
Submitted:
21 June 2023
Posted:
21 June 2023
You are already at the latest version
Abstract
The relationship between living things and their respective environments highly dependent on body temperature regulation. The human capacity to effectively thermoregulate evolved at a time when the environmental temperature was likely around 25°C during the Eocene epoch, some ~ 50-60 million years ago. This effectively meant that homeothermy settled on a core temperature balance point of ~ 37°C. When Homo split from chimpanzee around 5 million years ago the Earth was entering a cooling period where the balance point temperature was always well above that of the environment and body heat balance could be maintained. Following this cooling period, the Earth’s rewarming by 7 °C took over approximately 5,000 years, whereas the current estimates indicate 0.7 °C over the past 100 years; ten times the rate of ice-age-recovery warming, or 20 times faster compared with the last 2 million years. As such, if the predicted continued rise in global temperature continues, and surface temperature reaches values where heat load cannot be dumped as the body temperature balance point is at or near the environmental temperature, areas of the Earth would become inhospitable. This effectively means that we will need to deal with both physiological and behavioral limitations since our ability to adapt will be limited by a thermoregulatory strategy that evolved over millions of years for a different kind of environment, not one that is predicted to change rapidly over the next century. This paper outlines the basis on which Homo settled on a thermoregulatory balance point and what limitations this presents for us in the future.
Keywords:
Adaptation
; balance point
; Earth
; evolution
; Homo
; thermoregulation
1. Introduction
The relationship between living things and their respective environments is complex and highly dependent on their ability to regulate body temperature. For some animals such as fish and reptiles (ectotherms), body temperature is regulated based on the environment around them, whereas for mammals and birds (endotherms) a high body temperature is maintained by balancing internal heat production and offloading the excess heat to the environment. In essence thermoregulation can be characterised as a striking physiological and behavioural characteristic of all living creatures. As far as Homo is concerned, the ability to maintain a core body temperature within narrow limits is the hallmark of a system fined tuned to avoid the potential of a lethal outcome of either a having a very low or very high core temperature. As such, Homo has “settled” to regulate its body core temperature around a balance point of ~37°C [1]. The term balance point is preferred over the popular term “set point” given that there are inputs from multiple variables to establish thermal balance, whereas, set point relates to an engineering preference for a thermostat. The significance of 37 °C is seldom considered as a limitation other than the fact that serious thermal injury might ensue when the body core temperature deviates only a few degrees beyond either side of 37 °C. For this reason, a more considered and informed understanding with respect to the inputs and mechanisms which either contain or cause the deviation from the narrow range is warranted.
Given the relevance of maintaining a balance point for the sustainability of human health and functioning, it is interesting to note that the factors which might have determined how Homo settled on a balance point of 37 °C does not feature to any great extent in either the public health or the human performance literature. A more considered understanding of the evolutionary pressures that shaped the eventuality of Homo’s balance point temperature could also provide for a more meaningful understanding of the capability for human adaptation with respect to a either a changing environment or moving to inhabit a different or more extreme environment.
To this end, it is notable that an increase in global temperature of 5-6 °C is predicted based on present climate data trends, history and models, as the most likely scenario by 2100 [2]. Along with these global temperature predictions is the observation that 37.0% of deaths during warm seasons can be attributed to anthropogenic climate change [3]. In addition, however, is that from 1998 – 2017 the World Health Organization reports that more than 166,000 people died due to heat waves [4], exposing our frailties with respect to high ambient temperatures. Moreover, the risks to physical health associated with climate change are not particularly obvious, at least to the lay public, even though there is evidence that both cold and hot temperature extremes are associated with high mortality rates [5]. Although there is general consideration given to how we might manage the frequency of heat waves [6,7], there is somewhat less consideration given to how Homo might deal with a sustained higher global temperature in the future and whether we have the adaptive capability to manage this possible eventuality, given our strict operational thermoregulatory balance point parameters. Thus, the purpose of this narrative review is to present the possible evolutionary pressures which might have led Homo to settle on a balance point of ~37°C and to exam whether this adaptation will be useful in the face of a new climate with a sustained higher global temperature
2. Climate and the evolution of homeothermy
It is almost impossible to definitively know the precise pressures that determined the evolution of the thermoregulatory strategy employed by Homo. However, a retrospective analysis of the history of early Earth and the observed behaviour of primitive organisms underscores the relationship between the surrounding temperature and survivability. There is consensus that early life on Earth was subjected to a hot environment as outlined independently by Oparin and Haldane in the 1920s [8]. The early Earth was thought to be largely composed of nitrogen, ammonia, methane and hydrogen whereby by the addition of heat produced chemical reactions eventually giving rise to molecules, when combined in water formed what is termed the primordial soup of amino acids and the building blocks of life [9]. A key for the evolution of life was the incremental step from unicellular to multicellular organisms where cellular reactions were required to perform metabolic functions [10]. It follows that a pivotal evolutionary pressure must have been for organisms to respond to environmental challenges which could only occur if structures for cellular responsiveness were available.
In the classic experiment by Mendelssohn (1895) [11] which examined the relationship between the behaviour of unicellular organisms and surrounding temperature, the preferred temperature drove behaviour. Specifically, the paramecia that were bathed in a fluid medium would either disperse or congregate according to their seemingly preferred temperature medium. When their medium was set at 19 °C the paramecia randomly dispersed but when the medium was set at 38 °C congregation would occur at the cooler end between 25-26°C. There is nothing particularly remarkable about this behaviour other than the fact that these organisms do not poses a nervous system with connecting neurons per se but rather is an integrated organism that relates physiology with behaviour [12]. The most notable aspect of this thermoregulatory model is that the paramecia had a preferred temperature between 24-28 °C with avoidance behaviour for the extremes of 12 and 36 °C.
Much of the Earth’s surface temperature is thought to range from 0-50 °C with some species such as fish tolerating temperatures below 0 °C while some algae can function above temperatures of 70 °C [13]. Within this temperature range 0-50 °C , it is now apparent that the average temperature of the Earth has moved over the last 1300 years to now be at least 0.8 °C higher than around the 1980s [14,15]. Figure 1 are data redrawn from Hansen et al. [14] and shows the temperature anomalies between the years 1880-2010. It is evident that since around 1980, the surface temperature of the Earth has increased at a rate of ~0.2 °C per decade. If this trend continues the surface temperature of the Earth could increase by 2.0 °C within the next century. As such, dealing with heat waves may not necessarily be the only difficulty we face but the potential for a new average surface temperature.
