
Submitted:
25 June 2023
Posted:
26 June 2023
You are already at the latest version
Abstract
It is well known that the strong-evidence foodborne outbreaks of human campylobacteriosis are associated with the consumption of raw or incompletely thermally processed poultry meat, whereas broilers act as the main reservoir for Campylobacter species. Campylobacter jejuni and Campylobacter coli are the two main species of campylobacters detected in chicken meat, while they account for almost 90% of the reported cases of campylobacteriosis in humans. Over 80% of these cases are attributed to C. jejuni and about 10% of them are due to C. coli. Therefore, until recently the dominance of C. jejuni against all other Campylobacter spp. isolated from chicken meat samples was well established and unquestionable. Lately, however, C. coli has been increasingly recovered from chicken meat to such an extent that it is now evident that it often comprises the dominant species among the identified campylobacters in the meat samples. This work attempts for the first time a detailed review in the literature to deepen into this noteworthy epidemiological swift in the prevalence of C. jejuni and C. coli, along with the distribution of Campylobacter spp. in chicken meat. Factors such as the sampling method followed for screening campylobacters in broiler carcasses (e.g., swabs or carcass rinsates, skinned or skinless meat excised samples) and part of the animal carcass from which the sample is obtained (e.g., neck, breast, leg), seasonality of sampling (summer vs. winter) and environmental conditions (e.g., rainfall, relative humidity) at the farm level, the isolation procedure (enumeration or detection) and pathogen identification (biochemical or molecular), the enrichment and plating isolation media (e.g., Bolton vs. Preston broth, charcoal-based vs. chromogenic agars), as well as the biofilm-forming ability of different campylobacters, highlight the multivariate dimension of the phenomenon and are thoroughly discussed in the present review.
Keywords:
Campylobacter
; chicken
; epidemiology
; meat
; poultry
1. Introduction
According to the latest scientific report of the European Food Safety Authority (EFSA) and the European Centre for Disease Prevention and Control (ECDC) on the trends and sources of zoonoses and foodborne outbreaks in the European Union (EU), campylobacteriosis is the most commonly reported foodborne gastrointestinal infection in humans in the EU and has been so since 2007 [1]. Estimates of the overall human health impact of bacterial agents transmitted commonly through food, place Campylobacter as the first or second most common agent after nontyphoidal Salmonella in Europe, North America, Australia, and Japan [2,3,4]. The notable absence of notified Campylobacter outbreaks in China and some other populous countries, like India, could be attributed to the lack of mandatory surveillance by the established foodborne disease surveillance system or to the underreporting (e.g., mild symptoms and a smaller number of cases seeking health care) and/or underdiagnosis (e.g., lack of testing and diagnostic accuracy) of the disease, the lack of epidemiological surveillance data connecting causative agents of outbreaks, and the different dietary habits on those countries [5,6,7].
Major food categories of interest for Campylobacter occurrence include mostly meat and meat products (i.e., animal carcasses and fresh/ready-to-eat (RTE), cooked and fermented products), as well as milk and milk products (i.e., raw and pasteurized milk and dairy products including cheese) [1], although the prevalence of the disease-causing Campylobacter spp. is significantly increased in poultry meat samples compared to other types of meat or compared with milk and dairy products [8,9,10,11]. The strong-evidence foodborne association of campylobacteriosis outbreaks with the consumption of raw or incompletely thermally processed poultry meat is already well known and has been emphatically documented nowadays [8,10,12], while the foodborne illness due to the presence of Campylobacter spp. in poultry has been classified as the costliest pathogen-meat combination from an economic perspective [13]. Regardless the fact today poultry is considered the main reservoir for Campylobacter spp. (source of infection), latest epidemiological evidence suggests pathogen transmission to humans through a pathway implicating cattle as the primary reservoir of Campylobacter (source of contamination), infecting people via the fecal-oral route and the consumption of contaminated chickens [14].
World poultry meat consumption refers to the consumption of meat from chickens (broilers), turkey, and other avian species (e.g., ducks, geese). Available data compiled from the Food and Agriculture Organization (FAO) of the United Nations (UN) reveal an increase in worldwide annual poultry meat consumption per capita by more than 5.0 kg in the past 20 years; from 10.8 kg in 2000 to 16.2 kg in 2020 [15]. Chickens are by far the main protein source of animal origin for humans in terms of livestock animals reared and slaughtered for their meat [15], so the previous rates represent roughly the chicken meat being consumed on a global basis.
The different thermotolerant campylobacters validly described to date are summarized in Supplementary Table S1. Of these, Campylobacter jejuni and C. coli are the two most important species mainly detected in foods of animal origin [55], [56] (p. 1670). These two species account for almost 90% of the reported human campylobacteriosis cases, with over 80% of the occurring gastrointestinal infections being attributable to C. jejuni and the rest about 10% of infections attributed to C. coli [56] (pp. 1669–1670), [57]. Therefore, until recently it was well established and beyond any reasonable question that C. jejuni is the dominant species among all other Campylobacter spp. isolated from chicken meat samples. Lately, however, C. coli has been increasingly recovered from chicken samples to such an extent that it is now obvious it many times comprises the dominant species among the identified campylobacters in the meat samples [58,59,60,61,62]. To this end, in studies pertaining to the metropolitan area of Athens, Greece, and its suburbs in the Attica region, Andritsos et al. [63] reported isolation rates of 6% and 27% for C. jejuni and C. coli, respectively, during Campylobacter spp. detection in chicken meat samples, whereas the strict majority (87.5%) of the recovered campylobacters (16) from 830 fecal samples collected from five poultry farms by Marinou et al. [64] were identified as C. coli, without any of the strains being identified as C. jejuni whatsoever. Taking into account that in the latter case of Campylobacter presence in broilers’ litter, the positive predictive value in terms of microorganism’s occurrence in carcass skin samples is much greater, unless the pathogen cannot be detected in the intestinal content of the bird [65], C. coli dominance in the chicken flocks should be taken for granted.
Considering all the above, the present work attempts for the first time, to the best of the authors’ knowledge, a detailed review in the literature in order to elucidate the underlying epidemiological transition from C. jejuni to C. coli in chicken meat, along with the distribution of campylobacters in poultry. Figure 1 outlines the factors affecting the occurrence of Campylobacter spp. in poultry meat samples, while those factors are thoroughly being discussed below.
2. Sampling of Poultry
2.1. Carcass Sampling
Campylobacter which is originally associated with the bird’s feathers and contaminates the exterior of the animal might be transferred to the poultry skin during mechanical defeathering of broiler carcasses in the slaughterhouse [66]. Thus, the meat sampled with skin from poultry is more likely to contain the pathogen when compared to animal tissue samples obtained from broiler carcasses without the skin. Furthermore, due to the favorable conditions of humidity and temperature in the wings of poultry, there is a high Campylobacter load in the wings which could also be attributed to imperfect scalding, post-scalding contamination or a combination of both [67]. Hence, there is an increased prevalence of C. jejuni in the wings of sampled poultry [65,68,69,70]. Besides, an initial high Campylobacter contamination of the neck skin may occur when water excess drips down the carcass dragging along the bacteria during the slaughter line hanging (upside down) of carcasses [71]. This may also explain the comparatively lower number of campylobacters found on the back, breast and leg skin samples and other parts of the carcass than in the neck and wing skin samples (Figure 2). Moreover, breast and wing skin sites when sampled show a higher correlation in Campylobacter populations with the neck skin samples [71]. The variations in Campylobacter concentrations between skin sites of individual carcasses are often reflected in the non-homogeneous distribution of carcass contamination after post-chilling of broiler carcasses [71] (Figure 2).
2.2. Sampling Method, Type of Product and Refrigerated Storage
The method of sampling plays an important role in determining the microbiological quality of foods. Several non-destructive methods (e.g., surface swabs, contact plates, tissue excisions) are suitable for estimating bacterial populations anticipated in relatively high concentrations on the carcass, such as aerobic plate count (APC) and enterobacteria, while other –destructive in nature– methods (e.g., sampling/cutting of parts of the carcass, whole carcass rinses) are more suitable for detecting the presence of pathogens that may be more unevenly distributed and in low population on the carcass [72,73]. The former are fast and simple methods that can be used for comparative purposes, sanitation verification procedures, and shelf-life prediction, whereas the latter are time-consuming and labor-intensive methods preferable when the microbiological safety is of outmost importance. For instance, a sample area of at least 100 cm2 is swabbed for APC and Enterobacteriaceae enumeration in a cattle carcass after dressing but before chilling and a pooled neck skin sample is recommended for detecting Salmonella and enumerating Campylobacter in poultry carcasses after chilling [74,75]. In the case of broiler carcasses, counts for APC and hygiene indicators (total coliforms, Escherichia coli) from the microbiological analysis of swabs were generally more than 0.5 log units lower than the smallest values obtained through tissue excisions and carcass rinses [76].
The type of product or its preparation also affect Campylobacter incidence. Statistical analysis in a survey of Campylobacter spp. contamination in chicken meat preparations in Belgium indicated that the odds of Campylobacter presence are lower in minced meat than in portioned or cut meat and an unpredicted bias into prevalence and enumeration results could be plausible, unless proper sampling and a balanced selection of product types takes place [67]. Progressive increase of product’s surface through mincing, apart from increased levels of pathogen contamination, implies also increased microbial exposure to the air. For microaerophilic Campylobacter spp. this exposure to aerobic conditions could prove lethal since the degree of processing leads to an ever-increasing decrease in the number of campylobacters encountered in the products [55,68,77,78]. Finally, storage of product at refrigeration temperature may reduce Campylobacter count, as revealed by culturing methods during storage of refrigerated broiler breast and thigh meat at 4°C, packaged under aerobic, modified atmosphere packaging (MAP) or vacuum conditions [79,80]. Interestingly, the succession of Campylobacter species or strains, with a special emphasis on C. jejuni and C. coli, during refrigerated storage of artificially contaminated chicken meat packaged samples could be the subject of future research, as in the case of Listeria monocytogenes [81].
2.3. Seasonality of Sampling and Environmental Conditions
The seasonality of sampling and the effect environmental conditions may have on the recovery of Campylobacter spp. from broiler carcasses or chicken meat has been the subject of dozens of research papers, which present conflicting results as far as the effect of seasonality on Campylobacter prevalence is concerned and fail to reach an agreement on the matter. In this context, a large number of studies have indicated a significant seasonal increase in the prevalence of Campylobacter in broiler carcasses or in poultry meat during the hot summer months [10,68,82,83,84,85], even in tropical countries without marked seasons where other climatic conditions though, like increased rainfall and relative humidity, possibly reflect to the pathogen’s potential to colonize its host (e.g., chicken broilers) [86,87]. The seasonal effect on Campylobacter prevalence is inferred either by the increased number of reported campylobacteriosis cases during the summer months [10,82] or the increased fly activity [88] and the presence of flies in the summer, which play the role of the bacterial vector (mechanical/transport host) in the warm months [83]. In addition, the aforementioned increase in prevalence could reflect the level of environmental contamination due to more regular ventilation of the poultry houses in summer, resulting in increased contact of birds with the external environment [89,90]. To this end, higher wind speeds and the geographical location of the farm, surrounded by a more agricultural landscape, may also contribute to the extent of contamination in the poultry flock [84,91]. Wind may introduce contaminated material (e.g., soil, dried fecal particles) to the poultry house, which is more likely to exist in the rural setting, exposing the flock to Campylobacter spp. among other fecal pathogens, if birds have access to the external environment [91]. Biosecurity measures at the farm level greatly contribute towards prevention of Campylobacter colonization and reduce prevalence of the pathogen in broiler flocks [90,92].
