
Submitted:
25 May 2023
Posted:
29 May 2023
You are already at the latest version
Abstract
A biocatalytic system that could produce bioactive resveratrol poly-glucosides using sucrose as a low-cost source of UDP-glucose donors and amylosucrase DgAS from Deinococcus geothermalis was developed in this study. This system boasts several advantages, including fast and direct conversion of substrates to products, thermostable, and regio-stereospecific, and effectiveness in vitro and in vivo at 40°C. With a success rate of around 97.0% in a short period in vitro and 95.0% in vivo, resveratrol-O-glucosides showed exciting outcomes in cosmetic activities, such as antioxidant, anti-inflammatory, anti-aging, and whitening effects when tested with Raw 264.7, B16, and HS68 cell lines. DgAS is recognized as an important biocatalyst due to its higher thermostability, effectiveness, and specificity among all known amylosucrases (ASases) in the production of poly-glucosides in a chain of polyphenols, like resveratrol, making it an ideal candidate for industrial use to cost-effective production of cosmetic items.
Keywords:
Cosmetic activity
; resveratrol
; glucosylation
; amylosucrase
1. Introduction
Resveratrol is a polyphenol synthesized by plants as a secondary metabolite to protect themselves from environmental stresses and microbial infections [1,2]. Chemically, it exists in both trans- and cis-forms due to a double styrene bond between two phenolic rings [3]. Resveratrol has gained popularity in health-related products, dietary supplements, cosmetics, and medicines due to its multiple beneficial properties for human health, such as cardiovascular protection, anti-aging, anti-cancer, and antioxidant effects [3].
Resveratrol has been reported to be able to eliminate free radicals such as reactive nitrogen species (RNS) and reactive oxygen species (ROS) that can cause damage to sensitive organs and lead to lipid peroxidation. Resveratrol can enhance cholesterol efflux in the blood, improves cardiovascular and neurological activities, and displays anticancer activities by regulating the expression of small RNA and enhancing transcription factors [4,5,6]. Additionally, resveratrol is effective in treating obesity, type-II diabetes, Alzheimer’s disease, atherosclerosis, ischemic injury, hypertension, and various cancers [7,8]. However, its poor solubility in an aqueous medium remains a challenge for its effective utilization [9,10]. Therefore, different methods have been applied to overcome this limitation, including the use of analog natural products [11].
Resveratrol is very sensitive to pH, light, and high temperatures due to its double C-C bond and unstable hydroxyl groups. Therefore, many studies have been carried out on the stability of resveratrol and make its application more extensive. At room or body temperature, trans-resveratrol is stable under low pH. However, it degrades quickly in alkaline conditions [12,13]. Some techniques such as Zein-pectin core/shell nanoparticles, Co-encapsulation of α-tocopherol, PEG-modified liposomes, and modification with cyclodextrin can be used to enhance the stability of resveratrol successfully [14,15,16,17]. Recent studies have focused on cosmetic application of resveratrol as it has shown many positive cosmetic values in addition to its nutraceutical and pharmaceutical properties [3,7]. Quality cosmeceutical products are highly demanded around the world. Many cosmetic industries are working to meet the demand for quality make-up items. As one popular chemical, ascorbic acid (AA) is commonly included in anti-aging and beauty creams due to its efficacy and essential role. However, AA has limitations since it is a water-soluble, weak and very unstable. It easily deteriorates or oxidizes when exposed to light, air, high temperatures, alkali, copper, and heavy metals. Ravetti et al. and Boo et al. have found that glucosylation at the 2-position of ascorbic acid (2-glucosyl ascorbic acid) can increase its stability against oxidation and reduction [18,19]. Similarly, some researchers have focused on enzymatic production of 2-glucosyl glycerol(2GG) as it is a highly expensive, stable, soluble, and valuable cosmetic ingredient of many beauty creams [20,21].
Glycosylation is a popular method for modifying natural products, including plant polyphenols such as flavonoids, to enhance their pharmacological properties such as bioavailability, solubility, activity, and stability [22]. However, chemical synthesis of glycosylated flavonoids faces challenges such as strict reaction conditions, protection and deprotection groups, low stereospecificity and regioselectivity, and difficulty in purification, resulting in high production costs and time [23]. To overcome these obstacles, microbial cell factories have been engineered with appropriate sugar biosynthetic pathway genes and glycosyltransferases(GTs) from heterologous sources for the production of targeted glycosylated flavonoids [24,25]. However, high concentrations of some metabolites might be toxic to engineered cells [26,27].
An alternative to microbial cell factories is in vitro enzymatic glycosylation using amylosucrase ASase (EC 2.4.1.4), a versatile enzyme with hydrolysis, isomerization, and transglycosylation activities [28]. Amylosucrase from different sources has been used to transglycosylate different types of polyphenols including flavonoids [28,29,30]. However, glycodiversification of flavonoids using amylosucrase is challenging, despite attempts to engineer these enzymes to improve their tolerance to various sucrose analogs [31]. In this study, we used DgAS, an amylosucrase obtained from Deinococcus geothermalis DSM 11300, for the direct transglycosylation of resveratrol [30].
In this study, we employ a DgAS, amylosucrase, to produce resveratrol glucosides more effectively, easily, fastly, and environmentally friendly in vivo and in vitro systems, using sucrose as a cheap source of glucose (Figure 1) [32,33]. We focused on evaluating a wide range of cosmetic properties of resveratrol-O-glucosides using HS68, B16, and Raw264.7 cell lines for their potential industrial application.
2. Materials and Methods
2.1. Chemicals and Reagents
Resveratrol, starch, maltodextrin, cyclodextrin glucanotransferase (CGTase, from Bacillus sp.), and amyloglucosidase (from Aspergillus niger) were purchased from Sigma-Aldrich (USA). Sucrose was purchased from MBcell (Seoul, Korea). Luria-Bertani (LB) broth medium was bought from KinsanBio (Seoul, Korea). All other chemicals and required reagents were of the highest grade. They were purchased from different sources. High-performance liquid chromatography (HPLC) grade acetonitrile and water were purchased from Mallinckrodt Baker (Phillipsburg, NJ, USA). All restriction enzymes were purchased from Takara Bio (Japan) and Promega (USA).
2.2. Bacterial Strains, Cloning, and Culture Conditions
For the expression and production of DgAS in pHCE IIB (NdeI), Escherichia coli BL21 (DE3) (Stratagene, La Jolla, CA, USA) was used. For all DNA manipulation, E. coli XL1 blue was used as a host. Recombinant E. coli strains were grown in LB broth or an agar plate supplemented with ampicillin (100 μg/ml) at 37 °C. The recombinant strain E. coli (DE3) harboring pHCE IIB (NdeI)-DgAS was prepared by heat shock transformation. Nucleotide sequence of amylosucrase (GenBank Accession No. ABF44874.1) obtained from Deinococcus geothermalis DSM 11300 was codon-optimized and synthesized by General Biosystems (USA). With the restriction sites NdeI and HindIII, the DgAS gene was synthesized with a 1,984 bp size.
2.3. Protein Expression and Analysis
The seed culture of recombinant E. coli BL21 (DE3) harboring DgAS was prepared in LB broth medium supplemented with ampicillin antibiotic and incubated in a shaking incubator (200 rpm) at 37ºC overnight. Then 500 μL of seed culture was used to fresh 100 ml LB medium with the 100 µL ampicillin antibiotic in a 500 ml conical flask and incubated at 37 °C for 24 h in a shaking incubator without using IPTG for the induction. Cell pellets were then harvested by centrifugation at 842 x g (3,000 rpm) for 10 min, and washed (vortexed, followed by centrifugation) with buffer (200 mM Tris-HCl and 10% glycerol of pH 7) twice, and re-suspended with 1mL of the same buffer. Cell pellets were lysed using a Fisher Scientific Sonic Dismembrator Model 500 (5-9 sec pulse on and off, total 360 sec, at 20% amplitude) in an ice bath. Clear lysate was collected by high-speed centrifugation at 12,000 rpm (13,475 x g) for 30min at 4 °C. The crude protein obtained was further analyzed by 12% (w/v) sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). Soluble fraction of DgAS was purified using nickel nitrilotriacetic acid (Ni-NTA) affinity column chromatography (Qiagen Inc., USA) and eluted with different concentrations (10, 100, 200, and 500 mM) of imidazole, respectively, as reported previously [30]. Crude protein concentration was determined by using the Bradford method (Bradford et al. 1976), with bovine serum albumin as the standard.
2.4. Enzymatic Reaction
Glucosylation reaction was performed using resveratrol as an acceptor molecules in the presence of sucrose as a glucose donor catalyzed by amylosucrase (DgAS). A 200 μl of the total reaction mixture contained 0.2 mM resveratrol, 50 mM sucrose, 2 µg/ml amylosucrase enzyme, and 200 mM Tris-HCl buffer at pH 7. Resveratrol was dissolved in Dimethyl sulfoxide (DMSO) while sucrose was dissolved in water to make the required stock solution. Reactions were incubated at 40 °C. Reaction samples were taken out at 0.5 h, 1 h, 2 h, 3 h, and 4 h. Further reactions were stopped by treating the samples with chilled methanol and centrifuged at 13,475 × g for 30 min to remove denatured proteins. After that, samples were analyzed by high-performance liquid chromatography-photo diode array (HPLC-PDA) and confirmed by electrospray ionization mass spectrometry (ESI/MS) and nuclear magnetic resonance (NMR) analyses. Assay mixtures lacking enzymes and substrate resveratrol served as negative and positive controls, respectively.
2.5. Optimization in Vitro Reaction
Resveratrol concentration: Experiment conditions were changed by varying resveratrol concentrations (0.5, 1.0, 1.5, 2.0, 2.5, 5.0, 10.0, and 15.0 mM) while keeping other components constant as mentioned above. The concentrations of DgAS was maintained at 2 µg/ml in all reactions. Reactions were analyzed after incubation at 40ºC for 3h. Reaction samples were analyzed by reverse-phase HPLC after 3 h by quenching with chilled methanol.
Sucrose concentration: Experiment conditions were changed by varying sucrose concentrations (25, 50, 100, 150, 200, and 250 mM) while keeping other components constant as mentioned above. The concentrations of DgAS were maintained at 2 µg/ml in each reaction. Resveratrol was used at 0.5 mM concentration. Reactions were analyzed after 3h to find effect of changing concentrations of sucrose.
Enzyme concentration: Different identical sets of reaction conditions were set up by changing the concentration of the DgAS. Concentrations of purified DgAS enzyme were maintained as 0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 4.0, and 5.0 µg/ml in reaction sets separately. Reaction mixtures containing 0.5mM resveratrol, 100 mM sucrose, and 200 mM Tris-HCl (pH 7) buffer at pH 7, were incubated at 40ºC for 3 h.
Temperature: Similarly, identical sets of reactions were carried out in 200 µL total volume using 200 mM Tris-HCl (pH7), 0.5 mM resveratrol, and 10 0mM sucrose (final concentration) in the reaction mixture. During reactions, the concentration of DgAS was maintained at 2 µg/ml. Reaction assays were incubated at 25, 30, 35, 40, 45, 50, 55, and 60 °C for 3 h.
