Submitted:
27 May 2023
Posted:
30 May 2023
You are already at the latest version
Abstract
Legionella is an opportunistic pathogen with a biphasic life cycle and humans are occasional hosts of Legionella. The aim of the study was to assess the distribution of virulence genes and genetic diversity among L. pneumophila isolated from water-supply systems of residential buildings in Latvia. In total, 492 water samples from 200 residential buildings were collected. Identification of Legionella spp. was performed according to ISO 11731 and 58 isolates were subjected to whole genome sequencing. At least one Legionella positive sample was found in 112 out of 200 apartment buildings (56.0%). The study revealed extensive sequence type diversity where 58 L. pneumophila isolates fell into 36 different sequence types. A total of 420 virulence genes were identified of which 260 genes were found in all sequenced L. pneumophila isolates. Virulence genes enhC, htpB, omp28, and mip were detected in all isolates, suggesting that adhesion, at-tachment, and entry into host cells are enabled for all isolates. The relative frequency of viru-lence genes among L. pneumophila isolates was high. High prevalence, extensive genetic diver-sity, and the wide range of virulence genes indicated that the virulence potential of environ-mental Legionella is high and proper risk management is of key importance to public health.
Keywords:
Legionella
; residential buildings
; virulence factors
; virulence genes
; sequence type
1. Introduction
Legionella pneumophila is a gram-negative, intracellular bacterial pathogen that causes the potentially fatal but preventable Legionnaires' disease (LD), which manifests as severe pneumonia. L. pneumophila is also associated with a mild, flu-like illness Pontiac fever [1]. Although more than 16 serogroups of L. pneumophila are known, only half of them have been associated with LD, with the serogroup 1 (SG1) being responsible for the majority of LD cases in Europe. A large proportion of LD cases are identified by the urinary antigen test, which is the best for identifying illnesses caused by L. pneumophila SG1 [2]. However, widespread application of urinary antigen test may lead to improper identification and underreporting of other L. pneumophila SGs. This may result in delayed epidemiological investigation and implementation of preventive measures against L. pneumophila in water circulation systems [3].
Legionellae are ubiquitous and have been found in natural and artificial aquatic environments around the world [4]. The spectrum of habitats includes underground and surface water, moist soils, aquatic plants, and rainforests, although anthropogenic aquatic environment is considered as the main reservoir. Since Legionella can establish biofilms in faucets, shower heads, cooling towers, spas, fountains, and especially hot water systems, hotpots of Legionella infection have been reported in residential buildings, hospitals [5], nursing homes [6], hotels and in other private and public buildings [7].
The temperature and water stagnation seem to be the most important risk factors for Legionella colonization in building water supply systems [8]. The very low counts of Legionella characteristic of natural habitats can increase markedly within engineered hot water systems if the recommended temperature regime for the control of opportunistic pathogens has not been followed during routine maintenance in buildings [9]. Contamination of water with L. pneumophila in residential buildings has been strongly associated with temperature in the water circulation system [10]. Changes in the water flow may facilitate release of Legionella from biofilms localised on the inner surface of pipes in water supply network, while dead-ends with stagnant water or rarely used branches of the pipeline can significantly promote the proliferation of Legionella, causing further contamination of water [4,8].
Legionella is an opportunistic pathogen with a biphasic life cycle [11]. Like other intracellular bacteria such as Coxiella, Legionella can alternate between a transmissible (virulent) or a replicative (non-virulent) phases, which ensure the survival in both nutrient-rich and poor environments and enhance bacterial transfer between different ecological niches [12].
Humans are occasional hosts of Legionella and the ability of the pathogen to infect humans is considered to result from a long co-evolution between bacteria and protozoa. The development of LD depends not only on the host’s immunity, bacterial counts and SGs, but also on virulence factors that the pathogen may contain. The most important virulence factors of Legionella are related to the bacterial surface structure and effector protein secretion systems [13]. Surface virulence factors such as the outer membrane proteins, flagella, pili, and lipopolysaccharides promote the initial adhesion to the host cells and enhance infectivity, while toxin production increases the infectivity, survival, and multiplication in alveolar macrophages. Specific genes may regulate biofilm production and increase the ability of the pathogen to survive in water supply systems [14]. Proteins and effectors that provide Legionella with their natural ability to infect protozoa also provide the pathogen with the ability to infect mammalian cells. Through horizontal gene transfer from the host amoebae, Legionella has acquired numerous eukaryotic-like genes that regulate the host cell infection [15]. Some of the bacterial effectors can perform parallel functions in different organisms, and some of the effectors can overlap or replace each other in pathogenic and non-pathogenic species [16].
A multidisciplinary approach to the prevention can be essential for reducing the spread of L. pneumophila, hence decreasing the incidence of Legionnaires' disease. The lack of requirements and preventive actions leads us to assume that in Latvia we can expect a high prevalence and a high genetic diversity of Legionella in water-supply systems in residential buildings, which can pose a risk to residents. Thus, the aim of the present study was to assess the distribution of virulence genes and genetic diversity among L. pneumophila isolated from water-supply systems of residential buildings in Latvia.
2. Materials and Methods
2.1. Sampling
In total, 492 water samples from multi-storey residential buildings were collected in sterile bottles, including cold water (n=164) and hot water (n=328). Samples were collected from August 2016 until December 2022 at 200 residential buildings from 26 municipalities in Latvia. The association with LD cases had been epidemiologically confirmed for 132 apartment buildings, while for the rest of the buildings (n=69) no association with LD cases had been reported. Buildings that have been linked to LD cases reported in residents relied on thermal disinfection as the primary method of disinfection of water-supply systems. In buildings without LD history, the additional disinfection or Legionella monitoring has not been implemented. All the buildings included in the present study were older than 30 years.
Sampling was performed by a trained staff in accordance with the requirements of ISO 19458 [17]. At each sampling point, at least one hot water sample was taken from the shower head. Additional samples were taken depending on the size of the building and the responsiveness of the residents, and included a cold water sample from a shower head and a hot tap water sample. One litre of water was collected at each location. The water circulation temperature was measured during sampling. A specially equipped vehicle was used to transport the samples to the laboratory while maintaining the temperature between 0 and 6 °C during transportation. Testing of the samples was started no later than 6 hours after sampling.
2.2. Microbiological testing of Legionella spp.
Identification and enumeration of Legionella spp. was performed according to ISO 11731 [18]. One litre of water was concentrated using a 0.45 μm polyamide membrane filter (Millipore, Molsheim, France). The filter membranes were resuspended in sterile distilled water (5 mL), shaken for two minutes (Vortex Genius, IKA, Staufen, Germany). A total of three 0.1 mL aliquots (untreated, heat treated, and acid treated) were spread on Buffered charcoal yeast extract agar (BCYE, Biolife Italiana, Milan, Italy) and Glycine vancomycin polymyxin B cycloheximide agar (GVPC, Biolife Italiana, Milan, Italy). For samples taken before November 2017, only the GVPC medium was used.
The inoculated plates were incubated at 36 °C for 10 days. At least three presumptive colonies of Legionella from each plate were subcultured on BCYE agar medium (Biolife Italiana, Milan, Italy) and BCYE agar medium without L-cysteine (BCYE-Cys, Biolife Italiana, Milan, Italy), and incubated at 36 °C for at least 48 h.
Suspected Legionella colonies were identified by matrix-assisted laser desorption/ionisation time of flight mass spectrometry (MALDI-TOF MS, Bruker, Bremen, Germany). An agglutination test (Thermo Fisher Scientific, Bred, Netherlands) was used for the confirmation of L. pneumophila. Individual latex reagents (Pro-Lab Diagnostics, Richmond Hill, Canada) were used for the detection of L. pneumophila serogroups.
Presumptive colonies from all BCYE and BCYE-Cys plates were counted and confirmed, and the estimated number of Legionella was expressed as CFU/litre with an indication of serogroup. All confirmed Legionella isolates were obtained in pure cultures and transferred to the culture collection for long-term storage at −80 °C.
