Submitted:
13 June 2023
Posted:
13 June 2023
You are already at the latest version
Abstract
Fungal infections pose a significant threat to human health, particularly in immunocompromised individuals. Due to several factors, such as the inappropriate use of antimicrobial drugs, the emergence of resistance in microorganisms, and increasing demand for less cytotoxic antifungal agents for immunocompromised patients, antifungal research has gained significant attention in recent years. Antifungal peptides, and among them, cyclic peptides, are a class of antifungal agents that have been under development since the 1960s. Using cyclic peptides as a promising approach for combating antifungal resistance in pathogenic fungi has gained significant attention in recent years. The identification of antifungal cyclic peptides from various sources has been possible due to the widespread interest in peptide research in recent decades. It is increasingly important to evaluate the narrow to broad-spectrum antifungal activity and the mode of action of synthetic and natural cyclic peptides for both synthesized and extracted peptides. This short review aims to highlight some of the antifungal cyclic peptides isolated from bacteria, fungi, and plants, taking into consideration the progress made in peptide research. Additionally, it provides a summary of synthetic cyclic peptides with antifungal activity and discusses the potential future of utilizing combinations of antifungal peptides from different sources. The review underscores the need for further exploration of the novel therapeutic applications of these abundant and diverse cyclic peptides.
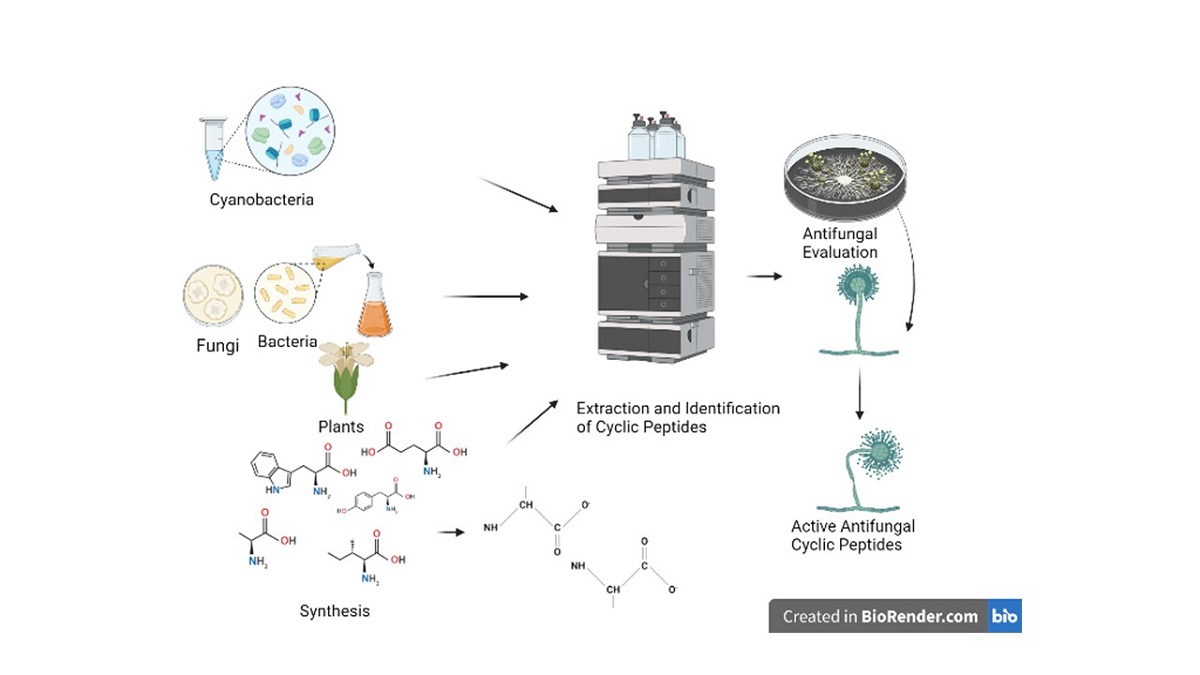
Keywords:
antifungal
; bacteria
; cyanobacteria
; cyclic peptides
; fungi
1. Fungal Infections
In the past few years, there has been a notable surge in the focus on antifungal research. This increased attention can be attributed to several factors, including the improper utilization of antimicrobial drugs, the rise of resistance among microorganisms, and the escalating need for less cytotoxic antifungal agents to cater to immunocompromised patients. A few fungal infections are a significant concern for human health, especially in immunocompromised individuals.
Candidiasis, the most common opportunistic yeast infection, is caused by Candida species. These complicated fungal infections are caused by Candida species and other microorganisms, with Candida albicans (C. albicans) being the most prevalent. Excessive growth in the gastrointestinal, urinary, and respiratory tracts is not limited to immunocompromised patients but also occurs in healthy individuals, frequently resulting in nosocomial infections [1].
Aspergillosis is caused by Aspergillus terreus, Aspergillus niger, Aspergillus flavus, and Aspergillus fumigatus. Allergic syndromes to chronic pulmonary conditions and invasive infections are the consequences of the disease spectrum produced by Aspergillus species. Immunocompromised patients often face significant morbidity and mortality due to invasive aspergillosis, which is a leading cause of these adverse outcomes. The risk of aspergillosis has been observed to increase in patients with post-influenza infection and after COVID-19 infection [2].
Fusarium species, comprising Fusarium solani, Fusarium oxysporum, Fusarium moniliforme, and Fusarium verticilloides, are the predominant filamentous pathogens found in high-risk patients. Among these species, Fusarium solani, originally a plant pathogen, is responsible for nearly 50% of fungal infections, which are also attributed to other opportunistic Fusarium species [3]. Thus, several antifungal agents have been developed to circumvent these pathogens and fungal infections.
2. Antifungal Agents
Antifungal agents are comparable to antibiotics in their ability to inhibit growth, either reversibly (fungistatic versus bacteriostatic) or irreversibly (fungicidal versus bactericidal). These agents work by disrupting essential reactions involved in fungal growth. This may include interference with biosynthetic pathways responsible for building blocks, coenzymes, and macromolecules such as nucleic acids and proteins. Additionally, antifungal agents may target the maintenance and synthesis of cellular structures, such as the cell wall or cell membrane of pathogenic fungi. Understanding the mode of action of an antifungal agent involves discovering the process by which growth is inhibited, and reactions are blocked [4]. Antifungal agents can target fungal sterols, nucleic acid, and β-glucan synthase, as well as act through protein synthesis pathways, inhibition of N-myristylation of fungal proteins, depolarization of mitochondrial membranes, nucleus or endoplasmic reticulum, and alteration of fungal cell membrane distribution [5].
Antifungal peptides primarily exert their mode of action on the cell membrane, cell wall, and other intracellular targets. Figure 1 summarizes the targets of antifungal peptides. The fungal cell wall is the first line of defense against the external environment and helps to regulate osmotic pressure. The inhibitory action of antifungal peptides on chitin synthase (CHS) and (1–3)-β-D-glucan synthase leads to the impairment of cell wall synthesis and disrupting normal cell morphology. This weakens the fungal cell's ability to regulate osmotic pressure. Antifungal peptides can be selective in targeting the fungal cell membrane due to structural differences between plant and fungal cell membranes. This contributes to their effectiveness as plant antifungal agents. Antifungal peptides can also target intracellular substances, including mitochondrial membranes, proteins, and nucleotides such as DNA and RNA. The selective targeting and characterization of antifungal targets provides a useful tool for designing effective antifungal agents, as noted by Zhang et al. [6].
3. Cyclic Peptides Development as Antifungal Agents
Cyclic peptides are a distinct group of cyclic compounds composed of amino acids renowned for their distinctive architectures and extensive repertoire of biological activities. They arise from the linkage of multiple natural or unnatural amino acids in a precise sequence, creating a cyclic structure. These amino acids are intricately connected through amide or other chemically stable bonds, which can form between the C- and N-termini of the peptide sequence or between the peptide's head and tail [7].
Compared to linear peptides, cyclic peptides offer several advantages. Firstly, under physiological conditions where spontaneous degradation may occur, peptide bonds are susceptible to proteolytic degradation. Converting peptides to their cyclic form enhances their in vivo stability when compared to their linear counterparts. This cyclization process has been observed to extend the integrity of peptides with unstable motifs in their sequence. Thus, cyclic peptides demonstrate slower metabolism due to their high resistance to proteases, resulting in a longer reservoir effect compared to linear peptides [8].
Moreover, although the conformational flexibility of macrocyclic structures is limited, it brings about a reduction in the entropy value of drug target binding. Interestingly, this limitation enhances the binding stability of cyclic peptides, leading to higher affinity and recognition specificity between the cyclic peptides and the target protein [9].
Cyclic peptides possess unique characteristics that make them suitable as antimicrobial agents, including high potency, low toxicity, and specificity. This review primarily emphasizes the identification of cyclic peptides derived from various sources and their preliminary evaluation of biological effects, along with future prospects and potential applications. This review will specifically highlights the potential of cyclic peptides from various sources, such as bacteria, fungi, plants, and synthesis, as antifungal agents. This concise review does not intend to provide an exhaustive compilation of all known antifungal cyclic peptides. Instead, its purpose is to present diverse examples of cyclic peptides with antifungal properties sourced from various origins. Our aim is to offer the scientific community insights into the potential applications of these compounds as antifungal agents. Moreover, the inclusion of commercially available cyclic antifungal peptides strengthens the notion that cyclic peptides can serve as a valuable resource for the development of antifungal drugs.
The antifungal peptide Bacillomycin was obtained from Bacillus subtilis in the year 1948 through isolation [10]. Since 1960, there has been ongoing development of antifungal peptides. Among these, Mycobacillin was discovered in 1963 from the biosyntheis in Bacillus subtilis [11]. Valinomycin was discovered in the early 1960s as a cyclodepsipeptide from a species of Streptomyces [12], while Syringomycin E, a lipodepsipeptide, was produced from Pseudomonas syringae pv. Syringae [13,14]. Another example is the antifungal cyclic decapeptide called calophycin [15], which was isolated from the terrestrial blue-green alga Calothrix fusca. Furthermore, Aculeacin A, a cyclopeptide that incorporates a long-chain fatty acid, was obtained from the mycelial cake of Aspergillus aculeatus M-4214 [16,17,18]. Chemical structures of Valinomycin, Syringomycin E, calophycin, Aculeacin A are illustrated in Figure 2.
In the last two decades, three cyclic hexapeptides, caspofungin, micafungin and anidulafungin, with antifungal properties have received approval for clinical use (Figure 2). All three antifungal agents have demonstrated clinical efficacy in treating invasive candidiasis and various types of systemic fungal infections [19]. These three agents target the 1,3-β-glucan synthase involved in the cell wall glucan synthesis.
Caspofungin received approval in the year 2001, while micafungin was granted approval in 2005. These compounds possess a common feature in their structure, which is a shared peptide core comprising six amino acids. Among these amino acids, two are threonine or threonine derivatives, while the remaining two are proline derivatives [20].
Micafungin, a semi-synthetic analog of the natural cyclic hexapeptide FR901379 [21], demonstrates remarkable antifungal potency against Aspergillus spp. and Candida spp. [22]. It is characterized by a linear pharmacokinetic profile and a significantly lower occurrence of adverse reactions compared to amphotericin B. These attributes make micafungin highly valuable for effectively treating deep-seated fungal infections. In both adult) and pediatric patients, micafungin exhibits absorption pharmacokinetics that are linear and dose-proportional across a wide dosage range [23]. However, micafungin differentiates itself from the other two compounds by having low oral bioavailability and being exclusively administered via parenteral routes. It possesses a half-life of approximately 15 h. The majority of micafungin undergoes metabolism and is excreted through the feces as a result of enzymatic modifications in its side chains [24].
Caspofungin, approved for the treatment of yeast and fungal infections under specific conditions in 2001 [25], is a semi-synthetic analog of pneumocandin B0. Pneumocandin B0 is a lipophilic cyclic peptide initially derived from the fungus Glarea lozoyensis in 1985 [26]. It exhibits a half-life of around nine days and is primarily metabolized through metabolic hydrolysis and N-acetylation, causing the cyclic ring to open. Micafungin, in contrast, is a semi-synthetic derivative of a natural cyclic hexapeptide. The enzymatic and chemical deacylation processes of the N-terminal palmitoyl group result in the formation of the optimal N-acyl isoxazole analog [27].
Anidulafungin, is a natural fermentation product of Aspergillus oryzae, and is derived from echinocandin B through diacylation to eliminate the linoleoyl side chain, followed by reacylation with the lipophilic terphenylacyl chain after three additional steps [20,28]. Anidulafungin exhibits clinical effectiveness in the inhibition of invasive candidiasis by employing a mechanism of noncompetitive inhibition on β-(1,3)-glucan synthase [29]. Furthermore, anidulafungin demonstrates a low oral bioavailability ranging from 2% to 7%, necessitating non-intestinal administration [30]. With a half-life of 27 h, anidulafungin undergoes metabolic conversion into linear peptides primarily through open-loop hydrolysis.
The sources of cyclic antifungal peptides vary and can include (a) cyanobacteria and other bacteria, (b) fungi, (c) plants, and (d) synthetic methods. The following sections will provide examples of cyclic peptides from each of these sources.
3.1. Cyclic Peptides Isolated From Cyanobacteria
Cyanobacteria are a type of blue-green algae that are known to produce biologically active cyclic peptides and depsipeptides, including microcystins and lyngbyatoxins, which have a structure similar to that found in marine invertebrates.