Buffer or pH optimization: To determine the optimal pH, identical sets of reaction conditions were made as in temperature optimization reactions while varying the pH of phosphate buffer in the range 5 and 6, working Tris-HCl buffer with pH 7 and 8, and glycine buffer with pH of 9. Reactions were carried out at 40ºC for 3 h.
2.6. Reaction with Different Sugar Donors and Commercial Enzyme
Separately identical sets of reactions were made at the same reaction conditions as above. Instead of sucrose, other sugar donors like starch, maltodextrin (4~7), maltodextrin (13~17), and maltodextrin (16.5~19.5) were used along with cyclodextrin glucanotransferase (CGTase, from Bacillus sp.), a commercial enzyme.
2.7. In Vivo Preparation of Resveratrol Glucosides
Seed culture of recombinant E. coli BL21 (DE3) harboring DgAS was prepared in LB broth medium supplemented with ampicillin antibiotic and incubated in a shaking incubator (200 rpm) at 37ºC overnight. Then 500 μL of seed culture was used to fresh 100 ml LB medium with the 100 µL ampicillin antibiotic in a 500 ml conical flask. The culture was incubated at 37 °C for 24 h in a shaking incubator without using IPTG for the induction. After that, 1mM resveratrol and 100 mM sucrose feeding were done and incubated for 36h at 37ºC in a shaking incubator at 200rpm. During incubation, samples (1mL each) were collected at 10 h and 36 h, and reactions were quenched by adding 1ml of chilled ethanol followed by vortexing for 10 min. Aliquots were then centrifuged at 12,000 rpm (13,475×g) to remove denatured protein. Samples were then analyzed by RP-HPLC.
2.8. Preparative Scale Production of Resveratrol Glucosides
large-scale production reaction was carried out by using 1 mM resveratrol, 100 mM sucrose, 200 mM Tris-HCl (pH7), and 2 µg/ml DgAS in the reaction mixture. The total volume of the reaction mixture was 20 ml in a 50 ml tube. The reaction was incubated in a shaking incubator at 40ºC for 5 h. Samples were analyzed at different timepoints of 3 h, 4 h, and 5 h. The reaction was stopped by boiling for 5mins in a water bath to denature enzymes. After that, the mixture was centrifuged at 12,000 rpm (13,475 x g) for 15 mins. The supernatant was concentrated using a rotatory evaporator and used for purification of products by preparative HPLC.
2.9. Analytical Methods
Reverse-phase HPLC-PDA analyses were performed at 280 nm UV absorbance using a C18 column (Mightysil RP-18 GP (4.6 × 250 mm, 5 μm) (Kanto Corporation, Oregon)) connected to a photodiode array (PDA). HPLC was performed using a binary condition of water (0.1% trifluoroacetic acid (TFA)), and 100% acetonitrile (ACN) maintained at a flow rate of 1 ml/min for the 25 mins program. The flow rate of ACN concentrations was set throughout as 0- 40% (0-15 min), 40-75% (15-20 min), and 75-0% (20-25 min). Products were quantified by a calibration curve of authentic standard substrate resveratrol curve created using different concentrations (0.625, 1.25, 2.5, 5, 10, 20, and 40 µM). The exact mass of products was analyzed using high-resolution quadrupole time-of-flight electrospray ionization mass spectrometry (HR-QTOF-ESI/MS) [ACQUITY (UPLC, Waters, Milford, MA)-SYNAPT G2-S (Waters)] in the positive ion mode. Compounds were purified using preparative-HPLC with a C18 column (YMC-PACK ODS-AQ (250×20 mm I.D., 10 μm particle size) connected to a UV detector (280 nm) under a binary condition of H2O (0.05% TFA) and 100% ACN at a flow rate of 10 ml/min for 35 min. The ACN concentrations were 10, 30, 50, 90, and 10% for 0-15, 15-20, 20-25, 25-32, and 32-35 min, respectively. The purified product was concentrated using a rotatory evaporator, followed by lyophilization. Purified compounds were structurally determined by Avance II 300 Bruker (Germany) BioSpin NMR spectrometer equipped with a TCI CryoProbe (5mm). All the samples were exchanged with D2O and dissolved in DMSO-d6 for nuclear magnetic resonance (NMR) analysis. Compounds were further characterized by a 700 MHz Avance II 900 Bruker BioSpin NMR spectrometer (Germany) using a Cryogenic TCi probe (5 mm). One-dimensional NMR (1H-NMR, 13C-NMR) was performed to elucidate the structure of the compounds. All the raw data were processed by using TopSpin 3.1 software (Bruker) and further analyzed by using MestReNova 8.0 software (Mestrelab Research S. L., Spain).
2.10. Assay of Resveratrol Glucosides for Cosmetic Activities
Cytotoxicity: Raw 264.7 cells, macrophage cell line, were seeded into 96-well cultures plates at a density of 1.0 × 104 cells/well. After 24 h, cells were treated with resveratrol and resveratrol-O-glucoside at indicated concentrations in a medium containing 10.0% of fetal bovine serum (FBS; Gibco, Thermo Fisher, USA) for 48h. HS68 cells (human fibroblast cells) were seeded into 96-well culture plates at a density of 0.5 × 104 cells/well. After 24 h, cells were treated with resveratrol and resveratrol-O-glucoside at various concentrations 1.563-400 μM) in a medium containing 2.0% of FBS (Samchun, Korea) for 48 h. B16 cells, a mouse melanoma cell line, were seeded into 96-well culture plates at a density of 8.0ⅹ103 cells/well abd cultured overnight.Cells were then treated with resveratrol and resveratrol-O-glucoside at various concentrations in a medium containing 5.0% of bovine serum (FBS;ATCC, Manassas, VA, USA) and 1% penicillin-Streptomycin mixtire (PS’ Lonza, Basel, Switzerland) for 72h.
Cell viability was determined using a Quanti-MAX™ WST-8 cell viability assay kit (Biomax, Korea) according to the manufacturer’s protocols. After treatment with 10.0%, WST-8 solution was added into each well from which the medium was removed. After incubation for 1h, the plate was measured at 450 nm using a fluorescence microplate reader.
Anti-inflammation: Raw 264.7 cells, a macrophage cell line, were seeded into 96-well culture plate at a density of 1.0 × 104 cells in 96-well culture plates. After 24 h, Raw 264.7 cells were treated with resveratrol and resveratrol-O-glucoside at various concentrations (0.625, 1.25, 2.5 μM) in the presence of LPS (100 ng/mL) for 48 h in the medium in the absence of phenol red containing 10.0% of fetal bovine serum (FBS; Gibco, Thermo Fisher, USA). After incubation, cell supernatants were collected from each well and NO contents were determined using a Griess reagent system kit (Promega, USA) per the manufacturer’s protocol. The plate was measured at 540 nm using a fluorescence microplate reader. Results were normalized to the NO standard.
Anti-aging: HS68 cells (human fibroblast cells) were seeded into 24-well culture plates at a density of 2.0 × 104 cells/well. After 24 h, cells were treated with resveratrol and resveratrol-O-glucoside at various concentrations (3, 6, 12 μM) in a medium containing 2.0% of FBS (Samchun, Korea) for 48h. For comparison, transforming growth factor beta 1 (TGF-β1) (20 ng/mL) was used as a positive control. After incubation, cell supernatant was collected from each well and pro-collagen content was determined using a human pro-collagen Ⅰ alpha 1 ELISA assay kit (R&D Systems Inc, US) followingh the manufacturer’s protocol. The plate was measured at 450 nm using a fluorescence microplate reader. Results were normalized against the pro-collagen standard.
Whitening: B16 melanoma cells were seeded into 48-well cell culture plates at a density of 2.5 × 10⁴ cells/well and incubated overnight. To induce melanin production, B16 cells were incubated with various concentrations of the test compounds in the presence of α-MSH (0.1 μM) in a phenol red-free cell culture medium in 48-well plates. After 72 h, extracellular melanin content was measured at 405nm using a Synergy™ HTX Multi-Mode Microplate Reader (Winooski, VT, USA) by transferring 100 µl of the medium in which B16 cells were cultured to 96-well plates. Intracellular melanin content was calculated and corrected according to protein concentration. Control cells considered to have 100% intercellular melanin content. To determine protein concentration, 200 µl of 1 N NaOH was added to dissolve cells at 60 °C for 30 mins. After that, 100 µl of cell lysate was added to a 96-well plate and absorbance was measured at 450nm using a microplate reader. Protein concentration of each sample was determined by Pierce™ BCA Protein Assay.
Antioxidant: DPPH((2,2-diphenyl-1-picrylhydrazyl) radical scavenging activity was measured to determine the antioxidant effect of each substance. After 100µl of 0.4 mM DPPH (Sigma Aldrich, USA) solution was added to 100 µl of sample, the plate was incubated in a dark room for 15 mins. Absorbance was then measured at 520 nm on a microplate reader. L-ascorbic acid (Samchun, Korea) was used for a positive control.
3. Results
3.1. In Vitro Reaction of Resvatrol
Protein expression and purification: To produce soluble recombinant proteins for, in vitro reaction, pCHE-DgAS (Figure S1 of the Supplementary Information) were transformed into E. coli BL21 (DE3) host. SDS-PAGE analysis of a soluble fraction of protein DgAS showed a clear band at around 72 kDa (Figure S2). The band size corresponds to the calculated molecular weight of a hexahistidine-tagged fusion protein. The soluble lysate containing protins was then subjected to purification using Ni++-NTA beads. Purified proteins were concentrated, quantified, and used for in vitro reactions.
Glucosylation of resveratrol: The reaction mixture of resveratrol with sucrose and DgAS was first analyzed by RP-HPLC-PDA. Four major different peaks, P4, P3, P2, and P1, in chromatograms appeared at retention time (tR) of 11.80, 12.03, 12.56, and 13.01 min respectively (Figure 2A). The UV-VIS analysis of the peaks resembled resveratrol with a subtle difference in the pattern (Figure 2B). The same sample was subjected to high-resolution QTOF-ESI/MS analysis. Further analysis observed that the exact mass [M+H]+ m/z+ of a product as 391.1381, which was a single glucose conjugated resveratrol. The calculated mass of single sugar-conjugated resveratrol with the formula C20H23O8+ in the protonated form [M+H]+ m/z+ was 391.1387 (Figure 2C(ii)). Along with this, di-glucosylated, tri-glucosylated, and tetra-glucosylated masses of resveratrol were seen as 553.1902, 715.2429, and 877.2952 respectively (Figure 3C). These masses exactly matched calculated [M+H]+ m/z+ masses for molecular formula of C26H33O13+, C32H43O18+, and C38H53O23+ with the masses of 553.1916, 715.2444, and 877.2972 respectively, which were corresponding masses of di-glucosylated, tri-glucosylated, and tetra-glucosylated of resveratrol (Figure 2C). The overall conversion of resveratrol to glucosides was about 97.0%.
During the reaction, samples taken at different time points were analyzed by RP-HPLC. The conversion of the resveratrol to its glucosides was increased till 3 h (~ 97.0%). It then remained the same even though further incubation of the reaction (Figure S3). Therefore, 3h incubation time had taken as the optimized time for the in vitro reaction.
