
Submitted:
05 June 2023
Posted:
05 June 2023
You are already at the latest version
Abstract
The orchid genus Brachystele Schltr. comprises 20 species distributed from Mexico to Argentina; 10 are found in Brazil. Anatomical studies of Orchidoideae Lindl. have been scarce, and the anatomy and histochemistry of Brachystele are still largely unknown. We characterized the vegetative organs of B. guayanensis (Lindl.) Schltr. using standard anatomical and histochemical microtechniques. Brachystele guayanensis was observed to display the anatomical characters commonly found in the vegetative organs of representatives of the Orchidaceae and Orchidoideae (including a uniseriate epidermis, thin cuticle, amphistomatic leaves, anomocytic, diacytic and tetracytic stomata, a homogeneous mesophyll, collateral vascular bundles, rhizomes with pericyclic fibers, roots with velamen, uniseriate exodermis, endodermis and pericycle). Histochemical tests confirmed the presence of lignin, proteins, and alkaloids, the lipidic nature of the cuticle, starch grains stored in spiranthosomes in the roots, and the composition of the raphides. Alkaloids were observed in great abundance, especially in the roots, and may have potentially useful medicinal activities, as has been observed in groups phylogenetically related to Brachystele.
Keywords:
Alkaloids
; Cells
; Cranichideae
; Ergastic substances
; Leaf
; Micromorphology
; Orchidoideae
; Rhizomes
; Roots
; Spiranthinae
1. Introduction
Brachystele Schltr. comprises 20 species distributed from southern Mexico to northern Argentina [1]; 10 species occur in Brazil, of which three are endemic [2]. The taxon is terrestrial, herbaceous, with short rhizomes, fleshy roots, leaves arranged in rosettes, lateral racemes with small flowers, resupinate, usually indumented, with a bilobed stigma [1,2].
Anatomical descriptions of the vegetative organs of Orchidoideae Lindl., the subfamily in which Brachystele is included, are still incipient and have mainly focused on species not occurring in Brazil [e.g., 3–20]. Anatomical studies of Brachystele are currently limited to the work of Bernal et al. [18], who examined the root trichomes of representatives of Spiranthinae Lindl. ex Meisn. as well as B. widgrenii (Rchb.f.) Schltr. There have been no histochemical studies of any Brachystele species, although some medicinal properties of B. dilatata (Lindl.) Schltr. and B. unilateralis (Poir.) Schltr. were mentioned (e.g., their diuretic and carminative property) in ethnobotanical studies [21,22].
We provide here anatomical and histochemical characterizations of the vegetative organs of B. guayanensis (Lindl.) Schltr. species (Figure 1) to expand available knowledge of that diversity in Orchidaceae, especially in Brachystele, and subsidize phylogenetic and micromorphological studies of that genus.
2. Results and Discussion
2.1. Anatomical data
2.1.1. Leaf anatomy
In frontal view, the leaf epidermis of B. guayanensis is covered by a striated cuticle and is composed of polygonal cells with both straight and curved walls (Figure 2a, b) – characters that have also been reported for Microchilus arietinus (Rchb. f. & Warm.) Ormerod and Zeuxine strateumatica (L.) Schltr.) by Andreota et al. [17] and Bona et al. [20] and described for Aa paleacea (Kunth) Rchb. f. and Pterichis multiflora (Lindl.) Schltr. by Corredor and Arias [15]. Anomocytic, diacytic and tetracytic stomata were observed on both faces of the leaf blade of B. guayanensis (Figure 2a, b), as seen in other species of Orchidaceae [3,14,16,19].
In cross-section, the epidermis of B. guayanensis is covered by a thin, striated cuticle and is uniseriate and composed of rounded, oblong cells with slightly thickened external periclinal walls (Figure 2c, d) – aspects reported for other members of Orchidoideae [e.g., 6,9,10,13–15,17,20]. The leaves are amphistomatic (Figure 2c-e), with stomata at the same level as common cells of the epidermis. The substomatal chambers are wider than the suprastomatal chambers (Figure 2c, d) – characteristics related to reducing water losses and evapotranspiration [23,24]. Amphistomatic leaves are commonly observed in plants having both high photosynthetic capacities and high stomatal conductance, especially those that grow in open and sunny environments [25,26], as studied here.
The mesophyll of B. guayanensis is homogeneous and consists of 6–11 layers of rounded cells having varying dimensions (Figure 2e). Those cells are interspersed with crystalliferous idioblasts containing raphides (Figure 2f) – a pattern cited for different groups of Orchidaceae [e.g., 6–10,13–17,19,20,27–29]. Collateral vascular bundles surrounded by a parenchyma sheath were observed in the median portion of the mesophyll (Figure 2e). The bundle corresponding to the midrib is flat-convex (Figure 2g) and has the largest caliber; the elements of that vessel are arranged in a “V” (Figure 2h); smaller bundles (Figure 2i), with smaller calibers are interspersed. This same pattern has been reported in other taxa by the authors cited above. The leaf margins of B. guayanensis are straight and rounded (Figure 2j).
2.1.2. Rhizome anatomy
The anatomy of the rhizomes of B. guayanensis was observed to be similar to other representatives of Orchidoideae [e.g., 6,9,10,13–16,19] in having rounded outlines and variable calibers (Figure 3a, b), with cataphylls in nodal regions or protecting the buds (Figure 3b, c). The epidermis is covered by a smooth, thin cuticle, is uniseriate, and composed of oblong, or occasionally, rounded common cells with thin cell walls (Figure 3d). Stomata were only observed on exposed portions of the rhizome and were arranged at the same level as the other common cells, with only tiny substomatal and suprastomatal chambers (Figure 3d).
The cortex of B. guayanensis is surrounded by a ring of pericyclic fibers (Figure 3h) and consists of 16–22 layers of rounded parenchyma cells of varying sizes (Figure 3a, b) with small triangular intercellular spaces; some of the parenchyma cells contain raphides (Figure 3e) and starch grains (Figure 3f). The arrangement is similar to that described by Stern [16] and by Stern and Judd [27,28] for the aerial stems of other Orchidaceae groups (such as Vanilloideae [27,28]; Orchidoideae, tribes Diseae Dressler and Ochideae Small; and Epidendroideae Lindl., tribes Sobralieae Pfitzer and Triphoreae Dressler [16,29]), as well as other groups of Monocotyledons [e.g., 30]. The endodermis and/or pericycle, as described here (Figure 3b), may participate in the genesis of adventitious roots, and therefore exhibit meristematic activity [30].
Fungal hyphae and pelotons were observed to be concentrated mainly in the more peripheral portions of the cortical parenchyma (Figure 3g), as was reported by Pereira et al. [31] for the roots of Bulbophyllum sp., Campylocentrum organense (Rchb.f.) Rolfe and Gomesa crispa (Lindl.) Klotzsch ex Rchb. f. According to these authors, the strategic positions of those structures contribute to the maintenance and (re)colonization of internal tissues and thus constitute important sources of inoculum for adventitious roots that will extend from the rhizome, corroborating the findings of Bougoure et al. [32] for the terrestrial orchids Rhizanthella gardneri R.S. Rogers and Suetsugu et al. [33] and Oreorchis indica (Lindl.) Hook. f.