Cyclic lipopeptides derived from cyanobacteria have shown remarkable antifungal activity against a range of human and plant fungal pathogens, alongside their antibiotic properties. The amphipathic structure of these peptides consists of a polar peptide cycle and a hydrophobic fatty acid side chain. Given their potential as new antifungal agents, there is growing interest in exploring the novel biotechnological and therapeutic applications of these abundant and diverse cyclic peptides derived from cyanobacteria.
One example of these compounds is Majusculamide C (Figure 3), a potent fungicidal agent extracted from Lungbya majuscula. This microfilament-depolymerizing agent has been shown to be effective against diseases in domestic plants and agricultural crops [31].
Tolypothrix byssoidea (EAWAG 195), a cyanobacterium, has been found to produce two cyclic tridecapeptides known as tolybyssidins A and B (Figure 3). Peptides A and B exhibit antifungal activity against the C. albicans at concentrations of 32 µg/mL and 64 µg/mL, respectively, and contain the non-natural amino acid dehydrohomoalanine (Dhha), along with proteinogenic amino acids, some of which have D- or L-configuration in their structure [32].
The cyanobacterium Anabaena cylindrica strain Bio33 was obtained from a water sample collected near Rügen Island, Germany, in the Baltic Sea. This isolate was found to contain four antifungal lipopeptides known as Balticidins A-D, which were characterized and extracted (Figure 3). These peptides have been shown to exhibit antifungal activity with generating inhibition zones from 21 to 32 mm using agar diffusion method against various strains including Candida maltosa, Candida krusei, C. albicans, Microsporum gypseum, Microsporum canis, Mucor sp., and Aspergillus fumigatus [33].
Cyclic antifungal lipopeptides derived from cyanobacteria can be categorized into four main structural classes: hassallidins, puwainaphycins, laxaphycins, and anabaenolysins (Figure 3). Hassallidins, produced by various cyanobacteria, are cyclic glycosylated lipopeptides consisting of a fatty acid chain, a peptide ring comprising eight amino acids, an exocyclic amino acid, and 1-3 sugar moieties. Puwainaphycins are amphipathic cyclic lipopeptides characterized by a β-amino fatty acid and a nine-membered peptide ring. Laxaphycins form a diverse group of cyclic lipopeptides, typically containing a rare β-amino fatty acid with a short linear chain of eight or ten carbons. Anabaenolysins, another class of cyclic lipopeptides, feature an uncommon unsaturated β-amino fatty acid with a conjugated triene structure and a four-membered peptide ring [34].
Hájek et al. [35] discovered that the cyclic lipopeptides from cyanobacteria, known as puwainaphycin/minutissamide (PUW/MIN) family (Figure 4) and a number of semisynthetic analogs, exhibited antifungal activity against both Alternaria alternata (a plant pathogen) and Aspergillus fumigatus (a human pathogen) with MIC values of 37 μM and 0.6 μM, respectively, suggesting a correlation between their antifungal activities. Moreover, the researchers observed an enhancement in the antifungal properties of these peptides through the generation of semi-synthetic lipopeptides.
3.2. Cyclic Peptides Isolated From Other Bacteria
Kaneda and Kajimura [36] obtained four strains of bacteria isolated from the rhizosphere of garlic with basal root due to plant pathogenic Fusarium oxysporum. Among these strains, Bacillus subtilis FR-2 was discovered to produce three cyclic lipopeptides, namely bacillopeptins A, B, and C (Figure 5), each incorporating a long-chain beta-amino acid. Compounds of bacillopeptins A, B and C showed MIC value of >100 µg/ml, >100 µg/ml, and 50 µg/ml, respectively, against C. albicans IFO 1594. For Saccharomyces cerevisiae HUT 7099, MIC values were >100 µg/ml, >100 µg/ml, and 25 µg/ml, respectively, for compounds A, B and C, respectively. MIC values for the two strains Fusarium oxysporum HF 8801 (pathogenic to garlic) and F. oxysporum HF 8835 (nonpathogenic to garlic) were >100 µg/ml and >100 µg/ml, respectively, for compounds A and B, while compound C exhibited a MIC value of 25 µg/ml for the pathogenic strain and 12.5 µg/ml for the nonpathogenic strain. Compounds A and B demonstrated similar activity against Aspergillus niger HUT 2016 with MIC value >100 µg/ml, whereas compound C showed MIC of 6.25 µg/ml. Compounds A, B, and C showed MIC values of >100 µg/ml, >100 µg/ml and 12.5 µg/ml against both A. oryzae IFO 4214. The same MIC values were also observed against and Penicillium thomii, respectively.
Bacillus polymyxa KT-8, another strain, was identified as a producer of highly effective antibiotic antifungal cyclic hexadepsipeptides, namely fusaricidins A, B, C, and D (Figure 5). These compounds all contain 15-guanidino-3-hydroxypentadecanoic acid as a side chain. Fusaricidins exhibited higher antifungal activity compared to bacillopeptins. Antifungal activity of fusaricidins A, B and mixture of C and D against Saccharomyces cerevisiae HUT 7099 showed MIC values of >100 µg/ml, 12.5 µg/ml and >100 µg/ml. The lowest MIC value of 1.56 µg/ml was observed against Fusarium oxysporum HF 8801 (pathogenic to garlic) and F. oxysporum HF 8835 (nonpathogenic to garlic). Fusaricidins A, B, and the mixture of C and D exhibited identical MIC values of 3.12 µg/ml against Aspergillus niger HUT 2016, A. oryzae IFO 4214, and Penicillium thomii.
Romano et al. [37] reported the discovery of two new cyclic lipopeptides (designated as 1 and 2) (Figure 6) belonging to the surfactins family, isolated from Bacillus amyloliquefaciens strain BO5A. The lipopeptides are composed of a heptapeptide chain with the amino acid sequence Glu-Val-Leu-Val-Asp-Leu-Leu, and are N-acylated at their N-terminal end by (R)-3-hydroxy fatty acids with linear alkyl chains of 16:0 and 15:0 for compounds 1 and 2, respectively, through the formation of a 25-membered lactone ring between the 3-hydroxyl group of the fatty acid and the carboxylic group of the C-terminal amino acid. Compound 2 displayed inhibitory antifungal activity against Aspergillus niger, Fusarium oxysporum, Penicillium italicum, and Trichoderma harzianum at three tested concentrations of 10, 50 and 100 ppm. Compound 1 demonstrated inhibition of 59% and 36% against F. oxysporum and A. niger, respectively. When a mixture of compounds 1 and 2 was evalauted, there was not any significant change in their antifungal activity.
Ma et al. [38] reported the isolation of three lipopeptides from the fermentation broth of Bacillus mojavensis B0621A that exhibited dose-dependent antifungal activity against a broad range of phytopathogens. Among these lipopeptides, there is one called anteiso-C17 mojavensin A (Figure 6), which belongs to the iturinic lipopeptide family. Antifungal activity of Anteiso-C17 mojavensin A was detected when concentration was over 2 mg/ml for Valsa mali, Fusarium oxysporum f. sp. cucumerinum, and Fusarium verticillioides. The compound possesses a distinctive peptide backbone composed of L-Asn₁, D-Tyr₂, D-Asn₃, L-Gln₄, L-Pro₅, D-Asn₆, and L-Asn₇. Additionally, it features an anteiso-type saturated β-fatty acid side chain.
The other two compounds, tentatively identified as iso-C16 fengycin B and anteiso-C17 fengycin B (Figure 6), were also assessed for their antifungal activities. The compounds resulted in ihbition zones ranging from 5.17 ± 0.19 to 11.88±0.47 mm when tested at concentations of 2 mg/ml and 3 mg/ml. These inhibitory effects observed against various fungal strains, including Valsa mali, Fusarium oxysporum f. sp. cucumerinum, Fusarium oxysporum f. sp. vasinfectum, Fusarium oxysporum f. sp., vasinfectum, SF2, Fusarium solani SF 130, Botryosphaeria berengriana f. sp. piricola, Botrytis cicrea, Rhizoctonia solani J. G. Kühn, Fusarium solani, Rhizoctonia solani, Valsa ceratosperma, Fusarium oxysporum f. sp., Cucumis melo L., Fusarium graminearum, Bipolaris maydis, Colletotrichum orbiculare, Fusarium verticillioides, and Fusarium verticillioid.
Zhang et al. [39] identified two cyclic lipopeptides, maribasins A and B (Figure 6), derived from the marine microorganism Bacillus marinus B-9987. The microorganism was isolated from Suaeda salsa, a plant species found along the Bohai coastline in the People's Republic of China. The structures of both compounds were determined to be cyclo (D-Pro-L-Gln-L-Asn-L-Ser-D-Asn¹-D-Tyr-D-Asn²-D-β-aminoisopentadecanoic acid) and cyclo (D-Pro-L-Gln-L-Asn-L-Ser-D-Asn-D-Tyr-D-Asn-D-β-aminoanteisopentadecanoic acid).
Compounds A and B exhibited broad-spectrum activities, demonstrating MIC values ranging from 25 to 200 µg/ml against various fungi, including Alternaria solani, Fusarium oxysporum, Verticillium alboatrum, F. graminearum, Sclerotium sp., Penicillium sp., Rhizoctonia solani, and Colletotrichum sp.
Routhu et al. [40] discovered that biofilm exopolymeric substances (EPS) surrounding marine microbes possess antifungal activity against Candida albicans pathogenic strains (susceptible) and one azole resistant Candida strain. They characterized five cyclic peptides CLPs (Figure 7) isolated from the marine microbe Neobacillus drentensis. The five CLP isoforms were recognized as new peptides with variants in the amino acid sequence and fatty acid chain. All the tested Candida sp. was inhibited with least concentrations from CLPS and depending on variety of strain. The MIC was found to be 7.8 µg/ml against two strains of C. albicans. The inhibition of C. albicans’s growth was characterized through preventing biofilm formation and disrupting the branching of filamentous hyphae. The cyclic peptide also plays a role in blocking the G1-S transition and inducing apoptotic cell death. The stereochemistry of the amino acids was not reported.
Konno et al. [41] reported that cyclic octapeptides derived from burkholdines exhibit antifungal activity against Saccharomyces cerevisiae, Aspergillus oryzae, and Candida viswanathii. The most potent analogues of burkholdines showed MIC values ranging from 25 to 50 µg/ml. The antifungal activity of BK-1097 was notably affected by both the lipid side chain and the stereochemistry of each amino acid present (see Figure 8).
Troskie and colleagues [42] reported the production of cationic cyclodecapeptides by Bacillus aneurinolyticus, including Tyrocidine A (TrcA), Tyrocidine B (TrcB), Tyrocidine C (TrcC), Phenycidine A (PhcA), Tryptocidine C (TpcC), and Gramicidin S [43], respectively. The sequences of these peptides are as follows: cyclo-(VOLfPFfNQY), cyclo-(VOLfPWfNQY), cyclo-(VOLfPWwNQY), cyclo-(VOLfPFfNQF), cyclo-(VOLfPWwNQW), and cyclo-(VOLfPVOLfP), respectively. The tyrocidines were found to exhibit antifungal activity against C. albicans and effectively inhibit biofilm formation in vitro. The mixture of Tyrocidines (TrcA, TrcB, TrcC, TpcC, and PhcA) displayed potent antifungal activity against planktonic C. albicans with a MIC value of 6.25 µg/ml. Purified tyrocidines, TpcC, and the analogous peptide GS showed MIC values of 6.25 µM, while a higher MIC of 12.5 µM was detected with PhcA. The inhibitory effect of tyrocidines on mature C. albicans’s biofilm cells is attributed to their disruptive impact on the membrane integrity.
Troskie and colleagues [44] evaluated the antifungal activity of the cationic cyclodecapeptides produced by Bacillus aneurinolyticus against Aspergillus fumigatis ATCC 204305, Fusarium solani STEU 6188, Fusarium oxysporum ATCC 10913, Fusarium verticilliodes CKJ1730, Botrytis cinerea CKJ1731, Cylindrocarpon liriodendri STEU 6170, Penicillium glabrum CKJ1732, Talaromyces ramulosus CKJ1735, Talaromyces mineoluteus CKJ1736, Penicillium expansum CKJ1733, and Penicillium digitatum CKJ1734. The tyrocidine peptide complex (Trc mixture) and purified tyrocidines exhibited antifungal activity, with MIC values of below 13 mg/ml ( ̴10 µM). The antifungal mechanism involved the inhibition of germination and the process of hyphal hyper-branching. The inhibitory impact of tyrocidines on the membrane integrity was found to be responsible for their inhibitory effect.
Rautenbach and colleagues [45] conducted a study on tyrocidines and their analogs, which are cyclic decapeptides produced by Brevibacillus parabrevis. These peptides shared a conserved sequence of cyclo(D-Phe1-Pro2-X3-X4-Asn5-Gln6-X7-Val8-X9-Leu10), where X3, X4, X7, and X9 represent variable amino acid residues. The aromatic dipeptide unit can be either Trp3,4/Phe3,4., and Lys9/Orn9 serves as the cationic residue. At position 7, the peptides contain either Tyr (tyrocidines), Trp (tryptocidines), or Phe (phenicidines). he researchers observed that replacing the Tyr residue at position 7 with Phe or Trp enhanced the antifungal activity of the peptides. The most active peptides displayed IC50 (µg/ml) values of 1.1, 2.6, and 2.6 against Botrytis cinerea CKJ1731, Fusarium solani STEU 6188, and Aspergillus fumigatis ATCC 204305 , respectively.
Overall, cyclic peptides isolated from cyanobacteria and other bacteria have great potential as new antifungal agents and for other biotechnological and therapeutic applications. There is growing interest in exploring the novel properties and applications of these abundant and diverse cyclic peptides.
3.3. Cyclic Peptides Isolated From Fungi
In general, several studies have reported the isolation of cyclic peptides from various fungal strains and their potential as antifungal agents. These findings suggest that cyclic peptides from fungi hold great promise as potential sources of novel antifungal agents.
