
Submitted:
31 May 2023
Posted:
01 June 2023
You are already at the latest version
Abstract
Active research of metal-containing compounds and enzymes as effective antifungal agents is currently noted. The interest in metals is due to the wide variety of ligands that can be used for metals, including chemically synthesized and naturally obtained variants as a result of the so-called "green synthesis". The main mechanism of antifungal action of metals is the triggering of generation and accumulation of reactive oxygen species (ROS). Further action of ROS on various biomolecules is nonspecific. This review highlights various hydrolytic enzymes (glucanases and proteases) that affect the structural elements of fungal cells (cell walls, membranes), fungal quorum sensing molecules, fungal own protective agents (mycotoxins and antibiotics), proteins responsible for the adhesion and formation of stable highly concentrated populations in the form of biofilms. A wide range of the substrates for enzymes allows the use of various mechanisms of their antifungal actions. The prospects of combining two different types of antifungal agents (metals and enzymes) for mycelial fungi and yeast cells are discussed in this review. Special attention is paid to the possible influence of metals on activity of the enzymes and the possible effects of proteins on antifungal activity of metal-containing compounds.
Keywords:
green synthesis
; MOFs
; amyloid proteins
; prionase
; mycotoxins
; growth inhibition
; quorum sensing
; quorum quenching
; lactamases
; biofilms
; chitinases
; lactonases
; His6-OPH
1. Introduction
Due to the accumulation of information about the role that microscopic fungi can play in the development of a number of negative processes affecting human health [1,2,3], interest is being formed in the antifungals of controlling and reducing growth, as well as the metabolic activity of these biological objects, especially those associated with pathogens [4]. A number of scientific tasks are currently related to the development of effective antifungals [5]. The seriousness of these tasks increases due to the fact that in some cases the formation of resistance of fungal cells to the chemical formulations used against them is noted [6,7,8].
Among the new trends in the development of effective antifungals, the prospects of a possible combination of various chemical compounds [8] with different mechanisms of action on fungal cells are considered, the combination of which can overcome the development of adaptive processes in fungi and, possibly, reduce the doses of the substances used, increasing the effectiveness of their action in such combinations. When implementing such a combined approach to suppressing the growth and metabolic activity of fungi, the main question arises about what is better to combine with what, and what may be unpromising. One of the possible answers to this question is based on the use of metal-containing compounds, resistance to which is not formed in most microorganisms, since the mechanism of suppression of biological processes is primarily associated with the generation of reactive oxygen species (ROS) in the cells. In the composition of such compounds with antifungal activity, the study of metals such as Ag, Cu, Fe, Zn, Se, Ni, Au, Zr, Ce, Ti and Pd is known [9,10,11,12]. At the same time, current scientific research on the antifungal properties of metals is mainly focused on the study of Ag and Au nanoparticles (NPs) [10,11,12,13,14,15], since antimicrobial effectiveness of their action has been known for a long time.
Among the works developing this direction, it should be noted the use, along with various organic synthetic ligands for the metals, of the so-called "green synthesized" metal-containing NPs. They are formed inside the cells of microorganisms in vivo or using plant extracts, polysaccharides of phototrophic microorganisms and extracellular enzymes of mycelial fungi [10,14,15,16,17]. Thus, these works focus the attention of researchers on the possibility of using natural biopolymers to obtain the desired metal-containing compounds. At the same time, it was found that cheaper and more environmentally friendly "green synthesis" makes it possible to obtain NPs with identical antifungal properties compared to similar chemically synthesized metal-containing compounds, and in some cases superior to them [17].
It is possible to increase the efficiency of the use of metal nanoparticles by obtaining combined preparations with cyclic peptides that exhibit antifungal properties [18]. The combination of metal NPs with known chemical fungicides makes it possible to reduce the minimal inhibiting concentration (MIC) of the latter by more than 8 times [17], but in this review we decided to consider the possibility of combining metal-containing compounds not with chemically synthesized fungicides, namely with biological molecules, having catalytic properties, in particular, with various enzymes exhibiting antifungal activity.
Since enzymes, unlike peptides that exhibit antimicrobial activity, have catalytic activity [19], which allows them not just to trigger destructive processes against fungi, but to repeatedly participate in these acts of biocatalysis, deepening antifungal processes. In addition, a wide range of substrate specificity of action for the enzymes themselves allows us to consider the possibility of their manifestation not only of destructive activity against fungal cells, but also against their most important molecules involved in the formation of resistance of fungal cells (quorum sensing (QS) molecules of adhesion proteins) [20], and molecules that ensure their own safety (antibiotics [21] and mycotoxins [22]).
In this regard, it was interesting from a scientific and applied point of view to analyze the current scientific background in the field of creating possible combinations of metal-containing compounds with enzymes possessing antifungal activity, as well as to evaluate the potential available in this area that can be used in practice. This was the purpose of this review. In order to better understand the existing prospects and limitations for the development of these combinations, initially in this review special attention was paid to recent information about metal-containing compounds and enzymes, which in recent years researchers have paid special attention to when studying their antifungal characteristics. At the same time, for enzymes, special attention was paid to the effect of different metals on their established antifungal activity, if such information was present in the analyzed publications.
2. Antifungal agents based on metal-nanoparticles, metal-organic frameworks and their derivatives
In current scientific investigations, antifungals became the widely used objects of developments, where metal was found in an oxidized forma as parts of components or organometallic frameworks (MOF) (Table 1, [11,12,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37]). The metal center in MOFs with antifungal activity is most often represented by metals such as Ce [11] and Cu [25,32,34,38]. In the composition of MOFs, 4,4’,4”-nitrilotribenzoic acid [11], 1,4-benzene dicarboxylate [35], 1,3,5-benzentricarboxylate [32,38], glutarate, 1,2-bis(4-pyridyl)ethane, 1,2-bis(4-pyridyl)ethylene, 1,3-bis(4-pyridyl) propane [34], and other [39] are mainly used as organic linkers for metals.
A special place among formulations with antifungal activity is occupied by artificially created nanomaterials with intrinsic enzyme-like characteristics, which are called "nanozymes" [11,36,41,42]. Nanozymes successfully combine the unique properties of nanomaterials and mimic the catalytic functions of enzymes [43]. The activity of scientists is mainly focused on research related to the generating of ROS. Known nanozymes with antifungal properties can inhibit the action of the following enzymes: peroxidase, catalase, superoxide dismutase [11,36,41,42]. The inclusion of Ce [11], Ag, Fe [36,40], Pt, Pd, Cu, Ti [12] and some other metals [41] as components of nanozyme compositions is under development.
Combined nanozymes are expected to have higher efficiency and an expanded spectrum of action in relation to biological objects. The combination of peroxidase-like activity in one formulation, manifested due to the presence of Fe3O4 [36,40] or TiO2 [12], and different biological activity of additional components makes it possible to expand the spectrum of potential pathogens, including fungal and bacterial cultures that can be destroyed during disinfection using such nanozymes.
Speaking about the future prospects for the development of new nanozymes, it is appropriate to pay attention to their potential target, such as the fungal cell wall. The composition of the cell wall of the most fungi contains such target components as chitin and β-1,3-glucan. In this regard, it is interesting to see the appearance of such antifungal nanozymes, which will have not peroxidase, but hydrolase-like activity [43]. It is interesting, but now such metal-containing catalysts are being developed, but for use in another field, in particular, for the destruction of microplastics [44]. It would be extremely interesting to test metal-containing nanozymes, already developed for other purposes, as antifungal agents. We assume the presence of a certain potential here, since among the mentioned nanozymes there are those that exhibit hydrolytic activity.
It is possible to note a certain variety of methods used for the synthesis of MOFs with antifungal activity [11,25,32,33,34], while the antifungal features of the formulations derived from metal NPs and their derivatives, as well as nanozymes [45,46], are due to several different mechanisms of their action. Metals and preparations based on them not only contribute to the appearance of ROS in cells [47], but also break up the integrity of the cell membrane of fungi, as well as processes associated with transmembrane transfer, they inhibit the activity of membrane proteins and intracellular enzymes, disrupt the structure of nucleotides, nucleosides and affect the gene expression.
For actively studied Ag nanoparticles, it was found that they mainly inhibit the expression of genes of proteins involved in the biosynthesis of fatty acids and lipids, which are components of membranes. It is believed that for a more successful manifestation of antifungal properties, metals must pass into an ionic state, for example, under the action of oxygen dissolved in the medium. Silver ions are able to bind almost irreversibly to thiol-containing cysteine and glutathione, as well as flavin groups, which are components of membrane proteins. By disrupting the three-dimensional structure of membrane proteins, metal ions inhibit membrane phosphorylation reactions, and the synthesis of adenosine triphosphate (ATP) is declined. In addition, the permeability of the membrane increases, protons enter from the external environment into the cell, the operation of the proton pump and the flow of transmembrane transfer processes are disrupted. In the presence of Ag+ silver ions, Fe(II) ions are released from Fe-S clusters present in large quantities in respiratory chain enzymes, which activate the Fenton reactions. As a result, the decomposition of hydrogen peroxide into hydroxyl and hydropyroxyl radicals proceeds more actively. When metal NPs are introduced into the medium, an increase in the expression of genes of systems responsible for oxidative stress can be observed, which is explained by the attempt of cells to remove toxic ions from the cytoplasmic space [48]. The advantage of using antifungal metal-containing agents in the form of NPs exists due to the fact that they adhere well to the surface of the cell membrane, in this way it is possible to increase their local concentrations on its surface, as well as facilitate the penetration of metal ions and NPs into the cell owing to violations of transmembrane transfer processes.
The use of metals in the composition of particles with larger than nonosize, such as some MOFs, seems promising as antifungal agents, since the reduction of metal ions in the composition of MOFs can generate superoxide species with further degradation of biomolecules and inhibition of cell growth [32]. Under certain conditions, metal ions are released from MOFs into the environment with fungi and possess a toxic effect on them [33]. However not all MOFs have anti-fungal properties. Apparently, the type of metal in the MOFs composition has a key role in this case.
Thus, NPs of UiO-66 (zirconium-based MOFs, 68.4±8.5 nm) when injected into a medium with white rot fungus Phanerochaete chrysosporium showed a weak effect on cells, did not penetrate them, but slightly damaged the cell wall and inhibited the activity of the laccase secreted by them [49].
It should be noted that some fungi, for example, representatives of the genera Penicillium and Aspergillus, show tolerance to the toxic effect of metals, in particular, to copper ions [33], whereas for many ecosystem participants, including humans, the high enough toxicity of same metals may be obvious. The strategies of tolerance of fungi to metals are based on the accumulation of metals in the composition of fungal biomass, the formation of complexes with various biomolecules and the active outflow or excretion of metals from the cells [46]. In this regard, it is interesting to search for those metal-containing compounds which antifungal doses remain ecologically acceptable for their use.
Green synthesis and biosynthesis of NPs can result in inclusion and/or adsorption of multiple organic compounds contained in the culture broth and cells, including proteins among others. Usually it is hard to distinguish the contribution of such protein(s) in antifungal activity of the end product. Thus, many authors even didn’t try to. However, multiple such (nano)formulations have been developed to date using bacterial [50,51], fungal [52], plant [53,54] and other isolates.
Little bit more accurate method applies, at least, partially purified proteins [55,56] and pure polypeptides [57]. For example, application of Ag lead to profound antibacterial activity of obtained formulation [50,54], thus giving additional functionality and opening an opportunity to treat multimicrobial associations (Table 2 [37,50,51,52,53,54,55,56,57]). Antioxidant activity is another famous mode of action and can be intrinsic to formulation due to metals/method used [53,54] or can be embedded by additional components [55,56].
It should be emphasized here that yeasts of genera Candida [11,24,26,29,31,32,34,37,50,52,53] have the ability to form biofilms, and the possible their degradation [52,53], and not the cells themselves, also allows to obtain an effective antifungals. At the same time, the presence of biofilm makes yeast cells more resistant to antifungal treatment(s) [59].
Yet another interesting option not considered to date is direct interaction of metal-containing nanomaterials with adhesins belonging to the family of agglutinin-like sequence (ALS) proteins and forming nanodomains on cell surface. Indeed, ALSs are long-known multipurpose utilities which participate in biofilm formation/adhesion and also capable to bind with ferritin [60] and even macroscopic metal surfaces [61]. Interestingly, interaction with metal surface similarly results in ROS generation and modulation of several genes, particularly issued in this work, being responsible for intracellular metal homeostasis.
Filamentation is the next step of biofilm maturation after initial cell attachment. It, at least partially, depends on cascades with ALSs and thus disturbing their regulation could lead to inability of (pseudo)hyphal formation. Strong evidence of such inhibition were shown with Ag NPs [52] and Fe3O4 NPs [53]. Noteworthily, medium-chain fatty acids (e.g., heptanoic and nonanoic acids) and, to a lesser extent, farnesol molecules not only downregulate genes responsible for hyphal formation, but also upregulate genes responsible for yeast-cell morphologic form [62]. Thus, cumulative effect can occur due to proteinaceous, fatty acid and other biologically active compounds forming a corona of green synthesized nanomaterials.
It should be noted that combination of formulation with conventional antifungals (e.g. fluconazole) acting on another biochemical target(s) within yeast resulted in synergetic effect and improved efficiency [52]. Another important point about possible toxicity of prepared formulations to humans and other non-target organisms is worth mentioning. Some toxicity is shown for as prepared Ag NPs towards mouse fibroblasts, human lung carcinoma epithelial cells and human red blood cells [52]; for Ti/Ag NPs towards human skin fibroblasts [51]. CuO formulation with poly-ε-lysine [57] has no influence on seed germination.
Many filamentous fungi have a spore germination stage which can be triggered by different biotic and abiotic factors [63]. This is a hard target for influence but is still possible to be affected using, for example, nanomaterials [64].
