
Submitted:
06 June 2023
Posted:
07 June 2023
You are already at the latest version
Abstract
Research efforts toward Philippine biodiversity have increased significantly due to concerns about its natural resources and biodiversity. However, there is an imbalance in the distribution of published studies, with specific territories and organisms receiving more attention. Mindanao, a southern Philippine Island, is known for its high levels of endemism for amphibians and reptiles. Despite this, research on these groups remains unconsolidated. To address this gap, we conducted a biodiversity synthesis focusing on amphibians and reptiles in the faunal region of Mindanao. We found an increasing effort towards herpetofauna over the two decades, yet there is a clear bias in the effort towards amphibians compared to reptiles. In addition, we found that amphibians have a significantly higher species threat index than reptiles and all other vertebrate groups, regardless of ecological status. Yet, threatened herpetofauna species and those facing a greater number of threats are poorly studied, in general. Understanding the factors driving research biases in Mindanao and the Philippines is essential to allow us to redirect research efforts to the most needed areas, particularly to understudied and neglected taxa in Mindanao. This is especially urgent in the face of changing climate, where diverse communities of herpetofauna are at increased risk of extinction. Consequently, by bridging these gaps in biodiversity information, we can effectively contribute to the conservation and understanding of biodiversity in the region.
Keywords:
Amphibians
; Conservation
; Islands
; Philippines
; Priorities.
1. Introduction
Ecosystems and human societies widely depend on biodiversity, but the number of species and their geographic ranges are diminishing [1,2]. The current estimate shows that the pace of species extinction could be up to a thousand times higher than in the pre-industrialized period [3]. Among tetrapods, amphibians and reptiles are particularly susceptible to environmental changes due to their traits sensitive to rapidly changing ecological conditions, such as increased temperature and changes in precipitation [4,5,6]. Land use, climate change, disease, and alien species have been negatively affecting the population of amphibians and reptiles for many years [7]. Additionally, direct anthropogenic activities, including hunting, trade, and pollution, negatively impact the ability of their populations to recover from loss [5]. The extreme sensitivity of amphibians to environmental changes makes them ideal bioindicators of the health of their environment and would help indicate conservation priorities [6,7].
The Philippine archipelago, consisting of more than 7000 islands, holds high herpetofauna biodiversity [8]. This high species diversity makes the country one of Southeast Asia’s hotspots for herpetofauna biodiversity [4,9]. However, many species and habitats in the Philippines are threatened, mainly by the growing human population and deforestation in lowland areas [10,11]. Therefore, identifying species and habitats at higher risk of loss and extinction is crucial to prioritize conservation efforts and reduce further negative impacts on biodiversity [12,13,14]. One of the approaches to identifying conservation priorities is to examine the proportion of threatened and endemic species, assess the extent of the threats they face and evaluate ongoing conservation efforts directed towards these species, and analyze how these threats interact with species traits [15]. Unfortunately, valuable information for comprehensive synthesis remains unconsolidated for many regions in the Philippines [16]. Moreover, the existing syntheses aimed at understanding extinction risks are often broad in scope and present challenges when it comes to their local or national integration. These challenges are particularly true in various parts of the Philippines.
The amount of research on Philippine biodiversity has increased significantly in recent years, commonly driven by concerns about the state of its biodiversity and natural resources, which have spurred funding and conservation efforts [11,17]. However, there is a noticeable imbalance in research focus, with certain territories and organisms receiving disproportionately more attention. Mindanao supports unusually high levels of endemism for both amphibians and reptiles [9,18,19]. Nevertheless, like any other taxonomic group, data for herpetofauna species in Mindanao remain sparse and unconsolidated, impeding the effective identification of key species and conservation priorities [8,20,21]. To address this gap, we have developed a biodiversity synthesis aimed at understanding the progress, patterns, and distribution of herpetofauna in Mindanao, Philippines. Specifically, our objective was to determine (i) the levels and factors of research effort on amphibians and reptiles, (ii) the relationship between their extinction risk and ecological status, (iii) the diversity and distribution of amphibians and reptiles across biodiversity sites in Mindanao faunal region, and (iv) determine the key threatening processes and levels of threats to amphibians and reptiles.
By identifying and addressing these biases, our goal is to bolster the overall understanding of herpetofauna's biodiversity in Mindanao. Furthermore, we intend to prioritize conservation efforts toward species in Mindanao that are both threatened and understudied.
2. Materials and Methods
2.1. Study area and synthesis framework
The focus of this study was on biodiversity hotspots in the Mindanao Faunal Region (MFR), which comprises the entire Mindanao archipelago, the Leyte and Samar islands, Bohol, Basilan, and the nearby minor islands (Figure 1). We adopted a framework to develop the synthesis by integrating the literature from 2000 to 2022 to assess species research efforts and levels of threat [22].
2.2. Literature search and databasing
We used the PRISMA approach to search key databases, including Google Scholar (https://scholar.google.com), ResearchGate (https://www.researchgate.net), and personal correspondence to create a database of amphibian and reptile literature from January 2000 to June 2022. We used the following keywords: (amphibian* or frog* or newt* or caecilian* or toads*) AND reptile OR snake* OR lizard* OR turtle* OR squamate*) AND (Philippine* OR Mindanao*) to search for literature [23]. Most Philippine academic institutions do not have an accessible online library or thesis repository [24]. We excluded unpublished theses and dissertations from the analysis to avoid incomplete and biased data sampling. All publications collated from the searched results were screened and classified by taxonomic group, year of publication, site, and thematic areas using standardized criteria [25].
We curated and updated species names from recent literature (e.g., [18,26], the International Union for the Conservation of Nature (IUCN) Red List (version 2022-2), the Amphibia Web database (https://amphibiaweb.org/search/) [27] and the Reptile Database (https://reptile-database.reptarium.cz/) [28]. We determined species endemism and conservation status using the IUCN Red List (v. 2022-2).
2.3. Diversity and Distribution of Amphibians and Reptiles
We investigated the relationship between species diversity, endemism, and species binomial extinction risk (threatened and non-threatened). To determine the risk of extinction, we used the IUCN red list classification [29]. We classified the species as ‘threatened’ when the species is ‘vulnerable’, ‘endangered’, or ‘critically endangered’. On the other hand, we classified species as ‘least threatened’ if they were categorized as ‘least concerned’ or ‘near threatened’. Additionally, we included species classified as ‘data deficient’ in the same category as ‘threatened’ when comparing species diversity. We made this assumption because species lacking information may have limited occurrences and are likely to be endangered [30,31]. We compared the species richness per site based on the number of species recorded.
The Chi-square test of independence (χ2) was then applied to assess the relationship between extinction risk, taxonomic groups, and ecological status. Specifically, we examined levels of endemism and trophic levels. We also employed this analysis to compare threatened categories and ecological status between biodiversity hotspots or sites.
2.4. Site-level comparison
We compared species diversity and ecology between sites using the Chi-square test of independence (χ2). Georeferenced species records were modeled in MaxEnt to broadly predict and visualize the potential spatial distribution of amphibians and reptiles in the MFR [32]. MaxEnt uses environmental data to estimate a probability distribution for the entire study area based on the available presence-only information. The modeling process incorporated nineteen bioclimatic variables obtained from WorldClim [33], which had a resolution of a 30 arc-second grid or 1 km2. Additionally, altitude was included as a factor in the modeling process. Furthermore, we estimated the distance and density of species recorded from terrestrial protected areas, which were then calculated using Quantum GIS.
Deforestation is a significant threat to terrestrial ecosystems in the Philippines, including key biodiversity sites. To assess the extent of deforestation and land use change and its relationship with species diversity and ecological status, we extracted data from [34] to measure tree cover loss and remaining forests. Kendall’s Tauτ B was used to examine the relationship between estimated species richness at the site level, %endemism and threatened species%, and the rate of change (e.g., rate of deforestation in protected areas). We used a generalized linear model (GLM) with Poisson distribution to determine the link between %remaining forest cover to %endemism and %threatened species in each key biodiversity site.