From 28 to 45 collateral vascular bundles can be observed internally, surrounded by a parenchyma sheath, in an atactostelic arrangement (Figure 3i) typical of Monocots [25,34,35]. Those bundles are observed in the central portion of the organ analyzed (Figure 3a, b, i), and have been observed in other groups of that family [e.g., 9,10,16,27–29]. The vessel elements of the xylem are arranged in “V” formations in the collateral vascular bundles (Figure 3j), consistent with the findings of Andreota [36] for the tribe Cranichideae.
2.1.3. Root anatomy
Brachystele guayanensis has transversely circular roots of variable calibers (Figure 4a) that are anatomically similar to those found in other taxa of Orchidoideae [e.g., 5,6,11,12,15–17], with uniseriate velamen of the “Spiranthes type”. The component cells are irregular, elliptic, and thin, with thin cell walls with helicoidal thickenings and small pores in the non-thickened regions (Figure 4b, c). Our observations were similar to those of Porembski and Barthlott [4] for Pelexia dolichorhiza Schltr. (= Pachygenium pteryganthum (Rchb. f. & Warm.) Szlach., R. González & Rutk.), Prescottia colorans Lindl. (= P. stachyoides (Sw.) Lindl.), and Sauroglossum elatum Lindl. Moreira and Isaias [12] reported that terrestrial orchids generally have thinner velamen than epiphytic or rupicolous species in terms of their numbers of layers. Pridgeon [37], Moreira and Isaias [12], and Chomicki et al. [38] noted that velamen protects the roots from high solar radiation levels and excessive evapotranspiration losses, helps prevent overheating, and provides mechanical support to internal tissues, among other functions.
Simple, unicellular trichomes, such as those observed here (Figure 4c), were reported for Brachystele widgrenii, Lankesterella caespitosa (Lindl.) Hoehne, L. ceracifolia (Barb. Rodr.) Mansf., Pelexia orthosepala (Rchb.f. & Warm.) Schltr, and Sacoila lanceolata (Aubl.) Garay. by Bernal et al. [18], for Cranichis candida (Barb.Rodr.) Cogn. by Andreota et al. [17], and for 11 species of Aspidogyne Garay and Microchilus C. Presl. by Bona et al. [20]. According to Stern et al. [6], Andreota et al. [17] and Bernal et al. [18], simple trichomes provide better attachment to the substrate and increase contact with that surface – thus facilitating the absorption of both water and mineral salts.
The root cortex is composed of 18–20 layers of rounded parenchyma cells of varying sizes with innumerable small, triangular intercellular spaces (Figure 4d); hyphae and pelotons are also observed, mainly concentrated in more peripheral portions and underlying the exodermis (Figure 4d, g, h). This pattern has been reported in the roots of other Orchidaceae taxa by Pereira et al. [31], who noted that the presence of such symbiotic microorganisms is of great importance to the germination of orchid seeds as they increase the surface area of the roots and thus facilitate water and nutrient absorption [31,39].
These characters are frequently observed in Orchidoideae [e.g., 11,15,16], with terrestrial members of the family (as was observed in the species studied here) generally having thick, fleshy roots with a more expressive cortex (in terms the numbers of layers) that is responsible for holding reserves of water, starch grains, and other nutrients, with a less expressive velamen. Some of the cells in the cortical parenchyma contain raphides (Figure 4e), as observed in different groups of Monocotyledons [e.g., 34], with polyhedral starch grains gathered in spiranthosomes (Figure 4f) – spherical specialized amyloplasts found in Cranichideae and interpreted as a synapomorphy of the tribe [See 5,17,40,41].
The exodermis (Figure 4b, c), endodermis and pericycle (Figure 4k) of B. guayanensis are uniseriate, and are composed of rounded, elliptic cells with thin walls; the cells of the exodermis are slightly thickened (Figure 4b, c), and endodermis cells have evident Casparian strips (Figure 4k). Those patterns are repeated in other terrestrial orchids, especially in the subfamily Orchidoideae [See 13,15,17,19,20]. Benzing et al. [42] and Sanford and Adanlawo [43] noted that the thickening of the exodermal cells, as well as presence of a Casparian strip, aid in minimizing water losses to the external environment and provide mechanical support to root tissues.
The vascular cylinders in the roots of B. guayanensis are of variable calibers, have circular or elliptic shapes, and 12–14 protoxylem poles (Figure 4i, j), categorizing those roots as polyarchs, which are common in Monocotyledons [25,35]. The xylem and phloem are interspersed (Figure 4i-k); the central portion of the vascular cylinder is composed of parenchyma cells of varying sizes and shapes with tiny triangular intercellular spaces (Figure 4i, j), similar to those observed in other groups of Orchidaceae [e.g., 11,13,15–17,19,20].
2.2. Histochemistry data
Coomassie blue and Xylidine Ponceau strongly stained the pelotons and hyphae, indicating their protein contents (Figure 5a, b), and Sudan IV staining confirm the lipidic nature of the waxy cuticle (Figure 5c); the raphides are composed of calcium oxalate. Lignin was identified in the walls of xylem vessel elements (Figure 5d, f, g) and pericyclic fibers (Figure 5e), as well as in Casparian strips (Figure 5g). Starch grains and alkaloids were identified within the stomata guard cells (Figure 5h, o), epidermal cells, mesophyll cells (especially in the vicinity of vascular bundles) (Figure 5i, j, p), the cortical parenchyma of rhizomes (and occasionally in the central portion) (Figure 5k, l, q, r), as well as in roots (where they were most abundant) (Figure 5m, n, s-u). The histochemical tests were negative for reducing sugars, phenolic compounds, and tannins.
The waxy cuticle performs several functions, including protection against solar radiation, overheating, and water losses from internal tissues; it confers protection against the entry and the attacks of pathogens and herbivores [44,45,46].
Calcium oxalate raphides that are found in different groups of Orchidaceae, as well as other Angiosperms [e.g., 16,47], serve as defenses against herbivores, act in osmoregulation and in other metabolic activities requiring calcium, and have a role in the detoxification of aluminum (an element very abundant savannas soils) [47,48].
The lignin commonly deposited in vessel elements, pericyclic fibers, and the Casparian strip of the vegetative organs of the studied species confer stability, rigidity and mechanical support to the cell walls and internal tissues (25, 35). The Casparian strip, together with suberin deposits (both hydrophobic substances), contribute to solute selectivity in the root endodermis and act as barriers against apoplast movement – therefore preventing the influx of ions from the vascular cylinder to the cortical region [49,50,51].
Starch grains, especially those found in the roots and rhizomes, constitute some of the principal plant reserves are used (among other purposes) for vegetative growth and propagation [52,53,54,55]. Starch grains inside the stomata guard cells of the species studied here have also been reported in Oxalis L. (Oxalidaceae), Sambucus australis Cham. & Schltdl. (Adoxaceae), and Socratea exorrhiza (Mart.) H. Wendl. (Arecaceae) by Nunes et al. [56], Kikuchi et al. [57] and Reis and Alvim [58]. According to Appezzato-da-Glória and Carmello-Guerreiro [35], potassium levels in those structures appear to be associated with starch hydrolysis, which provides those organic anions. There is also evidence that malate, the regulator responsible for guard cell movements, can be synthesized through starch degradation [59,60].