The question then is, does Homo have the thermoregulatory adaptive capability to manage this new environmental temperature? To interrogate this question appropriately, an understanding is required for the basis with which the balance point of 37 °C evolved. The basic principle of homeothermy, which is to maintain a constant internal temperature, is that heat loss must be balanced by the corresponding heat production and vice versa. For this kind of system to operate efficiently, the ability to turn heat production on and off is essential and can only occur if the temperature of the organism (i.e. Homo) is sufficiently higher than the surrounding temperature. The critical element of this system is that when the surrounding temperature rises above that of the organism’s then the system will fail. This simple physical requirement suggests that Homo’s choice for a balance point of 37 °C was based on having to deal with the surrounding temperature for the purpose of maintaining appropriate heat loss to the environment and, the capability to increase heat production if heat loss was high. Of course, if heat loss were to be drastic such as with submersion in cold water, then the system would also fail since heat production could not keep up with the heat loss. However, Homo is a land-dwelling mammal and typically avoids cold extreme exposure without sufficient protection.
A number of explanations have been posited as to how and why Homo settled on a balance point of 37 °C. These include preserving proteins from denaturation and thermal coagulation, inactivation of enzymes, reduced oxygen availability, metabolic reactions related to the Q10 effect and, the potential for temperature to effect cellular membrane structure [13,16,17]. These mechanisms all explain how different cellular process operate within the temperature range for Homo and other animals but individually and collectively they cannot explicate how we and other animals “settled” on 37°C rather than regulating at either a higher or lower temperature [18,19]. Further to this, any of these mechanisms would have had to have co-evolved with the “settling” of the balance point temperature.
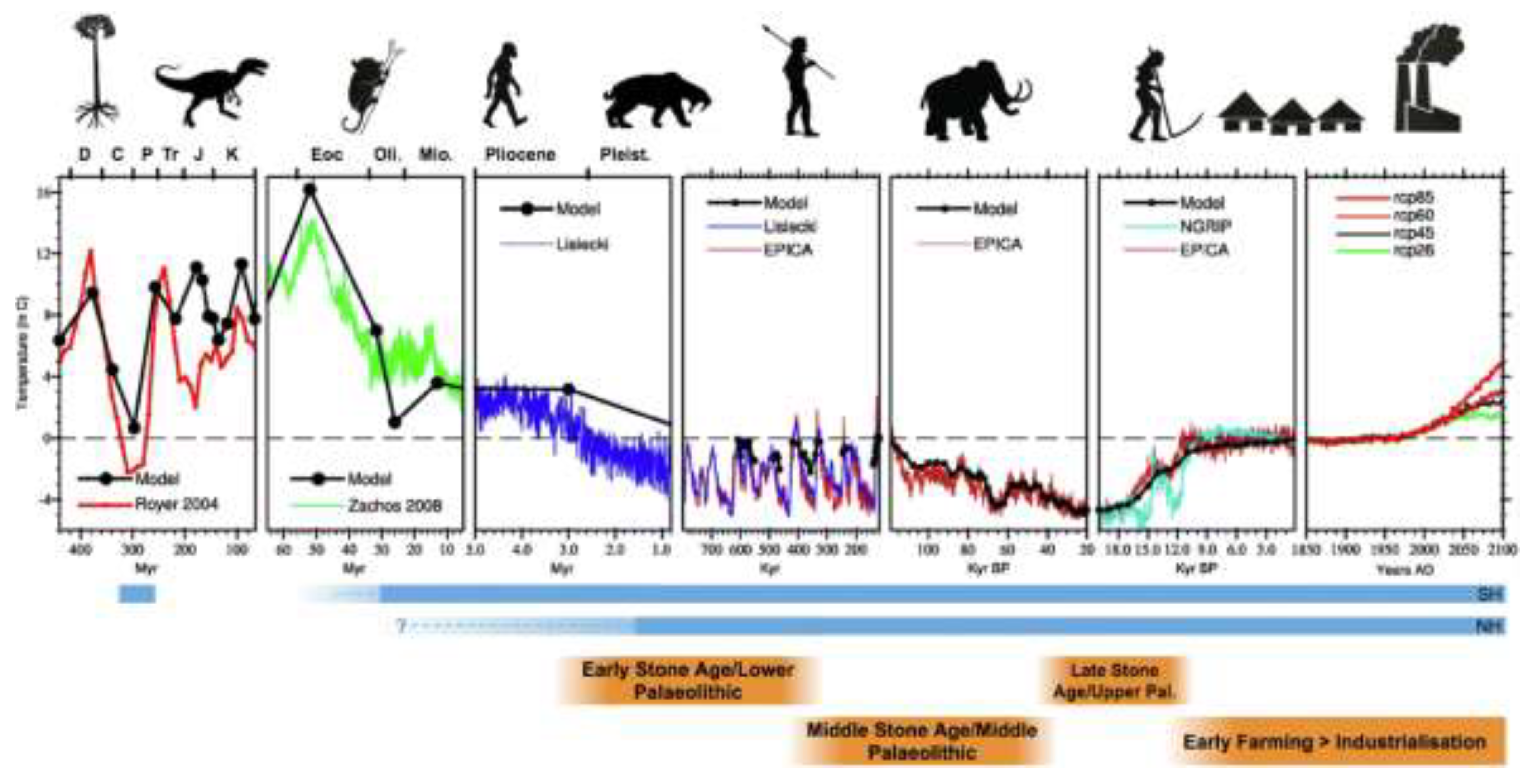
However, to arrive at common balance point of 37°C, an environmental assumption is required since as already indicated, homeothermy is dependent on the temperature of the organism being sufficiently higher than the surrounding temperature. Figure 2 depicts the temperature anomaly modelling from various sources dating back to 400 million years and up to the prediction for 2100 [20]. Since current evidence indicates that the most important evolutionary split occurred between primates with wet and dry noses during the Eocene (~40 million years ago), homeothermy must have developed during this epoch [21,22,23]. Figure 2 shows that the temperature anomaly during the Eocene was likely at least 8 – 12 °C higher than from ~700,000 years ago to the year 2000 which would make the ambient temperature ~ 25 °C. This figure is based on data which indicates that the Earth’s temperature is between 10-15 °C [24,25] (represented by 0 °C dotted line in Figure 2) Of course, the ambient conditions were likely interspersed with cooler periods over a timescale of approximately 50 - 60 million years (Figure 2).