2.5. Statistical Analyses
3. Results
3.1. Diversity and distribution of herpetofauna in Mindanao
A total of 42 published articles were collated between 2000 and 2022. We found that some studies overlap with other classifications and taxonomic groups, with the majority focused on areas of diversity and community surveys (Figure 2a). Regarding the number of studies per taxonomic group, there were 36 studies for amphibians and 26 studies for reptiles. The mean annual number of studies was higher among amphibians (mean = 1.48/yr.) compared to reptiles (mean = 1.09/yr.) (Figure 2b). Research in both taxonomic groups experienced an overall increase, albeit with an irregular pattern. Specifically, there was a notable increase in growth in 2015, followed by an inconsistent trend with peaks in 2020 and 2021 (Figure 2a). Interestingly, we did not find significant differences in the number of studies between amphibians and reptiles (Mann-Whitney U test = 229.5, p = 0.431). Furthermore, correlation analysis showed that amphibians and reptiles were studied extensively together, as shown in their positive and significant correlation in terms of the number of species recorded species per year (Kendall’s τ B = 0.845, p < 0.001).
Based on the database we built, we recorded 160 species of herpetofauna from the MFR. This number was represented by 52 species of amphibians belonging to nine families and two orders and 108 species of reptiles belonging to sixteen families of two orders. Among amphibians, Rhacophoridae and Dicroglossidae were the most species-rich families recorded (n = 12 spp., 23%), followed by Microhylidae (n = 7 spp., 13%), Ranidae (n = 6 spp., 11.5%), Bufonidae (n = 5 spp., 10%), Ceratobatrachidae (n = 4 spp., 8%), Megophyridae (n = 3 spp., 6%), Eleutherodactylidae and Ichthyophiidae both have the single species recorded (n = 1 spp., 2%). Whereas within reptile families, Scinidae was the most species-rich (n = 35 spp., 32%), followed by Gekkonidae (n = 15 spp., 14%), Colubridae (n = 13 spp., 12%), Agamidae (n = 11 spp., 10%), while other families were represented by fewer than ten species. Regarding species endemism, we found higher endemism among amphibians (63%, n = 33 spp.) relative to reptiles (58%, n = 63 spp.).
SREA was significantly higher among amphibians than among reptiles and all other taxonomic groups (Figure 3). Within amphibians, we found that non-threatened species (mean = 0.356 ± 0.299) had higher SREA compared to threatened groups (mean = 0.145 ± 0.145), but not statistically significant (H = 3.169, df = 1, p = 0.075). In terms of geopolitical endemism, we found a significantly higher SREA among Philippine endemic species (mean = 0.395 ± 0.316) versus non-endemic species (mean = 0.232 ± 0.220) (H = 4.186, df = 1, p = 0.041). However, among reptiles, we found that SREA did not differ significantly in terms of endemism but was higher between species endemic to the Philippines (mean = 0.183 ± 0.137) compared to their more widespread counterparts (Figure 3).
3.2. Conservation status across ecological status
Twenty percent of all herpetofauna species were considered in the threatened category (VU = 4 spp., EN = 3 spp., and DD = 14 spp.). The conservation status between taxonomic groups differed, with 14.8% and 9.6% of reptiles and amphibians being threatened, respectively. Among nine amphibian families, Dicroglossidae (25%, n = 3 spp.) and Ceratobatrachidae (20%, n = 1 spp.) had the highest proportions of threatened species (χ2 = 7.317, df = 8, p = 0.503). Although no significant differences were found in the distribution of threatened species between the 17 families of reptiles (χ2 = 22.458, df = 16, p = 0.129), we documented that at least six families had a remarkably higher proportion of threatened species, particularly Geomydidae (100%, n = 1 spp.), Elapidae (50%, n = 2 spp.), Typhlopidae (50%, n = 1), Agamidae (36.4%, n = 4), and Calamariidae (33.33%, n = 1). Furthermore, the proportion of threatened species between endemic species only differed significantly among reptiles (χ2 = 4.058, df = 1, p = 0.044) and was highest among those endemics in the Philippines (20.6%, n = 13 spp.). Although we did not find a significant relationship in the distribution of threatened species among amphibians (χ2 = 3.185, df = 1, p = 0.074), it was higher among Philippine endemic species (15.2%, n = 5 spp.) compared to those that are widespread (Figure 4a).
Among the key threats, bushmeat hunting, deforestation, and land use changes are the major threats to amphibians and reptiles (Figure 4b). Bushmeat consumption threatens 74% of amphibians and 55% of reptiles. Agriculture and aquaculture are the next significant threats, which threaten 64% of amphibians and 51% of reptiles (Figure 4b). Interestingly, urbanization (52%) and pollution (35%) threaten a high proportion of amphibians compared to reptiles.
We used the Species Threat Index (STI) to determine the level of threat each species faces according to its ecological status. Amphibians exhibited a significantly higher species threat index (SPI) compared to other taxonomic groups, such as birds and mammals, while reptiles displayed an exceptionally low SPI (Figure 7). Among ecological status, SPI was generally higher among threatened species of amphibians (0.500 ± 0.204) (H = 0.012, df = 1, p = 0.911) and reptiles (0.313 ± 0.175) (H = 6.285, df = 1, p = 0.012), but only statistically significant in the latter. We observed similar patterns of threat levels between Philippine endemic amphibians (0.556 ± 0.211) (H = 7.145, df = 1, p = 0.008) and reptiles (0.227 ± 0.166) (H = 4.534, df = 1, p = 0.033).
3.3. Relationship between Threat Index and Research Effort
We did not find a significant correlation between SREA and STI among amphibians (Kendall’s τ B = 0.007, p = 0.472) and reptiles (Kendall’s τ B = 0.062, p = 0.425). On the contrary, a positive and significant correlation between body mass and STI was specifically observed among reptiles (Figure 5). More importantly, our analysis revealed significant trends in the focus of research on the herpetofauna of Mindanao. Our findings revealed a negative correlation (β = -0.646, p < 0.001) between the allocation of the research effort and its threat status, indicating that greater efforts were directed towards non-threatened species (Table 1). Likewise, herpetofauna species with lower threat levels are studied more but were only significant for amphibians (β = -0.355, p < 0.001). Furthermore, Philippine endemic herpetofauna species exhibited a positive association with increased research effort (β = 0.415, p < 0.001), as did larger-bodied species (β = 0.272, p < 0.001). These trends were consistent in separate models for reptiles and amphibians, indicating an overall pattern in the relationship between the research effort and the ecological status of the species (Table 1).
3.4. Site-level Priorities
We recorded a total of 1060 (51%) amphibian and 1031 (49%) reptile occurrences at 28 biodiversity hotspot sites in Mindanao (Figure 8). Within this dataset, forty-four percent (44%) of the amphibian and 49% of the reptile records were within 10 km of protected areas. Among the 27 sites for amphibians, Mount Hibok-Hibok (n = 26 spp) and Mount Hilong-Hilong (n = 25 spp.) had the highest number of recorded species. While Mount Balatukan (100%, n = 25 spp.), Mount Lumot (77.4%, n = 24 spp.), Mount Magdiwata (78.1%, n = 25 spp.) had the highest number of Philippine endemic species (n = 30 spp.). However, among the sites included in our analysis, we observed a lower proportion of threatened amphibian species. Specifically, only two sites had a higher percentage of threatened amphibian species that exceeded 10%. Although 20 biodiversity hotspots for reptiles were studied, Mount Hamiguitan (n = 96 spp.) and Mount Magdiwata (n = 58 spp.) were the sites that had the highest species. Likewise, Mount Hamiguitan had the highest recorded endemic species (n = 63 spp., 66%), followed by Mount Magdiwata (n = 37 spp., 78%). Although reptile species exhibited a higher proportion in the distribution of threatened species among the analyzed sites. Specifically, three sites had more than 20% of their reptile species classified as threatened, while in five sites, at least 10% of the reptile species were in the threatened category (Figure 9). In general, the number of threatened species among both reptiles and amphibians did not show a significant difference between biodiversity sites.