According to Bulpitt et al. [61] and Sut et al. [62], many of the secondary metabolic substances identified in representatives of the Orchidaceae (such as flavonoids, alkaloids, terpenes, glycosides) act in plant defenses against herbivores and pathogens, and show bioactive pharmacological effects [63,64]. We believe that the presence of alkaloids in the stomata guard cells and in the cortical parenchyma of the rhizomes and roots together with the raphides identified in B. guayanensis, act to defend the species against attacks by pathogens and herbivores, as mentioned by Franceschi and Nakata [47] and Vizzotto et al. [65] for plants in general. Similar to what has been postulated by Li et al. [66] for the vegetative organs of other orchids, the abundant presence of raphides and secondary metabolites (e.g., alkaloids) in B. guayanensis, especially in the reserve organs of that species (e.g., the roots and rhizomes), are responsible for the renewal of their aerial portions, and help prevent pathogens and herbivores from reaching the vascular system (which is usually found in more internal regions) and causing local and/or systemic damage.
Among the bioactive substances found in Orchidaceae species are alkaloids (e.g., dendrobin, nobilonin, dendroxin) and nitrogenous heterocyclic organic molecules derived from the secondary metabolism of amino acids (e.g., phenylalanine, lysine, arginine, tyrosine, tryptophan). More than 100 types of bioactive alkaloids have been identified in more than 2,000 orchid species [67,68]. Those alkaloids have been found to be effective for treating gastrointestinal disorders and cardiovascular diseases, and also evidence anti-inflammatory, diuretic, analgesic, antioxidant, immunomodulatory, antipyretic, and antitumor activities [69,70,71]. Within this context, the alkaloids found in B. guayanensis indicate it as a potential medicinal species. (Bio)phytochemical investigations could therefore be informative, with the subsequent isolation and testing of the toxicity of any bioactive substances, and the characterization of their chemical natures.
This study was designed to provide initial information concerning the anatomy and histochemistry of the vegetative organs of the genus Brachystele and its medicinal potential and indicates the importance of anatomical and histochemical studies focusing on neotropical Orchidaceae species, especially those that have been only poorly studied.
3. Materials and Methods
For the anatomical studies, samples of vegetative organs (e.g., the mid-portions of the leaf blades, rhizomes, and roots) of five adult individuals of B. guayanensis (Figure 1) were collected in open fields, close to Bosque Auguste Saint-Hilaire, at Campus II (Samambaia) of the Federal University of Goiás (UFG), Goiânia, GO, Brazil. Collections of botanical material followed the recommendations of Mori et al. [72], and voucher specimens were deposited in the UFG Herbarium (registration numbers: I.S. Santos 1160 and 1161).
The samples collected for anatomical studies were fixed in 70% FAA (glacial acetic acid, formaldehyde, and 70% ethyl alcohol, 1:1:18) in hermetically sealed containers for 48 hours; after that time, they were preserved in 70% ethyl alcohol [73]. For the anatomical descriptions, cross sections were cut using a razor blade and clarified in a 20% aqueous solution of sodium hypochlorite (NaClO) (v/v) [74], stained with Astra Blue and Safranin (9:1) [75], and mounted in aqueous Glycerol solution (1:1). For analysis of the leaf surface in frontal view, the epidermis was dissociated using the Jeffrey method [73]. For the procedures mentioned above, the slides were sealed with a colorless sealant and subsequently photomicrographed using a Leica ICC50 HD® digital camera coupled to a Leica DM500® microscope, using Motic 2.0 Image Plus Software.
For histochemical studies, in natura samples obtained the time of collection were stained with the following reagents: Coomassie Blue and Xylidine Ponceau [76,77] for detecting proteins; Ferric chloride for detecting phenolic compounds; acidified Phloroglucinol for lignin; Sudan IV [73] for total lipids; Dittmar [78] for alkaloids: Fehling [74] for reducing sugars, Lugol [79] for starch; and hydrochloric vanillin [80] to detect tannins. To verify the chemical constitution of the crystals, 10% hydrochloric acid was used, following Chamberlain [81]. The descriptions of the analyzed organs were based on the terminologies used in the specialized literature [15,17,18,20].
Author Contributions
I.S.S. collected, processed, and identified the botanical material; M.J.S. supervised all laboratory work, and the analysis of data obtained by I.S.S.; M.J.S. guided the elaboration of the manuscript and provided a critical analysis of the data obtained; I.S.S and M.J.S. participated in the interpretation of the data, writing, conceptualization, theoretical basis, review and editing of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors would like to thank the Federal University of Goiás (UFG) and the São Paulo State University (Unesp) for the use of their facilities and logistical support, and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the PhD and Productivity scholarships in Research provided to authors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rutkowski, P.; Mytnik, J.; Szlachetko, D.L. New taxa and new combinations in Mesoamerican Spiranthinae (Orchidaceae, Spirantheae). Ann. Bot. Fenn. 2004, 41, 471–477. [Google Scholar] [CrossRef]
- Barros, F.; Vinhos, F.; Rodrigues, V.T.; Barberena, F.F.V.A.; Fraga, C.N.; Pessoa, E.M.; Forster, W.; Menini-Neto, L.; Furtado, S.G.; Nardy, C.; Azevedo, C.O.; Guimarães, L.R.S.; Orchidaceae. In Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. 2015. Available online: http://servicos.jbrj.gov.br/flora/search/Brachystele_guayanensis (accessed on 18-X-2021).
- Rasmussen, H. The diversity of stomatal development in Orchidaceae subfamily Orchidoideae. Bot. J. Linn. Soc. 1981, 82, 381–393. [Google Scholar] [CrossRef]
- Porembski, S.; Barthlott, W. Velamen radicum micromorphology and classification of Orchidaceae. Nord. J. Bot. 1988, 8, 117–137. [Google Scholar] [CrossRef]