2.3. DNA extraction from Legionella isolates
From all 197 L. pneumophila positive samples, 58 isolates were selected for genetic analysis. Isolates were selected, based on the frequency of L. pneumophila serogroup in the analysed water samples, the geographical origin of the samples, and the year of sampling. The isolates were thawed and cultured on BCYE agar at 37 °C for 48 h before DNA extraction. A single colony from each culture was subjected to manual DNA extraction using the NucleoSpin Tissue reagent kit (Macherey-Nagel, Düren, Germany) according to the manufacturer’s instructions.
2.4. Whole Genome Sequencing
All 58 isolates were subjected to whole genome sequencing. The wet-lab procedures for library preparation and the bioinformatics procedures for assembly, data quality control, sequence-based typing (SBT), and core genome MLST (cgMLST) typing were carried out as described previously [19]. Briefly, the trimmed reads were assembled into contigs using the SPAdes assembler v3.14.0 [20]. The contigs were then used as input for further SBT analysis according to the ESCMID Legionella Study Group (ESGLI) scheme [21,22] and cgMLST typing according to the scheme published by Moran-Gilad [23]. The collection of core genes among all isolates was determined using the pangenome analysis tool Roary v3.13.0 [24]. The core gene SNP alignment produced by Roary was then used by FastTree v2.1.10 [25] to build an approximate maximum-likelihood phylogeny.
The virulence factor database (VFDB, retrieved on 12 November 2021) [26] and separately the sequence of rtxA gene of L. pneumophila strain AA100 (nucleotide positions 949-4575 of GenBank ID AF057703.1) were used to identify virulence-encoding genes. The ABricate v1.0.1 tool (https://github.com/tseemann/abricate) was used to screen the assembled genomes for the presence of these virulence genes based on a nucleotide BLAST approach [27]. The thresholds of 80% BLAST sequence identity and 80% length coverage were set to qualify any gene as present in a genome. Additionally, in silico PCR with the rtx1/rtxA-rtx2/rtxA and rtx3/rtxA-rtx4/rtxA primer pairs [28] was simulated using the iPCRess tool from Exonerate v2.2.0 software package [29]. The presence of antimicrobial resistance (AMR) genes was determined using the ResFinder software v4.1.7 and its associated database (version 2022-05-24) [30]. The same identity and coverage thresholds were used for AMR genes as for virulence genes.
2.5. Data Analysis
R version 4.2.3 (15.03.2023. ucrt), 2023 (The R Foundation for Statistical Computing, Vienna, Austria) was used for data analysis. Chi-squared tests and ANOVA were used to calculate differences between variables. Genotypes were visualized in the form of dendrogram with iTOL v6.7.3 [42] and in the form of minimum spanning tree with GrapeTree v1.5.0 [43].
3. Results
3.1. Prevalence of L. pneumophila in residential buildings
Overall, 197 of 492 (40.0%) samples were Legionella spp. positive (Table 2). Only two Legionella species – L. pneumophila (n=196) and L. rubrilucens (n=1) were found.
At least one Legionella positive sample was found in 112 out of 200 apartment buildings (56.0%), however, there were no significant differences between buildings linked to known cases of LD (71 positive buildings of 132) and buildings without known LD cases (41 positive buildings of 69; p=0.54).
In buildings with known LD cases, the prevalence of Legionella spp. was significantly lower in cold water (p<0.0001) while in buildings without known LD cases differences in the prevalence of Legionella between the cold and the hot water were not observed (p=0.192). Overall, the prevalence of Legionella spp. was higher both in cold (p<0.01) and hot water samples in apartment buildings without known LD cases, although for hot water the difference was not statistically significant (p=0.056).
In total, six L. pneumophila serogroups (SG) and three combinations of them where identified. Out of all 196 L. pneumophila positive water samples, in 191 cases (97.4%) only one SG was identified. Overall, 55.1% (108/196) of isolates represented SG 2 and 28.1% (55/196) SG 3. In five water samples (2.6%) two SGs were found simultaneously (Table 3). No significant differences were observed in the prevalence of SGs in apartment buildings with and without known cases of LD (p>0.05).
The observed levels of L. pneumophila colonization varied from 50 CFU/L up to 1.7×104 CFU/L, with the average value of 1.8×10³ CFU/L. No significant differences were revealed in the levels of colonization with various SGs between buildings with or without known LD cases, with the exception of L. pneumophila SG 3, which showed a significantly higher level of colonization (p<0.05) in apartment buildings linked to LD cases (Table 4).
3.2. Whole-genome sequencing of L. pneumophila
Overall, 36 SBT sequence types (STs) were identified for 58 L. pneumophila isolates. Ten SBT sequence types have not been previously documented and are considered as new STs (Table 5).
ST 338 was the most frequent sequence type, which was determined for 10 out of 58 isolates (17%). None of ST 338 isolates represented L. pneumophila SG 1. ST 366 and ST 2002 were detected in three isolates each (5%), while the other STs were detected in no more than two isolates each.
Using cgMLST typing, all 58 sequenced L. pneumophila isolates fell into 56 different cgMLST types. No cgMLST types specific to geographic location, serogroup, or SBT type were detected. No distinct clusters have been identified, but clades around dominant STs such as ST 338 and ST 366 can be discerned, and the clade around ST 1104 is the most distant (Figure 1).
All sequenced L. pneumophila isolates had only one antibiotic resistance gene – aph(9)-la, encoding antibiotic resistance factor spectinomycin phosphotransferase.
In total, 420 virulence genes representing 59 gene families were found in 58 sequenced L. pneumophila genomes. The number of genes per one isolate varied from 312 to 415 (Table 5), with the average number of 375 virulence genes per isolate. A similar diversity of genes was observed between the isolates from buildings linked to LD cases and buildings without known LD cases. Notable differences between isolates of different serogroups were not found.
Genes enhC, htpB, omp28, mip, mavC, legK1, sidJ, lvhD4, lpnE, lspC, and rtxA were selected as objects of the greatest interest in this study and the relative frequency of these genes was evaluated in all 58 isolates (Table 6).
No significant differences were observed in the relative frequency of genes (p>0.05) between buildings linked to LD cases and buildings without known LD cases and between different serogroups, with the exceptions for sidJ, which was the less frequent in SG 9 isolates than in SG 1, SG 2, and SG 3 isolates (p<0.05) and for PCR simulated rtxA, which was less frequent in SG 1 isolates than in SG 3 isolates (p<0.05).
A total of 260 genes (62.1%), including enhC, htpB, omp28, mip, lpnE, and 11 genes of lsp family were observed in all of isolates (Figure 2). The Core Genome SNP dendrogram showed the same clades as the cgMLST minimum spanning tree. The individual leg family virulence genes were found only in seven isolates out of 58 which were present in the clade formed around ST 1104.
The mav family was represented by 13 identified genes, of which nine were found in all isolates while mavC was present in 54, mavG in 53, mavH in 55, and mavI in 57 isolates. Altogether, 29 virulence genes of the leg family have been identified. The frequency of leg genes in L. pneumophila isolates varied from 7 to 58. However, only six genes of the leg family had a frequency of less than 40: legU1 found in 23 isolates, legC1 in 13 isolates, as well as legA6, legL5, legL7, and legLC4 in 7 isolates each. Out of the 11 virulence genes representing the sid family, sidA, sidE, sidF, and sidK were found in all 58 isolates. Furthermore, sidG and sidH were the rarest and were found in six and 13 isolates, respectively. All 11 identified genes of the lvh family were found in 46 isolates, except for lvhB2, which was found only in 27 isolates.
4. Discussion
Evaluation of the presence and diversity of Legionella in 200 residential apartment buildings across Latvia revealed high prevalence of Legionella in water-supply systems of residential buildings. In general, Legionella was found in 56% of residential buildings and in 40% of water samples. Our study showed that the prevalence of L. pneumophila significantly exceeded the data reported in other studies: 20.7% in Germany [44] and 19.8% in Italy [45]. In the USA, at least one positive Legionella sample was found in 15% of single-family homes [46]. However, the prevalence of L. pneumophila reported in this study was similar to a previous report from Latvia, where 39% of water samples from apartment buildings were Legionella–positive [47]. The high prevalence of Legionella in water in Latvia could be related to ineffective water supply system maintenance strategies where water temperature requirements are not met while water temperature is one of the main factors for Legionella persistence and proliferation in building water supply systems [9,10].
