
Submitted:
29 June 2023
Posted:
30 June 2023
You are already at the latest version
Abstract
Much evidence reveals an important link between gut microbiota and the heart. In particular, the gut microbiota plays a key role in the onset of cardiovascular (CV) disease, including heart failure (HF). In HF, splanchnic hypoperfusion causes intestinal ischemia resulting in the translocation of bacteria and their metabolites into the blood circulation. Among these, the most important is the Trimethylamine N-Oxide (TMAO), which is responsible through various mechanisms for pathological processes in different organs and tissues. In this review, we summarise the complex interaction between gut microbiota and CV disease, particularly with HF, and the possible strategies to influence its composition and function. Finally, we highlight the potential role of TMAO as a novel prognostic marker and new therapeutic target of HF.
Keywords:
Gut microbiota
; Cardiovascular diseases
; Trimethylamine N-Oxide
; Heart failure
; Atherosclerosis
1. Introduction
The human gastrointestinal (GI) tract contains a rich and complex population of microorganisms constituting the gut microbiota [1].
Considering the symbiotic interaction between gut microbiota and the host, we can consider the host as a "superorganism" [2], whose metabolism is dependent on a combination of both microbial and human physiological processes, which perform immune and metabolic functions [1,3].
Gut bacteria play a role in the digestive process and contribute to the absorption of many nutrients and metabolites, including essential amino acids, vitamins, lipids, and short-chain fatty acids (SCFAs) [4]. The gut microbiota has a critical immune function that promotes host resistance to infectious diseases [5].
Everyone is endowed with a specific gut microbiota profile. It comprises different bacterial species taxonomically classified by phyla, classes, orders, families, genera and species [6,7].
Several factors can alter the gut microbiota, including genotype, dietary habits, age [8,9], mode of birth [10,11] and antibiotics [12,13]. Disruption of the gut microbiota is associated with several human diseases, including inflammatory bowel disease (IBD) [14,15], obesity, diabetes [15], allergies [16], autoimmune disease [17], CV disease [18,19] and hypertension [20]. Modifying its composition and diversity is a promising treatment option for these diseases. There are many ways to modulate the gut microbiota, including probiotics, prebiotics, and transplantation of the faecal microbiota, which can induce beneficial changes in its structure and functions.
Recent evidence suggests a link between the gut microbiota and CV disease, including HF. Despite the progress in pharmacological and non-pharmacological therapies, the latter is still burdened by very high mortality with poor prognosis.
This review aims to highlight the complex interplay between the microbiota, its metabolites, and CV disease's development and/or progression, emphasizing HF and giving exciting directions for possible future research.
2. The link between gut microbiota and heart
In the last decade, scientific awareness of the gut microbiota and its possible link to heart disease has grown. In fact, chemicals or processes associated with gut bacteria have been linked to a higher risk of HF, atherosclerosis, and major CV events such as heart attack and stroke [21].
While there is still much to learn about how the gut microbiota affects various disease risks, it is clear that an unhealthy gut negatively impacts health through inflammation, the immune system's response to an injury or foreign substance.
Seventy percent of the body's inflammatory cells are in gut-associated tissue [22]. So, gut inflammation possesses substances, including chemicals produced by gut bacteria, that leave the gut and enter the bloodstream. This causes an inflammatory response wherever they enter the body. When the inflammation affects the blood vessels, these lose their elasticity, the endothelial wall function is impaired, and pathophysiological conditions lead to atherosclerotic disease [23].
Many studies have shown that unhealthy gut microbiota and its active metabolites are involved in the onset and progression of CV disease. Among these, although the pathophysiological mechanisms are not yet fully understood, TMAO is the main substance able to trigger the inflammatory process that leads to the development of both metabolic and CV diseases.
In a meta-analysis study that included 19 clinical trials, the authors found that increased levels of TMAO were associated with an increased risk of 62% of major CV events, such as heart attack and stroke, and an increased risk of 63% of death from all causes. These results were homogeneous across study populations [24].
Another meta-analysis study that evaluated 17 trials showed that high blood levels of TMAO were associated with an increased risk of 67% and 91% of major CV events and death from all causes, respectively. The risk of death increased proportionally with the rise in levels of TMAO, not only at very high levels of this metabolite [25].
It's important to note that, according to the available data, even with adjustment for traditional risk factors - considering differences in blood pressure, cholesterol, or triglycerides - an elevated level of TMAO may predict an increased risk of CV events.
Another notable finding occurred in an experimental study of mice where the animals with atherosclerosis were fed a beneficial type of bacteria that uses TMAO for food. This kind of food resulted in lower levels of TMAO in their blood and reduced the amount of atherosclerotic plaque [26].
Two types of supplements are commonly recommended to support healthy gut microbiota: probiotics and prebiotics. Probiotics contain beneficial bacteria; the prebiotics contains substances that can promote the growth of beneficial bacteria. While there is evidence that some types of these supplements benefit gut health [27], it is not yet clarified whether taking probiotics or prebiotics can reduce inflammation and CV disease in the long term [28].
3. Gut microbiota and atherosclerosis
Infectious processes are associated with the development of CV disease and atherosclerosis. Recent research has identified microbial ecosystems in different parts of the human body as the cause of CV and metabolic disorders through multiple pathways [29]. First, local or systemic infections can trigger inflammation contributing to plaque development and/or destabilisation [30]. Second, cholesterol and lipid metabolism by the gut microbiota may lead to the development of atherosclerosis and, finally, dietary components metabolised by the gut microbiota may influence atherosclerosis differently [29]. Considering that the number of bacterial species differs between individuals, the microbiota is comparable at higher taxonomic levels (e.g., phyla). For example, Bacteroidetes and Firmicutes, which account for more than 90% of all taxa in the human intestine, are the two phyla that dominate the gut microbiota [31]. Plaque formation may be caused by an extended infection or a disease of the vessel wall cells; the discovery of bacterial DNA in plaques supports this mechanism [32]. The microorganisms of the atherosclerotic plaques are also found in other body parts, especially in the intestine. Among the infections caused by bacteria in atherosclerotic plaques, the gut microbiota affects CV disease by modulating host metabolism, including lipoprotein metabolism. Patients with atherosclerosis usually have an impairment of lipid metabolism, which can be modulated by the bacterial taxa of the gut microbiota [33]. Some microbiota species generate enzymes that facilitate the fermentation of indigestible carbohydrates into SCFAs [34]. These SCFAs have potential anti-inflammatory properties. The bacteria regulate intestinal permeability. Certain species may render the gut permeable, allowing microbe-associated metabolites to enter the circulatory system and elicit an inflammatory response following which, our body produces cytokines and other inflammatory mediators [35]. As mentioned, gastrointestinal bacteria can convert complex carbohydrates that the host otherwise undigested into SCFAs. SCFAs play a crucial function in the relationship between diet, gut microbiota and stimulation, or inhibition, of inflammatory cascades downstream. The gut microbiota can convert primary bile acids into secondary bile acids. Changes in gut microbiota affect the varieties of secondary bile acids synthesised. Particularly relevant is the fact that bile acids cause inflammation by triggering the farnesoid X receptor (FXR) signalling networks in enterocytes and adipocytes [36]. Choline, carnitine and betaine, found in red meat, fish and other animal sources, are converted by microorganisms in the gut into trimethylamine (TMA), which is converted by hepatic flavin monooxygenases into TMAO. When present in high serum concentrations, TMAO has been associated with adverse effects, including endothelial dysfunction, promoting vascular inflammation, atherosclerosis, and another CV disease, such as HF [37].
4. The gut hypothesis of HF
A lot of data supports the pathogenetic role of gut microbiota in the development of HF, so the “gut hypothesis of HF” has become interesting and even fascinating.
It is widely acknowledged that congestive HF (CHF) is a multisystemic disorder often associated with alterations in gut microbiota composition and function [38]. A well-balanced gut microbiota, an intact mucosa, and a normal functioning immune system are necessary to preserve the normal functionality of the gut barrier [39].
Several studies have shown that patients with CHF have an impairment of gut microbiota composition compared to healthy individuals. The gut of patients with CHF may present an intestinal overgrowth of pathogenic bacteria (adherent bacterial populations, such as Bacteroides and Fusobacterium) and fungal species (such as Candida), which can cause chronic intestinal wall inflammation that leads to malabsorption and decreased metabolic efficiency [40].