On the other hand, several papers highlight the non-statistically significant effect of seasonality on Campylobacter populations from broiler carcasses or chicken meat [61,63,90,93], where especially in the warmer countries of southern Europe, like the Mediterranean countries, a higher prevalence of Campylobacter spp. is reported in broiler flocks throughout the year compared to the colder northern European countries with the more pronounced and obvious seasonality in their bacterial recoveries of the pathogen [90,92]. Moreover, many times the inability to ascertain the degree of influence of the seasonality on the recovery rate of Campylobacter spp. is even related to the small number of samples analyzed which does not allow safe conclusions to be drawn [61,63].
Regarding the distribution of different Campylobacter species in poultry carcasses, Manfreda et al. [59] reported higher recovery rates for C. jejuni (75.2%) against C. coli (24.8%) in winter (i.e., December–March) for chicken carcasses sampled from a single slaughterhouse in Italy, which were processed from broilers coming from a dozen of different chicken farms, compared to other seasons where C. jejuni was either absent (in autumn), marginally (in spring) or slightly (in summer) dominant over C. coli. In contrast, in a retrospective study conducted in Poland for the five-year period 2014–2018, Wieczorek et al. [61] showed that the peak in the distribution of C. coli was clearly placed to the autumn-winter months (i.e., October–February).
3. Isolation of Campylobacter spp.
3.1. Detection and/or Enumeration Procedure
The choice of microbiological method that can be used to isolate campylobacters from a food commodity can significantly affect the prevalence or even the estimated concentration of the pathogen in food [73,94,95,96,97,98,99,100,101], and therefore the distribution of different Campylobacter species recovered from food during the microorganism’s detection or enumeration procedure followed. C. jejuni was mainly isolated during pathogen enumeration procedure, while C. coli was the predominant species recovered from all plating culture media following enrichment in Bolton broth during the detection of Campylobacter in chicken meat [63,98].
The possibility of combining different enrichment broths (e.g., Bolton broth or Preston broth) with different selective and/or differential (i.e., chromogenic) solid culture media (i.e., agars) (e.g., modified charcoal-cefoperazone-deoxycholate agar; mCCDA, Preston agar) could be utilized as an alternative to the standard ISO 10272-1 protocol [102] used for the detection of Campylobacter in food [103,104,105]. For pathogen enumeration, different agars than mCCDA proposed by ISO 10272-2 enumeration method [107], either selective (e.g., Preston agar or Karmali agar) or chromogenic (e.g., Campylobacter selective agar; CASA®, Brilliance™ Campy count agar, BCCA) which have been found to perform equally well to the recommended mCCDA in enumerating colonies of Campylobacter spp. [63,96,108,109,110,111,112], could be used after the initial dilution of the food sample. After all, the colored colonies of Campylobacter on chromogenic agars are easier to record and count than those on the charcoal-based agars, such as mCCDA.
Enrichment of chicken meat samples in Preston broth for 24 h followed by plating on mCCDA performed better than 48 h enrichment in Bolton broth and plating on mCCDA [96], while this alternative enrichment and plating combination was taken into account in the revised ISO method for Campylobacter spp. detection (i.e., procedure B in the ISO 10272-1 protocol) [102]. Furthermore, the parallel use of the detection and enumeration procedures improves the recovery of Campylobacter spp., whereas chromogenic agars, like CASA and BCCA, should be considered as secondary plating media for simultaneous or optional use together with the ISO recommended mCCDA and/or even as suitable alternatives to the latter selective agar [63]. In any case, it has been proved that the previously mentioned enrichment and/or plating alternatives can significantly affect the recovery of Campylobacter strains from food [73,94,95,96,97,98,99,100,101].
3.1.1. Composition of Culture Media for Detection/Enumeration
In food samples with an expected low number of Campylobacter spp. and low concentration levels of accompanying (background) microbiota and/or stressed campylobacters (e.g., cooked or frozen samples), the enrichment in Bolton broth is highly recommended for pathogen detection, whereas for samples with high concentration levels of background microbiota other than Campylobacter spp. (e.g., raw meats including poultry or raw milk), the use of Preston broth as a sample enrichment medium is suggested [96,102].
The reduced recovery of campylobacters from raw chicken meat after sample enrichment in Bolton broth compared to direct plating from the initial dilution of the food sample, is likely attributable to the presence of cefoperazone in the liquid medium. The latter is a third generation β-lactam antibiotic which is supplemented into Bolton enrichment broth, as well as in mCCDA plates utilized for pathogen isolation. Foods containing microbiota resistant to third generation β-lactams, including cefoperazone, such as raw chicken containing β-lactam-degrading E. coli, namely extended-spectrum β-lactamase-producing; ESBL-producing E. coli, may lead to the overgrowth of the accompanying flora during the enrichment procedure in Bolton broth, which in turns leads to the suppression of Campylobacter spp. rendering them non detectable following the subculturing on agar plates. Limited growth of Campylobacter co-cultured with ESBL-producing E. coli in Bolton broth is hypothesized to be due to oxygen availability during the growth of Campylobacter in the medium [113]. In such a case, different enrichment broth and plating agar combinations, based on different principles of selectivity (e.g., Preston broth combined with Preston agar), are considered more suitable for combating this type of resistant microbiota and allowing for better detection of Campylobacter spp. in chicken meat [96,102,103,105]. Alternatively, restoring the selectivity of Bolton broth and mCCDA can be achieved by supplementation of these media with β-lactamase inhibitors or inhibitory bacterial growth agents (e.g., antibiotics) to overcome the problem of ESBL-producing E. coli [114,115,116,117,118,119,120,121].
4. Confirmation and Identification of Campylobacter spp.
Using conventional microbiological methods and following the enrichment of food sample or direct plating from the initial dilution of the food sample and isolation of Campylobacter spp. on selective plating media, there is a need to confirm the presence of Campylobacter and then to proceed, if necessary, with the identification of Campylobacter species [102,107]. Biochemical tests are routinely used for confirmation and identification purposes, even though molecular methods, such as polymerase chain reaction (PCR) techniques (e.g., multiplex or quantitative PCR), are gaining more and more ground lately.
4.1. Biochemical Differentiation of Campylobacter species
Among the Campylobacter spp. present in food, the most frequently encountered species, as already highlighted, are C. jejuni and C. coli. However, other species have been described (e.g., C. lari, C. upsaliensis; Supplementary Table S1), some key characteristics of which permit their differentiation (Table 1) [102,107].
With a closer look, Table 1 reveals that the only biochemical test that can distinguish between C. jejuni and C. coli is hippurate hydrolysis, with the former species showing a positive reaction compared to the latter species which give a negative reaction to the presence of hippurate. Thence, this test is typically used to differentiate C. jejuni and C. coli. Nevertheless, given the fact that some hippurate-negative C. jejuni strains have been reported [122,123], hippurate hydrolysis is not a robust criterion for differentiating thermotolerant Campylobacter species.
4.2. Molecular Methods for Differentiating Campylobacter species
The inherent disadvantage that biochemical differentiation of C. jejuni and C. coli has because of the low specificity (ca. 20%) hippurate hydrolysis exhibits to some hippurate-negative strains of C. jejuni [122,123], together with the laborious and time-consuming character of biochemical identification tests, stimulated the development of molecular diagnostic methods and techniques as alternatives to the classical culture-dependent approach to differentiation of Campylobacter species.
Serological methods (i.e., immunoassays) (e.g., enzyme-linked immunosorbent assay; ELISA) [124,125,126,127,128], biosensors for the on-site detection of foodborne pathogens [129,130], DNA hybridization techniques (e.g., loop-mediated isothermal amplification; LAMP) [126,131,132], DNA fingerprinting techniques (e.g., multilocus sequence typing; MLST) [133,134,135] and above all PCR-based method and techniques (e.g., multiplex PCR; mPCR, quantitative or real time PCR; qPCR/rt-PCR), have been developed for the fastest and most efficient identification and differentiation of Campylobacter species among other foodborne pathogens. It should be noted though that some DNA fingerprinting techniques are more sophisticated (e.g., pulsed-field gel electrophoresis; PFGE, whole-genome sequencing; WGS) and require well-trained personnel with a know-how-to conduct the technique and interpret the data. Due to their enormous discriminatory power these molecular typing methods are preferable tools in outbreak investigations rather than the routine monitoring of pathogens [136,137,138,139,140]. For this reason, PCR techniques are the method of choice for Campylobacter differentiation during laboratory screening for the pathogen.
4.2.1. PCR-Based Methods and Techniques
The technique of mPRC has been widely studied by several researchers in an effort to find a fast and at the same time reliable means of identifying isolated Campylobacter species [141,142,143]. The combination of food sample enrichment with multiplex real-time PCR (mrt-PCR) results in a more rapid detection and identification of Campylobacter spp. isolated from food [144], compared to the standard ISO method utilizing the biochemical identification of the isolates [102]. Nevertheless, the enrichment of the food sample as well as the initial Campylobacter load in the matrix significantly affect the isolation frequency and the recovery rate of different subtypes of C. jejuni [145] and of many Campylobacter species. In such a case, combining mPCR with the pre-enrichment in Brucella broth and further enrichment in Preston broth of chicken meat samples led to the identification of the majority of isolated campylobacters as C. coli (53%) than C. jejuni (47%) [146].
The quantitative determination of Campylobacter spp. in contaminated foods and generally in the food chain is crucial, among others in order to comply with imposed EU microbiological criteria for Campylobacter in broiler carcasses [147]. Quantitative molecular PCR-based methods, such as rt-PCR techniques, have been developed for such quantification purposes [148,149,150,151], although currently only the culture-based ISO method is officially approved for pathogen enumeration [104]. Nonetheless, PCR-based methods suffer from the inability to differentiate between live and dead cells of the target microorganism and the apparent PCR signal that may occur from DNA originating from those dead cells. Therefore, quantitative viability rt-PCR assays have been proposed to normalize on the one hand the underestimation of Campylobacter spp. recovered from different food matrices, on the other to allow for reliable differentiation between live and dead Campylobacter and thus accurate estimations of pathogen concentration in foods [79,152,153,154,155].