Furthermore, we found a strong concordance between the species richness of amphibians and reptiles across sites (Kendall’s τ B = 0.626, p < 0.001). Similarly, we found a positive correlation between %endemism and %threatened among amphibians (Kendall’s τ B = 0.468, p = 0.007) and reptiles (Kendall’s τ B = 0.715, p < 0.001). Furthermore, we found that a higher proportion of threatened (β = 0.005, p = 0.034) and endemic (β = 0.006, p = 0.002) amphibian species, but not among reptiles, are related to the higher amount of remaining forest within biodiversity sites.
4. Discussion
Mindanao is home to a diverse range of unique and endangered taxa that provide valuable ecosystem services to humans. This study represents the first attempt to synthesize and analyze biodiversity conservation initiatives specifically targeting amphibians and reptiles in the megadiverse region of the Philippines. Our synthesis facilitated us to craft a consolidated understanding of the diversity and distribution of herpetofauna and their research gaps in the region.
Using our approach, we have accounted for 160 species of herpetofauna in the MFR, representing approximately 46% of the total herpetofauna species in the Philippines [18,26]. Our synthesis of the available data indicates that Mindanao has a high diversity of amphibians and reptiles. This is congruent with the broader patterns of distribution observed throughout the Philippines [8]. Various studies in the Philippines have shown that herpetofauna endemism is as high as 85%, with up to 23% of native amphibians being endemic to Mindanao [18,19,38]. Furthermore, our study found a positive correlation between research efforts and the number of herpetofauna species recorded in the last two decades. This growing record is due to the increased research efforts focused on herpetofauna in Mindanao, which advanced a better understanding of the distribution of amphibian and reptile biodiversity in the region. Recent comprehensive biodiversity assessments conducted in the various ecosystems of Mindanao have provided evidence of the exceptional levels of diversity and endemism of herpetofauna in the region. These assessments have shed light on the remarkable richness and unique composition of amphibians and reptiles found in Mindanao [26,39]. In Northeastern Mindanao and the surrounding islands, Sanguila et al. [26] recorded at least 126 amphibian and reptile species, including 40 species of frogs, one species of caecilian for amphibians, and 49 species of lizards, 35 species of snakes, one species of freshwater turtle, and one species of crocodile for reptiles. In addition, in the southern part of Mindanao, e.g., Mount Busa, which was poorly explored before, 68 species of herpetofauna were discovered, including 28 amphibians and 40 reptiles [40].
Yet, it is important to note that, despite the efforts made, individual contributions and efforts toward the conservation of amphibian and reptile species remain lacking, particularly for a significant proportion of these species. While amphibians and reptiles are often studied in tandem, we found a greater research focus on amphibians than on reptiles. A comparison of effort allocation measures across taxonomic groups revealed that amphibian species received more attention than birds and mammals [20]. Still, reptiles continue to be the most poorly studied group among terrestrial vertebrate groups. Our findings are consistent with a previous analysis showing that amphibians are studied and represented more in the literature than reptiles [41,42]. The observed bias in research between amphibians and reptiles may be explained by species traits and their probability of detection during field studies. For example, reptiles are often fossorial, making them more challenging to collect and study than those living aboveground, and this can lead to their poor representation in scientific collections and records relative to other groups [42,43,44]. In contrast, arboreal species, such as most amphibians, are easier to find and capture during visual surveys [45].
Furthermore, our analysis revealed a bias in research allocation toward larger herpetofauna species. This bias can be attributed to the practicality of detecting and studying larger species during fieldwork [42,46]. Their size and visibility make them more accessible and easier to observe and document than smaller, cryptic species. The preference to study larger herpetofauna species can inadvertently overlook the ecological importance and conservation needs of smaller species [47].
Contrary to our expectation endemic and rare species were more studied for both groups. Yet, we showed that threatened herpetofauna species were poorly studied, in general. This is because threatened species generally have narrower ranges and are therefore difficult to observe and study [48,49]. In addition, research on threatened species is often subject to more stringent regulations and permits, making research more difficult, costly, and time-consuming. This may contribute to the lower number of studies on threatened species. Our analysis also showed that amphibians face more threats than reptiles or any other group of vertebrates, regardless of their ecological status. Among the threats to herpetofauna are pollution, and habitat loss, which are likely driven by deforestation, shifting agriculture, and wildlife collection for consumption. Globally, amphibians and reptiles are especially susceptible to environmental changes and direct human activities [5,6]. Yet, amphibians are declining much faster than other terrestrial vertebrates, and there are multiple factors attributed to this trend [50,51]. Amphibians, besides being ectothermic, they have complex life cycles, making them usually more sensitive to rapid environmental disruption and threats such as habitat modification, pollution, and climatic change [7,52]. Furthermore, increasing levels of pollutants in freshwater ecosystems are also associated with direct mortality and physiological impacts on amphibian population decline [53]. Although there is limited research on reptiles, they are expected to be just as vulnerable to the same threats as amphibians [6,54,55], especially since the two groups share and occupy similar ecological requirements and habitats. Recent global analysis showed that 21% of global reptiles are in threatened categories and are facing significant threats similar to those of other tetrapods, including agriculture, logging, urban development, and invasive species [55]. Furthermore, our analysis showed that higher forest cover in key biodiversity sites supports more ecologically important species of herpetofauna. Tropical forest species of reptiles and amphibians, such as those of Mindanao forests, are more threatened and susceptible to many threats and population declines compared to species in other ecosystems [8,54,55]. This suggests that allocating more protection efforts in forest areas, especially in lowlands where threats are higher, will conserve more species and circumvent loss [20]. However, the clear potential impact of climate change on Mindanao herpetofauna remains unclear due to the lack of studies and investigations dedicated to the impacts of such threats [56].
This current state of herpetofauna biodiversity research in Mindanao highlights important trends and knowledge gaps that call for attention but also present an opportunity for future conservation efforts. By recognizing threats and their impacts on different species, we can strategically direct applied research efforts to identify the mechanisms driving these risks. The cryptic behavior of amphibians and reptiles requires long-term monitoring and effective consolidation of available information to effectively guide conservation strategies to mitigate and address the risk and challenges they face. Likewise, by understanding research biases toward different taxonomic groups in Mindanao, we can direct limited conservation funding to research efforts where they are most needed, that is, toward understudied and neglected taxa that may likely face more threats. Furthermore, considering the changing climate and the increasing human alterations of habitats, the need to address the issue becomes increasingly urgent, primarily due to the increased susceptibility of diverse herpetofauna communities to extinction [57,58]. Finally, efforts to increase knowledge and improve species inventory by promoting integrative taxonomic studies, building collaborative networks, and making biodiversity assessment data more accessible and open are among the few initiatives that should be sought to further advance herpetofauna research not only in Mindanao but throughout the Philippines.
Author Contributions
Conceptualization, K.C.T., S.S.A., and K.C.D.; methodology, K.C.T., S.S.A., and K.C.D.; validation, K.C.T., S.S.A., A.R.A. L.F.C. and K.C.D.; formal analysis, K.C.T., S.S.A., and K.C.D.; investigation, S.S.A.; resources, K.C.T.; data curation, K.C.T., S.S.A., A.R.A. L.F.C. and K.C.D.; writing—original draft preparation, S.S.A., A.R.A., and K.C.T.; writing—review and editing, K.C.T., S.S.A., A.R.A. L.F.C. and K.C.D; visualization, S.S.A., K.C.D., & K.C.T.; supervision, K.C.T., A.R.A., L.F.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research is self-funded and received no external funding from any institutions. The APC was funded by MDPI Conservation.