The decrease of cardiac output and the increase in sympathetic activity in CHF patients, cause a redistribution of blood flow away from the splanchnic circulation. These changes lead to reduced intestinal mucosal perfusion, intramucosal acidosis, mucosal oedema (thickened bowel wall), ischemia, epithelial dysfunction and increased mucosal permeability in the small and large intestines [41].
Impairment of gut barrier function allows bacteria and their products to cross the gut barrier -a process known as bacterial translocation- spreading to the mesenteric lymph nodes or entering the bloodstream and reaching more distant organs. Although it has been generally considered a pathologic process, it is common during early life and may help mucosal antigen sampling in the gut [42]. Three mechanisms have been suggested for the pathogenesis of bacterial translocation: bacterial overgrowth, an altered gut barrier function and impaired host defences [39] (Figure 1). The bacterial translocation increases the bacteria, as well as the endotoxin circulation (such as lipopolysaccharide, or LPS) that can activate monocytes and macrophages to release pro-inflammatory mediators [43], that take part in chronic systemic inflammation, which is a hallmark of CHF [44]. The increased levels of circulating cytokines further compromise the already dysfunctional gut barrier, promoting a more increase in bacterial translocation.
Niebauer et al. found that patients with CHF and recent onset of peripheral oedema presented increased serum concentrations of endotoxin and cytokines, and after diuretic treatment endotoxin levels (but not cytokines) normalised, suggesting that endotoxin may trigger immune activation and peripheral oedema [45]. During an acute or decompensated HF, endotoxin levels were higher in hepatic veins than in the left ventricle or pulmonary artery, suggesting endotoxin translocation from the gut to the bloodstream [46]. In addition, selective gut decontamination (aerobic gram-negative bacilli eradication) in patients with severe CHF reduced faecal endotoxin concentrations, monocyte CD14 expression, and intracellular pro-inflammatory cytokine production [47].
Bacterial translocation's role in HF pathogenesis is still being actively researched. However, it is recognised that the gut microbiota and its interaction with the host, significantly impact cardiovascular health. Strategies aimed at modulating the gut microbiota may have a potential therapeutic benefit in HF management.
5. TMAO, inflammation and CVD
TMAO can directly affect the heart by causing myocardial hypertrophy, fibrosis, inflammation, and mitochondrial dysfunction, indirectly promoting renal fibrosis and dysfunction, and platelet hyperreactivity [48,49]. Zhiye et al. demonstrated in vitro and in vivo studies that TMAO treatment directly caused cardiac hypertrophy and fibrosis, as evidenced by increased cardiomyocyte size and high levels of hypertrophic markers including atrial natriuretic peptide (ANP) and beta-myosin heavy chain (β-MHC). In addition, cardiac hypertrophy can be markedly inhibited by reducing the synthesis of TMAO, by antibiotics therapy [50]. The Smad3 signalling pathway was found to be activated in mice induced by transverse aortic constriction [51] and it would appear to play a key role in hypertrophy and fibrosis. In fact, its inhibition by SIS3 [50] markedly reduces cardiac remodelling. Moreover, it was found that an inhibitor of the synthesis of TMAO, the 3,3-dimethyl-1-butanol (DMB), prevents myocardial hypertrophy and fibrosis by regulating transforming growth factor-β1 (TGF-β1)/Smad3 and nuclear factor p65- κB (NF-κB) [52]. These findings confirm the role of TMAO in ventricular remodelling. The NF-kB signalling pathway is also responsible for inflammation in vascular smooth muscle cells. TMAO activates mitochondrial reactive oxygen species (mtROS) by inhibiting both sirtuin 3 (SIRT3) expression and superoxide dismutase 2 (SOD2) activity, which subsequently activates the inflammasome (NLRP3) leading to the extension of inflammation to endothelial cells [53]. Furthermore, TMAO promotes myocardial inflammation by increasing TNF-α levels and decreasing IL-10 levels leading to increased expression of pro-inflammatory cytokines and decreased expression of anti-inflammatory cytokines [54]. The effect of TMAO on endothelial dysfunction is partly attributable to the activation of PKC/NF-kB, resulting in increased VCAM-1 expression and monocyte adhesion [55]. TMAO decreases energy metabolism and mitochondrial function by influencing pyruvate and fatty acid oxidation, which are involved in the tricarboxylic acid cycle [56]. How does TMAO cause cardiac damage indirectly? TMAO directly contributes to renal interstitial fibrosis and dysfunction, promoting sodium and water retention. The kidney plays a key role in TMAO excretion [57], so impaired renal function causes its accumulation with the progression of cardiac injury. Finally, TMAO directly increases platelet hyperreactivity, promoting platelet adhesion to collagen and mobilization of cytoplasmic Ca 2+, promoting a prothrombotic phenotype with an increased risk of myocardial infarction [58] (Figure 2).
6. TMAO: a prognostic marker of HF
The first description of an association between increased TMAO levels and mortality risk in patients with HF dates back to 2014 [59]. In advanced CHF a common problem is venous congestion deriving from right-sided HF. The increase of venous pressure and splanchnic congestion in relation to the gut is associated with deleterious effects. The gut environmental alterations predispose the host to colonisation with harmful TMAO-producing bacteria, and the increase of TMAO levels, as in a negative loop, leads to the further worsening of HF [60]. Increased TMAO levels have also been shown to be a strong predictor of mortality in patients with HF with reduced ejection fraction (HFrEF), but not in those with preserved ejection fraction (HFpEF), over a mean follow-up of 9.7 years [61].
In a meta-analysis by Xingxing Li et al. the results deriving from 12 studies involving 13,425 participants from 2014 to 2021 demonstrated a strong association between elevated concentrations of TMAO and poor prognosis in patients with HF. Of the 12 studies of this meta-analysis, 9 included patients with chronic HF. From this analysis, high TMAO levels correlate with a greater risk of MACEs and an increase in all-cause mortality in patients with HF [62]. Several studies have reported that elevated TMAO levels are associated with poor prognoses in patients with HFrEF [59,63].
In a recent study including 1208 patients with chronic HF post myocardial infarction, by using a Cox regression analysis, it has been possible to demonstrate the association between plasma TMAO levels and cardiovascular outcomes. TMAO might be considered a predictor of MACE and could be used to improve risk stratification in patients with ischemic heart disease developing chronic HF. By investigating the association between HFpEF and TMAO, Zengxiang Dong et al. showed that the plasma levels of the microbial metabolite were 6.84 μmol per L in the HFpEF group and 1.63 μmol per L in the control group. This result showed that TMAO is significantly associated with HFpEF [64]. Moreover, pre-clinical studies showed that dietary supplementation of TMAO, in a generated mouse model of pressure overload-induced HF, aggravated myocardial fibrosis and left ventricular remodelling and dysfunction. Elevated levels of TMAO, therefore, could be associated with worsening of left ventricular diastolic function, an increased left atrial volume, thus impaired tissue mechanics which is a key feature of HFpEF [51].
Anyway, the exact role of TMAO on HFpEF is unclear. TMAO is a marker of HF as serum levels are higher in patients with HF than those without HF, but importantly, it does not discriminate HFpEF from HFrEF. Thus, TMAO cannot be considered a unique marker of HFpEF but can help identify high-risk patients when associated with other markers [65]. Acute HF is another important HF setting, in which TMAO is essential as a prognostic marker; in a study by Suzuki et al. the detection of elevated TMAO levels in patients admitted to the hospital for acute HF correlated with an increase in rehospitalization at 1 year and mortality. Moreover, high TMAO further strengthened the prediction of death or HF at 1 year when combined with another important HF marker, NT-proBNP. Patients with elevated levels in both markers were at the highest risk of death or rehospitalization for HF at 1 year [66]. In two different studies, Kinugasa et al. and Salzano et al. showed that elevated TMAO levels at discharge are associated with an increased risk of post-discharge cardiac events in patients hospitalized with acute HF and HFpEF, especially in those with low nutritional status [67,68].