Since the Law of Arrhenius states that heat production increases two-to-threefold with each 10 °C increase, coupled with the assumption that 25 °C was the environmental temperature during the evolution of homeothermy, we are able to arrive at the preferred balance point of 37 °C by the following mathematical model [26]:
The temperature gradient between the body core (Ct) and the assumed environmental temperature is given as:
Ct – 25 °C
If there is a 1 °C increase this would result in,
Ct + 1 °C – 25 °C = Ct – 24 °C
(Ct – 24 °C) / (Ct – 25 °C) = 1.086
In this example, 1.086 represents the Q10 effect where a 1 °C increase in Ct will increase heat production by 2.30.1 (or 1.086). By solving for Ct the result is 36.6 or ~37 °C. Beyond the ambient temperature assumption, the model also fits the underlying assumption that the most desirable core temperature would naturally be higher than the environmental temperature to facilitate transfer of heat away from the core. Figure 2 also indicates that current temperature anomalies commencing in the 1980s are similar to the temperatures at the time when Homo and chimpanzee split from the Last Common Ancestor around 5 million years ago [27]. As such, the current ambient temperature anomalies are likely to still be within our evolutionary limits. Although this scenario may not appear to be alarming, when considering the time taken in the past for global warming to occur, the temperature increased up to 7 °C over approximately 5,000 years, whereas the current estimates indicate 0.7 °C over the past 100 years; ten times the rate of ice-age-recovery warming or 20 times faster compared with the last 2 million years [28].
3. Public health, temperature and climate change
Much has been written regarding the predicted climate change scenario and consequences for human health, but as already alluded to, much of this is related to the frequency of extremes [29,30,31] and the relationship with mortality rates rather than a considered approach to how Homo is equipped to adapt to a new and sustained higher global temperature [32,33,34]. However, the data on the mortality rate during heat extremes is a window into the potential public health issues society might face with a new higher global temperature. The higher rate of mortality during extreme heat is not simply a consequence of a poor thermoregulatory response as it is to complications from previous or ongoing chronic sequalae.
During extreme heat events the main response which can lead to higher rates of mortality is the need for the redistribution of blood towards the skin for enhanced heat transfer from the skin to the environment via the evaporation of sweat. This increased skin blood flow will increase the demand for the heart to increase its rate and contractility to keep up with its own demand for oxygen. This scenario is particularly stressful for those individuals with pre-existing cardiovascular conditions which would place additional strain on the heart simply because there is a need to deal with an overwhelming environmental heat load for an extended time [35]. This increased cardiovascular strain is also a concern for the older population who overwhelmingly suffer from cardiovascular morbidity and are more likely to succumb to cardiovascular induced heat strain leading to mortality during extreme heat events [36]. Given the aging population in many countries where extreme heat waves are expected and the fact that cardiovascular disease is still a global health risk [37], the collision of climate change with these two factors alone pose a great risk to human health that cannot be ignored.
Beyond the cardiovascular morbidity are the potential effects of prolonged exposure to humid heat whereby much needed cooling via evaporation of sweat is minimised or even restricted, potentially leading to prolonged body fluid deficits [38,39]. Prolonged dehydration can have deleterious effects on multiple fronts, and in particular it can reduce blood volume [40] which could result in worsening cardiovascular symptoms. It is also possible that prolonged dehydration can have serious consequences on renal function with ensuing chronic kidney disease, including failure and fibrosis [41]. This has been highlighted as a serious concern for workers in outdoor environments as shown in field workers in Central America [42]. These authors make an important distinction which indicates that renal dysfunction with long exposure to high heat loads can be an outcome of strenuous work and not necessarily due to other comorbidities that might also be related to renal dysfunction (e.g. diabetes mellitus, hypertension).
An often-overlooked health implication associated with climate change is the effect that continuous exposure to high ambient temperatures have on the respiratory system. It is known for instance, that hyperthermia will increase ventilation resulting in the physiological response known as hyperpnea (deeper than normal breaths resulting in increased volume of air in the lungs) [43]. Although this response is typically observed from increases in metabolic demand with exercise, hyperpnea also occurs as a consequence of passive exposure to heat where the threshold for core temperature seems to be as low as ~ 37.8 °C [44]. Given that core temperatures during heat waves can reach as high as 39 °C in ill patients and to a lesser degree in health care workers (~37.5°C) [45], there is a likelihood that sustained exposure to higher heat loads for individuals with pre-existing respiratory conditions will experience respiratory distress due to heat related hyperpnea. An additional and indirect concern with respect to the potential effects of climate change on respiratory health is the rise in CO2 levels which have risen to 440 ppm and suggested to be the highest since recording commenced in 1958 [46]; a higher global temperature may come with a higher level of atmospheric CO2. Although the effect of exposure to high levels of CO2 (> 1000 ppm) on human health are well described [47], the effects of long-term low levels of increased atmospheric CO2 (<1000 ppm) on human respiratory health are not well established. It appears that physiological changes occur at CO2 exposures between 500 – 5000 ppm with cognitive effects evident at 1000 ppm [48]. Evidence from a mouse model now indicate that long term exposure at 890 ppm from preconception to adulthood resulted in significantly impacted respiratory structure and function in female mice [49]. These authors conclude that long-term exposure to the predicted climate change atmospheric CO2 levels can negatively impact lung structure and function in female mice, although the effects seem modest but biologically relevant. Nevertheless, these supposed modest effects might be more relevant in those individuals that have pre-existing respiratory ailments, since climate change will not just increase the chances of developing hyperpnea but exposure to higher CO2 levels, increased pollutants and aeroallergens which could increase the incidence of lung infections, asthma and exacerbations of chronic pulmonary obstructive disorder [50].
- i)
- Limitations for physiological adaptation
There are at least two considerations with respect to adaptive capability for a higher global temperature. The first consideration is related to Homo’s potential for physiological adaptation to a new climate, whereas the second is related to the behavioural adaptation which includes both personal and technological adaptations.