Resveratrol substrate tolerance: Eight sets of identical reactions were carried out at various resveratrol concentrations. Reactions contained 2 µg/mL DgAS. Other components of the reactions were fixed. Product formation and substrate conversion were monitored after 3 h. RP-HPLC analysis of the sample showed that when the concentration of resveratrol was 0.5 mM, maximum conversion of the substrate occurred (Figure S4). Therefore, 0.5 mM of resveratrol had taken as the optimized concentration for further reactions.
Optimization of sucrose concentration: Six sets of experiments were carried out using different concentrations of sucrose while keeping all other reaction components constant. Result showed that the maximum conversion of (~97.0%) resveratrol at 100 mM of sucrose. However, when the concentration of sucrose was 25 mM, the conversion was about 80.0% only. Moreover, with a concentration of sucrose of more than 100 mM, there was a decrease in the conversion of the resveratrol (Figure 3(i)). These result showed an efficient supply of glucose for the glucosylation reaction. For further reactions, 100 mM of sucrose was used.
Enzymes concentration: To identify the optimized concentration of the enzyme for reactions, eight sets of identical reactions were carried out and the outcome of the reaction was closely monitored for the optimal conversion of resveratrol. As expected, a different pattern of product profile was obtained while varying the enzyme concentration in the reaction. The conversion percentage of resveratrol at the concentration of 0.5 µg/ml was about 42.0% which increased at a higher concentration of DgAS and reached about 90.0% at 2 µg/ml and decreased slightly at 2.5 µg/ml, however, slightly increased at 3 µg/ml (Figure 3(ii)). Therefore, 2 µg/ml of DgAS was taken for further reactions.
Effects of temperature: Results obtained in the different ranges of temperature from 25 to 60°C showed multiple impacts on product formation (Figure 3(iii)). At 25ºC, only two products, P1 and P2 had formed while at 55 and 60°C, only one product P1 had formed. The maximum conversion of the resveratrol at 40ºC was found to be about 97.0%. All products (P1, P2, P3, and P4) formed at a temperature range of 30 to 50ºC. The optimal reaction temperature was 40°C.
Effects of buffer: To find out the optimal pH for the reaction assay, glucoside production was investigated at 40°C, keeping all other reaction ingredients intact while changing the buffer and pH. The pH of the reaction was agjusted to be 5 to 9 in 200 mM of phosphate, Tris-HCl, and glycine buffer with their respective pH as mentioned in the Methods and Materials. The reaction was monitored after incubating at 40°C for 3h. Results showed that at low pH 5 and 9, there was less conversion of resveratrol (~ 20.0%) to its glucosides (Figure 3(iv)). Conversion of the resveratrol was the highest at pH (6 and 7), which was about 97.0%. Therefore, the optimal pH for the reaction was 7. All products formed at pH 7 were found to be measurable. They were dominant under reaction conditions (Figure 3(iv)).
3.2. Effect of Using Different Sugar Donors and Commercial Enzyme
After analyzing results obtained from RP-HPLC chromatograms of samples incubated at 40ºC for 3h using different sources of sugar donors and commercial enzyme of CGTase, the maximum conversion of resveratrol was found with sucrose and DgAS (Figure S5). The conversion rate with DgAS and sucrose was found to be about 97.0%. With CGTase and different sugar donors (starch, maltodextrin, maltodextrin, and maltodextrin), conversion rates were around 20.0% only.
3.3. Preparative Scale Production of Resveratrol Glucosides
When product profiles at different time points were analyzed, four distinct peaks were initially observed. Among them, the peak at tR 13.01 min (P1) was initially the most dominant. However, with increasing incubation time from 3 h to 4 h and 5 h, peaks at tR 12.56 min (P2), 12.03 min (P3), and 11.80 min (P4) gradually increased, while that of P1 gradually decreased. The conversion of resveratrol to its glucosides was found to increase more with increasing incubation period in large-scale reactions (Figure 4 and Figure S6). Based on the nature of product formation, the amount of desired products can be controlled by changing the incubation period of the reaction. As time increased, mono-glucosylated products changed into di-glucosylated or poly-glucosylated products. The overall conversion of the resveratrol to its glucosides was about 97.0%. Calculated converstion rates for regular products of P1, P2, P3, and P4 at 5 h were 38.8, 28.8, 16.2, and 12.3%, respectively (Figure 4).
3.4. In Vivo Production of Resveratrol Glucoside
During in vivo production of resveratrol glucosides, the first sample was taken out at 10h and analyzed by RP-HPLC. The chromatogram of the sample showed a single peak at a retention time (tR) of 13.01 min (Figure 5A(ii)), which was exactly the same as the peak obtained in the in vitro reaction. Another sample was taken at 35h and analyzed by RP-HPLC as described earlier. It was found that the standard substrate was dcreased with more products peaking at retention time of 11.80, 12.03, and 12.56 min (Figure 5A(iv)).
When the same samples were subjected to high-resolution QTOF-ESI/MS analysis, the exact mass [M+H]+ m/z+ of a product was 391.1348, which resembled the single glucose conjugated resveratrol of the 10h sample. The calculated mass of single sugar-conjugated resveratrol with a formula C20H23O8+ in a protonated form [M+H]+ m/z+ was 391.1387 (Figure 5B,C). Moreover, with the 35h sample, exactly the same pattern of masses was obtained with peaks as seen in in vitro sample analysis (Figure 2C). These results clearly indicate that resveratrol glucosides by DgAS could also be formed in in vivo systems.
3.5. Structural Elucidation of Resveratrol Glycosides Products
Four major peaks produced during the preparative scale were purified using prep-HPLC. According to HPLC analysis, the purity level of each compound was more than 95.0%. Purified compounds were dried using a rotatory vacuum evaporator, followed by lyophilization for preparing NMR samples as described in the Materials and Methods section. HR-QTOF-ESI/MS peaks P1, P2, P3, and P4 showed the presence of single, di-, tri-, and tetra-glucosylated mass in resveratrol, respectively (Figure 2C and Figure 5B).
Furthermore, 1H-NMR analysis of P1 showed the absence of one proton peak at a chemical shift value δ 8.18 ppm (3-OH), meaning that a glucose molecule was attached at the 3-OH position of resveratrol. The presence of an anomeric proton peak at δ 5.34 ppm (d, J=3.7Hz) also confirmed the glucose unit at the alpha configuration. Similarly, in 13C-NMR analysis, an anomeric carbon peak appeared at δ 97.11ppm. Details of 1H- and 13C- data are shown in Table 1 and Table 2.
In the same way, in 1H-NMR analysis of P2, two proton peaks showed chemical shift values of δ 8.18 ppm (3-OH) and δ 6.14 ppm (4ˈ-OH), indicates glucose attached at 3-OH and 4ˈ-OH positions, respectively. In 13C-NMR, an anomeric proton at δ 5.34 ppm (d, J=3.7 Hz), an anomeric carbon peak at δ 97.76 ppm, and the protonated mass of two glucoses confirmed the structure of resveratrol as one-one glucose attached at 3-OH and 4ˈ-OH position of the resveratrol. However, after analyzing P3 and P4, proton peaks at δ 8.18 ppm (3-OH) and δ 6.94 ppm (4ˈ-OH), respectively, were missing. Further analysis of peaks of both products at the similar anomeric proton position at δ 4.85 (d, J=3.6 Hz) and their masses clearly indicated that P3 contained three glucose polymer chains at 4ˈ-OH and P4 had four glucose polymer chains at the 3-OH position with alpha configuration. The presence of anomeric carbon at δ 98.45 ppm further supported the structure of P3 and P4. All details of 1H-NMR and 13C-NMR are listed in Table 1 and Table 2.
According to the 1D NMR and HR-QTF-ESI/MS analyses, the four compounds were identified as resveratrol-3-O-α-glucoside (P1), resveratrol-3,4ˈ-O-α-diglucoside (P2), resveratrol-3-O-α-triglucoside (P3), and resveratrol-4ˈ-O-α-tetraglucoside (P4) (Figure S7).
3.6. Cosmetic Activities of Resveratrol Glucosides
Effects of resveratrol-O-glucosides and resveratrol on cell viability were determined using Raw 264.7 cells, HS68 cells, and B16 cells as mentioned in the above method and material. After incubation of 72h, samples were tested. Results revealed that resveratrol-O-glucosides were less toxic to B16 cells than resveratrol. After cells were treated with 100µM resveratrol-O-glucosides or 100µM resveratrol, cell viability was around 50.0% or around 30.0%, respectively (Figure 6A). When Raw 264.7 cells were treated with 200 µM resveratrol-O-glucosides or resveratrol for 48h, cell viability was more than 95.0% or only around 10.0%, respectively (Figure 6B). Similarly, when HS68 cells were treated with 400 µM resveratrol-O-glucoside or the orginal resveratrol, cell viability of around 100% (Figure 6C). Therefore, the cytotoxicity of resveratrol glucosides is much less than that of resveratrol.
Raw 264.7 cells (a macrophage cell line) were seeded into 96-well culture plates at a density of 1.0 ⅹ103 cells/well as mentioned in the method and material. After incubation for 48 h, samples were tested according to standard protocols and results were analyzed. Results clearly showed that both resveratrol and resveratrol-O-glucosides had anti-inflammatory effects with different IC50 values (Figure 7A).
As mentioned above, HS68 human fibroblast cells were seeded in a 24-well culture plate at a density of 20 ⅹ104 cells/well and incubated for 48h. Supernatant samples were then collected to determine pro-collagen contents. Results showed that both resveratrol and resveratrol-O-glucosides had anti-aging effects (Figure 7B).