- Stern, W.L.; Aldrich, H.C.; McDowell, L.M.; Morris, M.W.; Pridgeon, A.M. Amyloplasts from cortical root cells of Spiranthoideae (Orchidaceae). Protoplasma 1993a, 172, 49–55. [Google Scholar] [CrossRef]
- Stern, W.L.; Morris, M.W.; Judd, W.S.; Pridgeon, A.M.; Dressler, R.L. Comparative vegetative anatomy and systematics of Spiranthoideae (Orchidaceae). Bot. J. Linn. Soc. 1993b, 113, 161–197. [Google Scholar] [CrossRef]
- Pridgeon, A.M. Systematic leaf anatomy of Caladeniinae (Orchidaceae). Bot. J. Linn. Soc. 1994, 114, 31–48. [Google Scholar] [CrossRef]
- Kurzweil, H.; Linder, H.P.; Stern, W.L.; Pridgeon, A.M. Comparative vegetative anatomy and classification of Diseae (Orchidaceae). Bot. J. Linn. Soc. 1995, 117, 171–220. [Google Scholar] [CrossRef]
- Stern, W.L. Vegetative anatomy of subtribe Orchidinae (Orchidaceae). Bot. J. Linn. Soc. 1997a, 124, 121–136. [Google Scholar] [CrossRef]
- Stern, W.L. Vegetative anatomy of subtribe Habenariinae (Orchidaceae). Bot. J. Linn. Soc. 1997b, 125: 211–227. [CrossRef]
- Figueroa, C.; Salazar, G.A.; Zavaleta, H.A.; Engleman, E.M. Root character evolution and systematics in Cranichidinae, Prescottiinae and Spiranthinae (Orchidaceae, Cranichideae). Ann. Bot. 2008, 101, 509–520. [Google Scholar] [CrossRef]
- Moreira, A.S.F.P.; Isaias, R.M.S. Comparative anatomy of the absorption roots of terrestrial and epiphytic orchids. Braz. Arch. Biol. Technol. 2008, 51, 83–93. [Google Scholar] [CrossRef]
- Aybeke, M.; Sezik, E.; Olgun, G. Vegetative anatomy of some Ophrys, Orchis and Dactylorhiza (Orchidaceae) taxa in Trakya region of Turkey. Flora 2010, 205, 73–89. [Google Scholar] [CrossRef]
- Aybeke, M. Comparative anatomy of selected rhizomatous and tuberous taxa of subfamilies Orchidoideae and Epidendroideae (Orchidaceae) as an aid to identification. Plant Syst. Evol. 2012, 298, 1643–1658. [Google Scholar] [CrossRef]
- Corredor, B.A.D.; Arias, R.L. Morfoanatomía en Cranichideae (Orchidaceae) de la Estación Loma Redonda del Parque Nacional “Sierra Nevada”, Mérida, Venezuela. Lankesteriana 2012, 12, 61–75. [Google Scholar] [CrossRef]
- Stern, W.L. Anatomy of the Monocotyledons: X. Orchidaceae., 1st ed.; Oxford University Press: Oxford, England, 2014; pp. 1–288. [Google Scholar]
- Andreota, R.C.; Barros, F.; Sajo, M.G. Root and leaf anatomy of some terrestrial representatives of the Cranichideae tribe (Orchidaceae). Rev. Bras. Bot. 2015, 38, 367–378. [Google Scholar] [CrossRef]
- Bernal, A.A.; Smidt, E.C.; Bona, C. Spiral root hairs in Spiranthinae (Cranichideae: Orchidaceae). Rev. Bras. Bot. 2015, 38, 411–415. [Google Scholar] [CrossRef]
- Şenel, G.; Akbulut, M.K.; Şeker, Ş.S. Comparative anatomical properties of some Epidendroideae and Orchidoideae species distributed in NE Turkey. Protoplasma 2019, 256, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Bona, C.; Engels, M.E.; Pieczak, F.S.; Smidt, E.C. Comparative vegetative anatomy of Neotropical Goodyerinae Klotzsch (Orchidaceae Juss.: Orchidoideae Lindl.). Acta Bot. Bras. 2020, 34, 530–539. [Google Scholar] [CrossRef]
- Neumann, C. Orchideen als Arzneipflanzen: Ein Querschnitt durch ausgewählte medizinische und botanisch-pharmazeutische Literatur des 19. Jahrhunderts. Z. fur Phytother. 2009, 30, 1–29. [Google Scholar] [CrossRef]
- Verettoni, H.N. Contribución al conocimiesão bioativas de las plantas medicinales de la región de Bahía Blanca., 1st ed.; Harris y Cia: Bahía Blanca, Argentina, 1985; pp. 1–374. [Google Scholar]
- Oliveira, V.C.; Sajo, M.G. Leaf anatomy of epiphyte species of Orchidaceae. Rev. Bras. Bot. 1999, 22, 365–374. [Google Scholar] [CrossRef]
- Silva, C.I.; Milaneze-Gutierre, M.A. Caracterização morfoanatômica dos órgãos vegetativos de Cattleya walkeriana Gardner (Orchidaceae). Acta Sci. 2004, 26, 91–100. [Google Scholar] [CrossRef]
- Fahn, A. Plant anatomy., 4th ed.; Pergamon Press: Oxford, England, 1990; pp. 1–530. [Google Scholar]
- Smith, W.K.; Vogelmann, T.C.; DeLucia, E.H.; Bell, D.T.; Shepherd, K.A. Leaf form and photosynthesis. Bioscience 1997, 47, 785–793. [Google Scholar] [CrossRef]
- Stern, W.L.; Judd, W.S. Comparative vegetative anatomy and systematics of Vanilla (Orchidaceae). Bot. J. Linn. Soc. 1999, 131, 353–382. [Google Scholar] [CrossRef]
- Stern, W.L.; Judd, W.S. Comparative anatomy and systematics of the orchid tribe Vanilleae excluding Vanilla. Bot. J. Linn. Soc. 2000, 134, 179–202. [Google Scholar] [CrossRef]
- Carlsward, B.S.; Stern, W.L. Vegetative anatomy and systematics of Triphorinae (Orchidaceae). Bot. J. Linn. Soc. 2009, 159, 203–210. [Google Scholar] [CrossRef]
- Menezes, N.L.; Elbl, P.M.; Cury, G.; Appezzato-da-Glória, B.; Sasaki, K.L.; Silva, C.G.; Costa, G.R.; Lima, V.G. The meristematic activity of the endodermis and the pericycle and its role in the primary thickening of stems in monocotyledonous plants. Plant Ecol. Divers. 2012, 5, 153–165. [Google Scholar] [CrossRef]
- Pereira, O.L.; Kasuya, M.C.M.; Rollemberg, C.L.; Chaer, G.M. Isolamento e identificação de fungos micorrízicos rizoctonióides associados a três espécies de orquídeas epífitas neotropicais no Brasil. Rev. Bras. Cienc. Solo 2005, 29, 191–197. [Google Scholar] [CrossRef]
- Bougoure, J.; Ludwig, M.; Brundrett, M.; Grierson, P. Identity and specificity of the fungi forming mycorrhizas with the rare mycoheterotrophic orchid Rhizanthella gardneri. Mycol. Res. 2009, 113, 1097–1106. [Google Scholar] [CrossRef]
- Suetsugu, K.; Haraguchi, T.F.; Tanabe, A.S.; Tayasu, I. Specialized mycorrhizal association between a partially mycoheterotrophic orchid Oreorchis indica and a Tomentella taxon. Mycorrhiza 2021, 31, 243–250. [Google Scholar] [CrossRef]
- Metcalfe, C.R. Comparative anatomy as a modern Botanical discipline. In Advances in botanical research, 1st ed.; Metcalfe, C.R., Ed.; Academic Press: New York, United States, 1963; Volume VI, pp. 101–147. [Google Scholar]
- Appezzato-da-Glória, B.; Carmello-Guerreiro, S.M. Anatomia Vegetal., 2rd ed.; Editora UFV: Viçosa, Bahia, Brazil, 2006; pp. 1–438. [Google Scholar]
- Andreota, R.C. Anatomia dos órgãos vegetativos de representantes da tribo Cranichideae (Orchidoideae: Orchidaceae). Masters’ thesis, Universidade Estadual Paulista, Rio Claro, São Paulo, Brazil, 2013.
