoantigens of the nervous system and the membrane protein of human coronaviruses (Table 2) and for autoantigens the nervous system and nucleocapsid protein of human coronaviruses (Table 3) showed, that shared pentapeptides are located within immunoreactive epitopes of the virus, but outside of immunoreactive autoantigens of the nervous system (variant three).
The presented considerations are the rationale for formulating the following questions in relation to the phenomenon of molecular mimicry per se:
1. What is the need for the realization of the molecular mimicry phenomenon of location of shared pentapeptides exactly within autoreactive epitopes?
2. Is it possible to realize the molecular mimicry phenomenon when one of the shared pentapeptides is located outside the autoreactive epitope?
3. Is it possible to realize the molecular mimicry phenomenon when the shared pentapeptides in both the virus and the studied autoantigens are located outside the autoreactive epitopes?
4. What is the contribution to the realization of the molecular mimicry phenomenon of shared pentapeptides of various viral proteins (spike glycoprotein, membrane protein, nucleoprotein)?
5. To what extent are the questions posed directly related to post-viral pathology associated with the activation of autoantigens of the nervous system?
Considering these questions, 2 main interpretations may be given to the obtained results. The first is to consider all available pentapeptide similarities as potential candidates for the development of autoimmune processes. The second is to take into account only those of them that are located in immunoreactive epitopes.
Discussing the results based on the first interpretation, 27 common pentapeptides were identified among spike proteins of coronaviruses and autoantigens of the human nervous system. Seasonal coronaviruses had more such potential sites for cross-reactivity than highly pathogenic ones, and plexin D1 and sodium channel protein type 9 subunit alpha had the largest number of common with viruses pentapeptides. Among the membrane proteins, 4 such pentapeptides were identified, while almost all of them had similarities with the sodium channel protein. Among the nucleocapsid proteins, 11 potential candidates for cross-reactivity were identified, which were almost uniformly represented in both highly pathogenic and seasonal coronaviruses.
Considering the second interpretation, where only peptides located in immunologic epitopes are essential for the cross-reactivity, the number of potential candidates for autoimmune processes is narrowing. Such pentapeptides have been identified only in spike proteins of coronaviruses. For highly pathogenic coronaviruses, only one similarity was revealed - between the MERS–CoV virus and fibroblast growth factor receptor 3. For seasonal coronaviruses, five similarities were revealed. For HCoV-HKU-1 virus – with sodium channel protein type 9 subunit alpha and plexin-D1. For HCoV-OC43 virus – with plexin-D1. For HCoV-NL63 virus – with plexin-D1 and ubiquitin carboxyl-terminal hydrolase isozyme L1.
Fibroblast growth factor receptor 3 is promising primarily due to the role of growth factors in the pathophysiology of viral infection. Their increased concentrations were detected in patients suffering from COVID-19 [22,23]. The induction of growth factors possibly occurs due to endothelial dysfunction and may counter the adverse effects of cytokine storms and several other complications. Antifibroblast growth factor receptor 3 may be detected in patients with the idiopathic or autoimmune small fiber neuropathy pain, including corneal pain in the fibromyalgia-type post-COVID-19 syndrome [24,25].
The similar assumption is true for the plexin-D1 and seasonal coronaviruses, where the human protein is another promising target for the autoimmune aggression, especially in the face-related pain syndromes [26]. Plexin was primarily discovered in neurodevelopment, as a part of axonal guidance, but recently its role was described in cardiovascular development and diseases, endotheliocyte migration, angiogenesis and synapse formation [27]. Its autoantibodies were described in several cases of idiopathic pain, linking together endothelial dysfunction and autoimmune processes in the nervous system [28].
Seasonal coronaviruses share numerous amino acid sequences with the sodium channel protein type 9 subunit alfa, which disfunction can be responsible for neuropathic pain in humans [29]. It is infamous for being involved in pain pathophysiology both in hereditary and autoimmune disease, including small finer neuropathy, erythromelalgia and paroxysmal extreme pain disorder [30]. The main mechanism involves the electric conduction dysfunction, cased either by autoantibodies to this protein or impairment of its structure [31].
Ubiquitin carboxyl-terminal hydrolase isozyme L1, that may be a target for the molecular mimicry in coronaviruses, is known for its multiple roles in the inflammation, angiogenesis, neuronal injury and neurodegeneration [21], Hence, its alteration can be important in the development small fiber neuropathy and microvascular complications in the post-COVID-19 conditions.