Using reverse genetics technology and the C. albicans fitness test (CaFT), the cyclic desipeptide phaeofungin (Figure 9) was isolated from Phaeoshaeria sp. This compound consists of a 25-membered cyclic depsipeptide made up of seven amino acids and a β,γ-dihydroxy-γ-methylhexadecanoic acid. In a similar approach, the CaFT profile of the phaeofungin-containing extract was found to overlap with that of phomafungin, a structurally different cyclic lipodepsipeptide also produced by the same organism [46].
Functionally, phaeofungin and phomafungin are distinct from each other. Potentiation of Phomafungin's antifungal activity was observed when combined with cyclosporin A, an inhibitor of the calcineurin pathway. In contrast, Phaeofungin exhibited synergistic effects when combined with aureobasidin A 2, a sphingolipid biosynthesis inhibitor, and to some extent with caspofungin, a glucan synthase inhibitor. Interestingly, Phaeofungin induced ATP release in wild-type strains of C. albicans, while Phomafungin did not exhibit this effect. It displayed antifungal activity against Aspergillus fumigatus (MIC 8-16 µg/ml), C. albicans (MIC 16-32 µg/ml), and Trichophyton mentagrophytes (MIC 4 µg/ml). Notably, the inactivity of the linear peptide reflected the importance of the macrocyclic depsipeptide ring for target engagement and activity as antifungal compound [46].
Colisporifungin, a cyclic desilipopeptide with structural similarities to the linear peptides aselacins and cavinafungins A and B (as depicted in Figure 9), was isolated from the liquid culture broths of the Colispora cavincola fungus. In experimental studies, colisporifungin exhibited a remarkable enhancement of the antifungal growth activity of caspofungin against A. fumigatus (MIC 8 µg/ml), and to a lesser extent, C. albicans (MIC 0.5-4 µg/ml) [47].
Liang et al. [48] investigated the structure-bioactivity relationship of cyclic peptides derived from the deep-sea fungus strain Simplicillium obclavatum EIODSF 020. The study primarily examined the influence of the lactone linkage and the substituent group of the fatty acid chain fragment on the bioactivity of these cyclic peptides. These cyclic peptides included simplicilliumtides J–M, along with verlamelins A and B analogs. Out of these compounds, three peptides demonstrated antifungal activity against Aspergillus versicolor (IC50 = 14 µM) and Curvularia australiensis (IC50 = 16.7 µM).
In another study, the deep-sea-derived fungal strain Simplicillium obclavatum EIODSF 020 yielded two novel cyclopeptides, namely simplicilliumtides N and O (1 and 2), along with three previously identified analogs, verlamelins A, B, and simplicilliumtide J (3-5) (as depicted in Figure 9). These peptides exhibited notable antifungal activity against two phytopathogenic fungal strains, Colletotricum asianum and Alternaria solani (MIC= 0.195-6.25 µg/disc) [49].
A variety of peptides were isolated from the marine gorgonian-associated fungus Aspergillus sp. SCSIO 41501 (Trichocomaceae), including three cyclic lipopeptides (Maribasins C-E), four linear peptides (Aspergillipeptides H-K), and three previously identified cyclic lipopeptides (Maribasins A-B and Marihysin A). Notably, Maribasins C-E and Maribasins A-B exhibited remarkable antifungal activity against five phytopathogenic fungal strains: F. oxysporum, C. australiensis, P. oryzae, C. gloeosporioiles, and A. solani (MIC 3.12-50 µg/disc). The structure-bioactivity relationship revealed that the antifungal activity of these cyclic lipopeptides could be significantly affected by the β-amino fatty acid chain [50].
Clavariopsis aquatica, an aquatic hyphomycete, was found to contain seven cyclic depsipeptides - clavariopsins C-I, and two previously identified compounds, clavariopsins A and B (Figure 9), all of which possess antifungal properties against six plant pathogenic fungi: Magnaporthe oryzae, Botrytis cinerea, Fusarium oxysporum, Colletotrichum orbiculare, Aspergillus niger, and Alternaria alternata. These peptides are composed of nine amino acids and one α-hydroxy acid, and share the same structure. The antifungal activity showed values of minimum inhibitory dose ranged from 0.01 to 10 µg/disc. [51].
It was reported that Aureobasidium pullulans R106 produces aureobasidins, which possess antibiotic and antifungal properties. A member of this group is Aureobasidin A, which is a cyclic depsipeptide composed of eight alpha-amino acid units and one hydroxy acid unit. Studies have shown that in addition to having a broad spectrum of activity, Aureobasidin A is more effective against murine candidiasis than the aculeacin/echinocandins family. The antifungal activity against C. albicans , C. kefyr, C. glabrata, and Cryptococcus neoformans showed MIC values ranging from 0.05 to 25 µg/ml [52].
3.4. Cyclic Peptides Isolated from Plants
Antifungal activity was observed in the crude ethanolic extracts derived from cultured Anabaena laxa, a blue-green alga, which was attributed to the presence of Laxaphycins. Antifungal activity against Aspergillus oryzae, Candida albicans, Penicillium notatum, Saccharomyces cerevisiae, and Trichophyton mentagrophytes indicated with the inhibition zone ranging from 8 to 26 mm in agar diffusion. These cyclic peptides displayed an atypical biological synergism during the screening process for antifungal activity, as reported by Frankmölle et al. in [53].
Tunicyclins B-D (Figure 10), cyclic peptides extracted from the root of Psammosilene tunicoides, showed antifungal activity against various strains. Among them, tunicyclin D demonstrated a broad spectrum of antifungal activity against various strains. It showed a MIC80 value of 4 µg/ml against C.albicans (SC5314) and a slightly observed against Candida tropicalis. Tunicyclin D displayed the same MIC80 value of 1 µg/ml against both Candida parapsilosis and Cryptococcus neoformans (BLS108), as reported by Tian et al. [54].
In addition to cyclic peptides, conformationally constrained peptides have been shown to exhibit antifungal activity. The identification of small, basic, cysteine-rich peptides known as plant defensins has proven significant in the search for antifungal agents. These conformationally constrained peptides have demonstrated potent antifungal activity against various fungal and yeast species, at micromolar concentrations. Plant defensins exhibit selective activity and are non-phytotoxic, owing to the structural dissimilarities between the membrane components of plant and fungal cells. The antifungal activity of defensins in plants is a result of their interaction with specific lipid components in the plasma membrane of fungi, as reported by Thevissen et al. [55].
Orbitides and cyclotides are cyclic peptides that are derived from plants. They possess distinct structural arrangement and sequence. Cyclotides, in contrast to orbitides, do not contain a disulfide bond and have a higher proportion of hydrophobic residues. Integerrimides A and B are orbitides oisolated from the latex of Jatropha integerrima Jacq. In the antifungal assay, both compounds A and B exhibited no activity at concentrations of 64.1 µM and 65.3 µM, respectively [56,57].
Cyclotides, in contrast, possess a peptide backbone that forms a circular structure, typically composed of around 30 residues. They are characterized by the presence of six conserved cysteine residues, which form three disulfide bonds, creating a unique cyclic cystine knot motif [58]. Stromstedt et al. [59] evaluated the fungicidal activity of cycloviolacin O2, cycloviolacin O3, and cycloviolacin O19 against the fungi C. albicans. These cyclotides demonstrated a fungicidal activity of >99% with a Minimum Fungicidal Concentration (MFC) of 10 μM (MFC99−10 μM).
Thus, cyclic peptides and conformationally constrained peptides, such as defensins, isolated from plants have shown potent antifungal activity against various fungal and yeast species, at micromolar concentrations, owing to their interaction with specific lipid components in the plasma membrane of fungi. Furthermore, plant defensins exhibit selective activity and are non-phytotoxic, making them a promising avenue for antifungal drug development.
3.5. Synthetic Cyclic Peptides With Antifungal Activity
Grimaldi and colleagues [60] synthesized and characterized six peptides, namely AMT1, AMT2, AMT3, cyclo-AMT1, cyclo-AMT2, and cyclo-AMT3 (Figure 11), to assess their antifungal properties. The antifungal activity of these compounds was assessed against Candida species, including C. albicans, C. glabrata and C. tropicalis. The linear peptides exhibited MIC values ranging from 64 to >512 µg/mL, while the cyclic peptides showed MIC values ranging from 32 to 512 µg/mL Generally, the cyclic peptides displayed greater activity compared to their linear counterparts. Moreover, all the tested compounds displayed potent antifungal activity against C. neoformans ATCC 52817 and C. neoformans H99, with MIC values ranging from 4 to 16 µg/ml.
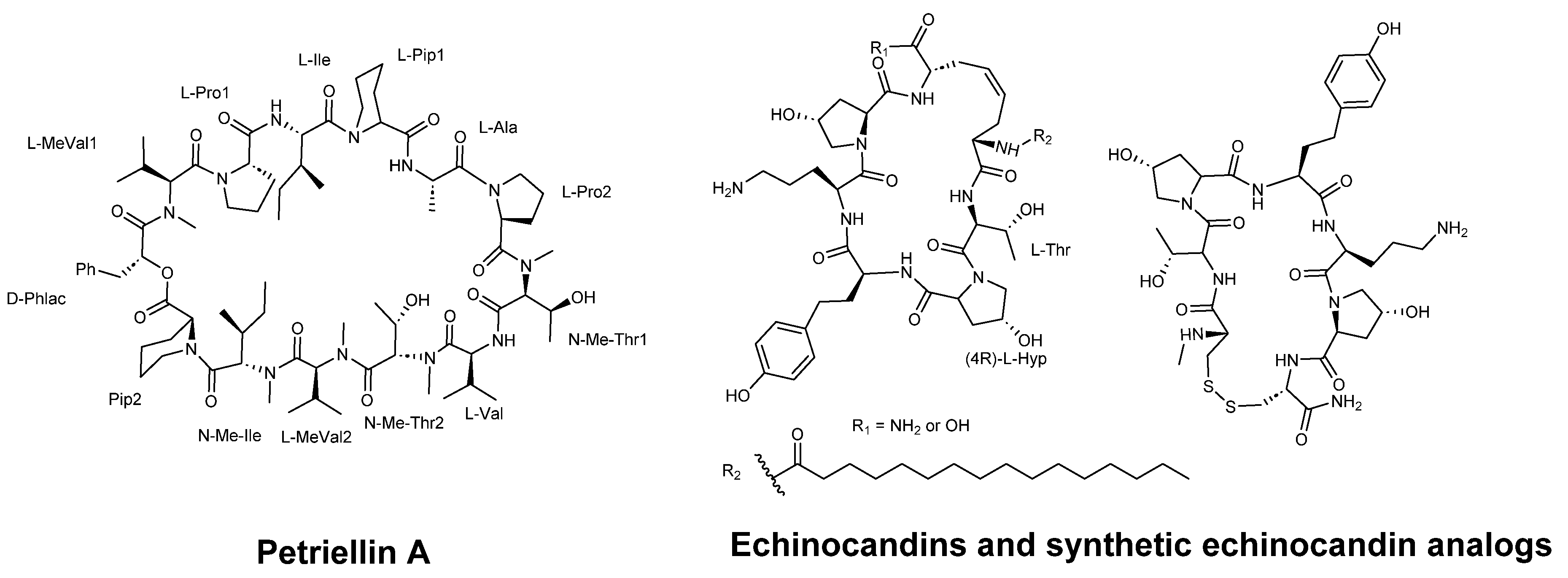
Sleebs et al. [41] described the synthesis of petriellin A (Figure 12), which was originally extracted from the organic extracts of Petriella sordida (UAMH 7493), an antagonistic coprophilous fungus. This cyclic depsipeptide is composed of one R-D-hydroxycarboxylic acid and 12 L-amino acids, five of which are N-methylated. The study demonstrated that petriellin A displayed strong antifungal activity against Ascobolus furfuraceous (NRRL 6460) and Sordaria fimicola (NRRL 6459).
Kurokawa and Ohfune [61] synthesized oligopeptides echinocandins C and D with fungicidal activity. Echinocandins as macrocyclic peptides inhibit the β-(1,3)-D-glucan synthase complex, a mechanism that provides selective activity as this complex is absent in mammalian cells. The antifungal activity of these peptides (see Figure 12) against C. albicans CBS 9975 evaluated, showing MIC values ranging from 0.025 to ˃100 µg/ml [62].
The glycosylic cyclic peptide antifungal drug caspofungin derivatives prepared from conjugation with β-d-glucopyranose, β-d-gal-actopyranose, β-d-xylopyranose, β-maltose, β-l-rhamnopyranose, and β-lactose units showed higher antifungal activity against in case of caspofungin–monosaccharide conjugates not the disaccharide ones. The MIC values against C. albicans Y0109, C. albicans SC5314, C. parapsilosis 22019, C. krusei 537,Cryptococcus neoformans 32609, Microsporum gypseum Cmccfmza, and Trichophyton rubrum Cmccftla ranged from 1 to 32 µm/L [63].
Antifungal cyclic and helical-stabilized analogues peptide Cm-p5 was synthesized and reported to be active against C. albicans and C. parapsilosis. Also, other derivatives showed antifungal activity against C. auris biofilms inhibiting 71–97% using concentrations between 10–21 µg/ml [64].
The natural phenylalanine-rich cycloheptapeptide segetalin C synthesis was conducted through coupling and cyclization of peptide units L-Phe-L-ala-L-phe-L-pro-OMe and Boc-gly-L-leu-L-his-OH. The synthesized cyclopeptide showed good antifungal activity against C. albicans [65].