Since the effect of TMAO on prognosis may interact with the patient’s nutritional status, the prognostic effect of TMAO may depend on the phenotype of HFpEF. Furthermore, it has been shown that racial and geographic differences could affect TMAO values and their effects on prognosis in HF [69,70].
We can conclude that TMAO plays an important prognostic role in HF and all its settings, but further studies are needed to validate its use in common clinical practice.
7. Gut microbiota analysis in clinical practice
In the last decade, a strong association between changes in microbiota composition and several host diseases has been found [71]. Especially, dysbiosis plays an important pathophysiological role in cardio-metabolic diseases [72].
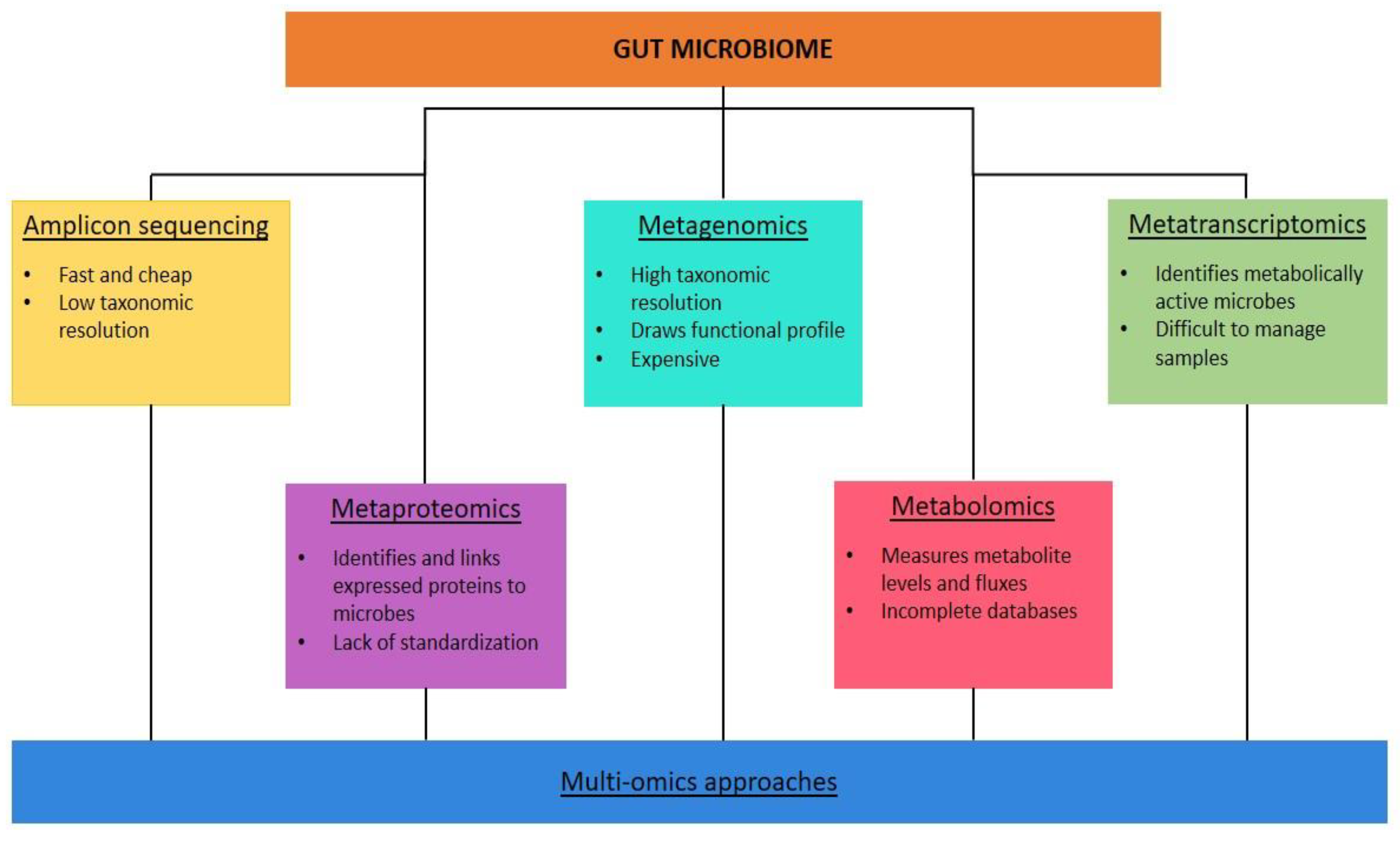
Advances in DNA sequencing technologies favoured the study of the microbiota and its contribution to cardiometabolic risk [73]. Meta-omics techniques characterise microbial communities, allowing us to perceive their influence on human physiology beyond simply profiling the taxonomic composition [74,75]. They are high-throughput sequencing (HTS) techniques performed at the microbe, DNA, and mRNA levels based on available samples and research goals [76]. The potential biological samples can be various, but the stool samples are the most used [77].
Sequencing and amplification of the marker gene (amplicon) is performed using a PCR primer specific for a particular hypervariable region of a microbial gene [71]. It uses 16S ribosomal ribonucleic acid (rDNA) gene for prokaryotes and 18S rDNA gene and internal transcribed spacers for eukaryotes, where the conserved and variable regions exist [76,78]. Therefore phylogenetic, or taxonomic profiles are produced using large public datasets for comparison [74]. The amplicon sequencing is the most used, fastest, and cheapest technique for samples with low biomass or host DNA contamination. Still, it provides low-resolution taxonomic information (only at the genus level) without information on the functional potential of the microbiota which can only be speculated [74,75]. Furthermore, bias may occur during amplification because it is sensitive to the specific primers and the number of PCR cycles chosen [73]. Therefore, the technique is often used in pioneering research [76].
Metagenomics uses short reads of DNA sequences and captures all DNA in the sample, including viral and eukaryotic DNA. Reference libraries and datasets give more detailed genomic information and taxonomic resolution at the species or strain level [73]. Furthermore, it draws a functional profile at the genetic level through bioinformatic analysis of metagenomic sequences [74,76]. Metagenomics is useful for profiling the entire genetic repertoire of a study group and in viral ecology where there aren’t specific marker genes to amplify. It could also reveal the interactions and mechanisms between the gut microbiota and disorders [78]. However, it is expensive and time-consuming, and it needs a much larger average genome size than 16S rDNA analysis and quality control protocols to remove human DNA contamination [74].
Meta-transcriptomics analyse gene expression through reverse transcription of RNA isolated from the sample [74]. It provides information on the microbiota's active functional output, discriminating metabolically active microbes from dormant or dead ones and extracellular DNA [73]. It also evaluates changes in microbial gene expression over time [74,75] and in response to external stimuli [73]. However, the sample collection, storage, and preparation are difficult to manage [74], the host mRNA contamination and rRNA must be removed and the data are biased for organisms with high transcription rates [73].
Metaproteomics can identify and quantify expressed proteins in a sample using higher-resolution mass spectrometry and quantitative proteomics techniques. Furthermore, it can link proteins to the microorganisms that encode them using databases and bioinformatics tools [74]. The limitations are the lack of standardization of the method [75]. Metabolomics measures metabolite levels and fluxes using mass spectroscopy and nuclear magnetic resonance [74]. It provides a readout of the host-microbiota interface using a range of known metabolites (targeted search, with higher sensitivity and semi-absolute quantifications), or covering as many metabolites as possible (untargeted search, harder to interpret) [75]. Incomplete databases and the difficulty of distinguishing metabolites from microbiota and host origin limit its application [74].
8. Therapeutic perspectives for CVD and HF
Data have been collected demonstrating that several therapeutic options based on the re-balancing of dysbiosis involving gut and liver microbiota may be helpful in better intervention strategies in counteracting metabolic disorders including atherosclerotic vascular disease states (ATVD). On the other hand, due to the huge correlation existing between microbiota modifications and the development of atherosclerosis and hypercholesterolemia, the effects of drugs commonly used for the treatment of these conditions may be affected [79]. Recently, gut microbiota alterations have also been shown to be involved in the pathogenesis and pathophysiology of HF and its comorbidities [80,81]. HF still represents a major clinical problem worldwide regarding mortality, hospitalization, and public health [82]. In the last decade, many advances have been made regarding the clinical approach to HF [83,84]. However, several unmet needs are still occurring, especially concerning the treatment of the early stages of the disease, mostly in the area of HF with HFpEF, in which an optimal therapeutic approach is still to be identified. Therefore, gut microbiota and its metabolites may represent an alternative therapeutic target to approach HFpEF.