5. Conclusions
The obtained results confirm the presence of a possible cross-reactivity between coronavirus proteins and antigens of small human nerve fibers. It is important to note that the greatest number of common amino acid sequences with components of the peripheral nervous system was detected in seasonal coronaviruses HCoV-HKU-1, HCoV-OC43, HCoV-NL63. The largest number of similar pentapeptides were found with sodium channel proteins. Autoantibodies to these structures can be detected in autoimmune lesions of the nervous system, leading to the development of polyneuropathy, characteristic of patients with post-COVID syndrome. Fibroblast growth factor receptor 3 is another example of a protein molecule to which autoantibodies can be produced. The presence of these antibodies is considered by some authors as one of the most promising methods for diagnosing neuropathy of small fibers [23]. Plexin-D1, which according to the results of this study has 10 amino acid sequences in common with coronavirus S-proteins, is involved in endotheliocyte migration, angiogenesis and synapse formation. The data obtained make it possible to identify new potential targets for the development of autoimmune reactions that occur against the background of the activity of highly pathogenic and seasonal coronaviruses.
Author Contributions
“Conceptualization, L.P.C. and N.Y.G.; methodology, L.P.C. and M.G.N.; software, M.G.N.; investigation, M.G.N.; resources, M.G.N.; data curation, V.J.U., L.A.S., T.V.F.; writing—original draft preparation, N.Y.G., M.G.N., V.J.U., L.A.S., T.V.F.; writing—review and editing, N.Y.G., V.J.U and L.P.C.; visualization, M.G.N.; supervision, L.P.C.; project administration, L.P.C. All authors have read and agreed to the published version of the manuscript.”
Funding
The research was supported by the grant of the Russian Science Foundation of the Russian Academy of Sciences No. 22-15-00113 dated 13.05.2022, https://rscf.ru/project/22-15-00113.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Spiteri, G.; Fielding, J.; Diercke, M.; Campese, C.; Enouf, V.; Gaymard, A.; Ciancio, B. C. First cases of coronavirus disease 2019 (COVID-19) in the WHO European Region, 24 January to 21 February 2020. Eurosurveillance 2020, 25, 2000178. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Cao, B. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. The lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Tan, W. Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding. The lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Al-Ramadan, A.; Rabab’h, O.; Shah, J.; Gharaibeh, A. Acute and Post-Acute Neurological Complications of COVID-19. Neurol. Int. 2021, 13, 102–119. [Google Scholar] [CrossRef] [PubMed]
- Halpert, G.; Shoenfeld, Y. SARS-CoV-2, the autoimmune virus. Autoimmun Rev. 2020, 19, 102695. [Google Scholar] [CrossRef] [PubMed]
- Tomar, B.S.; Singh, M.; Nathiya, D.; et al. Prevalence of Symptoms in Patients Discharged from COVID Care Facility of NIMS Hospital: Is RT PCR Negativity Truly Reflecting Recovery? A Single-Centre Observational Study. Int J Gen Med. 2021, 14, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Soldan, S.S.; Lieberman, P.M. Epstein-Barr virus and multiple sclerosis. Nat Rev Microbiol 2023, 21, 51–64. [Google Scholar] [CrossRef]
- Laman, J.D.; Huizinga, R.; Boons, G.J.; Jacobs, B.C. Guillain-Barré syndrome: expanding the concept of molecular mimicry. Trends Immunol. 2022, 43, 296–308. [Google Scholar] [CrossRef]
- Kanduc, D.; Shoenfeld, Y. Molecular mimicry between SARS-CoV-2 spike glycoprotein and mammalian proteomes: implications for the vaccine. Immunol Res. 2020, 68, 310–313. [Google Scholar] [CrossRef] [PubMed]
- Kanduc, D. From Anti-SARS-CoV-2 Immune Responses to COVID-19 via Molecular Mimicry. Antibodies 2020, 9, 33. [Google Scholar] [CrossRef] [PubMed]
- Novak, P. Post COVID-19 syndrome associated with orthostatic cerebral hypoperfusion syndrome, small fiber neuropathy and benefit of immunotherapy: a case report. eNeurologicalSci. 2020, 21, 100276. [Google Scholar] [CrossRef]
- Koralnik, I.J.; Tyler, K.L. COVID-19: a global threat to the nervous system. Annals of neurology 2020, 88, 1–11. [Google Scholar] [CrossRef]
- Neumann, B.; Schmidbauer, M.L.; Dimitriadis, K.; Otto, S.; Knier, B.; Niesen, W.D.; Bösel, J. Cerebrospinal fluid findings in COVID-19 patients with neurological symptoms. Journal of the neurological sciences 2020, 418. [Google Scholar] [CrossRef]
- Paterson, R.W.; Brown, R.L.; Benjamin, L.; Nortley, R.; Wiethoff, S.; Bharucha, T.;Zandi, M.S. The emerging spectrum of COVID-19 neurology: clinical, radiological and laboratory findings. Brain 3104, 143, 3104–3120. [Google Scholar] [CrossRef]
- Solomon, I.H.; Normandin, E.; Bhattacharyya, S.; Mukerji, S.S.; Keller, K.; Ali, A.S.; Sabeti, P. Neuropathological features of Covid-19. New England Journal of Medicine 2020, 393, 989–992. [Google Scholar] [CrossRef] [PubMed]
- UniProt Consortium. UniProt: a worldwide hub of protein knowledge. Nucleic acids research 2019, 47, D506––D515. [Google Scholar] [CrossRef]
- Berman, H. M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T. N.; Weissig, H.; Shindyalov, I, N; Bourne, P. E. The protein data bank. Nucleic acids research 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Velankar, S. AlphaFold Protein Structure Database: massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic acids research 2022, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger, L. The PyMOL molecular graphics system, version 2.5.4.