Our study showed that water contamination with Legionella was found more often (52,3% vs. 35,6%) in samples from buildings with no previous connection to LD cases. The average temperature of hot water was 50.7 °C, however, it was seven degrees lower on average in buildings without previous LD cases. Currently, the temperature of hot water circulation or at the points of consumption is not regulated in Latvia, and the maintenance companies and building managers are obliged to ensure only the temperature at the exit from the heat exchanger, which must not be lower than 55 °C according to the national legislation [48]. Due to the often considerable total length of water circulation pipelines and heat losses in the water supply system between the heat exchanger and the showerheads, the observed hot water temperature at the points of water consumption reached only 45.8 °C in buildings not related to LD cases. In contrast, managers are obliged to carry out disinfection procedures and Legionella monitoring in buildings with previous LD history, although guidelines for sampling frequency have not been set. Therefore, in buildings with a previous association with LD the managers are more aware and, perhaps for this reason, higher hot water temperatures and lower prevalence of culturable Legionella were observed in those buildings.
In the present study, six serogroups of L. pneumophila were identified, of which SG 2 (55.1%), SG 3 (27.0%), and SG 1 (9.7%) were predominant. However, these results differed from our recent study of L. pneumophila in Latvian hotels, where SG 3 was the predominant serogroup [19]. It is worth to note that the low prevalence of L. pneumophila SG 1 in residential buildings was in agreement with the results from hotels in Latvia [19]. Those results were consistent with low prevalence of L. pneumophila SG 1 antibodies (0.2%) in healthy blood donors in Latvia [49]. Residence in an apartment building with a centralized hot water supply had been identified as the main environmental risk factor, and the seroprevalence of 9.5% in residents of urban apartment buildings was reported [49]. Globally, L. pneumophila SG 1 is considered to be the main causative agent of LD [2] and, accordingly, diagnostic methods for clinical cases have been adapted to identify SG 1. The urine antigen test, which is specific only for L. pneumophila SG 1, is still the first-choice method for the diagnostics of LD [2]. Additionally, the incidence of LD could be underreported because the diagnostics and reports are biased towards SG1, thus we assume that only the most severe cases, which most likely initially had a higher bacterial load, were detected and reported, while other cases may remain unrecognized. Also, to the best of our knowledge, the absence of clinical isolates of other SGs in Latvia until the end of 2022 may indicate insufficient diagnostics of other SGs.
In five noteworthy cases, two different L. pneumophila strains belonging to two different serogroups were found simultaneously in the same water samples. At the time of infection, a person may encounter several Legionella strains with different immunological and antibacterial resistance characteristics, thus the choice of appropriate diagnostic methods for clinical cases can present a significant challenge.
Our study showed extensive sequence type diversity where 58 L. pneumophila isolates fell into 36 different sequence types, 10 of which have not been previously described. It must be admitted that the diversity of sequence types is not unusual for environmental Legionella, as confirmed by previous studies from Bosnia and Herzegovina [50], USA [51], China [52], and Latvia [19]. The higher diversity of STs and the isolation of new STs during the present study could be explained by the focus on residential buildings. In comparison, the lower diversity and lower number of new STs found in hotels during our previous study could be related to internationally distributed STs [19].
The 11 STs found in residential buildings matched those found in Latvian hotels, moreover, the predominant ST 338 and ST 336 were found both in hotels and in residential buildings. Several STs identified in our study have been associated with LD outbreaks and sporadic cases in other countries [53,54]. In addition, we did not find differences between the STs found in buildings with and without LD cases, and all these findings consequently suggest that Legionella strains may persist in residential buildings in Latvia and pose a long-term risk to residents.
In our study, 420 virulence genes were identified of which 260 genes were found in all sequenced L. pneumophila isolates. Genes enhC, htpB, omp28, and mip encoding virulence factors related to the bacterial surface structures were detected in all isolates, suggesting that adhesion, attachment and entry into the host cell are enabled for all isolates. The largest group of genes encoding T4SS effectors were quite variable, however, the relative frequency of virulence genes among L. pneumophila isolates was high. The wide range of genes encoding effectors demonstrated the high plasticity of the L. pneumophila genome and pointed to the possible redundancy of effectors, which is an important feature of Legionella [55]. The redundancy within the SidE effector family is well-established, where members of the SidE effector family exert the same function on specific host cell targets. SidE, SdeA, SdeB, and SdeC catalyse the ubiquitination of host proteins and the deletion of all four of these effectors together, but not individually, impairs intracellular growth, which can be restored by the insertion of just one of them [56].
New gene functions in Legionella are still being studied and not all of the lvh locus group genes have been assigned a function yet. However, the environment in which Legionella grows before contact with the host cell also plays a role. The lvh locus gene lvhB2 is closely related to the ability of the bacterium to infect macrophages or amoebae, depending on the temperature at which Legionella grew prior to contact [57].
During the initial analysis, we noticed a remarkable difference in the prevalence of rtxA-positive isolates between our study and other studies. The rtxA gene was absent in all isolates when screening the genomes against VFDB database. This was in stark contrast with other studies [58,59] where 20.69-100% of L. pneumophila isolates were rtxA-positive. However, those studies relied on PCR to determine the presence of this gene. The commonly used rtx1/rtxA-rtx2/rtxA and rtx3/rtxA-rtx4/rtxA primers have been developed based on the L. pneumophila strain AA100 DNA sequence [28] and they only target two approximately 540-630 bp long fragments of the gene. The rtxA itself is known to have a modular structure and to be highly variable in length and sequence similarity between different L. pneumophila strains [60]. Thereby, we hypothesized that the absence of these two PCR target sites did not necessarily indicate the absence of every possible variant of the rtxA gene.
In order to test this hypothesis, we simulated PCR in silico, using the two aforementioned primer pairs and L. pneumophila reference sequences that were used to characterize the modular structure of rtxA [60]. Only the sequence of strain AA100 produced both in silico PCR products, confirming our hypothesis. Furthermore, the rtxA reference (YP_123037) which was included in the respective release of VFDB also did not produce any of the two expected in silico PCR products. Since the rtxA sequence from strain AA100 was the shortest of available references and it contained the conserved regions that are located at the start and the end of rtxA, we used it as a reference for BLAST-based screening for rtxA in our Legionella genomes.
We concluded that the method for accurate determination of the presence of rtxA should be carefully assessed taking into account the apparent limitations of either the PCR methods or the alignment-based computational methods and their respective reference databases.
The high prevalence, extensive genetic diversity, and the wide range of virulence genes, which have been detected in all isolates from residential buildings in Latvia indicate that the virulence potential of environmental Legionella is high and that all Legionella strains persisting in the water-supply systems should be considered as potentially pathogenic. Virulence gene analysis has shown that Legionella strains persisting in residential buildings can acquire characteristics and increase the pathogenicity through horizontal gene transfer. Proper risk management, implementation of water safety plans, and microbiological monitoring would ensure the protection of residents by reducing the opportunities for Legionella to grow and evolve new virulence traits in man-made water systems.
Author Contributions
Conceptualization, O.V.; D.P., and J.Ķ..; methodology, O.V. and J.Ķ.; software, J.Ķ.; validation, O.V. and D.P.; formal analysis, O.V., M.T. and A.B.; investigation, L.L., J.Ķ.., O.V. and D.P.; resources, A.B.; data curation, D.P. and J.Ķ.; writing—original draft preparation, O.V. and D.P.; writing—review and editing, O.V., D.P., J.Ķ., M.T., L.L., A.K. and A.B.; visualization, D.P.; supervision, A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw fastq data obtained from all isolates have been deposited at the European Nucleotide Archive under the project accession number PRJEB61826.