There are several pathways, at least, to prevent activation of the process and, at most, to irreversibly disable it, for example, while degrading the spores. The latter case is rather limited now due to (bio)compatibility issues and is a matter of future research. Germination is initiated when outer and/or inner receptors are activated by the factor or when autoinhibition is eliminated by it. Due to their small size NPs can be adsorbed on the surface of spores and visualized there [51]. After that, physical damage and/or hindrances for penetration of activating factors, as well as chemical modification(s) of spore envelope, outer receptors and transport channels are possible. The internal resources of spores are limited and they can’t sustain a constant regeneration process. As a result, many of them may lose the ability to germinate properly, while the remaining ones may be severely depleted, which, for example, appeared in production of decreased amounts of extracellular enzymes [51]. Moreover, additional functionalities of antifungal formulations, e.g. ROS generation, can be very useful to improve the rate and efficiency of the process of spore inactivation [57]. It should be noted here that measured values of leaked proteins and nucleic acids from the spores [51] seem to be unreliable and should be verified.
Similar mechanism of action of nanomaterial formulations is even more profound for vegetative filamentous fungi [57]. Structurally mycelium is less durable than spores while visualization and/or measurement of inflicted damages is easier. Noteworthily, exactly proteinaceous corona multiply improves anti-sporulation, anti-germination and other effects of the antifungal formulations.
Prepared antifungal formulations were tested in planta in the soluble form for prophylaxis and therapy [57], in the coating films for fruits [55], in the nanofibrillar package material [56], etc. However, without special contamination by fungal cells some reports [55;56] appear to be rather preliminary. Application in prophylaxis and therapy modes lead to very similar antifungal activity [57] but removal of fungi was incomplete at applied doses.
Many authors still use disk diffusion test (Table 2) which is useful for medical diagnostics but insufficient for research field. (Micro)dilution test seem to be much more preferable, especially in the case of non-filamentous fungi. For filamentous fungi tests of spore germination and/or mycelium growth are applied. Nevertheless, all these results don’t allow distinguishing between true killing of cells to death and cells in the stasis state. It is highly needed to much wider implement modern tests determining true cellular state, like live/dead assay, ATP test [65], etc.
Discussing possible perspectives of future research, specific modification of metal NPs for targeted delivery of antifungal formulation, as it was realized in the case of organic liposomes [66], seems to be a very interesting direction.
3. Enzymes as antifungal agents
3.1. Antifungal enzymes using cell structural components of fungi as substrates
Discussing the possibilities and prospects for the use of various enzymes in the composition of antifungals, it should be noted that their diversity is determined by the spectrum of targets on which these enzymes can have notable effect, leading to a halt in the fungal growth, disruption of metabolism and death of fungi. Such targets for enzymatic action include: structural elements of fungal cells (cell walls, membranes)[67], nucleic acids [68], fungal Quorum Sensing molecules (QSMs) regulating fungal resistance to various negative factors and protect them (mycotoxins,antibiotics)[69]), peptides and proteins involved in the formation of stable forms such as biofilms (adhesives, hydrophobins) (Figure 1).
Enzymes as antifungal agents are interesting because they are proteins present in various natural sources (plants, microorganisms, animal tissues) (Table 3 [69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102]), which are designed to protect living objects from the effects of fungi. The necessary enzymes can be produced in various host cells as recombinant forms.
The analysis of enzymes exhibiting various antifungal activity indicates that most of them are hydrolases acting on polysaccharides present in the structure of the cell wall of fungi or involved in the formation of biofilms. The greatest effect was observed in the case of chitinases [69,70,71,72,73,74,75,76,77,78,79,80,81], among which there are both exo- and endochitinases, but their simultaneous presence in enzyme complexes used to pressure the growth of fungi is the most successful [80,82]. As a number of studies have shown [82,100,101], such a combination of chitinases with different substrate specificity can be successfully supplemented by the action of other hydrolytic enzymes (proteases and glucanases) [90,91,99,100,101], which in the general effect of the enzyme complex on fungal cells use molecules as substrates performing the role of structural elements of cell wall and membranes in fungi.
It is important to note that yeast cells are usually difficult to destroy, since the cell walls can form capsules or resistant spores. DNA can be extracted from yeast cells using a complex of lysing enzymes such as Lyticase, including mixture of β-(1-3)-glucan-laminar-ipentaohydrolase, β-(1-3)-glucanase, protease and mannanase [101]. The activity of this complex induces partial formation of spheroplasts; subsequently, the spheroplasts are lysed with the release of DNA.
However, it should be emphasized that not all chitinases known today [103] can be used as antifungal agents, since the diverse structure of fungal polysaccharides is characterized by the presence of various glycoside residues of different lengths and often does not correspond to the preferences of those substrate specificities possessed by most of these enzymes. In addition, the levels of biosynthesis of these enzymes cannot meet the needs that arise even when studying their properties, and in this regard, it is necessary to resort to obtaining their recombinant forms. E. coli BL21 (DE3) cells are most often used for this purpose [72,73,79,84,85].
Nucleases hydrolyzing RNA and DNA of fungi attract particular attention among enzymes that have antifungal activity [96,97,102]. The use of several nucleases at once [96,97,98] or nuclease in combination with glucanase [102] leads to the fact that not only the growth of fungal cells is stopped, but membrane destruction (permeabilization and depolymerization) is observed, lowering of mitochondrial membrane potential, degradation of target cellular nucleic acids and the death of microbial cells.
Oxidoreductases, in particular, peroxidases are standing at the second place after hydrolases in popularity among enzymes used as potential antifungal agents [94,95]. These enzymes catalyze the oxidation of fungal molecules by reducing hydrogen peroxide. The limitations in the use of these enzymes as antifungal agents are associated with a lower efficiency of their action in comparison with hydrolases and the need to introduce additional H2O2 into the medium with fungi.
Giving priority to hydrolases when necessary to influence fungi, it can be emphasized that they generally have the potential for antifungal effects in a fairly wide range of pH values (3.0-11.5) and temperatures (up to 80 oC) (Table 1). At the same time, it should be noted that the activity of hydrolases strongly depends on the presence of various metals in the media of their functioning [72,73,76,77,81,88,91]. In such media the most attractive options are those combinations of enzymes and metals that can significantly increase the level of antifungal action of hydrolases. Among the metal ions, which in the largest number of studies have shown a stimulating effect on the activity of hydrolases, Cu2+ [76,77] and Ca2+ [81,84,85,91] should be singled out, although their positive effect is not at all unambiguous, and in some cases they had the opposite (inhibitory) effect on the hydrolytic activity of enzymes. At the same time, the positive results obtained during enzymatic reactions directed against fungi in environments in the presence of metals indicate the expediency of searching for possible combinations of metals and enzymes in the development of new antifungal formulations.
3.2. Enzymes hydrolyzing fungal proteins with amyloid characteristics
Special attention should be paid to the fact that yeast and mycelial fungi are able to form amyloids, which are unbranched fibrils consisting of monomers stacked on top of each other and stabilized by intermolecular β-layers. For example, monomers of hydrophobins of class I, small surface-active proteins produced by fungi, form amyloid fibrils that perform many functions [104]. It is known that the specific functions of hydrophobins synthesized by fungi can enhance their pathogenicity. Thus, A. fumigatus can cause invasive aspergillosis in patients with weakened immunity due to the amyloid-forming ability of hydrophobin RodA [105,106]. The formation of amyloid by hydrophobin MPG1 in M. oryzae contributes to rice pyriculariosis [107]. One of the most well-described examples of amyloid proteins in yeast cells is the Cdc19 protein from S. cerevisiae, which, in the absence of glucose, self-aggregates into an amyloid-like aggregate to avoid degradation under stressful conditions [108].
It is known that the yeast cells of C. albicans, often used in studies of antifungals, also contain proteins with amyloid characteristics. Thus, the proteins As1, As3 and As5 from the ALS-type adhesion family have the ability to self-aggregate. The presence of an amyloid sequence in the monomers of these proteins leads to the formation of hydrophobic nanodomains that promote cell adhesion of C. albicans on biotic or abiotic surfaces and improve their ability to form biofilms [109,110]. It is assumed that Sap 6, Rbt 1, Page 59 and Pga62 proteins, as well as adhesives, play a significant role in the appearance of C. albicans biofilms due to the presence of an amyloid-forming sequence in their structures [111,112,113,114].
Today, due to their ability to be transmitted from “mother” cells to “daughter” cells, yeast prions are classified as infectious, for example [URE3] and [PSI+] in S. cerevisiae cells, HET in Podospora anserina, HELLP in Chaetomium globosum [115].
The presence of similar conditions for the formation of yeast prions and common molecular properties with pathogenic human amyloids has now led to the creation of models of neurodegenerative diseases based on yeast prions. The ways of their regulation are being investigated in order to develop new effective therapeutic agents and approaches to the treatment of diseases associated with prion proteins [116]. In this regard, the interest in enzymes that are capable of hydrolyzing amyloid aggregates formed by fungi is huge not only from the point of view of the development of antifungals that reduce the level of biofilm formation, but also from the point of view of the potential use of the enzyme-containing formulations for the treatment of neurodegenerative diseases in humans. Information about such proteases hydrolyzing amyloid proteins is presented in Table 4 [117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132].
Discussing the prospects for the possible use of enzymes hydrolyzing fungal amyloid proteins, it should be noted that so far there are a few such studies. The ability of several proteolytic enzymes, such as subtilisin, keratinases and proteinase K, to degrade yeast prion aggregates of protein Sup35NM under various conditions was investigated [127,128,129,130]. It has been shown that hexameric AAA+-ATPase (Hsp104), which is a yeast chaperone, is involved in the fragmentation of large fungal amyloid fibrils. It is believed that the direct binding of Hsp104 to amyloid fibrils prevents the reproduction of yeast prions. Since Hsp104 is absent in the cells of multicellular animals, including mammals, the possibility of constructing variants of Hsp104 with the potential for use for the degradation of abnormal human proteins is being investigated [116].
Despite the limited number of studies in the field of enzymatic degradation of yeast prions, a number of proteolytic enzymes are known today that can degrade prion proteins and amyloids associated with human diseases: subtilisin-like serine proteases TK-SP from hyperthermophilic archaeon T. kodakarensis [117], nattokinase from Bacillus subtilis natto [118], subtilisin 309 and protease from Bacillus lentus [119,121], two prionzymes from Bacillus subtilis and Bacillus lentus [120,122], subtilisin-like protease MSK103 from Bacillus licheniforms [123], enzyme E77 from Streptomyces sp. [124], subtilisin-homolog pernisine from the extremophile archaea Aeropyrum pernix [125], serine protease from lichens [126].
Multiple metalloenzymes have been reported to have an important role in the degradation of Aβ [131,132]: two metal-activated keratinases, Ker1 and Ker2, from an actinomycete Amycolatopsis sp. MBRL 40; NEP - a zinc-dependent metalloprotease, cleaving various vasoactive peptides; IDE - another zinc-dependent metallopeptidase, which could cleave insulin and amyloid Aβ. The ability to cleave amyloid precursor proteins has been confirmed in Zn-containing transmembrane metalloproteases [132]. At the same time, the influence of redox-active metals such as Cu and Fe (affecting the pathogenesis of Alzheimer's disease) was established, which consists in increasing the biosynthesis of the metalloproteases under discussion. The influence of the same metals on the activity of these enzymes has not yet been investigated, but is of great scientific and practical interest.
Although there is still no effective enzymatic formulation for the degradation of prion proteins, new proteolytic enzymes whose prionase activity should still be investigated continue to be discovered and studied [133,134,135]. In this review, we focus readers' attention on such enzymes as a potential basis for the development of new antifungals probably with some anti - neurodegenerative effect.
3.3. Enzymes hydrolyzing mycotoxins, antibiotics and QS molecules (QSMs) of fungi
To date, a significant amount of information has been accumulated about QS in the cells of various fungi and molecules that are produced by the fungi themselves in order to trigger the processes of cell transition to a state of stable intercellular communication, synchronization of the functions of multicellular populations and biochemical changes in the cells themselves, which are the response of fungi to an increase in their concentration per unit volume [136,137,138,139,140]. The ability of individual enzymes to catalyze the hydrolysis of QSMs fungi, allows them to be attributed to the so-called Quorum Quenching enzymes (QQE). Gluconolactonase [141]) and hexahistidine-containing organophosphorus hydrolase (His6-OPH) [142,143], esterases [144,145]) have been identified as such enzymes acting against fungi today (Table 5).
Discussing the potential of these enzymes as candidates for inclusion in combined antifungals with metal-containing compounds, it can be noted that for His6-OPH such possibilities have already been demonstrated and proved promising, while Ta NPs [149,150] proved to be the most effective option for such a combination. However, so far such combined antimicrobials have been investigated only against bacterial cells [151], and their effectiveness against fungal cells has yet to be confirmed.
Interesting use cases for combining with metal-containing compounds are enzymes that carry out the destruction of mycotoxins synthesized by fungi in the CFR state. At the same time, it should be noted that, as in the case of CSM hydrolysis, among the enzymes that carry out the destruction of various mycotoxins (zearalenone, patulin, deoxynivalenol, ochratoxin) there are all the same enzymes that are listed in Table 5: lactonases, esterases, lipases [22], His6-OPH [68,152]. In this regard, with their involvement in combined antifungal formulations, a very interesting option may turn out to provide a multi-targeted action due to the promiscuous activities of these enzymes.
Continuing to analyze possible variants of enzymes that can be considered as candidates for creating combined variants with metal NPs, it is undoubtedly necessary to pay attention to enzymes that are able to catalyze the hydrolysis of antibiotics synthesized by fungi among other secondary metabolites in their QS state. Here, the undisputed leaders are beta-lactamases, known to everyone due to studies of bacterial antibiotic resistance to natural and semi-synthetic penicillins and cephalosporins [153].
It is interesting to note that QQE including His6-OPH are close "relatives" for metallo-β-lactamases [154]. Moreover, the structural analogy revealed between phosphortriesterases (of the same His6-OPH) and some nucleases indicate that all these enzymes can catalyze to one degree or another same reactions with a certain preference for individual substrates. Since these enzymes have been mentioned here more than once in connection with their various targets of action in fungal cells (Table 3, Table 4 and Table 5), their use in research on the development of new antifungals may be not only new, but also promising. Surprisingly, an active search for data on the use of metallo-β-lactamases in the content of any antifungals to give them a number of catalytic activities, as discussed above, did not reveal any.
It should be noted here that many of these enzymes contain different transition metals, particularly Zn(II), Mn(II), and Fe(II)/Fe(III) in the active center [154], which can be positively taken into account when creating combinations with metal-containing compounds, since there are mushrooms, sensitive to these metals (Table 1 and Table 2). In addition, the combination of these enzymes with metal-containing compounds that are not embedded in the active center of enzymes, but can exhibit significant antimicrobial activity at low MIC values [149,150], looks interesting and promising.