- Vita, R.; Mahajan, S.; Overton, J. A.; Dhanda, S. K.; Martini, S.; Cantrell, J. R.; Peters, B. The immune epitope database (IEDB): 2018 update. Nucleic acids research 2019, 47, D339–D343. [Google Scholar] [CrossRef] [PubMed]
- Matuszczak, E.; Tylicka, M.; Komarowska, M.D.; Debek, W.; Hermanowicz, A. Ubiquitin carboxy-terminal hydrolase L1 - physiology and pathology. Cell Biochem Funct. 2020, 38, 533–540. [Google Scholar] [CrossRef]
- Gupta, A.; Jayakumar, M.N.; Saleh, M.A.; Kannan, M.; Halwani, R.; Qaisar, R.; Ahmad, F. SARS-CoV-2 infection- induced growth factors play differential roles in COVID-19 pathogenesis. Life Sci. 2022, 304, 120703. [Google Scholar] [CrossRef]
- Trevino, J.A.; Novak, P. TS-HDS and FGFR3 antibodies in small fiber neuropathy and Dysautonomia. Muscle Nerve. 2021, 64, 70–76. [Google Scholar] [CrossRef]
- Nagarajan, E.; Kang, S.A.; Holmes, C.; Govindarajan, R. Clinical features with anti fibroblast growth factor receptor 3 (FGFR3) antibody-related polyneuropathy: a retrospective study. BMC Neurol. 2021, 21, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bayraktutar, B.N.; Atocha, V.; Farhad, K.; Soto, O.; Hamrah, P. Autoantibodies Against Trisulfated Heparin Disaccharide and Fibroblast Growth Factor Receptor-3 May Play a Role in the Pathogenesis of Neuropathic Corneal Pain. Cornea. 2023, 42, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Zeidman, L.A.; Saini, P.; Mai, P. Immune-Mediated Small Fiber Neuropathy With Trisulfated Heparin Disaccharide, Fibroblast Growth Factor Receptor 3, or Plexin D1 Antibodies: Presentation and Treatment With Intravenous Immunoglobulin. J Clin Neuromuscul Dis. 2022, 24, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Holl, E.K.; O'Connor, B.P.; Holl, T.M.; Roney, K.E.; Zimmermann, A.G.; Jha, S.; Kelsoe, G.; Ting, J.P.-Y. Plexin-D1 is a novel regulator of germinal centers and humoral immune responses. J Immunol. 2011, 186, 5603–5611. [Google Scholar] [CrossRef]
- Fujii, T.; Yamasaki, R.; Iinuma, K.; Tsuchimoto, D.; Hayashi, Y.; Saitoh, B.Y.; Matsushita, T.; Kido, M.A.; Aishima, S.; Nakanishi, H.; Nakabeppu, Y.; Kira, J.I. A Novel Autoantibody against Plexin D1 in Patients with Neuropathic Pain. Ann Neurol. 2018, 84, 208–224. [Google Scholar] [CrossRef]
- Kelley, M.A.; Oaklander, A.L. Association of small-fiber polyneuropathy with three previously unassociated rare missense SCN9A variants. Can J Pain. 2020, 4, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Hisama, F.M.; Dib-Hajj, S.D.; Waxman, S.G. SCN9A Neuropathic Pain Syndromes. In GeneReviews®; Adam, M.P., Mirzaa, G.M., Pagon, R.A., et al., Eds.; University of Washington: Seattle (WA), 2006. [Google Scholar]
- Amaya, F.; Wang, H.; Costigan, M.; Allchorne, A.J.; Hatcher, J.P.; Egerton, J.; Stean, T.; Morisset, V.; Grose, D.; Gunthorpe, M.J.; Chessell, I.P.; Tate, S.; Green, P.J.; Woolf, C.J. The voltage-gated sodium channel Na(v)1.9 is an effector of peripheral inflammatory pain hypersensitivity. J Neurosci. 2006, 26, 12852–12860. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Confocal microscopy of the cornea (personal observation). (a) - 26 y.o. female with the post-COVID-19 syndrome. Reduced length and increased tortuosity of corneal nerve fibers (black arrows), as well as a large number of activated antigen-presenting cells (white arrows) are shown. (b) – 31 y.o. healthy volunteer.