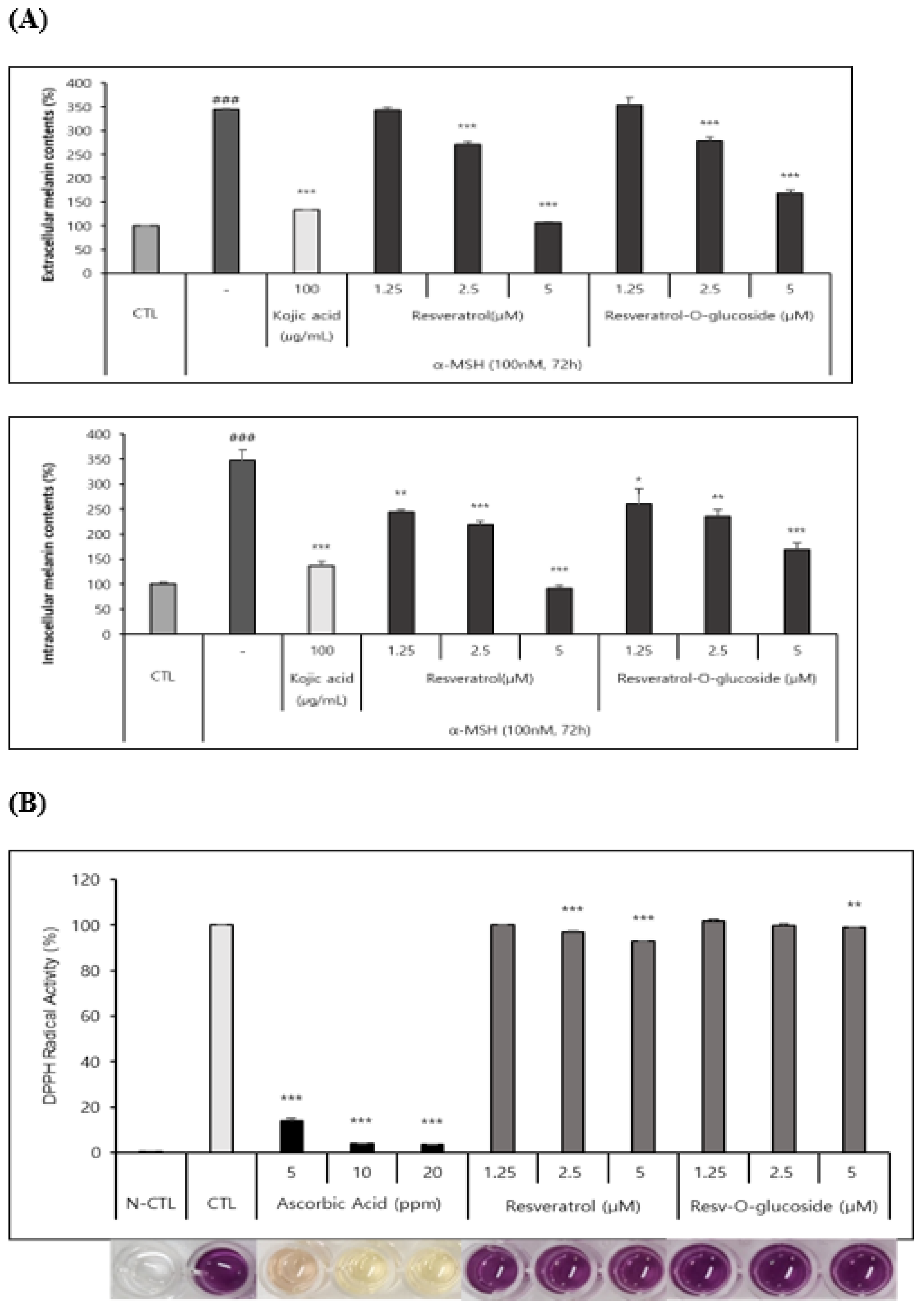
As described in the Materials and Methods section, B16 melanoma cells were seeded in a 48-well culture plate at a density of 2.5 ⅹ104 cells/well and cultured overnight. They were then incubated with control and resveratrol compounds for 72h. These samples were then analyzed according to the standard protocol mentioned above. It as found that both resveratrol and resveratrol-O-glucosides had whitening effects, although they had different IC50 values (Figure 8A).
DPPH radical scavenging assay was carried out as described in the Materials and Methods section to determines the anti-radical power of antioxidants. Samples were analyzed after incubation for 15 min at 515 nm wavelength in a microplate spectrophotometer. It was found that both resveratrol and resveratrol-O-glucosides had similar antioxidant activities with different IC50 values (Figure 8B).
4. Discussion
Resveratrol and its derivatives have gained considerable interest and demand due to their potent biological activities in medical and nutraceutical fields [3,5]. Recent advancement in biological tools and techniques has enabled cost-effective and rapid production of such bioactive plant products in microbial hosts. Although microbial hosts lack resveratrol biosynthetic genes, they offer a promising alternative source for large-scale production of resveratrol and its derivatives using metabolic engineering and synthetic biology approaches [33,34]. However, despite the use of latest techniques, the overall production of resveratrol and its derivatives remains insufficient [35,36]. Therefore, optimizing each gene involved in the biosynthetic pathways is necessary to improve enzyme activity in trans-located hosts by supplying sufficient precursors and regulating resveratrol concentration within the cell, which could enhance production [34,37]. Furthermore, research has demonstrated that resveratrol derivatives exhibit higher antioxidant activities than resveratrol [38]. Due to their higher stability against light, oxygen, and extreme pH than resveratrol, the production of resveratrol derivatives is increasing and becoming more demanding not only for their high medicinal, nutritional, and cosmetic values, but also for their superior properties.
In this research, we devised in vitro and in vivo biocatalytic systems using enzyme amylosucrase (DgAS) derived from Deinococcus geothermalis DSM 11300 for efficient transglycosylation of resveratrol with sucrose as the glycosyl donor [3,30]. Amylosucrase is a versatile enzyme with several industrial applications such as hydrolysis, polymerization, isomerization, and transglycosylation [39]. During the process of sucrose hydrolysis, the α-1, β-2-glycosidic bond of sucrose breaks down, leading to the formation of sucrose isomers of turanose and trehalulose. Similarly, in isomerase activity, sucrose isomerizes to produce turanose and trehalulose, while transglycosylation activity produces α-1,4-glucans such as amylose polymers [40,41]. DgAS can transfer the glucosyl moiety from sucrose to glucose or fructose without using costly UDP-glucose as a substrate [30]. However, the transglycosylation of resveratrol mostly occurs at the C-3 or C-4ˈ position due to position-specific transglycosylation rather than at the C-5 position, which is sterically hindered [42]. Studies have shown that DgAS attaches four or fewer glucose units to polyphenols during transglycosylation [42]. Our results indicated the formation of four major products, with product P4 exhibiting a four-glucose-unit chain attached to the 4ˈ-OH position of resveratrol, product P3 displaying a three-glucose-polymer chain at the 3-OH position, product P2 containing glucose moieties at both 3-OH and 4ˈ-OH positions with a di-glucosylated mass, and product P1 being identified as resveratrol-3-O-α-glucoside (Figure S7). Although DgAS exhibited high specific activity and thermostability, its polymerization activity was relatively lower than those of other ASases owing to its exceptional thermostable characteristics with a half-life of 69 h at 50°C [43,44]. Moreover, when DgAS was utilized to transglycosylate arbutin, it transferred a glucose unit of sucrose to the C-4 position in the glucose residue of salicin and arbutin, with the major arbutin glycoside transfer product being determined as glucosyl salicin [45,46].
Although both in vitro and in vivo systems are effective in producing resveratrol-O-glucosides, each has its advantages and disadvantages. In vitro systems using DgAS are fast, direct, easy to handle, environmentally friendly, and requiring fewer steps for product extraction and purification [30,33]. However, these systems require large amounts of protein and a longer time for the purification of enzymes for scale-up production. In contrast, in vivo systems do not require additional protein purification. Howefver, they require a longer time for culture to harvest products. They might have additional bacterial product peaks that might create an extra burden for purification. Ultimately, the choice between in vitro and in vivo systems for producing bioactive resveratrol-O-glucoside or resveratrol poly-glucosides depends on available facilities and specific production requirements.
Aging is a natural biological process that poses a threat to living organisms by making them susceptible to various diseases and disorders. The skin is the largest organ of the body and the most exposed. It is highly vulnerable to aging, resulting in cancer, skin damage, and other harmful conditions [47]. Resveratrol and related compounds have been found to exhibit significant effects against skin aging by inhibiting tyrosinase, an enzyme involved in melanin production. Melanin regulates vitamin D3 biosynthesis and enhances the skin’s resistance to tumors and sunburns [48,49]. Therefore, tyrosinase inhibitors such as resveratrol and its glucoside derivatives are highly sought-after in medicinal and cosmetic industries as depigmentation agents [50].
During melanin biosynthesis, intervention of L-cysteine can lead to the formation of pheomelanin, which can be cytotoxic as it produces reactive oxygen species (ROS) [51,52]. Hence, controlling excess melanin and intercellular ROS is a crucial biochemical step in developing a safe and effective skin-whitening agent [53]. Over ten skin-whitening substances such as ascorbic acid, kojic acid, and arbutin have been used as cosmetic ingredients [53]. Reports have suggested that resveratrol and its glucosylated derivatives possess antioxidant and free radical scavenging capabilities as they can promote activities of various antioxidative enzymes [54]. The presence of phenolic hydroxyl groups and electron delocalization across the molecules is the primary reason for their antioxidant properties [55].
Secretion of a biological response modifier in response to external or internal stimuli can result in allergies and inflammation in the body. Macrophages are the primary source of cytokines and growth factors that can affect epithelial, endothelial, and mesenchymal cells, leading to inflammation [56]. Under normal physiological conditions, a moderate increase in inducible NO synthase (NOS) activity leads to NO formation, which has bactericidal effects. However, high levels of NO and its derivatives such as peroxynitrite (ONOO-) can cause inflammation. Resveratrol preincubation with Raw 264.7 cells decreased inflammation by downregulating iNOS mRNA and protein [57,58]. Due to its high thermostability and specific gravity among all known ASases, D. geothermalis ASase (DgAS) is highly important in the industry through its transglucosylation activity among different acceptor substances [41]. Such variation in transglucosylation activity might be due to changes in active site conformation caused by intramolecular interactions including hydrogen bonds, disulfide bonds, and ionic interactions [41,46]. Although many flavone materials are available as potential pharmaceutical ca ndidates,their application is restricted because of their poor solubility. Thus, the main aim of this study was to synthesize flavone glycosides using DgAS on an industrial scale [59,60].