4. Combination of antifungal activity of enzymes with metal-containing compounds
It is known currently that many sources and types of enzymes can be used to prepare antifungal formulations with metal NPS: bacterial keratinase [155] and chitinase [156]; archaeal protease and lipase [157]; fungal β-1,3-glucanase, N-acetylglucosaminidase, chitinase and acid protease [158,159], etc. Such formulations can possess secondary antioxidant [155,156] and/or specific inhibitory activity [155]. Also additional antibacterial action mode of these combinations is widely present [155,157,159,160] (Table 6 [40,155,156,157,158,159,160,161]).
Green synthesized metal NPs are of great interest for production of enzyme formulations [155,157,158,159]. β-1,3-glucanase(s) and, to a lesser extent, N-acetylglucosaminidase(s) are prevalently adsorbed by Ag NPs as compared to chitinase(s) and acid protease(s) [158]. All together these enzymes on Ag NPs not only inhibit mycelium growth but also prevent formation of sclerotia thereby leading to lifecycle arrest.
Interestingly, “un-capping” of Ag NPs (i.e. desorption of enzymes) leads to detectable increase of their size and is likely to be a result of their aggregation [159]. At the same time, negative net charge of ‘uncapped’ Ag NPs argues for the substitution of enzymes by sodium dodecylsulfate used as solubilizer. That can contribute into increased toxicity of such ‘un-capped’ NPs towards non-target organisms and cell lines [159]. Surprisingly, ‘un-capped’ Ag NPs are ineffective in a mycelium growth test at all and only twofold decrease number of sclerotia as compared to control experiment without any effector.
Similar to germination, formation of sclerotia is known to be regulated by multiple genes though there are a lot of gaps in our knowledge about this process [162].
As a result, a biochemical composition of cell wall changes dramatically, for example: the most abundant components of Sclerotium rolfsii hyphae – polysaccharides and lipids – shift in 1.5-2 times (down and up, respectively) while unhydrolyzable compounds (so-called ‘melanin-like pigments’) increase numerously and become the second prevalent subclass (after polysaccharides). The last ones have been shown to propagate resistance of sclerotia towards environmental factors and, for example, to slaughter via hydrolytic action of extracellular glucanases and chitinases [163]. Moreover, the leakless thick rind can be formed from such melanized cells on the sclerotia surface [164], further limiting enzymatic hydrolysis and antifungal penetration. Thus, polyphenol-degrading activity may be useful in addition to antifungal formulation. Another rational functionality in such formulation(s) to treat sclerotia appears to be antioxidant activity discussed previously since ROS also affect sclerotial development somehow [162].
During field trials of chitinase-based formulation against filamentous fungi 156] it was found to be slightly less effective than the same formulation with live biocontrol agent (Streptomyces cellulosae). It can be a consequence of differing profile of protective gene modulation in the plant by these formulations.
As determined for peptide melittin, slow release of active compound from the Zn-MOF matrix occurs and maximal amounts (60%) is released at pH 6 during 24 h [161]. Antifungal activity of melittin is naturally decreased in 3 times during encapsulation within Zn-MOF at 30 wt.% loading. However, lactoferrin been added into such formulation almost twofold improves it. Altogether yeast adhesion to the surface during biofilm formation and (pseudo)hyphal transformation are inhibited.
Melittin is known to disturb membranes of different (micro)organisms, activate several transmembrane receptors, depolarize membranes, etc. Some of these effects are shown within composite formulation also [161]. Moreover, lactoferrin being transporter of iron ions and having other possible activities [165] greatly improves antifungal activity of melittin and especially towards pre-formed biofilms [161]. Also synergic action of lactoferrin and melittin is detectable using animal infection model in vivo. Lactoferrin can bind to fungal cell surface itself and affect biofilm formation and yeast-to-hyphal transition in combination with conventional drugs [166]. Thus, lactoferrin and melittin may interact with multiple and differing targets on yeast cell wall and within cell while amplifying antifungal activity of each other.
As mentioned previously, some toxicity is shown for as prepared Ag NPs towards lung fibroblasts of Chinese hamster, embryo fibroblasts of albino Swiss mouse, human aneuploid immortal keratinocytes as well as to the roots of onion [158,159]. Moreover, such formulations affects soil microbial (bacteria and fungus) community in situ after single exposure during, at least, 360 days [159]. Interesting that toxicity of such polypeptide as melittin towards macrophage cell line from a mouse tumor is greatly decreased within Zn-MOF formulation [161]. Since then it is necessary to hope but check the decrease in toxicity for people of the antifungal compositions contained both metal compounds and enzymes.
5. Conclusions
Analyzing the approaches to the development and combination of antifungals that were discussed 10 years ago [167] and those that are currently discussed [168], it can be noted that there is no much difference between them and there are no discussions of solutions based on the use of metal-containing compounds and enzymes, and even more so on their possible combinations. At the same time, the expediency and possible effectiveness of the combination of substances that enables the use of different mechanisms for suppressing the growth and metabolic activity of fungi have long been beyond doubt. It remains only to solve the problem of choosing partners for the most effective and safe combination of antifungal agents for humans. The emergence of new knowledge about possible targets for exposure to fungi and the analysis of the palette of known antifungal agents can form fresh ideas about possible useful combinations. Of course, those variants are interesting in which not just inhibitors of biochemical processes are used, which, as a rule, react with their targets in a one-to-one ratio and require specific binding, but namely biocatalysts are attractive for the processes of irreversible degradation of key fungal biomolecules which repeatedly enter into decisive catalytic acts.
Author Contributions
Conceptualization, E.E.; investigation, I.L., A.A., D.M., N.S., O.S., O.M. and E.E.; data curation I.L. and E.E.; writing—original draft preparation, I.L., A.A., D.M., N.S., O.S., O.M. and E.E.; writing—review and editing, I.L., A.A. and E.E.; supervision, E.E.. All authors have read and agreed to the published version of the manuscript.
Funding
The research was funded by Russian Science Foundation (23-14-00092).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fisher, M.C.; Gurr, S.J.; Cuomo, C.A.; Blehert, D.S.; Jin, H.; Stukenbrock, E.H.; Stajich, J.E.; Regine Kahmann, R.; Boone, C.; Denning, D.W.; Gow, N.A.R.; Klein, B.S.; Kronstad, J.W.; Sheppard, D.C.; Taylor, J.W.; Wright, C.D.; Heitman, J.; Casadevall, A.; Cowen L.E. Threats posed by the fungal kingdom to humans, wildlife, and agriculture. MBio 2020. 11(3), e00449-20. [CrossRef]
- Garg, D.; Muthu, V.; Sehgal, I.S.; Ramachandran, R.; Kaur, H.; Bhalla, A.; Puri, G.D.; Chakrabarti, A.; Agarwal, R. Coronavirus disease (COVID-19) associated mucormycosis (CAM): Case report and systematic review of literature. Mycopathologia 2021, 186, 289–298. [CrossRef]
- Raut, A.; Huy, N.T. Rising incidence of mucormycosis in patients with COVID-19: Another challenge for India amidst the second wave? Lancet Respir Med. 2021, 9(8), e77. [CrossRef]
- World Health Organization. WHO fungal priority pathogens list to guide research, development and public health action. World Health Organization, Geneva, Switzerland, 2022, pp 48. Available online: https://www.who.int/publications/i/item/9789240060241 (accessed on 29 May 2023).
- WHO. WHO releases first-ever list of health-threatening fungi, 2022. World Health Organization. Available online: https://www.who.int/news/item/25-10-2022-who-releases-first-ever-list-of-health-threatening-fungi (accessed on 30 May 2023).
- Robbins, N.; Caplan, T.; Cowen, L.E. Molecular evolution of antifungal drug resistance. Annu. Rev. Microbiol. 2017, 71, 753–775. [CrossRef]
- Fisher, M.C.; Hawkins, N.J.; Sanglard, D.; Gurr, S.J. Worldwide emergence of resistance to antifungal drugs challenges human health and food security. Science 2018, 360, 739–742. [CrossRef]
- Rabaan, A.A.; Sulaiman, T.; Al-Ahmed, S.H.; Buhaliqah, Z.A.; Buhaliqah, A.A.; AlYuosof, B.; Alfaresi, M.; Al Fares, M.A.; Alwarthan, S.; Alkathlan, M.S.; Almaghrabi, R.S.; Abuzaid, A.A.; Altowaileb, J.A.; Al Ibrahim, M.; AlSalman, E.M.; Alsalman, F.; Alghounaim, M.; Bueid, A.S.; Al-Omari, A.; Mohapatra, R.K. Potential strategies to control the risk of antifungal resistance in humans: A comprehensive review. Antibiotics 2023, 12, 608. [CrossRef]
- Cruz-Luna, A.R.; Cruz-Martínez, H.; Vásquez-López, A.; Medina, D.I. Metal nanoparticles as novel antifungal agents for sustainable agriculture: Current advances and future directions. J. Fungi 2021, 7, 1033. [CrossRef]
- Dananjaya, S.H.S.; Thao, N.T.; Wijerathna, H.M.S.M.; Lee, J.; Edussuriya, M.; Choi, D. Kumar, R.S. In vitro and in vivo anticandidal efficacy of green synthesized gold nanoparticles using Spirulina maxima polysaccharide. Process Biochem. 2020, 92, 138-148. [CrossRef]
- Abdelhamid, H.N.; Mahmoud, G.A.E.; Sharmouk, W. A cerium-based MOFzyme with multi-enzyme-like activity for the disruption and inhibition of fungal recolonization. J. Mater. Chem. B 2020, 8(33), 7548-7556. [CrossRef]
- Mukherjee, K.; Acharya, K.; Biswas, A.; Jana, N.R. TiO2 nanoparticles co-doped with nitrogen and fluorine as visible-light-activated antifungal agents. ACS Appl. Nano Mater. 2020, 3, 2016-2025. [CrossRef]
- Wen, H.; Shi, H.; Jiang, N.; Qiu, J.; Lin, F.; Kou, Y. Antifungal mechanisms of silver nanoparticles on mycotoxin producing rice false smut fungus. Iscience 2023, 26(1), 105763. [CrossRef]
- Malik, M.A.; Batterjee, M.G.; Kamli, M.R.; Alzahrani, K.A.; Danish, E.Y.; Nabi, A. Polyphenol-capped biogenic synthesis of noble metallic silver nanoparticles for antifungal activity against Candida auris. J. Fungi 2022, 8(6), 639. [CrossRef]
- Soleimani, P.; Mehrvar, A.; Michaud, J.P.; Vaez, N. Optimization of silver nanoparticle biosynthesis by entomopathogenic fungi and assays of their antimicrobial and antifungal properties. J. Invertebr. Pathol. 2022, 190, 107749. [CrossRef]
- Jamdagni, P.; Khatri, P.; Rana, J.S. Green synthesis of zinc oxide nanoparticles using flower extract of Nyctanthes arbor-tristis and their antifungal activity. J. King Saud Univ. Sci. 2018, 30(2), 168-175. [CrossRef]
- Jamdagni, P.; Rana, J.S.; Khatri, P.; Nehra, K. Comparative account of antifungal activity of green and chemically synthesized zinc oxide nanoparticles in combination with agricultural fungicides. Int. J. Nano Dimens. 2018, 9(2), 198-208.