Figure 1.
Confocal microscopy of the cornea (personal observation). (a) - 26 y.o. female with the post-COVID-19 syndrome. Reduced length and increased tortuosity of corneal nerve fibers (black arrows), as well as a large number of activated antigen-presenting cells (white arrows) are shown. (b) – 31 y.o. healthy volunteer.
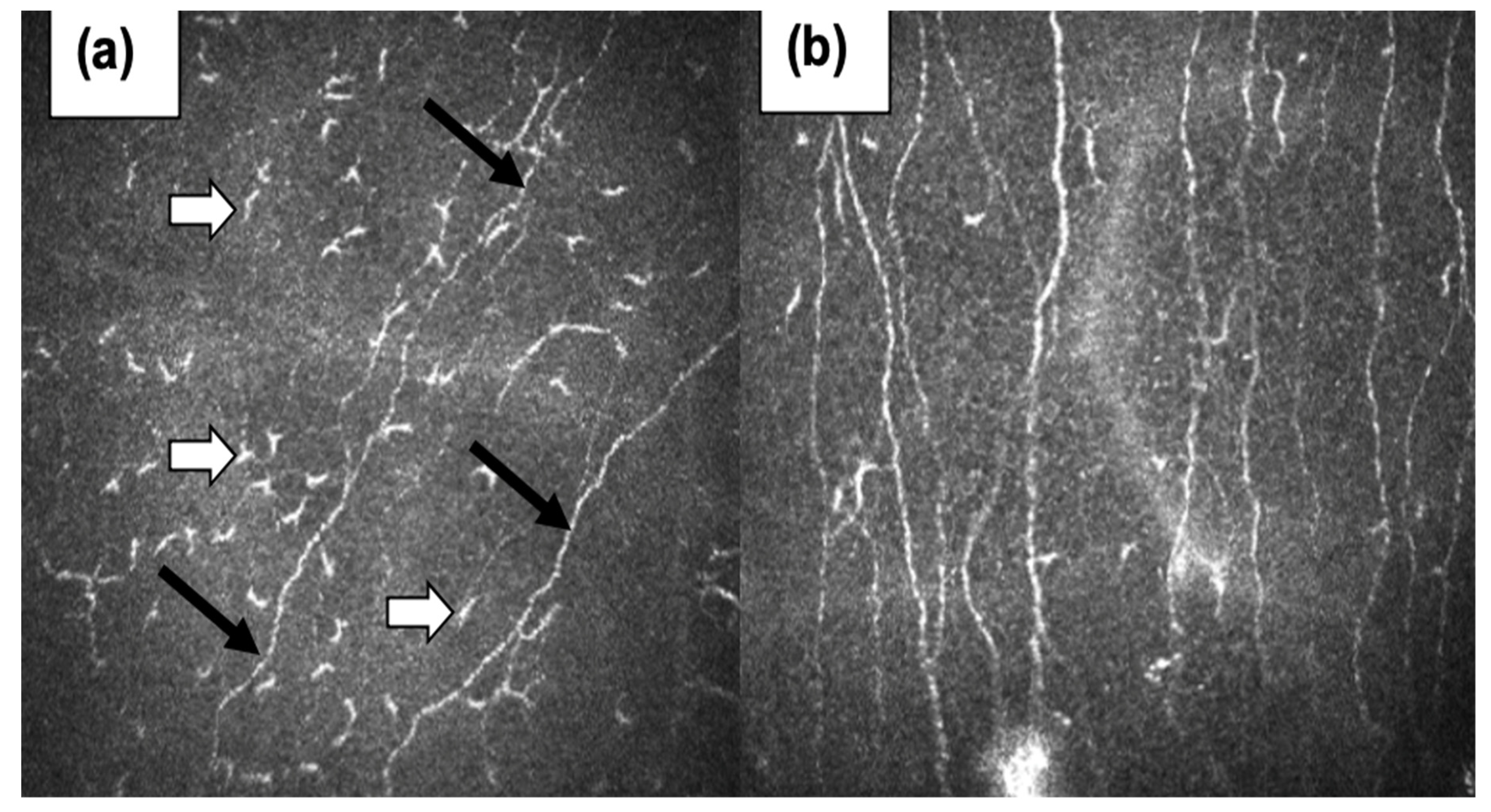
Figure 2.
Location of pentapeptides in 3D structure spike glycoproteins of all human coronaviruses. (According to databases: PDB). Pentapeptides are shown with different colors. Structural visualization generated with PyMoL v.2.5.4.