Data Availability Statement
Not Applicable.
Acknowledgments
Removed for peer-review.
Conflicts of Interest
The authors declare that the research was conducted without any commercial or financial relationship that could be construed as a potential conflict of interest.
References
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated Modern Human–Induced Species Losses: Entering the Sixth Mass Extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef]
- Wake, D.B.; Vredenburg, V.T. Are We in the Midst of the Sixth Mass Extinction? A View from the World of Amphibians. Proc. Natl. Acad. Sci. U.S.A. 2008, 105, 11466–11473. [Google Scholar] [CrossRef]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The Biodiversity of Species and Their Rates of Extinction, Distribution, and Protection. Science 2014, 344, 1246752. [Google Scholar] [CrossRef]
- Bickford, D.; Howard, S.D.; Ng, D.J.J.; Sheridan, J.A. Impacts of Climate Change on the Amphibians and Reptiles of Southeast Asia. Biodivers Conserv 2010, 19, 1043–1062. [Google Scholar] [CrossRef]
- Hoffmann, M.; Hilton-Taylor, C.; Angulo, A.; Böhm, M.; Brooks, T.M.; Butchart, S.H.M.; Carpenter, K.E.; Chanson, J.; Collen, B.; Cox, N.A.; et al. The Impact of Conservation on the Status of the World’s Vertebrates. Science 2010, 330, 1503–1509. [Google Scholar] [CrossRef]
- Böhm, M.; Collen, B.; Baillie, J.E.M.; Bowles, P.; Chanson, J.; Cox, N.; Hammerson, G.; Hoffmann, M.; Livingstone, S.R.; Ram, M.; et al. The Conservation Status of the World’s Reptiles. Biological Conservation 2013, 157, 372–385. [Google Scholar] [CrossRef]
- Hof, C.; Araújo, M.B.; Jetz, W.; Rahbek, C. Additive Threats from Pathogens, Climate and Land-Use Change for Global Amphibian Diversity. Nature 2011, 480, 516–519. [Google Scholar] [CrossRef]
- Diesmos, A.; Brown, R.; Alcala, A.; Sison, R.; Afuang, L.; Gee, G. Philippine Amphibians and Reptiles: An Overview of Species Diversity, Biogeography, and Conservation. In Philippine Biodiversity Conservation Priorities: a Second Iteration of the National Biodiversity Strategy and Action Plan; Department of the Environment and Natural Resources– Protected Areas and Wildlife Bureau, Conservation International Philippines, Biodiversity Conservation Program–University of the Philippines Center for Integrative and Developmental Studies, and Foundation for the Philippine Environment: Quezon City, Philippines, 2002; pp. 26–44. [Google Scholar]
- Diesmos, A.; Brown, R.; Alcala, A.; Sison, R.; Afuang, L.; Gee, G. Philippine Amphibians and Reptiles: An Overview of Species Diversity, Biogeography, and Conservation. 2023. [Google Scholar]
- Alcala, E.L.; Alcala, A.C.; Dolino, C.N. Amphibians and Reptiles in Tropical Rainforest Fragments on Negros Island, the Philippines. Environmental Conservation 2004, 31, 254–261. [Google Scholar] [CrossRef]
- Posa, M.R.C.; Diesmos, A.C.; Sodhi, N.S.; Brooks, T.M. Hope for Threatened Tropical Biodiversity: Lessons from the Philippines. BioScience 2008, 58, 231–240. [Google Scholar] [CrossRef]
- Hoffmann, M.; Brooks, T.M.; Fonseca, G.A.B. da; Gascon, C.; Hawkins, A.F.A.; James, R.E.; Langhammer, P.; Mittermeier, R.A.; Pilgrim, J.D.; Rodrigues, A.S.L.; et al. Conservation Planning and the IUCN Red List. Endangered Species Research 2008, 6, 113–125. [Google Scholar] [CrossRef]
- Rondinini, C.; Rodrigues, A.S.L.; Boitani, L. The Key Elements of a Comprehensive Global Mammal Conservation Strategy. Philosophical Transactions of the Royal Society B: Biological Sciences 2011, 366, 2591–2597. [Google Scholar] [CrossRef]
- Tanalgo, K.C.; Oliveira, H.F.M.; Hughes, A.C. Mapping Global Conservation Priorities and Habitat Vulnerabilities for Cave-Dwelling Bats in a Changing World. Science of The Total Environment 2022, 843, 156909. [Google Scholar] [CrossRef] [PubMed]
- Cooper, N.; Bielby, J.; Thomas, G.H.; Purvis, A. Macroecology and Extinction Risk Correlates of Frogs. Global Ecology and Biogeography 2008, 17, 211–221. [Google Scholar] [CrossRef]
- Heberling, J.M.; Miller, J.T.; Noesgaard, D.; Weingart, S.B.; Schigel, D. Data Integration Enables Global Biodiversity Synthesis. PNAS 2021, 118. [Google Scholar] [CrossRef]
- Mercado, D. Compilation & Synthesis of Valuation Studies on Philippine Biodiversity 2016.
- Diesmos, A.; Watters, J.; Huron, N.; Davis, D.; Alcala, A.; Crombie, R.; Afuang, L.; Gee-Das, G.; Sison, R.; Sanguila, M.; et al. Amphibians of the Philippines, Part 1: Checklist of the Species. 2015, 62.
- Sanguila, M.B.; Cobb, K.A.; Siler, C.D.; Diesmos, A.C.; Alcala, A.C.; Brown, R.M. The Amphibians and Reptiles of Mindanao Island, Southern Philippines, II: The Herpetofauna of Northeast Mindanao and Adjacent Islands. Zookeys 2016, 1–132. [Google Scholar] [CrossRef]
- Dela Cruz, K.C.; Abdullah, S.S.; Agduma, A.R.; Tanalgo, K.C. Early Twenty-First Century Biodiversity Data Pinpoint Key Targets for Bird and Mammal Conservation in Mindanao, Southern Philippines. Biodiversity 2023, 0, 1–18. [Google Scholar] [CrossRef]
- Agduma, A.R.; Garcia, F.G.; Cabasan, M.T.; Pimentel, J.; Ele, R.J.; Rubio, M.; Murray, S.; Hilario-Husain, B.A.; Dela Cruz, K.C.; Abdullah, S.; et al. Overview of Priorities, Threats, and Challenges to Biodiversity Conservation in the Southern Philippines. Regional Sustainability 2023, 4, 203–213. [Google Scholar] [CrossRef]
- Tanalgo, K.C.; Hughes, A.C. Priority-Setting for Philippine Bats Using Practical Approach to Guide Effective Species Conservation and Policy-Making in the Anthropocene. Hystrix 2019, 30, 0. [Google Scholar] [CrossRef]
- McInnes, M.D.F.; Moher, D.; Thombs, B.D.; McGrath, T.A.; Bossuyt, P.M.; Clifford, T.; Cohen, J.F.; Deeks, J.J.; Gatsonis, C.; Hooft, L.; et al. Preferred Reporting Items for a Systematic Review and Meta-Analysis of Diagnostic Test Accuracy Studies: The PRISMA-DTA Statement. JAMA 2018, 319, 388–396. [Google Scholar] [CrossRef]
- Guerrero, J.; Tanalgo, K.; Landscape Ecology Group, Center for Integrative Conservation, Xishuangbanna Tropical Botanical Garden, Menglun, Mengla 666303, Yunnan Province, P.R. China; Abante, C. Bicol University Research and Development Management Division, Legazpi City 4500, the Republic of the Philippines Role of Academic Institution to Inform Local and Regional Scale Biodiversity in the Eastern Philippines. J.Trop.Life.Science 2021, 11, 241–250. [Google Scholar] [CrossRef]
- Tanalgo, K.C.; Hughes, A.C. Bats of the Philippine Islands—A Review of Research Directions and Relevance to National-Level Priorities and Targets. Mammalian Biology 2018, 91, 46–56. [Google Scholar] [CrossRef]
- Sanguila, M.B.; Cobb, K.A.; Siler, C.D.; Diesmos, A.C.; Alcala, A.C.; Brown, R.M. The Amphibians and Reptiles of Mindanao Island, Southern Philippines, II: The Herpetofauna of Northeast Mindanao and Adjacent Islands. Zookeys 2016, 1–132. [Google Scholar] [CrossRef]
- AmphibiaWeb Database AmphibiaWeb Database Search . Available online: https://amphibiaweb.org/search/ (accessed on 5 April 2023).