- Zhou, L.; Zhao, X.; Li, M.; Lu, Y.; Ai, C.; Jiang, C.; Liu, Y.; Pan, Z.; Shi, J. Antifungal activity of silver nanoparticles synthesized by iturin against Candida albicans in vitro and in vivo. Appl. Microbiol. Biotechnol. 2021, 105(9), 3759-3770. [CrossRef]
- Shamraychuk, I.L.; Belyakova, G.A.; Eremina, I.M.; Kurakov, A.V.; Belozersky, M.A.; Dunaevsky, Y.E. Fungal proteolytic enzymes and their inhibitors as perspective biocides with antifungal action. Moscow Univ. Biol. Sci. Bull. 2020, 75, 97–103. [CrossRef]
- Padder, S.A.; Prasad, R.; Shah, A.H. Quorum sensing: A less known mode of communication among fungi. Microbiol. Res. 2018, 210, 51-58. [CrossRef]
- Baier, F.; Tokuriki, N. Connectivity between catalytic landscapes of the metallo-β-lactamase superfamily. J. Molec. Biol. 2014, 426 (13), 2442-2456. [CrossRef]
- Lyagin, I.; Efremenko, E. Enzymes for detoxification of various mycotoxins: Origins and mechanisms of catalytic action. Molecules 2019, 24, 2362. [CrossRef]
- Ayanwale, A.P.; Estrada-Capetillo, B.L.; Reyes-López, S.Y. Evaluation of antifungal activity by mixed oxide metallic nanocomposite against Candida spp. Processes 2021, 9, 773. [CrossRef]
- Bhatt, V.K.; Patel, M.; Pataniya, P.M.; Iyer, B.D.; Sumesh, C.K.; Late, D.J. Enhanced antifungal activity of WS2/ZnO nanohybrid against Candida albicans. ACS Biomater. Sci. Eng. 2020, 6(11), 6069-6075. [CrossRef]
- Abdelhamid, H.N.; Mahmoud, G.A.E. Antifungal and nanozyme activities of metal–organic framework-derived CuO@C. Appl. Organomet. Chem. 2023, 37(3), e7011. [CrossRef]
- Pillai, A.M.; Sivasankarapillai, V.S.; Rahdar, A.; Joseph, J.; Sadeghfar, F.; Rajesh, K.; Kyzas, G.Z. Green synthesis and characterization of zinc oxide nanoparticles with antibacterial and antifungal activity. J. Mol. Struct. 2020, 1211, 128107. [CrossRef]
- Arciniegas-Grijalba, P.A.; Patiño-Portela, M.C.; Mosquera-Sánchez, L.P.; Guerrero-Vargas, J.A.; Rodríguez-Páez, J.E. ZnO nanoparticles (ZnO-NPs) and their antifungal activity against coffee fungus Erythricium salmonicolor. Appl. Nanosci. 2017, 7, 225-241. [CrossRef]
- Ilkhechi, N.N.; Mozammel, M.; Khosroushahi, A.Y. Antifungal effects of ZnO, TiO2 and ZnO-TiO2 nanostructures on Aspergillus flavus. Pestic. Biochem. Phys. 2021, 176, 104869. [CrossRef]
- Miri, A.; Khatami, M.; Ebrahimy, O.; Sarani, M. Cytotoxic and antifungal studies of biosynthesized zinc oxide nanoparticles using extract of Prosopis farcta fruit. Green Chem. Lett. Rev. 2020, 13(1), 27-33. [CrossRef]
- Parveen, S.; Wani, A.H.; Shah, M.A.; Devi, H.S.; Bhat, M.Y.; Koka, J.A. Preparation, characterization and antifungal activity of iron oxide nanoparticles. Microb. Pathog. 2018, 115, 287-292. [CrossRef]
- Golipour, F.; Habibipour, R.; Moradihaghgou, L. Investigating effects of superparamagnetic iron oxide nanoparticles on Candida albicans biofilm formation. Med. Lab. J. 2019, 13(6), 44-50. [CrossRef]
- Bouson, S.; Krittayavathananon, A.; Phattharasupakun, N.; Siwayaprahm, P.; Sawangphruk, M. Antifungal activity of water-stable copper-containing metal-organic frameworks. R. Soc. Open Sci. 2017, 4(10), 170654. [CrossRef]
- Celis-Arias, V.; Loera-Serna, S.; Beltrán, H.I.; Álvarez-Zeferino, J.C.; Garrido, E.; Ruiz-Ramos, R. The fungicide effect of HKUST-1 on Aspergillus niger, Fusarium solani and Penicillium chrysogenum. New J. Chem. 2018, 42(7), 5570-5579. [CrossRef]
- Veerana, M.; Kim, H.C.; Mitra, S.; Adhikari, B.C.; Park, G.; Huh, S.; Kim, S.; Kim, Y. Analysis of the effects of Cu-MOFs on fungal cell inactivation. RSC Adv. 2021, 11(2), 1057-1065. [CrossRef]
- Tella, A.C.; Okoro, H.K.; Sokoya, S.O.; Adimula, V.O.; Olatunji, S.O.; Zvinowanda, C.; Ngila, J. C.; Shaibu, R.O.; Adeyemi, O.G. Synthesis, characterization and antifungal activity of Fe(III)metal–organic framework and its nano-composite. Chemistry Africa 2020, 3, 119-126. [CrossRef]
- Wei, F.; Cui, X.; Wang, Z.; Dong, C.; Li, J.; Han, X. Recoverable peroxidase-like Fe3O4@MoS2-Ag nanozyme with enhanced antibacterial ability. Chem. Eng. J. 2021, 408, 127240. [CrossRef]
- Liu, J.; Cheng, W.; Wang, Y.; Fan, X.; Shen, J.; Liu, H.; Wang, A.; Hui, A.; Nichols, F.; Chen, S. Cobalt-doped zinc oxide nanoparticle–MoS2 nanosheet composites as broad-spectrum bactericidal agents. ACS Appl. Nano Mater. 2021, 4, 4361-4370. [CrossRef]
- Chiericatti, C.; Basilico, J.C.; Basilico, M.L.Z.; Zamaro, J.M. Novel application of HKUST-1 metal–organic framework as antifungal: Biological tests and physicochemical characterizations. Microporous Mesoporous Mater. 2012, 162, 60-63. [CrossRef]
- Livesey, T.C.; Mahmoud, L.A.M.; Katsikogianni, M.G.; Nayak, S. Metal–organic frameworks and their biodegradable composites for controlled delivery of antimicrobial drugs. Pharmaceutics 2023, 15, 274. [CrossRef]
- Ji, Y.; Han, Z.; Ding, H.; Xu, X.; Wang, D.; Zhu, Y.; An, F.; Tang, S.; Zhang, H.; Deng, J.; Zhou, Q. Enhanced eradication of bacterial/fungi biofilms by glucose oxidase-modified magnetic nanoparticles as a potential treatment for persistent endodontic infections. ACS Appl. Mater. Interfaces 2021, 13, 17289-17299. [CrossRef]
- Wang, Q.; Jiang, J.; Gao, L. Catalytic antimicrobial therapy using nanozymes. Wiley Interdiscip. Rev.: Nanomed. Nanobiotechnol. 2022, 14(2), e1769. [CrossRef]
- Li, X.; Wu, X.M.; Yuan, T.; Zhu, J.R.; Yang, Y.L. Influence of the iodine content of nitrogen- and iodine-doped carbon dots as a peroxidase mimetic nanozyme exhibiting antifungal activity against C. albicans. Biochem. Eng. J. 2021, 175, 108139. https:// doi.org/10.1016/j.bej.2021.108139.
- Qingzhi, W.; Zou, S.; Wang, Q.; Chen, L.; Yan, X.; Gao, L. Catalytic defense against fungal pathogens using nanozymes. Nanotechnol. Rev. 2021, 10(1), 1277-1292. [CrossRef]
- Efremenko, E.N.; Lyagin, I.V.; Maslova, O.V.; Senko, O.V.; Stepanov, N.A.; Aslanli, A.G. Catalytic degradation of microplastics. Rus. Chem. Reviews 2023, 92(2), RCR5069. [CrossRef]
- Liang, M.; Yan, X. Nanozymes: From new concepts, mechanisms, and standards to applications. Acc. Chem. Res. 2019, 52(8), 2190-2200. [CrossRef]
- Priyadarshini, E.; Priyadarshini, S.S.; Cousins, B.G.; Pradhan, N. Metal-fungus interaction: Review on cellular processes underlying heavy metal detoxification and synthesis of metal nanoparticles. Chemosphere 2021, 274, 129976. [CrossRef]
- Jian, Y.; Chen, X.; Ahmed, T.; Shang, Q.; Zhang, S.; Ma, Z.; Yin, Y. Toxicity and action mechanisms of silver nanoparticles against the mycotoxin-producing fungus Fusarium graminearum. J. Adv. Res. 2022, 38, 1-12. [CrossRef]
- Khina, A.G.; Krutyakov, Y.A. Similarities and differences in the mechanism of antibacterial action of silver ions and nanoparticles. Appl. Biochem. Microbiol. 2021, 57, 683-693. [CrossRef]
- Yuan, H.; Li, J.; Pan, L.; Li, X.; Yuan, Y.; Zhong, Q.; Wu, X.; Luo, J.;Yang, S.T. Particulate toxicity of metal-organic framework UiO-66 to white rot fungus Phanerochaete chrysosporium. Ecotoxicol. Environ. Saf. 2022, 247, 114275. [CrossRef]
- Sholkamy, E.N.; Ahamd, M.S.; Yasser, M.M.; Eslam, N. Anti-microbiological activities of bio-synthesized silver Nano-stars by Saccharopolyspora hirsute. Saudi J. Biol. Sci. 2019, 26, 195–200. [CrossRef]
- Abd El-Ghany, M.N.; Hamdi, S.A.; Korany, S.M.; Elbaz, R.M.; Farahat, M.G. Biosynthesis of novel tellurium nanorods by Gayadomonas sp. TNPM15 isolated from mangrove sediments and assessment of their impact on spore germination and ultrastructure of phytopathogenic fungi. Microorganisms 2023, 11, e558. [CrossRef]
- Różalska, B.; Sadowska, B.; Budzyńska, A.; Bernat, P.; Różalska, S. Biogenic nanosilver synthesized in Metarhizium robertsii waste mycelium extract - As a modulator of Candida albicans morphogenesis, membrane lipidome and biofilm. PLoS ONE 2018, 13, e0194254. [CrossRef]
- Malik, M.A.; AlHarbi, L.; Nabi, A.; Alzahrani, K.A.; Narasimharao, K.; Kamli, M.R. Facile synthesis of magnetic nigella sativa seeds: Advances on nano-formulation approaches for delivering antioxidants and their antifungal activity against Candida albicans. Pharmaceutics 2023, 15, e642. [CrossRef]
- Akinola, P.O.; Lateef, A.; Asafa, T.B.; Beukes, L.S.; Abbas, S.H.; Irshad, H.M. Phytofabrication of titanium-silver alloy nanoparticles (Ti-AgNPs) by Cola nitida for biomedical and catalytic applications. Inorg. Chem. Commun. 2022, 139, e109357. [CrossRef]
- Li, J.; Sun, Q.; Sun, Y.; Chen, B.; Wu, X.; Le, T. Improvement of banana postharvest quality using a novel soybean protein isolate/cinnamaldehyde/zinc oxide bionanocomposite coating strategy. Sci. Hortic. 2019, 258, e108786. [CrossRef]
- Yuan, Y.; Tian, H.; Huang, R.; Liu, H.; Wu, H.; Guo, G.; Xiao, J. Fabrication and characterization of natural polyphenol and ZnO nanoparticles loaded protein-based biopolymer multifunction electrospun nanofiber films, and application in fruit preservation. Food Chem. 2023, 418, e135851. [CrossRef]
- Zhu, X.; Ma, X.; Gao, C.; Mu, Y.; Pei, Y.; Liu, C.; Zou, A.; Sun, X. Fabrication of CuO nanoparticles composite ε-polylysine-alginate nanogel for high-efficiency management of Alternaria alternate. Int. J. Biol. Macromol. 2022, 223(Pt A), 1208–1222. [CrossRef]
- Fernandes, L.; Ribeiro, R.; Henriques, M.; Rodrigues, M.E. Candida auris, a singular emergent pathogenic yeast: Its resistance and new therapeutic alternatives. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 41, 1371–1385. [CrossRef]
- Mourer, T.; El Ghalid, M.; d'Enfert, C.; Bachellier-Bassi, S. Involvement of amyloid proteins in the formation of biofilms in the pathogenic yeast Candida albicans. Res. Microbiol. 2021, 172, e103813. [CrossRef]
- Liu, Y.; Filler, S.G. Candida albicans Als3, a multifunctional adhesin and invasion. Eukaryot. Cell 2011, 10, 168–173. [CrossRef]
- Zheng, S.; Chang, W.; Li, C.; Lou, H. Als1 and Als3 regulate the intracellular uptake of copper ions when Candida albicans biofilms are exposed to metallic copper surfaces. FEMS Yeast Res. 2016, 16, fow029. [CrossRef]
- Lee, J.-H.; Kim, Y.-G.; Khadke, S.K.; Lee, J. Antibiofilm and antifungal activities of medium-chain fatty acids against Candida albicans via mimicking of the quorum-sensing molecule farnesol. Microb. Biotechnol. 2021, 14, 1353–1366. [CrossRef]
- Sephton-Clark, P.C.S.; Voelz, K. Spore germination of pathogenic filamentous fungi. Adv. Appl. Microbiol. 2018, 102, 117–157. [CrossRef]
- Slavin, Y.N.; Bach, H. Mechanisms of antifungal properties of metal nanoparticles. Nanomaterials 2022, 12, 4470. [CrossRef]
- Efremenko, E.; Senko, O.; Stepanov, N.; Maslova, O.; Lomakina, G.Y.; Ugarova, N. Luminescent analysis of ATP: Modern objects and processes for sensing. Chemosensors 2022, 10, 493. [CrossRef]
- Ambati, S.; Ferarro, A.R.; Kang, S.E.; Lin, J.; Lin, X.; Momany, M.; Lewis, Z.A.; Meagher, R.B. Dectin-1-targeted antifungal liposomes exhibit enhanced efficacy. mSphere 2019, 4, e00025-19. [CrossRef]
- Gow, N.A.R.; Latge, J.P.; Munro, C.A. The fungal cell wall: Structure, biosynthesis, and function. Microbiol. Spectr. 2017, 5, FUNK-0035-2016. [CrossRef]
- Kühbacher, A.; Burger-Kentischer, A.; Rupp, S. Interaction of Candida species with the skin. Microorganisms 2017, 5, 32. [CrossRef]
- Lyagin, I.; Stepanov, N.; Maslova, O.; Senko, O.; Aslanli, A.; Efremenko E. Not a mistake but a feature: Promiscuous activity of enzymes meeting mycotoxins. Catalysts 2022, 12, 1095. [CrossRef]
- Li, C.; Li, X.; Bai, C.; Zhang, Y.; Wang, Z. A chitinase with antifungal activity from naked oat (Avena chinensis) seeds. J. Food Biochem. 2019; 43, e12713. [CrossRef]
- Dikbaş, N.; Uçar, S.; Tozlu, E.; Kotan, M.S.; ·Kotan, R. Antifungal activity of partially purified bacterial chitinase against Alternaria alternata. Erwerbs-Obstbau 2022. [CrossRef]