We know from a multitude of exercise acclimatization studies that the ability for Homo to adapt to higher environmental temperatures is significant and can occur within nine days of exposure [51,52,53,54,55,56]. Exercise acclimatization to a higher heat load brings with it many physiological changes which provide improved thermoregulatory responses. These include reduced heart rate for similar workload, increased blood volume and sweat rate, enhanced evaporation potential, decreased salt loss in sweat and a lessened potential to develop heat related illness. It also appears that higher levels of physical fitness provide for higher levels of heat tolerance with acclimation [57]. Therefore, one strategy that can be employed is to improve the physical fitness of the general population which would not only improve general health and wellbeing, but it would also improve the resilience and tolerance for a higher, more sustained environmental heat load.
However, even if the difficulties with deploying a mass population fitness strategy were overcome, it may only serve as a short to medium term intervention. For if we project to the level of heat load expected with a global-mean warming of ~ 7°C, metabolic heat dissipation would be severely hampered, and with warming of 11–12 °C there would be inhospitable zones for humans [58]. The reasons for this reduced evaporative capacity are not particularly obvious to the public but are related to the maximum heat dissipation capacity of the environment and the limitations of our thermoregulatory strategy. When we consider the key environmental inputs (humidity, radiation and air movement) and individual variables such as physical activity and clothing; which determine the physiological strain during any given circumstance [59], it is the air humidity that will ultimately limit the human ability to thermoregulate effectively [60]. This is because heat balance as given by equation 1:
where heat storage (S) is gained or lost (±) through convection and/or conduction (C), radiation (R), metabolic heat (M) and loss through evaporation (E) of sweat, establishes that the major way of losing heat is through evaporation of sweat. The evaporative capacity of the environment is determined by the wet bulb temperature (Tw) and represents the potential upper limit for human heat tolerance [58]. This relationship is determined by the fact that heat cannot be lost to the environment when the object’s temperature is exceeded by the Tw. Since human skin temperature is well regulated at ~35°C or slightly lower, it is hypothesised that a Tw of 35 °C for long periods would be intolerable because the temperature gradient from core-skin-environment would not be sufficient to offload metabolic heat. However, recent experimental modelling across six different humid environments has determined that uncompensable heat stress occurs at Tw significantly lower than 35°C [61]. In simple terms, when the skin temperature rises to be equivalent or above core temperature, offloading metabolic heat is not possible since the water vapour pressure gradient will limit the evaporation of sweat and heat storage will result. Although early modelling suggested that global surface temperatures will increase by an additional 2-2.5°C by 2100 [62], more recent estimates suggest this figure is closer to 5-6°C [2]. Regardless, these estimates indicate that there is a real possibility that the environmental temperatures will not be tolerable, and adaptability will be limited with our current thermoregulatory capability since the rate of change for the environmental temperature is far in excess of what it was when homeothermy was thought to have evolved. Finally, even though Homo’s ability to acclimatize to the heat is well documented and appears to be quite responsive, this is only the case within the environment we currently inhabit. Beyond this, it is very doubtful that our adaptive capacity will serve us if the predicted high global temperatures eventuate [63].
S = C ± R ± C ± M – E
- ii)
- Limitations for behavioural adaptation
It is not controversial to claim that humans will alter their behaviour to accommodate the environment they inhabit. This altered behaviour extends to strategies which will accommodate thermal comfort [64,65,66]. The classic understanding of thermal comfort was proposed by Fanger (1970) [67] who combined six key parameters (ambient and core temperatures, relative humidity, wind velocity, clothing units and metabolic rate) as determinants. However, the behavioural strategies that might be employed are highly dependent on thermal inputs from the core, skin and air temperature along with length of exposure as historically suggested by Gagge et al. 1967 [64]. More recent data indicate that the initiation of thermoregulatory behaviour is not necessarily dependent on changes in temperature but on thermal sensation and discomfort as behavioural controllers [68]. Regardless of the weighting or value of the determinants of thermal comfort, there are limited choices with respect to our behavioural adaptation. That is, manipulating the air flow to affect skin temperature [7] or removing or donning clothing will assist. However, there is complexity in modifying the surroundings as recently highlighted by Jay et al. [33]. For instance, these authors outline that the use of air conditioning is likely to be unsustainable in the future since this will add to power usage and adding further burdens to electricity demands and to the causes of anthropogenic climate change. As an alternative, these authors outline strategies which address the urban landscape and building possibilities which are posited to augment our adaptive capacity to heat extremes and hot weather. The question then is whether society has a) the lead time to make these changes for predicted new thermal environment and, b) the financial capability to enact these physical and structural changes.
Nevertheless, there are individual behavioural strategies that can be enacted which could assist. First, it is known that reducing intensity of physical activity will reduce the rate of rise in body temperature [69] which for manual and outdoor workers would be essential in improving comfort, alleviating the likelihood of thermal injury and loss of productivity [70]. Second, the use of cooling strategies either ahead of, or during exposure to higher ambient temperatures is known to reduce the rate of rise in body temperature and attenuate thermal discomfort [71,72,73,74]. Altering the times when work is conducted could negate the exposure to daytime temperatures or increasing breaks during work time could improve the productivity and reduce the chances of the deleterious effects of exposure to higher heat loads [75]. Any of these strategies and perhaps others could improve health and wellbeing when exposed to higher heat loads. However, their applicability as a long term and sustainable strategy if and when we reach a new higher global temperature is questionable since our capacity to store and lose heat will not have likely been altered.
A final consideration and a less studied area is the effect of night-time temperature on human sleep, which has been shown to be significantly disrupted during summer and among both lower-income and older individuals [76]. It is not uncontroversial to suggest that sleep is paramount for health and that either disrupted, or not enough sleep may lead to other illnesses and psychological distress [77]. The fact that insufficient sleep can negatively influence a myriad of human functions and is associated with increasing prevalence of chronic conditions such as Type 2 diabetes mellitus, obesity and eating disorders [78,79] alone should be cause for concern. If this is overlaid with additional mental health consequences and if sleep patterns are shown to be altered due to temperature anomalies as a corollary of climate change, there is potentially a more complex chronic disorder which intersects both climate, chronic health conditions and mental health. Moreover, recent findings indicate that from 28 cities across Japan, South Korea and China, the increased frequency of hot nights was significantly associated with increased mortality risks, with a prediction that the frequency of hot nights would increase more than 30% and the intensity of hot night would increase by 50% by 2100s [80]. These authors concluded that there is a growing role of night-time warming in heat- related health effects in a changing climate. Therefore, it is not only necessary to modify daytime behaviour, but it appears that behaviour and modifications to our surroundings are also required to avoid the possibility of sleep disruption and its potential for poor health and wellbeing.