5. Biofilm-forming ability of Campylobacter spp.
The survival of campylobacter in the food chain remains a paradox since the bacterium is a fastidious organism with characteristic special growth requirements for successful subculturing in the laboratory (e.g., heat-resistant, microaerophilic organism requiring the presence of blood in its culture medium). Recently, biofilm formation has been proposed as the main mechanism of maintenance and transmission for the pathogen from animals to humans [156]. In general, the biofilm-forming ability of Campylobacter is strain-dependent and varies among organism’s isolated strains [157,158], as well as between different Campylobacter spp. [159,160,161], while it is also affected by the presence of other bacterial species [162,163,164,165,166,167]. Regarding the biofilm-forming ability of C. jejuni and C. coli, the latter isolates seem able to form biofilms significantly better compared to C. jejuni isolates (p < 0.05) [157,160] and that could be another reason for the increased prevalence of C. coli against C. jejuni in the chicken meat samples.
The ability of C. jejuni to form a biofilm is highly dependent on the strain and the type of abiotic surface on which it is found [159]. Teh et al. [165] concluded that C. jejuni exhibits a much weaker biofilm-forming ability compared to other bacteria, such as Pseudomonas spp., Staphylococcus aureus, Salmonella spp., and E. coli. However, in controlled mixed-microbial populations of a specific C. jejuni strain (sequence type; ST-474) with Enterococcus faecalis and/or Staphylococcus sp., optical intense biofilms for the two species were developed when they were grown with C. jejuni, while C. jejuni cells were recovered from most of the biofilms containing E. faecalis and/or Staphylococcus sp. [165]. That was the case and in the studies of Ica et al. [166] and Sterniša et al. [167], where the co-cultivation of C. jejuni with P.aeruginosa and P. fragi, respectively, resulted in the increased determined number of culturable biofilm C. jejuni cells. In contrast to monoculture biofilms, the mixed-culture biofilms of C. jejuni with pseudomonads had significantly enhanced mechanical strength [166]. Enhanced biofilm formation was also observed for C. jejuni and C. coli in the presence of S. aureus, with increased aerotolerance and survivability in parallel for the Campylobacter strains [162].
6. Conclusions
The prevalence and distribution of Campylobacter spp. in raw poultry meat from broiler carcasses depends on a variety of factors, such as the sampling method, part of the animal carcass from which the meat sample is obtained, seasonality of sampling, the isolation procedure followed with the different enrichment and plating media utilized for pathogen isolation along with the methods and techniques used for Campylobacter spp. differentiation, as well as the biofilm-forming ability of the isolated Campylobacter strains with regards to their co-culture with other bacterial species. All these factors should be considered when conducting field surveys or monitoring for Campylobacter presence in naturally contaminated poultry meat samples. At the same time, the indicated number of factors highlights the multifactorial dimension and complexity of the phenomenon when interpreting results for the recovery of Campylobacter spp. from poultry meat. Thus, the noticed epidemiological transition from the established predominance of C. jejuni to the ever-increasing recovery of C. coli in raw chicken meat could be the result of such a versatile effect. The present review attempted for the first time to elucidate the causes of this noteworthy epidemiological swift in prevalence and distribution of Campylobacter species on the food matrix itself, without extending to the interaction between pathogen and human host.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: List of thermotolerant Campylobacter species currently identified.
Author Contributions
Conceptualization, N.D.A.; methodology, N.D.A. and M.M.; X.X.; validation, N.D.A., N.T. and M.M.; formal analysis, N.D.A. and M.M.; investigation, N.D.A. and N.T.; resources, N.D.A. and M.M.; data curation, N.D.A. and M.M.; writing—original draft preparation, N.D.A.; writing—review and editing, N.D.A., N.T. and M.M.; visualization, N.D.A.; supervision, N.T.; project administration, N.D.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- EFSA (European Food Safety Authority); ECDC (European Centre for Disease Prevention and Control). The European Union one health 2021 zoonoses report. ESFA J. 2022, 20, e07666. [CrossRef]
- Kumagai, Y.; Pires, S.M.; Kubota, K.; Asakura, H. Attributing human foodborne diseases to food sources and water in Japan using analysis of outbreak surveillance data. J. Food Prot. 2020, 83, 2087–2094. [CrossRef]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.-A.; Roy, S.L.; Jones, J.L.; Grifiin, P.M. Foodborne illness acquired in the United States – Major pathogens, Emerg. Infect. Dis. 2011, 17, 7–15. [CrossRef]
- Walter, E.J.S.; Griffin, P.M.; Bruce, B.B.; Hoekstra, R.M. Estimating the number of illnesses caused by agents transmitted commonly through food: A scoping review. Foodborne Pathog. Dis. 2021, 18, 841–858. [CrossRef]
- Bisht, A.; Kamble, M.P.; Choudhary, P.; Chaturvedi, K.; Kohli, G.; Juneja, V.K.; Sehgal, S.; Taneja, N.K. A surveillance of food borne disease outbreaks in India: 2009–2018. Food Control 2021, 121, 107630. [CrossRef]
- Li, W.; Pires, S.M.; Liu, Z.; Ma, X.; Liang, J.; Jiang, Y.; Chen, J.; Liang, J.; Wang, S.; Wang, L.; Wang, Y.; Meng, C.; Huo, X.; Lan, Z.; Lai, S.; Liu, C.; Han, H.; Liu, J. Fu, P.; Guo, Y. Surveillance of foodborne disease outbreaks in China, 2003–2017. Food Control 2020, 118, 107359. [CrossRef]
- Zhao, J.; Cheng, H.; Wang, Z.; Fu, P.; Guo, Y.; Yang, S. Attribution analysis of foodborne disease outbreaks related to meat and meat products in China, 2002–2017. Foodborne Pathog. Dis. 2022, 19, 839–847. [CrossRef]
- Batz, M.B.; Hoffmann, S.; Morris, Jr., J.G. Ranking the disease burden of 14 pathogens in food sources in the United States using attribution data from outbreak investigations and expert elicitation. J. Food Prot. 2012, 75, 1278–1291. [CrossRef]
- Batz, M.B.; Richardson, L.C.; Bazaco, M.C.; Parker, C.C.; Chirtel, S.J.; Cole, D.; Golden, N.J.; Griffin, P.M.; Gu, W.; Schmitt, S.K.; Wolpert, B.J.; Kufel, J.S.Z.; Hoekstra, R.M. Recency-weighted statistical modeling approach to attribute illnesses caused by 4 pathogens to food sources using outbreak data, United States. Emerg. Infect. Dis. 2021, 27, 214–222. [CrossRef]
- Silva, J.; Leite, D.; Fernandes, M.; Mena, C.; Gibbs, P.A.; Teixeira, P. Campylobacter spp. as a foodborne pathogen: a review. Front. Microbiol. 2011, 2, 200. [CrossRef]
- Stern, N.J.; Hernandez, M.P.; Blankenship, L.; Deibel, K.E.; Doores, S.; Doyle, M.P.; Ng, H.; Pierson, M.D.; Sofos, J.N.; Sveum, W.H.; Westhoff, D.C. Prevalence and distribution of Campylobacter jejuni and Campylobacter coli in retail meats. J. Food Prot. 1985, 48, 595–599. [CrossRef]
- Mataragas, M.; Skandamis, P.N.; Drosinos, E.H. Risk profiles of pork and poultry meat and risk ratings of various pathogen/product combinations. Int. J. Food. Microbiol. 2008, 126, 1–12. [CrossRef]
- Scharff, R.L. Food attribution and economic cost estimates for meat- and poultry-related illnesses. J. Food Prot. 2020, 80, 959–967. [CrossRef]
- Teixeira, J.S.; Boras, V.F.; Hetman, B.J.; Taboada, E.N.; Inglis, G.D. Molecular epidemiological evidence implicates cattle as a primary reservoir of Campylobacter jejuni infecting people via contaminated chickens. Pathogens 2022, 11, 1366. [CrossRef]
- Ritchie, H.; Rosado, P.; Roser, M. Our World in Data: Meat and Dairy Production. 2017, Available online: https://ourworldindata.org/meat-production#citation (accessed on 12 April 2023).
- Parte, A.C.; Sardà Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2022, 70, 5607–5612. [CrossRef]
- Available online: https://www.bacterio.net/genus/campylobacter (accessed on 7 May 2023).
- Aydin, F.; Abay, S.; Kayman, T.; Karakaya, E.; Mustak, H.K.; Mustak, I.B.; Bilgen, N.; Goncuoglu, M.; Duzler, A.; Guran, O.; Sahin, O.; Saticioglu, I.B. Campylobacter anatolicus sp. nov., a novel member of the genus Campylobacter isolated from feces of Anatolian Ground Squirrel (Spermophilus xanthoprymnus) in Turkey. Syst. Appl. Microbiol. 2021, 44, 126265. [CrossRef]
- Boukerb, A.M.; Penny, C.; Serghine, J.; Walczak, C.; Cauchie, H.M.; Miller, W.G.; Losch, S.; Ragimbeau, C.; Mossong, J.; Mégraud, F.; Lehours, P.; Bénéjat, L.; Gourmelot, M. Campylobacter armoricus sp. nov., a novel member of the Campylobacter lari group isolated from surface water and stools from humans with enteric infection. Int. J. Syst. Evol. Microbiol. 2019, 69, 3969–3979. [CrossRef]
- Bryant, E.; Shen, Z.; Mannion, A.; Patterson, M.; Buczek, J.; Fox, J.G. Campylobacter taeniopygiae sp. nov., Campylobacter aviculae sp. nov., and Campylobacter estrildidarum sp. nov., novel species isolated from laboratory-maintained zebra finches. Avian Dis. 2020, 64, 457–466. [CrossRef]
- Rossi, M.; Debruyne, L.; Zanoni, R.G.; Manfreda, G.; Revez, J.; Vandamme, P. Campylobacter avium sp. nov., a hippurate-positive species isolated from poultry. Int. J. Syst. Evol. Microbiol. 2009, 59, 2364–2369. [CrossRef]
- Phung, C.; Scott, P.C.; Dekiwadia, C.; Moore, R.J.; Van T.T.H. Campylobacter bilis sp. nov., isolated from chickens with spotty liver disease. Int. J. Syst. Evol. Microbiol. 2022, 72, 5314. [CrossRef]
- Gilbert, M.J.; Zomer, A.L.; Timmerman, A.J.; Spaninks, M.P.; Rubio-Garcia, A.; Rossen, J.W.; Duim, B.; Wagenaar, J.A. Campylobacter blaseri sp. nov., isolated from common seals (Phoca vitulina). Int. J. Syst. Evol. Microbiol. 2018, 68, 1787–1794. [CrossRef]
- Inglis, G.D.; Hoar, B.M.; Whiteside, D.P.; Morck, D.W. Campylobacter canadensis sp. nov., from captive whooping cranes in Canada. Int. J. Syst. Evol. Microbiol. 2007, 57, 2636–2644. [CrossRef]
- Véron, M.; Chatelain, R. Taxonomic study of the genus Campylobacter Sebald and Véron and designation of the neotype strain for the type species, Campylobacter fetus (Smith and Taylor) Sebald and Véron. Int. J. Syst. Bact. 1973, 23:122–134. [CrossRef]
- Tanner, A.C.R.; Badger, S.; Lai, C.H.; Listgarten, M.A.; Visconti, R.A.; Socransky, S.S. Wolinella gen. nov., Wolinella succinogenes (Vibrio succinogenes Wolin et al.) comb. nov., and description of Bacteroides gracilis sp. nov., Wolinella recta sp. nov., Campylobacter concisus sp. nov., and Eikenella corrodens from humans with periodontal disease. Int. J. Syst. Bacteriol. 1981, 31, 432–445. [CrossRef]
- Koziel, M.; O'Doherty, P.; Vandamme, P.; Corcoran, G.D.; Sleator R.D.; Lucey, B. Campylobacter corcagiensis sp. nov., isolated from faeces of captive lion-tailed macaques (Macaca silenus). Int. J. Syst. Evol. Microbiol. 2014; 64, 2878–2883. [CrossRef]
- Zanoni, R.G.; Debruyne, L.; Rossi, M.; Revez, J.; Vandamme, P. Campylobacter cuniculorum sp. nov., from rabbits. Int. J. Syst. Evol. Microbiol. 2009, 59, 1666–1671. [CrossRef]
- Vandamme, P.; Falsen, E.; Rossau, R.; Hoste, B.; Segers, P.; Tytgat, R.; De Ley, J. Revision of Campylobacter, Helicobacter, and Wolinella taxonomy: emendation of generic descriptions and proposal of Arcobacter gen. nov. Int. J. Syst. Bacteriol. 1991, 41, 88–103. [CrossRef]
- Sebald, M.; Veron, M. [BASE DNA CONTENT AND CLASSIFICATION OF VIBRIOS]. Ann. Inst. Pasteur (Paris) 1963, 105, 897–910.