- Pridgeon, A.M. The velamen and exodermis of orchid roots. In Orchid biology, reviews and perspectives, 1st ed.; Arditti, J., Ed.; Cornell University Press: Ithaca, New York, United States, 1987; Volume IV, pp. 139–192. [Google Scholar]
- Chomicki, G.; Bidel, L.P.R.; Ming, F.; Coiro, M.; Zhang, X.; Wang, Y.; Baissac, Y.; Jay-Allemand, C.; Renner, S.S. The velamen protects photosynthetic orchid roots against UV-B damage, and a large dated phylogeny implies multiple gains and losses of this function during the Cenozoic. New Phytol. 2015, 205, 1330–1341. [Google Scholar] [CrossRef]
- Cameron, D.D.; Johnson, I.; Leake, J.R.; Read, D.J. Mycorrhizal acquisition of inorganic phosphorus by the green-leaved terrestrial orchid Goodyera repens. Ann. Bot. 2007, 99, 831–834. [Google Scholar] [CrossRef]
- Salazar, G.A.; Chase, M.W.; Arenas, M.S.; Ingrouille, M. Phylogenetics of Cranichideae with emphasis on Spiranthinae (Orchidaceae, Orchidoideae): evidence from plastid and nuclear DNA sequences. Am. J. Bot. 2003, 90, 777–795. [Google Scholar] [CrossRef] [PubMed]
- Şeker, Ş.S. What does the quantitative morphological diversity of starch grains in terrestrial orchids indicate? Microsc. Res. Tech. 2022, 85, 2931–2942. [Google Scholar] [CrossRef] [PubMed]
- Benzing, D.H.; Friedman, W.E.; Peterson, G.; Renfrow, A. Shootlessness, velamentous roots, and the pre-eminence of Orchidaceae in the epiphytic biotype. Am. J. Bot. 1983, 70, 121–33. [Google Scholar] [CrossRef] [PubMed]
- Sanford, W.W.; Adanlawo, I. Velamen and exodermis characters of west african epiphytic orchids in relation to taxonomic grouping and habitat tolerance. Bot. J. Linn. Soc. 1973, 66, 307–321. [Google Scholar] [CrossRef]
- Juniper, B.E.; Jeffree, C.E. Plant surfaces., 1st ed.; Edward Arnold Publishers: London, England, 1983; pp. 1–93. [Google Scholar]
- Dickison, W.C. Integrative plant anatomy., 1st ed.; Academic Press: San Diego, United States, 2000; pp. 1–533. [Google Scholar]
- Evert, R.F. Esau’s plant anatomy: meristems, cells and tissues of the plant body: their structure, function and development., 3rd ed.; John Wiley & Sons, Inc.: New Jersey, United States, 2006; pp. 1–601. [Google Scholar]
- Franceschi, V.R.; Nakata, P.A. Calcium oxalate in plants: formation and function. Annu. Rev. Plant Biol. 2005, 56, 41–71. [Google Scholar] [CrossRef] [PubMed]
- Haridasan, M. Alumínio é um elemento tóxico para as plantas nativas do cerrado. In Fisiologia Vegetal: práticas em relações hídricas, fotossíntese e nutrição mineral, 1st ed.; Prado, C.H.B.A., Casali, C.A., Eds.; Editora Manole: Barueri, São Paulo, Brazil, 2006; Volume 1, pp. 1–10. [Google Scholar]
- Peterson, C.A.; Murrmann, M.; Steudle, E. Location of the major barriers to water and ion movement in young roots of Zea mays L. Planta 1993, 190, 127–136. [Google Scholar] [CrossRef]
- Enstone, D.E.; Peterson, C.A.; Ma, F. Root endodermis and exodermis: structure, function, and responses to the environment. J. Plant Growth Regul. 2003, 21, 335–351. [Google Scholar] [CrossRef]
- Lux, A.; Luxová, M. Growth and differentiation of root endodermis in Primula acaulis Jacq. Biol. Plant. 2003, 47, 91–97. [Google Scholar] [CrossRef]
- Holttum, R. Growth habitats of monocotyledons - variations on a theme. Phytomorphology 1955, 5, 399–413. [Google Scholar]
- Beck, E.; Ziegler, P. Biosynthesis and degradation of starch in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 95–117. [Google Scholar] [CrossRef]
- Oliveira, F.; Akisue, G. Fundamentos de farmacobotânica., 1st ed.; Atheneu: São Paulo, Brazil, 1989; pp. 1–216. [Google Scholar]
- Salisbury, F.B.; Ross, C.W. Plant physiology., 4th ed.; Wadsworth: Belmont, United States, 1992; pp. 1–267. [Google Scholar]
- Nunes, E.; Scopel, M.; Vignoli-Silva, M.; Vendruscolo, G.S.; Henriques, A.T.; Mentz, L.A. Caracterização farmacobotânica das espécies de Sambucus (Caprifoliaceae) utilizadas como medicinais no Brasil. Parte II. Sambucus australis Cham. & Schltdl. Braz. J. Pharmacog. 2007, 17, 414–425. [Google Scholar] [CrossRef]
- Kikuchi, T.Y.S.; Braga, Z.V.; Potiguara, R.C.V. Anatomia foliar de Socratea exorrhiza (Mart.) H. Wendl. (Arecaceae). Biota Amazôn. 2016, 6, 73–79. [Google Scholar] [CrossRef]
- Reis, R.E.; Alvim, M.N. Anatomia foliar comparada de três espécies do gênero Oxalis L. (Oxalidaceae). NBC 2013, 3, 59–72. [Google Scholar] [CrossRef]
- Vavasseur, A.; Raghavendra, A.S. Guard cell metabolism and CO2 sensing. New Phytol. 2005, 165, 665–682. [Google Scholar] [CrossRef] [PubMed]
- Araújo, W.L.; Nunes-Nesi, A.; Osorio, S.; Usadel, B.; Fuentes, D.; Nagy, R.; Balbo, I.; Lehmann, M.; Studart-Witkowski, C.; Tohge, T.; Martinoia, E.; Jordana, X.; DaMatta, F.M.; Fernie, A.R. Antisense inhibition of the iron-sulphur subunit of succinate dehydrogenase enhances photosynthesis and growth in tomato via an organic acid mediated effect on stomatal aperture. Plant Cell. 2011, 23, 600–627. [Google Scholar] [CrossRef] [PubMed]