- The Reptile Database The Reptile Database. Available online: https://reptile-database.reptarium.cz/ (accessed on 5 April 2023).
- Atwood, T.B.; Valentine, S.A.; Hammill, E.; McCauley, D.J.; Madin, E.M.; Beard, K.H.; Pearse, W.D. Herbivores at the Highest Risk of Extinction among Mammals, Birds, and Reptiles. Science advances 2020, 6, eabb8458. [Google Scholar] [CrossRef] [PubMed]
- Bland, L.M.; Collen, B.; Orme, C.D.L.; Bielby, J. Predicting the Conservation Status of Data-Deficient Species: Predicting Extinction Risk. Conservation Biology 2015, 29, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Tanalgo, K.C.; Tabora, J.A.G.; Hughes, A.C. Bat Cave Vulnerability Index (BCVI): A Holistic Rapid Assessment Tool to Identify Priorities for Effective Cave Conservation in the Tropics. Ecological Indicators 2018, 89, 852–860. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the Black Box: An Open-Source Release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very High Resolution Interpolated Climate Surfaces for Global Land Areas. International Journal of Climatology 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Daipan, B.P. Patterns of Forest Cover Loss in the Terrestrial Key Biodiversity Areas in the Philippines: Critical Habitat Conservation Priorities. Journal of Threatened Taxa 2021, 13, 20019–20032. [Google Scholar] [CrossRef]
- The Jamovi Project Jamovi (Version 2.3.22) [Computer Software]. 2023.
- GraphPad Prism GraphPad Software, Inc., San Diego, CA 2022.
- QGIS Development Team QGIS Geographic Information System. Open Source Geospatial Foundation Project. Http://Qgis.Osgeo.Org 2022.
- DIESMOS, A.C. Ecology and Diversity of Herpetofaunal Communities in Fragmented Lowland Rainforests in the Philippines. 2009. [Google Scholar]
- Plaza, J.; Sanguila, M. Preliminary Report on the Anurans of Mount Hilong-Hilong, Agusan Del Norte, Eastern Mindanao, Philippines. Asian Herpetological Research 2015, 6, 18–33. [Google Scholar] [CrossRef]
- Pitogo, K.M.; Saavedra, A.J.; Afuang, L. Amphibians and Reptiles of Mount Busa, Sarangani Province: A Glimpse of the Herpetological Community of Southern Mindanao, Philippines. Philippine Journal of Science 2021, 150, 1279–1306. [Google Scholar] [CrossRef]
- Christoffel, R.A.; Lepczyk, C.A. Representation of Herpetofauna in Wildlife Research Journals. The Journal of Wildlife Management 2012, 76, 661–669. [Google Scholar] [CrossRef]
- Guedes, J.J.M.; Moura, M.R.; Alexandre, F. Diniz-Filho, J. Species out of Sight: Elucidating the Determinants of Research Effort in Global Reptiles. Ecography 2023, 2023, e06491. [Google Scholar] [CrossRef]
- Colli, G.R.; Fenker, J.; Tedeschi, L.G.; Barreto-Lima, A.F.; Mott, T.; Ribeiro, S.L.B. In the Depths of Obscurity: Knowledge Gaps and Extinction Risk of Brazilian Worm Lizards (Squamata, Amphisbaenidae). Biological Conservation 2016, 204, 51–62. [Google Scholar] [CrossRef]
- Moura, M.R.; Costa, H.C.; Abegg, A.D.; Alaminos, E.; Angarita-Sierra, T.; Azevedo, W.S.; Cabral, H.; Carvalho, P.; Cechin, S.; Citeli, N.; et al. Unwrapping Broken Tails: Biological and Environmental Correlates of Predation Pressure in Limbless Reptiles. Journal of Animal Ecology 2023, 92, 324–337. [Google Scholar] [CrossRef]
- Petrovan, S.O.; Al-Fulaij, N.; Christie, A.; Andrews, H. Why Link Diverse Citizen Science Surveys? Widespread Arboreal Habits of a Terrestrial Amphibian Revealed by Mammalian Tree Surveys in Britain. PLOS ONE 2022, 17, e0265156. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, X.; Shine, R.; Lourdais, O. Taxonomic Chauvinism. Trends in Ecology & Evolution 2002, 17, 1–3. [Google Scholar] [CrossRef]
- Ruggieri-Mitchell, C. The More Things Changed, the More They Stayed the Same: Trends in Conservation Focus 2010–2019. Journal for Nature Conservation 2023, 73, 126403. [Google Scholar] [CrossRef]
- Ducatez, S.; Shine, R. Drivers of Extinction Risk in Terrestrial Vertebrates. Conservation Letters 2017, 10, 186–194. [Google Scholar] [CrossRef]
- Chen, C.; Chen, C.; Wang, Y. Ecological Correlates of Extinction Risk in Chinese Amphibians. Diversity and Distributions 2019, 25, 1586–1598. [Google Scholar] [CrossRef]
- Catenazzi, A. State of the World’s Amphibians. Annual Review of Environment and Resources 2015, 40, 91–119. [Google Scholar] [CrossRef]
- Harfoot, M.B.J.; Johnston, A.; Balmford, A.; Burgess, N.D.; Butchart, S.H.M.; Dias, M.P.; Hazin, C.; Hilton-Taylor, C.; Hoffmann, M.; Isaac, N.J.B.; et al. Using the IUCN Red List to Map Threats to Terrestrial Vertebrates at Global Scale. Nat Ecol Evol 2021, 5, 1510–1519. [Google Scholar] [CrossRef]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischman, D.L.; Waller, R.W. Status and Trends of Amphibian Declines and Extinctions Worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef]
- Polo-Cavia, N.; Burraco, P.; Gomez-Mestre, I. Low Levels of Chemical Anthropogenic Pollution May Threaten Amphibians by Impairing Predator Recognition. Aquatic Toxicology 2016, 172, 30–35. [Google Scholar] [CrossRef]
- Gibbons, J.W.; Scott, D.E.; Ryan, T.J.; Buhlmann, K.A.; Tuberville, T.D.; Metts, B.S.; Greene, J.L.; Mills, T.; Leiden, Y.; Poppy, S.; et al. The Global Decline of Reptiles, Déjà Vu Amphibians: Reptile Species Are Declining on a Global Scale. Six Significant Threats to Reptile Populations Are Habitat Loss and Degradation, Introduced Invasive Species, Environmental Pollution, Disease, Unsustainable Use, and Global Climate Change. BioScience 2000, 50, 653–666. [Google Scholar] [CrossRef]
- Cox, N.; Young, B.E.; Bowles, P.; Fernandez, M.; Marin, J.; Rapacciuolo, G.; Böhm, M.; Brooks, T.M.; Hedges, S.B.; Hilton-Taylor, C.; et al. A Global Reptile Assessment Highlights Shared Conservation Needs of Tetrapods. Nature 2022, 605, 285–290. [Google Scholar] [CrossRef]
- Alcala, A.; Bucol, A.; Diesmos, A.; Brown, R. Vulnerability of Philippine Amphibians to Climate Change. Philippine Journal of Science 2012, 141. [Google Scholar]
- Iglesias-Carrasco, M.; Medina, I.; Ord, T.J. Global Effects of Forest Modification on Herpetofauna Communities. Conservation Biology 2023, 37, e13998. [Google Scholar] [CrossRef]
- Tan, W.C.; Herrel, A.; Rödder, D. A Global Analysis of Habitat Fragmentation Research in Reptiles and Amphibians: What Have We Done so Far? Biodivers Conserv 2023, 32, 439–468. [Google Scholar] [CrossRef]
Figure 1.