Acknowledgments
Laboratory of Medical Microbiology, Microbiological Culture Collection, Molecular Biology Division, and Customer Service experts are acknowledged for their technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mondino, S.; Schmidt, S.; Rolando, M.; Escoll, P.; Gomez-Valero, L.; Buchrieser, C. Legionnaires’ Disease: State of the Art Knowledge of Pathogenesis Mechanisms of Legionella. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 439–466. [Google Scholar] [CrossRef] [PubMed]
- Legionnaires’ Disease-Annual Epidemiological Report for 2020 Available online: https://www.ecdc.europa.eu/en/publications-data/legionnaires-disease-annual-epidemiological-report-2020.
- Byrne, B.G.; McColm, S.; McElmurry, S.P.; Kilgore, P.E.; Sobeck, J.; Sadler, R.; Love, N.G.; Swanson, M.S. Prevalence of Infection-Competent Serogroup 6 Legionella Pneumophila within Premise Plumbing in Southeast Michigan. mBio 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.T.; Kamali, A.; Vugia, D.J. Legionella Epidemiologic and Environmental Risks. Current Epidemiology Reports 2019, 6, 310–320. [Google Scholar] [CrossRef]
- Kessler, M.A.; Osman, F.; Marx, J.; Pop-Vicas, A.; Safdar, N. Hospital-Acquired Legionella Pneumonia Outbreak at an Academic Medical Center: Lessons Learned. American Journal of Infection Control 2021. [Google Scholar] [CrossRef]
- De Filippis, P.; Mozzetti, C.; Messina, A.; D'Alò, G.L. Prevalence of Legionella in Retirement Homes and Group Homes Water Distribution Systems. Science of The Total Environment 2018, 643, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Mentula, S.; Kääriäinen, S.; Jaakola, S.; Niittynen, M.; Airaksinen, P.; Koivula, I.; Lehtola, M.; Mauranen, E.; Mononen, I.; Savolainen, R.; et al. Tap Water as the Source of a Legionnaires’ Disease Outbreak Spread to Several Residential Buildings and One Hospital, Finland, 2020 to 2021. Eurosurveillance 2023, 28. [Google Scholar] [CrossRef]
- Nisar, M.A.; Ross, K.E.; Brown, M.H.; Bentham, R.; Whiley, H. Water Stagnation and Flow Obstruction Reduces the Quality of Potable Water and Increases the Risk of Legionelloses. Frontiers in Environmental Science 2020, 8. [Google Scholar] [CrossRef]
- Singh, R.; Chauhan, D.; Fogarty, A.; Rasheduzzaman, M.; Gurian, P.L. Practitioners’ Perspective on the Prevalent Water Quality Management Practices for Legionella Control in Large Buildings in the United States. Water 2022, 14, 663. [Google Scholar] [CrossRef]
- Kruse, E.-B.; Wehner, A.; Wisplinghoff, H. Prevalence and Distribution of Legionella Spp in Potable Water Systems in Germany, Risk Factors Associated with Contamination, and Effectiveness of Thermal Disinfection. American Journal of Infection Control 2016, 44, 470–474. [Google Scholar] [CrossRef]
- Ge, Z.; Yuan, P.; Chen, L.; Chen, J.; Shen, D.; She, Z.; Lu, Y. New Global Insights on the Regulation of the Biphasic Life Cycle and Virulence via ClpP-Dependent Proteolysis in Legionella Pneumophila. Molecular & Cellular Proteomics 2022, 21, 100233. [Google Scholar] [CrossRef]
- Chauhan, D.; Shames, S.R. Pathogenicity and Virulence Of Legionella: Intracellular Replication and Host Response. Virulence 2021, 12, 1122–1144. [Google Scholar] [CrossRef] [PubMed]
- Gattuso, G.; Rizzo, R.; Lavoro, A.; Spoto, V.; Porciello, G.; Montagnese, C.; Cinà, D.; Cosentino, A.; Lombardo, C.; Mezzatesta, M.L.; et al. Overview of the Clinical and Molecular Features of Legionella Pneumophila: Focus on Novel Surveillance and Diagnostic Strategies. Antibiotics 2022, 11, 370. [Google Scholar] [CrossRef] [PubMed]
- Abu Khweek, A.; Amer, A.O. Factors Mediating Environmental Biofilm Formation by Legionella Pneumophila. Frontiers in Cellular and Infection Microbiology 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Scheithauer, L.; Karagöz, M.S.; Mayer, B.E.; Steinert, M. Protein Sociology of ProA, Mip and Other Secreted Virulence Factors at the Legionella Pneumophila Surface. Frontiers in Cellular and Infection Microbiology 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-L.; Li, D.; Zhan, X.-Y. Concept about the Virulence Factor of Legionella. Microorganisms 2022, 11, 74. [Google Scholar] [CrossRef] [PubMed]
- International Organization for Standardization ISO 19458:2006 Water Quality—Sampling for Microbiological Analysis. Available online: https://www.iso.org/standard/33845.html. (accessed on 15 November 2022).
- International Organization for Standardization ISO 11731:2017 Water Quality—Enumeration of Legionella. Available online: https://www.iso.org/standard/61782.html. (accessed on 15 November 2022).
- Valciņa, O.; Pūle, D.; Ķibilds, J.; Lazdāne, A.; Trofimova, J.; Makarova, S.; Konvisers, G.; Ķimse, L.; Krūmiņa, A.; Bērziņš, A. Prevalence and Genetic Diversity of Legionella Spp. In Hotel Water-Supply Systems in Latvia. Microorganisms 2023, 11, 596. [Google Scholar] [CrossRef]
- Prjibelski, A.D.; Puglia, G.D.; Antipov, D.; Bushmanova, E.; Giordano, D.; Mikheenko, A.; Vitale, D.; Lapidus, A. Extending RnaSPAdes Functionality for Hybrid Transcriptome Assembly. BMC Bioinformatics 2020, 21. [Google Scholar] [CrossRef]
- Gaia, V.; Fry, N.K.; Afshar, B.; Luck, P.C.; Meugnier, H.; Etienne, J.; Peduzzi, R.; Harrison, T.G. Consensus Sequence-Based Scheme for Epidemiological Typing of Clinical and Environmental Isolates of Legionella Pneumophila. Journal of Clinical Microbiology 2005, 43, 2047–2052. [Google Scholar] [CrossRef]
- Mentasti, M.; Underwood, A.; Lück, C.; Kozak-Muiznieks, N.A.; Harrison, T.G.; Fry, N.K. Extension of the Legionella Pneumophila Sequence-Based Typing Scheme to Include Strains Carrying a Variant of the N-Acylneuraminate Cytidylyltransferase Gene. Clinical Microbiology and Infection 2014, 20, 435–441. [Google Scholar] [CrossRef]
- Moran-Gilad, J.; Prior, K.; Yakunin, E.; Harrison, T.G.; Underwood, A.; Lazarovitch, T.; Valinsky, L.; Lück, C.; Krux, F.; Agmon, V.; et al. Design and Application of a Core Genome Multilocus Sequence Typing Scheme for Investigation of Legionnaires’ Disease Incidents. Eurosurveillance 2015, 20. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid Large-Scale Prokaryote Pan Genome Analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2–Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. VFDB 2022: A General Classification Scheme for Bacterial Virulence Factors. Nucleic Acids Research 2021, 50, D912–D917. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A Greedy Algorithm for Aligning DNA Sequences. Journal of Computational Biology 2000, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Samrakandi, M.M.; Cirillo, S.L.G.; Ridenour, D.A.; Bermudez, L.E.; Cirillo, J.D. Genetic and Phenotypic Differences between Legionella Pneumophila Strains. Journal of Clinical Microbiology 2002, 40, 1352–1362. [Google Scholar] [CrossRef] [PubMed]
- Slater, G.; Birney, E. Automated Generation of Heuristics for Biological Sequence Comparison. BMC Bioinformatics 2005, 6, 31. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. Journal of Antimicrobial Chemotherapy 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Conover, G.M.; Isberg, R.R. Legionella Pneumophila EnhC Is Required for Efficient Replication in Tumour Necrosis Factor α-Stimulated Macrophages. Cellular Microbiology 2008, 10, 1906–1923. [Google Scholar] [CrossRef]
