Submitted:
26 May 2023
Posted:
29 May 2023
You are already at the latest version
Abstract
Chitooligosaccharides (COS) are -1,4-linked homo-oligosaccharides of N-acetylglucosamine (GlcNAc) or glucosamine (GlcN), and also include hetero-oligosaccharides composed of GlcNAc and GlcN. These sugars are of practical importance because of their various biological activities, such as anti-microbial, anti-inflammatory, anti-oxidant and anti-tumor activities, as well as triggering the innate immunity in plants. The reported data on bioactivities of COS used to contain some uncertainties or contradictions, because the experiments were conducted with poorly characterized COS mixtures. Recently, COS have been satisfactorily characterized with respect to their structures, especially the degree of polymerization (DP) and degree of N-acetylation (DA); thus, the structure-bioactivity relationship of COS has become more unambiguous. To date, various green-chemical strategies involving enzymatic synthesis of COS with designed sequences and desired biological activities have been developed. The enzymatic strategies could involve transglycosylation or glycosynthase reactions using reducing end-activated sugars as the donor substrates and chitinase/chitosanase and their mutants as the biocatalysts. Site-specific chitin deacetylases were also proposed to be applicable for this purpose. Furthermore, to improve the yields of the COS products, metabolic engineering techniques could be applied. The above-mentioned approaches will provide the opportunity to produce tailor-made COS, leading to the enhanced utilization of chitin biomass.
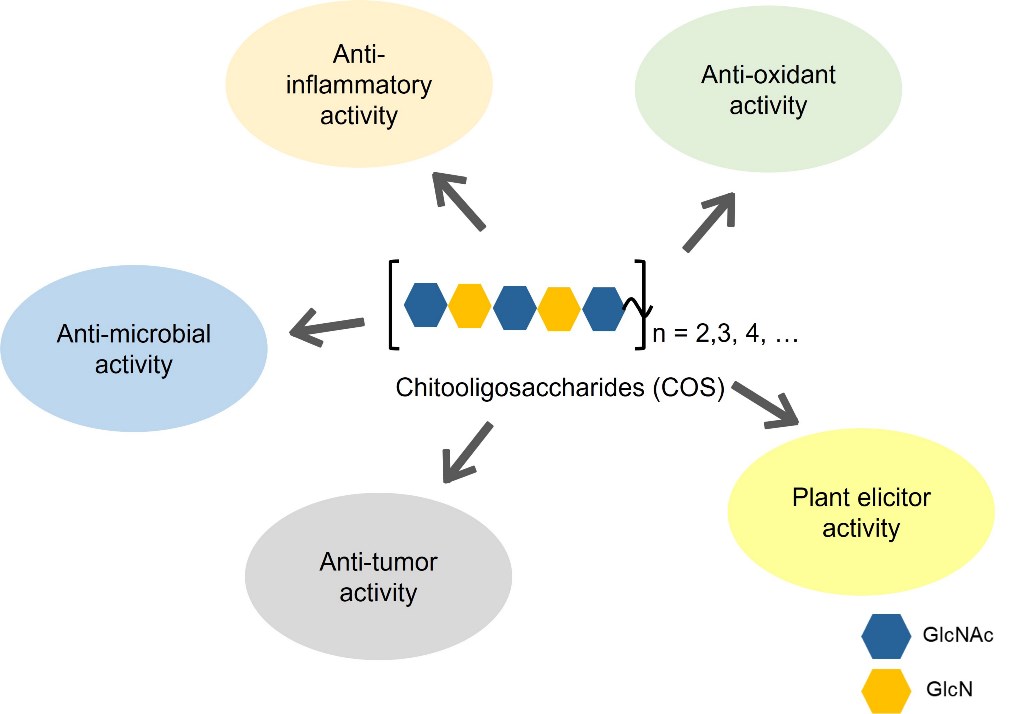
Keywords:
chitooligosaccharides
; biological activities
; chitinase
; chitosanase
; transglycosylation
; glycosynthase
1. Introduction
Chitin, a b-1,4-linked polysaccharide of N-acetylglucosamine (GlcNAc), is the second most abundant biomass on the earth, next to cellulose. Although cellulose is most efficiently utilized as the essential material in our daily life, the utilization of chitin is much less intensive. Many attempts have been conducted to enhance the utilization of chitin [1], but effective approaches still remain to be developed. In previous attempts at enhancing chitin utilization, chitin and its de-N-acetylated derivatives, chitosan, were degraded to chitooligosaccharides (COS) by acid or enzymatic hydrolysis [2], and the oligosaccharides characterized with respect to their anti-microbial, anti-inflammatory, antioxidant, immune-potentiating and anti-tumor activities [3]. Nowadays, COS are well known to exhibit such biological activities in vitro or in vivo. These activity data were obtained using COS mixtures, which included b-1,4-linked oligosaccharides, composed of N-acetylglucosamine (GlcNAc), glucosamine (GlcN) or both (Figure 1).
However, recent developments in analytical methods for carbohydrate structures have enabled exact determinations of the degrees of polymerization (DP) and of N-acetylation (DA), as well as sequences, which can be analyzed by NMR, IR and mass spectrometry [4,5]. In fact, the biological activities of COS were found to be controlled by the three structural factors: DP, DA and sequence. For example, the antioxidant and anti-inflammatory activities of COS were dependent on their DP, as well as on DA [6,7]. Similar dependencies were also observed for other biological activities, as described previously [3]. Furthermore, it was proposed that the specific sequences composed of GlcNAc and GlcN, so called “ChitoCode”, were recognized by some proteins with significant functions in fungal interactions with other organisms [8]. Similar mechanisms of molecular recognition may exist in other interaction systems composed of living organisms that have chitin/chitosan as structural components. To obtain COS with desired biological activities, it has now become possible to design synthesized COS with specified sequences, so called tailor-made COS. Here, we have made a comprehensive review of the biological activities of COS and discussed their structure-function relationship. The review underlines the green-chemical methods reported for the synthesis of pure COS with designated sequences. Especially, enzymatic and metabolic engineering are considered promising approaches for enhanced production of high quality COS with useful bioactive properties (Figure 2).
2. Biological activities of COS
2.1. Anti-microbial activity
COS have been recognized to exhibit antimicrobial activities, inhibiting the growth of a variety of phytopathogenic and animal-pathogenic fungi, bacteria and viruses. The antimicrobial effects of COS with higher, medium and lower molecular weights were respectively examined, revealing that COS with higher molecular weights had greater anti-microbial effects than those with medium and lower ones [9]. Higher DPs were also reported to be beneficial for exerting the antibacterial activities of COS [10,11]. It is likely that the positive charges of COS interact with negative charges on the bacterial cell surfaces, leading to inhibition of cell growth. In contrast, the shorter COS (DA = 0.15-0.5; DP = 2, 3, and 4) and low molecular weight chitosan produced by a chitosanase from Bacillus thuringiensis B-387 from chitosan polysaccharide, of which the average MWs were in a range of 14-46 kDa, exhibited greater antifungal and antibacterial activities in vitro in comparison with the initial polysaccharides [12]. Ganan et al. analyzed the antifungal activities of soluble and well-defined COS. Determination of the antifungal activities of COS fractions with varying average DPs (17, 31, 54, and 62) indicated that fractions with an intermediate DPs (31 and 54) had the strongest inhibitory effects [13]. Another report proposed that short-chain COS exhibited high fungicidal activities toward Candida species [14]. The contradictions in these findings may arise from differences in COS-preparation methods and the targeted microorganisms, as well as in experimental conditions. The antimicrobial activities may depend not only on the electrostatic interactions of free amino groups of COS but also on the interaction of GlcNAc residues with the hydrophobic regions of microbial cell surfaces. A summary of the antimicrobial activities of COS is presented in Table 1.
2.2. Anti-inflammatory activity
Four COS with different DAs (0, 0.12, 0.5, and 0.85) and the same DP distributions were employed for assays of anti-inflammatory activity [15]. The results revealed that COS with a DA of 0.12 had greater anti-inflammatory activity than COS with other DAs. Another study demonstrated that low-MW COS with DP of 3-5 obtained from chitosan were found to be effective against allergic inflammation in vivo [16]. In support of the fact that low-MW COS possess the highest anti-inflammatory activity, Pangestuti et al. reported that depolymerized products from chitosan (MW < 10 kDa) reduced the production of nitric oxide and prostaglandin E2 by inhibiting inducible nitric oxide synthase (iNOS) and cyclooxygenase expression [17]. The effect of COS (MW < 1 kDa) on nitric oxide production was further studied by Wei et al. [18]. Pretreatment with COS could inhibit the production of nitric oxide by suppressing iNOS expression in activated microglial cells. Sanchez et al. [19] prepared COS composed of 42% fully de-N-acetylated and 54% mono-N-acetylated oligomers through a single enzymatic hydrolysis step. This combination of COS demonstrated anti-inflammatory effects [19]. Taken together, COS with lower DPs (3-5) and a lower DA significantly attenuated the inflammation. It is notable, however, that completely de-N-acetylated COS (DA = 0) exhibited no effective anti-inflammatory activity. The anti-inflammatory effects of COS are summarized in Table 2.
2.3. Anti-oxidant activity
Fully N-acetylated COS (MW = 229.21–593.12 Da; DP = 1, 2 and 3) produced by the acid hydrolysis of crab chitin, which may contain a small fraction of de-N-acetylated residues, was found to inhibit myeloperoxidase activity and decrease free radicals in human myeloid cells, HL-60 or suppress oxidation of DNA and membrane proteins in mouse macrophages Raw 264.7 [20].In the same work, COS were shown to also directly scavenge free radical-induced DNA oxidation generated by 2′,7′-dichlorofluorescein, as well as increase intracellular glutathione level, which confirms the antioxidant properties of COS in living cells. Enzymatic hydrolysates from housefly larvae powder (HLP) contained mainly hetero-COS composed of GlcNAc and GlcN (DP = 2–6) [21]. Maximum production of COS with a DP of 5 (51.2 μg·mL−1) was achieved by hydrolysis of HLP for 72 h and COS with a DP of 5 exhibited the strongest hydroxyl-scavenging ability and reducing power. COS obtained from chitosan with a DA of 0.05 have a protective effect on H2O2-induced human umbilical vein endothelial cell apoptosis, and also from H2O2-induced oxidative damage in endothelial cells [22,23]. Five COS from chitosan with increasing DPs (3-7) were examined and showed effects on the antioxidant activity of soybean (Glycine max) seeds during germination [24]. The COS of different DPs exhibited varying influences on the antioxidant activity in soybean seeds. Especially, COS with DP = 6 exerted a strong effect and significantly increased the antioxidant activity. Hao et al. [25] produced COS dimers with different sequences, GlcNAc-GlcNAc, GlcN-GlcNAc, GlcNAc-GlcN and GlcN-GlcN, by the enzymatic method, and the antioxidant activities of all four dimers were studied. The amino group at the reducing end played a vital role in scavenging superoxide radicals. Furthermore, they found that GlcN-GlcN (DD) showed the highest 2,2-diphenyl-1-picrylhydrazyl scavenging activity. Although antioxidant activities were dependent on the radical species used in the experiments, GlcN residues in COS were likely to dominate their antioxidant activity. COS with a DP of 5-6 appeared to have higher antioxidant activity [21,24]. Antioxidant activity of COS are summarized in Table 3.