3.6. Combination Antifungal Activity
Antifungal agents exhibit synergistic effects by acting through various mechanisms to develop their destructive effect against pathogenic fungi. Combination therapy between antifungal drugs has become necessary due to the emergence of antifungal resistance, enhancing their effectiveness against pathogenic fungi that are resistant to single drugs. In their study, Pesee et al. [66] conducted a study investigating the combination of the lipopeptide antifungal drug caspofungin (CAS) with fluconazole. In vitro experiments revealed a notable influence on the biofilm biomass, cultivable viability, and microstructure of mixed biofilms composed of C. albicans and Candida glabrata.
To develop effective and specific antifungal compounds, Rodríguez López et al. [67] conducted a study in which they designed amphiphilic, cationic, 14-helical β-peptides. These β-peptides were created to emulate the structural features of natural antimicrobial α-peptides and act as antifungal agents against planktonic C. albicans cells. The researchers combined these β-peptides with isoamyl alcohol, which inhibits hyphal formation, resulting in reduced minimum biofilm prevention concentrations (MBPCs) of the β-peptides. These findings suggest that combining peptides from different sources may be a promising approach for combating antifungal resistance in pathogenic fungi.
4. Future Perspectives
The rising prevalence of fungal infections, particularly among immunocompromised individuals, has created a need for effective and safe antifungal treatments [68]. In this context, the utilization of cyclic peptides as antifungal agents holds significant promise, and there is a growing interest in exploring their therapeutic applications. Cyclic peptides offer several advantages over conventional antifungal agents, including high potency, low toxicity, and target specificity as shown with caspofungin, micafungin and anidulafungin [19,20,23]. Recently FDA approved Rezafungin [69] (a novel echinocandin antifungal agent against invasive candidiasis and candidemia. Given the success of these drugs, further exploration of other cyclic antifungal peptides is warranted.
Cyclic peptides can be obtained from diverse origins, including plants, fungi, and bacteria, and can also be synthesized and they exhibit diverse antifungal activities. Cyanobacteria, for example, have been found to produce a wide range of biologically active cyclic peptides that have potential applications in biotechnology and therapeutics [18,19,20,21,22]. The outlook for the isolation of cyclic peptides from fungi is also encouraging. Fungal cyclic peptides have high potential as sources of novel antifungal agents [30,31,32,33,34], and ongoing research is expected to unveil new compounds with different mechanisms of action. Additionally, marine fungi have been identified as rich sources of cyclic peptides with antifungal properties.
Future research in this field could focus on identifying new strains of cyanobacteria and fungi that produce unique cyclic peptides with novel structures and biological activities. With advancements in biotechnology and synthetic methods, it is highly likely that more cyclic peptides with potent antifungal activity will be discovered and developed. This could be accomplished by using genomic and transcriptomic techniques to identify biosynthetic gene clusters responsible for cyclic peptide production, as well as employing high-throughput assays to screen for new bioactive compounds [70,71]. Moreover, petidomics is an emerging discipline that integrates genomics, proteomics, analytical chemistry techniques such as liquid chromatography and mass spectrometry, and computational biology to qualitatively and quantitatively analyze biologically significant cyclic peptides [72].
Furthermore, the utilization of machine learning algorithms in artificial intelligence (AI) [73,74] enables the extraction of valuable insights and patterns from existing data on the characteristics and functions of acknowledged cyclic peptides. This ability streamlines the search for novel cyclic peptides with specific antifungal properties, derived from both natural and synthetic sources. Moreover, AI aids in accurately predicting the structural properties, including stability and folding patterns, of cyclic peptides. This guidance assists researchers in identifying the most promising candidates for further examination and analysis.
Moreover, the enhancement of existing cyclic peptides derived from cyanobacteria, fungi, and plants via semi-synthetic approaches holds promise in establishing the structure-activity relationship and the creation of more potent and selective analogs with improved pharmacological characteristics with appropriate pharmacokinetics properties and stability [75,76]. Studying the relationship between structure and bioactivity of synthesized compounds can provide insights into how different structural features affect the antifungal activity of these compounds. Further investigation of these semi-synthesized compounds as antifungal agents could be pursued through in vivo studies, while advanced biochemical and biophysical methodologies could be employed to unravel their mechanisms of action. Furthermore, these compounds could serve as scaffolds for the development of new drug candidates optimizing them through medicinal chemistry techniques. Overall, the optimization of cyclic peptides extracted from different sources presents a tremendous potential for the identification of novel therapeutic agents and the advancement of biotechnological applications in the coming years.
In the face of antifungal-resistant pathogens [77], the importance of developing combination antifungal therapy has escalated. With the emergence of new strains of resistant fungi, it is crucial to persist in investigating and developing fresh drug combinations that effectively combat these pathogens, thereby reducing the likelihood of resistance development [78]. This approach holds particular significance in the treatment of chronic or recurrent fungal infections, where the emergence of resistance is a notable concern [79]. One avenue for advancing combination antifungal therapy involves merging natural products and synthetic drugs [80]. For instance, researchers can explore the potential of synergizing plant extracts or essential oils with synthetic antifungal agents to create more potent treatment options. Another area of research focuses on combination therapies that target multiple mechanisms of action. By combining antifungal agents with distinct modes of action, researchers can heighten the probability of pathogen eradication while minimizing the risk of resistance development.
The combination of peptides derived from natural, microbial, and synthetic pathways with other drugs displays promising potential in the development of exceptionally effective antifungal agents [81,82,83]. In the future, personalized combination antifungal therapy could be explored. By analyzing specific fungal strains and patient characteristics, researchers may develop tailored combination therapies that are more effective and have fewer side effects.
In summary, cyclic peptides exhibit considerable potential as effective and safe antifungal agents, characterized by their high potency, low toxicity, and specificity. The clinical approval of Caspofungin, Micafungin, and Anidulafungin emphasizes the promising prospects of cyclic peptides in antifungal therapy. However, further investigation is needed to fully exploit the potential of cyclic peptides derived from diverse sources, including cyanobacteria, bacteria, fungi, and plants. Ongoing research and development in this field hold the promise of discovering new cyclic peptides from various origins, significantly broadening the range of available antifungal treatments. Computational methods can aid in the identification and optimization of cyclic peptides with improved antifungal properties. These novel cyclic peptides offer innovative therapeutic options for combating fungal infections in both immunocompromised and healthy individuals, thereby addressing the challenge posed by the widespread prevalence of antifungal-resistant pathogenic fungi. Through continued exploration and advancement, cyclic peptides have the potential to revolutionize the treatment of fungal infections, whether used as standalone therapies or in combination with other drugs, thus contributing to the advancement of global health.
Author Contributions
Conceptualization, N.M.H. and K.P.; R.K.T.; writing—original draft preparation, N.M.H. and K.P., review and editing, N.M.H. and K.P.
Funding
The authors acknowledge financial support from Chapman University School of Pharmacy.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Martins, N.; Ferreira, I.C.; Barros, L.; Silva, S.; Henriques, M. Candidiasis: predisposing factors, prevention, diagnosis and alternative treatment. Mycopathologia 2014, 177, 223–240. [Google Scholar] [CrossRef]
- Cadena, J.; Thompson, G.R., 3rd; Patterson, T.F. Aspergillosis: Epidemiology, Diagnosis, and Treatment. Infect Dis Clin North Am 2021, 35, 415–434. [Google Scholar] [CrossRef] [PubMed]
- Rautenbach, M.; Troskie, A.M.; Vosloo, J.A. Antifungal peptides: To be or not to be membrane active. Biochimie 2016, 130, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Zähner, H.; Maas, W.K. Mode of Action. In Biology of Antibiotics; Springer US: New York, NY, 1972; pp. 62–96. [Google Scholar]
- Carrillo-Muñoz, A.J.; Giusiano, G.; Ezkurra, P.A.; Quindós, G. Antifungal agents: mode of action in yeast cells. Rev Esp Quimioter 2006, 19, 130–139. [Google Scholar]
- Zhang, D.; Lu, Y.; Chen, H.; Wu, C.; Zhang, H.; Chen, L.; Chen, X. Antifungal peptides produced by actinomycetes and their biological activities against plant diseases. J Antibiot (Tokyo) 2020, 73, 265–282. [Google Scholar] [CrossRef] [PubMed]
- Ramadhani, D.; Maharani, R.; Gazzali, A.M.; Muchtaridi, M. Cyclic Peptides for the Treatment of Cancers: A Review. Molecules 2022, 27, 4428. [Google Scholar] [CrossRef] [PubMed]
- Evans, B.J.; King, A.T.; Katsifis, A.; Matesic, L.; Jamie, J.F. Methods to Enhance the Metabolic Stability of Peptide-Based PET Radiopharmaceuticals. Molecules 2020, 25, 2314. [Google Scholar] [CrossRef]
- Vinogradov, A.A.; Yin, Y.; Suga, H. Macrocyclic Peptides as Drug Candidates: Recent Progress and Remaining Challenges. Journal of the American Chemical Society 2019, 141, 4167–4181. [Google Scholar] [CrossRef]
- Landy, M.; Warren, G.H.; RosenmanM, S.B.; Colio, L.G. Bacillomycin: An Antibiotic from Bacillus subtilis Active against Pathogenic Fungi∗. Proceedings of the Society for Experimental Biology and Medicine 1948, 67, 539–541. [Google Scholar] [CrossRef]
- Banerjee, N.; Bose, S.K. MODE OF ACTION OF MYCOBACILLIN, A NEW ANTIFUNGAL ANTIBIOTIC. J Bacteriol 1963, 86, 387–391. [Google Scholar] [CrossRef]
- Rose, A.H.; Wilkinson, J.F. Advances in Microbial Physiology; Elsevier Science: 1970.