Map of the Mindanao faunal region (hereafter ‘MFR’) [20].
Figure 1.
Map of the Mindanao faunal region (hereafter ‘MFR’) [20].
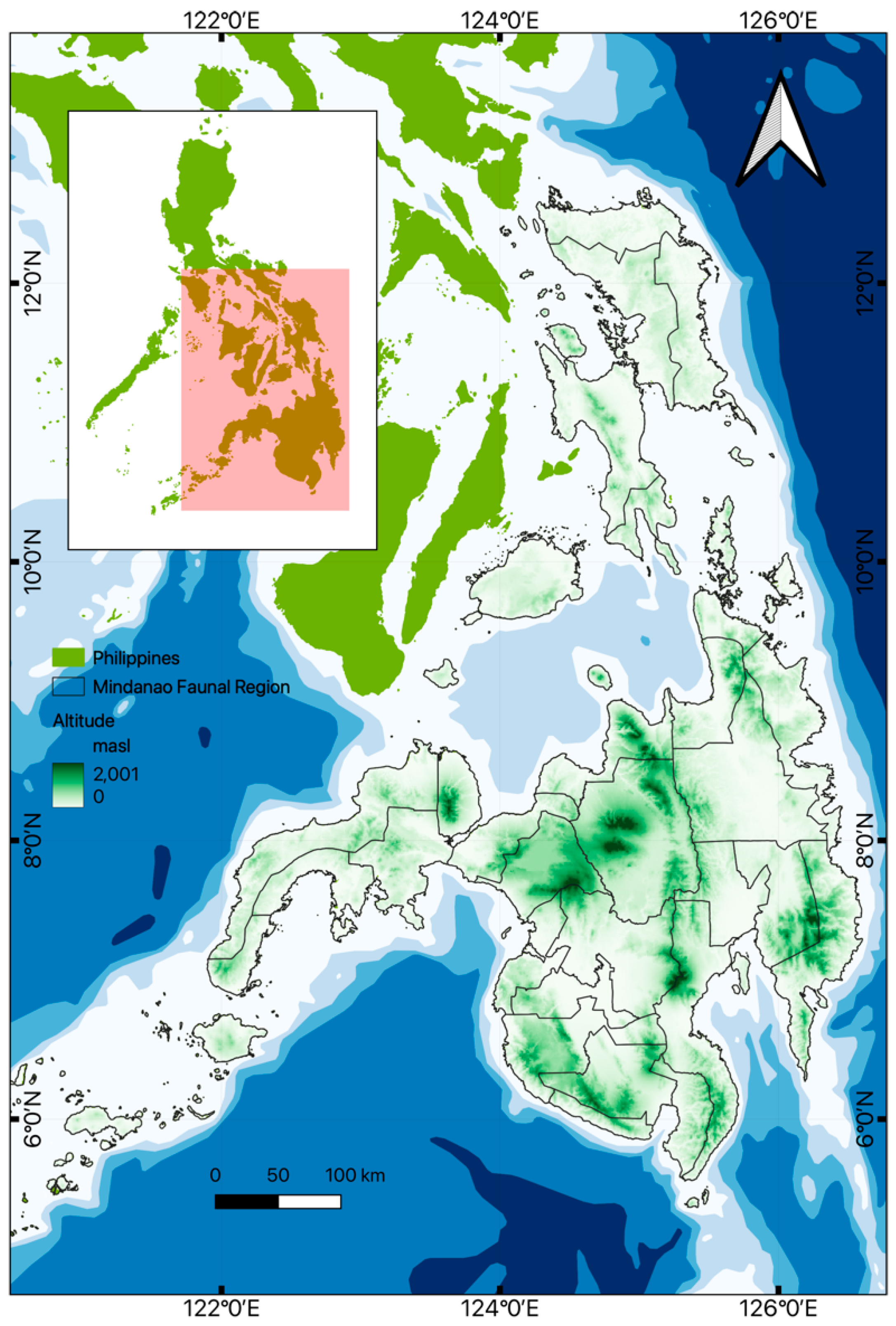
Figure 2.
Annual trends in the number (a) and average study (b) of amphibian and reptile research published in journals from 2000-2022, compared to other taxonomic groups. Broken lines indicate the average number of studies per year for birds and mammals.
Figure 2.
Annual trends in the number (a) and average study (b) of amphibian and reptile research published in journals from 2000-2022, compared to other taxonomic groups. Broken lines indicate the average number of studies per year for birds and mammals.
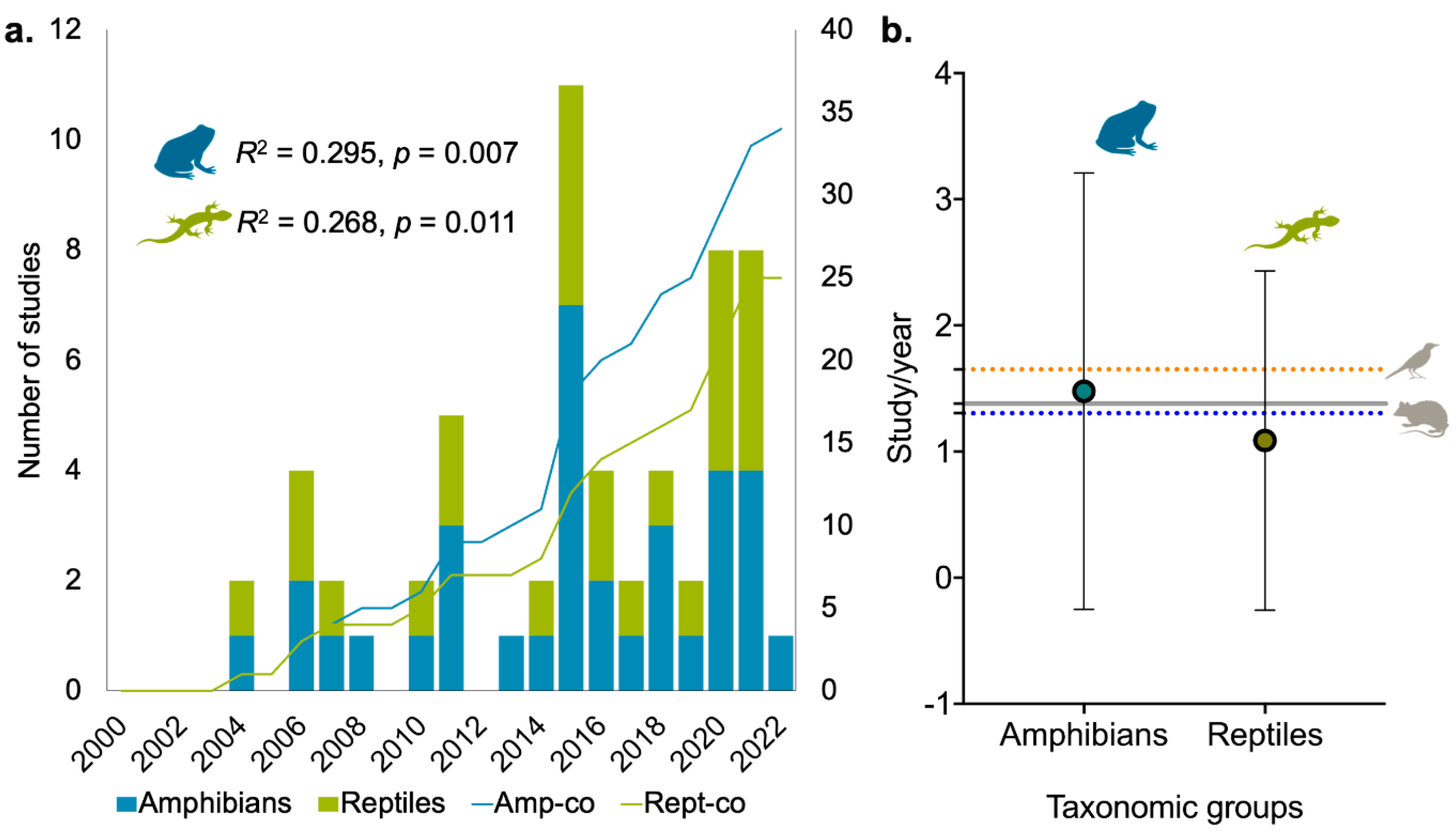
Figure 3.