4. Conclusions
Homeothermy represents a fine-tuned system which has enabled Homo to interact with the environment and balance heat loss with heat production. This thermoregulatory ~37 °C balance point evolved over millions of years and served well during an extended cooling period of the Earth. However, the magnitude of the rate of increase in warming is precited to be close to 20 times faster than what it was when the Earth was rewarming from the Ice Age. This effectively means that when the surrounding temperature reaches values which impeded the potential to offload heat, areas of the Earth may be uninhabitable by humans and probably other animal life. Our adaptability for this new environment is limited by the thermoregulatory capability and therefore we will require other strategies to mitigate this limitation.
Acknowledgments
The author’s research is supported by the School of Allied Health, Exercise & Sports Sciences, Charles Sturt University.
Conflicts of Interest
There are no conflicts to declare
References
- Romanovsky, A.A. Thermoregulation: some concepts have changed. Functional architecture of the thermoregulatory system. Am J Physiol Regul Integr Comp Physiol. 2007, 292, R37–46. [Google Scholar] [CrossRef] [PubMed]
- Valone, T.F. Linear global temperature correlation to carbon dioxide level, sea level, and innovative solutions to a projected 6 °C Warming by 2100. J Geosci Env Prot. 2021, 09, 84–135. [Google Scholar] [CrossRef]
- Vicedo-Cabrera, A.M.; Scovronick, N.; Sera, F.; et al. The burden of heat-related mortality attributable to recent human-induced climate change. Nat Clim Change 2021, 11, 492–500. [Google Scholar] [CrossRef] [PubMed]
- WHO. Heatwaves. Available online: https://www.who.int/health-topics/heatwaves#tab=tab_1 (accessed on 18 May 2023).
- Gasparrini, A.; Guo, Y.; Hashizume, M.; et al. Mortality risk attributable to high and low ambient temperature: a multi country observational study. Lancet 2015, 386, 369–375. [Google Scholar] [CrossRef]
- Vargas, N.T.; Schlader, Z.J.; Jay, O. , Hunter, A. Prioritize research on human behaviour during extreme heat. Nature Hum Behav. 2023, 7, 473–74. [Google Scholar] [CrossRef] [PubMed]
- Ravanelli, N.M.; Hodder, S.G.; Havenith, G. , Jay, O. Heart rate and body temperature responses to extreme heat and humidity with and without electric fans. JAMA. 2015, 313, 724–25. [Google Scholar] [CrossRef]
- Miller, S.L.; Schopf, J.W.; Lazcano, A. Oparin’s "Origin of Life": Sixty Years Later. J Mol Evol. 1997, 44, 351–353. [Google Scholar]
- Miller, S.L. A production of amino acids under possible primitive earth conditions. Sci. 1953, 117, 528–29. [Google Scholar] [CrossRef]
- Schopf, J.W. The evolution of the earliest cells. Sci Am. 1978, 239, 110–38. [Google Scholar] [CrossRef]
- Mendelssohn, M. Ueber den thermotropismus einzelliger organismen. Pflügers Archiv. 1895, 60, 1–27. [Google Scholar] [CrossRef]
- Brette, R. Integrative neuroscience of Paramecium, a “swimming neuron”. eNeuro 2021, 8, ENEURO.0018–21.2021. [Google Scholar] [CrossRef]
- Schmidt-Nielsen, K. Animal physiology: adaptation and environment. Cambridge: Cambridge University Press; 1997.
- Hansen, J.; Sato, M.; Ruedy, R.; Lo, K.; Lea, D.W. , Medina-Elizade, M. Global temperature change. PNAS. 2006, 103, 14288–93. [Google Scholar] [CrossRef] [PubMed]
- Mann, M.E.; Zhang, Z.; Hughes, M.K.; et al. Proxy-based reconstructions of hemispheric and global surface temperature variations over the past two millennia. PNAS. 2008, 105, 13252–57. [Google Scholar] [CrossRef]
- Crompton, A.W.; Taylor, C.R. , Jagger, J.A. Evolution of homeothermy in mammals. Nat. 1978, 272, 333. [Google Scholar] [CrossRef] [PubMed]
- Pörtner, H.O. Climate variability and the energetic pathways of evolution: the origin of endothermy in mammals and birds. Physiol Biochem Zool. 2004, 77, 959–81. [Google Scholar] [CrossRef] [PubMed]
- Marino, F.E. The evolutionary basis of thermoregulation and exercise performance. In: Marino, F.E., editor. Thermoregulation and human performance: physiological and biological aspects. Basel: Karger Publishers; 2008:1–13.
- Marino, F.E. Human Fatigue: evolution, health and performance. London: Routledge; 2019.
- Haywood, A.M.; Valdes, P.J.; Aze, T.; et al. What can Palaeoclimate Modelling do for you. Earth Sys Environ. 2019, 3, 1–18. [Google Scholar] [CrossRef]
- Zachos, J.; Pagani, M.; Sloan, L.; Thomas, E.; Billups, K. Trends, rhythms, and aberrations in global climate 65 Ma to present. Sci. 2001, 292, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Franzen, J.L.; Gingerich, P.D.; Habersetzer, J.; Hurum, J.H.; von Koenigswald, W.; Smith, B.H. Complete primate skeleton from the Middle Eocene of Messel in Germany: morphology and paleobiology. PLOS ONE 2009, 4, e5723. [Google Scholar] [CrossRef]
- Tudge, C. The link: uncovering our earliest ancestor. Little, Brown; 2009.
- A. Muller, R.; Rohde, R.; Jacobsen, R.; Muller, E., Wickham, C. A new estimate of the average Earth surface land temperature spanning 1753 to 2011. A new estimate of the average Earth surface land temperature spanning 1753 to 2011. 2013, 01.