- Bulpitt, C.J.; Li, Y.; Bulpitt, P.F.; Wang, J. The use of orchids in Chinese medicine. J. R. Soc. Med. 2007, 100, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Sut, S.; Maggi, F.; Dall'Acqua, S. Bioactive secondary metabolites from orchids (Orchidaceae). Chem. Biodivers. 2017, 14, e1700172. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, T. Chemical ecology of pyrrolizidine alkaloids. Planta 1999, 207, 483–495. [Google Scholar] [CrossRef]
- Sepúlveda-Jiménez, G.; Porta-Ducoing, H.; Rocha-Sosa, M. La participación de los metabolitos secundarios en la defensa de las plantas. Rev. Mex. Fitopatol. 2003, 21, 355–363. [Google Scholar]
- Vizzotto, M.; Krolow, A.C.R.; Weber, G.E.B. Metabólitos secundários encontrados em plantas e sua importância., 1st ed.; Embrapa Clima Temperado: Pelotas, Rio Grande do Sul, Brazil, 2010; pp. 1–17. [Google Scholar]
- Li, J.W.; Zhang, Z.B.; Zhang, S.B. Widely targeted metabolic, physical and anatomical analyses reveal diverse defensive strategies for pseudobulbs and succulent roots of orchids with industrial value. Ind. Crop. Prod. 2022, 177, 114510. [Google Scholar] [CrossRef]
- Gutierrez, R.M.P. Orchids: A review of uses in traditional medicine, its phytochemistry and pharmacology. J. Med. Plant Res. 2010, 4, 592–638. [Google Scholar] [CrossRef]
- Hossain, M.M. Therapeutic orchids: Traditional uses and recent advances—An overview. Fitoterapia 2011, 82, 102–140. [Google Scholar] [CrossRef] [PubMed]
- Brito, H.O.; Noronha, E.P.; França, L.M. Phytochemical analysis composition from Annona squamosa (ATA) ethanolic extract leaves. Rev. Bras. Farm. 2008, 89, 180–184. [Google Scholar]
- Ng, T.B.; Liu, J.; Wong, J.H.; Ye, X.; Sze, S.C.W.; Tong, Y.; Zhang, K.Y. Review of research on Dendrobium, a prized folk medicine. Appl. Microbiol. Biotechnol. 2012, 93, 1795–1803. [Google Scholar] [CrossRef]
- Li, R.; Liu, T.; Liu, M.; Chen, F.; Liu, S.; Yang, J. Anti-influenza a virus activity of dendrobine and its mechanism of action. J. Agric. Food Chem. 2017, 65, 3665–3674. [Google Scholar] [CrossRef]
- Mori, S.A.; Silva, L.A.; Lisboa, G.; Coradin, L. Manual de Manejo do Herbário Fanerogâmico. 2nd. ed.; CEPLAC: Ilhéus, Bahia, Brazil, 1989; pp. 1–104. [Google Scholar]
- Johansen, D.A. Plant microtechnique., 1st ed.; McGraw-Hill Book Company, Inc.: New York, United States, 1940; pp. 1–523. [Google Scholar]
- Kraus, J.E.; Arduin, M. Manual básico de métodos em morfologia vegetal., 1st ed.; EDUR: Rio de Janeiro, Brazil, 1997; pp. 1–198. [Google Scholar]
- Bukatsch, F. Bermerkungen zur Doppelfrbung Astrablau-Safranin. Mikrokosmos 1972, 61, 255. [Google Scholar]
- Fisher, D.B. Protein staining of ribboned epon sections for light microscopy. Histochemie 1968, 16, 92–96. [Google Scholar] [CrossRef]
- O’Brien, T.P.; McCully, M.E. The study of plant structure principles and selected methods., 1st ed.; Termarcarphi: Melbourne, Australia, 1981; pp. 1–344. [Google Scholar]
- Furr, M.; Mahlberg, P.G. Histochemical analyses of laticifers and glandular trichomes in Cannabis sativa. J. Nat. Prod. 1981, 44, 153–159. [Google Scholar] [CrossRef]
- Jensen, W.A. Botanical histochemistry: principles and practice., 1st ed.; W.H. Freeman: San Francisco, United States, 1962; pp. 1–408. [Google Scholar]
- Mace, M.E.; Howell, C.R. Histological and identification of condensed tannin precursor in roots of cotton seedlings. Canad. J. Bot. 1974, 52, 2423–2426. [Google Scholar] [CrossRef]
- Chamberlain, C.J. Methods in plant histology., 5th ed.; University of Chicago Press: Chicago, United States, 1932; pp. 1–416. [Google Scholar]
Figure 1.
Brachystele guayanensis (Lindl.) Schltr. (a) Habit; (b, c) Details of leaf, rhizome, roots, and base of the inflorescence axis; (d) Detail of the inflorescence; (e) Flower frontal view; (f) Flower lateral view; (g) Capsules. Photographs by Igor Soares dos Santos.
Figure 1.
Brachystele guayanensis (Lindl.) Schltr. (a) Habit; (b, c) Details of leaf, rhizome, roots, and base of the inflorescence axis; (d) Detail of the inflorescence; (e) Flower frontal view; (f) Flower lateral view; (g) Capsules. Photographs by Igor Soares dos Santos.

Figure 2.
Leaf anatomy of Brachystele guayanensis (Lindl.) Schltr.; (a, b) Epidermis in frontal view; (c-j) Cross sections; (a, c) Detail of the adaxial surface; (b, d) Detail of the abaxial surface; (e) Leaf blade; (f) Detail of raphides under polarized light; (g) Midrib; (h) Detail of the vascular bundle of the midrib; (i) Detail of the smaller-caliber vascular bundle; (j) Margin. ep = Epidermis; fl = Phloem; fv = Vascular bundle; me = Mesophyll; xi = Xylem; * = Substomatic chamber; Arrows = Stomata. Scales: a-d, f, h-j = 50 µm; e, g = 200 µm. Photomicrographs by Igor Soares dos Santos.
Figure 2.
Leaf anatomy of Brachystele guayanensis (Lindl.) Schltr.; (a, b) Epidermis in frontal view; (c-j) Cross sections; (a, c) Detail of the adaxial surface; (b, d) Detail of the abaxial surface; (e) Leaf blade; (f) Detail of raphides under polarized light; (g) Midrib; (h) Detail of the vascular bundle of the midrib; (i) Detail of the smaller-caliber vascular bundle; (j) Margin. ep = Epidermis; fl = Phloem; fv = Vascular bundle; me = Mesophyll; xi = Xylem; * = Substomatic chamber; Arrows = Stomata. Scales: a-d, f, h-j = 50 µm; e, g = 200 µm. Photomicrographs by Igor Soares dos Santos.
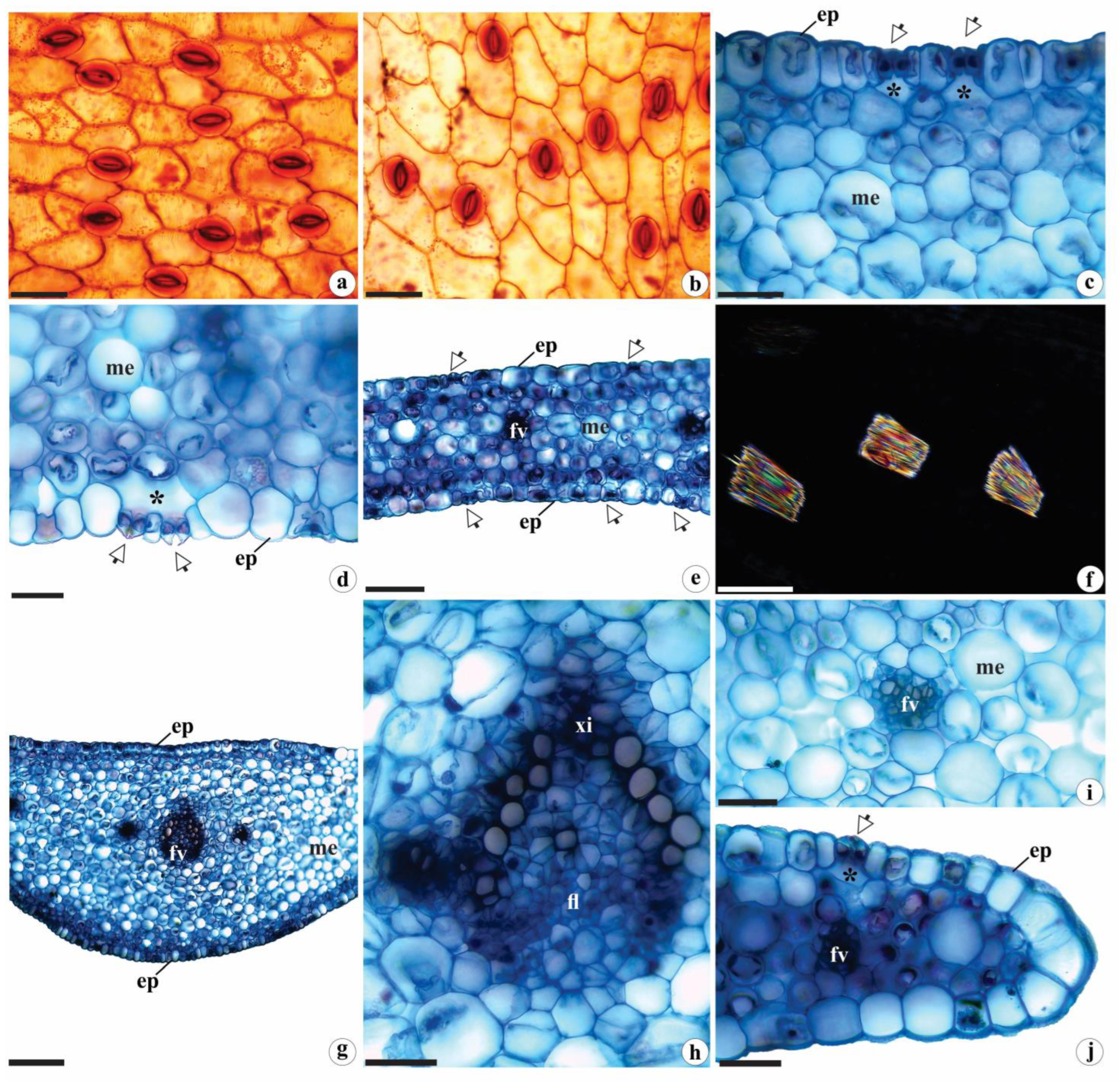
Figure 3.