The mechanisms leading to gut microbiota-related changes in cardiac performance are still to be clarified. However, evidence shows that oxidative damage and inflammation after the modification of gut microbiota seem to play a major role [85]. Subsequently, modulation of inflammatory states [86] and counteracting oxidative damage due to metabolic imbalance appear to represent promising solutions in approaching the treatment of HFpEF.
Recently, nutritional interventions have been found to affect the composition of gut microbiota [87] significantly. In particular, many studies have shown that in patients undergoing HF, a diet characterised by a high fibre intake and short-chain fatty acids is associated with reduced gut dysbacteriosis [87]. Merques et al, demonstrated that a diet characterised by a low fibre intake is associated with negative cardiac remodelling that leads to hypertension and the progression of cardiac fibrosis [88]. Probiotics may represent an additional therapeutic strategy to prevent cardiovascular disease. In fact, it is known that probiotics are microorganisms whose ingestion and consequent interaction with gut flora may result in several benefits in terms of myocardial protection and leading anti-inflammatory response [89].
However, in this context, the limitation is represented by the fact that most of the data obtained on probiotics in treating CV disease, particularly of HF, are derived from animal studies [90]. GutHeart is a multicentre prospective randomized open-label trial in which the authors randomized HF patients with left ventricular ejection fraction (LVEF) <40% and NYHA class II-III, to treatment (1:1:1) with Saccharomyces boulardii, the antibiotic rifaximin, or standard of care only. The results at three months have not shown the difference in terms of LVEF, microbiota composition, laboratory biomarker and functional assessment [91].
Gut Microbiota enzymes, which promote negative CV effects, may be the subject of the therapeutic target. In particular, TMAO levels correlate with markers of inflammation, endothelial dysfunction in HF and diastolic impairment [92]. Therefore, targeting the molecular reactions leading to the increase in circulating TMAO, for example, the conversion of choline to TMAO, may be a valid plan of action to reduce the negative effects of HF and its comorbidities [93]. Finally, faecal transplantation may be the last therapeutic strategy, although still, no data are available regarding its application outside the treatment of Clostridium difficile infection [94].
9. Conclusions
It is increasingly evident that gut microbiota plays a role in the pathogenesis of many disorders, including CV disease and CHF. Therefore, a more detailed investigation of the link between the gut microbiota and the CV system is necessary. The common drugs that we routinely use for the treatment of HF, have revolutionized the course of patients with extremely compromising situations. Still, these have proved insufficient to control the early stage of HF. Scientific evidence suggests that intervening in comorbidities and lifestyle modification is the only and most important way to impact the disease positively and precociously. In this context, therapeutic intervention on dysbiosis of gut microbiota could represent a new and innovative instrument to improve the clinical course of HF. Further randomized clinical trials are mandatory to define the correlation between gut microbiota and CHF and to indicate the most appropriate therapeutic approach.
Author Contributions
Conceptualization, M.B., F.B. and M.F.; writing—original draft preparation, M.B., F.R.P., L.B., S.L., D. L., S.M., R.M., D.M., D.S.; writing—review and editing, M.B., M.F. and F.B.; figures, M.B, S.L., F.R.P.; supervision, M.F. and F.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.R.; Pop, M.; DeBoy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic Analysis of the Human Distal Gut Microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef] [PubMed]
- Brestoff, J.R.; Artis, D. Commensal bacteria at the interface of host metabolism and the immune system. Nat. Immunol. 2013, 14, 676–684. [Google Scholar] [CrossRef]
- Backhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Science 307, 1915 (2005).
- Khosravi, A.; Mazmanian, S.K. Disruption of the gut microbiome as a risk factor for microbial infections. Curr. Opin. Microbiol. 2013, 16, 221–227. [Google Scholar] [CrossRef]
- Laterza, L.; Rizzatti, G.; Gaetani, E.; Chiusolo, P.; Gasbarrini, A. The gut microbiota and immune system relationship in human graft-versus-host disease. Mediterr. J. Hematol. Infect. Dis. 2016, 8, e2016025. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Odamaki, T.; Kato, K.; Sugahara, H.; Hashikura, N.; Takahashi, S.; Xiao, J.-Z.; Abe, F.; Osawa, R. Age-related changes in gut microbiota composition from newborn to centenarian: A cross-sectional study. BMC Microbiol. 2016, 16, 90. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Nageshwar Reddy, D. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787–8803. [Google Scholar] [CrossRef]
- Nagpal, R.; Tsuji, H.; Takahashi, T.; Nomoto, K.; Kawashima, K.; Nagata, S.; Yamashiro, Y. Ontogenesis of the Gut Microbiota Composition in Healthy, Full-Term, Vaginally Born and Breast-Fed Infants over the First 3 Years of Life: A Quantitative Bird’s-Eye View. Front. Microbiol. 2017, 8, 1388. [Google Scholar] [CrossRef]
- Wen, L.; Duffy, A. Factors Influencing the Gut Microbiota, Inflammation, and Type 2 Diabetes. J. Nutr. 2017, 147, 1468S–1475S. [Google Scholar] [CrossRef]
- Goodrich, J.K.; Waters, J.L.; Poole, A.C.; Sutter, J.L.; Koren, O.; Blekhman, R.; Beaumont, M.; Van Treuren, W.; Knight, R.; Bell, J.T.; et al. Human Genetics Shape the Gut Microbiome. Cell 2014, 159, 789–799. [Google Scholar] [CrossRef]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef]
- Bien, J.; Palagani, V.; Bozko, P. The intestinal microbiota dysbiosis and Clostridium difficile infection: Is there a relationship with inflammatory bowel disease? Ther. Adv. Gastroenterol. 2013, 6, 53–68. [Google Scholar] [CrossRef]
- Nishino, K.; Nishida, A.; Inoue, R.; Kawada, Y.; Ohno, M.; Sakai, S.; Inatomi, O.; Bamba, S.; Sugimoto, M.; Kawahara, M.; et al. Analysis of endoscopic brush samples identified mucosa-associated dysbiosis in inflammatory bowel disease. J. Gastroenterol. 2018, 53, 95–106. [Google Scholar] [CrossRef]
- Bunyavanich, S.; Shen, N.; Grishin, A.; Wood, R.; Burks, W.; Dawson, P.; Jones, S.M.; Leung, D.Y.; Sampson, H.; Sicherer, S.; et al. Early-life gut microbiome composition and milk allergy resolution. J. Allergy Clin. Immunol. 2016, 138, 1122–1130. [Google Scholar] [CrossRef]