Figure 2.
Location of pentapeptides in 3D structure spike glycoproteins of all human coronaviruses. (According to databases: PDB). Pentapeptides are shown with different colors. Structural visualization generated with PyMoL v.2.5.4.
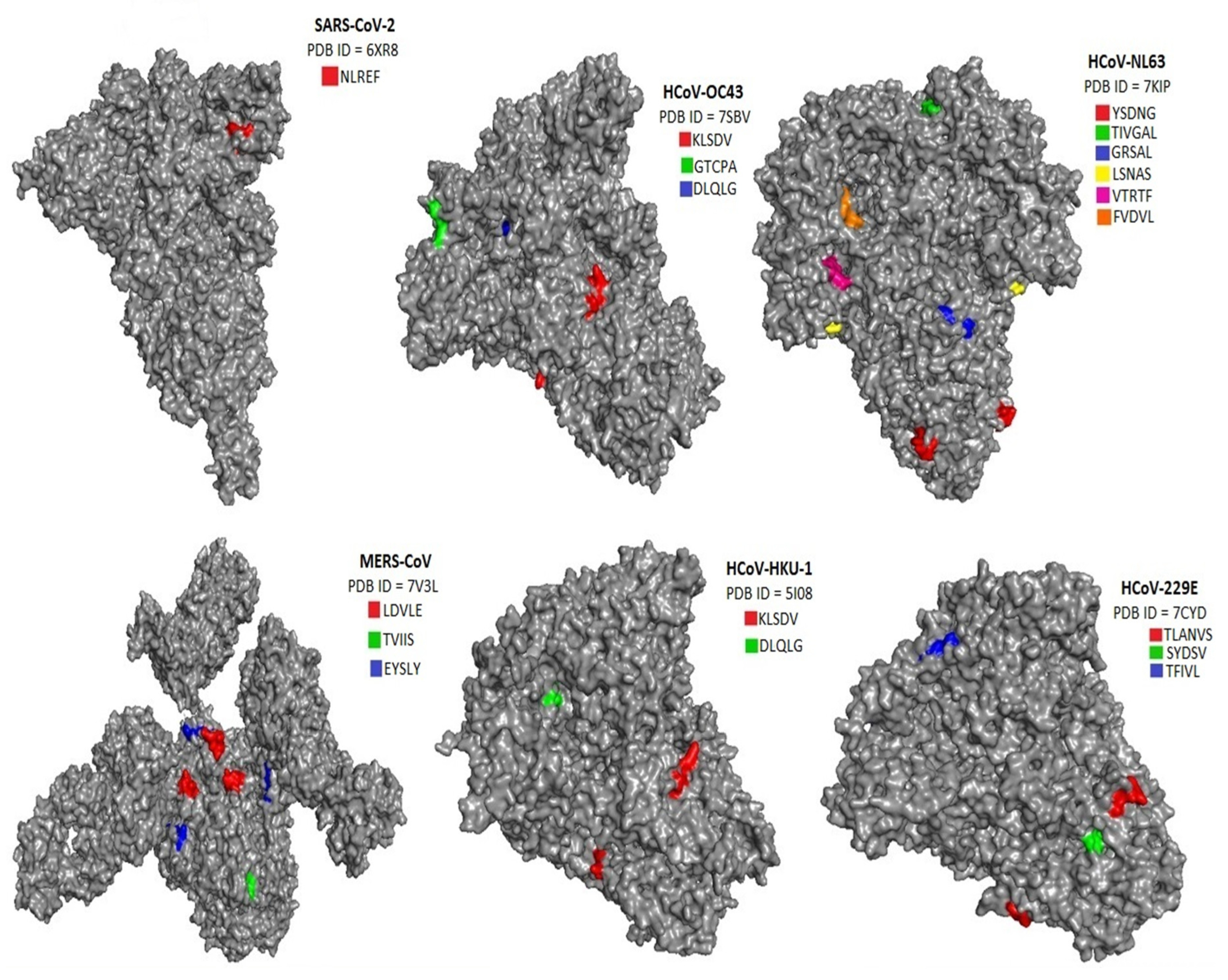
Figure 3.
Location of pentapeptides in 3D structure autoantigens of human nervous system. (According to databases: AlphaFold). Pentapeptides are shown with different colors. Structural visualization generated with PyMoL .2.5.4.
Figure 3.
Location of pentapeptides in 3D structure autoantigens of human nervous system. (According to databases: AlphaFold). Pentapeptides are shown with different colors. Structural visualization generated with PyMoL .2.5.4.