- Piccirillo, A.; Nier,o G.; Calleros, L.; Perez, R.; Naya, H.; Iraola, G. Campylobacter geochelonis sp. nov. isolated from the western Hermann's tortoise (Testudo hermanni hermanni). Int. J. Syst. Evol. Microbiol. 2016 66, 3468–3476. [CrossRef]
- Vandamme, P.; Daneshvar, M.I.; Dewhirst, F.E.; Paster, B.J.; Kersters, K.; Goossens, H.; Moss, C.W. Chemotaxonomic analyses of Bacteroides gracilis and Bacteroides ureolyticus and reclassification of B. gracilis as Campylobacter gracilis comb. nov. Int. J. Syst. Bacteriol. 1995, 45, 145–152. [CrossRef]
- Stanley, J.; Burnens, A.P.; Linton, D.; On, S.L.; Costas, M.; Owen, R.J. Campylobacter helveticus sp. nov., a new thermophilic species from domestic animals: characterization, and cloning of a species-specific DNA probe. J. Gen. Microbiol. 1992, 138, 2293–2303. [CrossRef]
- Van, T.T.; Elshagmani, E.; Gor, M.C.; Scott, P.C.; Moore, R.J. Campylobacter hepaticus sp. nov., isolated from chickens with spotty liver disease. Int. J. Syst. Evol. Microbiol. 2016, 66, 4518–4524. [CrossRef]
- Lawson, A.J.; On, S.L.; Logan, J.M.; Stanley, J. Campylobacter hominis sp. nov., from the human gastrointestinal tract. Int. J. Syst. Evol. Microbiol. 2001, 51, 651–660. [CrossRef]
- Gebhart, C.J.; Edmonds, P.; Ward, G.E.; Kurtz, H.J.; Brenner, D.J. "Campylobacter hyointestinalis" sp. nov.: a new species of Campylobacter found in the intestines of pigs and other animals. J. Clin. Microbiol. 1985, 21, 715–720. [CrossRef]
- Gilbert, M.J.; Kik, M.; Miller, W.G.; Duim, B.; Wagenaar, J.A. Campylobacter iguaniorum sp. nov., isolated from reptiles. Int. J. Syst. Evol. Microbiol. 2015, 65, 975–982. [CrossRef]
- Foster, G.; Holmes, B.; Steigerwalt, A.G.; Lawson, P.A.; Thorne, P.; Byrer, D.E.; Ross, H.M.; Xerry, J.; Thompson, P.M.; Collins, M.D. Campylobacter insulaenigrae sp. nov., isolated from marine mammals. Int. J. Syst. Evol. Microbiol. 2004, 54, 2369–2373. [CrossRef]
- Logan, J.M.; Burnens, A.; Linton, D.; Lawson, A.J.; Stanley, J. Campylobacter lanienae sp. nov., a new species isolated from workers in an abattoir. Int. J. Syst. Evol. Microbiol. 2000, 50, 865–872. [CrossRef]
- Benjamin, J.; Leaper, S.; Owen, R.J.; Skirrow, M.B. Description of Campylobacter laridis, a new species comprising the nalidixic acid resistant thermophilic Campylobacter (NARTC) group. Curr. Microbiol. 1983, 8, 231–238. [CrossRef]
- Lynch, C.; Peeters, C.; Walsh, N.; McCarthy, C.; Coffey, A.; Lucey, B.; Vandamme, P. Campylobacter majalis sp. nov. and Campylobacter suis sp. nov., novel Campylobacter species isolated from porcine gastrointestinal mucosa. Int. J. Syst. Evol. Microbiol. 2022, 72,5510. [CrossRef]
- Antezack, A.; Boxberger, M.; Rolland, C.; Ben Khedher, M.; Monnet-Corti, V.; La Scola, B. Isolation and characterization of Campylobacter massiliensis sp. nov., a novel Campylobacter species detected in a gingivitis subject. Int. J. Syst. Evol. Microbiol. 2021, 71, 5039. [CrossRef]
- Roop Ii, R.M.; Smibert, R.M.; Johnson, J.L.; Krieg, N.R. Campylobacter mucosalis (Lawson, Leaver, Pettigrew, and Rowland 1981) comb. nov.: emended description. Int. J. Syst. Bacteriol. 1985, 35, 189–192. [CrossRef]
- Bloomfield, S.; Wilkinson, D.; Rogers, L.; Biggs, P.; French, N.; Mohan, V.; Savoian, M.; Venter, P.; Midwinter, A. Campylobacter novaezeelandiae sp. nov., isolated from birds and water in New Zealand. Int. J. Syst. Evol. Microbiol. 2020, 70, 3775–3784. [CrossRef]
- Caceres, A.; Munoz, I.; Iraola, G.; Diaz-Viraque, F.; Collado, L. Campylobacter ornithocola sp. nov., a novel member of the Campylobacter lari group isolated from wild bird faecal samples. Int. J. Syst. Evol. Microbiol. 2017, 67, 1643–1649. [CrossRef]
- Debruyne, L.; On, S.L.; De Brandt, E.; Vandamme, P. Novel Campylobacter lari-like bacteria from humans and molluscs: description of Campylobacter peloridis sp. nov., Campylobacter lari subsp. concheus subsp. nov. and Campylobacter lari subsp. lari subsp. nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 1126–1132. [CrossRef]
- Gilbert, M.J.; Miller, W.G.; Leger, J.S.; Chapman, M.H.; Timmerman, A.J.; Duim, B.; Foster, G.; Wagenaar, J.A. Campylobacter pinnipediorum sp. nov., isolated from pinnipeds, comprising Campylobacter pinnipediorum subsp. pinnipediorum subsp. nov. and Campylobacter pinnipediorum subsp. caledonicus subsp. nov. Int. J. Syst. Evol. Microbiol. 2017, 67, 1961–1968. [CrossRef]
- Silva, M.F.; Pereira, G.; Carneiro, C.; Hemphill, A.; Mateus, L.; Lopes-da-Costa, L.; Silva, E. Campylobacter portucalensis sp. nov., a new species of Campylobacter isolated from the preputial mucosa of bulls. PLoS One 2020, 15, e0227500. [CrossRef]
- Etoh, Y.; Dewhirst, F.E.; Paster, B.J.; Yamamoto, A.; Goto, N. Campylobacter showae sp. nov., isolated from the human oral cavity. Int. J. Syst. Bacteriol. 1993, 43, 631–639. [CrossRef]
- Debruyne, L.; Broman, T.; Bergstrom, S.; Olsen, B.; On, S.L.; Vandamme, P. Campylobacter subantarcticus sp. nov., isolated from birds in the sub-Antarctic region. Int. J. Syst. Evol. Microbiol. 2010, 60, 815–819. [CrossRef]
- Sandstedt, K.; Ursing, J. Description of Campylobacter upsaliensis sp. nov. previously known as the CNW group. Syst. Appl. Microbiol. 1991, 14, 39–45. [CrossRef]
- Vandamme, P.; Debruyne, L.; De Brandt, E.; Falsen, E. Reclassification of Bacteroides ureolyticus as Campylobacter ureolyticus comb. nov., and emended description of the genus Campylobacter. Int. J. Syst. Evol. Microbiol. 2010, 60, 2016–2022. [CrossRef]
- Debruyne, L.; Broman, T.; Bergstrom, S.; Olsen, B.; On, S.L.; Vandamme, P. Campylobacter volucris sp. nov., isolated from black-headed gulls (Larus ridibundus). Int. J. Syst. Evol. Microbiol. 2010, 60, 1870–1875. [CrossRef]
- Parisi, A.; Chiara, M.; Caffara, M.; Mion, D.; Miller, W.G.; Caruso, M.; Manzari, C.; Florio, D.; Capozzi, L.; D'Erchia, A.M.; Manzulli, V.; Zanoni, R.G. Campylobacter vulpis sp. nov. isolated from wild red foxes. Syst. Appl. Microbiol. 2021, 44, 126204. [CrossRef]
- Ghafir, Y.; China, B.; Dierick, K.; De Zutter, L.; Daube, G. A seven-year survey of Campylobacter contamination in meat at different production stages in Belgium. Int. J. Food Microbiol. 2007, 116, 111–120. [CrossRef]
- Wagenaar, J.A.; van der Graaf-van Blools, L. Chapter 3.10.4. Infection with Campylobacter jejuni and C. coli. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, OIE; World Organization for Animal Health, 2018; pp. 1669–1677. Available online: https://www.woah.org/en/what-we-do/standards/codes-and-manuals/terrestrial-manual-online-access/ (accessed on 11 April 2023).