- Chu, D.M.; Ma, J.; Prince, A.L.; Antony, K.M.; Seferovic, M.D.; Aagaard, K.M. Maturation of the infant microbiome community structure and function across multiple body sites and in relation to mode of delivery. Nat. Med. 2017, 23, 314–326. [Google Scholar] [CrossRef]
- Cui, L.; Zhao, T.; Hu, H.; Zhang, W.; Hua, X. Association Study of Gut Flora in Coronary Heart Disease through High-Throughput Sequencing. BioMed Res. Int. 2017, 2017, 3796359. [Google Scholar] [CrossRef]
- Jie, Z.; Xia, H.; Zhong, S.-L.; Feng, Q.; Li, S.; Liang, S.; Zhong, H.; Liu, Z.; Gao, Y.; Zhao, H.; et al. The gut microbiome in atherosclerotic cardiovascular disease. Nat. Commun. 2017, 8, 845. [Google Scholar] [CrossRef]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; et al. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef]
- Kazemian, N.; Mahmoudi, M.; Halperin, F.; Wu, J.C.; Pakpour, S. Gut microbiota and cardiovascular disease: Opportunities and challenges. Microbiome 2020, 8, 36. [Google Scholar] [CrossRef]
- Barrows, I.R.; Ramezani, A.; Raj, D.S. Inflammation, Immunity, and Oxidative Stress in Hypertension—Partners in Crime? Adv. Chronic Kidney Dis. 2019, 26, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, A.L.; Bäckhed, F. Role of gut microbiota in atherosclerosis. Nat. Rev. Cardiol. 2017, 14, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Heianza, Y.; Masi, W.; Manson, J.E.; Rexrode, K.M.; Qi, L. Gut Microbiota Metabolites and Risk of Major Adverse Cardiovascular Disease Events and Death: A Systematic Review and Meta-Analysis of Prospective Studies. J. Am. Heart Assoc. 2017, 6, e004947. [Google Scholar] [CrossRef]
- Schiattarella, G.G.; Sannino, A.; Toscano, E.; Giugliano, G.; Gargiulo, G.; Franzone, A.; Trimarco, B.; Esposito, G.; Perrino, C. Gut microbe-generated metabolite trimethylamine-N-oxide as cardiovascular risk biomarker: A systematic review and dose-response meta-analysis. Eur. Heart J. 2017, 38, 2948–2956. [Google Scholar] [CrossRef] [PubMed]
- Ramezani, A.; Nolin, T.D.; Barrows, I.R.; Serrano, M.G.; Buck, G.A.; Regunathan-Shenk, R.; West, R.E.; Latham, P.S.; Amdur, R.; Raj, D.S. Gut Colonization with Methanogenic Archaea Lowers Plasma Trimethylamine N-oxide Concentrations in Apolipoprotein e−/− Mice. Sci. Rep. 2018, 8, 14752. [Google Scholar] [CrossRef]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef]
- Oniszczuk, A.; Oniszczuk, T.; Gancarz, M.; Szymańska, J. Role of Gut Microbiota, Probiotics and Prebiotics in the Cardiovascular Diseases. Molecules 2021, 26, 1172. [Google Scholar] [CrossRef]
- Cimmino, G.; Muscoli, S.; De Rosa, S.; Cesaro, A.; Perrone, M.A.; Selvaggio, S.; Selvaggio, G.; Aimo, A.; Pedrinelli, R.; Mercuro, G.; et al. Evolving concepts in the pathophysiology of atherosclerosis: From endothelial dysfunction to thrombus formation through multiple shades of inflammation. J. Cardiovasc. Med. 2023, 24 (Suppl. 2), e156–e167. [Google Scholar] [CrossRef]
- Monteleone, I.; Muscoli, S.; Terribili, N.; Zorzi, F.; Mariano, E.; Mehta, J.L.; Pallone, F.; Monteleone, G.; Romeo, F. Local immune activity in acute coronary syndrome: oxLDL abrogates LPS-tolerance in mononuclear cells isolated from culprit lesion. Int. J. Cardiol. 2013, 169, 44–51. [Google Scholar] [CrossRef]
- Human Microbiome Project, C. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef]
- Ott, S.J.; El Mokhtari, N.E.; Musfeldt, M.; Hellmig, S.; Freitag, S.; Rehman, A.; Kühbacher, T.; Nikolaus, S.; Namsolleck, P.; Blaut, M.; et al. Detection of Diverse Bacterial Signatures in Atherosclerotic Lesions of Patients With Coronary Heart Disease. Circulation 2006, 113, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Koren, O.; Spor, A.; Felin, J.; Fak, F.; Stombaugh, J.; Tremaroli, V.; et al. Human oral, gut, and plaque microbiota in patients with atherosclerosis. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4592–4598. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The Role of Short-Chain Fatty Acids in Health and Disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar] [CrossRef] [PubMed]
- Brandsma, E.; Kloosterhuis, N.J.; Koster, M.; Dekker, D.C.; Gijbels, M.J.; van der Velden, S.; Ríos-Morales, M.; van Faassen, M.J.; Loreti, M.G.; de Bruin, A.; et al. A Proinflammatory Gut Microbiota Increases Systemic Inflammation and Accelerates Atherosclerosis. Circ. Res. 2019, 124, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Just, S.; Mondot, S.; Ecker, J.; Wegner, K.; Rath, E.; Gau, L.; Streidl, T.; Hery-Arnaud, G.; Schmidt, S.; Lesker, T.R.; et al. The gut microbiota drives the impact of bile acids and fat source in diet on mouse metabolism. Microbiome 2018, 6, 134. [Google Scholar] [CrossRef] [PubMed]
- Janeiro, M.H.; Ramírez, M.J.; Milagro, F.I.; Martínez, J.A.; Solas, M. Implication of Trimethylamine N-Oxide (TMAO) in Disease: Potential Biomarker or New Therapeutic Target. Nutrients 2018, 10, 1398. [Google Scholar] [CrossRef]
- Nagatomo, Y.; Tang, W.H.W. Intersections Between Microbiome and Heart Failure: Revisiting the Gut Hypothesis. J. Card. Fail. 2015, 21, 973–980. [Google Scholar] [CrossRef]
- Krack, A.; Sharma, R.; Figulla, H.R.; Anker, S.D. The importance of the gastrointestinal system in the pathogenesis of heart failure. Eur. Heart J. 2005, 26, 2368–2374. [Google Scholar] [CrossRef]
- Pasini, E.; Aquilani, R.; Testa, C.; Baiardi, P.; Angioletti, S.; Boschi, F.; Verri, M.; Dioguardi, F. Pathogenic Gut Flora in Patients With Chronic Heart Failure. JACC Heart Fail. 2016, 4, 220–227. [Google Scholar] [CrossRef]
- Sandek, A.; Bauditz, J.; Swidsinski, A.; Buhner, S.; Weber-Eibel, J.; von Haehling, S.; Schroedl, W.; Karhausen, T.; Doehner, W.; Rauchhaus, M.; et al. Altered Intestinal Function in Patients With Chronic Heart Failure. J. Am. Coll. Cardiol. 2007, 50, 1561–1569. [Google Scholar] [CrossRef]
- Sedman, P.C.; Macfie, J.; Sagar, P.; Mitchell, C.J.; May, J.; Mancey-Jones, B.; Johnstone, D. The prevalence of gut translocation in humans. Gastroenterology 1994, 107, 643–649. [Google Scholar] [CrossRef]
- Anker, S.D.; Egerer, K.R.; Volk, H.-D.; Kox, W.J.; Poole-Wilson, P.A.; Coats, A.J. Elevated soluble CD14 receptors and altered cytokines in chronic heart failure. Am. J. Cardiol. 1997, 79, 1426–1430. [Google Scholar] [CrossRef]
- Sandek, A.; Bjarnason, I.; Volk, H.-D.; Crane, R.; Meddings, J.B.; Niebauer, J.; Kalra, P.R.; Buhner, S.; Herrmann, R.; Springer, J.; et al. Studies on bacterial endotoxin and intestinal absorption function in patients with chronic heart failure. Int. J. Cardiol. 2012, 157, 80–85. [Google Scholar] [CrossRef]