2.4. Anti-tumor activity
Fully N-acetylated COS with a DP of 6, (GlcNAc)6, was found to display an anti-metastatic effect on tumor cells. It was reported that approximately 1 mg.kg−1 of (GlcNAc)6 administered intravenously resulted in a significant decrease in the pulmonary nodules formed from the Lewis lung carcinoma in mice [26]. However, the antitumor actions of COS in vivo included various biological processes, such as absorption of COS through the intestinal epithelium, distribution and accumulation in the kidney, liver and spleen, enzymatic transformation, and elimination from the body. The DP, DA and sequence of COS may significantly affect the individual processes. Therefore, at present it is impossible to define clearly the structure-activity relationship for antitumor activity in vivo. Salah et al. [27] reported that low-MW chitin (2480 Da) obtained from shrimp shells was more effective against a human leukemia cell line than the higher MW products (338 kDa chitin and 12 kDa chitosan). Similarly, COS with high MW and high DA were 50% less effective in tumor suppression than COS with low MW and low DA [28]. A decrease in MW or DA of COS resulted in enhancement of the in vitro anticancer activity. However, it appeared that a DP of at least 6 is required to exert significant antitumor activity [3,29]. These findings on COS antitumor activities were recently reviewed by Zhai et al. [30]. The anti-tumor activities of COS are outlined in Table 4.
2.5. Plant elicitor activity
Vander et al. [31] evaluated the capabilities of (GlcNAc)n (n = 4, 5, 6, 7, 8, 9, and 10), (GlcN)n (n = 5, 6, and 7) and partially N-acetylated chitosan to induce phenylalanine ammonia-lyase (PAL) and peroxidase (POD) in healthy wheat leaves. They found that purified (GlcN)n were not active as plant elicitors, whereas purified (GlcNAc)n (n ≥ 7) strongly elicited POD activities, but not PAL activities. Partially N-acetylated chitosans elicited both PAL and POD activities, and maximum elicitation was observed with chitosan oligosaccharides with intermediate DAs. Ramakrishna et al. [32] reported the elicitor activities in rice seedlings of (GlcNAc)n (n = 5-7) produced by a hypertransglycosylating mutant of a chitinase from Serratia proteamaculans. (GlcNAc)7 strongly induced an oxidative burst response, as well as peroxidase and PAL activities. These reports confirmed that (GlcNAc)n (n = 5-7) are most effective in eliciting an immune response in plants. A chitin elicitor receptor kinase 1 (CERK1) was found to be essential for chitin perception by plant cells [33]. CERK1 was composed of an extracellular domain (ectodomain) with three-contiguous LysM motifs (LysM1, LysM2, and LysM3), a transmembrane domain and an intracellular kinase domain. Crystal structure analysis of the COS-bound ectodomain of CERK1 from Arabidopsis thaliana (AtCERK1-ECD) [34] suggested that (GlcNAc)8 acted as a bivalent ligand that bound to the two AtCERK1-ECD proteins through a continuous groove formed between one LysM2 and another LysM2, inducing the homo-dimerization of two AtCERK1-ECD proteins. This dimerization formed an active receptor complex and was regarded as crucial for the immune response. In A. thaliana, (GlcNAc)8 binding to AtLYK5, another receptor containing three LysM motifs, induced the homo-dimerization, recruiting the two AtCERK1 proteins and eventually forming an active receptor complex [35]. In rice, a chitin elicitor binding protein (OsCEBiP) composed of three LysM motifs [36] may first bind the (GlcNAc)8 elicitor, inducing the dimerization of OsCEBiP, and subsequently forming a hetero-tetramer receptor complex composed of two OsCEBiP and two OsCERK1. Hayafune et al. [37] found that hetero-COS DP-8 containing alternating GlcN and GlcNAc (DADADADA) did not elicit an immune response in rice plants, and hence was likely to inhibit formation of the hetero-tetramer complex. A hetero-COS with a DP of 6, ADAAAA, elicited the immune response in A. thaliana, but the response was in a lower level caused by (GlcNAc)6. In the case of another hetero-COS with a DP of 6, ADAADA, the immune response was eliminated almost completely [38]. Although the binding mode of COS elicitor to chitin receptor complex is still controversial [38], the N-acetylated residues were confirmed to be essential for the immune response in plants. The biological activities of COS s as plant elicitors are summarized in Table 5.
3. Synthetic strategies of COS with promising functions
Although the structure-function relationship of COS is complex, the available data on COS of various bioactivities provides information useful for the strategic design of tailor-made COS with improved biological activities. To obtain COS with desired sequences different green-chemical strategies have been reported, as described below.
3.1. Use of transglycosylation (TG) activity in chitinolytic enzymes
Many glycoside hydrolases with anomeric-retaining mechanism have been recognized to catalyze transglycosylation (TG) reactions to some extent [39]. Hen egg white lysozyme is one of the enzymes that catalyze TG in addition to hydrolysis [40]. TG in the reaction catalyzed by lysozyme was estimated to be efficient from the HPLC-based reaction time-course of COS degradation [41,42,43,44] and was utilized for synthesis of COS with longer chains and higher degree of crystallinity [45]. Lysozyme TG was also applied to the synthesis of a novel inhibitor of hen egg-white lysozyme [46], and a new substrate for assaying lysozyme activity [47]. Like lysozyme, chitinases belonging to the GH18 family were reported to catalyze TG reactions with COS [48,49]. In these enzymatic TG reactions, native enzymes with full intrinsic hydrolytic activities were used. Therefore, COS with longer chains produced by TG activity were again hydrolyzed to COS with shorter chains. It was highly desirable to suppress the hydrolytic activity of these enzymes, although this case complete loss of the glycosidic bond cleavage should be avoided, because the cleavage of b-1,4-glycosidic bond activates the anomeric center to the transition state, which is essential for receiving the attack of acceptor molecule [42,44]. It is necessary to employ some technique that suppresses hydration of the anomeric center in the transition state, while maintaining glycosidic bond cleavage activity. Fukamizo et al. reported enhanced TG activity in a chemically modified lysozyme, in which bulky residues, such as glucosamine and p-nitrophenyl-sulfenyl moieties, were introduced into the subsites -4, -3, and -2 of the lysozyme binding cleft [50,51]. They explained that the enhancement of TG activity was derived from the lower binding ability of the glycone - binding site (negatively numbered subsites), which, in turn, enhanced the acceptor binding to the aglycone - binding site (positively numbered subsites). However, the chemical methods seemed to be applicable only to inexpensive enzymes that are easy to isolate, such as lysozyme.
A GH2 exo-b-glucosaminidase from Amycolatopsis orientalis was reported to split off the non-reducing end GlcN unit from the (GlcN)n (n = 2, 3, 4, , , , ) substrates. This enzyme was found to catalyze a TG reaction in addition to hydrolysis; that is, from the (GlcN)4 substrate, (GlcN)5 and (GlcN)6 were formed as TG products [52]. After hydrolysis, the transition state of the GlcN monomer was transferred to the remaining (GlcN)4, which in turn acted as an acceptor, producing (GlcN)5. Similarly, the TG product (GlcN)5 subsequently acted as an acceptor, producing (GlcN)6. A GH5 chitosanase from Streptomyces griseus HUT 6037 was also reported to catalyze a TG reaction. When incubated with this enzyme, (GlcN)5 was hydrolyzed to (GlcN)2 + (GlcN)3 or (GlcNAc)3 + (GlcN)2. When the same reaction was conducted in the presence of excess amount of (GlcNAc)3, TG products, including (GlcN)2-(GlcNAc)3 and (GlcN)3-(GlcNAc)3, were formed [53].
3.2. Mutation strategies for enhancing TG activity
Various mutations have been attempted to enhance the TG activities of GH18 chitinases. Aronson et al. [54] reported for the first time that the mutation of a tryptophan residue in the substrate-binding cleft of Serratia marcescens chitinase A strongly enhanced the transglycosylation activity. The mutated tryptophan residue (Trp167) was located at subsite -3; therefore, the mechanism of TG enhancement was similar to that in the chemically modified lysozyme mentioned above [51]. Mutation of the middle Asp residue of the catalytic DxDxE motif of GH18 chitinases was also found to be useful for obtaining an efficiently transglycosylating chitinase [55,56]. Electrostatics in the catalytic cleft may be affected by the Asp mutation, probably resulting in the change in the state of the catalytic water molecule. Podile and his coworkers [57,58] thoroughly mutated the amino acids localized in the catalytic center and the groove of a bacterial GH18 chitinase, indicating that the triple mutations, which reduce the hydrolytic activity, binding affinity and stability of intermediate states, provided hypertransglycosylating mutants. Furthermore, they suggested the importance of optimal positioning of the catalytic water molecule and the acceptor molecule surrounding the catalytic center. In addition to these bacterial chitinases, plant chitinases were also employed in producing transglycosylating chitinases. Structure/function studies of GH18 chitinases from Nicotiana tobaccum (NtChiV), Arabidopsis thaliana (AtChiC) and Cycas revoluta (CrChiA) revealed that CrChiA possesses stronger TG activity as compared with NtChiV and AtChiC [59]. Introducing the Trp side chain into the upper portion of the catalytic center of AtChiC and CrChiA was found to strongly enhance TG activities of the enzymes [60,61]. The enhanced hydrophobicity of this region may change the state of the catalytic water, resulting in the suppression of attack by the water molecule and thus enhancing the TG reaction. Especially, the CrChiA mutant, in which the Trp side chain was introduced, exhibited hyper-transglycosylating activity [61].
3.3. Converting chitinolytic enzymes to glycosynthase
Glycosynthase was first developed from anomer-retaining glycoside hydrolases by mutation of the catalytic nucleophile [62,63]. In 2006, Honda and Kitaoka reported the first glycosynthase from an inverting glycoside hydrolase [64]. Since the retaining GH18 chitinases did not have a corresponding nucleophile in the catalytic center [65], inverting chitinases belonging to GH19 were employed for developing glycosynthase [66,67]. GH19 Bryum coronatum chitinase (BcChiA) mutants, in which the serine residue fixing a catalytic water molecule was mutated to alanine, cysteine or glycine, successfully catalyzed glycosyl transfer of the activated COS fluoride (COS-F, Figure 3A) to the acceptor COS. Using these BcChiA mutants, (GlcNAc)4 was synthesized from (GlcNAc)2-fluoride (donor) and (GlcNAc)2 (acceptor). The substrate-binding groove of BcChiA consists of four subsites, -2, -1, +1 and +2 [68], while that of a GH19 chitinase from rye seeds (RSC-c) consists of eight subsites, -4, -3, -2, -1, +1, +2, +3 and +4 [69]. It appeared that glycosynthase derived from RSC-c produces COS with longer chains. In fact, the double mutants, in which Glu89, acting as a catalytic base and Ser120, fixing a catalytic water molecule, were mutated, produced (GlcNAc)7 from (GlcNAc)3-fluoride (donor) and (GlcNAc)4 (acceptor). Glycosynthase derived from hen egg-white lysozyme (HEL-D52S) was produced and found to catalyze the glycosyl transfer. Using GlcN-(GlcNAc)2-fluoride obtained from (GlcNAc)3-fluoride by the action of a site-specific chitooligosaccharide N-deacetylase, the HEL-D52S glycosynthase allowed the size-controlled synthesis of GlcN-(GlcNAc)5, GlcN-(GlcNAc)6 and GlcN-(GlcNAc)7 from the acceptors (GlcNAc)3, (GlcNAc)4 and (GlcNAc)5, respectively. The use of the site-specific deacetylase avoided a condensation reaction of the donor substrate itself [70].