- Hoffman, P.C.; Garduno, R.A. Surface-Associated Heat Shock Proteins Of Legionella Pneumophila and Helicobacter Pylori: Roles in Pathogenesis and Immunity. Infectious Diseases in Obstetrics and Gynecology 1999, 7, 58–63. [Google Scholar] [CrossRef]
- Helbig, J.H.; König, B.; Knospe, H.; Bubert, B.; Yu, C.; Lück, C.P.; Riboldi-Tunnicliffe, A.; Hilgenfeld, R.; Jacobs, E.; Hacker, J.; et al. The PPIase Active Site of Legionella Pneumophila Mip Protein Is Involved in the Infection of Eukaryotic Host Cells. Biological Chemistry 2003, 384. [Google Scholar] [CrossRef]
- Bellinger-Kawahara, C.; Horwitz, M.A. Complement Component C3 Fixes Selectively to the Major Outer Membrane Protein (MOMP) of Legionella Pneumophila and Mediates Phagocytosis of Liposome-MOMP Complexes by Human Monocytes. The Journal of Experimental Medicine 1990, 172, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- Gan, N.; Nakayasu, E.S.; Hollenbeck, P.J.; Luo, Z.-Q. Legionella Pneumophila Inhibits Immune Signalling via MavC-Mediated Transglutaminase-Induced Ubiquitination of UBE2N. Nature Microbiology 2018, 4, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Losick, V.P.; Haenssler, E.; Moy, M.-Y.; Isberg, R.R. LnaB: A Legionella Pneumophila Activator of NF-ΚB. Cellular Microbiology 2010, 12, 1083–1097. [Google Scholar] [CrossRef] [PubMed]
- Bhogaraju, S.; Bonn, F.; Mukherjee, R.; Adams, M.; Pfleiderer, M.M.; Galej, W.P.; Matkovic, V.; Lopez-Mosqueda, J.; Kalayil, S.; Shin, D.; et al. Inhibition of Bacterial Ubiquitin Ligases by SidJ–Calmodulin Catalysed Glutamylation. Nature 2019, 572, 382–386. [Google Scholar] [CrossRef]
- Bandyopadhyay, P.; Lang, E.A.S.; Rasaputra, K.S.; Steinman, H.M. Implication of the VirD4 Coupling Protein of the Lvh Type 4 Secretion System in Virulence Phenotypes of Legionella Pneumophila. Journal of Bacteriology 2013, 195, 3468–3475. [Google Scholar] [CrossRef]
- Newton, H.J.; Browning, G.F.; Dao, J.; McAlister, A.D.; Sloan, J.; Cianciotto, N.P.; Hartland, E.L. Sel1 Repeat Protein LpnE Is a Legionella Pneumophila Virulence Determinant That Influences Vacuolar Trafficking. 2007, 75, 5575–5585. [Google Scholar] [CrossRef]
- Rossier, O.; Starkenburg, S.R.; Cianciotto, N.P. Legionella Pneumophila Type II Protein Secretion Promotes Virulence in the A/J Mouse Model of Legionnaires’ Disease Pneumonia. Infection and Immunity 2003, 72, 310–321. [Google Scholar] [CrossRef]
- Cirillo, S.L.G.; Bermudez, L.E.; El-Etr, S.H.; Duhamel, G.E.; Cirillo, J.D. Legionella Pneumophila Entry Gene RtxA Is Involved in Virulence. Infection and Immunity 2001, 69, 508–517. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) V5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Research 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Zhou, Z.; Alikhan, N.-F.; Sergeant, M.J.; Luhmann, N.; Vaz, C.; Francisco, A.P.; Carriço, J.A.; Achtman, M. GrapeTree: Visualization of Core Genomic Relationships among 100,000 Bacterial Pathogens. Genome Research 2018, 28, 1395–1404. [Google Scholar] [CrossRef]
- Dilger, T.; Melzl, H.; Gessner, A. Legionella Contamination in Warm Water Systems: A Species-Level Survey. International Journal of Hygiene and Environmental Health 2018, 221, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Felice, A.; Franchi, M.; De Martin, S.; Vitacolonna, N.; Iacumin, L.; Civilini, M. Environmental Surveillance and Spatio-Temporal Analysis of Legionella Spp. In a Region of Northeastern Italy (2002–2017). PLoS ONE 2019, 14, e0218687. [Google Scholar] [CrossRef] [PubMed]
- Gleason, J.A.; Conner, L.E.; Ross, K.M. Associations of Household Factors, Hot Water Temperature, and Chlorine Residual with Legionella Occurrence in Single-Family Homes in New Jersey. Science of The Total Environment 2023, 870, 161984. [Google Scholar] [CrossRef] [PubMed]
- Valciņa, O.; Pūle, D.; Mališevs, A.; Trofimova, J.; Makarova, S.; Konvisers, G.; Bērziņš, A.; Krūmiņa, A. Co-Occurrence of Free-Living Amoeba and Legionella in Drinking Water Supply Systems. Medicina 2019, 55, 492. [Google Scholar] [CrossRef] [PubMed]
- Cabinet of Ministers of Latvia MK Nr. 906, 2010.28.09. Rules of Sanitary Maintenance of the Residential House. Available online: https://likumi.lv/ta/id/218830-dzivojamas-majas-sanitaras-apkopes-noteikumi (accessed on 11 May 2023).
- Valciņa, O.; Pūle, D.; Lucenko, I.; Krastiņa, D.; Šteingolde, Ž.; Krūmiņa, A.; Bērziņš, A. Legionella Pneumophila Seropositivity-Associated Factors in Latvian Blood Donors. International Journal of Environmental Research and Public Health 2015, 13, 58. [Google Scholar] [CrossRef] [PubMed]
- Bešić, A.; Karakaš, S.; Obradović, Z.; Mušović, A.; Hrapović, E. Travel-Related Epidemiological Studies of Legionellosis in Federation of Bosnia and Herzegovina. Health and Technology 2021, 11, 971–979. [Google Scholar] [CrossRef]
- Raphael, B.H.; Huynh, T.; Brown, E.; Smith, J.C.; Ruberto, I.; Getsinger, L.; White, S.; Winchell, J.M. Culture of Clinical Specimens Reveals Extensive Diversity of Legionella Pneumophila Strains in Arizona. mSphere 2019, 4. [Google Scholar] [CrossRef]
- Jiang, L.; Zhao, S.; Cai, X.; Mu, D.; Zhang, X.; Kang, J.; Zhao, L.; Chen, Y. Sequence-Based Typing of Clinical and Environmental Legionella Pneumophila Isolates in Shenyang, China. Enfermedades infecciosas y microbiologia clinica (English ed.) 2021, 39, 383–389. [Google Scholar] [CrossRef]
- Keše, D.; Obreza, A.; Rojko, T.; Kišek, T.C. Legionella Pneumophila—Epidemiology and Characterization of Clinical Isolates, Slovenia, 2006–2020. Diagnostics 2021, 11, 1201. [Google Scholar] [CrossRef]
- Lévesque, S.; Lalancette, C.; Bernard, K.; Pacheco, A.L.; Dion, R.; Longtin, J.; Tremblay, C. Molecular Typing of Legionella Pneumophila Isolates in the Province of Quebec from 2005 to 2015. PLoS ONE 2016, 11, e0163818. [Google Scholar] [CrossRef]
- Best, A.; Abu Kwaik, Y. Evolution of the Arsenal of Legionella Pneumophila Effectors to Modulate Protist Hosts. mBio 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; O’Connor, T.J. Beyond Paralogs: The Multiple Layers of Redundancy in Bacterial Pathogenesis. Frontiers in Cellular and Infection Microbiology 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Ridenour, D.A.; Cirillo, S.L.G.; Feng, S.; Samrakandi, M.M.; Cirillo, J.D. Identification of a Gene That Affects the Efficiency of Host Cell Infection by Legionella Pneumophila in a Temperature-Dependent Fashion. Infection and Immunity 2003, 71, 6256–6263. [Google Scholar] [CrossRef] [PubMed]
- Sawczyn-Domańska, A. Detection of Legionella Spp. And Occurrence of Virulence Genes: Lvh, RtxA and EnhC in Water Samples from Artificial Water Systems. Annals of Agricultural and Environmental Medicine 2021, 28, 617–620. [Google Scholar] [CrossRef]
- Zeng, L.Z.; Liao, H.Y.; Luo, L.Z.; He, S.S.; Qin, T.; Zhou, H.J.; Li, H.X.; Chen, D.L.; Chen, J.P. An Investigation on the Molecular Characteristics and Intracellular Growth Ability among Environmental and Clinical Isolates of Legionella Pneumophila in Sichuan Province, China. Biomedical and Environmental Sciences 2019, 32, 520–530. [Google Scholar] [CrossRef]
- D’Auria, G.; Jiménez, N.; Peris-Bondia, F.; Pelaz, C.; Latorre, A.; Moya, A. Virulence Factor Rtx in Legionella Pneumophila, Evidence Suggesting It Is a Modular Multifunctional Protein. BMC Genomics 2008, 9. [Google Scholar] [CrossRef]
Figure 1.