- Niebauer, J.; Volk, H.D.; Kemp, M.; Dominguez, M.; Schumann, R.R.; Rauchhaus, M.; Poole-Wilson, P.A.; Coats, A.J.; Anker, S.D. Endotoxin and immune activation in chronic heart failure: A prospective cohort study. Lancet 1999, 353, 1838–1842. [Google Scholar] [CrossRef]
- Peschel, T.; Schönauer, M.; Thiele, H.; Anker, S.; Schuler, G.; Niebauer, J. Invasive assessment of bacterial endotoxin and inflammatory cytokines in patients with acute heart failure [published correction appears in Eur J Heart Fail. 2004 Mar 1;6(2):245]. Eur. J. Heart Fail. 2003, 5, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Conraads, V.M.; Jorens, P.G.; De Clerck, L.S.; Van Saene, H.K.; Ieven, M.M.; Bosmans, J.M.; Schuerwegh, A.; Bridts, C.H.; Wuyts, F.; Stevens, W.J.; et al. Selective intestinal decontamination in advanced chronic heart failure: A pilot trial. Eur. J. Heart Fail. 2004, 6, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.W.; Bäckhed, F.; Landmesser, U.; Hazen, S.L. Intestinal Microbiota in Cardiovascular Health and Disease: JACC state-of-the-art review. J. Am. Coll. Cardiol. 2019, 73, 2089–2105. [Google Scholar] [CrossRef]
- Zhu, W.; Buffa, J.A.; Wang, Z.; Warrier, M.; Schugar, R.; Shih, D.M.; Gupta, N.; Gregory, J.C.; Org, E.; Fu, X.; et al. Flavin monooxygenase 3, the host hepatic enzyme in the metaorganismal trimethylamine N-oxide-generating pathway, modulates platelet responsiveness and thrombosis risk. J. Thromb. Haemost. 2018, 16, 1857–1872. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wu, Z.; Yan, J.; Liu, H.; Liu, Q.; Deng, Y.; Ou, C.; Chen, M. Gut microbe-derived metabolite trimethylamine N-oxide induces cardiac hypertrophy and fibrosis. Lab. Investig. 2019, 99, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Organ, C.L.; Otsuka, H.; Bhushan, S.; Wang, Z.; Bradley, J.; Trivedi, R.; Polhemus, D.J.; Tang, W.W.; Wu, Y.; Hazen, S.L.; et al. Choline Diet and Its Gut Microbe–Derived Metabolite, Trimethylamine N-Oxide, Exacerbate Pressure Overload–Induced Heart Failure. Circ. Heart Fail. 2016, 9, e002314. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Kong, B.; Shuai, W.; Fu, H.; Jiang, X.; Huang, H. 3,3-Dimethyl-1-butanol attenuates cardiac remodeling in pressure-overload-induced heart failure mice. J. Nutr. Biochem. 2020, 78, 108341. [Google Scholar] [CrossRef]
- Sun, X.; Jiao, X.; Ma, Y.; Liu, Y.; Zhang, L.; He, Y.; Chen, Y. Trimethylamine N-oxide induces inflammation and endothelial dysfunction in human umbilical vein endothelial cells via activating ROS-TXNIP-NLRP3 inflammasome. Biochem. Biophys. Res. Commun. 2016, 481, 63–70. [Google Scholar] [CrossRef]
- Zhang, H.; Meng, J.; Yu, H. Trimethylamine N-oxide Supplementation Abolishes the Cardioprotective Effects of Voluntary Exercise in Mice Fed a Western Diet. Front. Physiol. 2017, 8, 944. [Google Scholar] [CrossRef]
- Ma, G.; Pan, B.; Chen, Y.; Guo, C.; Zhao, M.; Zheng, L.; Chen, B. Trimethylamine N-oxide in atherogenesis: Impairing endothelial self-repair capacity and enhancing monocyte adhesion. Biosci. Rep. 2017, 37, BSR20160244. [Google Scholar] [CrossRef] [PubMed]
- Makrecka-Kuka, M.; Volska, K.; Antone, U.; Vilskersts, R.; Grinberga, S.; Bandere, D.; Liepinsh, E.; Dambrova, M. Trimethylamine N-oxide impairs pyruvate and fatty acid oxidation in cardiac mitochondria. Toxicol. Lett. 2017, 267, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.W.; Wang, Z.; Kennedy, D.J.; Wu, Y.; Buffa, J.A.; Agatisa-Boyle, B.; Li, X.S.; Levison, B.S.; Hazen, S.L.; X, Z.; et al. Gut Microbiota-Dependent Trimethylamine N -Oxide (TMAO) Pathway Contributes to Both Development of Renal Insufficiency and Mortality Risk in Chronic Kidney Disease. Circ. Res. 2015, 116, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Gregory, J.C.; Org, E.; Buffa, J.A.; Gupta, N.; Wang, Z.; Li, L.; Fu, X.; Wu, Y.; Mehrabian, M.; et al. Gut Microbial Metabolite TMAO Enhances Platelet Hyperreactivity and Thrombosis Risk. Cell 2016, 165, 111–124. [Google Scholar] [CrossRef]
- Tang, W.W.; Wang, Z.; Fan, Y.; Levison, B.; Hazen, J.E.; Donahue, L.M.; Wu, Y.; Hazen, S.L. Prognostic Value of Elevated Levels of Intestinal Microbe-Generated Metabolite Trimethylamine-N-Oxide in Patients With Heart Failure: Refining the gut hypothesis. J. Am. Coll. Cardiol. 2014, 64, 1908–1914. [Google Scholar] [CrossRef]
- Polsinelli, V.B.; Marteau, L.; Shah, S.J. The role of splanchnic congestion and the intestinal microenvironment in the pathogenesis of advanced heart failure. Curr. Opin. Support. Palliat. Care 2019, 13, 24–30. [Google Scholar] [CrossRef]
- Schuett, K.; Kleber, M.E.; Scharnagl, H.; Lorkowski, S.; März, W.; Niessner, A.; Marx, N.; Meinitzer, A. Trimethylamine-N-oxide and Heart Failure With Reduced Versus Preserved Ejection Fraction. J. Am. Coll. Cardiol. 2017, 70, 3202–3204. [Google Scholar] [CrossRef]
- Li, X.; Fan, Z.; Cui, J.; Li, D.; Lu, J.; Cui, X.; Xie, L.; Wu, Y.; Lin, Q.; Li, Y. Trimethylamine N-Oxide in Heart Failure: A Meta-Analysis of Prognostic Value. Front. Cardiovasc. Med. 2022, 9, 817396. [Google Scholar] [CrossRef] [PubMed]
- Trøseid, M.; Ueland, T.; Hov, J.R.; Svardal, A.; Gregersen, I.; Dahl, C.P.; Aakhus, S.; Gude, E.; Bjørndal, B.; Halvorsen, B.; et al. Microbiota-dependent metabolite trimethylamine-N-oxide is associated with disease severity and survival of patients with chronic heart failure. J. Intern. Med. 2014, 277, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Zheng, S.; Shen, Z.; Luo, Y.; Hai, X. Trimethylamine N-Oxide is Associated with Heart Failure Risk in Patients with Preserved Ejection Fraction. Lab. Med. 2021, 52, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Gui, X.Y.; Rabkin, S.W. C-Reactive Protein, Interleukin-6, Trimethylamine-N-Oxide, Syndecan-1, Nitric Oxide, and Tumor Necrosis Factor Receptor-1 in Heart Failure with Preserved Versus Reduced Ejection Fraction: A Meta-Analysis. Curr. Heart Fail. Rep. 2023, 20, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Heaney, L.M.; Bhandari, S.S.; Jones, D.J.L.; Ng, L.L. TrimethylamineN-oxide and prognosis in acute heart failure. Heart 2016, 102, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Kinugasa, Y.; Nakamura, K.; Kamitani, H.; Hirai, M.; Yanagihara, K.; Kato, M.; Yamamoto, K. Trimethylamine N-oxide and outcomes in patients hospitalized with acute heart failure and preserved ejection fraction. ESC Heart Fail. 2021, 8, 2103–2110. [Google Scholar] [CrossRef]
- Salzano, A.; Israr, M.Z.; Yazaki, Y.; Heaney, L.M.; Kanagala, P.; Singh, A.; Arnold, J.R.; Gulsin, G.S.; Squire, I.B.; McCann, G.P.; Ng, L.L.; Suzuki, T. Combined use of trimethylamine N-oxide with BNP for risk stratification in heart failure with preserved ejection fraction: Findings from the DIAMOND-HFpEF study. Eur J Prev Cardiol 2020; 27: 2159–2162.