5. Conclusion
In summary, enzymatic production of resveratrol glucosides using DgAS in both in vitro and in vivo systems is highly effective, with a conversion rate of approximately 97.0% in a short period. DgAS can add glucose to multiple hydroxyl positions and conjugate glucose in the chain, resulting in highly soluble glucosylated products. Furthermore, positive outcomes obtained from in vitro assays using Raw 264.7, HS68, and B16 cell lines highlight the potential application of resveratrol glucosides in the cosmetics industry. Therefore, this system holds promise for cost-effective industrial production of resveratrol glucosides.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
References
- Pandey, R.P.; Parajuli, P.; Shin, J.Y.; Lee, J.; Lee, S.; Hong, Y.S.; Park, Y. Il; Kim, J.S.; Sohng, J.K. Enzymatic biosynthesis of novel resveratrol glucoside and glycoside derivatives. Appl. Environ. Microbiol. 2014, 80, 7235–7243. https://doi.org/10.1128/AEM.02076-14. [CrossRef]
- Yang, Y.; Lin, Y.; Li, L.; Linhardt, R.J.; Yan, Y. Regulating malonyl-CoA metabolism via synthetic antisense RNAs for enhanced biosynthesis of natural products. Metab. Eng. 2015, 29, 217–226. https://doi.org/10.1016/j.ymben.2015.03.018. [CrossRef]
- Gambini, J.; Inglés, M.; Olaso, G.; Lopez-Grueso, R.; Bonet-Costa, V.; Gimeno-Mallench, L.; Mas-Bargues, C.; Abdelaziz, K.M.; Gomez-Cabrera, M.C.; Vina, J.; et al. Properties of Resveratrol: In Vitro and In Vivo Studies about Metabolism, Bioavailability, and Biological Effects in Animal Models and Humans. Oxid. Med. Cell. Longev. 2015, 2015. https://doi.org/10.1155/2015/837042. [CrossRef]
- Pangeni, R.; Sahni, J.K.; Ali, J.; Sharma, S.; Baboota, S. Resveratrol: Review on therapeutic potential and recent advances in drug delivery. Expert Opin. Drug Deliv. 2014, 11, 1285–1298. https://doi.org/10.1517/17425247.2014.919253. [CrossRef]
- Park, S.-J.; Ahmad, F.; Philp, A.; Baar, K.; Williams, T.; Luo, H.; Ke, H.; Rehmann, H.; Taussig, R.; Brown, A.L.; et al. Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phosphodiesterases. BMC Proc. 2012, 6, 6561. https://doi.org/10.1186/1753-6561-6-s3-p73. [CrossRef]
- Bai, X.; Yu, W.; Ji, W.; Lin, Z.; Tan, S.; Duan, K.; Dong, Y.; Xu, L.; Li, N. Early versus delayed administration of norepinephrine in patients with septic shock. Crit. Care 2014, 18, 1–8. https://doi.org/10.1186/s13054-014-0532-y. [CrossRef]
- Petrovski, G.; Gurusamy, N.; Das, D.K. Resveratrol in cardiovascular health and disease. Ann. N. Y. Acad. Sci. 2011, 1215, 22–33. https://doi.org/10.1111/j.1749-6632.2010.05843.x. [CrossRef]
- Bhatt, S.R.; Lokhandwala, M.F.; Banday, A.A. Resveratrol prevents endothelial nitric oxide synthase uncoupling and attenuates development of hypertension in spontaneously hypertensive rats. Eur. J. Pharmacol. 2011, 667, 258–264. https://doi.org/10.1016/j.ejphar.2011.05.026. [CrossRef]
- Ko, K.P. Isoflavones: Chemistry, analysis, functions and effects on health and cancer. Asian Pacific J. Cancer Prev. 2014, 15, 7001–7010. https://doi.org/10.7314/APJCP.2014.15.17.7001. [CrossRef]
- Zaheer, K.; Humayoun Akhtar, M. An updated review of dietary isoflavones: Nutrition, processing, bioavailability and impacts on human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 1280–1293. https://doi.org/10.1080/10408398.2014.989958. [CrossRef]
- Szeja, W. la.; Grynkiewicz, G.; Rusin, A. Isoflavones, their Glycosides and Glycoconjugates. Synthesis and Biological Activity. Curr. Org. Chem. 2016, 21, 218–235. https://doi.org/10.2174/1385272820666160928120822. [CrossRef]
- Tian, B.; Liu, J. Resveratrol: a review of plant sources, synthesis, stability, modification and food application. J. Sci. Food Agric. 2020, 100, 1392–1404. https://doi.org/10.1002/jsfa.10152. [CrossRef]
- Zupančič, Š.; Lavrič, Z.; Kristl, J. Stability and solubility of trans-resveratrol are strongly influenced by pH and temperature. Eur. J. Pharm. Biopharm. 2015, 93, 196–204. https://doi.org/10.1016/j.ejpb.2015.04.002. [CrossRef]
- Huang, X.; Liu, Y.; Zou, Y.; Liang, X.; Peng, Y.; McClements, D.J.; Hu, K. Encapsulation of resveratrol in zein/pectin core-shell nanoparticles: Stability, bioaccessibility, and antioxidant capacity after simulated gastrointestinal digestion. Food Hydrocoll. 2019, 93, 261–269. https://doi.org/10.1016/j.foodhyd.2019.02.039. [CrossRef]
- Liu, Y.; Fan, Y.; Gao, L.; Zhang, Y.; Yi, J. Enhanced pH and thermal stability, solubility and antioxidant activity of resveratrol by nanocomplexation with α-lactalbumin; 2018; Vol. 9; ISBN 1021382620.
- Zhang, F.; Khan, M.A.; Cheng, H.; Liang, L. Co-encapsulation of α-tocopherol and resveratrol within zein nanoparticles: Impact on antioxidant activity and stability. J. Food Eng. 2019, 247, 9–18. https://doi.org/10.1016/j.jfoodeng.2018.11.021. [CrossRef]
- Caddeo, C.; Pucci, L.; Gabriele, M.; Carbone, C.; Fernàndez-Busquets, X.; Valenti, D.; Pons, R.; Vassallo, A.; Fadda, A.M.; Manconi, M. Stability, biocompatibility and antioxidant activity of PEG-modified liposomes containing resveratrol. Int. J. Pharm. 2018, 538, 40–47. https://doi.org/10.1016/j.ijpharm.2017.12.047. [CrossRef]
- Ravetti, S.; Clemente, C.; Brignone, S.; Hergert, L.; Allemandi, D.; Palma, S. Ascorbic acid in skin health. Cosmetics 2019, 6, 6–13. https://doi.org/10.3390/COSMETICS6040058. [CrossRef]
- Boo, Y.C. Ascorbic Acid (Vitamin C) as a Cosmeceutical to Increase Dermal Collagen for Skin Antiaging Purposes: Emerging Combination Therapies. Antioxidants 2022, 11. https://doi.org/10.3390/antiox11091663. [CrossRef]
- Gull, M.; Pasek, M.A. The role of glycerol and its derivatives in the biochemistry of living organisms, and their prebiotic origin and significance in the evolution of life. Catalysts 2021, 11, 1–32. https://doi.org/10.3390/catal11010086. [CrossRef]
- Kruschitz, A.; Nidetzky, B. Biocatalytic Production of 2-α- d -Glucosyl-glycerol for Functional Ingredient Use: Integrated Process Design and Techno-Economic Assessment. ACS Sustain. Chem. Eng. 2022, 10, 1246–1255. https://doi.org/10.1021/acssuschemeng.1c07210. [CrossRef]
- Khan, A.W.; Kotta, S.; Ansari, S.H.; Sharma, R.K.; Ali, J. Enhanced dissolution and bioavailability of grapefruit flavonoid Naringenin by solid dispersion utilizing fourth generation carrier. Drug Dev. Ind. Pharm. 2015, 41, 772–779. https://doi.org/10.3109/03639045.2014.902466. [CrossRef]
- Xie, L.; Zhang, L.; Wang, C.; Wang, X.; Xu, Y. ming; Yu, H.; Wu, P.; Li, S.; Han, L.; Gunatilaka, A.A.L.; et al. Methylglucosylation of aromatic amino and phenolic moieties of drug-like biosynthons by combinatorial biosynthesis. Proc. Natl. Acad. Sci. U. S. A. 2018, 115, E4980–E4989. https://doi.org/10.1073/pnas.1716046115. [CrossRef]
- Kim, B.G.; Yang, S.M.; Kim, S.Y.; Cha, M.N.; Ahn, J.H. Biosynthesis and production of glycosylated flavonoids in Escherichia coli: current state and perspectives. Appl. Microbiol. Biotechnol. 2015, 99, 2979–2988. https://doi.org/10.1007/s00253-015-6504-6. [CrossRef]
- Chouhan, S.; Sharma, K.; Zha, J.; Guleria, S.; Koffas, M.A.G. Recent advances in the recombinant biosynthesis of polyphenols. Front. Microbiol. 2017, 8, 1–16. https://doi.org/10.3389/fmicb.2017.02259. [CrossRef]
- Kim, B.; Park, H.; Na, D.; Lee, S.Y. Metabolic engineering of Escherichia coli for the production of phenol from glucose. Biotechnol.J.2013, 9(5), 621-629.
- Shomar, H.; Gontier, S.; Van Den Broek, N.J.F.; Tejeda Mora, H.; Noga, M.J.; Hagedoorn, P.L.; Bokinsky, G. Metabolic engineering of a carbapenem antibiotic synthesis pathway in Escherichia coli. Nat. Chem. Biol. 2018, 14, 794–800. https://doi.org/10.1038/s41589-018-0084-6. [CrossRef]
- Kim, K.-T.; Rha, C.-S.; Jung, Y.S.; Kim, Y.-J.; Jung, D.-H.; Seo, D.-H.; Park, C.-S. Comparative study on amylosucrases derived from Deinococcus species and catalytic characterization and use of amylosucrase derived from Deinococcus wulumuqiensis. Amylase 2019, 3, 19–31. https://doi.org/10.1515/amylase-2019-0002. [CrossRef]
- Jung, Y.S.; Kim, Y.; Kim, A.T.; Jang, D.; Kim, M.S.; Seo, D.H.; Nam, T.G.; Rha, C.S.; Park, C.S; Kim, D.O. Enrichment of Polyglucosylated Isoflavones from soybean isoflavone aglycones using optimized amylosucrase transglycosylation. Molecules, 2020, 25(1), 181. https://doi.org/10.3390/molecules25010181. [CrossRef]
- Lee, H.S.; Kim, T.S.; Parajuli, P.; Pandey, R.P.; Sohng, J.K. Sustainable production of dihydroxybenzene glucosides using immobilized amylosucrase from Deinococcus geothermalis. J. Microbiol. Biotechnol. 2018, 28, 1447–1456. https://doi.org/10.4014/jmb.1805.05054. [CrossRef]
- Xu, L.; Qi, T.; Xu, L.; Lu, L.; Xiao, M. Recent progress in the enzymatic glycosylation of phenolic compounds. J. Carbohydr. Chem. 2016, 35, 1–23. https://doi.org/10.1080/07328303.2015.1137580. [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. ScientificWorldJournal. 2013, 2013, 162750. https://doi.org/10.1155/2013/162750. [CrossRef]
- Mei, Y.Z.; Liu, R.X.; Wang, D.P.; Wang, X.; Dai, C.C. Biocatalysis and biotransformation of resveratrol in microorganisms. Biotechnol. Lett. 2015, 37, 9–18. https://doi.org/10.1007/s10529-014-1651-x. [CrossRef]
- Wu, J.; Liu, P.; Fan, Y.; Bao, H.; Du, G.; Zhou, J.; Chen, J. Multivariate modular metabolic engineering of Escherichia coli to produce resveratrol from l-tyrosine. J. Biotechnol. 2013, 167, 404–411. https://doi.org/10.1016/j.jbiotec.2013.07.030. [CrossRef]
- Li, M.; Schneider, K.; Kristensen, M.; Borodina, I.; Nielsen, J. Engineering yeast for high-level production of stilbenoid antioxidants. Sci. Rep. 2016, 6, 1–8. https://doi.org/10.1038/srep36827. [CrossRef]
- Choi, O.; Wu, C.Z.; Kang, S.Y.; Ahn, J.S.; Uhm, T.B.; Hong, Y.S. Biosynthesis of plant-speciWc phenylpropanoids by construction of an artificial biosynthetic pathway in Escherichia coli. J. Ind. Microbiol. Biotechnol. 2011, 38, 1657–1665. https://doi.org/10.1007/s10295-011-0954-3. [CrossRef]
- Shrestha, A.; Pandey, R.P.; Sohng, J.K. Biosynthesis of resveratrol and piceatannol in engineered microbial strains: achievements and perspectives. Appl. Microbiol. Biotechnol. 2019, 103, 2959–2972. https://doi.org/10.1007/s00253-019-09672-8. [CrossRef]
- Fauconneau, B.; Waffo-teguop, P.; Huguet, F.; Barrier, L.; Decendit, A.; Merillon, J.M. Comparative study of radical scavenger and antioxidant properties of phenolic compounds from Vitis vinifera cell cultures using in vitro tests. Life Sci. 1997, 61, 2103–2110.