- Zhang, W.; Ma, J.; Yan, Q.; Jiang,Z.; Yang, S. Biochemical characterization of a novel acidic chitinase with antifungal activity from Paenibacillus xylanexedens Z2–4. Int. J. Biol. Macromol. 2021, 182, 1528-1536. [CrossRef]
- Rajninec, M.; Jopcik, M.; Danchenko, M.; Libantova, J. Biochemical and antifungal characteristics of recombinant class I chitinase from Drosera rotundifolia. Int. J. Biol. Macromol. 2020, 161, 854–863. [CrossRef]
- Wang, N-N.; Gao, K-Y.; Han, N.; Tian, R-Z.; Zhang, J-L.; Yan, X.; Huang, L-L. ChbB increases antifungal activity of Bacillus amyloliquefaciens against Valsa mali and shows synergistic action with bacterial chitinases. Biol. Control. 2020, 142, 104150. [CrossRef]
- Li, Q.; Hou, Z.; Zhou, D.; Jia, M.; Lu, S.; Yu, J. Antifungal activity and possible mechanism of Bacillus amyloliquefaciens FX2 against the postharvest apple ring rot pathogen. Phytopathology 2022, 112, 2486-2494. [CrossRef]
- Lu, Y.; Wang, N.; He, J.; Li, Y.; Gao, X.; Huang, L.; Yan, X. Expression and characterization of a novel chitinase with antifungal activity from a rare actinomycete Saccharothrix yanglingensis Hhs.015. Protein Expr. Purif. 2018, 143, 45–51. [CrossRef]
- Brzezinska, M.S.; Jankiewicz, U.; Kalwasinska, A.; Swiatczak, J.; Zero, K. Characterization of chitinase from Streptomyces luridiscabiei U05 and its antagonist potential against fungal plant pathogens. J. Phytopathol. 2019, 167, 404–412. [CrossRef]
- Le, B.; Yang, S.H. Characterization of a chitinase from Salinivibrio sp. BAO-1801 as an antifungal activity and a biocatalyst for producing chitobiose. J. Basic Microbiol. 2018, 58, 848–856. [CrossRef]
- Li, Z.; Xia, C.; Wang, Y.; Li, X.; Qiao, Y.; Li, C.; Zhou, J.; Zhang, L.; Ye, X.; Huang, Y.; Cui, Z. Identification of an endo-chitinase from Corallococcus sp. EGB and evaluation of its antifungal properties. Int. J. Biol. Macromol. 2019, 132, 1235–1243. [CrossRef]
- Moon, C.; Seo, D.J.; Song, Y.S.; Hong, S.H.; Choi, S.H.; Jung, W.J. Antifungal activity and patterns of N-acetyl-chitooligosaccharide degradation via chitinase produced from Serratia marcescens PRNK-1. Microb. Pathog. 2017, 113, 218–224. [CrossRef]
- Deng, J.-J.; Shi, D.; Mao, H.-H.; Li, Z.-W.; Liang, S.; Ke, Y.; Luo, X.-C. Heterologous expression and characterization of an antifungal chitinase (Chit46) from Trichoderma harzianum GIM 3.442 and its application in colloidal chitin conversion. Int. J. Biol. Macromol. 2019, 134, 113-121. [CrossRef]
- Yilmaz, G.; Cadirci, B. Comparison of in vitro antifungal activity methods using Aeromonas sp. BHC02 chitinase, whose physicochemical properties were determined as antifungal agent candidate. Res. Sq. 2022. [CrossRef]
- Wang, Y.; Li, D.; Xia, C.; Fan, Q.; Li, X.; Lan, Z.; Shi, G.; Dong, W.; Li, Z.; Cui, Z. Preparation of the active chitooligosaccharides with a novel chitosanase AqCoA and their application in fungal disease protection. J. Agric. Food Chem. 2021, 69, 3351−3361. [CrossRef]
- Zhang, W.; Liu, Y.; Ma, J.; Yan, Q.; Jiang, Z.; Yang, S. Biochemical characterization of a bifunctional chitinase/lysozyme from Streptomyces sampsonii suitable for N-acetyl chitobiose production. Biotechnol. Lett. 2020, 42, 1489–1499. [CrossRef]
- Li, S.; Zhang, B.; Zhu, H.; Zhu, T. Cloning and expression of the chitinase encoded by ChiKJ406136 from Streptomyces sampsonii (Millard & Burr) Waksman KJ40 and its antifungal effect. Forests 2018, 9, 699. [CrossRef]
- Bamford, N.C.; Le Mauff, F.; Subramanian, A.S.; Yip, P.; Millán, C.; Zhang, Y.; Zacharias, C.; Forman, A.; Nitz, M.; Codée, J.D.C.; Usón, I.; Sheppard, D.C.; Howell, P.L. Ega3 from the fungal pathogen Aspergillus fumigatus is an endo-α-1,4-galactosaminidase that disrupts microbial biofilms. J. Biol. Chem. 2019, 294(37), 3833-13849. [CrossRef]
- Ostapska, H.; Raju, D.; Lehoux, M.; Lacdao, I.; Gilbert, S.; Sivarajah, P.; Bamford, N.C.; Baker, P.; Nguyen, T.T.M.; Zacharias, C.A.; Gravelat, F.N.; Howell, P.L.; Sheppard, D.C. Preclinical evaluation of recombinant microbial glycoside hydrolases in the prevention of experimental invasive aspergillosis. mBio 2021, 12, e02446-21. [CrossRef]
- Vidhate, R.P.; Bhide, A.J.; Gaikwad, S.M.; Giri, A.P. A potent chitin-hydrolyzing enzyme from Myrothecium verrucaria affects growth and development of Helicoverpa armigera and plant fungal pathogens. Int. J. Biol. Macromol. 2019, 141, 517-528. [CrossRef]
- Zhang, L.; Li, W.; Tao, Y.; Zhao, S.; Yao, L.; Cai, Y.; Niu, Q. Overexpression of the key virulencef 1,3-1,4-β-d-glucanase in the endophytic bacterium Bacillus halotolerans Y6 to improve Verticillium resistance in cotton. J. Agric. Food Chem. 2019, 67(24), 6828-6836. https:// doi.org/10.1021/acs.jafc.9b00728.
- Ling, L.; Cheng, W.; Jiang, K.; Jiao, Z.; Luo, H.; Yang, C.; Pang, M.; Lu. L. The antifungal activity of a serine protease and the enzyme production of characteristics of Bacillus licheniformis TG116. Arch. Microbiol. 2022, 204, 601. [CrossRef]
- Deng, J.-J.; Huang, W.Q.; Li, Z.-W.; Lu, D.-L.; Zhang, Y.; Luo, X.-C. Biocontrol activity of recombinant aspartic protease from Trichoderma harzianum against pathogenic fungi. Enzyme Microb. Technol. 2018, 112, 35-42. [CrossRef]
- Wang, A.; Paul, J.; Weldrick, P.J.; Madden, L.A.; Paunov, V.N. Enhanced clearing of Candida biofilms on a 3D urothelial cell in vitro model using lysozyme-functionalized fluconazole-loaded shellac nanoparticles. Biomater. Sci. 2021, 9, 6927–6939. [CrossRef]
- Hernández-Téllez, C.N.; Rodríguez-Córdova, F.J.; Rosas-Burgos, E.C.; Cortez-Rocha, M.O.; Burgos-Hernández, A.; Lizardi-Mendoza, J.; Torres-Arreola, W.; Martínez-Higuera, A.; Plascencia-Jatomea, M. Activity of chitosan-lysozyme nanoparticles on the growth, membrane integrity, and β-1,3-glucanase production by Aspergillus parasiticus. 3 Biotech 2017, 7, e279. [CrossRef]
- Silva, F.A.; Albuquerque, L.M.; Martins, T.F.; de Freitas, J.A.; Vasconcelos I.M.; de Freitas,D. Q.; Moreno, F.B.M.B.; Monteiro-Moreira, A.C.O.; Oliveira, J. T.A. A peroxidase purified from cowpea roots possesses high thermal stability and displays antifungal activity against Colletotrichum gloeosporioides and Fusarium oxysporum. Biocatal. Agric. Biotechnol. 2022, 42, 102322. [CrossRef]
- Zhang, L.; Tao, Y.; Zhao, S. A novel peroxiredoxin from the antagonistic endophytic bacterium Enterobacter sp. V1 contributes to cotton resistance against Verticillium dahliae. Plant Soil 2020, 454, 395–409. [CrossRef]
- Salazar, V.A.; Arranz-Trullén, J.; Navarro, S.; Blanco, J.A.; Sánchez, D.; Moussaoui, M.; Boix, E. Exploring the mechanisms of action of human secretory RNase 3 and RNase 7 against Candida albicans. MicrobiologyOpen 2016, 5(5), 830-845. [CrossRef]
- Salazar, V.A.; Arranz-Trullén, J.; Prats-Ejarque, G.; Torrent, M.; Andreu, D.; Pulido, D.; Boix, E. Insight into the antifungal mechanism of action of human RNase N-terminus derived peptides. Int. J. Mol. Sci. 2019, 20, 4558. [CrossRef]
- Kosgey, J.C.; Jia, L.; Nyamao, R.M.; Zhao, Y.; Xue, T.; Yang, J.; Fang, Y.; Zhang, F. RNase 1, 2, 5 & 8 role in innate immunity: Strain specific antimicrobial activity. Int. J. Biol. Macromol. 2020, 160, 1042-1049. [CrossRef]
- Philip, N.V.; Koteshwara, A.; Kiran, G.A.; Raja, S.; Subrahmanyam, V.M.; Chandrashekar, H.R. Statistical optimization for coproduction of chitinase and beta 1,4-endoglucanase by chitinolytic Paenibacillus elgii PB1 having antifungal activity. Appl. Biochem. Biotechnol. 2020, 191(1), 135-150. [CrossRef]
- Sinitsyna, O.A.; Rubtsova, E.A.; Sinelnikov, I.G.; Osipov, D. O.; Rozhkova, A. M.; Matys, V. Yu.; Bubnova, T.V.; Nemashkalov, V.A.; Sereda, A. S.; Tcsherbakova, L. A.; Sinitsyn, A.P. Creation of chitinase producer and disruption of micromycete cell wall with the obtained enzyme preparation. Biochem. (Mosc.) 2020, 85, 717–724. [CrossRef]
- Sachivkina, N.; Lenchenko, E.; Blumenkrants, D.; Ibragimova, A.; Bazarkina, O. Effects of farnesol and lyticase on the formation of Candida albicans biofilm. Vet. World 2020, 13(6), 1030-1036. [CrossRef]
- Tan, Y.; Ma, S.; Leonhard, M.; Moser, D.; Ludwig, R.; Schneider-Stickler, B. Co-immobilization of cellobiose dehydrogenase and deoxyribonuclease I on chitosan nanoparticles against fungal/bacterial polymicrobial biofilms targeting both biofilm matrix and microorganisms. Mater Sci. Eng. C 2020, 108, 110499. [CrossRef]
- Oyeleye, A.; Normi, Y.M. Chitinase: Diversity, limitations, and trends in engineering for suitable applications. Biosci. Rep. 2018, 38, BSR2018032300. [CrossRef]
- Pham, C.L.L.; de Francisco, B.R.; Valsecchi, I.; Dazzoni, R.; Pillé, A.; Lo, V.; Ball, S.R.; Cappai, R.; Wien, F.; Kwan, A.H.; Guijarro, J.I.; Sunde, M. Probing structural changes during self-assembly of surface-active hydrophobin proteins that form functional amyloids in fungi. J. Mol. Biol. 2018, 430, 3784–3801. [CrossRef]
- Valsecchi, I.; Dupres, V.; Stephen-Victor, E.; Guijarro, J.I.; Gibbons, J.; Beau, R.; Bayry, J.; Coppee, J.-Y.; Lafont, F.; Latgé, J. P.; Beauvais, A. Role of hydrophobins in Aspergillus fumigatus. J. Fungi 2017, 4, 2. [CrossRef]
- Valsecchi, I.; Lai, J.I.; Stephen-Victor, E.; Pillé, A.; Beaussart, A.; Lo, V.; Pham, C.L.L.; Aimanianda, V.; Kwan, A.H.; Duchateau, M.; Gianetto, Q.G.; Matondo, M.; Lehoux, M.; Sheppard, D.C.; Dufrene, Y.F.; Bayry, J.; Guijarro, J.I.; Sunde, M.; Latgé, J. P. Assembly and disassembly of Aspergillus fumigatus conidial rodlets. Cell Surf. 2019, 5, 100023. [CrossRef]
- Pham, C.L.L.; Rey, A.; Lo, V.; Soulès, M.; Ren, Q.; Meisl, G.; Knowles, T.P.J.; Kwan, A.H.; Sunde, M. Self-assembly of MPG1, a hydrophobin protein from the rice blast fungus that forms functional amyloid coatings, occurs by a surface-driven mechanism. Sci. Rep. 2016, 6. [CrossRef]
- Saad, S.; Cereghetti, G.; Feng, Y.; Picotti, P.; Peter, M.; Dechant, R. Reversible protein aggregation is a protective mechanism to ensure cell cycle restart after stress. Nat. Cell Biol. 2017, 19, 1202e13. [CrossRef]
- Beaussart, A.; Alsteens, D.; El-Kirat-Chatel, S.; Lipke, P.N.; Kucharíkova, S.; Dijck, P.V.; Dufrene, Y.F. Single-molecule imaging and functional analysis of Als adhesins and mannans during Candida albicans morphogenesis. ACS Nano 2012, 6 (12), 10950–10964. [CrossRef]
- Ho, V.; Herman-Bausier, P.; Shaw, C.; Conrad, K.A.; Garcia-Sherman, M.C.; Draghi, J.; Dufrene, Y.F.; Lipke, P.N.; Rauceo, J.M. An amyloid core sequence in the major Candida albicans adhesin Als1p mediates cell-cell adhesion. mBio 2019, 10. [CrossRef]
- Kumar, R.; Breindel, C.; Saraswat, D.; Cullen, P.J.; Edgerton, M. Candida albicans Sap6 amyloid regions function in cellular aggregation and zinc binding, and contribute to zinc acquisition. Sci. Rep. 2017, 7, 2908. [CrossRef]
- Monniot, C.; Boisrame, A.; Costa, G.D.; Chauvel, M.; Sautour, M.; Bougnoux, M-E.; Bellon-Fontaine, M.-N.; Dalle, F.; d’Enfert, C.; Richard, M.L. Rbt1 protein domains analysis in Candida albicans brings insights into hyphal surface modifications and Rbt1 potential role during adhesion and biofilm formation. PLoS ONE 2013, 8, e82395. [CrossRef]
- Cabral, V.; Znaidi, S.; Walker, L.A.; Martin-Yken, H.; Dague, E.; Legrand, M.; Lee, K.; Chauvel, M.; Firon, A.; Rossignol, T.; Munro, A.C.; Bachelier-Bassi, S.; d’Enfert, C. Targeted changes of the cell wall proteome influence Candida albicans ability to form single- and multi-strain biofilms. PLoS Pathog. 2014, 10, e1004542. [CrossRef]
- Moreno-Ruiz, E.; Ortu, G.; de Groot, P.W.J.; Cottier, F.; Loussert, C.; Prevost, M-C.; de Koster, C.; Klis, M.F., Goyard, S.; d’Enfert, C. The GPI-modified proteins Pga59 and Pga62 of Candida albicans are required for cell wall integrity. Microbiology 2009, 155, 2004e20. [CrossRef]
- Shanmugam, N.; Baker, M.O.; Ball, S.R.; Steain, M.; Pham, C.L.; Sunde, M. Microbial functional amyloids serve diverse purposes for structure, adhesion and defence. Biophys. Rev. 2019, 11, 287-302. [CrossRef]