- Jorgensen, F.; Ellis-Iversen, J.; Rushton, S.; Bull, S.A.; Harris; S.A.; Bryan, S.J.; Gonzalez, A.; Humphrey, T.J. Influence of season and geography on Campylobacter jejuni and C. coli subtypes in housed broiler flocks reared in Great Britain. Appl. Environ. Microbiol. 2011, 77, 3741–3748. [CrossRef]
- Habib, I.; Mohamed, M.-Y.I.; Lakshmi, G.B.; Khan, M.; Li, D. Quantification of Campylobacter contamination on chicken carcasses sold in retail markets in the United Arab Emirates. Int. J. Food Contam. 2022, 9, 9. [CrossRef]
- Manfreda, G.; De Cesare, A.; Bondioli, V.; Stern, J.N.; Franchini, A. Enumeration and identity of Campylobacter spp. in Italian broilers. Poult. Sci. 2006, 85, 556–562. [CrossRef]
- Repérant, E.; Laisney, M.J.; Nagard, B.; Quesne, S.; Rouxel, S.; Le Gall, F.; Chemaly, M.; Denis, M. Influence of enrichment and isolation media on the detection of Campylobacter spp. in naturally contaminated chicken samples. J. Microbiol. Methods 2016, 128, 42–47. [CrossRef]
- Wieczorek, K.; Bocian, Ł.; Osek, J. Prevalence and antimicrobial resistance of Campylobacter isolated from carcasses of chickens slaughtered in Poland – a retrospective study. Food Control 2020, 112, 107159. [CrossRef]
- Wieczorek, K.; Szewczyk, R.; Osek, J. Prevalence, antimicrobial resistance, and molecular characterization of Campylobacter jejuni and C. coli isolated from retail raw meat in Poland. Vet. Med. 2012, 57, 293–299.
- Andritsos, N.D.; Tzimotoudis, N.; Mataragas, M. Estimating the performance of four culture media used for enumeration and detection of Campylobacter species in chicken meat. LWT 2020, 118, 108808. [CrossRef]
- Marinou, I.; Bersimis, S.; Ioannidis, A.; Nicolau, C.; Mitroussia-Ziouva, A.; Legakis, N.J.; Chatzipanagiotou, S. Identification and antimicrobial resistance of Campylobacter species isolated from animal sources. Front. Microbiol. 2012, 3, 58. [CrossRef]
- Jeffrey, J.S.; Tonooka, K.H.; Lozano, J. (2001). Prevalence of Campylobacter spp. from skin, crop, and intestine of commercial broiler chicken carcasses at processing. Poult. Sci. 2001, 80, 1390–1392. [CrossRef]
- Buhr, R.J.; Berrang, M.E.; Cason, J.A. Bacterial recovery from breast skin of genetically feathered and featherless broiler carcasses immediately following scalding and picking. Poult. Sci. 2003, 82, 1641–1647.
- Cason, J.A.; Hinton, Jr., A.;Buhr, R.J. Impact of feathers and feather follicles on broiler carcass bacteria. Poult. Sci. 2004, 83, 1452–1455. [CrossRef]
- Habib, I.; Sampers, I.; Uyttendaelle, M.; Berkvens, D.; De Zutter, L. Baseline data from a Belgium-wide survey of Campylobacter species contamination in chicken meat preparations and considerations for a reliable monitoring program. Appl. Environ. Microbiol. 2008, 74, 5483–5489. [CrossRef]
- Kinde, H.; Genigeorgis, C.A.; Pappaioanou, M. Prevalence of Campylobacter jejuni in chicken wings. Appl. Environ. Microbiol. 1983, 45, 1116–1118. [CrossRef]
- Rayes, H.M.; Genigeorgis, C.A.; Farver, T.B. Prevalence of Campylobacter jejuni on turkey wings at the supermarket level. J. Food Prot. 1983, 46, 292–294. [CrossRef]
- Baré, J.; Uyttendaele, M.; Depraetere, O.; Houf, K.; De Zutter, L. Variation in Campylobacter distribution on different sites of broiler carcasses. Food Control 2013, 32, 279–282. [CrossRef]
- Russell, S.M.; Cox, N.A.; Bailey, J.S. Sampling poultry carcasses and parts to determine bacterial levels. J. Appl. Poult. Res. 1997, 6, 234–237. [CrossRef]
- Sproston, E.L.; Carrillo, C.D.; Boulter-Bitzer, J. The quantitative and qualitative recovery of Campylobacter from raw poultry using USDA and Health Canada methods. Food Microbiol. 2014, 44, 258–263. [CrossRef]
- Commission Regulation (EC) 2073/2005. Microbiological criteria for foodstuffs. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32005R2073 (accessed on 12 April 2023).
- Commission Regulation (EU) 2017/1495. Amending Regulation (EC) No 2073/2005 as regards Campylobacter in broiler carcasses. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32017R1495 (accessed on 12 April 2023).
- Gill, C.O.; Badoni, M.; Moza, L.F.; Barbut, S.; Griffiths, M.W. Microbiological sampling of poultry carcass portions by excision, rinsing, or swabbing. J. Food Prot. 2005, 68, 2718–2720. [CrossRef]
- Bishop. H.; Evans, J.; Eze, J.I.; Webster, C.; Humphry, R.W.; Beattie, R.; White, J.; Couper, J.; Allison, L.; Brown, D.; Tongue, S.C. Bacteriological survey of fresh minced beef on sale at retail outlets in Scotland in 2019: Three foodborne pathogens, hygiene process indicators, and phenotypic antimicrobial resistance. J. Food Prot. 2022, 85, 1370–1379. [CrossRef]
- Stella, S.; Soncini, G.; Ziino, G.; Panebianco, A.; Pedonese, F.; Nuvoloni, R.; Di Giannatale, E.; Colavita, G.; Alberghini, L.; Giaccone, V. Prevalence and quantification of thermophilic Campylobacter spp. in Italian retail poultry meat: Analysis of influencing factors. Food Microbiol. 2017, 62, 232–238. [CrossRef]
- Beterams, A.; Tolksdorf, T.; Martin, A.; Stingl, K.; Bandick, N.; Reich, F. Change of Campylobacter, Escherichia coli and Salmonella counts in packaged broiler breast meat stored under modified atmosphere and vacuum conditions at 4 and 10 °C based on cultural and molecular biological quantification. Food Control 2023, 145, 109337. [CrossRef]
- Proietti, P.C.; Pergola, S.; Bellucci, S.; Menchetti, L.; Miraglia, D.; Franciosini, M.P. Occurrence and antimicrobial susceptibility of Campylobacter spp. on fresh and refrigerated chicken meat products in central Italy. Poult. Sci. 2018, 97, 2895–2901. [CrossRef]
- Andritsos, N.D.; Paramithiotis, S.; Mataragas, M.; Drosinos, E.H. Listeria monocytogenes serogroup 1/2 strains have a competitive growth advantage over serotype 4b during refrigerated storage of an artificially contaminated ready-to-eat pork meat product. Appl. Sci. 2021, 11, 6096. [CrossRef]
- Djennad, A.; Lo lacono, G.; Sarran, C.; Lane, C.; Elson, R.; Höser, C.; Lake, I.R.; Colón-González, F.J.; Kovats, S.; Semenza, J.C.; Bailey, T.C.; Kessel, A.; Fleming, L.E.; Nichols, G.L. Seasonality and the effects of weather on Campylobacter infections. BMC Infect. Dis. 2019, 19, 255. [CrossRef]
- Guerrin, M.T.; Sir, C.; Sargeant, J.M.; Wadell, L.; O’Connor, A.M.; Wills, R.M.; Bailey, R.H.; Byrd, J.A. The change in prevalence of Campylobacter on chicken carcasses during processing. Poult. Sci. 2010, 89, 1070–1084. [CrossRef]
- Jorgensen, F.; Ellis-Iversen, J.; Rushton, S.; Bull, S.A.; Harris, S.A.; Bryan, S.J.; Gonzalez, A.; Humphrey, T.J. Influence of season and geography on Campylobacter jejuni and C. coli subtypes in housed broiler flocks reared in Great Britain. Appl. Environ. Microbiol. 2011, 77, 3741–3748. [CrossRef]
- Mäesaar, M.; Praakle, K.; Meremäe, K.; Kramarenko, T.; Sõgel, J.; Viltrop, A.; Muutra, K.; Kovalenko, K.; Matt, D.; Hörman, A.; Hänninen, M.-L.; Roasto, M. Prevalence and counts of Campylobacter spp. in poultry meat at retail level in Estonia. Food Control 2014, 44, 72–77. [CrossRef]
- Kalupahana, R.S.; Mughini-Gras, L.; Kottawatta, S.A.; Somarathne, S.; Gamage, C.; Wagenaar, J.A. Weather correlates of Campylobacter prevalence in broilers at slaughter under tropical conditions in Sri Lanka. Epidemiol. Infect. 2018, 146, 972–979. [CrossRef]
- Prachantasena, S.; Charununtakorn, P.; Muangnoicharoen, S.; Hankla, L.; Techawal, N.; Chaveerach, P.; Tuitemwong, P.; Chokesajjawatee, N.; Williams, N.; Humphrey, T.; Luangtongkum, T. Climatic factors and prevalence of Campylobacter in commercial broiler flocks in Thailand. Poult. Sci. 2017, 96, 980–985. [CrossRef]
- Jonsson, M.E.; Chriél, M.; Norström, M.; Hofshagen, M. Effect of climate and farm environment on Campylobacter spp. colonization in Norwegian broiler flocks. Prev. Vet. Med. 2012, 107, 95–104. [CrossRef]
- Nylen, G.; Dunstan, F.; Palmer, S.R.; Andersson, Y.; Bager, F.; Cowden, J.; Feierl, G.; Galloway, Y.; Kapperud, G.; Megraud, F.; Molbak, K.; Petersen, L.R.; Ruutu, P. The seasonal distribution of campylobacter infection in nine European countries and New Zealand. Epidemiol. Infect. 2002, 128, 383–390. [CrossRef]
- Urdaneta, S.; Lorca-Oró, C.; Dolz, R.; López-Soria, S.; Cerdà-Cuéllar, M. In a warm climate, ventilation, indoor temperature and outdoor relative humidity have significant effects on Campylobacter spp. colonization in chicken broiler farms which can occur in only 2 days. Food Microbiol. 2023, 109, 104118. [CrossRef]
- Smith, O.M.; Cornell, K.A.; Crossley, M.S.; Crespo, R.; Jones, M.S.; Snyder, W.E.; Owen, J.P. Wind speed and landscape context mediate Campylobacter risk among poultry reared in open environments. Animals 2023, 13, 492. [CrossRef]
- Sommer, H.M.; Borck Høg, B.; Larsen, L.S.; Sørensen, A.I.V.; Williams, N.; Merga, J.Y.; Cerdà-Cuéllae, M.; Urdaneta, S.; Dolz, R.; Wieczorek, K.; Osek, J.; David, B.; Hofshagen, M.; Jonsson, M.; Wagenaar, J.A.; Bolder, N.; Rosenquist, H. Analysis of farm specific risk factors for Campylobacter colonization of broilers in six European countries. Microb. Risk Anal. 2016, 2–3, 16–26. [CrossRef]
- Wong, T.L.; Hollis, L.; Cornelius, A.; Nicol, C.; Cook, R.; Hudson, J.A. Prevalence, numbers, and subtypes of Campylobacter jejuni and Campylobacter coli in uncooked retail meat samples. J. Food. Prot. 2007, 70, 566–573. [CrossRef]
- Baylis, C.L.; MacPhee, S.; Martin, K.W.; Humphrey, T.J.; Betts, R.P. Comparison of three enrichment media for the isolation of Campylobacter spp. from foods. J. Appl. Microbiol. 2000, 89, 884–891. [CrossRef]
- Carrillo, C.D.; Plante, D.; Iugovaz, I.; Kenwell, R.; Bélanger, G.; Boucher, F.; Poulin, N.; Trottier, Y.-L. Method-dependent variability in determination of prevalence of Campylobacter jejuni and Campylobacter coli in Canadian retail poultry. J. Food Prot. 2014, 77, 1682–1688. [CrossRef]
- Habib, I.; Uyttendaele, M.; De Zutter, L. Evaluation of ISO 10272:2006 standard versus alternative enrichment and plating combinations for enumeration and detection of Campylobacter in chicken meat. Food Microbiol. 2011, 28, 1117–1123. [CrossRef]
- Jasson, V.; Sampers, I.; Botteldoorn, N.; López-Gálvez, F.; Baert, L.; Denayer, S.; Rajkovic, A.; Habib, I.; De Zutter, L.; Debevere, J.; Uyttendaele, M. Characterization of Escherichia coli from raw poultry in Belgium and impact on the detection of Campylobacter jejuni using Bolton broth. Int. J. Food Microbiol. 2009, 135, 248–253. [CrossRef]
- Repérant, E.; Laisney, M.J.; Nagard, B.; Quesne, S.; Rouxel, S.; Le Gall, F.; Chemaly, M.; Denis, M. Influence of enrichment and isolation media on the detection of Campylobacter spp. in naturally contaminated chicken meat samples. J. Microbiol. Methods 2016, 128, 42–47. [CrossRef]
- Seliwiorstow, T.; De Zutter, L.; Houf, K.; Botteldoorn, N.; Baré, J.; Van Damme, I. Comparative performance of isolation methods using Preston broth, Bolton broth and their modifications for the detection of Campylobacter spp. from naturally contaminated fresh and frozen raw poultry meat. Int. J. Food Microbiol. 2016, 234, 60–64. [CrossRef]
- Stern, N.J.; Line, J.E. Comparison of three methods for recovery of Campylobacter spp. from broiler carcasses. J. Food Prot. 1992, 55, 663–666. [CrossRef]
- Williams, L.K.; Jørgensen, F.; Grogono-Thomas, R.; Humphrey, T.J. Enrichment culture for the isolation of Campylobacter spp.: Effects of incubation conditions and the inclusion of blood in selective broths. Int. J. Food Microbiol. 2009, 130, 131–134. [CrossRef]
- ISO 10272-1, G: Food Chain–Horizontal Method for Detection and Enumeration of Campylobacter spp.–Part 1: Detection Method. ISO (International Organization for Standardization), 2017.