- Choi, O.; Lee, J.K.; Kang, S.Y.; Pandey, R.P.; Sohng, J.K.; Ahn, J.S.; Hong, Y.S. Construction of artificial biosynthetic pathways for resveratrol glucoside derivatives. J. Microbiol. Biotechnol. 2014, 24, 614–618. https://doi.org/10.4014/jmb.1401.01031. [CrossRef]
- Moulis, C.; André, I.; Remaud-Simeon, M. GH13 amylosucrases and GH70 branching sucrases, atypical enzymes in their respective families. Cell. Mol. Life Sci. 2016, 73, 2661–2679. https://doi.org/10.1007/s00018-016-2244-8. [CrossRef]
- Seo, D.H.; Jung, J.H.; Jung, D.H.; Park, S.; Yoo, S.H.; Kim, Y.R.; Park, C.S. An unusual chimeric amylosucrase generated by domain-swapping mutagenesis. Enzyme Microb. Technol. 2016, 86, 7–16. https://doi.org/10.1016/j.enzmictec.2016.01.004. [CrossRef]
- Rha, C.S.; Jung, Y.S.; Seo, D.H.; Kim, D.O.; Park, C.S. Site-specific α-glycosylation of hydroxyflavones and hydroxyflavanones by amylosucrase from Deinococcus geothermalis. Enzyme Microb. Technol. 2019, 129, 109361. https://doi.org/10.1016/j.enzmictec.2019.109361. [CrossRef]
- Ha, S.J.; Seo, D.H.; Jung, J.H.; Cha, J.; Kim, T.J.; Kim, Y.W.; Park, C.S. Molecular cloning and functional expression of a new amylosucrase from Alteromonas macleodii. Biosci. Biotechnol. Biochem. 2009, 73, 1505–1512. https://doi.org/10.1271/bbb.80891. [CrossRef]
- Wang, J.; Yang, Y.; Yan, Y. Bioproduction of resveratrol. Biotechnol. Nat. Prod. 2017, 61–79. https://doi.org/10.1007/978-3-319-67903-7_3. [CrossRef]
- Jung, J.H.; Seo, D.H.; Ha, S.J.; Song, M.C.; Cha, J.; Yoo, S.H.; Kim, T.J.; Baek, N.I.; Baik, M.Y.; Park, C.S. Enzymatic synthesis of salicin glycosides through transglycosylation catalyzed by amylosucrases from Deinococcus geothermalis and Neisseria polysaccharea. Carbohydr. Res. 2009, 344, 1612–1619. https://doi.org/10.1016/j.carres.2009.04.019. [CrossRef]
- Seo, D.H.; Jung, J.H.; Ha, S.J.; Song, M.C.; Cha, J.; Yoo, S.H.; Kim, T.J.; Baek, N.I.; Park, C.S. Highly selective biotransformation of arbutin to arbutin-α-glucoside using amylosucrase from Deinococcus geothermalis DSM 11300. J. Mol. Catal. B Enzym. 2009, 60, 113–118. https://doi.org/10.1016/j.molcatb.2009.04.006. [CrossRef]
- Abbas, H.; Kamel, R.; El-Sayed, N. Dermal anti-oxidant, anti-inflammatory and anti-aging effects of Compritol ATO-based Resveratrol colloidal carriers prepared using mixed surfactants. Int. J. Pharm. 2018, 541, 37–47. https://doi.org/10.1016/j.ijpharm.2018.01.054. [CrossRef]
- Honisch, C.; Osto, A.; Dupas de Matos, A.; Vincenzi, S.; Ruzza, P. Isolation of a tyrosinase inhibitor from unripe grapes juice: A spectrophotometric study. Food Chem. 2020, 305, 125506. https://doi.org/10.1016/j.foodchem.2019.125506. [CrossRef]
- Skoczynska, A., Budzisz, E., Trznadel-Grodzka, E., and Rotsztehn, H. Melanin and Lipofuscin. Adv. dermatology Allergol. 2017, 34 (2), 1.
- Zolghadri, S.; Bahrami, A.; Hassan Khan, M.T.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2019, 34, 279–309. https://doi.org/10.1080/14756366.2018.1545767. [CrossRef]
- Napolitano, A.; Panzella, L.; Monfrecola, G.; d’Ischia, M. Pheomelanin-induced oxidative stress: Bright and dark chemistry bridging red hair phenotype and melanoma. Pigment Cell Melanoma Res. 2014, 27, 721–733. https://doi.org/10.1111/pcmr.12262. [CrossRef]
- Smit, N.P.M.; Van Nieuwpoort, F.A.; Marrot, L.; Out, C.; Poorthuis, B.; Van Pelt, H.; Meunier, J.R.; Pavel, S. Increased melanogenesis is a risk factor for oxidative DNA damage - Study on cultured melanocytes and atypical nevus cells. Photochem. Photobiol. 2008, 84, 550–555. https://doi.org/10.1111/j.1751-1097.2007.00242.x. [CrossRef]
- Okura, M.; Yamashita, T.; Ishii-Osai, Y.; Yoshikawa, M.; Sumikawa, Y.; Wakamatsu, K.; Ito, S. Effects of rhododendrol and its metabolic products on melanocytic cell growth. J. Dermatol. Sci. 2015, 80, 142–149. https://doi.org/10.1016/j.jdermsci.2015.07.010. [CrossRef]
- De La Lastra, C.A.; Villegas, I. Resveratrol as an anti-inflammatory and anti-aging agent: Mechanisms and clinical implications. Mol. Nutr. Food Res. 2005, 49, 405–430. https://doi.org/10.1002/mnfr.200500022. [CrossRef]
- López-Vélez, M.; Martínez-Martínez, F.; Valle-Ribes, C. Del The Study of Phenolic Compounds as Natural Antioxidants in Wine. Crit. Rev. Food Sci. Nutr. 2003, 43, 233–244. https://doi.org/10.1080/10408690390826509. [CrossRef]
- Fritsch, C.; Simon-Assmann, P.; Kedinger, M.; Evans, G.S. Cytokines modulate fibroblast phenotype and epithelial-stroma interactions in rat intestine. Gastroenterology 1997, 112, 826–838. https://doi.org/10.1053/gast.1997.v112.pm9041244. [CrossRef]
- Tsai, S.H.; Lin-Shiau, S.Y.; Lin, J.K. Suppression of nitric oxide synthase and the down-regulation of the activation of NFκB in macrophages by resveratrol. Br. J. Pharmacol. 1999, 126, 673–680. https://doi.org/10.1038/sj.bjp.0702357. [CrossRef]
- Wadsworth, T.L.; Koop, D.R. Effects of the wine polyphenolics quercetin and resveratrol on pro-inflammatory cytokine expression in RAW 264.7 macrophages. Biochem. Pharmacol. 1999, 57, 941–949. https://doi.org/10.1016/S0006-2952(99)00002-7. [CrossRef]
- Tommasini, S.; Raneri, D.; Ficarra, R.; Calabrò, M.L.; Stancanelli, R.; Ficarra, P. Improvement in solubility and dissolution rate of flavonoids by complexation with β-cyclodextrin. J. Pharm. Biomed. Anal. 2004, 35, 379–387. https://doi.org/10.1016/S0731-7085(03)00647-2. [CrossRef]
- Moon, K.; Lee, S.; Park, H.; Cha, J. Enzymatic Synthesis of Resveratrol α-Glucoside by Amylosucrase of Deinococcus geothermalis. J. Microbiol. Biotechnol. 2021, 31, 1692–1700. https://doi.org/10.4014/jmb.2108.08034. [CrossRef]
Figure 1.
Representative reaction scheme of resveratrol glucosides formation by the enzyme DgAS, Amylosucrase.
Figure 1.
Representative reaction scheme of resveratrol glucosides formation by the enzyme DgAS, Amylosucrase.
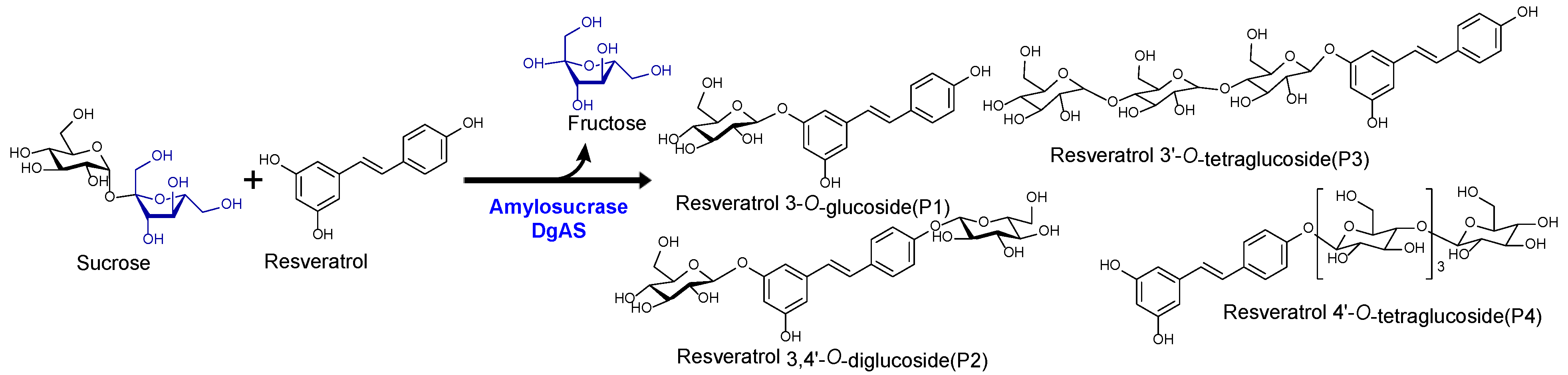
Figure 2.
(A) HPLC analysis of resveratrol reaction mixture. (i) Control reaction for regular glucosylation reaction of DgAS; (ii) Standard resveratrol; and (iii) Regular glucosylation reaction carried out using DgAS. (B) UV-VIS analysis of major each peak from HPLC (i) Standard Resveratrol, (ii) P1, (iii) P2, (iv) P3,and (v) P4. (C) HRQTOF-ESI/MS analysis of (i) Resveratrol standard, (ii) Product P1, (iii) Product P2, (iv) product P3,and (v) Product P4.
Figure 2.
(A) HPLC analysis of resveratrol reaction mixture. (i) Control reaction for regular glucosylation reaction of DgAS; (ii) Standard resveratrol; and (iii) Regular glucosylation reaction carried out using DgAS. (B) UV-VIS analysis of major each peak from HPLC (i) Standard Resveratrol, (ii) P1, (iii) P2, (iv) P3,and (v) P4. (C) HRQTOF-ESI/MS analysis of (i) Resveratrol standard, (ii) Product P1, (iii) Product P2, (iv) product P3,and (v) Product P4.