- Chernova, T.A.; Chernoff, Y.O.; Wilkinson, K.D. Yeast models for amyloids and prions: Environmental modulation and drug discovery. Molecules 2019, 24, 3388. [CrossRef]
- Hirata, A.; Hori, Y.; Koga, Y.; Okada, J.; Sakudo, A.; Ikuta, K.; Kanaya, S.; Takano, K. Enzymatic activity of a subtilisin homolog, Tk-SP, from Thermococcus kodakarensis in detergents and its ability to degrade the abnormal prion protein. BMC Biotech. 2013, 13, 1-7. [CrossRef]
- Dabbagh, F.; Negahdaripour, M.; Berenjian, A.; Behfar, A.; Mohammadi, F.; Zamani, M.; Irajie, C.; Ghasemi, Y. Nattokinase: Production and application. Appl. Microbiol. Biotechnol. 2014, 98, 9199–9206. [CrossRef]
- Pilon, J.L.; Nash, P.B.; Arver, T.; Hoglund, D.; VerCauteren, K.C. Feasibility of infectious prion digestion using mild conditions and commercial subtilisin. J. Virol. Methods. 2009, 161, 168–172. [CrossRef]
- Saunders, S.E.; Bartz, J.C.; Vercauteren, K.C.; Bartelt-Hunt, S.L. Enzymatic digestion of chronic wasting disease prions bound to soil. Environ. Sci. Technol. 2010, 44, 4129–4135. [CrossRef]
- McLeod, A.H.; Murdoch, H.; Dickinson, J.; Dennis, M.J.; Hall, G.A.; Buswell, C.M.; Carr, J.; Taylor, D.M.; Sutton, J.M.; Raven, N.D.H. Proteolytic inactivation of the bovine spongiform encephalopathy agent. Biochem. Biophys. Res. Commun. 2004, 317, 1165e1170. [CrossRef]
- Dickinson, J.; Murdoch, H.; Dennis, M.J.; Hall, G.A.; Bott, R.; Crabb, W.D.; Penet, C.; Sutton, J.M.; Raven, N.D.H. Decontamination of prion protein (BSE301V) using a genetically engineered protease. J. Hosp. Infect. 2009, 72, 65e70. [CrossRef]
- Yoshioka, M.; Miwa, T.; Horii, H.; Takata, M.; Yokoyama, T.; Nishizawa, K.; Watanabe, M.; Shinagawa, M.; Murayama, Y. Characterization of a proteolytic enzyme derived from a Bacillus strain that effectively degrades prion protein. J. Appl. Microbiol. 2007, 102, 509e515. [CrossRef]
- Hui, Z.; Doi, H.; Kanouchi, H.; Matsuura, Y.; Mohri, S.; Nonomura, Y.; Oka, T. Alkaline serine protease produced by Streptomyces sp. degrades PrPSc. Biochem. Biophys. Res. Commun. 2004, 321, 45e50. [CrossRef]
- Bahun, M.; Šnajder, M.; Turk, D.; Poklar Ulrih, N. Insights into the maturation of pernisine, a subtilisin-like protease from the hyperthermophilic archaeon Aeropyrum pernix. Appl. Environ. Microbiol. 2020, 86, e00971-20. [CrossRef]
- Johnson, C.J.; Bennett, J.P.; Biro, S.M.; Duque-Velasquez, J.C.; Rodriguez, C.M.; Bessen, R.A.; Rocke, T.E. Degradation of the disease-associated prion protein by a serine protease from lichens. PLoS ONE. 2011, 11, e19836. [CrossRef]
- Chen, C.Y.; Rojanatavorn, K.; Clark, A.C.; Shih, J.C. Characterization and enzymatic degradation of Sup35NM, a yeast prion-like protein. Prot. Sci. 2005, 14, 2228-2235. [CrossRef]
- Wang, J.J.; Borwornpinyo, R.; Odetallah, N.; Shih, J.C. Enzymatic degradation of a prion-like protein, Sup35NM-His6. Enzyme Microb. Technol. 2005, 36, 758-765. [CrossRef]
- Sharma, R.; Gupta, R. Coupled action of γ-glutamyl transpeptidase-glutathione and keratinase effectively degrades feather keratin and surrogate prion protein, Sup 35NM. Biores. Tech. 2012, 120, 314-317. [CrossRef]
- Rajput, R.; Gupta, R. Thermostable keratinase from Bacillus pumilus KS12: Production, chitin crosslinking and degradation of Sup35NM aggregates. Biores. Tech. 2013, 133, 118-126. [CrossRef]
- Ningthoujam, D.S.; Mukherjee, S., Devi, L.J.; Singh, E.S.; Tamreihao, K.; Khunjamayum, R.; Banerjee, S.; Mukhopadhyay, D. In vitro degradation of β-amyloid fibrils by microbial keratinase. Alzheimers Dement. 2019, 5, 154-163. [CrossRef]
- Kim, N.; Lee, H.J. Redox-active metal ions and amyloid-degrading enzymes in Alzheimer’s disease. Int. J. Mol. Sci. 2021, 22, 7697. [CrossRef]
- Manikandan, P.; Moopantakath, J.; Imchen, M.; Kumavath, R.; SenthilKumar, P. K. Identification of multi-potent protein subtilisin A from halophilic bacterium Bacillus firmus VE2. Microb. Pathog. 2021, 157, 105007. [CrossRef]
- Kokwe, L.; Nnolim, N. E.; Ezeogu, L.I.; Sithole, B.; Nwodo, U.U. Thermoactive metallo-keratinase from Bacillus sp. NFH5: Characterization, structural elucidation, and potential application as detergent additive. Heliyon. 2023, 9. [CrossRef]
- Efremenko, E.; Aslanli, A.; Lyagin, I. Advanced situation with recombinant toxins: Diversity, production and application purposes. Int. J. Mol. Sci. 2023, 24, 4630. [CrossRef]
- Efremenko, E.; Senko, O.; Stepanov, N.; Aslanli, A.; Maslova, O.; Lyagin, I. Quorum sensing as a trigger that improves characteristics of microbial biocatalysts. Microorganisms 2023, 11, 1395. [CrossRef]
- Willaert, R.G. Adhesins of yeasts: Protein structure and interactions. J. Fungi 2018, 4, 119. [CrossRef]
- Tian, X.; Ding, H.; Ke,W.; Wang, L. Quorum sensing in fungal species. Annu. Rev. Microbiol. 2021, 75, 449–469. [CrossRef]
- Mehmood, A.; Liu, G.;Wang, X.; Meng, G.;Wang, C.; Liu, Y. Fungal quorum-sensing molecules and inhibitors with potential antifungal activity: A review. Molecules 2019, 24, 1950. [CrossRef]
- Lee, K.; Lee, S.; Lee, S.H.; Kim, S.R.; Oh, H.S.; Park, P.K.; Choo, K.H.; Kim, Y.W.; Lee, J.K.; Lee, C.H. Fungal quorum quenching: A paradigm shift for energy savings in membrane bioreactor (MBR) for wastewater treatment. Environ. Sci. Technol. 2016, 50(20), 10914-10922. [CrossRef]
- Ogawa, K.; Nakajima-Kambe, T.; Nakahara, T.; Kokufuta, E. Coimmobilization of gluconolactonase with glucose oxidase for improvement in kinetic property of enzymatically induced volume collapse in ionic gels. Biomacromolecules 2002; 3(3), 625-631. [CrossRef]
- Aslanli, A.; Domnin, M.; Stepanov, N.; Efremenko, E. “Universal” antimicrobial combination of bacitracin and His6-OPH with lactonase activity, acting against various bacterial and yeast cells. Int. J. Mol. Sci. 2022, 23, 9400. [CrossRef]
- Aslanli, A.; Domnin, M.; Stepanov, N.; Efremenko, E. Synergistic antimicrobial action of lactoferrin-derived peptides and quorum quenching enzymes. Int. J. Mol. Sci. 2023, 24, 3566. [CrossRef]
- Hogan, D. Talking to themselves: Autoregulation and quorum sensing in fungi. Eukaryoitic Cell 2006, 5(4), 613–619. [CrossRef]
- Bu'LocK, J.D.; Jones, B.E.; Winskill, N. The apocarotenoid system of sex hormones and prohormones in mucorales. Pure AppL. Chem. 1976, 47, 191-202. [CrossRef]
- Alves, D.R.; de Morais, S.M.; Tomiotto-Pellissier, F.; Vasconcelos, F.R.; Francisco das Chagas Oliveira Freire, F.C.O.; da Silva, I.N.G.; Cataneo, A.H.D.; Miranda-Sapla, M.M.; Gustavo Adolfo Saavedra Pinto, G.A.S.; Conchon-Costa, I.; Arlindo de Alencar Araripe Noronha, A.A.A.; Pavanelli, W.R. Leishmanicidal and fungicidal activity of lipases obtained from endophytic fungi extracts. PLoS ONE 2018, 13(6), e0196796. [CrossRef]
- Singh, A.; Poeta, M. Lipid signalling in pathogenic fungi. Cell. Microbiol. 2011, 13, 177–185. [CrossRef]
- Beccaccioli,M.; Reverberi,M.; Scala, V. Fungal lipids: Biosynthesis and signalling during plant-pathogen interaction. Front. Biosci. 2019, 24(1), 168–181. [CrossRef]
- Frolov, G.; Lyagin, I.; Senko, O.; Stepanov, N.; Pogorelsky, I.; Efremenko, E. Metal nanoparticles for improving bactericide functionality of usual fibers. Nanomaterials 2020, 10, 1724. [CrossRef]
- Lyagin, I.; Stepanov, N.; Frolov, G.; Efremenko, E. Combined modification of fiber materials by enzymes and metal nanoparticles for chemical and biological protection. Int. J. Mol. Sci. 2022, 23, 1359. [CrossRef]
- Lyagin, I.; Maslova, O.; Stepanov, N.; Presnov, D.; Efremenko, E. Assessment of composite with fibers as a support for antibacterial nanomaterials: A case study of bacterial cellulose, polylactide and usual textile. Fibers 2022, 10, 70. [CrossRef]
- Lyagin, I.; Maslova, O.; Stepanov, N.; Efremenko, E. Degradation of mycotoxins in mixtures by combined proteinous nanobiocatalysts: In silico, in vitro and in vivo. Int. J. Biol. Macromol. 2022, 218, 866–877. [CrossRef]
- Garces, F.; Fernández, F.J.; Montellà, C.; Penya-Soler, Prohens, E.R.; Aguilar, J.; Baldomà, L.; Coll, M.; Badia, J.; Vega, M.C. Molecular architecture of the Mn2+-dependent lactonase UlaG reveals an RNase-like metallo-β-lactamase fold and a novel quaternary structure. J. Mol. Biol. 2010, 39(5), 715-729. [CrossRef]
- González, J.M. Visualizing the superfamily of metallo-β-lactamases through sequence similarity network neighborhood connectivity analysis. Heliyon 2021, 7(1), e05867. [CrossRef]
- Jang, E.-Y.; Son, Y.-J.; Park, S.-Y.; Yoo, J.-Y.; Cho, Y.-N.; Jeong, S.-Y.; Liu, S.; Son, H.-J. Improved biosynthesis of silver nanoparticles using keratinase from Stenotrophomonas maltophilia R13: Reaction optimization, structural characterization, and biomedical activity. Bioprocess Biosyst. Eng. 2018, 41, 381–393. [CrossRef]
- Abo-Zaid, G.; Abdelkhalek, A.; Matar, S.; Darwish, M.; Abdel-Gayed, M. Application of bio-friendly formulations of chitinase-producing Streptomyces cellulosae Actino 48 for controlling peanut soil-borne diseases caused by Sclerotium rolfsii. J. Fungi 2021, 7, e167. [CrossRef]
- Gaonkar, S.K.; Furtado, I.J. Biorefinery-fermentation of agro-wastes by Haloferax lucentensis GUBF-2 MG076878 to haloextremozymes for use as biofertilizer and biosynthesizer of AgNPs. Waste Biomass Valori. 2022, 13, 1117–1133. [CrossRef]
- Guilger-Casagrande, M.; Germano-Costa, T.; Pasquoto-Stigliani, T.; Fraceto, L.F.; de Lima, R. Biosynthesis of silver nanoparticles employing Trichoderma harzianum with enzymatic stimulation for the control of Sclerotinia sclerotiorum. Sci. Rep. 2019, 9, e14351. [CrossRef]
- Guilger-Casagrande, M.; Germano-Costa, T.; Bilesky-José, N.; Pasquoto-Stigliani, T.; Carvalho, L.; Fraceto, L.F.; de Lima, R. Influence of the capping of biogenic silver nanoparticles on their toxicity and mechanism of action towards Sclerotinia sclerotiorum. J. Nanobiotechnology 2021, 19, e53. [CrossRef]
- Dror, Y.; Ophir, C.; Freeman, A. Silver-enzyme hybrids as wide-spectrum antimicrobial agents. Innovations and merging technologies in wound care (Amit Gefen, Ed.). Elsevier Inc. 2020, 293–307. [CrossRef]
- Yu, D.; Wang, Y.; Zhang, J.; Yu, Q.; Liu, S.; Li, M. Synthesis of the ternary nanocomposites composed of zinc 2-methylimidazolate frameworks, lactoferrin and melittin for antifungal therapy. J. Mater. Sci. 2022, 57, 16809–16819. [CrossRef]
- Xia, S.; Xu, Y.; Hoy, R.; Zhang, J.; Qin, L.; Li, X. The notorious soilborne pathogenic fungus Sclerotinia sclerotiorum: An update on genes studied with mutant analysis. Pathogens 2020, 9, e27. [CrossRef]
- Chet, I.; Henis, Y. Effect of catechol and disodium EDTA on melanin content of hyphal and sclerotial walls of Sclerotium rolfsii sacc. and the role of melanin in the susceptibility of these walls to β-(1→3) glucanase and chitinase. Soil Biol. Biochem. 1969, 1, 131–138. [CrossRef]
- Melo, B.S.; Voltan, A.R.; Arruda, W.; Lopes, F.A.C.; Georg, R.C.; Ulhoa, C.J. Morphological and molecular aspects of sclerotial development in the phytopathogenic fungus Sclerotinia sclerotiorum. Microbiol. Res. 2019, 229, e126326. [CrossRef]
- Babina, S.E.; Kanyshkova, T.G.; Buneva, V.N.; Nevinsky, G.A. Lactoferrin is the major deoxyribonuclease of human milk. Biochemistry (Mosc.) 2004, 69, 1006–1015. [CrossRef]
- Fernandes, K.E.; Weeks, K.; Carter, D.A. Lactoferrin is broadly active against yeasts and highly synergistic with amphotericin B. Antimicrob. Agents Chemother. 2020, 64, e02284-19. [CrossRef]
- Espinel-Ingroff, A. Novel antifungal agents, targets or therapeutic strategies for the treatment of invasive fungal diseases: A review of the literature (2005-2009). Rev. Iberoam. Micol. 2009, 26(1), 15-22. [CrossRef]
- Ivanov, M.; Ćirić, A.; Stojković, D. Emerging antifungal targets and strategies. Int. J. Mol. Sci. 2022, 23, 2756. [CrossRef]
Figure 1.