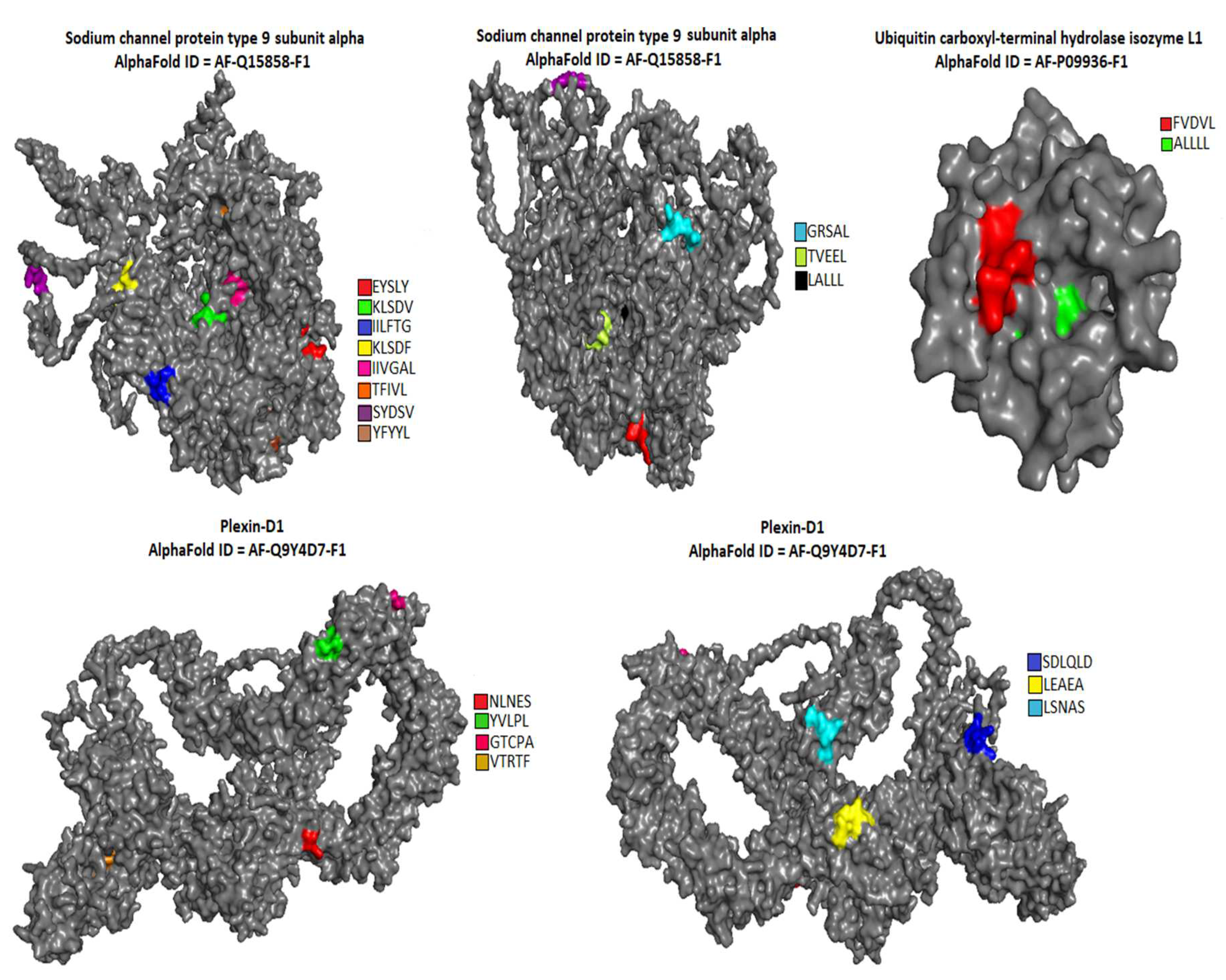
Figure 4.
Location of pentapeptides in 3D structure autoantigens of human nervous system. (According to databases: AlphaFold). Pentapeptides are shown with different colors. Structural visualization generated with PyMoL v.2.5.4.
Figure 4.
Location of pentapeptides in 3D structure autoantigens of human nervous system. (According to databases: AlphaFold). Pentapeptides are shown with different colors. Structural visualization generated with PyMoL v.2.5.4.