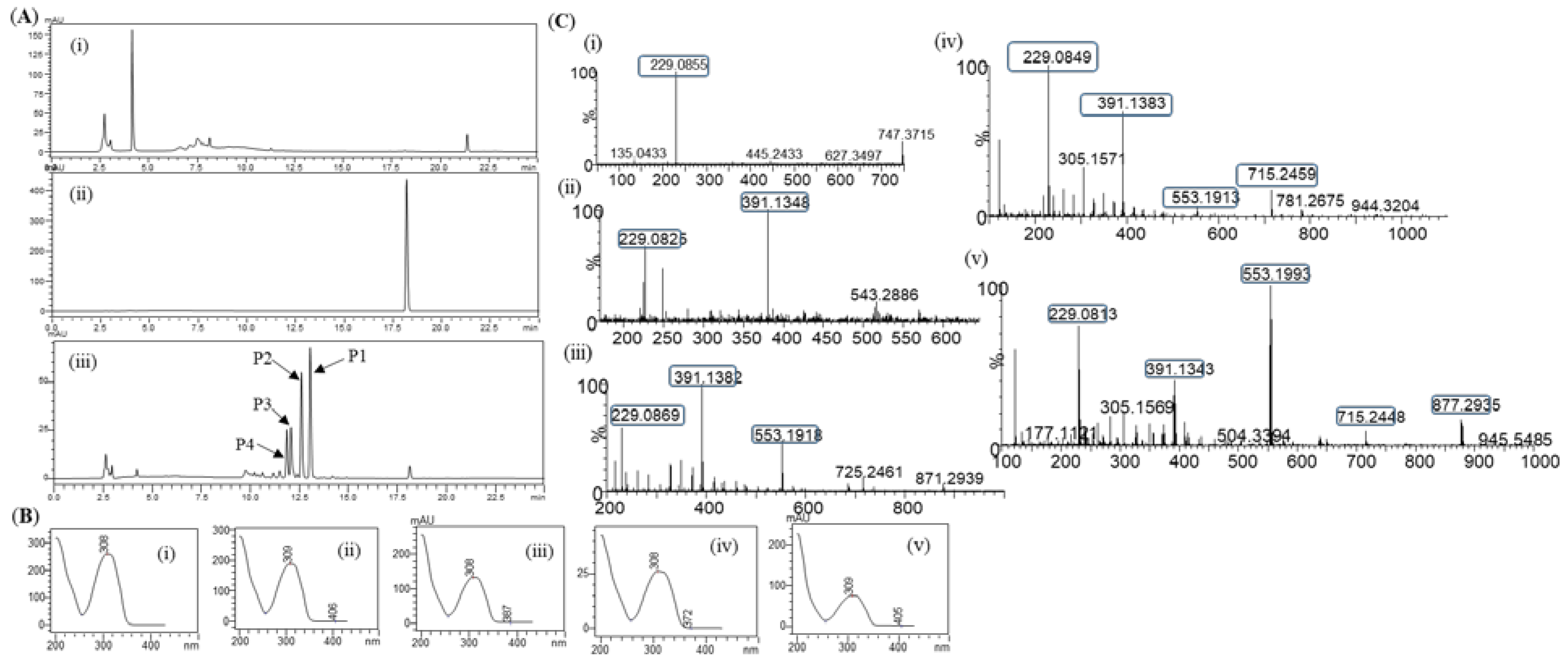
Figure 3.
Conversion percentage of resveratrol to resveratrol glucosides in the optimization cascade reaction;- (i) Variation of sucrose concentration, (ii) Different enzymes concentration of DgAS, (iii) Variation reaction temperature, and (iv) Variation of buffer and pH.
Figure 3.
Conversion percentage of resveratrol to resveratrol glucosides in the optimization cascade reaction;- (i) Variation of sucrose concentration, (ii) Different enzymes concentration of DgAS, (iii) Variation reaction temperature, and (iv) Variation of buffer and pH.
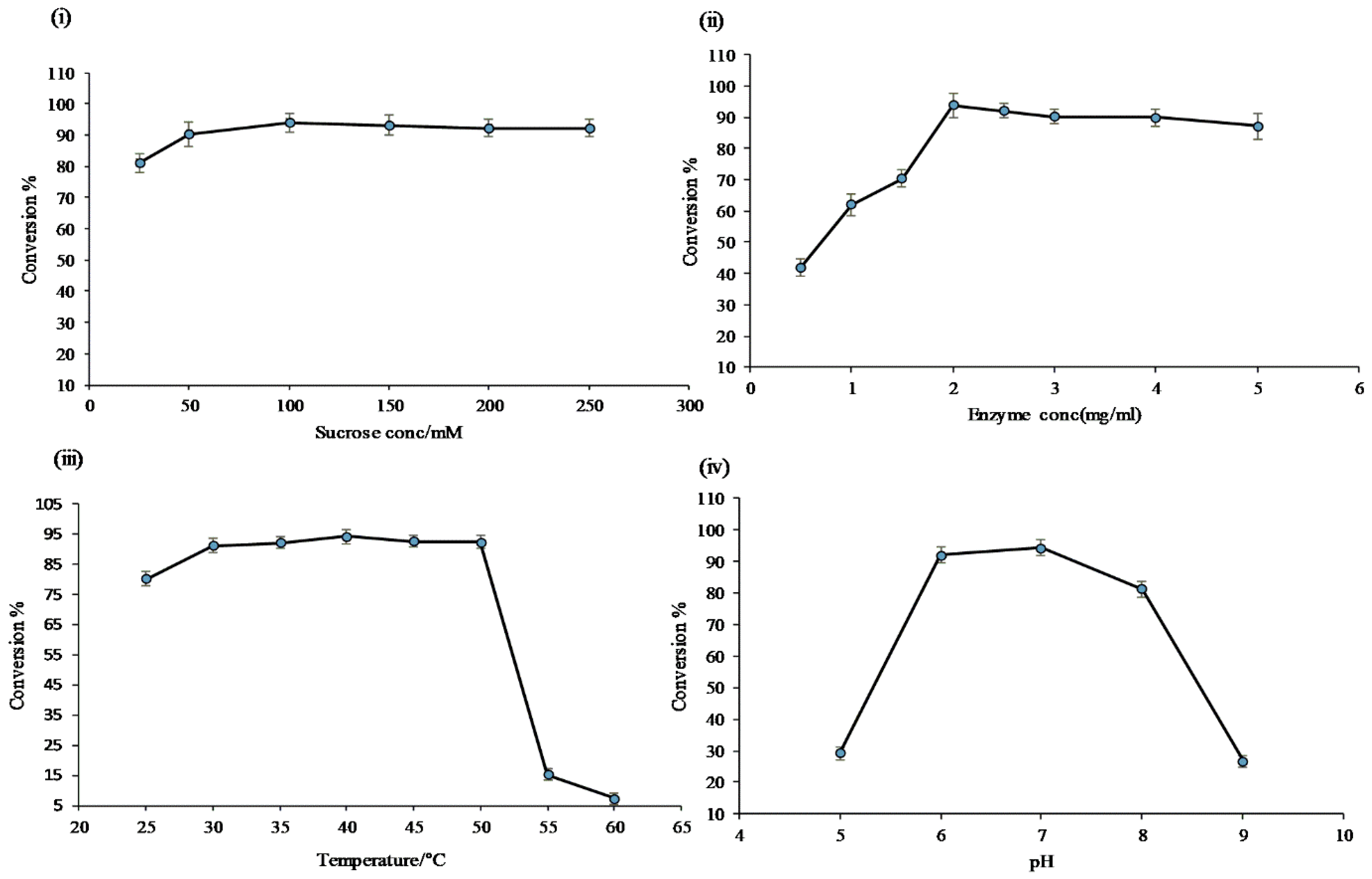
Figure 4.
Conversion of resveratrol to its glucosides in preparative scale in vitro reaction at different times.
Figure 4.
Conversion of resveratrol to its glucosides in preparative scale in vitro reaction at different times.
Figure 5.
(A) HPLC-PDA chromatogram for resveratrol and resveratrol glucosides in vivo system; (i) Standard resveratrol; (ii) Control; (iii) 10h sample; and (iv) 35h sample. (B) HR-QTOF ESI/MS analysis of (i) Resveratrol standard and (ii) Resveratrol glucoside at 10h sample, P1, (iii) P2, (iv) P3, and (iv) P4. (C) UV-VIS analysis of (i) Standard resveratrol, (ii) Product, P1, (iii) P2, (iv) P3, and (iv) P4.
Figure 5.
(A) HPLC-PDA chromatogram for resveratrol and resveratrol glucosides in vivo system; (i) Standard resveratrol; (ii) Control; (iii) 10h sample; and (iv) 35h sample. (B) HR-QTOF ESI/MS analysis of (i) Resveratrol standard and (ii) Resveratrol glucoside at 10h sample, P1, (iii) P2, (iv) P3, and (iv) P4. (C) UV-VIS analysis of (i) Standard resveratrol, (ii) Product, P1, (iii) P2, (iv) P3, and (iv) P4.
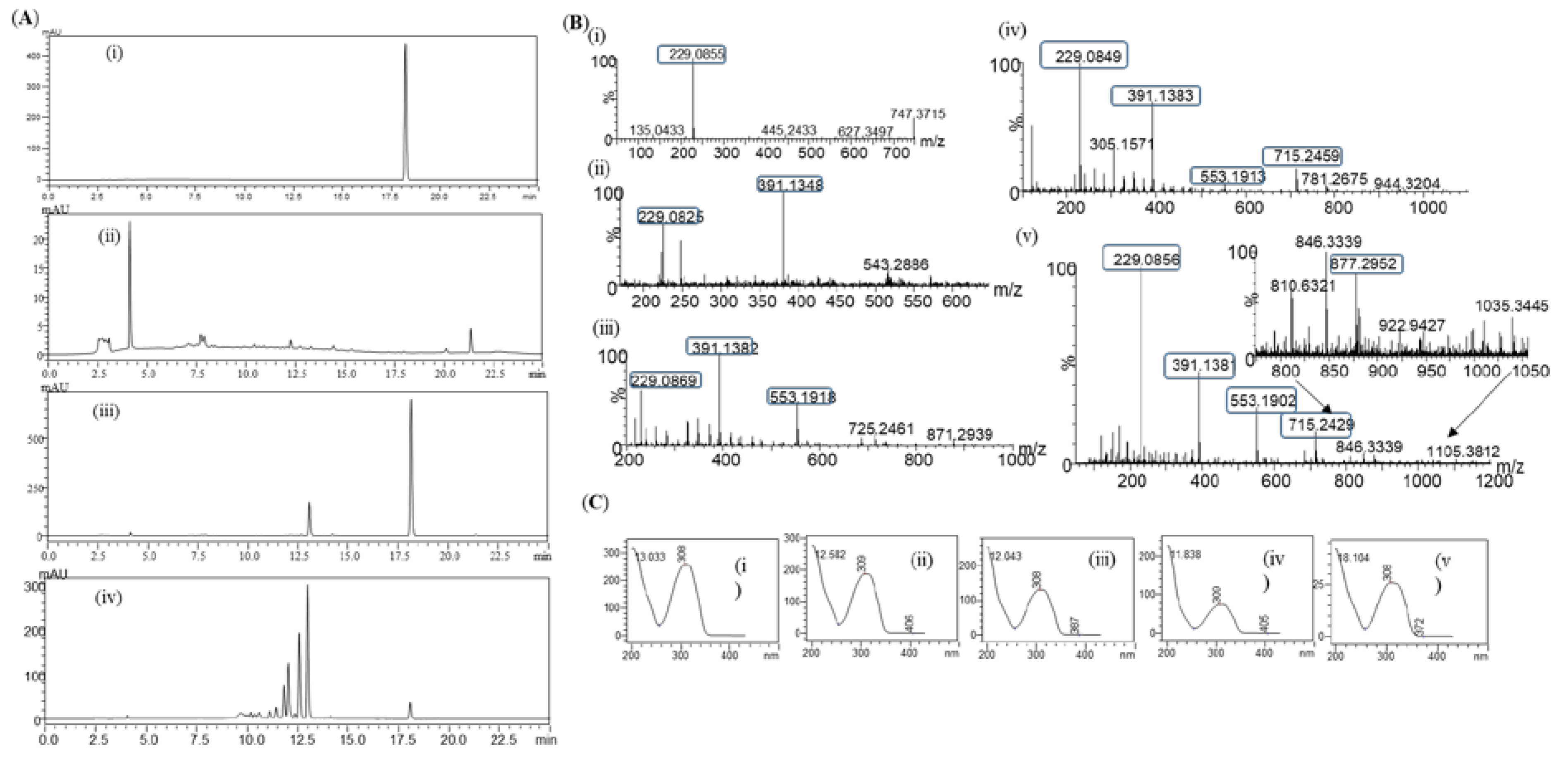
Figure 6.