3.4. Use of activated sugars as donor substrates
It is well known that p-nitrophenyl glycosides are efficient donor substrates in glycoside synthesis [71]. Harmsen et al. [72] reported the efficient synthesis of COS with alternating GlcNAc and GlcN using p-nitrophenyl GlcN-GlcNAc dimer (GlcN-GlcNAc-pNP, Figure 3B), which was obtained by enzymatic deacetylation of GlcNAc-GlcNAc-pNP, and Serratia GH18 chitinase mutants as the catalysts. The mutants efficiently produced the alternating COS composed of GlcN and GlcNAc, i.e., (DA)2, (DA)3, (DA)4 and (DA)5, which are possible candidates for inhibitors of chitin-related enzymes and proteins, such as human chitotriosidase, a therapeutic target for systemic sclerosis [72]. (DA)4 was also obtained from oxazoline derivatives of DA and was confirmed to be an inhibitor of chitin elicitor binding protein in plants [37].
Production of reducing-end activated COS, such as glycosyl fluoride and p-nitrophenyl glycoside, used to require protection/deprotection procedures unfamiliar to enzymologists. Nowadays however, protection-free methods for obtaining activated sugars have been developed and used for reducing end-activation of COS. Tanaka et al. reported the protection-free synthesis of reducing-end-activated sugar derivatives, in which the 4.6-dimethoxy-1,3,5-triazin-2-yl (DMT) group was introduced at the anomeric center of the reducing end [73]. GlcN-DMT (Figure 3C) was synthesized and used as the glycosyl donor for synthesizing (GlcN)n with longer chains using GH2 exo-b-glucosaminidase as a template [74]. Furthermore, (GlcNAc)2-DMT was synthesized and used as a glycosyl donor for synthesizing (GlcNAc)4. A glycosynthase mutant derived from GH19 BcChiA [66] successfully synthesized (GlcNAc)4 from (GlcNAc)2-DMT and (GlcNAc)2 [75].
Sugar oxazolines have been frequently used as the glycosyl donors and are synthesized by the protection-free method using the water-soluble dehydrating agents, carbodiimides. Noguchi et al. reported that 2-chloro-1,3-dimethylimidazolinium chloride (DMC) was the most suitable for oxazoline synthesis from COS. The yields were 70-80% [76]. The oxazoline derivatives from (GlcNAc)n (Figure 3D; (GlcNAc)n-oxa, n = 2, 3, 4 and 5) were purified by HPLC, and used as the donor substrates for synthesizing (GlcNAc)n with longer chains using an activity-reduced mutant from bacterial GH18 chitinase as a template [77]. Various mutations were introduced into bacterial GH18 chitinases, SpChiD, and (GlcNAc)10 was produced from (GlcNAc)5-oxa in a good yield [78]. Hypertransglycosylating mutants of GH18 chitinases from plant origins, Arabidopsis thaliana and Cycas revoluta, were also used for (GlcNAc)n synthesis from the sugar oxazolines [79]. The product distribution was controlled by using different combinations of the substrate size of donor/acceptor.
3.5. Use of site-specific chitin deacetylases
Hembach et al. [80] reported the enzymatic production of a full set of partially N-acetylated chitosan tetramers of all possible sequences consisting of GlcN and GlcNAc, through de-N-acetylation of (GlcNAc)4 using various site-specific chitin deacetylases and reverse N-acetylation of (GlcN)4 in the presence of acetate (2 M) by the same deacetylases. The structures of the products were successfully identified using ultra high-performance liquid chromatography-electrospray ionization-mass spectrometry (UHPLC/ESI/MS) [81,82], which enabled the full separation of the enzymatic products. In UHPLC/ESI/MS, a hydrophilic interaction chromatography (HILIC) column (Acquity UHPLC BEH Amide) was used to separate COS tetramers with different degrees of acetylation and sequences, and gradient elution was performed with 80 - 20% acetonitrile in water containing 10 mM NH4HCO3 and 0.1% (v/v) formic acid, at a flow rate of 0.8 mL.min−1 and 75 ºC using an appropriate gradient program controlled by an UHPLC system purchased from Dionex Co. The effluents were detected with an ESI-MS detector in positive mode. The sequencing methods of the COS tetramers consists of the following procedures: (1) acetylation of free amino groups in COS using a deuterated reagent, (2) labeling the reducing end with H218O; (3) quantifying COS using [13C2, 2H3]-labeled internal standards; (4) sequencing by MS/MS [81]. The enzymatic de-N-acetylation/N-acetylation of the COS, combined with the state-of-the-art separation/analysis methods, enabled the production of COS with desired sequences. This may subsequently lead to defining the structure-function relationships of COS. This strategy of using site-specific deacetylases was also utilized for producing chitosan oligosaccharides with random and non-random acetylation patterns [83].
3.6. Metabolic engineering approaches
Studies of the biological activities of COS usually need large amounts of COS. However, the enzymatic methods described thus far provided only limited amounts of COS. As an alternative to the current enzymatic methods, microbial fermentation using recombinant Escherichia coli has now attracted researchers’ attention, because the method is capable of producing larger amounts of COS and is environmentally friendly [84]. Recently, a non-pathogenic bacterium, Bacillus subtilis, also provided an additional opportunity for fermentative production of COS [85]. B. subtilis was engineered by introducing a combinatorial pathway for the production of well-defined COS. Specifically, an exogenous COS synthase was overexpressed in B. subtilis, then the GlcNAc synthesis module was also introduced to enhance the intracellular GlcNAc supply. Furthermore, both the de novo pathway and the salvage pathway of UDP-GlcNAc were engineered to further promote the biosynthesis of (GlcNAc)n. The metabolic engineering finally attained the production of (GlcNAc)n of 4.8 g·L−1 including (GlcNAc)5 (86%), (GlcNAc)4 (7%), (GlcNAc)3 (5%), and (GlcNAc)2 (2%). Further engineering may provide a cell factory for producing required amounts of COS with desired DP, DA and sequences [86].
4. Conclusions and future prospects
Recently, the relationship between the structures and the biological activities of COS has become more explicit since the biological activities of COS were satisfactorily characterized with respect to DPs, DAs and sequences. Based on the data listed in Table 1, Table 2, Table 3, Table 4 and Table 5, it is possible to design COS sequences with desired biological functions. A combination of the strategies summarized in Table 6 enabled the synthesis of tailor-made COS with specified sequences.
Recently, we successfully produced a large amount of high-quality (GlcNAc)2 from chitin food wastes using an in-house Vibrio chitinase [87], while (GlcN)2 was most efficiently produced by endo-splitting GH46 chitosanases from the chitosan wastes [88,89]. Starting from these chitobioses, various green-chemical strategies are proposed, as shown in Figure 4. COS of differing chain length can be elongated from a DP of 2 to a DP of 4, 6, 8, or 10 by means of transglycosylation reactions, using chitinase/chitosanase mutants as effective catalysts. Glycosynthase mutants were also useful for chain-length elongation using reducing-end activated COS as the donor substrates. Subsequent site-specific de-N-acetylation/N-acetylation may possibly produce tailor-made COS that possess most plausible sequences with desired biological activities. The concept of tailor-made COS will further enhance the exploitation of chitin biomass.
Author Contributions
Conceptualization, R.T. and W.S.; Data curation, R.T. and T.F.; original draft preparation, R.T. and T.F.; review and editing, T.F. and W.S.; project administration, W.S.; funding acquisition, W.S. All authors have read and agreed to the published version of the manuscript.
Funding
R.T. received a full-time PhD scholarship from Vidyasirimedhi Institute of Science and Technology (VISTEC). W.S. was funded by Thailand Science Research and Innovation (TSRI) and Vidyasirimedhi Institute of Science and Technology (Grant nos. B22WIS-TSR010, and N42A660311).
Acknowledgments
We would like to thank David Apps for valuable and critical proofreading of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
COS, chitooligosaccharide; DP, degree of polymerization; DA, degree of N-acetylation; MW, molecular weight; GlcNAc (A), 2-acetamido-2-deoxy-D-glucopyranose; GlcN (D), 2-amino-2-deoxy-D-glucopyranose; (GlcNAc)n and (GlcN)n, homo-oligosaccharides composed of GlcNAc and GlcN, respectively, with a DP of n; AtChiC, a GH18 chitinase from Arabidopsis thaliana; NtChiV, a GH18 chitinase from Nicotiana tabacum; CrChiA, a GH18 chitinase from Cycas revoluta; BcChiA, a GH19 chitinase from Bryum coronatum; RSC-c, a GH19 chitinase from Secale cereale; TG, transglycosylation.
References
- Tharanathan, R.N.; Kittur, F.S. Chitin-the undisputed biomolecule of great potential. Crit. Rev. Food Sci. Nutr. 2003, 43, 61–87. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.J.; Park, R.D. Bioproduction of chitooligosaccharides: present and perspectives. Mar. Drugs. 2014, 2014 12, 5328–5356. [Google Scholar] [CrossRef]
- Liaqat, F.; Eltem, R. Chitooligosaccharides and their biological activities: A comprehensive review. Carbohydr. Polym. 2018, 184, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Kasaai, M.R. The use of various types of NMR and IR spectroscopy for structural characterization of chitin and chitosan. in “Chitin, Chitosan, oligosaccharides, and their derivatives: Biological activities and application”, ed. by Kim, Se-K., CRC Press (Taylor & Francis Group), 2011, pp. 149-170.
- Peter, M.G.; Eberlin, M.N. Application of Mass spectrometry to analyze the structure and Bioactivity of chitooligosaccharides. in “Chitin, Chitosan, oligosaccharides, and their derivatives: Biological activities and application”, ed. by Kim, Se-K., CRC Press (Talor & Francis Group), 2011, pp. 127-148.