Rhizome anatomy of Brachystele guayanensis (Lindl.) Schltr.; (a, b, d-j) Cross-sections; (c) Longitudinal sections; (a, b) General aspect; (c) Detail of the buds and cataphylls; (d) Detail of the epidermis and portion of the cortical parenchyma; (e) Raphides; (f) Starch grains; (g) Detail of hyphae and pelotons; (h) Detail of pericyclic fibers; (i) Detail of the central portion of the rhizomes and arrangement of the vascular bundles; (j) Detail of the vascular bundle. am = Starch grains; ca = Cataphylls; ct = Cortex; ep = Epidermis; fl = Phloem; fi = Pericyclic fibers; fv = Vascular bundles; ge = buds; hi = Hyphae; pa = Parenchyma; pe = Pelotons; ra = Adventitious root; xi = Xylem; arrows = Stomata. Scales: a, b = 1000 µm; c = 500 µm; i = 200 µm; d, f-h, j = 50 µm; e = 20 µm. Photomicrographs by Igor Soares dos Santos.
Figure 3.
Rhizome anatomy of Brachystele guayanensis (Lindl.) Schltr.; (a, b, d-j) Cross-sections; (c) Longitudinal sections; (a, b) General aspect; (c) Detail of the buds and cataphylls; (d) Detail of the epidermis and portion of the cortical parenchyma; (e) Raphides; (f) Starch grains; (g) Detail of hyphae and pelotons; (h) Detail of pericyclic fibers; (i) Detail of the central portion of the rhizomes and arrangement of the vascular bundles; (j) Detail of the vascular bundle. am = Starch grains; ca = Cataphylls; ct = Cortex; ep = Epidermis; fl = Phloem; fi = Pericyclic fibers; fv = Vascular bundles; ge = buds; hi = Hyphae; pa = Parenchyma; pe = Pelotons; ra = Adventitious root; xi = Xylem; arrows = Stomata. Scales: a, b = 1000 µm; c = 500 µm; i = 200 µm; d, f-h, j = 50 µm; e = 20 µm. Photomicrographs by Igor Soares dos Santos.
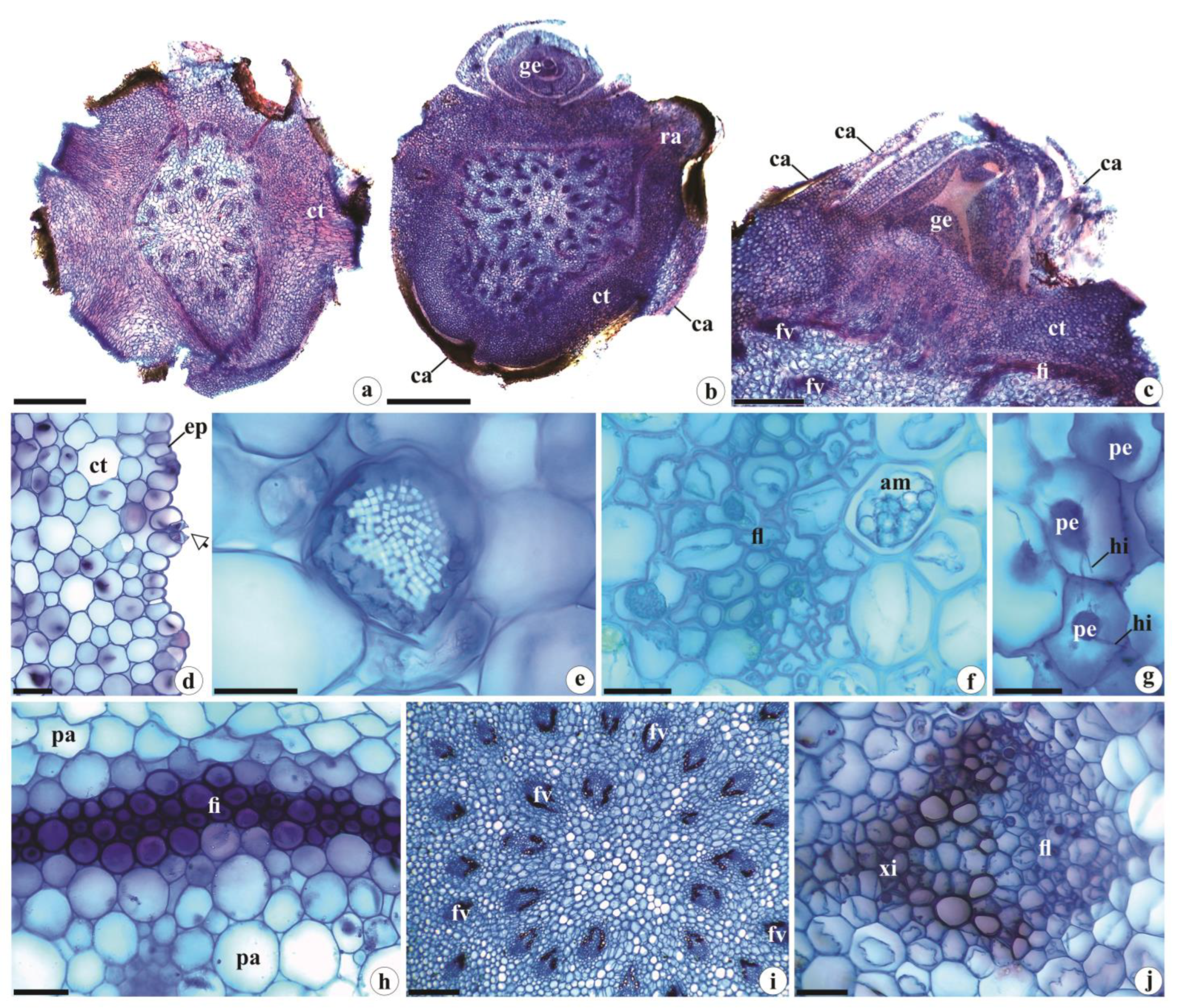
Figure 4.
Root anatomy of Brachystele guayanensis (Lindl.) Schltr.; (a-k) Cross sections; (a) General aspect; (b, c) Detail of the velamen, exodermis and trichomes; (d) Detail of the cortex. Note the presence of hyphae and pelotons in more peripheral portions; (e) Raphides under polarized light; (f) Starch grains gathered in spiranthosomes; (g, h) Detail of hyphae and pelotons in the cortical parenchyma; (i, j) Vascular cylinder; (k) Detail of the endodermis, pericycle, xylem and phloem. am = Starch grains gathered in spiranthosomes; ct = Cortex; en = Endodermis; ex = Exodermis; fl = Phloem; hi = Hyphae; pe = Pelotons; tr = Trichomes; ve = Velamen; xi = Xylem; * = Pericycle; White arrows = Casparian strips. Scales: a = 1000 µm; b, c, e, g, h, k = 50 µm; f = 20 µm; d, i, j = 200 µm. Photomicrographs by Igor Soares dos Santos.