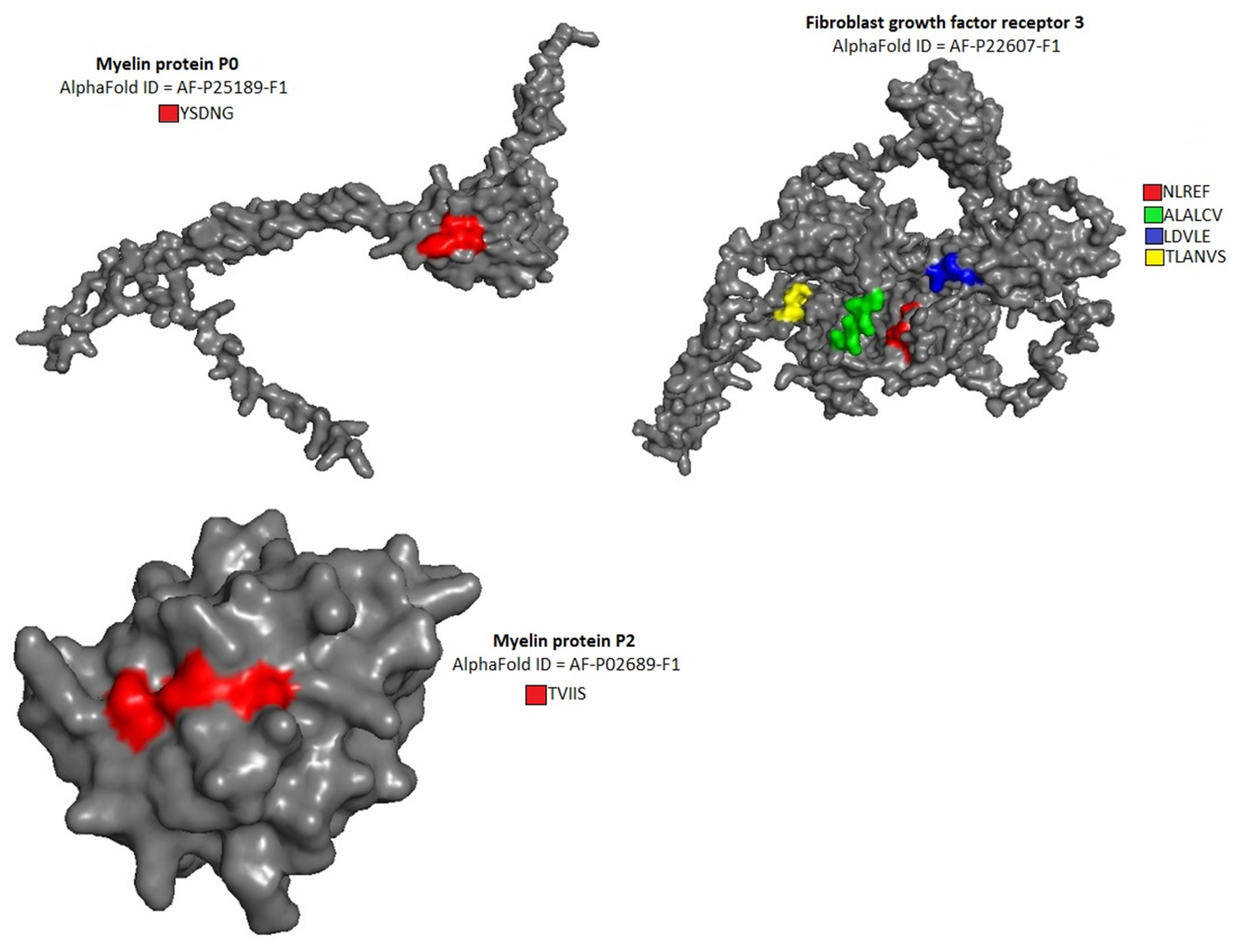

Table 1.
Shared pentapeptides of human autoantigens of nervous system and of spike glycoprotein of human coronaviruses. In square brackets - the number of immunoreactive epitopes containing shared pentapeptides in human coronaviruses. In parentheses - the number of immunoreactive epitopes containing shared pentapeptides in human autoantigens of nervous system.
Table 1.
Shared pentapeptides of human autoantigens of nervous system and of spike glycoprotein of human coronaviruses. In square brackets - the number of immunoreactive epitopes containing shared pentapeptides in human coronaviruses. In parentheses - the number of immunoreactive epitopes containing shared pentapeptides in human autoantigens of nervous system.