A minimum spanning tree of 58 L. pneumophila isolates from water samples taken in apartment buildings in Latvia, based on cgMLST. The node sizes are proportional to the numbers of isolates sharing an identical pattern, node colors represent geographical origin of the isolates and node labels indicate STs of the L. pneumophila isolates.
Figure 1.
A minimum spanning tree of 58 L. pneumophila isolates from water samples taken in apartment buildings in Latvia, based on cgMLST. The node sizes are proportional to the numbers of isolates sharing an identical pattern, node colors represent geographical origin of the isolates and node labels indicate STs of the L. pneumophila isolates.
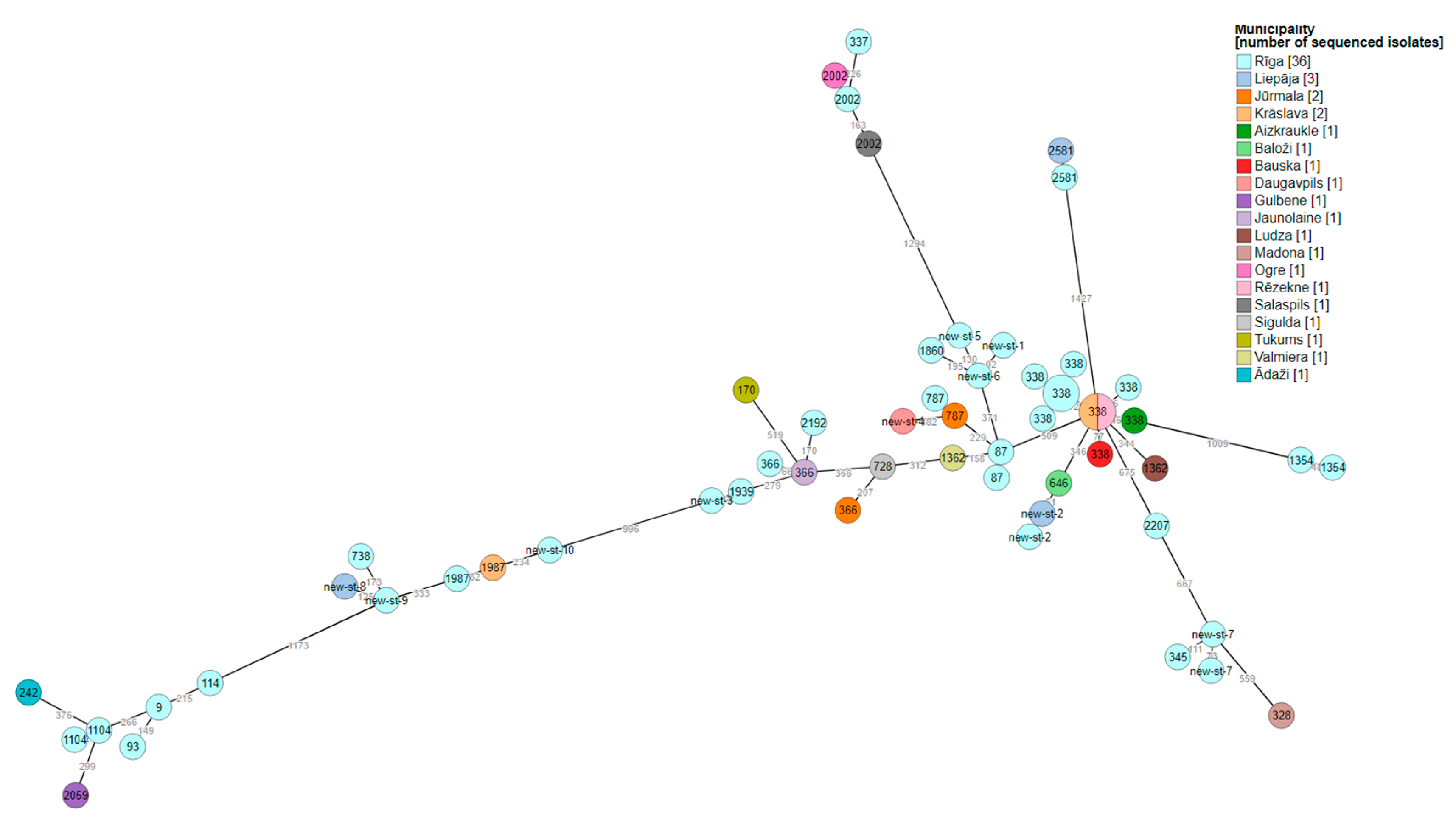
Figure 2.
A maximum-likelihood tree built from core gene alignment of 58 L. pneumophila strains. The municipality of sample origin and SBT type is given in brackets after the name of each node, with strains from buildings linked to LD cases highlighted in bold. Coloured squares indicate the serogroup of each isolate. The panel of green squares shows the distribution of L. pneumophila virulence and antimicrobial resistance genes among all isolates. Green filled squares indicate the presence of the gene while empty squares indicate the absence of the gene in the respective L. pneumophila isolate.
Figure 2.
A maximum-likelihood tree built from core gene alignment of 58 L. pneumophila strains. The municipality of sample origin and SBT type is given in brackets after the name of each node, with strains from buildings linked to LD cases highlighted in bold. Coloured squares indicate the serogroup of each isolate. The panel of green squares shows the distribution of L. pneumophila virulence and antimicrobial resistance genes among all isolates. Green filled squares indicate the presence of the gene while empty squares indicate the absence of the gene in the respective L. pneumophila isolate.