- Mengíbar, M.; Mateos-Aparicio, I.; Miralles, B.; Heras, A. Influence of the physico-chemical characteristics of chitooligosaccharides (COS) on antioxidant activity. Carbohydr. Polym. 2013, 97, 776–782. [Google Scholar] [CrossRef]
- Hao, W.; Li, K.; Ge, X.; Yang, H.; Xu, C.; Liu, S. Yu, H.; Li, P.; Xing, R. The effect of N-acetylation on the anti-inflammatory activity of chitooligosaccharides and its potential for relieving endotoxemia. Int. J. Mol. Sci. 2022, 23, 8205. [Google Scholar] [CrossRef]
- Cord-Landwehr, S.; Moerschbacher, B.M. Deciphering the ChitoCode: fungal chitins and chitosans as functional biopolymers. Fungal Biol. Biotechnol. 2021, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.J.; Park, P.J.; Kim, S.K. Antimicrobial effect of chitooligosaccharides produced by bioreactor. Carbohydr. Polym. 2001, 44, 71–76. [Google Scholar] [CrossRef]
- Li, K.; Xing, R.; Liu, S.; Qin, Y.; Yu, H.; Li, P. Size and pH effects of chitooligomers on antibacterial activity against Staphylococcus aureus. Int. J. Biol. Macromol. 2014, 64, 302–305. [Google Scholar] [CrossRef]
- No, H.K.; Park, N.Y.; Lee, S.H.; Meyers, S.P. Antibacterial activity of chitosans and chitosan oligomers with different molecular weights. Int. J. Food Microbiol. 2002, 74, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Aktuganov, GE.; Safina, V.R.; Galimzianova, N.F.; Elena A Gilvanova, Lyudmila Yu Kuzmina, Alexander I Melentiev, Andrei H Baymiev, Sergey A Lopatin.. Constitutive chitosanase from and its potential for preparation of antimicrobial chitooligomers World J Microbiol Biotechnol 2022 Jul 22;38(10):167.
- Ganan, M.; Silje B Lorentzen, S.B.; Jane W Agger, J.W.; Heyward, C.A.; Bakke, O.; Knutsen, S.H.; Aam, B.B.; Eijsink, V.G.H.; Gaustad, P.; Sørlie, M. Antifungal activity of well-defined chitooligosaccharide preparations against medically relevant yeasts. PLoS One 2019, 14, e0210208. [Google Scholar] [CrossRef]
- Kulikov, S.N.; Lisovskaya, S.A.; Zelenikhin, P.V.; Bezrodnykh, E.A.; Shakirova, D.R.; Blagodatskikh, I.V.; Tikhonov, V.E. Antifungal activity of oligochitosans (short-chain chitosans) against some Candida species and clinical isolates of Candida albicans: Molecular weight-activity relationship. Eur. J. Med. Chem. 2014, 74, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Li, K.; Ge, X.; Yang, H.; Xu, C.; Liu, S.; Yu, H.; Li, P.; Xing, R. The Effect of N-acetylation on the anti-inflammatory activity of chitooligosaccharides and its potential for relieving endotoxemia. Int. J. Mol. Sci. 2022, 23, 8205. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.J.; Park, J.K.; Il Park, Y. Anti-inflammatory effects of low-molecular weight chitosan oligosaccharides in IgE–antigen complex-stimulated RBL-2H3 cells and asthma model mice. Int. Immunopharmacol. 2012, 12, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Pangestuti, R.; Bak, S.-S.; Kim, S.-K. Attenuation of pro-inflammatory mediators in LPS-stimulated BV2 microglia by chitooligosaccharides via the MAPK signaling pathway. Int. J. Biol. Macromol. 2011, 49, 599–606. [Google Scholar] [CrossRef]
- Wei, P.; Ma, P.; Xu, Q.-S.; Bai, Q.-H.; Gu, J.-G.; Xi, H.; Du, Y.-G.; Yu, C. Chitosan oligosaccharides suppress production of nitric oxide in lipopolysaccharide-induced N9 murine microglial cells in Vitro. Glycoconj.. J. 2012, 29, 285–295. [Google Scholar] [CrossRef]
- Sánchez, Á.; Mengíbar, M.; Fernández, M.; Alemany, S.; Heras, A.; Acosta, N. Influence of preparation methods of chitooligosaccharides on their physicochemical properties and their anti-inflammatory effects in mice and in RAW264.7 Macrophages. Mar. Drugs 2018, 16, 430. [Google Scholar] [CrossRef]
- Ngo, D.-N.; Kim, M.-M.; Kim, S.-K. Chitin oligosaccharides inhibit oxidative stress in live cells. Carbohydr. Polym. 2008, 74, 228–234. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, X.; Ji, L.; Du, X.; Sang, Q.; Chen, F. Enzymatic single-step preparation and antioxidant activity of hetero-chitooligosaccharides using non-pretreated housefly larvae powder. Carbohydr. Polym. 2017, 172, 113–119. [Google Scholar] [CrossRef]
- Liu, H.T.; Li, W.M.; Xu, G.; Li, X.Y.; Bai, X.F.; Wei, P.; Yu, C.; Du, Y.G. Chitosan oligosaccharides attenuate hydrogen peroxide-induced stress injury in human umbilical vein endothelial cells. Pharmacol. Res. 2009, 59, 167–175. [Google Scholar] [CrossRef]
- Liu, H.T.; He, J.L.; Li, W.M.; Yang, Z.; Wang, Y.X.; Bai, X.F.; Yu, C.; Du, Y.G. Chitosan oligosaccharides protect human umbilical vein endothelial cells from hydrogen peroxide-induced apoptosis. Carbohydr. Polym. 2010, 80, 1062–1071. [Google Scholar] [CrossRef]
- Jia, Y.; Ma, Y.; Zou, P.; Cheng, G.; Zhou, J.; Cai, S. Effects of different oligochitosans on isoflavone metabolites, antioxidant activity, and isoflavone biosynthetic genes in soybean (Glycine max) seeds during germination. J. Agric. Food Chem. 2019, 67, 4652–4661. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Li, K.; Ma, Y.; Li, R.; Xing, R.; Yu, H.; Li., P. Preparation and antioxidant activity of chitosan dimers with different sequences. Mar. Drugs 2021, 19, 366. [Google Scholar] [CrossRef] [PubMed]
- Tsukada, K.; Matsumoto, T.; Aizawa, K.; Tokoro, A.; Naruse, R.; Suzuki, S.; Suzuki, M. Antimetastatic and growth-inhibitory effects of N-acetylchitohexaose in mice bearing Lewis lung Carcinoma. Jap. J. Cancer Res. 1990, 81, 259–265. [Google Scholar] [CrossRef]
- Salah, R.; Michaud, P.; Mati, F.; Harrat, Z.; Lounici, H.; Abdi, N.; Drouiche, N.; Mameri, N. Anticancer activity of chemically prepared shrimp low molecular weight chitin evaluation with the human monocyte leukaemia cell line, THP-1. Int. J. Biol. Macromol. 2013, 52, 333–339. [Google Scholar] [CrossRef]
- Park, J.K.; Chung, M.J.; Choi, H.N.; Park, Y.I. Effects of the molecular weight and the degree of deacetylation of chitosan oligosaccharides on antitumor activity. Int. J. Mol. Sci. 2011, 12, 266–277. [Google Scholar] [CrossRef]
- Li, X.; Wang, J.; Chen, X.; Tian, J.; Li, L.; Zhao, M.; Jiao, Y.; Zhou, C. Effect of chitooligosaccharides on cyclin D1, bcl-xl and bcl-2 mRNA expression in A549 cells using quantitative PCR. Chinese Sci. Bull. 2011, 56, 1629–1632. [Google Scholar] [CrossRef]
- Zhai, X.; Li, C.; Ren, D.; Wang, J.; Ma, C.; Abd El-Aty, A.M. The impact of chitooligosaccharides and their derivatives on the in vitro and in vivo antitumor activity: A comprehensive review. Carbohydr. Polym. 2021, 266, 118132. [Google Scholar] [CrossRef]
- Vander, P.; Km, V.R.; Domard, A.; Eddine, E.G.N.; Moerschbacher, B.M. Comparison of the ability of partially N-acetylated chitosans and chitooligosaccharides to elicit resistance reactions in wheat leaves. Plant Physiol. 1998, 118, 1353–1359. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, B.; Sarma, P.V.S.R.N.; Ankati, S.; Bhuvanachandra, B.; Podile, A.R. Elicitation of defense response by transglycosylated chitooligosaccharides in rice seedlings. Carbohydr. Res. 2021 510, 108459. [CrossRef]
- Miya, A.; Albert, P.; Shinya, T.; Desaki, Y.; Ichimura, K.; Shirasu, K.; Narusaka, Y.; Kawakami, N.; Kaku, H.; Shibuya, N. CERK1, a LysM receptor kinase, is essential for chitin elicitor signaling in Arabidopsis. Proc. Natl. Acad. Sci. U. S. A. 2007, 104, 19613–19618. [Google Scholar] [CrossRef]