- Hayashi, M.; Kubota-Hayashi, S.; Natori, T.; Mizuno, T.; Miyata, M.; Yoshida, S.; Zhang, J.; Kawamoto, K.; Ohkusu, K.; Makino, S.; Ezaki, T. Use of blood-free enrichment broth in the development of a rapid protocol to detect Campylobacter in twenty-five grams of chicken meat. Int. J. Food Microbiol. 2013, 163, 41–46. [CrossRef]
- Hinton, A.; Cox, N.A.; Selective medium for aerobic incubation of Campylobacter. J. Food Microbiol. Saf. Hyg. 2018, 3, 131. [CrossRef]
- Seliwiorstow, T.; De Zutter, L.; Houf, K.; Botteldoorn, N.; Baré, J.; Van Damme, I. Comparative performance of isolation methods using Preston broth, Bolton broth and their modifications for the detection of Campylobacter spp. from naturally contaminated fresh and frozen raw poultry meat. Int. J. Food Microbiol. 2016, 234, 60–64. [CrossRef]
- Yoo, J.-H.; Choi, N.-Y.; Bae, Y.-M.; Lee, J.-S.; Lee, S.-Y. Development of a selective agar plate for the detection of Campylobacter spp. in fresh produce. Int. J. Food Microbiol. 2014, 189, 67–74. [CrossRef]
- ISO 10272-2, G: Food Chain–Horizontal Method for Detection and Enumeration of Campylobacter spp.–Part 2: Enumeration Method. ISO (International Organization for Standardization), 2017.
- Ahmed, León-Velarde, C.G.; Odumeru, J.A. Evaluation of novel agars for the enumeration of Campylobacter spp. in poultry retail samples. J. Microbiol. Methods 2012, 88, 304–310. [CrossRef]
- Le Bars, H.; Kayal, S.; Bonnaure-Mallet, M.; Minet, J. CASA chromogenic medium for enteric Campylobacter species. J. Clin. Microbiol. 2011, 49, 3675–3677. [CrossRef]
- Levican, A.; Hinton, Jr., A. CAMPYAIR, a new selective, differential medium for Campylobacter spp. isolation without the need for microaerobic atmosphere. Microorganisms 2022, 10, 1403. [CrossRef]
- Seliwiorstow, T.; Baré, J.; Verhaegen, B.; Uyttendaele, M.; De Zutter, L. Evaluation of a new chromogenic medium for direct enumeration of Campylobacter in poultry meat samples. J. Food Prot. 2014, 77, 2111–2114. [CrossRef]
- Teramura, H.; Iwasaki, M.; Ogihara, H. Development of a novel chromogenic medium for improved Campylobacter detection from poultry samples. J. Food Prot. 2015, 78, 1750–1755. [CrossRef]
- Lanzl, M.I.; van Mastrigt, O.; Zwietering, M.H.; Abee, T.; den Besten, H.M.W. Role of substrate availability in the growth of Campylobacter co-cultured with extended spectrum beta-lactamase-producing Escherichia coli in Bolton broth. Int. J. Food Microbiol. 2022, 363, 109518. [CrossRef]
- Chon, J.-W.; Hyeon, J.-Y.; Yim, J.-H.; Kim, J.-H.; Song, K.-Y.; Seo, K.-H. Improvement of modified charcoal-cefoperazone-deoxycholate agar by supplementation with a high concentration of polymyxin B for detection of Campylobacter jejuni and C. coli in chicken carcass rinses. Appl. Environ. Microbiol. 2012, 78, 1624–1626. [CrossRef]
- Chon, J.-W.; Kim, H.; Kim, H.-S.; Seo, K.-H. Improvement of modified charcoal-cefoperazone-deoxycholate agar by addition of potassium clavulanate for detecting Campylobacter spp. in chicken carcass rinse. Int. J. Food Microbiol. 2013, 165, 7–10. [CrossRef]
- Chon, J.-W.; Kim, H.; Yim, J.-H.; Song, K.-Y.; Moon, J.-S.; Kim, Y.-J.; Seo, K.-H. Improvement of Karmali agar by addition of polymyxin B for the detection of Campylobacter jejuni and C. coli in whole-chicken carcass rinse. J. Food Sci. 2013, 78, 752–755. [CrossRef]
- Chon, J.-W.; Kim, H.-S.; Kim, D.-H.; Kim, H.; Choi, I.-S.; Oh, D.-H.; Seo, K.-H. Modification of Karmali agar by supplementation with potassium clavulanate for the isolation of Campylobacter from chicken carcass rinses. J. Food Prot. 2014, 77, 1207–1211. [CrossRef]
- Chon, J.-W.; Kim, Y.-J.; Kim, H.-S.; Kim, D.-H.; Kim, H.; Song, K.-W.; Seo, K.-H. Supplementation of Bolton broth with triclosan improves detection of Campylobacter jejuni and Campylobacter coli in chicken carcass rinse. Int. J. Food Microbiol. 2014, 181, 37–39. [CrossRef]
- Chon, J.-W.; Kim, Y.-J.; Kim, Y.-J.; Jung, J.Y.; Bae, D.; Khan, S.; Seo, K.-H.; Sung, K. Addition of rifampicin to Bolton broth to inhibit extended-spectrum β-lactamase-producing Escherichia coli for the detection of Campylobacter. J. Food Sci. 2017, 82, 1688–1692. [CrossRef]
- Kim, Y.-J.; Whan, C.-J.; Kim, H.-S.; Kim, K.-Y.; Yim, J.-H.; Cho, S.-H.; Seo, K.-H. Improvement of Karmali agar by supplementation with tazobactam for detecting Campylobacter in raw poultry. J. Food Prot. 2016, 79, 1982–1985. [CrossRef]
- Moran, L.; Kelly, C.; Cormican, M.; McGettrick, S.; Madden, R.H. Restoring the selectivity of Bolton broth during enrichment for Campylobacter spp. from raw chicken. Lett. Appl. Microbiol. 2011, 52, 614–618. [CrossRef]
- Rautelin, H.; Jusufovic, J.; Hänninen, M.-L. Identification of hippurate-negative thermophilic campylobacters. Diagn. Microbiol. Infect. Dis. 1999, 35, 9–12. [CrossRef]
- Totten, P.A.; Patton, C.M.; Tenover, F.C.; Barrett, T.J.; Stamm, W.E.; Steigerwalt, A.G.; Lin, J.Y.; Holmes, K.K.; Brenner, D.J. Prevalence and characterization of hippurate-negative Campylobacter jejuni in King County, Washington. J. Clin. Microbiol.1987, 25, 1747–1752. [CrossRef]
- Bessède, E.; Asselineau, J.; Perez, P.; Valdenaire, G.; Richer, O.; Lehours, P.; Mégraud, F. Evaluation of the diagnostic accuracy of two immunochromatographic tests detecting campylobacter in stools and their role in campylobacter infection diagnosis. J. Clin. Microbiol. 2018, 56, e01567-17. [CrossRef]
- Franco, J.; Bénejat, L.; Ducournau, A.; Mégraud, F.; Lehours, P.; Bessède, E. Evaluation of CAMPYLOBACTER QUIK CHEK™ rapid membrane enzyme immunoassay to detect Campylobacter spp. antigen in stool samples. Gut Pathog. 2021, 13, 4. [CrossRef]
- Li, C.; Chen, X.; Wen, R.; Ma, P.; Gu, K.; Li, C.; Zhou, C.; Lei, C.; Tang, Y.; Wang, H. Immunocapture magnetic beads enhanced the LAMP-CRISP/Cas12a method for the sensitive, specific, and visual detection of Campylobacter jejuni. Biosensors 2022, 12, 154. [CrossRef]
- Muralidharan, C.; Anwar, A.; Wilson, T.B.; Scott, P.C.; Moore, R.J.; Van, T.T.H. Development of an enzyme-linked immunosorbent assay for detecting Campylobacter hepaticus specific antibodies in chicken sera – a key tool in spotty liver disease screening and vaccine development. Avian Pathol. 2020, 49, 658–665. [CrossRef]
- Schnee, A.E.; Haque, R.; Taniuchi, M.; Uddin, J.; Petri, W.A., Jr. Evaluation of two new membrane-based and microtiter plate enzyme-linked immunosorbent assays for detection of Campylobacter jejuni in stools of Bangladeshi children. J. Clin. Microbiol. 2018, 56, e00702-18. [CrossRef]
- He, D.; Du, Z.; Wang, Y.; Xu, E.; Jin, Z.; Wu, Z. Quantitative detection of Campylobacter jejuni with a core-satellite assemblies-based dual-modular aptasensor. Food Control 2022, 135, 108828. [CrossRef]
- Quintela, I.A.; Vasse, T.; Lin, C.-S.; Wu, V.C.H. Advances, applications, and limitations of portable and rapid detection technologies for routinely encountered foodborne pathogens. Front. Microbiol. 2022, 13, 1054782. [CrossRef]
- Babu, U.S.; Harrison, L.M.; Mammel, M.K.; Bigley III, E.C.; Hiett, K.L.; Balan, K.V. A loop-mediated isothermal amplification (LAMP) assay for the consensus detection of human pathogenic Campylobacter species. J. Microbiol. Methods 2020, 176, 106009. [CrossRef]
- Phaneuf, C.R.; Mangadu, B.; Tran, H.M.; Light, Y.K.; Sinha, A.; Charbonier, F.W.; Eckles, T.P.; Singh, A.K.; Koh, C.-Y. Integrated LAMP and immunoassay platform for diarrheal disease detection. Biosens. Bioelectron. 2018, 120, 93–101. [CrossRef]
- Asakura, H.; Yamamoto, S.; Kazuhiro, Y.; Kawase, J.; Nakamura, H.; Abe, K.; Sasaki, Y.; Ikeda, T.; Nomoto, R. Quantitative detection and genetic characterization of thermotolerant Campylobacter spp. in fresh chicken meats at retail in Japan. Front. Microbiol. 2022, 13, 1014212. [CrossRef]
- Aydin, F.; Kayman, T.; Abay, S.; Hizlisoy, H.; Saticioğlu, I.B.; Karakaya, E.; Sahin, O. MLST genotypes and quinolone resistance profiles of Campylobacter jejuni isolates from various sources in Turkey. Int. J. Food Microbiol. 2023, 391-393, 110137. [CrossRef]
- Joseph, L.A.; Griswold, T.; Vidyaprakash, E.; Im, S.B.; Williams, G.M.; Pouseele, H.A.; Hise, K.B.; Carleton, H.A. Evaluation of core genome and whole genome multilocus sequence typing schemes for Campylobacter jejuni and Campylobacter coli outbreak detection in the USA. Microb. Genom. 2023, 9, 001012. [CrossRef]
- Bakhshi, B.; Kalantar, M.; Rastegar-Lari, A.; Fallah, F. PFGE genotyping and molecular characterization of Campylobacter spp. isolated from chicken meat. Iran J. Vet. Res. 2016, 17, 177–183.