- Yazaki, Y.; Aizawa, K.; Israr, M.Z.; Negishi, K.; Salzano, A.; Saitoh, Y.; Kimura, N.; Kono, K.; Heaney, L.; Cassambai, S.; et al. Ethnic differences in association of outcomes with trimethylamine N-oxide in acute heart failure patients. ESC Heart Fail. 2020, 7, 2373–2378. [Google Scholar] [CrossRef]
- Yazaki, Y.; Salzano, A.; Nelson, C.P.; Voors, A.A.; Anker, S.D.; Cleland, J.G.; Lang, C.C.; Metra, M.; Samani, N.J.; Ng, L.L.; et al. Geographical location affects the levels and association of trimethylamine N-oxide with heart failure mortality in BIOSTAT-CHF: A post-hoc analysis. Eur. J. Heart Fail. 2019, 21, 1291–1294. [Google Scholar] [CrossRef]
- Longo, S.; Rizza, S.; Federici, M. Microbiota-gut-brain axis: Relationships among the vagus nerve, gut microbiota, obesity, and diabetes. Acta Diabetol. 2023, 60, 1007–1017. [Google Scholar] [CrossRef]
- Longo, S.; Legramante, J.M.; Rizza, S.; Federici, M. Vasovagal syncope: An overview of pathophysiological mechanisms. Eur. J. Intern. Med. 2023, 112, 6–14. [Google Scholar] [CrossRef]
- Knight, R.; Vrbanac, A.; Taylor, B.C.; Aksenov, A.; Callewaert, C.; Debelius, J.; Gonzalez, A.; Kosciolek, T.; McCall, L.-I.; McDonald, D.; et al. Best practices for analysing microbiomes. Nat. Rev. Microbiol. 2018, 16, 410–422. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yang, Y. Gut microbiome and its meta-omics perspectives: Profound implications for cardiovascular diseases. Gut Microbes 2021, 13, 1936379. [Google Scholar] [CrossRef] [PubMed]
- Valles-Colomer, M.; Menni, C.; Berry, S.E.; Valdes, A.M.; Spector, T.D.; Segata, N. Cardiometabolic health, diet and the gut microbiome: A meta-omics perspective. Nat. Med. 2023, 29, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-X.; Qin, Y.; Chen, T.; Lu, M.; Qian, X.; Guo, X.; Bai, Y. A practical guide to amplicon and metagenomic analysis of microbiome data. Protein Cell 2021, 12, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.X.; Wang, S.-Y.; Kuo, C.-H.; Tsai, I.-L. Metabolome analysis for investigating host-gut microbiota interactions. J. Formos. Med Assoc. 2019, 118 (Suppl. 1), S10–S22. [Google Scholar] [CrossRef]
- Whon, T.W.; Shin, N.-R.; Kim, J.Y.; Roh, S.W. Omics in gut microbiome analysis. J. Microbiol. 2021, 59, 292–297. [Google Scholar] [CrossRef]
- Tuteja, S.; Ferguson, J.F. Gut Microbiome and Response to Cardiovascular Drugs. Circ. Genom. Precis. Med. 2019, 12, 421–429. [Google Scholar] [CrossRef]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef]
- Sandek, A.; Swidsinski, A.; Schroedl, W.; Watson, A.; Valentova, M.; Herrmann, R.; Scherbakov, N.; Cramer, L.; Rauchhaus, M.; Grosse-Herrenthey, A.; et al. Intestinal Blood Flow in Patients With Chronic Heart Failure: A Link With Bacterial Growth, Gastrointestinal Symptoms, and Cachexia. J. Am. Coll. Cardiol. 2014, 64, 1092–1102. [Google Scholar] [CrossRef]
- Roger, V.L. Epidemiology of Heart Failure. A Contemporary Perspective. Circ. Res. 2021, 128, 1421–1434. [Google Scholar] [CrossRef]
- McMurray, J.J.V.; Packer, M.; Desai, A.S.; et al. "Angiotensin-neprilysin inhibition versus enalapril in heart failure". N Engl JMed2014;371:993-1004.
- Solomon, S.D.; McMurray, J.J.V.; Anand, I.S.; Junbo Ge, D.P.; Lam, C.S.P.; Maggioni, A.P.; Martinez, F.; Packer, M.; Pfeffer, M.A.; Pieske, B.; et al. Angiotensin–Neprilysin Inhibition in Heart Failure with Preserved Ejection Fraction. N. Engl. J. Med. 2019, 381, 1609–1620. [Google Scholar] [CrossRef] [PubMed]
- Finamore, A.; Ambra, R.; Nobili, F.; Garaguso, I.; Raguzzini, A.; Serafini, M. Redox role of Lactobacillus casei Shirota against the cellular damage induced by 2, 2’-azobis (2-amidinopropane) dihydrochloride-induced oxidative and inflammatory stress in enterocytes-like epithelial cells. Front. Immunol. 2018, 9, 1131. [Google Scholar] [CrossRef] [PubMed]
- Pasini, E.; Aquilani, R.; Testa, C.; Baiardi, P.; Angioletti, S.; Boschi, F.; Verri, M.; Dioguardi, F. Pathogenic Gut Flora in Patients With Chronic Heart Failure. JACC Heart Fail. 2016, 4, 220–227. [Google Scholar] [CrossRef]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Muller, A.; Young, V.B.; et al. A dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell 2016, 167, 1339–1353.e21. [Google Scholar] [CrossRef]
- Marques, F.Z.; Nelson, E.; Chu, P.-Y.; Horlock, D.; Fiedler, A.; Ziemann, M.; Tan, J.K.; Kuruppu, S.; Rajapakse, N.W.; El-Osta, A.; et al. High-Fiber Diet and Acetate Supplementation Change the Gut Microbiota and Prevent the Development of Hypertension and Heart Failure in Hypertensive Mice. Circulation 2017, 135, 964–977. [Google Scholar] [CrossRef] [PubMed]
- Gan, X.T.; Ettinger, G.; Huang, C.X.; Burton, J.P.; Haist, J.V.; Rajapurohitam, V.; Sidaway, J.E.; Martin, G.; Gloor, G.B.; Swann, J.R.; et al. Probiotic Administration Attenuates Myocardial Hypertrophy and Heart Failure After Myocardial Infarction in the Rat. Circ. Heart Fail. 2014, 7, 491–499. [Google Scholar] [CrossRef]
- Lam, V.; Su, J.; Koprowski, S.; Hsu, A.; Tweddell, J.S.; Rafiee, P.; Gross, G.J.; Salzman, N.H.; Baker, J.E. Intestinal microbiota determine severity of myocardial infarction in rats. FASEB J. 2012, 26, 1727–1735. [Google Scholar] [CrossRef]
- Awoyemi, A.; Mayerhofer, C.; Felix, A.S.; Hov, J.R.; Moscavitch, S.D.; Lappegård, K.T.; Hovland, A.; Halvorsen, S.; Halvorsen, B.; Gregersen, I.; et al. Rifaximin or Saccharomyces boulardii in heart failure with reduced ejection fraction: Results from the randomized GutHeart trial. EBioMedicine 2021, 70, 103511. [Google Scholar] [CrossRef]
- Tang, W.W.; Wang, Z.; Shrestha, K.; Borowski, A.G.; Wu, Y.; Troughton, R.W.; Klein, A.L.; Hazen, S.L. Intestinal Microbiota-Dependent Phosphatidylcholine Metabolites, Diastolic Dysfunction, and Adverse Clinical Outcomes in Chronic Systolic Heart Failure. J. Card. Fail. 2015, 21, 91–96. [Google Scholar] [CrossRef]
- Organ, C.L.; Li, Z.; Sharp, T.E., 3rd; Polhemus, D.J.; Gupta, N.; Goodchild, T.T.; Tang, W.H.W.; Hazen, S.L.; Lefer, D.J. Nonlethal Inhibition of Gut Microbial Trimethylamine N-oxide Production Improves Cardiac Function and Remodeling in a Murine Model of Heart Failure. J. Am. Heart Assoc. 2020, 9, e016223. [Google Scholar] [CrossRef]
- McDonald, L.C.; Gerding, D.N.; Johnson, S.; Bakken, J.S.; Carroll, K.C.; E Coffin, S.; Dubberke, E.R.; Garey, K.W.; Gould, C.V.; Kelly, C.; et al. Clinical Practice Guidelines for Clostridium difficile Infection in Adults and Children: 2017 Update by the Infectious Diseases Society of America (IDSA) and Society for Healthcare Epidemiology of America (SHEA). Clin. Infect. Dis. 2018, 66, 987–994. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The 3 mechanisms responsible for bacterial translocation and systemic inflammation. LPS= lipopolysaccharide.
Figure 1.
The 3 mechanisms responsible for bacterial translocation and systemic inflammation. LPS= lipopolysaccharide.