- Communication, N.A.S.A.P.S. Solar system temperatures. Available online: https://solarsystem.nasa.gov/resources/681/solar-system-temperatures/#:~:text=Earth%20%2D%2059%C2%B0F%20(15%C2%B0C) (accessed on 13 June 2023).
- Gisolfi, C.V.; Mora, M.T.; Mora, F., Teruel, F.M. The hot brain: survival, temperature, and the human body. Cambridge, MA: MIT Press; 2000.
- Lieberman, D.E. Human locomotion and heat loss: an evolutionary perspective. Comp Physiol. 2015, 5, 99–117. [Google Scholar]
- Observatory, N.A.S.A.E. How is today’s warming different from the past. Available online: https://earthobservatory.nasa.gov/features/GlobalWarming/page3.php#:~:text=As%20the%20Earth%20moved%20out,ice%2Dage%2Drecovery%20warming (accessed on 12 June 2023).
- Tong, S.; Wang, X.Y.; Yu, W.; Chen, D. , Wang, X. The impact of heatwaves on mortality in Australia: a multicity study. BMJ Open 2014, 4, e003579. [Google Scholar]
- Sun, Z.; Chen, C.; Yan, M.; et al. Heat wave characteristics, mortality and effect modification by temperature zones: a time-series study in 130 counties of China. In J Epidemiol 2021, 49, 1813–1822. [Google Scholar]
- Franklin, R.C.; Mason, H.M.; King, J.C.; et al. Heatwaves and mortality in Queensland 2010-2019: implications for a homogenous state-wide approach. Int J Biometeriol 2023, 67, 503–515. [Google Scholar]
- Ebi, K.L.; Capon, A.; Berry, P.; et al. Hot weather and heat extremes: health risks. Lancet 2021, 398, 698–708. [Google Scholar]
- Jay, O.; Capon, A.; Berry, P.; et al. Reducing the health effects of hot weather and heat extremes: from personal cooling strategies to green cities. Lancet 2021, 398, 709–724. [Google Scholar]
- Vanos, J.K.; Baldwin, J.W.; Jay, O. , Ebi, K.L. Simplicity lacks robustness when projecting heat-health outcomes in a changing climate. Nat Comm 2020, 11, 6079. [Google Scholar]
- Kenny, G.P.; Yardley, J.; Brown, C.; Sigal, R.J. , Jay, O. Heat stress in older individuals and patients with common chronic diseases. CMAJ. 2010, 182, 1053–60. [Google Scholar] [CrossRef]
- Bunker, A.; Wildenhain, J.; Vandenbergh, A.; et al. Effects of air temperature on climate-sensitive mortality and morbidity outcomes in the elderly; a systematic review and meta-analysis of epidemiological evidence. Ebio Med 2016, 6, 258–268. [Google Scholar]
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990-2019: Update From the GBD 2019 Study. J Am Coll Card 2020, 76, 2982–3021. [Google Scholar]
- Kamijo, Y.-i. , Nose, H. Heat illness during working and preventive considerations from body fluid homeostasis. Indust Hlth. 2006, 44, 345–58. [Google Scholar] [CrossRef]
- Patterson, M.J.; Stocks, J.M. , Taylor, N.A. Whole-body fluid distribution in humans during dehydration and recovery, before and after humid-heat acclimation induced using controlled hyperthermia. Acta Physiol. 2014, 210, 899–912. [Google Scholar] [CrossRef] [PubMed]
- Coyle, E.F. Cardiovascular drift during prolonged exercise and the effects of dehydration. Int J Sports Med. 1998, 19, S121–24. [Google Scholar] [CrossRef] [PubMed]
- Roncal-Jimenez, C.; Lanaspa, M.A.; Jensen, T.; Sanchez-Lozada, L.G. , Johnson, R.J. Mechanisms by which dehydration may lead to chronic kidney disease. Ann Nut Metab. 2015, 66 Suppl 3, 10–13. [Google Scholar] [CrossRef]
- García-Trabanino, R.; Jarquín, E.; Wesseling, C.; et al. Heat stress, dehydration, and kidney function in sugarcane cutters in El Salvador–a cross-shift study of workers at risk of Mesoamerican nephropathy. Env Res. 2015, 142, 746–55. [Google Scholar] [CrossRef]
- Cabanac, M. , White, M.D. Core temperature thresholds for hyperpnea during passive hyperthermia in humans. Eur J Appl Physiol. 1995, 71, 71–76. [Google Scholar] [CrossRef]
- Fujii, N.; Honda, Y.; Hayashi, K.; Kondo, N.; Koga, S. , Nishiyasu, T. Effects of chemoreflexes on hyperthermic hyperventilation and cerebral blood velocity in resting heated humans. Exp Physiol. 2008, 93, 994–1001. [Google Scholar] [CrossRef]
- Stéphan, F.; Ghiglione, S.; Decailliot, F.; Yakhou, L.; Duvaldestin, P. , Legrand, P. Effect of excessive environmental heat on core temperature in critically ill patients. An observational study during the 2003 European heat wave. Br J Anaesth. 2005, 94, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Showstack, R. Carbon dioxide tops 400 ppm at Mauna Loa, Hawaii. Eos Trans Am Geophys Un. 2013, 94, 192–192. [Google Scholar] [CrossRef]
- Jacobson, T.A.; Kler, J.S.; Hernke, M.T.; Braun, R.K.; Meyer, K.C. , Funk, W.E. Direct human health risks of increased atmospheric carbon dioxide. Nat Sustain. 2019, 2, 691–701. [Google Scholar] [CrossRef]
- Azuma, K.; Kagi, N.; Yanagi, U.; Osawa, H. Effects of low-level inhalation exposure to carbon dioxide in indoor environments: A short review on human health and psychomotor performance. Environ Int. 2018, 121, 51–56. [Google Scholar] [CrossRef]
- Larcombe, A.N.; Papini, M.G.; Chivers, E.K.; Berry, L.J.; Lucas, R.M.; Wyrwoll, C.S. Mouse Lung Structure and Function after Long-Term Exposure to an Atmospheric Carbon Dioxide Level Predicted by Climate Change Modeling. Environ Health Perspect. 2021, 129, 17001. [Google Scholar] [CrossRef] [PubMed]
- Joshi, M.; Goraya, H.; Joshi, A.; Bartter, T. Climate change and respiratory diseases: a 2020 perspective. Curr Opin Pulmon Med. 2020, 26, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Pandolf, K.B.; Burse, R.L.; Goldman, R.F. Role of physical fitness in heat acclimatisation, decay and reinduction. Ergo 1977, 20, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Havenith, G.; van Middendorp, H. The relative influence of physical fitness, acclimatization state, anthropometric measures and gender on individual reactions to heat stress. Eur J Appl Physiol. 1990, 61, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, B.; Hales, J.R.; Strange, S.; Christensen, N.J.; Warberg, J.; Saltin, B. Human circulatory and thermoregulatory adaptations with heat acclimation and exercise in a hot, dry environment. J Physiol. 1993, 460, 467–485. [Google Scholar] [CrossRef] [PubMed]