- Segre, A.; Bachmann, R.C.; Ballio, A.; Bossa, F.; Grgurina, I.; Iacobellis, N.S.; Marino, G.; Pucci, P.; Simmaco, M.; Takemoto, J.Y. The structure of syringomycins A1, E and G. FEBS Lett 1989, 255, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, J.Y.; Zhang, L.; Taguchi, N.; Tachikawa, T.; Miyakawa, T. Mechanism of action of the phytotoxin syringomycin: a resistant mutant of Saccharomyces cerevisiae reveals an involvement of Ca2+ transport. Microbiology 1991, 137, 653–659. [Google Scholar] [CrossRef]
- Moon, S.S.; Chen, J.L.; Moore, R.E.; Patterson, G.M.L. Calophycin, a fungicidal cyclic decapeptide from the terrestrial blue-green alga Calothrix fusca. The Journal of Organic Chemistry 1992, 57, 1097–1103. [Google Scholar] [CrossRef]
- Mizuno, K.; Yagi, A.; Satoi, S.; Takada, M.; Hayashi, M. Studies on aculeacin. I. Isolation and characterization of aculeacin A. J Antibiot (Tokyo) 1977, 30, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Gordee, R.S.; Zeckner, D.J.; Ellis, L.F.; Thakkar, A.L.; Howard, L.C. In vitro and in vivo anti-Candida activity and toxicology of LY121019. J Antibiot (Tokyo) 1984, 37, 1054–1065. [Google Scholar] [CrossRef] [PubMed]
- Mizoguchi, J.; Saito, T.; Mizuno, K.; Hayano, K. On the mode of action of a new antifungal antibiotic, aculeacin A: inhibition of cell wall synthesis in Saccharomyces cerevisiae. J Antibiot (Tokyo) 1977, 30, 308–313. [Google Scholar] [CrossRef]
- Balkovec, J.M.; Hughes, D.L.; Masurekar, P.S.; Sable, C.A.; Schwartz, R.E.; Singh, S.B. Discovery and development of first in class antifungal caspofungin (CANCIDAS®)—A case study. Natural Product Reports 2014, 31, 15–34. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, S. Cyclic peptide drugs approved in the last two decades (2001-2021). RSC Chem Biol 2022, 3, 18–31. [Google Scholar] [CrossRef]
- Abdalla, M.A.; McGaw, L.J. Natural Cyclic Peptides as an Attractive Modality for Therapeutics: A Mini Review. Molecules 2018, 23. [Google Scholar] [CrossRef]
- Higashiyama, Y.; Kohno, S. Micafungin: a therapeutic review. Expert Rev Anti Infect Ther 2004, 2, 345–355. [Google Scholar] [CrossRef]
- Scott, L.J. Micafungin: a review of its use in the prophylaxis and treatment of invasive Candida infections. Drugs 2012, 72, 2141–2165. [Google Scholar] [CrossRef] [PubMed]
- Wasmann, R.E.; Muilwijk, E.W.; Burger, D.M.; Verweij, P.E.; Knibbe, C.A.; Brüggemann, R.J. Clinical Pharmacokinetics and Pharmacodynamics of Micafungin. Clin Pharmacokinet 2018, 57, 267–286. [Google Scholar] [CrossRef] [PubMed]
- Kofla, G.; Ruhnke, M. Pharmacology and metabolism of anidulafungin, caspofungin and micafungin in the treatment of invasive candidosis: review of the literature. Eur J Med Res 2011, 16, 159–166. [Google Scholar] [CrossRef]
- Gang, D.; Kim, D.W.; Park, H.S. Cyclic Peptides: Promising Scaffolds for Biopharmaceuticals. Genes (Basel) 2018, 9. [Google Scholar] [CrossRef]
- Hashimoto, S. Micafungin: a sulfated echinocandin. The Journal of Antibiotics 2009, 62, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Shao, L.; Li, J.; Liu, A.; Chang, Q.; Lin, H.; Chen, D. Efficient bioconversion of echinocandin B to its nucleus by overexpression of deacylase genes in different host strains. Appl Environ Microbiol 2013, 79, 1126–1133. [Google Scholar] [CrossRef]
- Davis, S.L.; Vazquez, J.A. Anidulafungin: an evidence-based review of its use in invasive fungal infections. Core Evid 2008, 2, 241–249. [Google Scholar]
- Alarcón-Manoja, E.; Cardozo-Espinola, C.; Puerta-Alcalde, P.; García-Vidal, C. Comments on practice guidelines for the diagnosis and management of aspergillosis made by the IDSA in 2016. Rev Esp Quimioter 2017, 30 Suppl 1, 26–29. [Google Scholar]
- Moore, R.E. Cyclic peptides and depsipeptides from cyanobacteria: a review. J Ind Microbiol 1996, 16, 134–143. [Google Scholar] [CrossRef]
- Jaki, B.; Zerbe, O.; Heilmann, J.; Sticher, O. Two novel cyclic peptides with antifungal activity from the cyanobacterium Tolypothrix byssoidea (EAWAG 195). J Nat Prod 2001, 64, 154–158. [Google Scholar] [CrossRef]
- Bui, T.H.; Wray, V.; Nimtz, M.; Fossen, T.; Preisitsch, M.; Schröder, G.; Wende, K.; Heiden, S.E.; Mundt, S. Balticidins A-D, antifungal hassallidin-like lipopeptides from the Baltic Sea cyanobacterium Anabaena cylindrica Bio33. J Nat Prod 2014, 77, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Fewer, D.P.; Jokela, J.; Heinilä, L.; Aesoy, R.; Sivonen, K.; Galica, T.; Hrouzek, P.; Herfindal, L. Chemical diversity and cellular effects of antifungal cyclic lipopeptides from cyanobacteria. Physiol Plant 2021, 173, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Hájek, J.; Bieringer, S.; Voráčová, K.; Macho, M.; Saurav, K.; Delawská, K.; Divoká, P.; Fišer, R.; Mikušová, G.; Cheel, J.; et al. Semi-synthetic puwainaphycin/minutissamide cyclic lipopeptides with improved antifungal activity and limited cytotoxicity. RSC Adv 2021, 11, 30873–30886. [Google Scholar] [CrossRef]
- Kaneda, M.; Kajimura, Y. [New antifungal antibiotics, bacillopeptins and fusaricidins]. Yakugaku Zasshi 2002, 122, 651–671. [Google Scholar] [CrossRef] [PubMed]
- Romano, A.; Vitullo, D.; Senatore, M.; Lima, G.; Lanzotti, V. Antifungal cyclic lipopeptides from Bacillus amyloliquefaciens strain BO5A. J Nat Prod 2013, 76, 2019–2025. [Google Scholar] [CrossRef]
- Ma, Z.; Wang, N.; Hu, J.; Wang, S. Isolation and characterization of a new iturinic lipopeptide, mojavensin A produced by a marine-derived bacterium Bacillus mojavensis B0621A. J Antibiot (Tokyo) 2012, 65, 317–322. [Google Scholar] [CrossRef]
- Zhang, D.J.; Liu, R.F.; Li, Y.G.; Tao, L.M.; Tian, L. Two new antifungal cyclic lipopeptides from Bacillus marinus B-9987. Chem Pharm Bull (Tokyo) 2010, 58, 1630–1634. [Google Scholar] [CrossRef]
- Routhu, S.R.; Ragi, N.C.; Yedla, P.; Shaik, A.B.; Venkataraman, G.; Cheemalamarri, C.; Chityala, G.K.; Amanchy, R.; Sripadi, P.; Kamal, A. Identification, characterization and evaluation of novel antifungal cyclic peptides from Neobacillus drentensis. Bioorg Chem 2021, 115, 105180. [Google Scholar] [CrossRef]
- Konno, H.; Otsuki, Y.; Matsuzaki, K.; Nosaka, K. Synthesis and antifungal activities of cyclic octa-lipopeptide burkholdine analogues. Bioorg Med Chem Lett 2013, 23, 4244–4247. [Google Scholar] [CrossRef]
- Troskie, A.M.; Rautenbach, M.; Delattin, N.; Vosloo, J.A.; Dathe, M.; Cammue, B.P.; Thevissen, K. Synergistic activity of the tyrocidines, antimicrobial cyclodecapeptides from Bacillus aneurinolyticus, with amphotericin B and caspofungin against Candida albicans biofilms. Antimicrob Agents Chemother 2014, 58, 3697–3707. [Google Scholar] [CrossRef]
- van Eijk, M.; Boerefijn, S.; Cen, L.; Rosa, M.; Morren, M.J.H.; van der Ent, C.K.; Kraak, B.; Dijksterhuis, J.; Valdes, I.D.; Haagsman, H.P.; et al. Cathelicidin-inspired antimicrobial peptides as novel antifungal compounds. Med Mycol 2020, 58, 1073–1084. [Google Scholar] [CrossRef] [PubMed]
- Troskie, A.M.; de Beer, A.; Vosloo, J.A.; Jacobs, K.; Rautenbach, M. Inhibition of agronomically relevant fungal phytopathogens by tyrocidines, cyclic antimicrobial peptides isolated from Bacillus aneurinolyticus. Microbiology (Reading) 2014, 160, 2089–2101. [Google Scholar] [CrossRef]
- Rautenbach, M.; Troskie, A.M.; Vosloo, J.A.; Dathe, M.E. Antifungal membranolytic activity of the tyrocidines against filamentous plant fungi. Biochimie 2016, 130, 122–131. [Google Scholar] [CrossRef]
- Singh, S.B.; Ondeyka, J.; Harris, G.; Herath, K.; Zink, D.; Vicente, F.; Bills, G.; Collado, J.; Platas, G.; González del Val, A.; et al. Isolation, structure, and biological activity of Phaeofungin, a cyclic lipodepsipeptide from a Phaeosphaeria sp. Using the Genome-Wide Candida albicans Fitness Test. J Nat Prod 2013, 76, 334–345. [Google Scholar] [CrossRef]
- Ortíz-López, F.J.; Monteiro, M.C.; González-Menéndez, V.; Tormo, J.R.; Genilloud, O.; Bills, G.F.; Vicente, F.; Zhang, C.; Roemer, T.; Singh, S.B.; et al. Cyclic colisporifungin and linear cavinafungins, antifungal lipopeptides isolated from Colispora cavincola. J Nat Prod 2015, 78, 468–475. [Google Scholar] [CrossRef]
- Liang, X.; Nong, X.H.; Huang, Z.H.; Qi, S.H. Antifungal and Antiviral Cyclic Peptides from the Deep-Sea-Derived Fungus Simplicillium obclavatum EIODSF 020. J Agric Food Chem 2017, 65, 5114–5121. [Google Scholar] [CrossRef]
- Firmansyah, K. Bioassay-guided isolation of antifungal cyclopeptides from the deep-sea-derived fungus Simplicillium obclavatum EIODSF 020. Phytochemistry letters 2022, 48, 68–71. [Google Scholar] [CrossRef]
- Yao, F.H.; Liang, X.; Cheng, X.; Ling, J.; Dong, J.D.; Qi, S.H. Antifungal peptides from the marine gorgonian-associated fungus Aspergillus sp. SCSIO41501. Phytochemistry 2021, 192, 112967. [Google Scholar] [CrossRef]
- Soe, T.W.; Han, C.; Fudou, R.; Kaida, K.; Sawaki, Y.; Tomura, T.; Ojika, M. Clavariopsins C-I, Antifungal Cyclic Depsipeptides from the Aquatic Hyphomycete Clavariopsis aquatica. J Nat Prod 2019, 82, 1971–1978. [Google Scholar] [CrossRef] [PubMed]
- Ikai, K.; Takesako, K.; Shiomi, K.; Moriguchi, M.; Umeda, Y.; Yamamoto, J.; Kato, I.; Naganawa, H. Structure of aureobasidin A. J Antibiot (Tokyo) 1991, 44, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Frankmölle, W.P.; Larsen, L.K.; Caplan, F.R.; Patterson, G.M.; Knübel, G.; Levine, I.A.; Moore, R.E. Antifungal cyclic peptides from the terrestrial blue-green alga Anabaena laxa. I. Isolation and biological properties. J Antibiot (Tokyo) 1992, 45, 1451–1457. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Shen, Y.; Yang, X.; Liang, S.; Shan, L.; Li, H.; Liu, R.; Zhang, W. Antifungal cyclic peptides from Psammosilene tunicoides. J Nat Prod 2010, 73, 1987–1992. [Google Scholar] [CrossRef] [PubMed]
- Thevissen, K.; Ferket, K.K.; François, I.E.; Cammue, B.P. Interactions of antifungal plant defensins with fungal membrane components. Peptides 2003, 24, 1705–1712. [Google Scholar] [CrossRef] [PubMed]
- Mongkolvisut, W.; Sutthivaiyakit, S.; Leutbecher, H.; Mika, S.; Klaiber, I.; Möller, W.; Rösner, H.; Beifuss, U.; Conrad, J. Integerrimides A and B, cyclic heptapeptides from the latex of Jatropha integerrima. J Nat Prod 2006, 69, 1435–1441. [Google Scholar] [CrossRef] [PubMed]
- Ramalho, S.D.; Pinto, M.E.F.; Ferreira, D.; Bolzani, V.S. Biologically Active Orbitides from the Euphorbiaceae Family. Planta Med 2018, 84, 558–567. [Google Scholar] [CrossRef]
- Grover, T.; Mishra, R.; Bushra; Gulati, P.; Mohanty, A. An insight into biological activities of native cyclotides for potential applications in agriculture and pharmaceutics. Peptides 2021, 135, 170430. [Google Scholar] [CrossRef]
- Strömstedt, A.A.; Park, S.; Burman, R.; Göransson, U. Bactericidal activity of cyclotides where phosphatidylethanolamine-lipid selectivity determines antimicrobial spectra. Biochim Biophys Acta Biomembr 2017, 1859, 1986–2000. [Google Scholar] [CrossRef]
- Grimaldi, M.; De Rosa, M.; Di Marino, S.; Scrima, M.; Posteraro, B.; Sanguinetti, M.; Fadda, G.; Soriente, A.; D'Ursi, A.M. Synthesis of new antifungal peptides selective against Cryptococcus neoformans. Bioorg Med Chem 2010, 18, 7985–7990. [Google Scholar] [CrossRef]
- Kurokawa, N.; Ohfune, Y. Synthetic studies on antifungal cyclic peptides, echinocandins. Stereoselective total synthesis of echinocandin D via a novel peptide coupling. Tetrahedron 1993, 49, 6195–6222. [Google Scholar] [CrossRef]
- Mulder, M.P.; Kruijtzer, J.A.; Breukink, E.J.; Kemmink, J.; Pieters, R.J.; Liskamp, R.M. Synthesis and evaluation of novel macrocyclic antifungal peptides. Bioorg Med Chem 2011, 19, 6505–6517. [Google Scholar] [CrossRef]
- Guo, J.; Hu, H.; Zhao, Q.; Wang, T.; Zou, Y.; Yu, S.; Wu, Q.; Guo, Z. Synthesis and antifungal activities of glycosylated derivatives of the cyclic peptide fungicide caspofungin. ChemMedChem 2012, 7, 1496–1503. [Google Scholar] [CrossRef]
- Kubiczek, D.; Raber, H.; Gonzalez-García, M.; Morales-Vicente, F.; Staendker, L.; Otero-Gonzalez, A.J.; Rosenau, F. Derivates of the Antifungal Peptide Cm-p5 Inhibit Development of Candida auris Biofilms In Vitro. Antibiotics (Basel) 2020, 9. [Google Scholar] [CrossRef]
- Dahiya, R.; Kaur, K. Synthesis and pharmacological investigation of segetalin C as a novel antifungal and cytotoxic agent. Arzneimittelforschung 2008, 58, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Pesee, S.; Angkananuwat, C.; Tancharoensukjit, S.; Muanmai, S.; Sirivan, P.; Bubphawas, M.; Tanarerkchai, N. In vitro activity of Caspofungin combined with Fluconazole on mixed Candida albicans and Candida glabrata biofilm. Med Mycol 2016, 54, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez López, A.L.; Lee, M.R.; Wang, N.B.; Dunn, K.K.; Sanchez, H.; Raman, N.; Andes, D.R.; Lynn, D.M.; Palecek, S.P. Small-Molecule Morphogenesis Modulators Enhance the Ability of 14-Helical β-Peptides To Prevent Candida albicans Biofilm Formation. Antimicrob Agents Chemother 2019, 63. [Google Scholar] [CrossRef] [PubMed]
- Muhaj, F.F.; George, S.J.; Nguyen, C.D.; Tyring, S.K. Antimicrobials and resistance part II: Antifungals, antivirals, and antiparasitics. J Am Acad Dermatol 2022, 86, 1207–1226. [Google Scholar] [CrossRef]
- Neoh, C.F.; Jeong, W.; Kong, D.C.; Slavin, M.A. The antifungal pipeline for invasive fungal diseases: what does the future hold? Expert Rev Anti Infect Ther 2023, 21, 577–594. [Google Scholar] [CrossRef]
- Kersten, R.D.; Weng, J.K. Gene-guided discovery and engineering of branched cyclic peptides in plants. Proc Natl Acad Sci U S A 2018, 115, E10961–e10969. [Google Scholar] [CrossRef]
- Gerlach, S.L.; Göransson, U.; Kaas, Q.; Craik, D.J.; Mondal, D.; Gruber, C.W. A systematic approach to document cyclotide distribution in plant species from genomic, transcriptomic, and peptidomic analysis. Biopolymers 2013, 100, 433–437. [Google Scholar] [CrossRef]
- Hellinger, R.; Sigurdsson, A.; Wu, W.; Romanova, E.V.; Li, L.; Sweedler, J.V.; Süssmuth, R.D.; Gruber, C.W. Peptidomics. Nature Reviews Methods Primers 2023, 3, 25. [Google Scholar] [CrossRef]
- Gupta, R.; Srivastava, D.; Sahu, M.; Tiwari, S.; Ambasta, R.K.; Kumar, P. Artificial intelligence to deep learning: machine intelligence approach for drug discovery. Mol Divers 2021, 25, 1315–1360. [Google Scholar] [CrossRef] [PubMed]
- Melo, M.C.R.; Maasch, J.R.M.A.; de la Fuente-Nunez, C. Accelerating antibiotic discovery through artificial intelligence. Communications Biology 2021, 4, 1050. [Google Scholar] [CrossRef] [PubMed]
- Ermert, P.; Luther, A.; Zbinden, P.; Obrecht, D. Frontier Between Cyclic Peptides and Macrocycles. Methods Mol Biol 2019, 2001, 147–202. [Google Scholar] [CrossRef] [PubMed]
- Mulder, M.P.C.; Fodran, P.; Kemmink, J.; Breukink, E.J.; Kruijtzer, J.A.W.; Minnaard, A.J.; Liskamp, R.M.J. Mutual influence of backbone proline substitution and lipophilic tail character on the biological activity of simplified analogues of caspofungin. Organic & Biomolecular Chemistry 2012, 10, 7491–7502. [Google Scholar] [CrossRef]
- Logan, A.; Wolfe, A.; Williamson, J.C. Antifungal Resistance and the Role of New Therapeutic Agents. Curr Infect Dis Rep 2022, 24, 105–116. [Google Scholar] [CrossRef]
- Johnson, M.D.; MacDougall, C.; Ostrosky-Zeichner, L.; Perfect, J.R.; Rex, J.H. Combination antifungal therapy. Antimicrob Agents Chemother 2004, 48, 693–715. [Google Scholar] [CrossRef]
- Lignieres, G.; Guitard, J.; Alby-Laurent, F.; Rambaud, J.; Bigot, J.; Morand, K.; Leverger, G.; Tabone, M.D. Antifungal combination therapy for invasive fungal infections in a paediatric oncology and haematology department: A retrospective analysis of practice. J Mycol Med 2022, 32, 101276. [Google Scholar] [CrossRef]
- Su, S.; Yan, H.; Min, L.; Wang, H.; Chen, X.; Shi, J.; Sun, S. The antifungal activity of caspofungin in combination with antifungals or non-antifungals against Candida species in vitro and in clinical therapy. Expert Rev Anti Infect Ther 2022, 20, 161–178. [Google Scholar] [CrossRef]
- Chassot, F.; Venturini, T.P.; Piasentin, F.B.; Santurio, J.M.; Svidzinski, T.I.E.; Alves, S.H. Activity of antifungal agents alone and in combination against echinocandin-susceptible and -resistant Candida parapsilosis strains. Revista Iberoamericana de Micología 2019, 36, 44–47. [Google Scholar] [CrossRef]
- Hatipoglu, N.; Hatipoglu, H. Combination antifungal therapy for invasive fungal infections in children and adults. Expert Rev Anti Infect Ther 2013, 11, 523–535. [Google Scholar] [CrossRef]
- Wang, Y.; Yan, H.; Li, J.; Zhang, Y.; Wang, Z.; Sun, S. Antifungal activity and potential mechanism of action of caspofungin in combination with ribavirin against Candida albicans. Int J Antimicrob Agents 2023, 61, 106709. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mode of action of antifungal peptides [6].