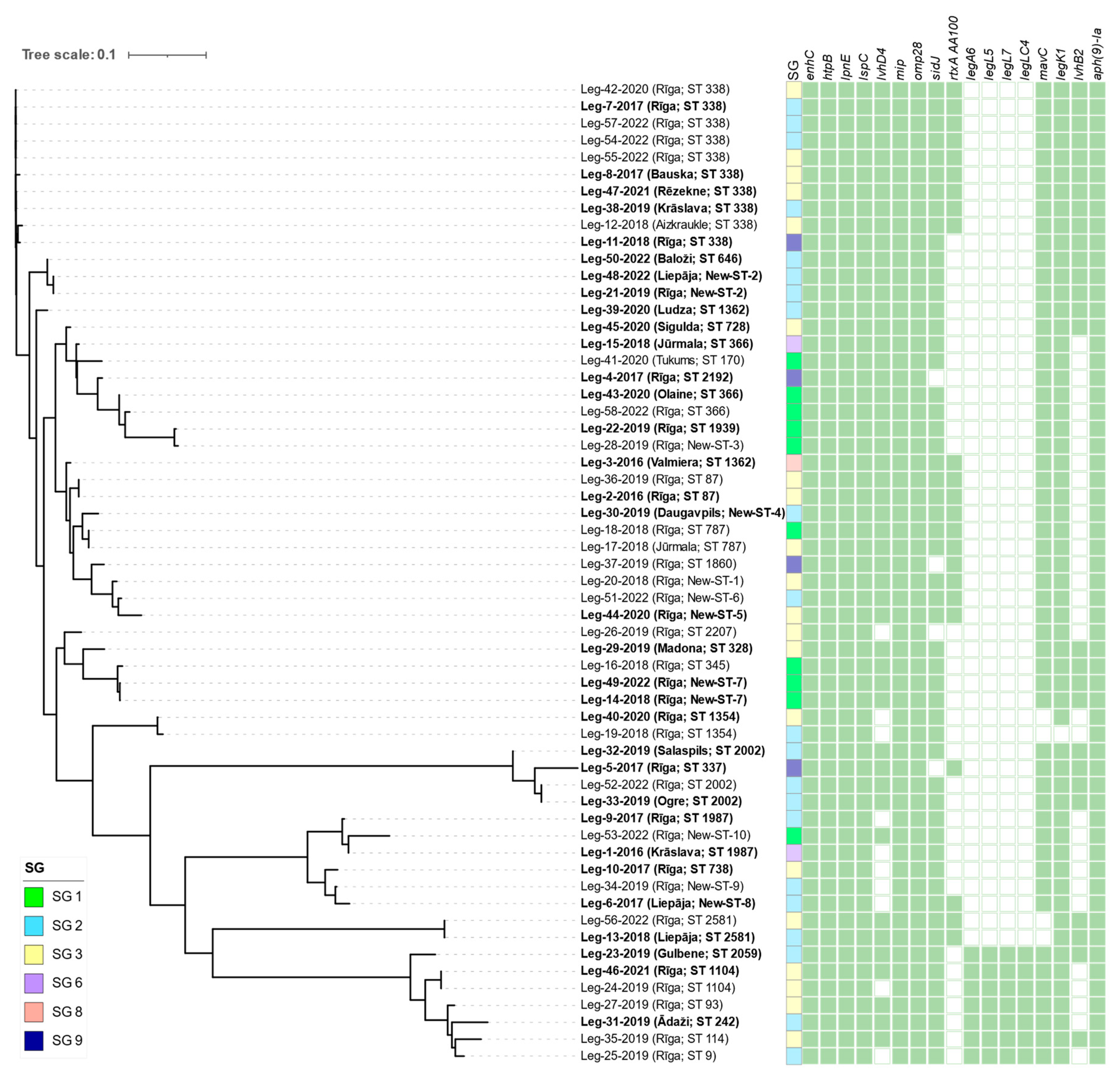

Table 1.
Legionella virulence factors.
| Gene | Virulence factor | Roles | |
| Bacterial surface structures | enhC | EnhC | Enhanced entry, trafficking of Legionella-containing vacuole [31] |
| htpB | Hsp60 | Attachment, modulation of invasion [32] | |
| mip | MIP | Penetration of the lung epithelial barrier [33] | |
| omp28 | MOMP | Mediating phagocytosis [34] | |
| T4SS effectors | mavC | MavC | Inhibiting host immunity [35] |
| legK1 | LegK1 | Activation of NF-kB [36] | |
| sidJ | SidJ | Calmodulin-activated glutamylase for SidE [37] | |
| lvhD4 | VirD4 | Coupling protein, reversing virulence defects [38] | |
| lpnE | LpnE | Entry into macrophages and epithelial cells; manipulate host cell trafficking [39] | |
| T2SS effectors | lsp | Lsp | Transport proteins from the periplasm to the extracellular space [40] |
| T1SS effectors | rtxA | RtxA | Ensures adherence and entry into host and enhances replication and cytotoxicity and pore forming [41] |
Table 2.
The prevalence of L. pneumophila in water samples from residential buildings.
| Buildings with known LD cases | Buildings without known LD cases | Total | ||||
| Samples tested/ positive samples (%) | Average water temperature, °C | Samples tested/ positive samples (%) | Average water temperature, °C | Samples tested/ positive samples (%) | Average water temperature, °C | |
| Cold water | 120/22 (18.3%) | 12.9 ± 0.4 | 44/19 (43.2%) | 15.6 ± 0.7 | 164/41 (25%) | 13.5 ± 0.3 |
| Hot water | 242/107 (44.2%) | 52.1 ± 0.4 | 86/49 (57.0%) | 45.8 ± 1.1 | 328/156 (47.6%) | 50.7 ± 0.4 |
| Total | 362/129 (35.6%) | -- | 130/68 (52.3%) | -- | 492/197 (40.0%) | -- |
Table 3.
The prevalence of L. pneumophila serogroups in water samples from buildings with and without previous episodes of LD.
Table 3.
The prevalence of L. pneumophila serogroups in water samples from buildings with and without previous episodes of LD.
| Serogroup | No. of L. pneumophila isolates (%) | ||
|---|---|---|---|
| Buildings with known LD cases | Buildings without known LD cases | Total | |
| SG 1 | 15 (11.6%) | 4 (6.0%) | 19 (9.7%) |
| SG 2 | 69 (53.5%) | 39 (58.2%) | 108 (55.1%) |
| SG 3 | 35 (27.1%) | 20 (29.9%) | 55 (28.1%) |
| SG 6 | 3 (2.3%) | 1 (1.5%) | 4 (2.0%) |
| SG 8 | 1 (0.8%) | -- | 1 (0.5%) |
| SG 9 | 2 (1.6%) | 2 (3.0%) | 4 (2.0%) |
| SG 3, SG 2 | 3 (2.3%) | -- | 3 (1.5%) |
| SG 3, SG 1 | -- | 1 (1.5%) | 1 (0.5%) |
| SG 3, SG 9 | 1 (0.8%) | -- | 1 (0.5%) |
Table 4.
The levels of colonization in water supply systems by L. pneumophila.
| Serogroup |
Levels of L. pneumophila colonization (min – max (average)), CFU/L |
||
| Buildings with known LD cases | Buildings without known LD cases | p-value | |
| SG 1 | 50 – 4.0×103 (8.4×102) | 4.0×102 – 5.5×103 (2.6×103) |
0.056 |
| SG 2 | 50 – 1.3×103 (1.5×103) |
50 – 6.4×103 (2.1×103) |
0.158 |
| SG 3 | 50 - 1.7×104 (2.8×103) |
1.0×102 – 3.9×103 (8.2×102) |
0.033 |
| Total | 50 - 1.7×104 (2.0×103) |
50 – 6.4×103 (1.7×103) |
0.574 |
Table 5.
Characteristics of L. pneumophila isolated from water-supply systems in Latvia between 2016 and 2022.
Table 5.
Characteristics of L. pneumophila isolated from water-supply systems in Latvia between 2016 and 2022.