Comparison of Species Research Effort Allocation (SREA) between the ecological status of amphibians and reptiles. Broken lines indicate the average study per year for birds and mammals.
Figure 3.
Comparison of Species Research Effort Allocation (SREA) between the ecological status of amphibians and reptiles. Broken lines indicate the average study per year for birds and mammals.
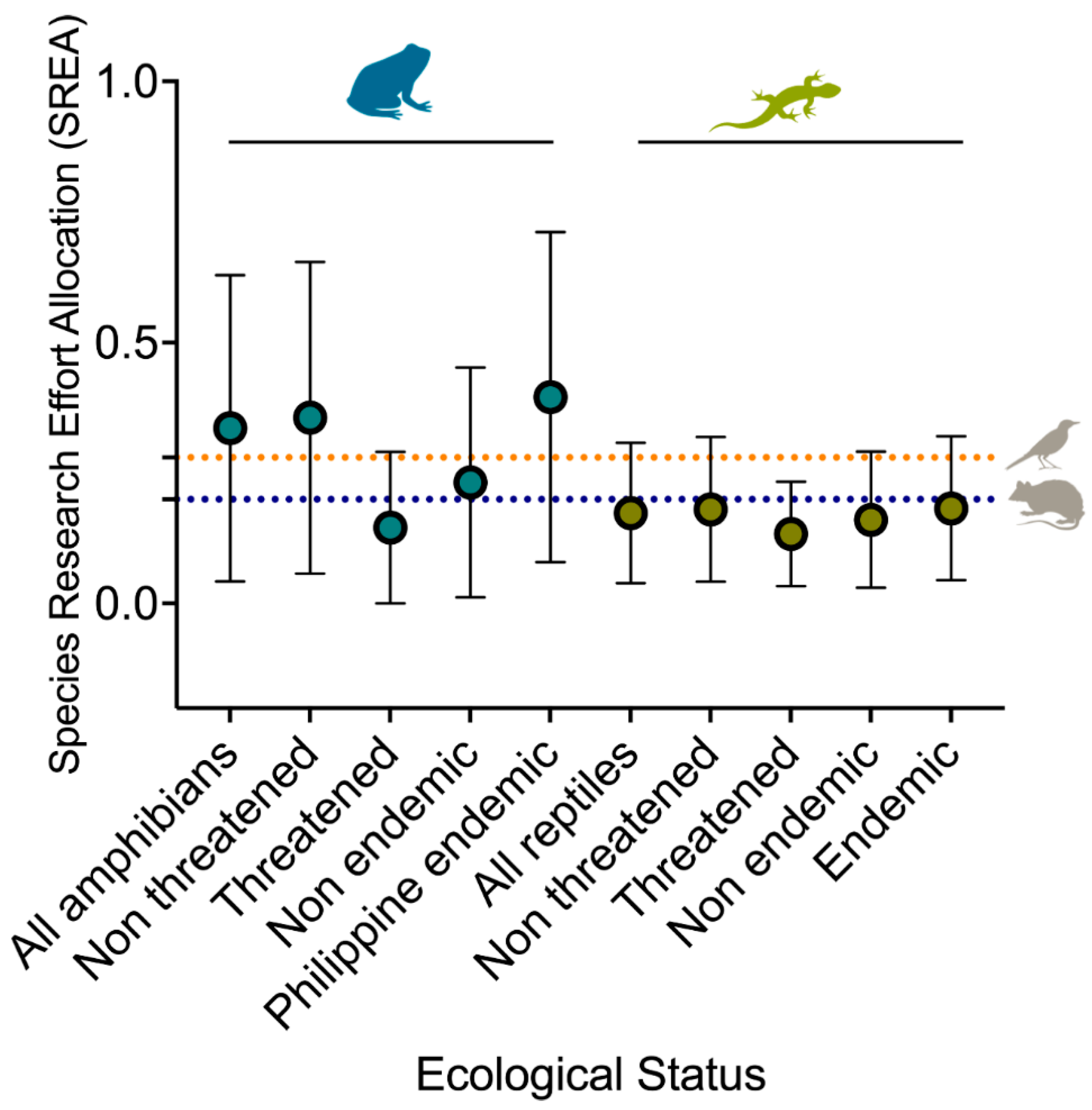
Figure 4.
Distribution and proportion of endemic and threatened amphibian and reptile species (a). Sankey plot showing the distribution of threats according to taxonomic group, endemism, and threatened categories (b).
Figure 4.
Distribution and proportion of endemic and threatened amphibian and reptile species (a). Sankey plot showing the distribution of threats according to taxonomic group, endemism, and threatened categories (b).
Figure 5.
Comparison of Species Research Effort Allocation (SREA) between the ecological status of amphibians and reptiles. Broken lines indicate mean SREA for birds and mammals.
Figure 5.
Comparison of Species Research Effort Allocation (SREA) between the ecological status of amphibians and reptiles. Broken lines indicate mean SREA for birds and mammals.
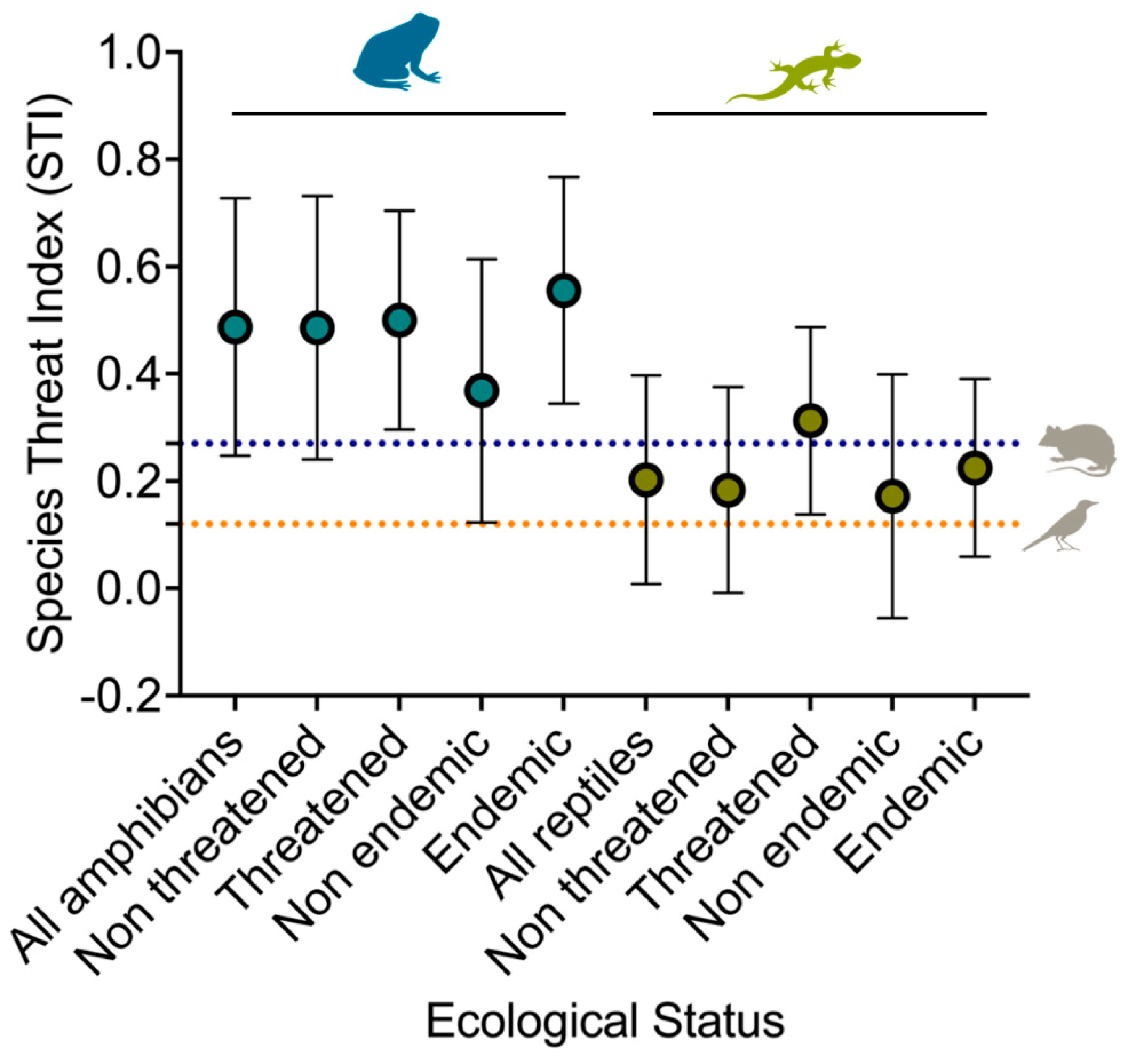
Figure 6.