Figure 1.
Mode of action of antifungal peptides [6].
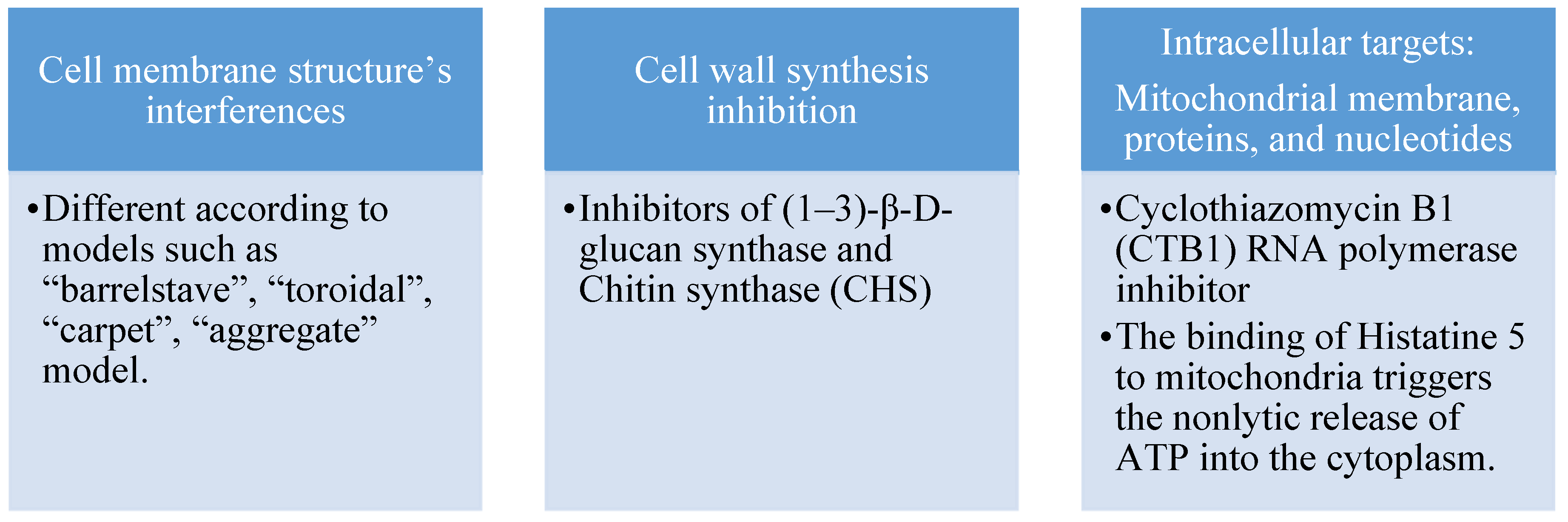
Figure 2.
Chemical structures of Valinomycin, Syringomycin E, calophycin, Aculeacin A, and three antifungal cyclic peptide drugs (Micafungin, Caspofungin, and Anidulafungin) approved for clinical use in the last two decades.
Figure 2.
Chemical structures of Valinomycin, Syringomycin E, calophycin, Aculeacin A, and three antifungal cyclic peptide drugs (Micafungin, Caspofungin, and Anidulafungin) approved for clinical use in the last two decades.
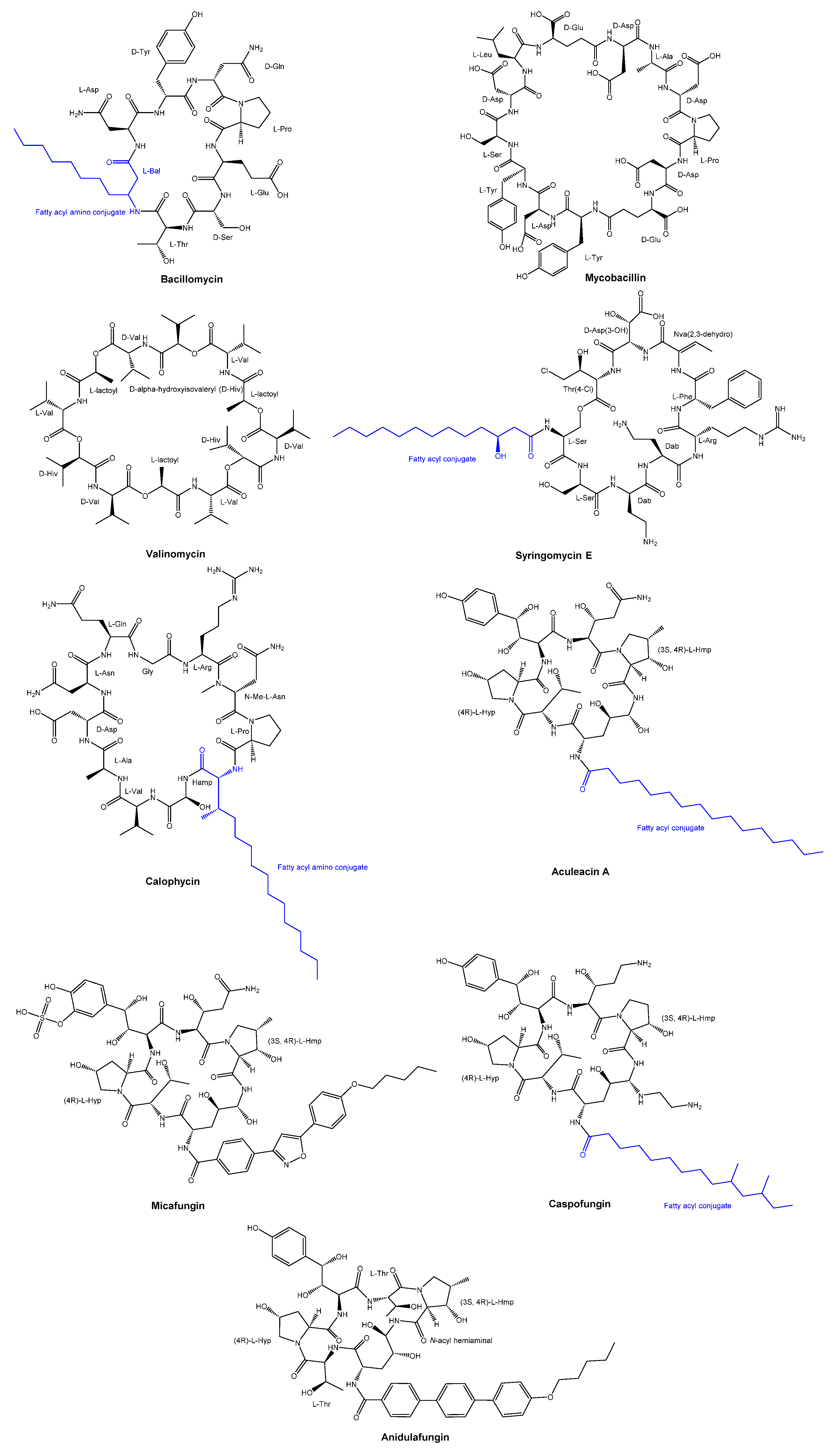
Figure 3.
Some cyclic peptides isolated from Cyanobacteria includes Tolybyssidins A and B, Majusculamide C, Anabaenolysin A, Balticidins A-D, Puwainaphycin E, laxaphycin A and B, and Hassallidin A.
Figure 3.
Some cyclic peptides isolated from Cyanobacteria includes Tolybyssidins A and B, Majusculamide C, Anabaenolysin A, Balticidins A-D, Puwainaphycin E, laxaphycin A and B, and Hassallidin A.
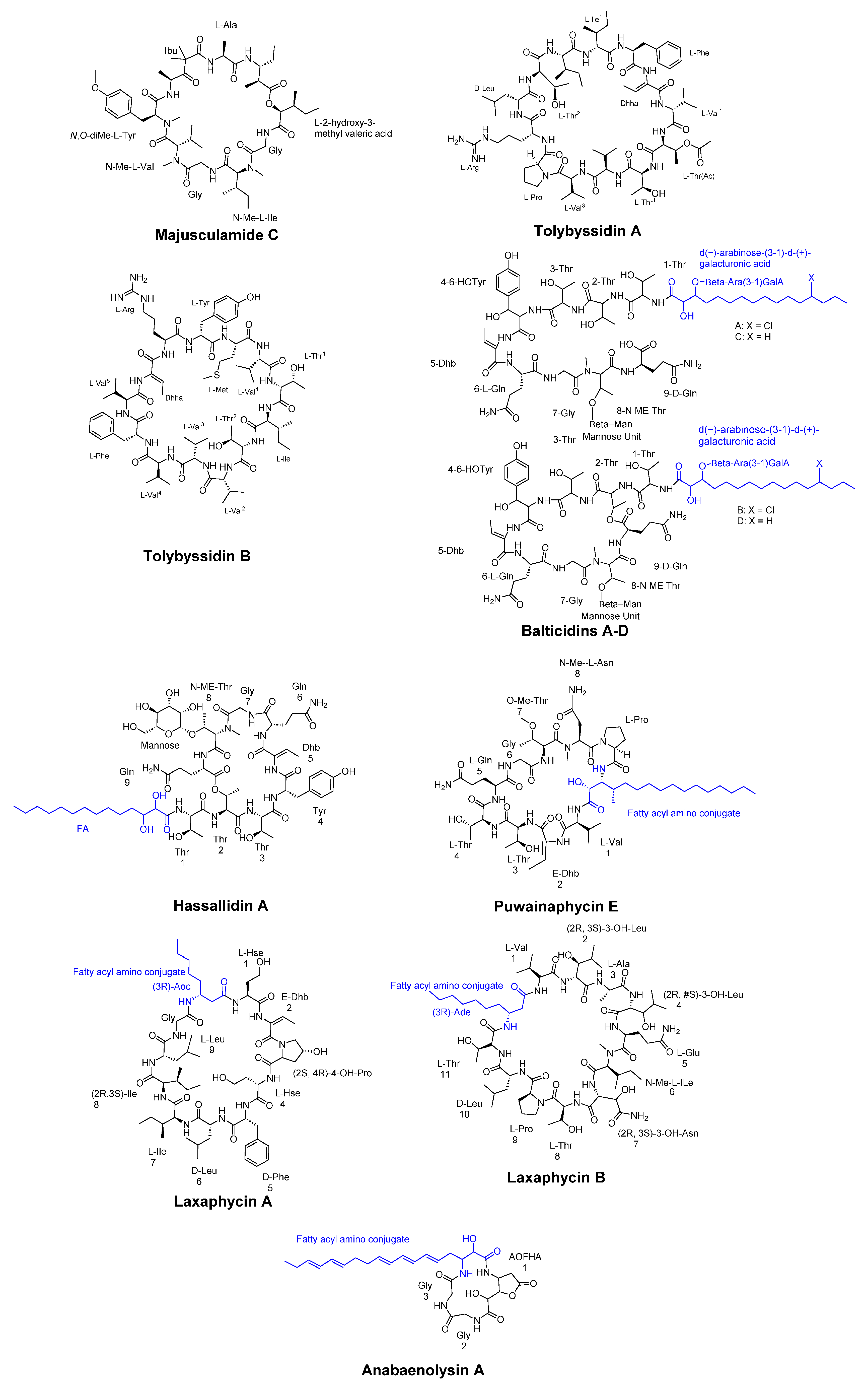
Figure 4.