- Liu, T.; Liu, Z.; Song, C.; Hu, Y.; Han, Z.; She, J.; Fan, F.; Wang, J.; Jin, C.; Chang, J.; Zhou, J.M.; Chai, J. Chitin-induced dimerization activates a plant immune receptor. Science. 2012, 336, 1160–1164. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Liang, Y.; Tanaka, K.; Nguyen, C.T.; Jedrzejczak, R.P.; Joachimiak, A.; Stacey, G. The kinase LYK5 is a major chitin receptor in Arabidopsis and forms a chitin-induced complex with related kinase CERK1. eLife. 2014, 3, e03766. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, J.; Han, Z.; Gong, X.; Zhang, H.; Chai, J. Molecular mechanism for fungal cell wall recognition by rice chitin receptor OsCEBiP. Structure. 2016, 24, 1192–1200. [Google Scholar] [CrossRef] [PubMed]
- Hayafune, M.; Berisio, R.; Marchetti, R.; Silipo, A.; Kayama, M.; Desaki, Y.; Arima, S.; Squeglia, F.; Ruggiero, A.; Tokuyasu, K.; Molinaro, A.; Kaku, H.; Shibuya, N. Chitin-induced activation of immune signaling by the rice receptor CEBiP relies on a unique sandwich-type dimerization. Proc. Natl. Acad. Sci. U.S.A. 2014, 111, E404–E413. [Google Scholar] [CrossRef]
- Gubaeva, E.; Gubaev, A.; Melcher, R.L.J.; Cord-Landwehr, S.; Singh, R.; El Gueddari, N.E.; Moerschbacher, B.M. ‘Slipped Sandwich’ model for chitin and chitosan perception in Arabidopsis. Mol. Plant Microbe Interact. 2018, 31, 1145–1153. [Google Scholar] [CrossRef]
- Mészáros, Z.; Nekvasilová, P.; Bojarová, P.; Křen, V.; Slámová, K. Advanced glycosidases as ingenious biosynthetic instruments. Biotechnol Adv. 2021, 49, 107733. [Google Scholar] [CrossRef]
- Chipman, D.M.; Pollock, J.J.; Sharon, N. Lysozyme-catalyzed hydrolysis and transglycosylation reactions of bacterial cell wall oligosaccharides. J. Biol. Chem. 1968, 243, 487–496. [Google Scholar] [CrossRef]
- Masaki, A.; Fukamizo, T.; Otakara, A.; Torikata, T.; Hayashi, K.; Imoto, T. Lysozyme-catalyzed reaction of chitooligosaccharides. J. Biochem. 1981, 90, 527–533. [Google Scholar] [CrossRef]
- Masaki, A.; Fukamizo, T.; Otakara, A.; Torikata, T.; Hayashi, K.; Imoto, T. Estimation of rate constants in lysozyme-catalyzed reaction of chitooligosaccharides. J. Biochem. 1981, 90, 1167–1175. [Google Scholar] [CrossRef]
- Fukamizo, T.; Torikata, T.; Nagayama, T.; Minematsu, T.; Hayashi, K. Enzymatic activity of avian egg-white lysozymes. J. Biochem. 1983, 94, 115–122. [Google Scholar] [CrossRef]
- Fukamizo, T.; Minematsu, T.; Yanase, Y.; Hayashi, K.; Goto., S. Substrate size dependence of lysozyme-catalyzed reaction. Arch. Biochem. Biophys. 1986, 250, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Hattori, T.; Sakabe, Y.; Ogata, M.; Michishita, K.; Dohra, H.; Kawagishi, H.; Totani, K.; Nikaido, M.; Nakamura, T.; Koshino, H.; Usui, T. Enzymatic synthesis of an α-chitin-like substance via lysozyme-mediated transglycosylation. Carbohydr. Res. 2012, 347, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Ogata, M.; Umemoto, N.; Ohnuma, T.; Numata, T.; Suzuki, A.; Usui, T.; Fukamizo, T. A novel transition-state analogue for lysozyme, 4-O-β-tri-N-acetylchitotriosyl moranoline, provided evidence supporting the covalent glycosyl-enzyme intermediate. J. Biol. Chem. 2013, 288, 6072–6082. [Google Scholar] [CrossRef] [PubMed]
- Ogata, M.; Matsui, M.; Kono, H.; Matsuzaki, Y.; Kato, Y.; Usui, T. A novel analytical procedure for assaying lysozyme activity using an end-blocked chitotetraose derivative as substrate. Anal. Biochem. 2017, 538, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Madhuprakash, J.; Singh, A.; Kumar, S.; Sinha, M.; Kaur, P.; Sharma, S.; Podile, A.R.; Singh, T.P. Structure of chitinase D from Serratia proteamaculans reveals the structural basis of its dual action of hydrolysis and transglycosylation. Int. J. Biochem. Mol. Biol. 2013, 4, 166–178. [Google Scholar]
- Bhuvanachandra, B.; Podile, A.R. A transglycosylating chitinase from Chitiniphilus shinanonensis (CsChiL) hydrolyzes chitin in a processive manner. Int. J. Biol. Macromol. 2020, 145, 1–10. [Google Scholar] [CrossRef]
- Fukamizo, T.; Goto, S.; Torikata, T.; Araki, T. State of binding subsites in Asp 101-modified lysozymes. J. Biochem. 1990, 107, 445–451. [Google Scholar] [CrossRef]
- Fukamizo, T.; Goto, S.; Torikata, T.; Araki, T. Enhancement of transglycosylation of lysozyme by chemical modification. Agric. Biol. Chem. 1989, 53, 2641–2651. [Google Scholar] [CrossRef]
- Fukamizo, T.; Fleury, A.; Côté, N.; Mitsutomi, M.; Brzezinski, R. Exo-b-D-glucosaminidase from Amycolatopsis orientalis: catalytic residues, sugar recognition specificity, kinetics, and synergism. Glycobiol. 2006, 16, 1064–1072. [Google Scholar] [CrossRef]
- Tanabe, T.; Morinaga, K.; Fukamizo, T.; Mitsutomi, M. Novel chitosanase from Streptomyces griseus HUT 6037 with transglycosylation activity. Biosci. Biotechnol. Biochem. 2003, 67, 354–364. [Google Scholar] [CrossRef]
- Aronson, N.N., Jr.; Halloran, B.A.; Alexeyev, M.F.; Zhou, X.E.; Wang, Y.; Meehan, E.J.; Chen, L. Mutation of a conserved tryptophan in the chitin-binding cleft of Serratia marcescens chitinase A enhances transglycosylation. Biosci. Biotechnol. Biochem. 2006, 70, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Zakariassen, H.; Hansen, M.C.; Jøranli, M.; Eijsink, V.G.H.; Sørlie, M. Mutational effects on transglycosylating activity of family 18 chitinases and construction of a hypertransglycosylating mutant. Biochemistry 2011, 50, 5693–5703. [Google Scholar] [CrossRef]
- Sirimontree, P.; Suginta, W.; Sritho, N.; Kanda, Y.; Shinya, S.; Ohnuma, T.; Fukamizo, T. Mutation strategies for obtaining chitooligosaccharides with longer chains by transglycosylation reaction of family GH18 chitinase. Biosci. Biotechnol. Biochem. 2014, 78, 2014–2021. [Google Scholar] [CrossRef] [PubMed]
- Madhuprakash, J.; Dalhus, B.; Rani, T.S.; Podile, A.R.; Eijsink, V.G.H.; Sørlie, M. Key residues affecting transglycosylation activity in family 18 chitinases: Insights into donor and acceptor subsites. Biochemistry. 2018, 57, 4325–4337. [Google Scholar] [CrossRef] [PubMed]
- Madhuprakash, J.; Tanneeru, K.; Purushotham, P.; Guruprasad, L.; Podile, A.R. Transglycosylation by chitinase D from Serratia proteamaculans improved through altered substrate interactions. J. Biol. Chem. 2012, 287, 44619–44627. [Google Scholar] [CrossRef]
- Umemoto, N.; Kanda, Y.; Ohnuma, T.; Osawa, T.; Numata, T.; Sakuda, S.; Taira, T.; Fukamizo, T. Crystal structures and inhibitor binding properties of plant class V chitinases: the cycad enzyme exhibits unique structural and functional features. Plant J. 2015, 82, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Umemoto, N.; Ohnuma, T.; Mizuhara, M.; Sato, H.; Skriver, K.; Fukamizo, T. Introduction of a tryptophan side chain into subsite +1 enhances transglycosylation activity of a GH-18 chitinase from Arabidopsis thaliana, AtChiC. Glycobiology. 2013, 23, 81–90. [Google Scholar] [CrossRef]
- Umemoto, N.; Ohnuma, T.; Osawa, T.; Numata, T.; Fukamizo, T. Modulation of the transglycosylation activity of plant family GH18 chitinase by removing or introducing a tryptophan side chain. FEBS Lett. 2015, 589, 2327–2333. [Google Scholar] [CrossRef]
- Faijes, M.; Planas, A. In vitro synthesis of artificial polysaccharides by glycosidases and glycosynthases. Carbohydr. Res. 2007, 342, 1581–1594. [Google Scholar] [CrossRef]
- Danby, P.M.; Withers, S.G. Advances in enzymatic glycoside synthesis. ACS Chem. Biol. 2016, 11, 1784–1794. [Google Scholar] [CrossRef]
- Honda, Y.; Kitaoka, M. The first glycosynthase derived from an inverting glycoside hydrolase. J. Biol. Chem. 2006, 281, 1426–1431. [Google Scholar] [CrossRef]