- Andres, T.; Hexamer, M.; Werner, J. Heat acclimation of humans: hot environment versus physical exercise. J Therm Biol. 2000, 25, 139–142. [Google Scholar] [CrossRef]
- Wingfield, G.L.; Gale, R.; Minett, G.M.; Marino, F.E. , Skein, M. The effect of high versus low intensity heat acclimation on performance and neuromuscular responses. J Therm Biol. 2016, 58, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.H.; Fujiwara, M.; Gerrett, N.; et al. The effect of seasonal acclimatization on whole body heat loss response during exercise in a hot humid environment with different air velocity. J Appl Physiol. 2021, 131, 520–31. [Google Scholar] [CrossRef]
- Cheung, S.S.; McLellan, T.M. Heat acclimation, aerobic fitness, and hydration effects on tolerance during uncompensable heat stress. J Appl Physiol. 1998. [Google Scholar] [CrossRef]
- Sherwood, S.C. , Huber, M. An adaptability limit to climate change due to heat stress. PNAS. 2010, 107, 9552–55. [Google Scholar] [CrossRef]
- Budd, G.M. Wet-bulb globe temperature (WBGT)--its history and its limitations. J Sci Med Sport. 2008, 11, 20–32. [Google Scholar] [CrossRef]
- Nielsen, B. Olympics in Atlanta: a fight against physics. Med Sci Sports Exerc. 1996, 28, 665–68. [Google Scholar] [CrossRef]
- Vecellio, D.J.; Wolf, S.T.; Cottle, R.M.; Kenney, W.L. Evaluating the 35°C wet-bulb temperature adaptability threshold for young, healthy subjects (PSU HEAT Project). J Appl Physiol 2022, 132, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, A.P.; Stone, P.H.; Forest, C.E.; et al. Probabilistic forecast for twenty-first-century climate based on uncertainties in emissions (without policy) and climate parameters. J Clim. 2009, 22, 5175–5204. [Google Scholar] [CrossRef]
- Hanna, E.; Tait, P. Limitations to thermoregulation and acclimatization challenge: human adaptation to global warming. Int J Environ Res Public Health 2015, 12, 8034–8074. [Google Scholar] [CrossRef] [PubMed]
- Gagge, A.P.; Stolwijk, J.A.J. , Hardy, J.D. Comfort and thermal sensations and associated physiological responses at various ambient temperatures. Environ Res. 1967, 1, 1–20. [Google Scholar] [CrossRef] [PubMed]
- van Hoof, J. Forty years of Fanger’s model of thermal comfort: comfort for all. Indoor Air. 2008, 18, 182–201. [Google Scholar] [CrossRef]
- Kingma, B.R.; Frijns, A.J.; Schellen, L.; van Marken Lichtenbelt, W.D. Beyond the classic thermoneutral zone: Including thermal comfort. Temp. 2014, 1, 142–149. [Google Scholar] [CrossRef]
- Fanger, P.O. Thermal comfort. Analysis and applications in environmental engineering. Copenhagen: Danish Technical Press.; 1970.
- Schlader, Z.J.; Simmons, S.E.; Stannard, S.R. , Mündel, T. The independent roles of temperature and thermal perception in the control of human thermoregulatory behavior. Physiol Behav. 2011, 103, 217–24. [Google Scholar] [CrossRef]
- Gant, N.; Williams, C.; King, J.; Hodge, B.J. Thermoregulatory responses to exercise: relative versus absolute intensity. J Sports Sci. 2004, 22, 1083–1090. [Google Scholar] [CrossRef]
- Zander, K.K.; Botzen, W.J.W.; Oppermann, E.; Kjellstrom, T. , Garnett, S.T. Heat stress causes substantial labour productivity loss in Australia. Nat Clim Change. 2015, 5, 647–51. [Google Scholar] [CrossRef]
- Booth, J.; Marino, F. , Ward, J.J. Improved running performance in hot humid conditions following whole body precooling. Med Sci Sports Exerc. 1997, 29, 943–49. [Google Scholar] [CrossRef] [PubMed]
- Duffield, R.; Green, R.; Castle, P.; Maxwell, N. Precooling can prevent the reduction of self-paced exercise intensity in the heat. Med Sci Sports Exerc. 2010, 42, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Minett, G.M.; Duffield, R.; Marino, F.E. , Portus, M. Volume-dependent response of precooling for intermittent-sprint exercise in the heat. Med Sci Sports Exerc. 2011, 43, 1760–69. [Google Scholar] [CrossRef]
- Choo, H.C.; Nosaka, K.; Peiffer, J.J.; Ihsan, M.; Abbiss, C.R. Ergogenic effects of precooling with cold water immersion and ice ingestion: A meta-analysis. Eur J Sport Sci 2018, 18, 170–181. [Google Scholar] [CrossRef]
- Hunt, A.P.; Brearley, M.; Hall, A. , Pope, R. Climate Change Effects on the Predicted Heat Strain and Labour Capacity of Outdoor Workers in Australia. Int J Environ Res Public Health. 2023, 20, 5675. [Google Scholar] [CrossRef]
- Obradovich, N.; Migliorini, R.; Mednick, S.C. , Fowler, J.H. Nighttime temperature and human sleep loss in a changing climate. Sci Adv. 2017, 3, e1601555. [Google Scholar] [CrossRef]
- Benjamin, C.L.; Curtis, R.M.; Huggins, R.A.; et al. Sleep Dysfunction and Mood in Collegiate Soccer Athletes. Sports Health. 2020, 12, 234–40. [Google Scholar] [CrossRef]
- Knutson, K.L.; Ryden, A.M.; Mander, B.A. , Van Cauter, E. Role of sleep duration and quality in the risk and severity of type 2 diabetes mellitus. Arch Int Med. 2006, 166, 1768–74. [Google Scholar] [CrossRef]
- Allison, K.C.; Spaeth, A. , Hopkins, C.M. Sleep and eating disorders. Curr Psych Rep. 2016, 18, 1–8. [Google Scholar] [CrossRef]
- He, C.; Kim, H.; Hashizume, M.; et al. The effects of night-time warming on mortality burden under future climate change scenarios: a modelling study. Lancet Planet Health 2022, 6, e648–57. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Global land-ocean temperature anomaly (°C) relative to 1951–1980 from surface air measurements at meteorological stations and ship and satellite measurements. Points represent the annual mean. Data in the figure are redrawn from [14].