Enzymes with antifungal activities due to their catalytic action on different targets as substrates.
Figure 1.
Enzymes with antifungal activities due to their catalytic action on different targets as substrates.
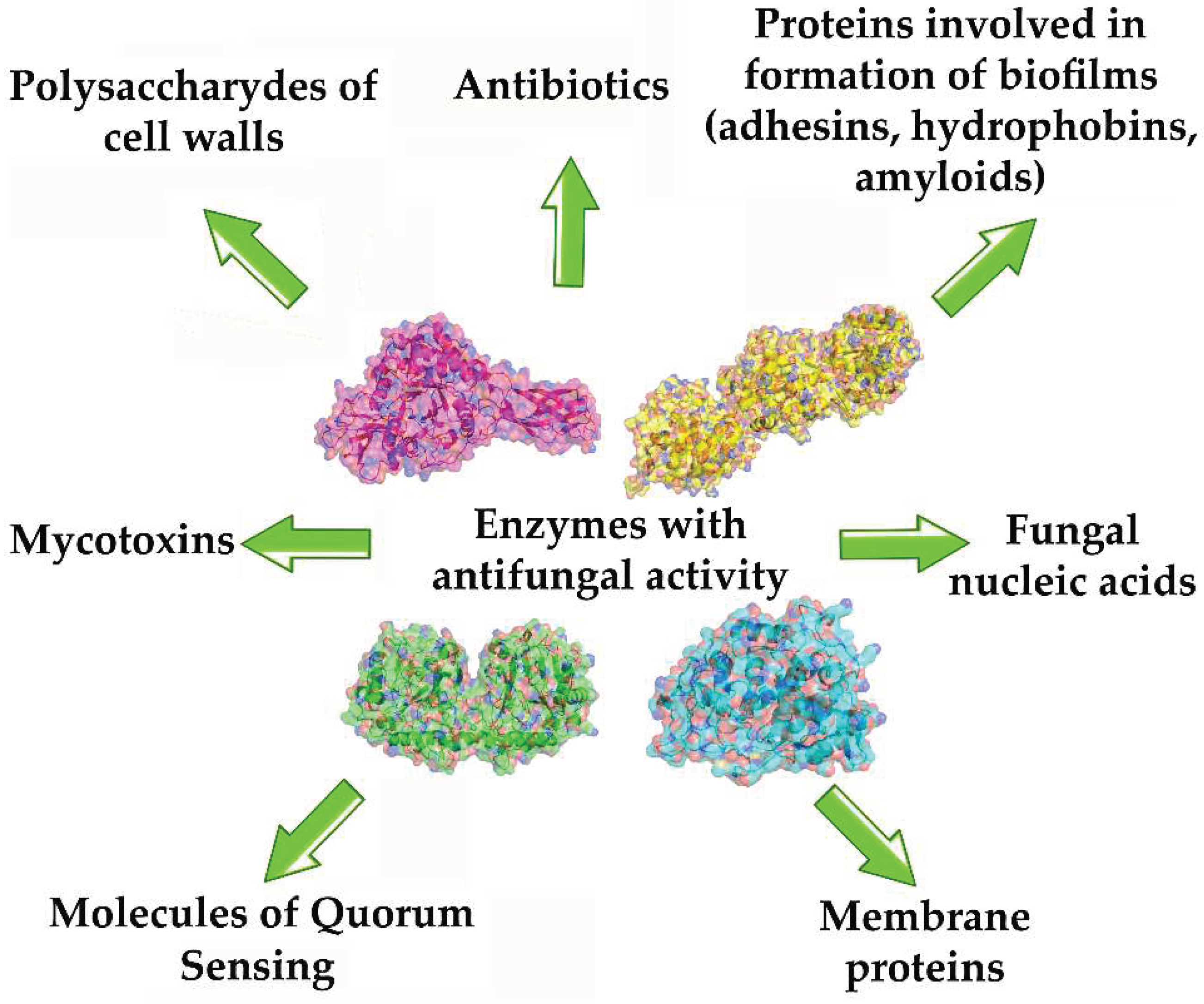

Table 1.
Antifungals based on use of metal-nanoparticles (Me-NPs), metal–organic frameworks (MOFs), and their derivatives*.
Table 1.
Antifungals based on use of metal-nanoparticles (Me-NPs), metal–organic frameworks (MOFs), and their derivatives*.
| Antifungal agent [Reference] | Target of action | Antifungal activity | Efficiency of antifungal action |
|---|---|---|---|
| ZrO2-Ag2O (14-42 nm)[23] |
Candida albicans, C. dubliniensis, C.glabrata, C.tropicalis |
The growth rate inhibition | 89-97% inhibition |
| WS2/ZnO nano- hybrids [24] | C. albicans | Inhibition of biofilm formation | 91% inhibition |
| CuO@C (36–123 nm) [25] |
Alternaria alternata, Fusarium oxysporum, Penicillium digitatum, Rhizopus oryzae | Inhibition of the hydrolytic activity of fungal enzymes used by them for their own metabolism | Inhibition (100 μg/mL) of cellulases and amylases secreted by fungi: 38% and 42% for A. alternata, 39% and 45% for F. oxysporum, 24% and 67% for P. digitatum, and 20% and 24%for R. oryzae, respectively |
| ZnO-NPs [26] |
C. albicans, Aspergillus niger |
Inhibition of growth | Large enough zone of growth absence (8-9 mm) |
| ZnO-NPs (20-45 nm) [27] |
Erythricium salmonicolor | Notable thinning of the hyphae and cell walls, liquefaction of the cytoplasmic content with decrease in presence of a number of vacuoles | Significant inhibition (9-12 mmol/L) of cell growth |
| ZnO–TiO2 (8-33 nm) [28] |
A. flavus | High level of ROS production and oxidative stress induction. Treated objects have a lower count of spores and damaged tubular filaments and noticeably thinner hyphae compared to the untreated fungi | Fungicidal inhibition (150 μg/mL) zone is 100 % |
| ZnO (40-50 nm) [29] |
C. albicans | High level of ROS production | MIC=32-64 mkg/mL MFC=128-512 mg/mL |
| Fe2O3 (10–30 nm) [30] |
Trichothecium roseum, Cladosporium herbarum, P. chrysogenum, A. alternate, A. niger. |
Inhibition of spore germination | MIC=0.063-0.016 mg/mL |
| Fe3O4 (70 nm) [31] | C. albicans | Inhibition of cell growth and biofilm formation | MIC=100 ppm MFC=200 ppm |
| Cu-BTC (10–20 µm)[32] |
C. albicans, A. niger, Aspergillus oryzae, F. oxysporum |
ROS producing, the damage of the cell membrane | Inhibition of C. albicans colonies is 96% by 300 ppm and up to 100% by 500 ppm. Inhibition growth of F. oxysporum and A. oryzae is is 30% with 500 ppm. No significant effect on the A. niger growth. |
| HKUST-1 or HKUST-1NP (doped with NPs of Cu(I)) (49-51 nm) [33] | A. niger, Fusarium solani, Penicillium chrysogenum | Appearance of Cu+2 inhibiting of cell growth | 100% growth inhibition of F. solani by 750-1000 ppm and P. chrysogenum by 1000 ppm; for A. niger - no inhibition |
| [Cu2(Glu)2(LIGAND)] x(H2O) [34] |
C. albicans , A. niger spores |
The apoptosis-like fungal cell death | 50–80% fungal death at 2 mg/mL of the MOFs |
| MIL-53(Fe) and Ag@MIL-53(Fe) composite [35] | Aspergillus flavus | Inhibition of cell growth | MIC= 40 μg/mL for the MIL-53(Fe); MIC=15 μg/mL for the Ag@MIL-53(Fe) |
| Ce-MOF on the base of 4,4’,4”-nitrilo- tribenzoic acid [11] |
A. flavus, A. niger, Aspergillus terreus, C. albicans, Rhodotorula glutinis |
Enzymatic-like activity: catalase, superoxide dismutase, and peroxidase | Inhibition efficiency of 93.3–99.3% based on the colony-forming unit method |
| NPs of TiO2 co-doped with nitrogen and fluorine (200–300 nm) [12] | F. oxysporum | Peroxidase-like enzymatic activity, production of ROS under light irradiation | 100% inhibition of fungal growth |
| Fe3O4@MoS2-Ag (~428.9 nm) [36] | C. albicans | Peroxidase-like enzymatic activity | 80% damage of cell membranes |
| CoZnO/MoS2 nanocomposite [37] | A. flavus | Peroxidase-like photocatalytic activity | MIC=1.8 mg/mL |
*BTC - 1,3,5-benzenetricarboxylate; Glu – glutarate; HKUST – Hong Kong University of Science and Technology; HKUST-1 = [Cu3(BTC)2(H2O)3]n with three-dimensional structure; MFC - minimum fungicidal concentrations; MIC - minimum inhibitory concentration; MIL - Material Institute Lavoiser, MIL-53(Fe) - iron−benzenedicarboxylate constructed from a combination of 1,4-benzenedicarboxylate and FeO4(OH); LIGAND - 1,2-bis(4-pyridyl)ethane, 1,2-bis(4-pyridyl)ethylene, or 1,3-bis(4-pyridyl) propane.
Table 2.
Metal-containing compounds with biomolecules as ligands.
| Antifungal formulation [References] |
Target fungi | *MFC, mg/L | Comment |
|---|---|---|---|
| Green synthesized Au NPs in Spirulina maxima [37] |
C. albicans |
0.064 (μDT) |
Damage of cell wall |
| Green synthesized Ag NPs (30 nm) [50] |
C. albicans | n/a (DDT) | Antibacterial agent |
| Bio-synthesized Te nanorods (ca. 50×500 nm) [51] |
F. oxysporum Alternaria alternata |
60 (SGT) 40 (SGT) |
Germination inhibitor |
| Green synthesized Ag NPs (30 nm) [52] |
C. albicans Candida glabrata Candida parapsilosis |
1.6–6.3 (μDT) 3.1 (μDT) 12.5 (μDT) |
Antibiofilm agent, yeast-to-hyphal inhibitor |
| Green synthesized Fe3O4 NPs (0.5–2 μm) [53] |
C. albicans | 6.3 (μDT) | Antibiofilm agent, yeast-to-hyphal inhibitor |
| Green synthesized Ti/Ag NPs (20–120 nm) [54] |
A. niger A. flavus F. solani |
>100 (MGT) | Antibacterial agent |
| ZnO microparticles (2–5 μm) in film of soybean proteins with cinnamaldehyde [55] | Aspergillus niger | n/a (DDT) | Coating for surfaces, field trial |
| ZnO NPs (20–40 nm) in zein/gelatin nanofibers with (poly)phenolic acids [56] | Botrytis cinerea | n/a (MGT) | Coating for surfaces, field trial |
| CuO NPs in Ca-alginate nanogel coated by poly-ε-lysine (60 nm) [57] |
A. alternate, B. cinerea Phytophthora capsica, Thanatephorus cucumeris Fusarium graminearum |
>1000 (MGT) | Germination inhibitor used in field trials |
* MFC – minimum fungicidal concentration; n/a – not available; DDT – disk diffusion test; μDT – microdilution test; MGT – mycelium growth test; SGT – germination test; **EC50 and EC90 - the effective concentrations for 50% and 90% inhibition.
Table 3.
Enzymes from various sources with antifungal activity.