- van Aalten, D.M.; Komander, D.; Synstad, B.; Gåseidnes, S.; Peter, M.G.; Eijsink, V.G. Structural insights into the catalytic mechanism of a family 18 exo-chitinase. Proc. Natl. Acad. Sci. U.S.A. 2001, 98, 8979–8984. [Google Scholar] [CrossRef] [PubMed]
- Ohnuma, T.; Fukuda, T.; Dozen, S.; Honda, Y.; Kitaoka, M.; Fukamizo, T. A glycosynthase derived from an inverting GH19 chitinase from the moss Bryum coronatum. Biochem. J. 2012, 444, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Ohnuma, T.; Dozen, S.; Honda, Y.; Kitaoka, M.; Fukamizo, T. A glycosynthase derived from an inverting chitinase with an extended binding cleft. J. Biochem. 2016, 160, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Ohnuma, T.; Umemoto, N.; Nagata, T.; Shinya, S.; Numata, T.; Taira, T.; Fukamizo, T. Crystal structure of a “loopless” GH19 chitinase in complex with chitin tetrasaccharide spanning the catalytic center. Biochim. Biophys. Acta. 2014, 1844, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Ohnuma, T.; Umemoto, N.; Kondo, K.; Numata, T.; Fukamizo, T. Complete subsite mapping of a “loopful” GH19 chitinase from rye seeds based on its crystal structure. FEBS Lett. 2013, 587, 2691–2697. [Google Scholar] [CrossRef]
- Rousseau, A.; Armand, S.; Cottaz, S.; Fort, S. Size-controlled synthesis of β(1→4)-GlcNAc oligosaccharides using an endo-glycosynthase. Chemistry. 2021, 27, 17637–17646. [Google Scholar] [CrossRef] [PubMed]
- Rastelli, R.A.; Bucke, C. Enzymatic synthesis of oligosaccharides, Biotechnol. Genet. Eng. Rev., 1992, 10, 253–282. [Google Scholar] [CrossRef]
- Harmsen, R.A.G.; Aam, B.B.; Madhuprakash, J.; Hamre, A.G.; Goddard-Borger, E.D.; Withers, S.G.; Eijsink, V.G.H.; Sørlie, M. Chemoenzymatic synthesis of chito-oligosaccharides with alternating N-D-acetylglucosamine and D-glucosamine. Biochemistry, 2020, 59, 4581–4590. [Google Scholar] [CrossRef]
- Tanaka, T.; Noguchi, M.; Kobayashi, A.; Shoda, S. A novel glycosyl donor for chemo-enzymatic oligosaccharide synthesis: 4,6-dimethoxy-1,3,5-triazin-2-yl glycoside. Chem. Commun. (Camb). 2008, 17, 2016–2018. [Google Scholar] [CrossRef]
- Tanaka, T.; Wada, T.; Noguchi, M.; Ishihara, M.; Kobayashi, A.; Ohnuma, T.; Fukamizo, T.; Brzezinski, R.; Shoda, S.-I. 4,6-Dimethoxy-1,3,5-triazin-2-yl β-D-glycosaminides: novel substrates for transglycosylation reaction catalyzed by exo-β-D-glucosaminidase from Amycolatopsis orientalis. J. Carbohydr. Chem., 2012, 31, 634–646. [Google Scholar] [CrossRef]
- Ohnuma, T.; Tanaka, T.; Urasaki, A.; Dozen, S.; Fukamizo, T. A novel method for chemo-enzymatic synthesis of chitin oligosaccharide catalyzed by the mutant of inverting family GH19 chitinase using 4,6-dimethoxy-1,3,5-triazin-2-yl α-chitobioside as a glycosyl donor. J. Biochem. 2019, 165, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, M.; Tanaka, T.; Gyakushi, H.; Kobayashi, A.; Shoda, S. Efficient synthesis of sugar oxazolines from unprotected N-acetyl-2-amino sugars by using chloroformamidinium reagent in water. J. Org. Chem. 2009, 74, 2210–2212. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, N.; Tanaka, T.; Noguchi, M.; Kobayashi, A.; Ishikura, K.; Ikenuma, T.; Seno, H.; Watanabe, T.; Kohri, M.; Shoda, S.-I. One-pot chemoenzymatic route to chitoheptaose via specific transglycosylation of chitopentaose-oxazoline on chitinase-template. Chem. Lett. 2012, 41, 689–690. [Google Scholar] [CrossRef]
- Alsina, C.; Sancho-Vaello, E.; Aranda-Martínez, A.; Faijes, M.; Planas, A. Auxiliary active site mutations enhance the glycosynthase activity of a GH18 chitinase for polymerization of chitooligosaccharides. Carbohydr. Polym. 2021, 252, 117121. [Google Scholar] [CrossRef]
- Umemoto, N.; Saito, N.; Noguchi, M.; Shoda, S.-I.; Ohnuma, T.; Watanabe, T.; Sakuda, S.; Fukamizo, T. Plant chitinase mutants as the catalysts for chitooligosaccharide synthesis using the sugar oxazoline derivatives. J. Agric. Food Chem. 2022, 70, 12897–12906. [Google Scholar] [CrossRef]
- Hembach, L.; Cord-Landwehr, S.; Moerschbacher, B.M. Enzymatic production of all fourteen partially acetylated chitosan tetramers using different chitin deacetylases acting in forward or reverse mode. Sci. Rep. 2017, 7, 17692. [Google Scholar] [CrossRef]
- Cord-Landwehr, S.; Ihmor, P.; Niehues, A.; Luftmann, H.; Moerschbacher, B.M.; Mormann, M. Quantitative mass-spectrometric sequencing of chitosan oligomers revealing cleavage sites of chitosan hydrolases. Anal. Chem. 2017, 89, 2893–2900. [Google Scholar] [CrossRef]
- Hamer, S.N.; Cord-Landwehr, S.; Biarnés, X.; Planas, A.; Waegeman, H.; Moerschbacher, B.M.; Kolkenbrock, S. Enzymatic production of defined chitosan oligomers with a specific pattern of acetylation using a combination of chitin oligosaccharide deacetylases. Sci.Rep. 2015, 5, 8716. [Google Scholar] [CrossRef]
- Sreekumar, S.; Wattjes, J.; Niehues, A.; Mengoni, T.; Mendes, A.C.; Morris, E.R.; Goycoolea, F.M.; Moerschbacher, B.M. Biotechnologically produced chitosans with nonrandom acetylation patterns differ from conventional chitosans in properties and activities. Nat. Commun. 2022, 13, 7125. [Google Scholar] [CrossRef]
- Ling, M.; Li, J.; Du, G.; Liu, L. Metabolic engineering for the production of chitooligosaccharides: advances and perspectives. Emerg. Top Life Sci. 2018, 2, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Ling, M.; Wu, Y.; Tian, R.; Liu, Y.; Yu, W.; Tao, G.; Lv, X.; Li, J.; Du, G.; Amaro, R.L.; Liu, L. Combinatorial pathway engineering of Bacillus subtilis for production of structurally defined and homogeneous chitooligosaccharides. Metab. Eng. 2022, 70, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Weyer, R.; Hellmann, M.J.; Hamer-Timmermann, S.N.; Singh, R.; Moerschbacher, B.M. Customized chitooligosaccharide production-controlling their length via engineering of rhizobial chitin synthases and the choice of expression system. Front. Bioeng. Biotechnol. 2022, 10, 1073447. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.; Fukamizo, T.; Suginta, W. Bioeconomic production of high-quality chitobiose from chitin food wastes using an in-house chitinase from Vibrio campbellii. Biores. Bioprocess. 2022, 9, 86. [Google Scholar] [CrossRef]
- Chen, T.; Cheng, G.; Jiao, S.; Ren, L.; Zhao, C.; Wei, J.; Han, J.; Pei, M.; Du, Y.; Li, J.J. Expression and biochemical characterization of a novel marine chitosanase from Streptomyces niveus suitable for preparation of chitobiose. Mar. Drugs. 2021, 19, 300. [Google Scholar] [CrossRef]
- Li, Y.; Gou, Y.; Liu, Z.; Xie, T.; Wang, G. Structure-based rational design of chitosanase CsnMY002 for high yields of chitobiose. Colloids Surf. B Biointerfaces. 2021, 202, 111692. [Google Scholar] [CrossRef]
Figure 1.
The molecular structures of COS with different bioactivities. (A) chitooligosaccharides (GlcNAc)n, n = 1,2,3, …, (B) partially deacetylated chitooligosaccharides (GlcNAc.GlcN)n; n = 1,2,3,…, (C) chitosan oligosaccharides (GlcN)n, n = 1,2,3….
Figure 1.
The molecular structures of COS with different bioactivities. (A) chitooligosaccharides (GlcNAc)n, n = 1,2,3, …, (B) partially deacetylated chitooligosaccharides (GlcNAc.GlcN)n; n = 1,2,3,…, (C) chitosan oligosaccharides (GlcN)n, n = 1,2,3….
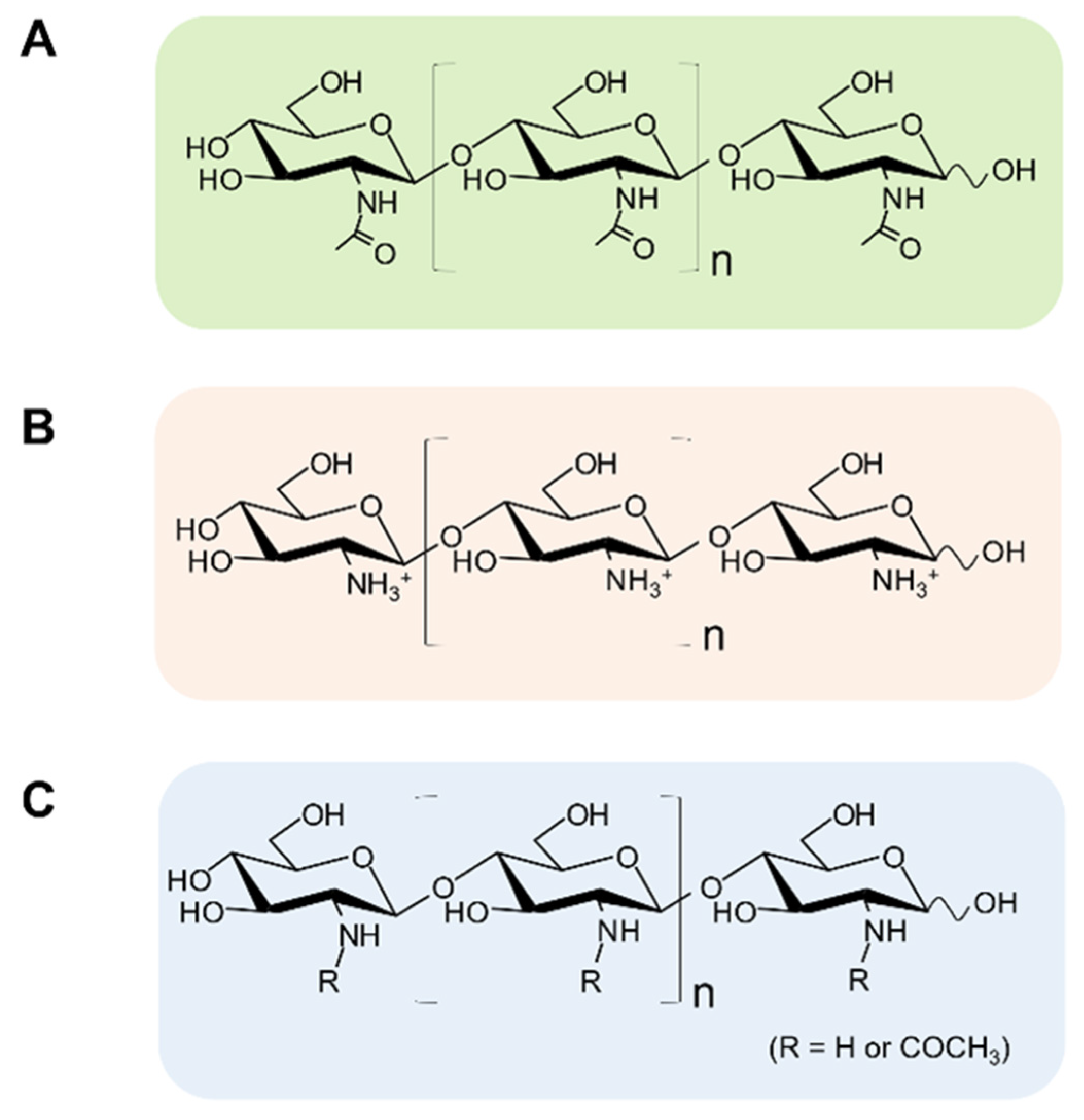
Figure 2.
A summary of biological activity of COS.
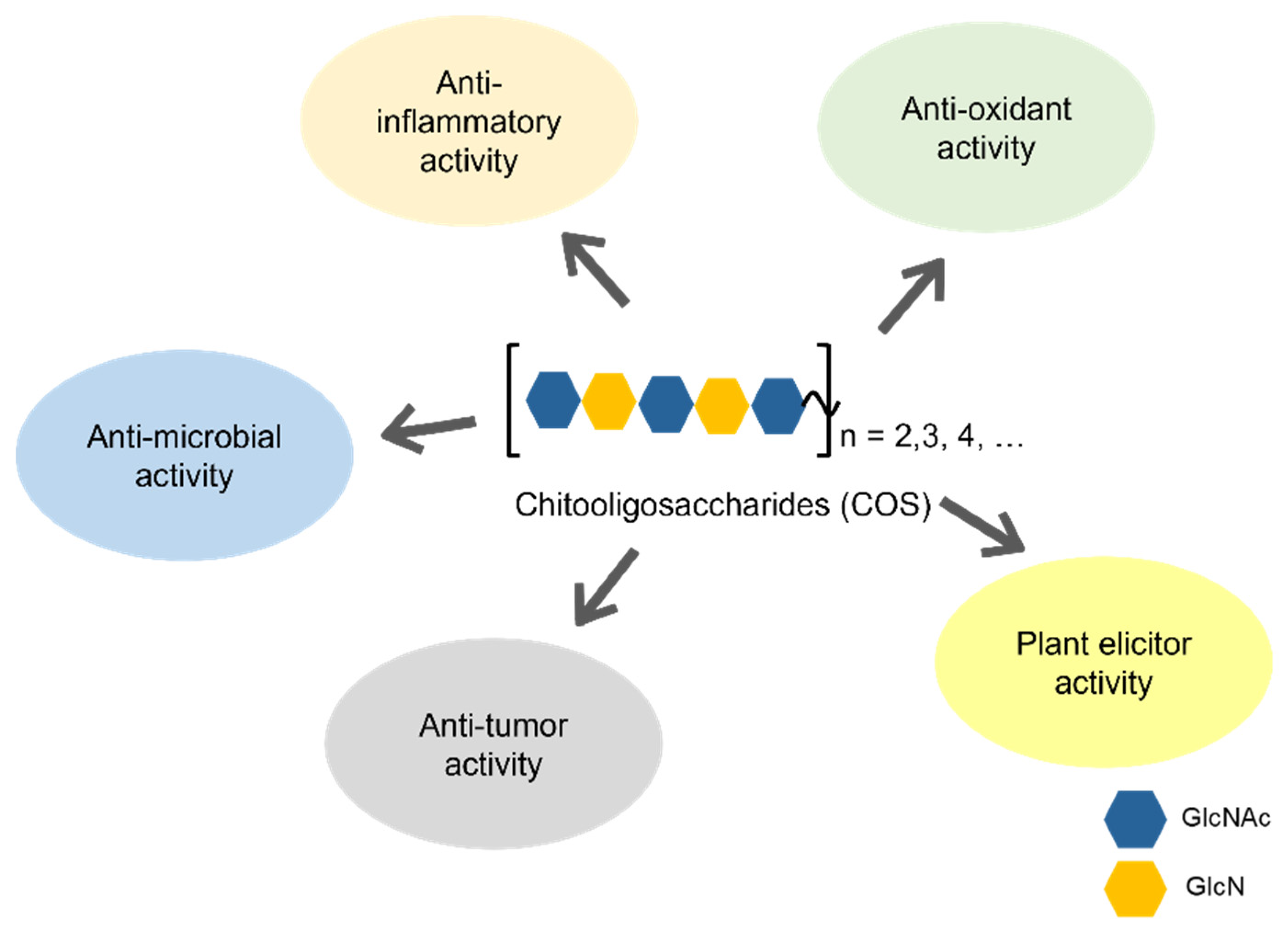
Figure 3.