Visualized predicted potential distribution of amphibians (a) and reptiles (b) in the faunal region of Mindanao based on the Maxent model. The dots represent the presence-only distribution of species recorded in the literature. Distribution of amphibians and reptiles across key biodiversity sites in Mindanao faunal region (c).
Figure 6.
Visualized predicted potential distribution of amphibians (a) and reptiles (b) in the faunal region of Mindanao based on the Maxent model. The dots represent the presence-only distribution of species recorded in the literature. Distribution of amphibians and reptiles across key biodiversity sites in Mindanao faunal region (c).
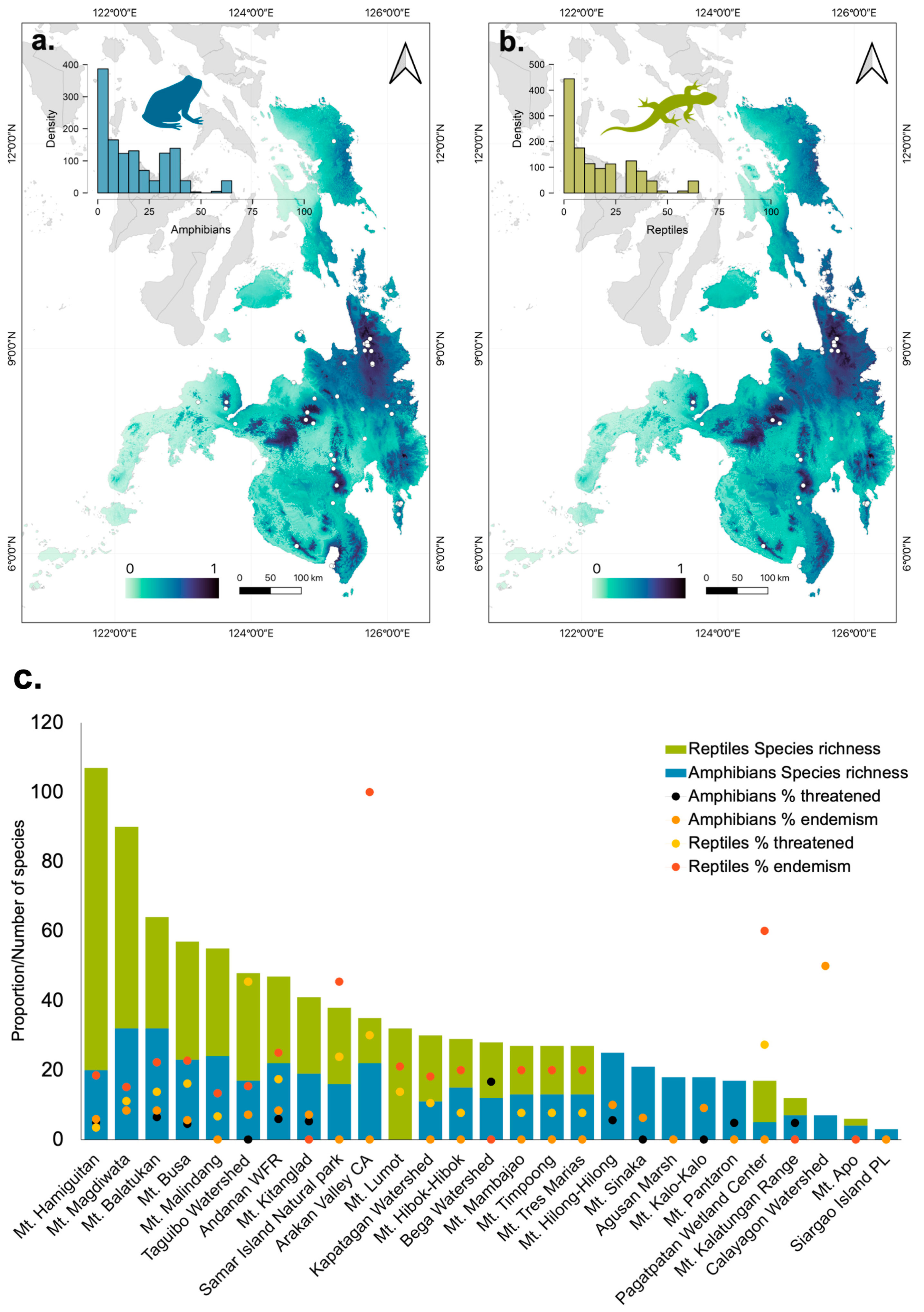

Table 1.
Results of generalized linear mixed modeling (GLMM) predicting the correlation of species ecological status and traits and threats to research efforts toward herpetofauna species.
Table 1.
Results of generalized linear mixed modeling (GLMM) predicting the correlation of species ecological status and traits and threats to research efforts toward herpetofauna species.
| (A) All herpetofauna | ||||
| Variables | β | SE | z | p |
| (Intercept) | 2.39 | 0.203 | 11.758 | < .001 |
| Threatened - Non threatened | -0.646 | 0.077 | -8.42 | < .001 |
| Philippine endemic - Non-endemic | 0.417 | 0.048 | 8.69 | < .001 |
| Body size (log10) | 0.273 | 0.064 | 4.256 | < .001 |
| Species Threat Index (SPI) | -0.02 | 0.116 | -0.172 | 0.863 |
| (B) Amphibians | ||||
| (Intercept) | 1.961 | 0.35 | 5.604 | < .001 |
| Threatened - Non threatened | -1.424 | 0.16 | -8.916 | < .001 |
| Philippine endemic - Non-endemic | 0.858 | 0.081 | 10.614 | < .001 |
| Body size (log10) | 0.544 | 0.152 | 3.591 | < .001 |
| Species Threat Index (SPI) | -0.355 | 0.158 | -2.243 | 0.025 |
| (C) Reptiles | ||||
| (Intercept) | 2.427 | 0.252 | 9.641 | < .001 |
| Threatened - Non threatened | -0.351 | 0.092 | -3.811 | < .001 |
| Philippine endemic - Non-endemic | 0.173 | 0.061 | 2.821 | 0.005 |
| Body size (log10) | 0.293 | 0.07 | 4.2 | < .001 |
| Species Threat Index (SPI) | 0.199 | 0.179 | 1.109 | 0.267 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