| Hum. Coronaviruses // autoantigens of nervous system | SARS-CoV-2 (P0DTC2) |
SARS-CoV (P59594) |
MERS-CoV (R9UQ53) |
HCoV-HKU-1 (S5YA28) |
HCoV-OC43 (P36334) |
HCoV-NL63 (Q6Q1S2) |
HCoV-229E (P15423) |
|---|---|---|---|---|---|---|---|
| Fibroblast growth factor receptor 3 (P22607) | NLREF [26], (-) |
- | ALALC [-], (-), LALCV [-], (-), LDVLE [14], (1) |
- | - | - | TLANV [7], (-), LANVS [6], (-) |
| Myelin protein P0 (P25189) | - | - | - | - | - | YSDNG [-], (-) |
- |
| Myelin protein P2 (P02689) |
- | - | TVIIS [12], (2) |
- | - | - | - |
| Sodium channel protein type 9 subunit alpha (Q15858) | - | - | EYSLY [1], (-) |
KLSDV [5], (-), IILFT [-], (-), ILFTG [3], (-), KLSDF [15], (1) |
KLSDV [10], (-) |
TIVGA [2], (-), IVGAL [2], (-), GRSAL [16], (-) |
TFIVL [3], (1), SYDSV [11], (-) |
| Plexin-D1 (Q9Y4D7) |
NLNES [32], (-) |
NLNES [2], (-) |
- | YVLPL [1], (7), SDLQL [4], (-), DLQLG [7], (-) |
LEAEA [6], (1), GTCPA [11], (1), DLQLG [13], (-) |
LSNAS [6], (-), VTRTF [1], (1) |
- |
| Ubiquitin carboxyl-terminal hydrolase isozyme L1 (P09936) | - | - | - |
- | - | FVDVL [2], (3) |
- |
Table 2.
Shared pentapeptides of human autoantigens of nervous system and of membrane protein of human coronaviruses. In square brackets – the number of immunoreactive epitopes containing shared pentapeptides in human coronaviruses. In parentheses – the number of immunoreactive epitopes containing shared pentapeptides in human autoantigens of nervous system.
Table 2.
Shared pentapeptides of human autoantigens of nervous system and of membrane protein of human coronaviruses. In square brackets – the number of immunoreactive epitopes containing shared pentapeptides in human coronaviruses. In parentheses – the number of immunoreactive epitopes containing shared pentapeptides in human autoantigens of nervous system.
| Hum. Coronaviruses // autoantigens of nervous system | SARS-CoV-2 (P0DTC5) |
SARS-CoV (P59596) |
MERS-CoV (R9UNX5) |
HCoV-HKU-1 (Q0ZJ82) |
HCoV-OC43 (Q01455) |
HCoV-NL63 (U3M6Q8) |
HCoV-229E (P15422) |
||
|---|---|---|---|---|---|---|---|---|---|
| Fibroblast growth factor receptor 3 (P22607) | - |
- | - | - | - | - | - | ||
| Myelin protein P0 (P25189) | - | - | - | - | - | - | - | ||
| Myelin protein P2 (P02689) |
- | - | - | - | - | - | - | ||
| Sodium channel protein type 9 subunit alpha (Q15858) | TVEEL [25], (-) |
TVEEL [4], (-) |
- | IISIV [-], (-) |
- | ILTVF [1], (-) |
- | ||
| Plexin-D1 (Q9Y4D7) |
- | - | - | - | - | VGQLP [2], (-) |
- | ||
| Ubiquitin carboxyl-terminal hydrolase isozyme L1 (P09936) | - | - | - | - | - | - | - | ||
Table 3.
Shared pentapeptides of human autoantigens of nervous system and of nucleocapsid protein of human coronaviruses. In square brackets – the number of immunoreactive epitopes containing shared pentapeptides in human coronaviruses. In parentheses – the number of immunoreactive epitopes containing shared pentapeptides in human autoantigens of nervous system.
Table 3.
Shared pentapeptides of human autoantigens of nervous system and of nucleocapsid protein of human coronaviruses. In square brackets – the number of immunoreactive epitopes containing shared pentapeptides in human coronaviruses. In parentheses – the number of immunoreactive epitopes containing shared pentapeptides in human autoantigens of nervous system.
| Hum. Coronaviruses // autoantigens of nervous system | SARS-CoV-2 (P0DTC9) |
SARS-CoV (P59595) |
MERS-CoV (T2BBK0) |
HCoV-HKU-1 (S5ZBQ7) |
HCoV-OC43 (P33469) |
HCoV-NL63 (H9EJA4) |
HCoV-229E (P15130) |
|---|---|---|---|---|---|---|---|
| Fibroblast growth factor receptor 3 (P22607) | - |
- | TDAPS [7], (-) |
- | - | VLNAS [4], (-), VAAVT [4], (-), AAVTL [4], (-) |
- |
| Myelin protein P0 (P25189) | - | - | - | - | - | - | - |
| Myelin protein P2 (P02689) |
- | - | - | - | - | - | - |
| Sodium channel protein type 9 subunit alpha (Q15858) | YFYYL [10], (-), LALLL [4], (-) |
YFYYL [1], (-), LALLL [5], (-) |
- | YFYYL [2], (-), GKDSK [1], (-) |
YFYYL [2], (-), PIAPG [6], (-) |
- |
- |
| Plexin-D1 (Q9Y4D7) |
- | - | - | LGKDS [2], (-) |
- | - | - |
| Ubiquitin carboxyl-terminal hydrolase isozyme L1 (P09936) | - | - | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