| Enzyme, its molecular weight and origin [Reference] | Object of action | Conditions of action | Target of action |
|---|---|---|---|
| Chitinases | |||
| Chitinase (35 kDa) from seeds of naked oat Avena chinensis [70] | Panus conchatus, Trichoderma reesei | pH 7.0; 30-50 °C | Hydrolysis of β-1, 4-glycosidic linkages in chitin (an insoluble linear homopolymer of β-1,4-linked N-acetyl-glucosamine residues |
| Chitinase (33 kDa) from Lactobacillus coryniformis 3N11[71] | Alternaria alternata | pH 5.0-7.0; 50-80 °C | Inhibition of fungal cell growth detected by the method of disks on agar-containing medium |
| Acidic chitinase (52.8 kDa) from Paenibacillus xylanexedens Z2–4 was produced in Escherichia coli BL21 (DE3) cells [72] |
Alternaria alstroemeriae, Botrytis cinerea,Rhizoctonia solani,Sclerotinia sclerotorum, Valsa mali |
pH 4.0-13.0; 50-65 °C; pHoptimum 4.5; Inhibition of the enzyme was detected by the metals: 22-24% by Cu2+and Co2+; 15% by Cr3+ and Mn2+; 17-18% by Sr2+,Ni2+, Fe2+. |
The highest specific activity has towards colloidal chitin, followed by ethylene glycol chitin and ball milled chitin. It inhibits the hyphal extension. |
| Chitinase (30 kDa) from Drosera rotundifolia was produced in Escherichia coli BL21-CodonPlus (DE3)-RIPL [73] | Fusarium poae, Trichoderma viride, Alternaria solani | pH 5.0-7.0; 30-45 °C; Inhibition of the enzyme was detected by the metals: 17% by Fe2+, 40% by Pb2+, 48% by Cu2+ and 58% by Cd2+ |
40-52.6% decrease of fungal growth, whereas the stimulation (up to 50%) of growth of R. solani sp. |
| Chitinase (64.1 kDa) from Bacillus amyloliquefaciens [74,75] |
Botryosphaeria dothidea, B. cinerea, F. graminearum, Rhizoctonia cerealis,S. sclerotiorum, Ustilaginoidea virens |
pH 8.0; 37 °C |
60-68% decrease of fungal growth, degeneration of hyphae morphology |
| Chitinase (77.9 kDa) from a rare actinomycete Saccharothrix yanglingensis Hhs.015 (isolated from the roots of cucumber) [76] |
V. mali | pH 6.0; 30-45 °C; Inhibition of the enzyme was detected by the metals: 80% by Zn2+, 65-70% by Ca2+, Mn2+, Fe2+. Ions Cu2+, Cr3+ and Mg2+ significantly promoted chitinase activity (by 187.3%, 167.5% and 111.9%, respectively). |
Multiple deformations of fungal hyphae. |
| Chitinase (45 kDa) from Streptomyces luridiscabiei U05 (isolated from wheat rhizosphere) [77] |
A. alternata, F. oxysporum, F. solani, F. culmorum, B. cinerea, Penicillium verrucosum |
pH 6.0–8.0; 35–40 °C; 98% inhibition of the enzyme was detected in presence of Hg2+and Pb2+. The chitinase activity was stimulated by Ca2+ (120%) and Mg2+ (140%) ions. |
Inhibition of fungal growth due to the demonstration of both endo- and exo-chitinase activity by the enzyme. |
| Chitinase (94.2 kDa) from Salinivibrio sp. BAO-1801 [78] |
A.niger, F. oxysporum, R. solani |
pH 6.0–8.0; 40–55 °C |
100% inhibitory effect on spore germination of fungal cellss |
| Endochitinase (52.9 kDa) from Corallococcus sp. EGB was produced in Escherichia coli BL21 (DE3) [79] | Magnaporthe oryzae | pH 5.0–8.0; 30–55 °C |
Inhibition of conidia germination and appressorium formation due to the hydrolysis of chitin into N-acetylated chitohexaose |
| Endo- and exochitinases (34, 41 and 48 kDa) from Serratia marcescens PRNK-1 isolated from cockroaches Periplaneta americana [80] | R. solani, F. oxysporum | pH 4.5-7.0; 40-60 °C; pHoptimum 5.5, toptimum 55 °C |
Strong inhibition of fungal hyphae growth |
| Chitinase (46 kDa) from Trichoderma harzianum GIM 3.442 [81] | B. cinerea | pH 5.0-8.0; 40-55 °C; pHoptimum 6.0, toptimum 45 °C 49.4% and 66.6% inhibition of the enzyme was detected in presence of Zn2+and Cu2+ respectively. The chitinase activity was stimulated by Ca2+ (115.2%) and Sr2+ (112.6%) ions. |
Up to 80% inhibition of fungal growth |
| Complex of chitinases (25, 37 and 110 kDa) from Aeromonas sp.[82] |
F. solani, A. alternate, B. cinerea, Penicillium sp. |
pH 5.0-8.0; 30-50 °C |
Inhibition of fungal growth |
| Endochitosanasa (50.7 kDa) from Aquabacterium sp. [83] |
M. oryzae, F. oxysporum |
pH 5.0; 40 °C | The enzyme inhibits appressorium formation of M. oryzae and hydrolyzes 95%-deacetylated chitosan with accumulation of chitooligosaccharides inhibiting the growth of fungal cells of M. oryzae and F. oxysporum. |
| Chitinase (30 kDa or 48 kDa) from Streptomyces sampsonii with bifunctional activity was produced in Escherichia coli BL21 (DE3) [84,85] |
Cylindrocladium scoparium, Cryphonectria parasitica,, Neofusicoccum parvum, F. oxysporum |
pH 3.0-11.5; 30-60 °C pHoptimum 6.0, toptimum 55 °C Enzymatic activity was stimulated by Ca2+ (132%), Mg2+ and Mn2+ slightly (9%) decrease the activity, whereas Ag+ and Cr3+ notably inhibited enzymatic activity (80% and 42% respectively). |
The enzyme possessed the dual enzymatic activity of chitinase and lysozyme and its action results in complete destruction of the mycelial morphology. |
| Glucanases | |||
| Endo-1,4-galactosaminidase from Aspergillus fumigatus was produced in E. coli BL21 (DE3) [86] | A. fumigatus | pH 6.0-7.0; 28 °C |
Enzyme catalyzes the hydrolysis of exopolysaccharide galactosaminogalactan being integral component of A. fumigatus matrix |
| Endo-a-1,4-N-acetyl-D- galactosaminidase (produced in E.coli) and Endo-a-1,4-D- galactosaminidase (produced in Pichia pastoris) [87] |
A. fumigatus | pH 7.4; 37 °C | The enzymes catalyses destruction of adhesive exopolysaccharides in biofilms formed by fungi. |
| Endo-β-1,3-glucanase (46.6 kDa) from M. oryzae was produced in E. coli BL21 (DE3) [88] | M. oryzae,U. maydis | pH 5.0-8.0; 20-45 °C Relative activity was following in presence of K+ (39%), Ba2+ (46.2%), Ca2+ (47.1%), Co2+ (22.2%), Cr2+ (55.1%), Cu2+ (30%), Mg2+ (31.6%), Mn2+ (23.1%), Ni2+ (20.7%), Zn2+ (1.9%), Fe3+ (60%) and Fe2+ (103.1%) |
The enzyme inhibits formation of germ tubes and appressoria. |
| β-(1-3)-glucanase (32 kDa) from Bacillus halotolerans was produced in E. coli BL21 [89] | V. dahliae | pH 7.0; 28 °C |
Strong inhibition of spore germination and mycelial growth of the fungal cells. |
| Proteases | |||
| Serine protease (87.16 kDa) from B. licheniformis TG116 [90] |
P. capsica, R. solani, F. graminearum, F. oxysporum, Botrytis cinerea, Cescospora capsici |
pH 7.3; 30 °C | The most notable inhibition of fungal growth was revealed in case of C. capsici. |
| Aspartic protease P6281 (38 kDa) from T. harzianum was produced in Pichia pastoris cells [91] |
B. cinerea, Mucor circinelloides, A. fumigatus, A. flavus, R. solani, C. albicans |
pH 2.5-4.0; 30-45 °C; 49.4% and 66.6% inhibition of the enzyme was detected in presence of Zn2+and Cu2+ respectively. The aspartic protease activity was stimulated by Mn2+ (140.1%) and Cu2+ (151.2%) ions. Ca2+, Mg2+ and Ni2+ slightly (7-10%) increase the activity, whereas Fe2+ and Zn2+ ions decrease activity of the enzyme. | Inhibition of spore germination and growth of fungal cells: 57.3% B. cinerea, 30.9% 26.1% M. circinelloides, 27.2% A. fumigatus, 34.8% A. flavus, R. solani, C. albicans |
| Lysozyme | |||
| Lysozyme with fluconazole in shellac NPs [92] | Biofilm of C. albicans | pH 5.5 | Biofilm clearing effect was observed. |
| Lysozyme in NPs of chitosane [93] | Aspergillus parasiticus | 28 °C | The decrease in fungal cell viability, 100%- inhibitory effect on the germination of spores was confirmed. |
| Peroxidases | |||
| Peroxidase (58 kDa) from cowpea (Vigna unguiculata) roots [94] |
Colletotrichum gloeosporioides, F. oxysporum |
pH 4-7; 37-75 °C |
Enzyme catalyzes redox reactions and inhibits the conidia germination of fungal cells by altering the permeabilization of membranes. |
| Peroxiredoxin (38 kDa) from Enterobacter sp. was produced in E. coli DH5α [95] |
Verticillium dahlia, F. solani |
pH 4-7; 37-75 °C |
Peroxidase inhibits the growth of fungi. |
| Nucleases | |||
| RNase 3 from eosinophils and the skin-derived RNase 7 were produced in E. coli BL21 (DE3) [96,97] | C. albicans | pH 5.0-7.2; 20-37 °C | RNases demonstrated dual mechanism of action: an overall yeast membrane-destabilization (permeabilization and depolymerization) and degradation of target cellular RNA. |
| Bovine pancreas RNase A1, human recombinant ribonucleases A2, A5 and A8 [98] |
C. albicans, C. glabrata |
pH 5.0-7.2; 30-37 °C | Action of RNase A1 was the most pronounced, it completely killed Candida cells by lowering the mitochondrial membrane potential but did not damage the cell membrane. |
| Enzymatic comlexes | |||
| Chitinase and β-1,4-endoglucanase co-synthesized by Paenibacillus elgii PB1[99] |
A. niger, Trichophyton rubrum, Microsporum gypseum, C. albicans, Saccharomyces cerevisiae |
pH 5.0; 30 °C Urea had significant negative effect on β-1,4-endoglucanase, Zn2+ positively affected both enzymatic activities. |
Inhibition of fungal growth: 88% A. niger, 92% T. rubrum, 52% M. gypseum, 55% C. albicans, 71% S. cerevisiae |
| Enzymatic complex from Penicillilum verruculosum containing chitinase (43 kDa, gene from Myceliophtora thermophyla), cellobiohydrolase (66 kDa), endoglucanase (39 kDa) and xylanase (32 kDa) [100] |
Fusarium culmorum; F. sambucinum; F. graminearum; Stagonospora nodorum; S. tritici; A. flavus |
pH 4.5-6.2; 52-65 °C |
The enzymatic complex catalyzed hydrolysis of fungal mycelium. |
| Lyticase (enzymatic complex with activity of β-(1-3)-glucan laminaripentaohydrolase, β-(1-3)-glucanase, protease and mannanase) from Arthrobacter luteus [101] | C. albicans | pH 7.3; 25-37 °C | Lyticase provides disruption of yeast cell walls and spores with formation of spheroplasts and further release of DNA from them. |
| Cellobiose dehydrogenase (CDH) and deoxyribonuclease I (DNase) co-immobilized on positively charged chitosan nanoparticles [102] | Polymicrobial biofilms of C. albicans and Staphylococcus aureus | pH 7.5; 37 °C | The action of two enzymes provides a violation of biofilm formation due to the degradation of eDNA, a decrease in the thickness of the biofilm and the death of microbial cells. |
Table 4.
Different proteases applied for prion degradation.
| Enzyme; Origin; Reference | Protease Type | Prion/Amyloid |
|---|---|---|
| Subtilisin homolog Tk-SP from Thermococcus kodakarensis [117] | Serine protease |
abnormal human prion protein |
| Nattokinase from Bacillus subtilis natto [118] | amyloid β fibrils / recombinant human prion protein | |
| Subtilisin 309 from Bacillus lentus [119] | mouse-adapted scrapie prion protein | |
| Prionzyme from Bacillus subtilis [120] | hamster prion protein | |
| Properase from Bacillus lentus [121] | mouse-adapted prion protein | |
| MC3 (Prionzyme) from Bacillus lentus [122] | 301V prion | |
| MSK103 from Bacillus licheniforms [123] | hamster-adapted scrapie prion protein | |
| E77 from Streptomyces sp. [124] | ||
| Pernisine from Aeropyrum pernix [125] | abnormal human prion protein | |
| Protease from lichens (Parmelia sulcata, Cladonia rangiferina and Lobaria pulmonaria) [126] | hamster prion protein | |
| Keratinase from Bacillus licheniformis [127] | yeast prion protein, Sup35NM | |
| Proteinase K [128] | ||
| Keratinase rKP2 from Pseudomonas aeruginosa KS-1 [129] | ||
| Keratinase from Bacillus pumilus KS12 [130] | ||
| Keratinases, Ker1 and Ker2, from Amycolatopsis sp. MBRL 40 [131] |
Metal-activated serine protease | amyloid β fibrills |
| Neprilysin [132] | Zn-dependent metalloprotease | amyloid β fibrills |
| Insulin-degrading enzyme [132] | ||
| A disintegrin and metalloproteinase (ADAM10) [132] |
Table 5.
Enzymes hydrolyzing the QSMs of various fungal cells.
| QQE [Reference] | Objects of action | Target QSMs |
|---|---|---|
| Gluconolactonase [141] | A. niger | Lactone-containing QSMs |
| His6-OPH [142,143] |
Trichosporon beigelii, Candida sp., S. cerevisiae, Pachysolen tannophilus, Kluyveromyces marxianus |
Lactone-containing QSMs |
| Esterases [144,145] |
Mucor mucedo, Blakeslea trispor Phycomyces blakesleeanus |
Trisporic acids |
| Lipases [146,147,148] |
Malassezia sp., Microsporum canis Leishmania amazonensi |
Lipids with long carbon chain fatty (oleic, linoleic, and linolenic) acids |
Table 6.
Combined application of enzymes possessing antifungal activity with metal NPs.
| Antifungal formulation [Reference] |
Target fungi | MFC*, mg/L | Comment, [Reference] |
|---|---|---|---|
| Glucose oxidase with NPs of Fe3O4 [40] | C. albicans | 1 mg/mL | Antimicrobial activity |
| Keratinase on green synthesized Ag NPs (5–25 nm) [155] | C. albicans | n/a (DDT) | Antibacterial agent |
| Chitinase on talc (0.2–3 μm) combined with chitin [156] | Sclerotium rolfsii | n/a | Passed field trial |
| Protease and lipase on green synthesized Ag NPs (10–45 nm) [157] |
C. albicans | n/a (DDT) | Antibacterial agent |
| β-1,3-glucanase, chitinase, N-acetylglucosaminidase and acid protease on green synthesized Ag NPs (20–200 nm) [158] |
Sclerotinia sclerotiorum |
n/a (MGT) | Variable sorption capacityof Ag NPs for different enzymes |
| β-1,3-glucanase, chitinase, N-acetylglucosaminidase and acid protease on green synthesized Ag NPs (20–200 nm) [159] |
S. sclerotiorum Beauveria bassiana |
n/a (MGT) | Antibacterial agent |
| Glucose oxidase conjugated with polyglutaraldehyde–β-alanin and covered by Ag shell [160] |
C. albicans Microsporum canis T. rubrum |
n/a (DDT) | Antibacterial agent, cytochrome inhibition |
| Lactoferrin and melittin in Zn–MOF (0.5 μm) [161] | C.albicans | >100 (μDT) | Antibiofilm agent, yeast-to-hyphal inhibitor used in vivo |
*MFC – minimum fungicidal concentration; n/a – not available; DDT – disk diffusion test; MGT – mycelium growth test; μDT – microdilutin test;.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