Figure 1.
Global land-ocean temperature anomaly (°C) relative to 1951–1980 from surface air measurements at meteorological stations and ship and satellite measurements. Points represent the annual mean. Data in the figure are redrawn from [14].
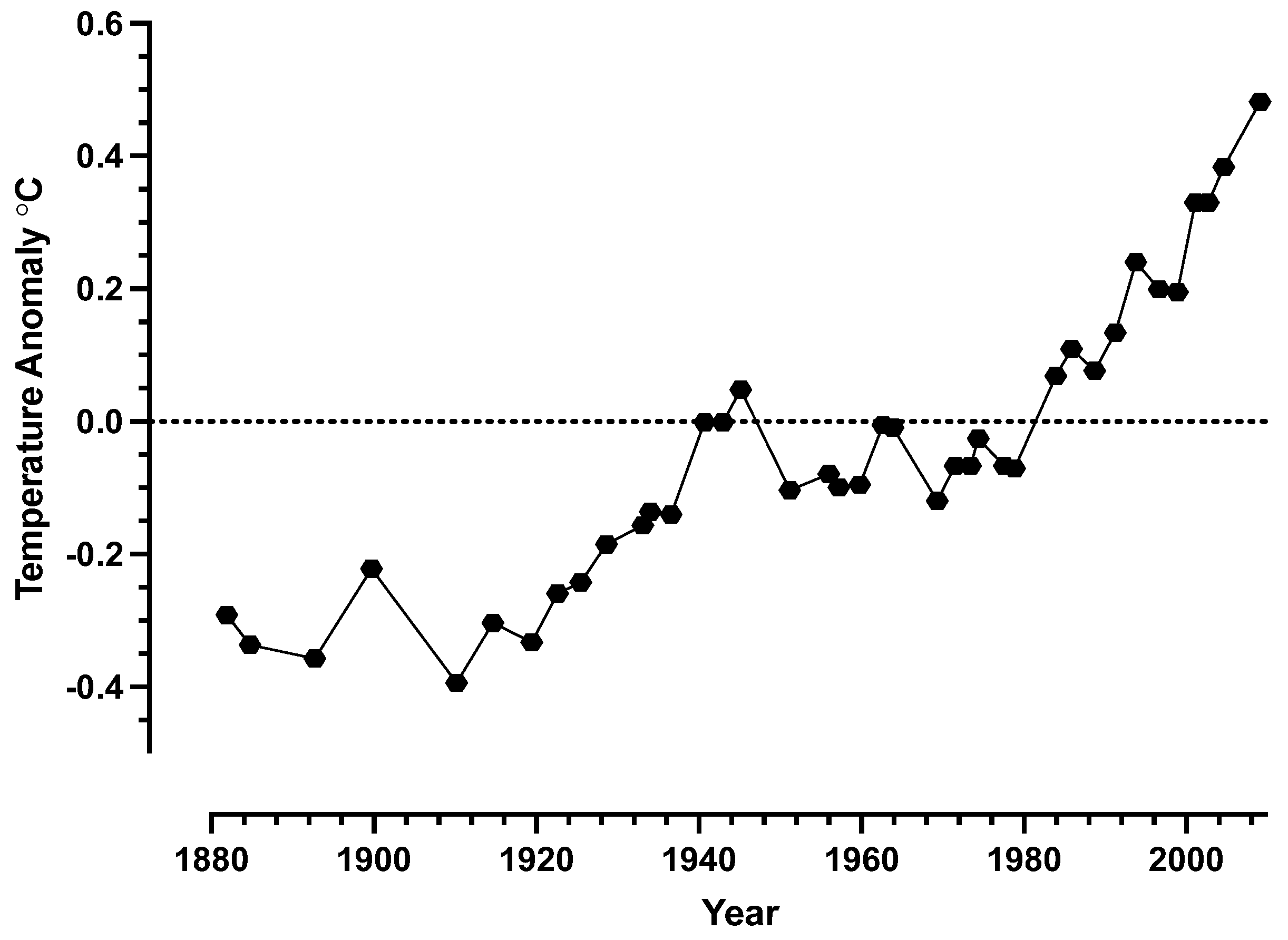
Figure 2.
Global annual mean temperature variation of the Earth through time (last 400 million years) predicted by the Hadley Centre Coupled Climate Model, compared with geologically derived estimates of temperature variability over the same period. Geological epochs include the Devonian (D), Carboniferous (C) Permian (P), Triassic (Tr), Jurassic (J) Cretaceous (K), Eocene (Eoc), Oligocene (Oli.), Miocene (Mio), Pliocene and Pleistocene (Pleist.). Major evolutionary characteristics and events over the last 400 million years represented by cartoon silhouettes. Figure reproduced from [20] under the Creative Commons CC BY license.
Figure 2.
Global annual mean temperature variation of the Earth through time (last 400 million years) predicted by the Hadley Centre Coupled Climate Model, compared with geologically derived estimates of temperature variability over the same period. Geological epochs include the Devonian (D), Carboniferous (C) Permian (P), Triassic (Tr), Jurassic (J) Cretaceous (K), Eocene (Eoc), Oligocene (Oli.), Miocene (Mio), Pliocene and Pleistocene (Pleist.). Major evolutionary characteristics and events over the last 400 million years represented by cartoon silhouettes. Figure reproduced from [20] under the Creative Commons CC BY license.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