Figure 4.
Root anatomy of Brachystele guayanensis (Lindl.) Schltr.; (a-k) Cross sections; (a) General aspect; (b, c) Detail of the velamen, exodermis and trichomes; (d) Detail of the cortex. Note the presence of hyphae and pelotons in more peripheral portions; (e) Raphides under polarized light; (f) Starch grains gathered in spiranthosomes; (g, h) Detail of hyphae and pelotons in the cortical parenchyma; (i, j) Vascular cylinder; (k) Detail of the endodermis, pericycle, xylem and phloem. am = Starch grains gathered in spiranthosomes; ct = Cortex; en = Endodermis; ex = Exodermis; fl = Phloem; hi = Hyphae; pe = Pelotons; tr = Trichomes; ve = Velamen; xi = Xylem; * = Pericycle; White arrows = Casparian strips. Scales: a = 1000 µm; b, c, e, g, h, k = 50 µm; f = 20 µm; d, i, j = 200 µm. Photomicrographs by Igor Soares dos Santos.
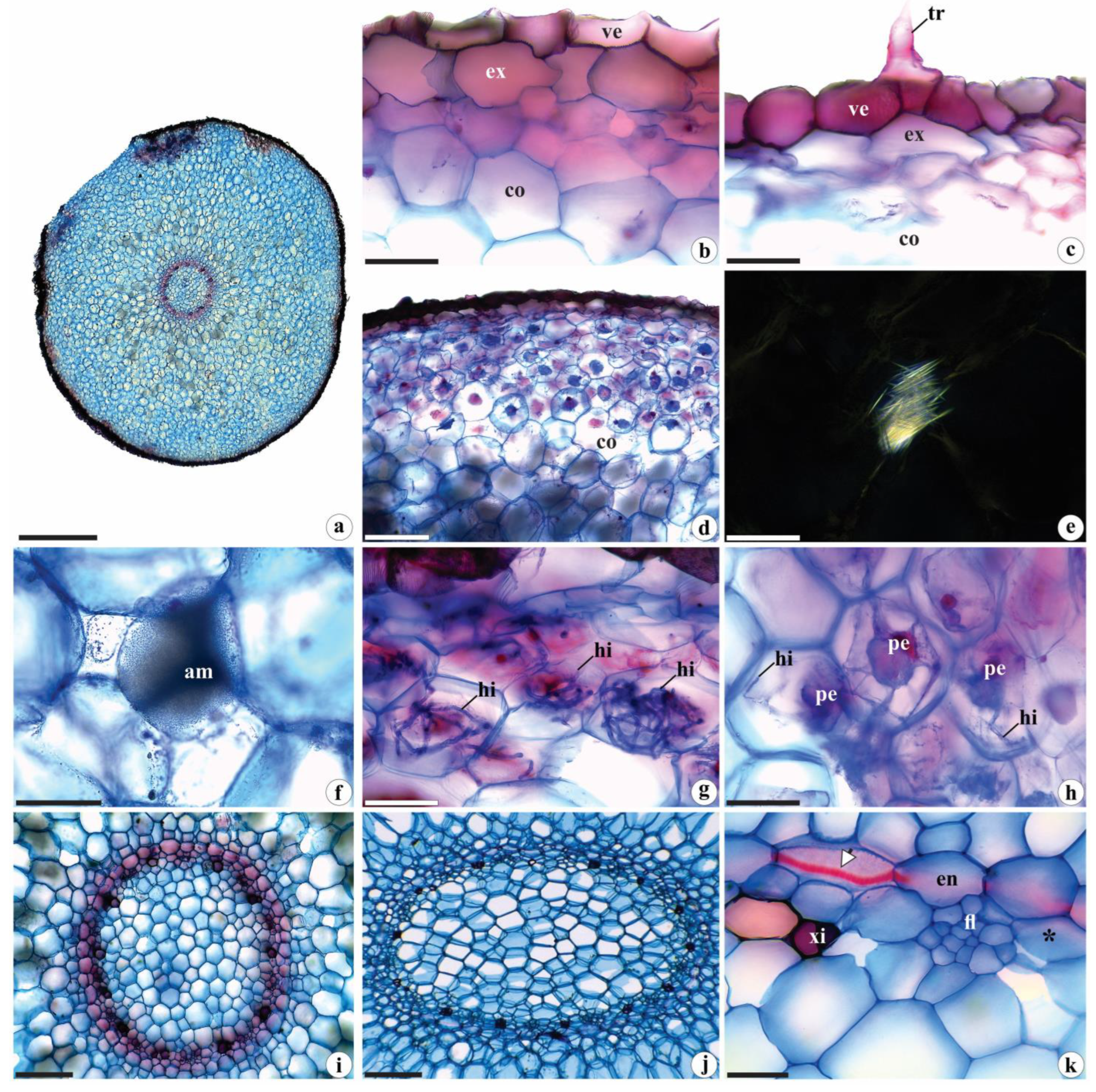
Figure 5.
Histochemical tests in the vegetative organs of Brachystele guayanensis (Lindl.) Schltr.; (a-g, i-n, p-u) Cross sections; (h, o) Epidermis in frontal view; (a, b, g, m, n, s-u) Root; (c, d, h-j, o, p) Leaf; (e, f, k, l, q, r) Rhizome; (a, b) Protein nature of fungal hyphae and pelotons; (c) Detail of the cuticle (total lipids) under polarized light; (d-g) Lignin (polarized light in e); (h-n) Starch grains. Note the spiranthosomes in n; (o) Total alkaloids. c = Cuticle; ct = Cortex; en = Endodermis; ep = Epidermis; fi = Pericyclic fibers; fl = Phloem; fv = Vascular bundle; hi = Hyphae; pe = Pelotons; xi = Xylem; * = Pericycle; Black arrows = Casparian strips; White arrows = starch grains and alkaloids. Scales: a, b, d-h, j, l, o, q-t = 50 µm; c, n, u = 20 µm; i, m, p = 200 µm; k = 500 µm. Photomicrographs by Igor Soares dos Santos.
Figure 5.
Histochemical tests in the vegetative organs of Brachystele guayanensis (Lindl.) Schltr.; (a-g, i-n, p-u) Cross sections; (h, o) Epidermis in frontal view; (a, b, g, m, n, s-u) Root; (c, d, h-j, o, p) Leaf; (e, f, k, l, q, r) Rhizome; (a, b) Protein nature of fungal hyphae and pelotons; (c) Detail of the cuticle (total lipids) under polarized light; (d-g) Lignin (polarized light in e); (h-n) Starch grains. Note the spiranthosomes in n; (o) Total alkaloids. c = Cuticle; ct = Cortex; en = Endodermis; ep = Epidermis; fi = Pericyclic fibers; fl = Phloem; fv = Vascular bundle; hi = Hyphae; pe = Pelotons; xi = Xylem; * = Pericycle; Black arrows = Casparian strips; White arrows = starch grains and alkaloids. Scales: a, b, d-h, j, l, o, q-t = 50 µm; c, n, u = 20 µm; i, m, p = 200 µm; k = 500 µm. Photomicrographs by Igor Soares dos Santos.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