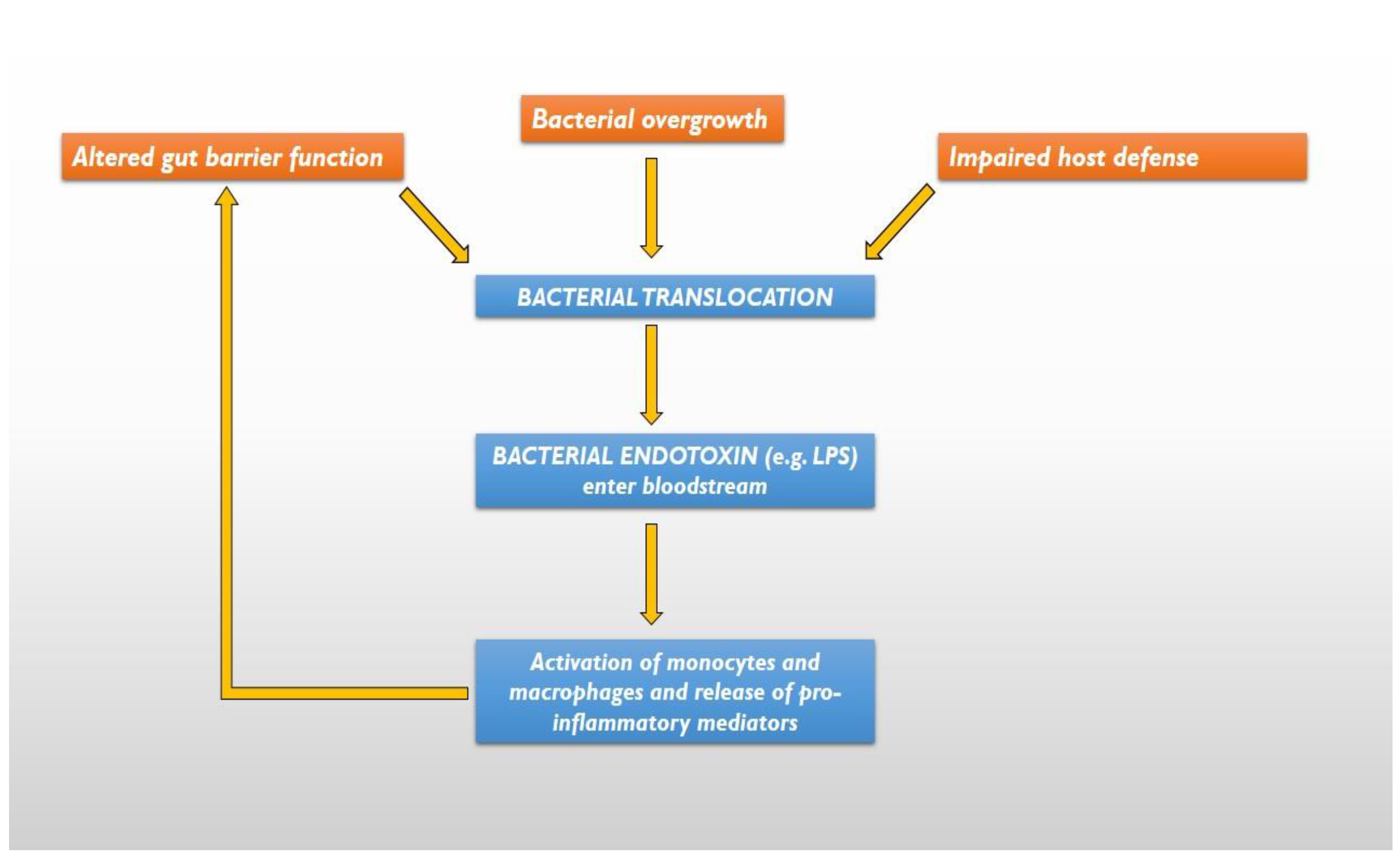
Figure 2.
TMA produced by the gut microbiota enters the circulatory system and is oxidized to TMAO by FMO3 in the liver. Circulating TMAO causes endothelial dysfunction, platelet hyperreactivity, heart failure and progression of renal disease.
Figure 2.
TMA produced by the gut microbiota enters the circulatory system and is oxidized to TMAO by FMO3 in the liver. Circulating TMAO causes endothelial dysfunction, platelet hyperreactivity, heart failure and progression of renal disease.
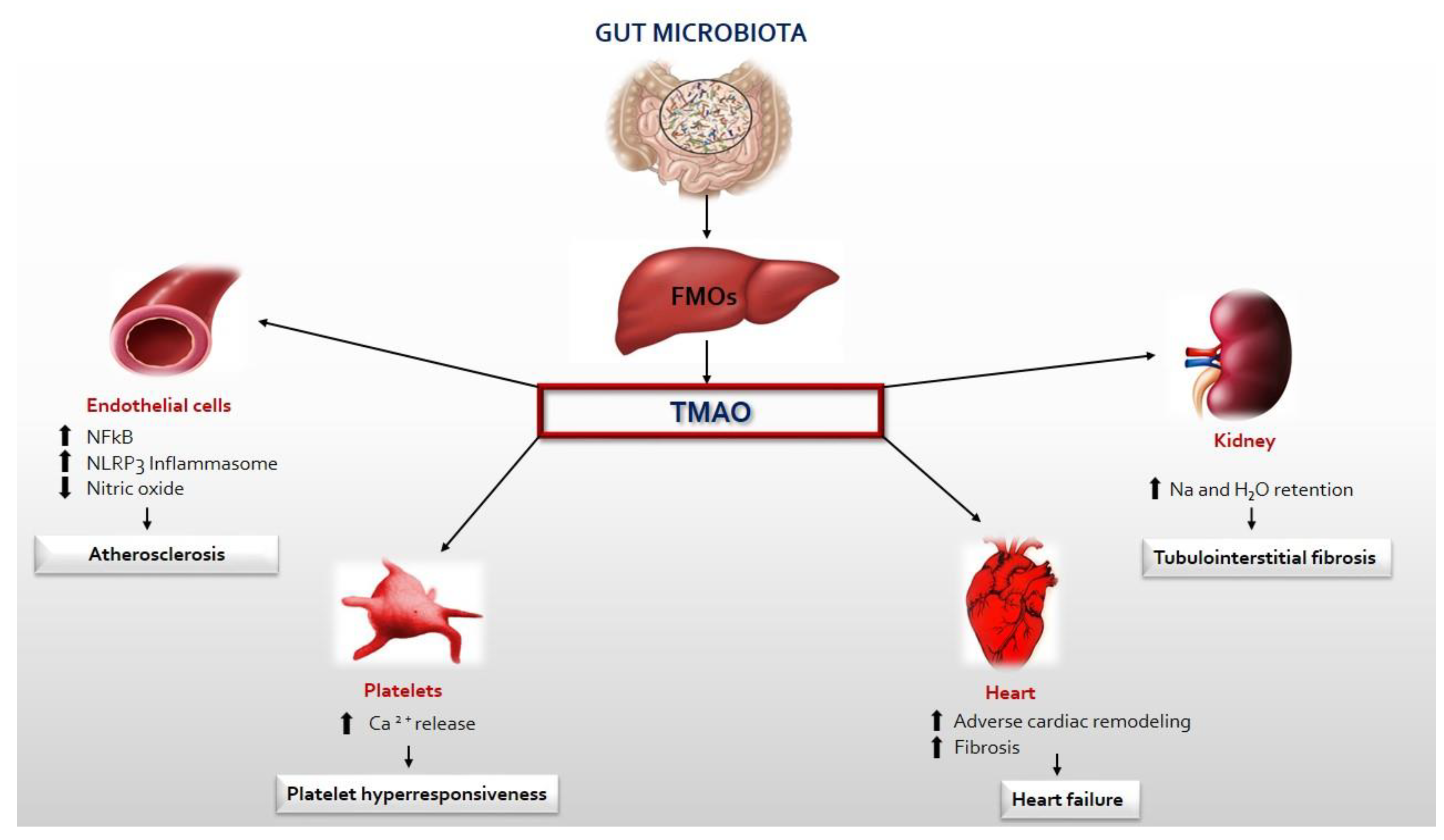
Figure 3.
Different high-throughput sequencing techniques performed at the level of microbes, DNA, mRNA, proteins and metabolites.
Figure 3.
Different high-throughput sequencing techniques performed at the level of microbes, DNA, mRNA, proteins and metabolites.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