Structures of isolated compounds MIN A, PUW F, and MIN C.
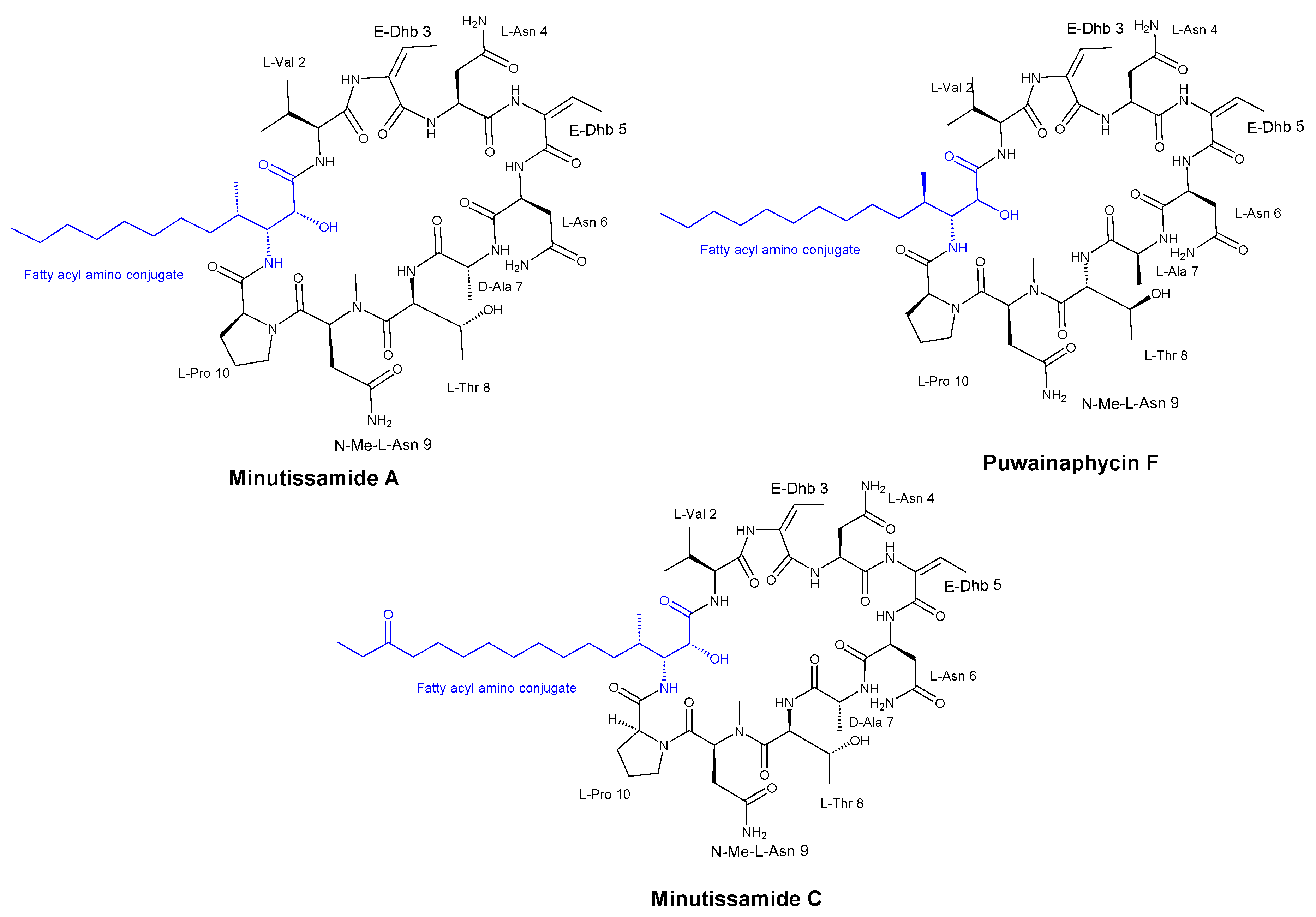
Figure 5.
Chemical structures of bacillopeptins A, B, C and fusaricidins A, C, and D.
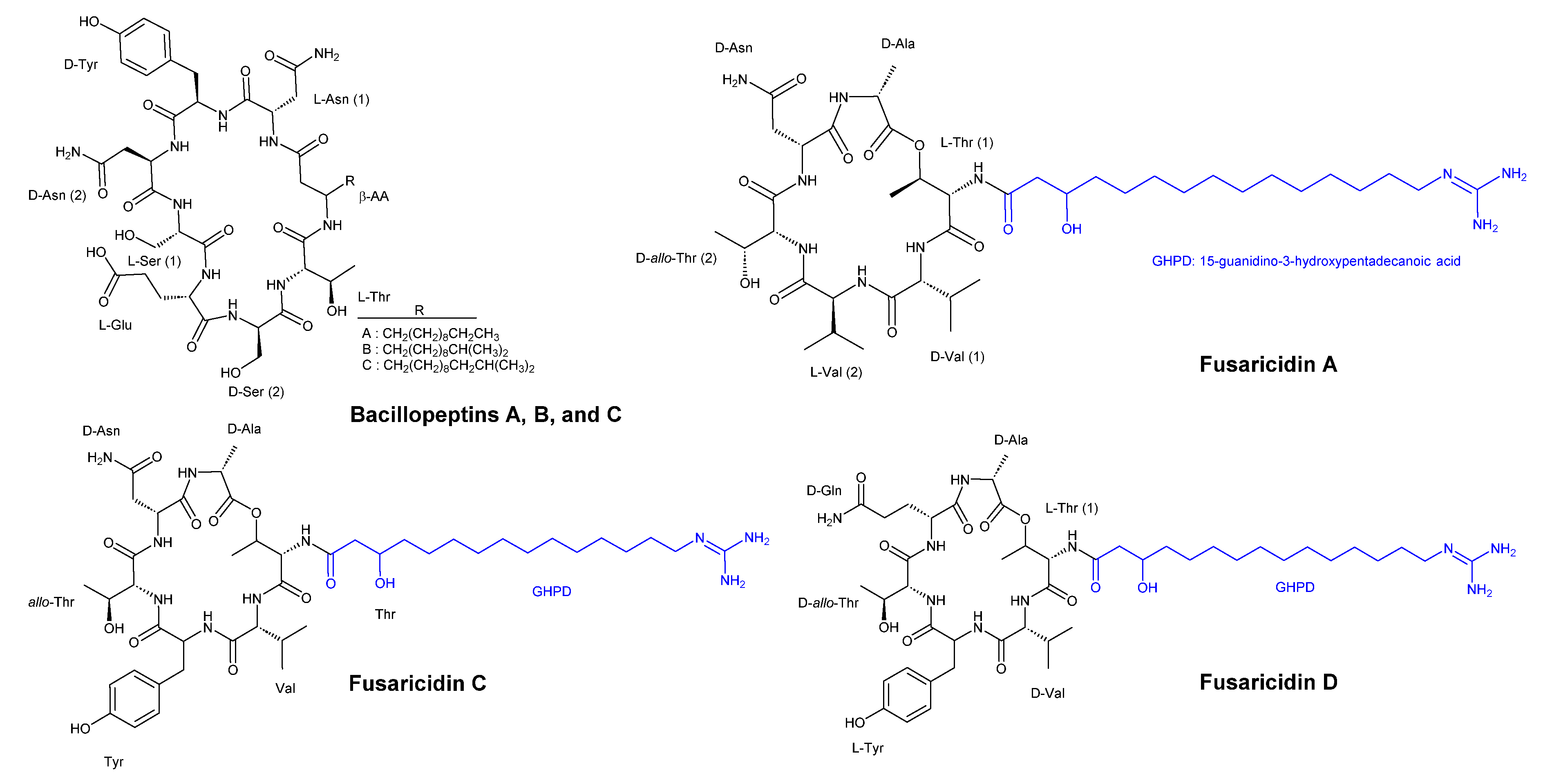
Figure 6.
Structures of compounds 1 and 2 from the surfactins family, anteiso-C17 mojavensin A (1), iso-C16 fengycin B, and anteiso-C17 fengycin B, and two cyclic lipopeptides, maribasins A and B.
Figure 6.
Structures of compounds 1 and 2 from the surfactins family, anteiso-C17 mojavensin A (1), iso-C16 fengycin B, and anteiso-C17 fengycin B, and two cyclic lipopeptides, maribasins A and B.
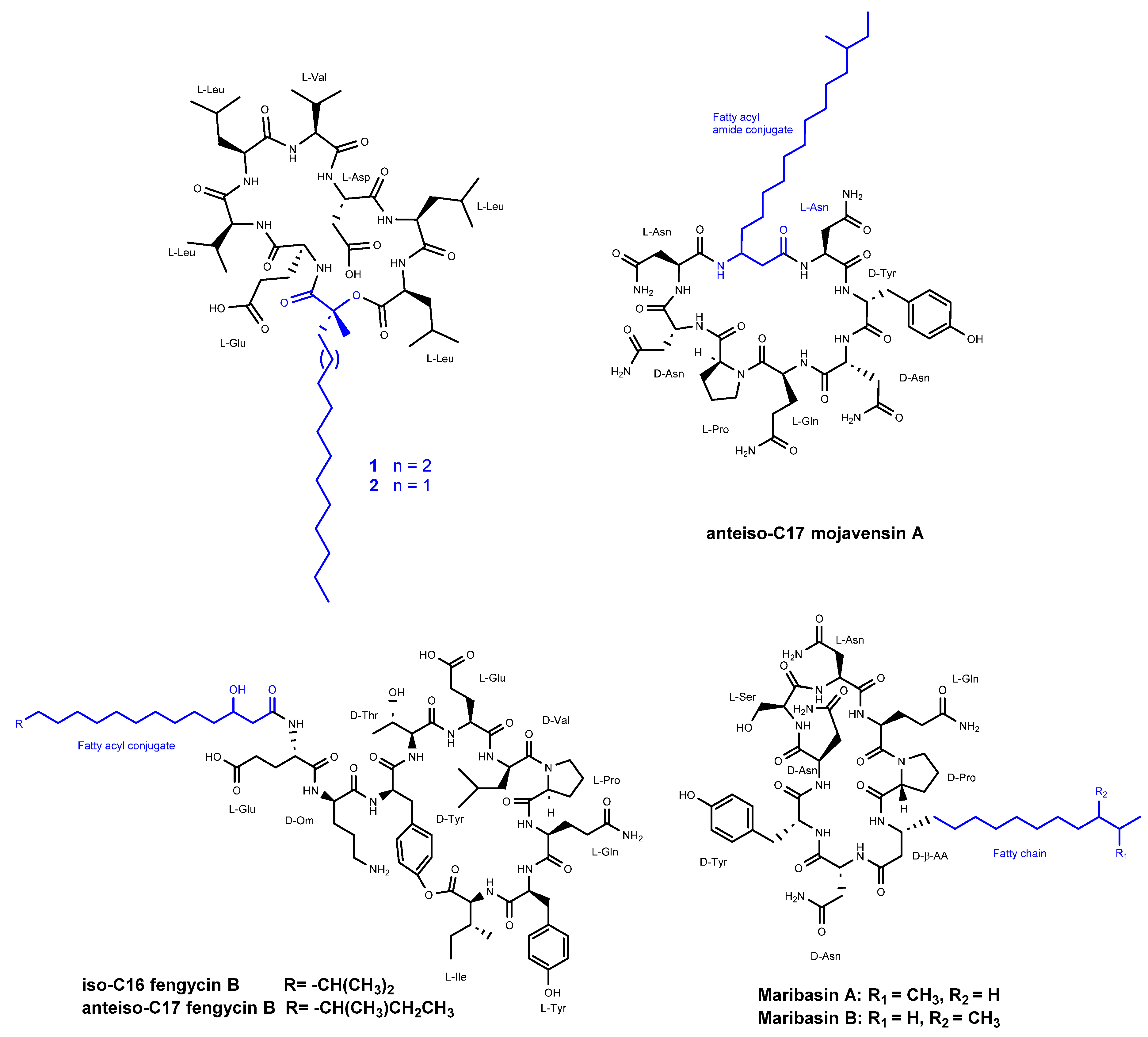
Figure 7.
Structure of five cyclic lipopeptides (CLPS) obtained from Neobacillus drentensis AKLSR2. Stereochemistry of these compounds were not reported.
Figure 7.
Structure of five cyclic lipopeptides (CLPS) obtained from Neobacillus drentensis AKLSR2. Stereochemistry of these compounds were not reported.