The chemical structures of activated COS used for different green chemical synthesis. (A) COS-fluoride, (B) p-nitrophenyl GlcN-GlcNAc dimer, (C) GlcN-DMP, (D) GlcNAc)n-oxazoline.
Figure 3.
The chemical structures of activated COS used for different green chemical synthesis. (A) COS-fluoride, (B) p-nitrophenyl GlcN-GlcNAc dimer, (C) GlcN-DMP, (D) GlcNAc)n-oxazoline.
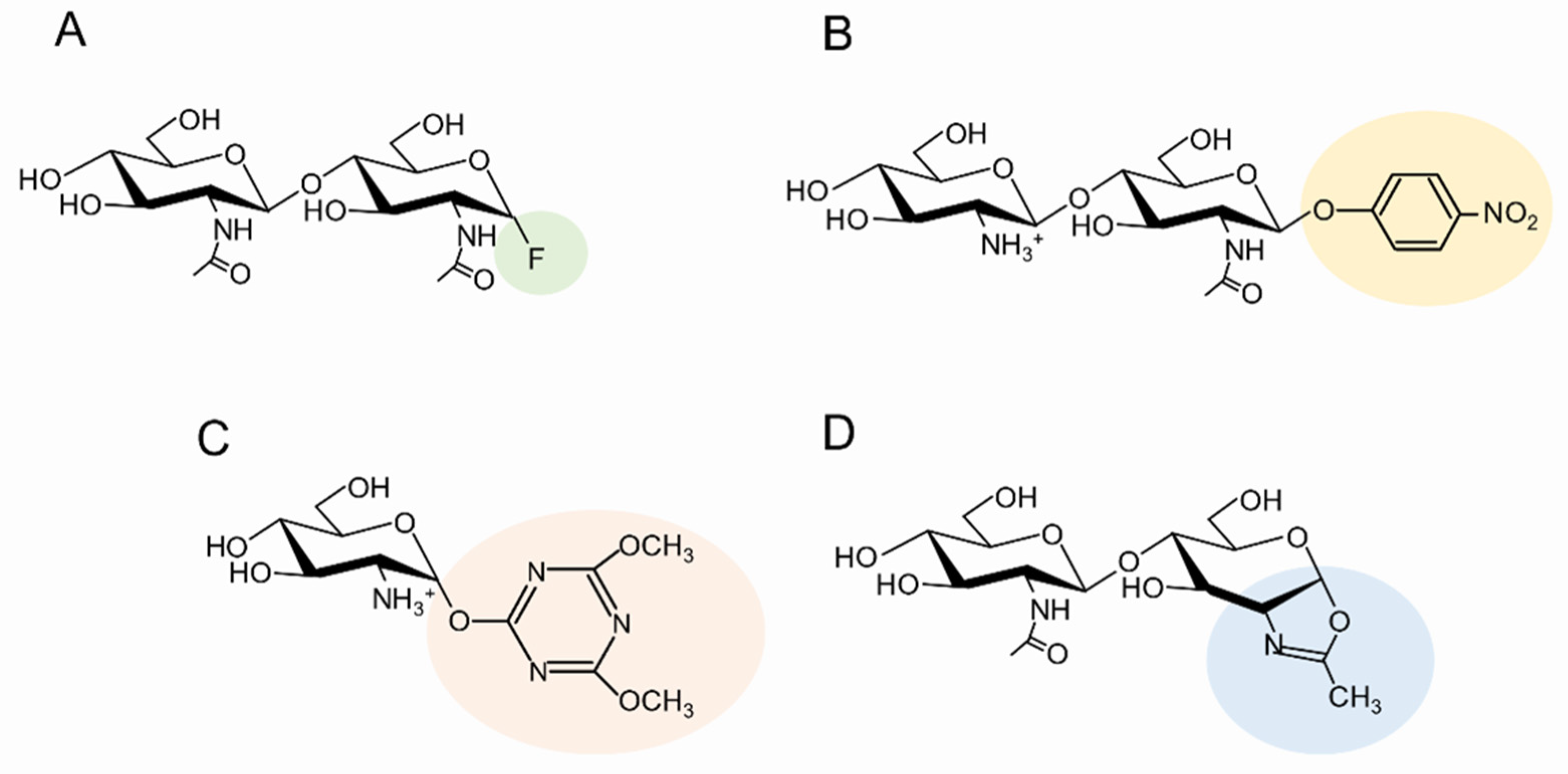
Figure 4.
Proposed enzymatic synthesis of tailor-made COS (DP4–10) through transglycosylation/glycosynthase reactions. Chitobiose and chitosan dimer are used as sugar acceptors and the reducing-end activated COS are used as donor substrates. Site-specific de-N-acetylation/N-acetylation are subsequently used in the final step to generate tailor-made COS.
Figure 4.
Proposed enzymatic synthesis of tailor-made COS (DP4–10) through transglycosylation/glycosynthase reactions. Chitobiose and chitosan dimer are used as sugar acceptors and the reducing-end activated COS are used as donor substrates. Site-specific de-N-acetylation/N-acetylation are subsequently used in the final step to generate tailor-made COS.
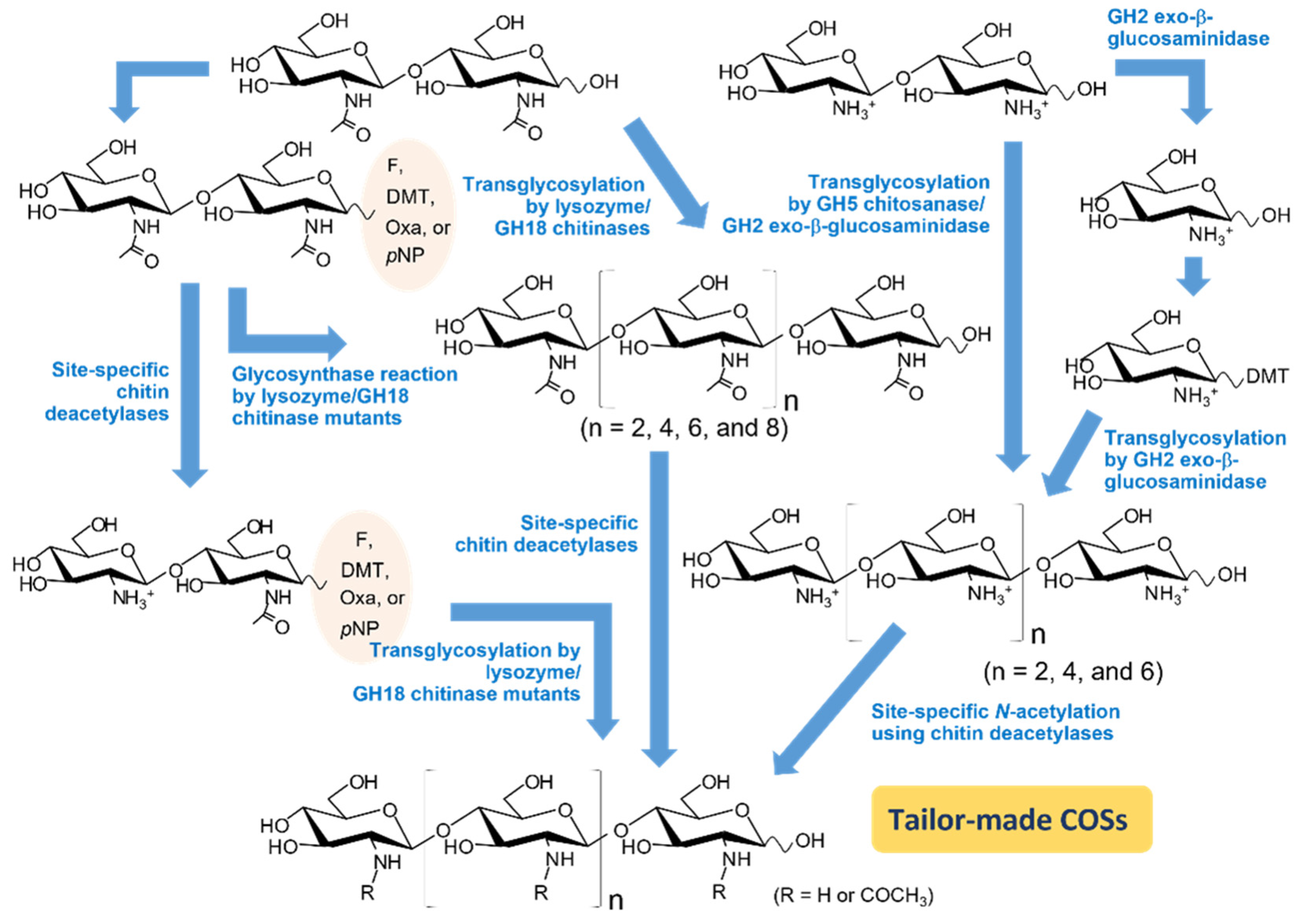

Table 1.
A summary of anti-microbial activity of COS.
| MW or DP | Source, DA or sequence | Activity | Assay | Ref | |
|---|---|---|---|---|---|
| Anti-microbial activity | Higher MW (10 kDa), Medium MW (5 kDa), and Lower MW (1 kDa) |
from chitosan | Higher activity (10 kDa) Lower activity (1 and 5 kDa) |
Growth inhibition toward Gram-positive, Gram-negative, and Lactic acid bacteria |
[9] |
| Chitosan polymers and Chitosan oligomers |
from chitosan | Higher activity (polymer) Lower activity (oligomer) |
Growth inhibition toward Staphyllococcus aureus Gram-positive and Gram-negative bacteria |
[10,11] | |
| DPs of 2, 3, and 4, and Lower MW Initial polymer |
DA, 0.15-0.5 chitosan |
Higher activity (DP of 2-4 and Lower MW) Lower activity (polymer) |
Growth inhibition toward fungi and bacteria |
[12] | |
| Average DPs of 17, 31, 54, and 62 | from chitosan (DA, 0.15) |
Higher activity (DP of 31 and 54) Lower activity (DP of 17 and 62) |
Growth inhibition toward yeast, fungi, including Candida |
[13] | |
| Chitosans (70 and 600 kDa) Chitosan oligomers |
from chitosan | Higher activity (oligomers) |
Growth inhibition toward Candida | [14] |
Table 2.