- Davedow, T.; Carleton, H.; Kubota, K.; Palm, D.; Schroeder, M.; Gerner-Smidt, P.; Al-Jardani, A.; Chinen, I.; Kam, K.M.; Smith, A.M.; Nadon, C. PulseNet international survey on the implementation of whole genome sequencing in low and middle-income countries for foodborne disease surveillance. Foodborne Pathog. Dis. 2022, 19, 332–340. [CrossRef]
- Hodges, L.M.; Taboada, E.N.; Koziol, A.; Mutschall, S.; Blais, B.W.; Inglis, G.D.; Leclair, D.; Carrillo, C.D. Systematic evaluation of whole-genome sequencing based prediction of antimicrobial resistance in Campylobacter jejuni and C. coli. Front. Microbiol. 2021, 12, 776967. [CrossRef]
- Lakicevic, B.; Jankovic, V.; Pietzka, A.; Ruppitsch, W. Wholegenome sequencing as the gold standard approach for control of Listeria monocytogenes in the food chain. J. Food Prot. 2023, 86, 100003. [CrossRef]
- Natsos, G.; Mouttotou, N.K.; Ahmad, S.; Kamran, Z.; Ioannidis, A.; Koutoulis, K.C. The genus Campylobacter: detection and isolation methods, species identification & typing techniques. J. Hellenic Vet. Med. Soc. 2019, 70, 1327–1338. [CrossRef]
- Al Amri, A.; Senok, A.C.; Ismaeel, A.Y.; Al-Mahmeed, A.E.; Botta, G.A. Multiplex PCR for direct identification of Campylobacter spp. in human and chicken stools. J. Med. Microbiol. 2007, 56, 1350–1355. [CrossRef]
- Linton, D.; Lawson, A.J.; Owen, R.J.; Stanley, J. PCR detection, identification to species level, and fingerprinting of Campylobacter jejuni and Campylobacter coli direct from diarrheic samples. J. Clin. Microbiol. 1997, 35, 2568–2572. [CrossRef]
- Persson, S.; Olsen, K.E.P. Multiplex PCR for identification of Campylobacter coli and Campylobacter jejuni from pure cultures and directly on stool samples. J. Med. Microbiol. 2005, 54, 1043–1047. [CrossRef]
- Lanzl, M.I.; Zwietering, M.H.; Abee, T.; den Besten, H.M.W. Combining enrichment with multiplex real-time PCR leads to faster detection and identification of Campylobacter spp. in food compared to ISO 10272-1:2017. Food Microbiol. 2022, 108, 104117. [CrossRef]
- Hetman, B.M.; Mutschall, S.K.; Carrillo, C.D.; Thomas, J.E.; Gannon, V.P.J.; Inglis, G.D.; Taboada, E.N. “These aren’t the strains you’re looking for”: Recovery bias of common Campylobacter jejuni subtypes in mixed cultures. Front. Microbiol. 2020, 11, 541. [CrossRef]
- Nayak, R.; Stewart, T.M.; Nawaz, M.S. PCR identification of Campylobacter coli and Campylobacter jejuni by partial sequencing of virulence genes. Moll. Cell. Probes 2005, 19, 187–193. [CrossRef]
- Commission Regulation (EU) 2017/1495. Campylobacter in broiler carcasses. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32017R1495 (accessed on 18 June 2023).
- Dawson, P.; Buyukyavuz, A.; Ionita, C.; Northcutt, J. Effects of DNA extraction methods on the real time PCR quantification of Campylobacter jejuni, Campylobacter coli, and Campylobacter lari in chicken feces and ceca contents. Poult. Sci. 2023, 102, 102369. [CrossRef]
- Dubovitskaya, O.; Seinige, D.; Valero, A.; Reich, F.; Kehrenberg, C. Quantitative assessment of Campylobacter spp. levels with real-time PCR methods at different stages of the broiler food chain. Food Microbiol. 2023, 110, 104152. [CrossRef]
- Ito, S.; Kishimoto, M. Development of a sampling and real-time PCR method for the quantitative detection of Campylobacter spp. in retail chicken meat without DNA extraction. J. Food Prot. 2023, 86, 100028. [CrossRef]
- Reese, K.R.; Elkins, K.M. Simultaneous detection of foodborne pathogens using real-time PCR triplex high-resolution melt assay. Appl. Microbiol. 2022, 2, 453–459. [CrossRef]
- Pacholewicz, E.; Buhler, C.; Wulsten, I.F.; Kraushaar, B.; Luu, H.Q.; Iwobi, A.N.; Huber, I.; Stingl, K. Internal sampling process control improves cultivation-independent quantification of thermotolerant Campylobacter. Food Microbiol. 2019, 78, 53–61. [CrossRef]
- Stingl, K.; Heise, J.; Thieck, M.; Wulsten, I.F.; Pacholewicz, E.; Iwobi, A.N.; Govindaswamy, J.; Zeller-Pérronet, V.; Scheuring, S.; Luu, H.Q.; Fridriksdottir, V.; Gölz, G.; Priller, F.; Gruntar, I.; Jorgensen, F.; Koene, M.; Kovac, J.; Lick, S.; Répérant, E.; Rohlfing, A.; Zawilak-Pawlik, A.; Rossow, M.; Schlierf, A.; Frost, K.; Simon, K.; Uhlig, S.; Huber, I. Challenging the “gold standard” of colony-forming units – Validation of a multiplex real-time PCR for quantification of viable Campylobacter spp. in meat rinses. Int. J. Food Microbiol. 2021, 359, 109417. [CrossRef]
- Wulsten, I.F.; Galeev, A.; Stingl, K. Underestimated survival of Campylobacter in raw milk highlighted by viability real-time PCR and growth recovery. Front. Microbiol. 2020, 11, 1107. [CrossRef]
- Wulsten, I.F.; Thieck, M.; Göhler, A.; Schuh, E.; Stingl, K. Chicken skin decontamination of thermotolerant Campylobacter spp. and hygiene indicator Escherichia coli assessed by viability real-time PCR. Pathogens 2022, 11, 706. [CrossRef]
- Ma, L.; Feng, J.; Zhang, J.; Lu, X. Campylobacter biofilms. Microbiol. Res. 2022, 264, 127149. [CrossRef]
- Kim, S.-H.; Park, C.; Lee, E.-J.; Bang, W.-S.; Kim, Y.-J.; Kim, J.-S. Biofilm formation of Campylobacter strains isolated from raw chickens and its reduction with DNase I treatment. Food Control 2017, 71, 94–100. [CrossRef]
- Laconi, A.; Tolosi, R.; Drigo, I.; Bano, L.; Piccirillo, A. Association between ability to form biofilm and virulence factors of poultry extra-intestinal Campylobacter jejuni and Campylobacter coli. Vet Microbiol. 2023, 282, 109770. [CrossRef]
- Gunther IV, N.W.; Chen, C.-Y. The biofilm forming potential of bacterial species in the genus Campylobacter. Food Microbiol. 2009, 26, 44–51. [CrossRef]
- Kostoglou, D.; Mucka, E.; Andritsos, N.; Giaouris, E. A Combined Study on the Antibiotic Resistance and Biofilm-Forming Abilities of Campylobacter jejuni and Campylobacter coli Isolates from Retail Raw Chicken Samples. In Proceedings of the 4th International Conference on Biofilms (Asia-Pacific Biofilms 2022), Guangzhou, China, 18–23 October 2022.