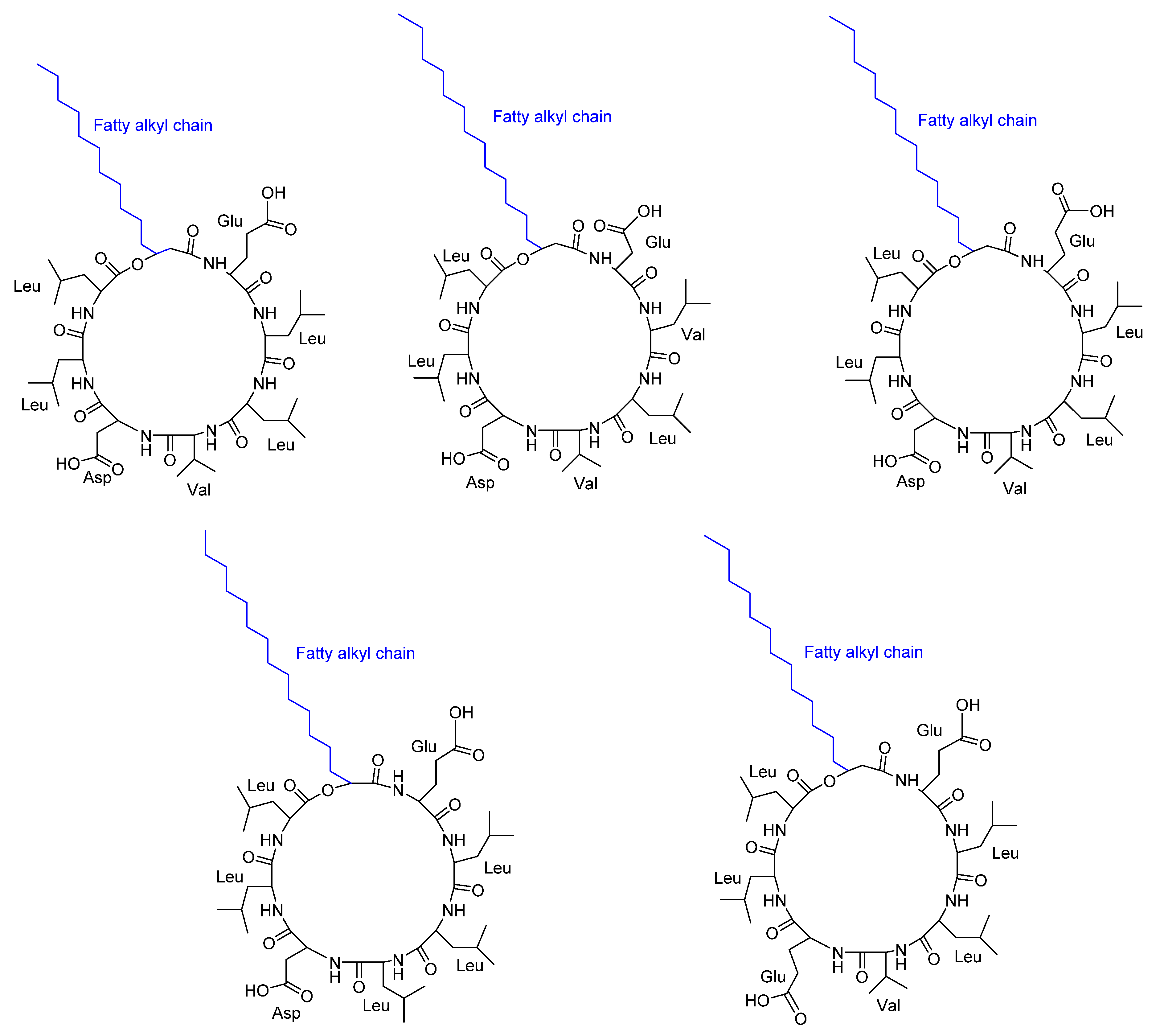
Figure 8.
BK-1097 and BK-1097 analogs.
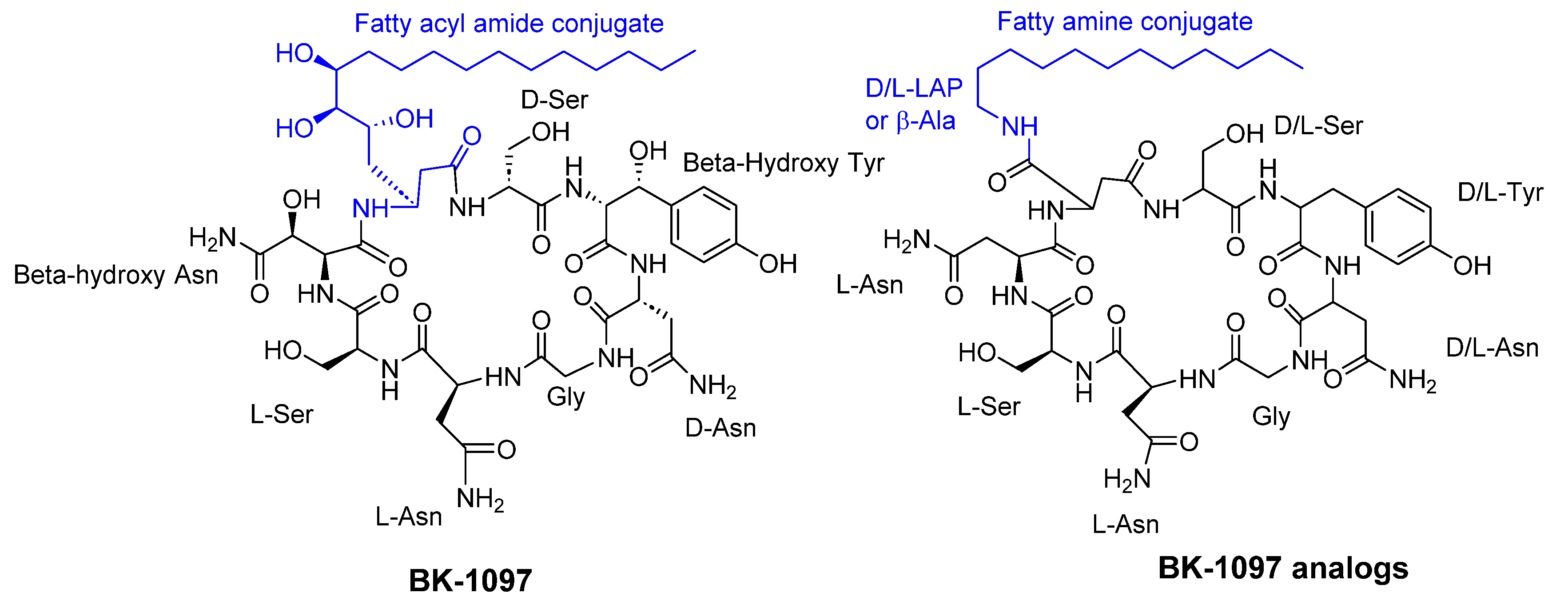
Figure 9.
The structures of cyclic peptides isolated from fungi.
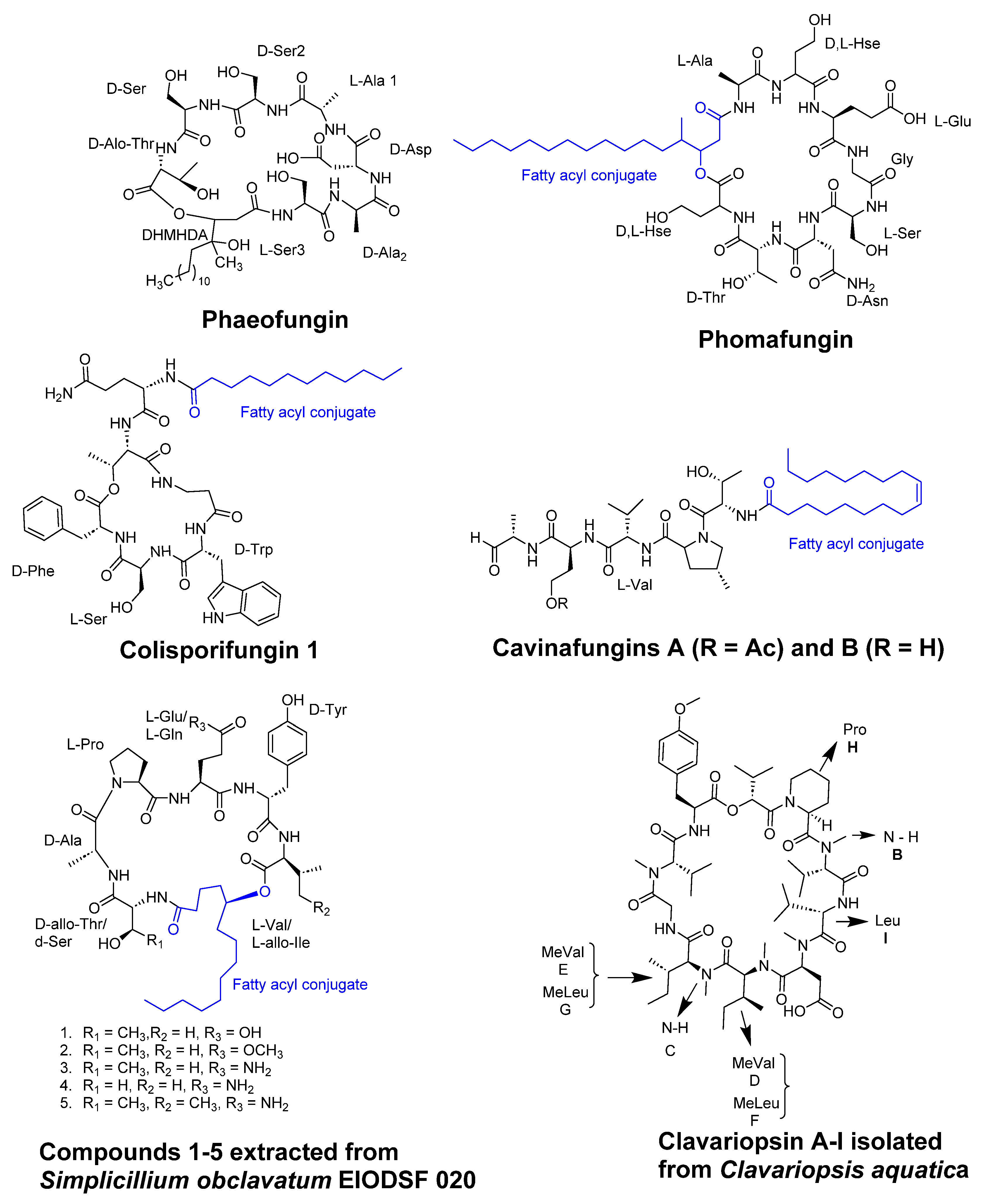
Figure 10.
Structure of Tunicyclins B-D.
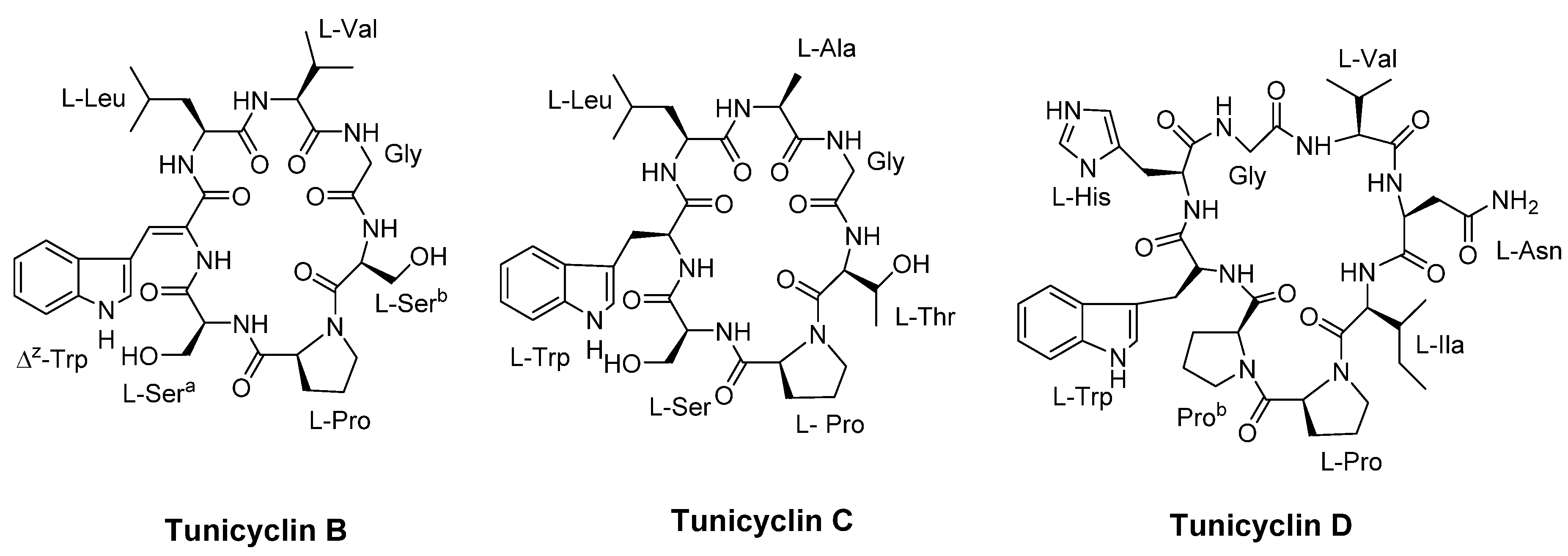
Figure 11.
Structures of AMT1, AMT2, AMT3, cyclo-AMT1, cyclo-AMT2 and cyclo-AMT3.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