A summary of anti-inflammatory bioactivity of COS.
| MW or DP | Source, DA or sequence | Activity | Assay | Ref. | |
|---|---|---|---|---|---|
| Anti-inflammatory activity | COS with similar DP distribution | Different DAs of 0,0.12,0.50, and 0.85 | COS with DA of 0.12 has the highest activity | Inhibition of LPS-induced inflammatory cytokines burst, down-regulating its mRNA expression, and reducing phosphorylation of IκBα. | [15] |
| Chitosan oligomer mixture (DPs of 3-5) |
DA of 0.0 | Active toward allergic asthma inflammation in vivo | Inhibition of degranulation, cytokine generation in RBL-2H3 cells, and lung inflammation in allergic asthma model mice | [16] | |
| Chitosan oligomers with MW <1 kDa, 1-3 kDa, 3-5 kDa, and 5-10 kDa | from chitosan |
The lower the MW, the higher the activity. | Inhibition of iNOS and cyclooxygenase expressions | [17] | |
| MW<1 kDa (DP2-6; 4, 16, 29,37, and 14%, respectively) | DA <0.05 | Active toward N9 microglia cells | Inhibition of NO production by suppressing iNOS expression | [18] | |
| 42 %fully de-N-acetylated/54% mono-N-acetylated oligomers (42/54) and 50% fully de-N—acetylated/27% mono-N-acetylated oligomers (50/27) | from chitosan |
42/54 attenuated the inflammation both in vivo and in vitro, but 50/27 promoted the inflammatory response | Effects of COS preparations on the inflammation in lipopolysaccharide-induced mice and in RAW264.7 macrophages | [19] |
Table 3.
A summary of anti-oxidant bioactivity of COS.
| MW or DP | Source, DA or sequence | Activity | Assay | Ref | |
|---|---|---|---|---|---|
| Anti-oxidant activity | COS with DP of 1,2, and 3 | fully N-acetylated crab chitin (DA about 1.0) | Attenuated peroxidase activity and oxidation of DNA/membrane proteins, and exhibited free-radical scavenging effects | Experiments using human myeloid cells, HL-60 and mouse macrophages RAW 264.7 | [20] |
| Hetero-COS with DP of 2-6 | from housefly larvae powder | The COS with major DP of 5 exhibited the highest activity | Hydroxyl-scavenging effects | [21] | |
| COS | from chitosan (DA = 5%) |
Protective effects on H2O2-induced apoptosis or oxidative damage | Experiments using human umbilical vein endothelial cell | [22,23] | |
| COS with different DPs (3-7) | from chitosan | COS with DP of 6 has the highest activity | Effects on isoflavone contents and antioxidant activity in soyabean seeds during germination | [24] | |
| COS dimers, AA, DA, AD, and DD (A, GlcNAc; D, GlcN) |
Site-specific chitin deacetylase treatments of the purified (GlcNAc)2 |
DD>AD>>DA, AA toward superoxide radicals AA>DA>>AD, DD toward hydroxyl radicals DD>>AD, DA, AA toward DPPH |
All scavenging effects were examined in vitro | [25] |
Table 4.
A summary of anti-tumor bioactivities of COS.
| MW or DP | Source, DA or sequence | Activity | Assay | Ref | |
|---|---|---|---|---|---|
| Anti-tumor activity | (GlcNAc)6 | from crab chitin | Decreased the pulmonary nodules | Experiments using Lewis lung carcinoma mice | [26] |
| Chitin, chitosan 2.5-338 kDa |
from shrimp shell chitin | Activity of chitin (2.5 kDa) was higher than those of chitin (338 kDa) and chitosan (12 kDa) | Experiments using human leukemia cells | [27] | |
| COS from chitosan hydrolysis (DPs of 2,3, and 4 are the major components) | Enzymatic hydrolysis of high molecular weight chitosan | COS with lower MW> COS with higher MW | Experiments using prostate and lung cancer cells and hepatoma cells | [28] | |
| Chitosan oligomers DP 2-6 | Chemical and enzymatic hydrolysis of chitosan | For effective inhibition of the cell proliferation, DP should be at least 6 | Inhibitory effect on A549 cell proliferation | [29] |
Table 5.
A summary of COS as plant elicitors.
| MW or DP | Source, DA or sequence | Activity | Assay | Ref | |
|---|---|---|---|---|---|
| Plant elicitor | (GlcNAc)n (n=4,5,6,7,8,9, and 10), (GlcN)n (n=5,6, and 7) and Partially N-acetylated chitosans |
Chemical hydrolysis of chitin (fluorolysis) deacetylation of the high MW chitin |
(GlcN)n were not active as elicitors. (GlcNAc)n with a DP ≥7 strongly elicited POD activities but not PAL activities. Partially N-acetylated chitosans elicited both PAL and POD activities. |
Induction of phenylalanine ammonia-lyase (PAL), peroxidase (POD) in healthy wheat leaves | [31] |
| (GlcNAc)n (n=5,6, and 7) |
Enzymatic transglycosylation | (GlcNAc)7 strongly induced oxidative burst response as well as peroxidase and PAL activities. | Induction of phenylalanine ammonia-lyase (PAL), peroxidase (POD), PR protein gene expression in rice seedlings | [32] | |
| (GlcNAc)8 and hetero-COS with a DP of 8 (A, GlcNAc; D, GlcN) |
Enzymatic synthesis | AAAAAAAA was elicitor active, but DADADADA inactive | Inhibition of CEBiP-dimerization and Reactive oxygen generation | [37] | |
| (GlcNAc)6 and two hetero-COS with DP of 6 (A, GlcNAc; D, GlcN) |
Enzymatic deacetylation of (GlcNAc)6 |
AAAAAA > ADAAAA > ADAADA |
Inhibition of reactive oxygen generation | [38] |
Table 6.
Strategies for synthesizing COS with desired sequences.
| Catalysts | Substrates | Products | Ref. | |
|---|---|---|---|---|
| Donor | acceptor | |||
| Hen egg white lysozyme Wild type |
(GlcNAc)3 | (GlcNAc)n (n = 3–15) | [45] | |
| Hen egg white lysozyme Wild type |
(GlcNAc)4 | Moranoline (1-deoxynojirimycin) | 4-O-b-di(tri)-N-acetylchitobi(tri)osyl moranoline | [46] |
| Hen egg white lysozyme Wild type |
(GlcNAc)4 | 2-acetamide-2,3-dideoxy-glucopyranose | 44-O-β-d-galactosyl-β-tri-N-acetylchitotriosyl 2-acetamide-2,3-dideoxy-glucopyranose | [47] |
| Hen egg white lysozyme Asp101, Trp62-modified |
(GlcNAc)5 | (GlcNAc)9 | [51] | |
| Amycolatopsis orientalis GH2 exo-b-D-glucosaminidase | (GlcN)4 | (GlcN)5 and (GlcN)6 | [52] | |
| Streptomyces griseus HUT6037 GH5 endo-chitosanase | (GlcN)5 | (GlcNAc)3 | (GlcN)2-(GlcNAc)3 (GlcN)3-(GlcNAc)3 |
[53] |
|
Serratia marcescens GH18 chitinase A mutated at Trp at site -3. |
(GlcNAc)4 or (GlcNAc)5 | (GlcNAc)6 or (GlcNAc)7 | [54] | |
|
Serratia marcescens GH18 chitinases A and B mutated at the middle Asp in the DxDxE motif. |
(GlcNAc)4 | (GlcNAc)3 produced through the transglycosylation product (GlcNAc)6 | [55] | |
| Vibrio harveyi GH18 chitinase A mutated at the middle Asp in the DxDxE motif. | (GlcNAc)4 (GlcNAc)6 |
(GlcNAc)6 (GlcNAc)8 |
[56] | |
|
Serratia proteamaculans GH18 chitinase D triple-mutated at the glycon- or aglycon-binding aromatic residues as well as at the middle Asp in the DxDxE motif. |
(GlcNAc)4 | (GlcNAc)5 or (GlcNAc)6 | [57] | |
|
Serratia proteamaculans GH18 chitinase D single-mutated at the catalytic center and the binding groove. |
(GlcNAc)4 | (GlcNAc)5 or (GlcNAc)6 | [58] | |
| Arabidopsis thaliana GH18 chitinase C mutant, in which tryptophan side chain was introduced into the upper portion of the catalytic center. | (GlcNAc)4 | (GlcNAc)3 produced through the transglycosylation product (GlcNAc)6 | [60] | |
| Cycas revoluta GH18 chitinase A mutant, in which tryptophan side chain was introduced into the upper portion of the catalytic center. | (GlcNAc)4 | (GlcNAc)3 produced through the transglycosylation product (GlcNAc)6 | [61] | |
| A glycosynthase derived from Bryum coronatum GH19 chitinase A | (GlcNAc)2-fluoride | (GlcNAc)2 | (GlcNAc)4 | [66] |
| A glycosynthase derived from Secale cereale GH19 chitinase C | (GlcNAc)3-fluoride | (GlcNAc)4 | (GlcNAc)7 | [67] |
| A chitin-oligosaccharide N-deacetylase (NodB) and a glycosynthase derived from hen egg white lysozyme (Asp52→Ser) |
GlcN-(GlcNAc)2-fluoride | (GlcNAc)3 (GlcNAc)4 (GlcNAc)5 |
GlcN-(GlcNAc)5 GlcN-(GlcNAc)6 GlcN-(GlcNAc)7 |
[70] |
| Hyper-transglycosylating mutants from Serratia marcescens GH18 chitinases A and Serratia proteamaculans GH18 chitinase D | GlcN-GlcNAc-pNP (p-nitorophenylated) obtained by enzymatic de-N-acetylation of (GlcNAc)2-pNP | (GlcN-GlcNAc)2 (GlcN-GlcNAc)3 (GlcN-GlcNAc)4 (GlcN-GlcNAc)5 |
[72] | |
| Amycolatopsis orientalis GH2 exo-b-glucosaminidase | GlcN-DMT (4,6-dimethoxy-1,3,5-triazin-2-yl) | (GlcNAc)2 | GlcN-(GlcNAc)2 | [74] |
| A glycosynthase derived from Bryum coronatum GH19 chitinase A | (GlcNAc)2-DMT | (GlcNAc)2 | (GlcNAc)4 | [75] |
| An activity-suppressed mutant from Bacillus circulans GH18 chitinase A1 | (GlcNAc)2-oxa | (GlcNAc)5 | (GlcNAc)7 | [77] |
| Catalytic-site mutants from Serratia proteamaculans GH18 chitinase D | (GlcNAc)5-oxa | (GlcNAc)10 | [78] | |
| Hyper-transglycosylating mutants from Nicotiana tobaccum GH18 chitinase C and Cycas revoluta GH18 chitinase A | (GlcNAc)2-oxa (GlcNAc)3-oxa (GlcNAc)4-oxa (GlcNAc)5-oxa |
(GlcNAc)5 (GlcNAc)4 (GlcNAc)3 (GlcNAc)2 |
(GlcNAc)7 | [79] |
| Site-specific chitin dectylases from fungi | (GlcNAc)4 | A full lineup of partially N-acetylated chitotetraoses | [80] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