| Isolate Id | Year of sampling |
Municipality | SG | Linked with LD cases | Allelic profile | Sequence type | Number of observed virulence genes |
|---|---|---|---|---|---|---|---|
| Leg-1-2016 | 2016 | Krāslava | 6 | Yes | 7,6,17,3,50,11,9 | 1987 | 377 |
| Leg-2-2016 | 2016 | Rīga | 3 | Yes | 2,10,3,28,9,4,13 | 87 | 372 |
| Leg-3-2016 | 2016 | Valmiera | 8 | Yes | 2,10,3,28,9,4,207 | 1362 | 372 |
| Leg-4-2017 | 2017 | Rīga | 9 | Yes | 2,10,24,3,9,4,6 | 2192 | 370 |
| Leg-5-2017 | 2017 | Rīga | 9 | Yes | 10,22,7,28,16,18,6 | 337 | 358 |
| Leg-6-2017 | 2017 | Liepāja | 2 | Yes | 7,10,17,6,9,11,9 | New-ST-8 | 378 |
| Leg-7-2017 | 2017 | Rīga | 2 | Yes | 2,10,15,28,9,4,13 | 338 | 375 |
| Leg-8-2017 | 2017 | Bauska | 3 | Yes | 2,10,15,28,9,4,13 | 338 | 378 |
| Leg-9-2017 | 2017 | Rīga | 2 | Yes | 7,6,17,3,50,11,9 | 1987 | 378 |
| Leg-10-2017 | 2017 | Rīga | 3 | Yes | 7,10,17,28,17,11,9 | 738 | 376 |
| Leg-11-2018 | 2018 | Rīga | 9 | Yes | 2,10,15,28,9,4,13 | 338 | 378 |
| Leg-12-2018 | 2018 | Aizkraukle | 3 | No | 2,10,15,28,9,4,13 | 338 | 378 |
| Leg-13-2018 | 2018 | Liepāja | 2 | Yes | 2,32,20,38,34,35,219 | 2581 | 344 |
| Leg-14-2018 | 2018 | Rīga | 1 | Yes | 6,10,19,3,98,4,novel neuA allele | New-ST-7 | 374 |
| Leg-15-2018 | 2018 | Jūrmala | 6 | Yes | 2,10,3,3,9,4,6 | 366 | 374 |
| Leg-16-2018 | 2018 | Rīga | 1 | No | 6,10,19,3,19,4,11 | 345 | 374 |
| Leg-17-2018 | 2018 | Jūrmala | 3 | No | 2,10,1,3,9,4,3 | 787 | 372 |
| Leg-18-2018 | 2018 | Rīga | 1 | No | 2,10,1,3,9,4,3 | 787 | 372 |
| Leg-19-2018 | 2018 | Rīga | 2 | No | 2,10,24,28,4,4,207 | 1354 | 312 |
| Leg-20-2018 | 2018 | Rīga | 3 | No | 2,10,3,3,50,4,3 | New-ST-1 | 372 |
| Leg-21-2019 | 2019 | Rīga | 2 | Yes | 2,10,21,28,9,4,6 | New-ST-2 | 374 |
| Leg-22-2019 | 2019 | Rīga | 1 | Yes | 2,10,1,3,9,4,6 | 1939 | 374 |
| Leg-23-2019 | 2019 | Gulbene | 2 | Yes | 3,4,1,6,35,9,220 | 2059 | 415 |
| Leg-24-2019 | 2019 | Rīga | 3 | No | 3,13,1,28,14,9,13 | 1104 | 397 |
| Leg-25-2019 | 2019 | Rīga | 2 | No | 3,10,1,3,14,9,11 | 9 | 396 |
| Leg-26-2019 | 2019 | Rīga | 3 | No | 2,10,19,28,19,4,3 | 2207 | 360 |
| Leg-27-2019 | 2019 | Rīga | 3 | No | 3,10,1,28,14,9,13 | 93 | 415 |
| Leg-28-2019 | 2019 | Rīga | 1 | No | 2,10,17,3,9,4,6 | New-ST-3 | 373 |
| Leg-29-2019 | 2019 | Madona | 3 | Yes | 6,10,19,28,19,4,9 | 328 | 375 |
| Leg-30-2019 | 2019 | Daugavpils | 2 | Yes | 2,10,17,3,9,4,9 | New-ST-4 | 369 |
| Leg-31-2019 | 2019 | Ādaži | 2 | Yes | 3,10,1,28,1,9,3 | 242 | 415 |
| Leg-32-2019 | 2019 | Salaspils | 2 | Yes | 10,22,7,28,16,18,8 | 2002 | 363 |
| Leg-33-2019 | 2019 | Ogre | 2 | Yes | 10,22,7,28,16,18,8 | 2002 | 357 |
| Leg-34-2019 | 2019 | Rīga | 2 | No | 7,10,17,6,17,11,9 | New-ST-9 | 377 |
| Leg-35-2019 | 2019 | Rīga | 3 | No | 3,6,1,6,14,11,9 | 114 | 415 |
| Leg-36-2019 | 2019 | Rīga | 3 | No | 2,10,3,28,9,4,13 | 87 | 373 |
| Leg-37-2019 | 2019 | Rīga | 9 | No | 2,10,3,3,9,4,207 | 1860 | 371 |
| Leg-38-2019 | 2019 | Krāslava | 2 | Yes | 2,10,15,28,9,4,13 | 338 | 378 |
| Leg-39-2020 | 2020 | Ludza | 2 | Yes | 2,10,3,28,9,4,207 | 1362 | 376 |
| Leg-40-2020 | 2020 | Rīga | 3 | Yes | 2,10,24,28,4,4,207 | 1354 | 328 |
| Leg-41-2020 | 2020 | Tukums | 1 | Yes | 2,10,3,10,9,4,11 | 170 | 375 |
| Leg-42-2020 | 2020 | Rīga | 3 | No | 2,10,15,28,9,4,13 | 338 | 371 |
| Leg-43-2020 | 2020 | Olaine | 1 | Yes | 2,10,3,3,9,4,6 | 366 | 379 |
| Leg-44-2020 | 2020 | Rīga | 3 | Yes | 2,22,3,28,50,4,3 | New-ST-5 | 371 |
| Leg-45-2020 | 2020 | Sigulda | 3 | Yes | 2,10,3,28,9,4,3 | 728 | 373 |
| Leg-46-2021 | 2021 | Rīga | 3 | Yes | 3,13,1,28,14,9,13 | 1104 | 412 |
| Leg-47-2021 | 2021 | Rēzekne | 3 | Yes | 2,10,15,28,9,4,13 | 338 | 378 |
| Leg-48-2022 | 2022 | Liepāja | 2 | Yes | 2,10,21,28,9,4,6 | New-ST-2 | 372 |
| Leg-49-2022 | 2022 | Rīga | 1 | Yes | 6,10,19,3,17,4,11 | New-ST-7 | 374 |
| Leg-50-2022 | 2022 | Baloži | 2 | Yes | 2,10,21,28,9,4,13 | 646 | 375 |
| Leg-51-2022 | 2022 | Rīga | 2 | No | 2,10,3,3,50,4,6 | New-ST-6 | 372 |
| Leg-52-2022 | 2022 | Rīga | 2 | No | 10,22,7,28,16,18,8 | 2002 | 360 |
| Leg-53-2022 | 2022 | Rīga | 1 | No | 7,6,17,3,50,4,9 | New-ST-10 | 390 |
| Leg-54-2022 | 2022 | Rīga | 2 | No | 2,10,15,28,9,4,13 | 338 | 375 |
| Leg-55-2022 | 2022 | Rīga | 3 | No | 2,10,15,28,9,4,13 | 338 | 375 |
| Leg-56-2022 | 2022 | Rīga | 3 | No | 2,32,20,38,34,35,219 | 2581 | 344 |
| Leg-57-2022 | 2022 | Rīga | 2 | No | 2,10,15,28,9,4,13 | 338 | 375 |
| Leg-58-2022 | 2022 | Rīga | 1 | No | 2,10,3,3,9,4,6 | 366 | 375 |
Table 6.
The relative frequency of 11 virulence genes in L. pneumophila isolates.
| Virulence gene | No. of positive / No. of sequenced isolates | Relative frequency, % | ||||||||
| Overall | Isolates from buildings linked to LD cases N=34 |
Isolates from buildings not linked to LD cases N=24 |
SG 1 isolates N=10 |
SG 2 isolates N=21 |
SG 3 isolates N=20 |
SG 6 isolates N=2 |
SG 8 isolates N=1 |
SG 9 isolates N=4 |
||
| enhC | 58/58 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| htpB | 58/58 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| omp28 | 58/58 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| mip | 58/58 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| mavC | 54/58 | 93 | 94 | 92 | 100 | 90 | 90 | 100 | 100 | 100 |
| legK1 | 57/58 | 98 | 100 | 96 | 100 | 95 | 100 | 100 | 100 | 100 |
| sidJ | 54/58 | 93 | 94 | 92 | 100 | 100 | 95 | 100 | 100 | 25 |
| lvhD4 | 48/58 | 83 | 85 | 79 | 100 | 76 | 80 | 50 | 100 | 100 |
| lpnE | 58/58 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| lspC | 58/58 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| rtxA AA100 | 23/58 | 40 | 32 | 50 | 10 | 38 | 55 | 0 | 100 | 50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