- Sulaeman, S.; Le Bihan, G.; Rossero, A.; Federighi, M.; Dé, E.; Tresse, O. Comparison between the biofilm initiation of Campylobacter jejuni and Campylobacter coli strains to an inert surface using BioFilm Ring Test. J. Appl. Microbiol. 2010, 108, 1303–1312. [CrossRef]
- Karki, A.B.; Ballard, K.; Harper, C.; Sheaff, R.J.; Fakhr, M.K. Staphylococcus aureus enhances biofilm formation, aerotolerance, and survival of Campylobacter strains isolated from retail meats. Sci. Rep. 2021, 11, 13837. [CrossRef]
- Klančnik, A.; Gobin, I.; Jeršek, B.; Možina, S.S.; Vučkovic, D.; Žnidarič, M.T.; Abram, M. Adhesion of Campylobacter jejuni is increased in association with foodborne bacteria. Microorganisms 2020, 8, 201. [CrossRef]
- Ramić, D.; Klančnik, A.; Možina, S.S.; Dogsa, I. Elucidation of the AI-2 communication system in the food-borne pathogen Campylobacter jejuni by whole-cell-based biosensor quantification. Biosens. Bioelectron. 2022, 212, 114439. [CrossRef]
- The, K.H.; Flint, S.; French, N. Biofilm formation by Campylobacter jejuni in controlled mixed-microbial populations. Int. J. Food Microbiol. 2010, 143, 118–124. [CrossRef]
- Ica, T.; Caner, V.; Istanbullu, O.; Nguyen, H.D.; Ahmed, B.; Call, D.R.; Beyenal, H. Characterization of mono- and mixed-culture Campylobacter jejuni biofilms. Appl. Environ. Microbiol. 2012, 78, 1033–1038. [CrossRef]
- Sterniša, M.; Centa, U.G.; Drnovšek, A.; Remškar, M.; Možina, S.S. Pseudomonas fragi biofilm on stainless steel 9at low temperatures) affects the survival of Campylobacter jejuni and Listeria monocytogenes and their control by a polymer molybdenum oxide nanocomposite coating. Int. J. Food Microbiol. 2023, 394, 110159. [CrossRef]
Figure 1.
Factors affecting the prevalence and distribution of Campylobacter spp. in poultry meat. Factors that are related to each other are grouped together under the same color. As a result, distinct groups of factors referring to sampling (light green), isolation procedure (light blue), confirmation and/or identification (light purple), and biofilm production (light brown) for Campylobacter species are presented. Created with BioRender.com.
Figure 1.
Factors affecting the prevalence and distribution of Campylobacter spp. in poultry meat. Factors that are related to each other are grouped together under the same color. As a result, distinct groups of factors referring to sampling (light green), isolation procedure (light blue), confirmation and/or identification (light purple), and biofilm production (light brown) for Campylobacter species are presented. Created with BioRender.com.
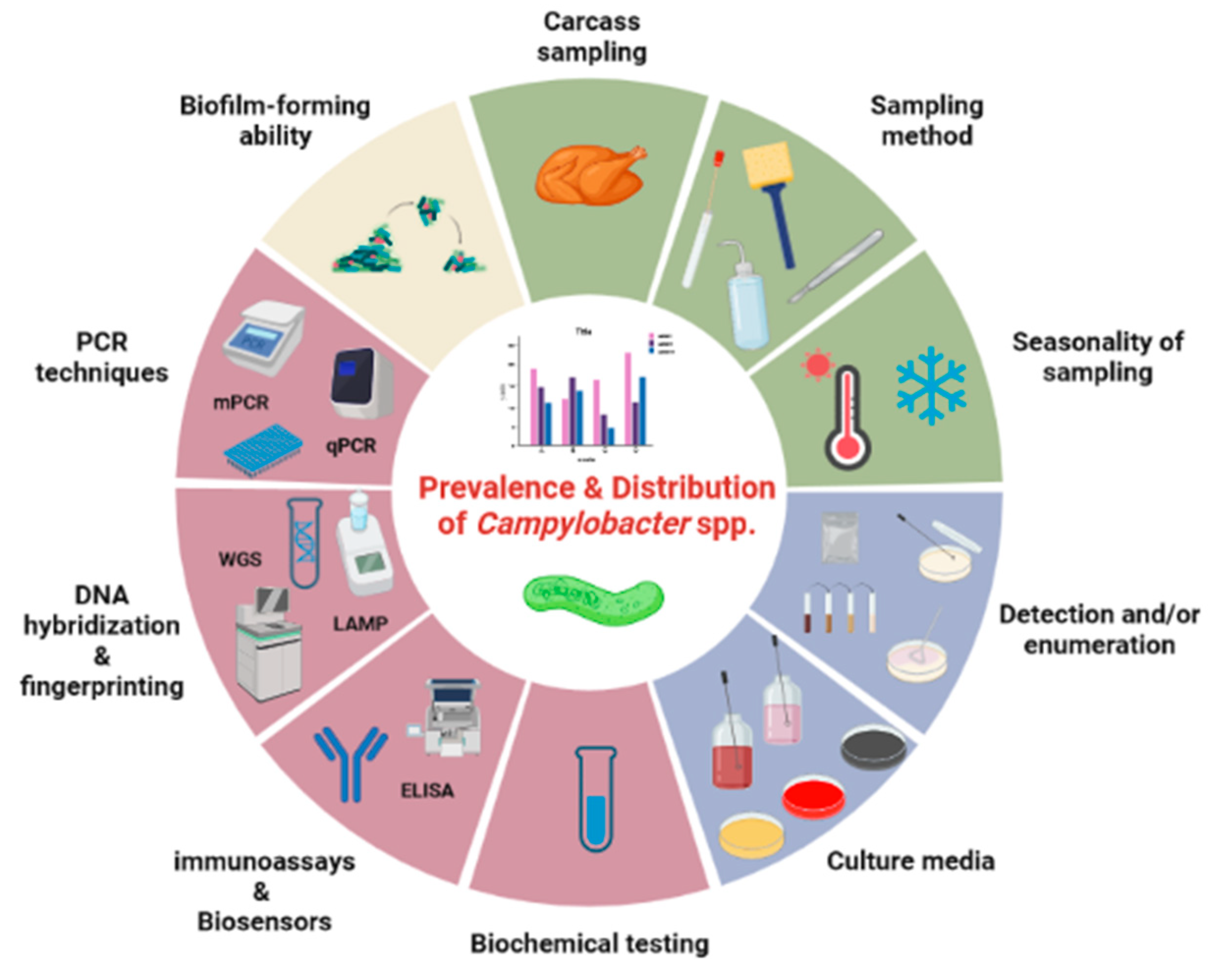
Figure 2.
Sampling sites of a broiler carcass with variations in Campylobacter contamination depicted with the relative concentration of campylobacters (green curved bacteria). Campylobacter concentration in descending order from the more heavily contaminated site to the less contaminated one, when sampled with skin, is as follows: (neck ≥ wing ≥ breast ≥ leg) > (abdomen ≥ back). Parentheses designate the sites that are highly correlated to each other in terms of Campylobacter concentration. Created with BioRender.com.
Figure 2.
Sampling sites of a broiler carcass with variations in Campylobacter contamination depicted with the relative concentration of campylobacters (green curved bacteria). Campylobacter concentration in descending order from the more heavily contaminated site to the less contaminated one, when sampled with skin, is as follows: (neck ≥ wing ≥ breast ≥ leg) > (abdomen ≥ back). Parentheses designate the sites that are highly correlated to each other in terms of Campylobacter concentration. Created with BioRender.com.
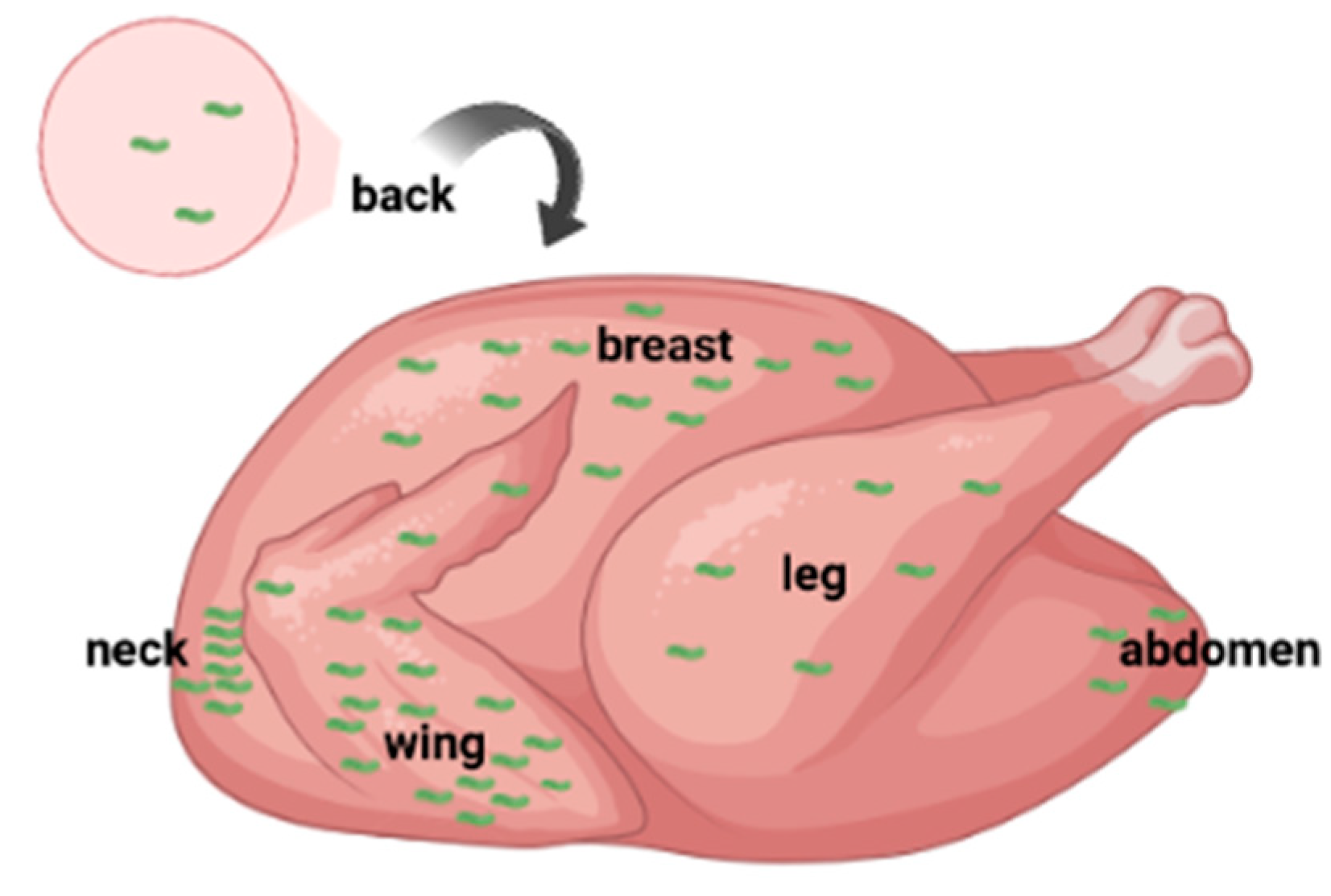

Table 1.
Key biochemical characteristics of Campylobacter species encountered in foods.
| Characteristic | C. jejuni | C. coli | C. lari | C. upsaliensis |
| Catalase activity | + 1 | + | + | - 2 or weak |
| Hippurate hydrolysis | + 3 | - | - | - |
| Indoxyl acetate hydrolysis | + | + | - | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