Cytotoxicity evaluation of raw resveratrol and resveratrol-O-glucoside. (A) Experimental conditions; B16/ 8 ⅹ 103 cells/ 96 well plate 24h incubation period after sample treatment. (B) Experimental conditions; Raw 264.7/ 1.0 ⅹ 104 cells/ 96 well plate incubation period after sample treatment (48h). (C) Experimental conditions: HS68 cells/ 0.5 ⅹ 104 cells/ 96 well plate. Incubation period after sample treatment (48h). Note-Used media containing 5% ATCC FBS for cell culture. Significance is indicated only when there is toxicity relative to CTL ***; p < 0.001 / **; p < 0.01.
Figure 6.
Cytotoxicity evaluation of raw resveratrol and resveratrol-O-glucoside. (A) Experimental conditions; B16/ 8 ⅹ 103 cells/ 96 well plate 24h incubation period after sample treatment. (B) Experimental conditions; Raw 264.7/ 1.0 ⅹ 104 cells/ 96 well plate incubation period after sample treatment (48h). (C) Experimental conditions: HS68 cells/ 0.5 ⅹ 104 cells/ 96 well plate. Incubation period after sample treatment (48h). Note-Used media containing 5% ATCC FBS for cell culture. Significance is indicated only when there is toxicity relative to CTL ***; p < 0.001 / **; p < 0.01.
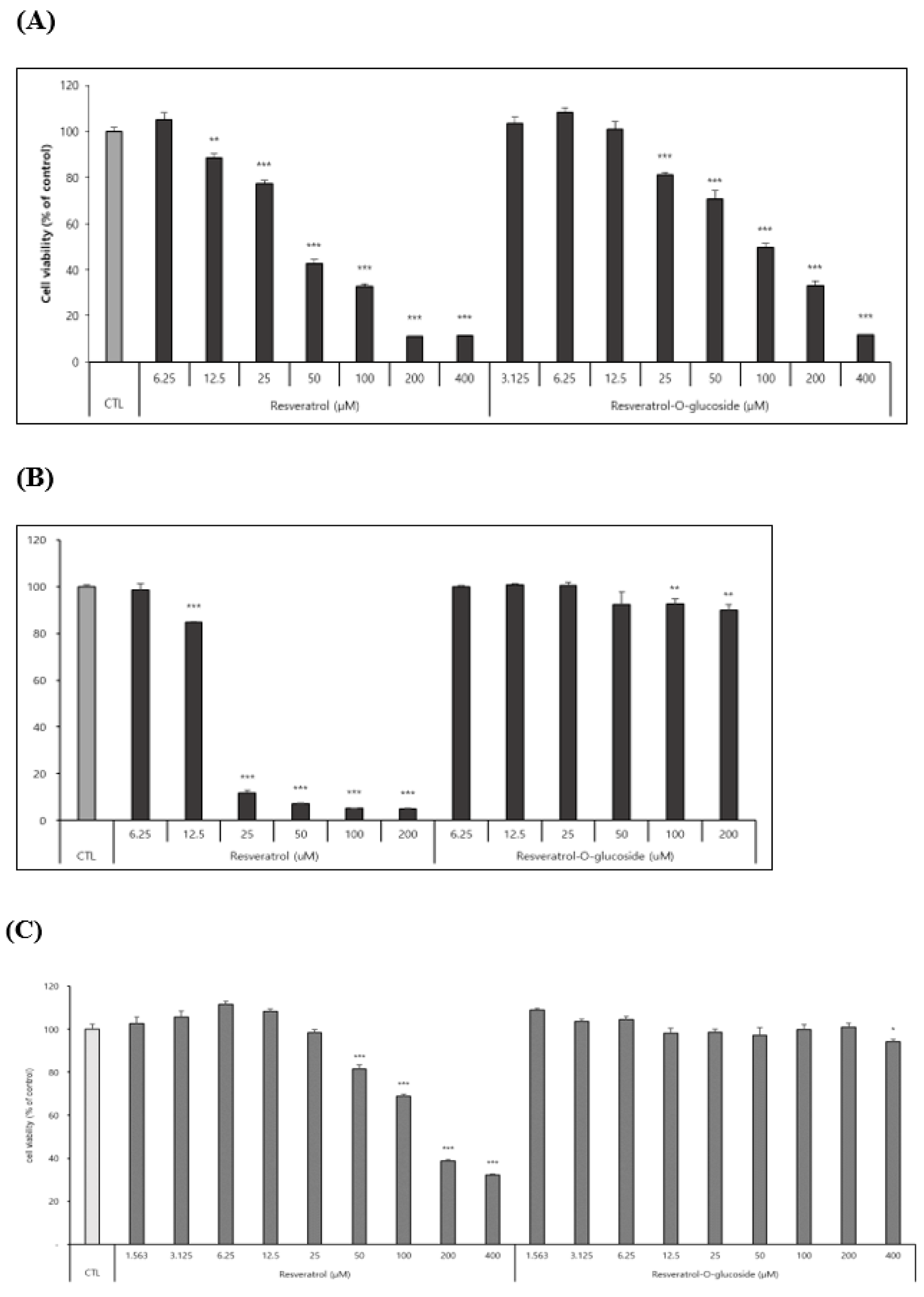
Figure 7.
Anti-inflammation and anti-aging test of resveratrol and resveratrol-O-glucosides. (A) Experimental condition: raw 264.7 /1.0ⅹ 103 cells/ 96 well plate. Incubation period after sample treatment (48h). (B) Experimental condition: HS68 /3.0ⅹ 104 cells/24 well plate. Incubation period after sample treatment (48h). Note: For cell culture, use triple FBS (10%)/ For drug treatment, use triple FBS (2%). Significance is indicated only when there is efficacy over CTL. *** or ###; p < 0.001 / ** or ##; p < 0.01 / * or #; p < 0.5.TGF-β is used as a positive control for procollagen production.
Figure 7.
Anti-inflammation and anti-aging test of resveratrol and resveratrol-O-glucosides. (A) Experimental condition: raw 264.7 /1.0ⅹ 103 cells/ 96 well plate. Incubation period after sample treatment (48h). (B) Experimental condition: HS68 /3.0ⅹ 104 cells/24 well plate. Incubation period after sample treatment (48h). Note: For cell culture, use triple FBS (10%)/ For drug treatment, use triple FBS (2%). Significance is indicated only when there is efficacy over CTL. *** or ###; p < 0.001 / ** or ##; p < 0.01 / * or #; p < 0.5.TGF-β is used as a positive control for procollagen production.
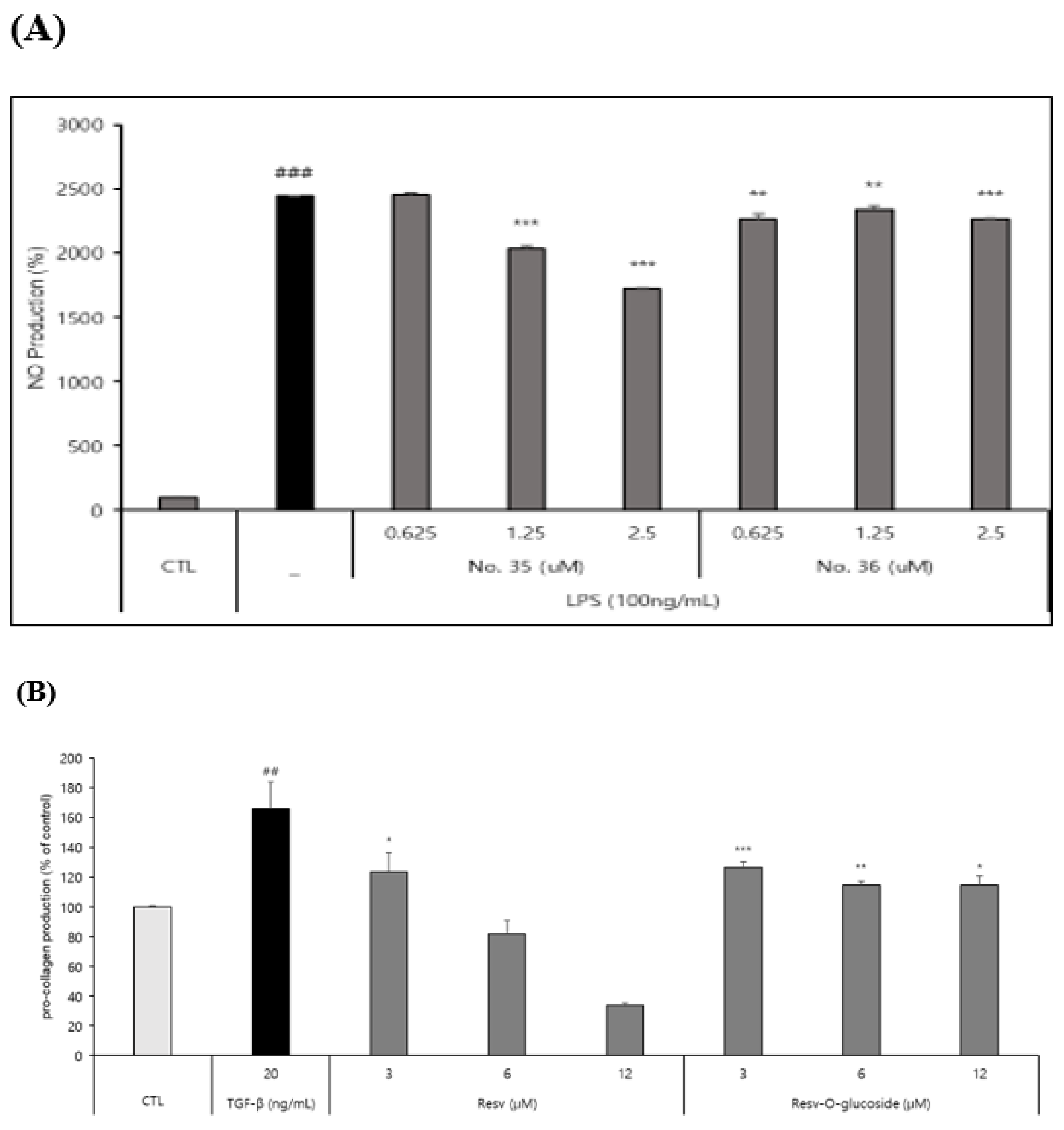
Figure 8.
Whitening and antioxidant test of raw resveratrol and resveratrol-O-glucoside. (A) (i) Extracellular melanin contents (%) and (ii) Intracellular melanin contents (%). Experimental conditions; B16/ 25 ⅹ 103 cells/ 48 well plate/ 72h incubation period after sample treatment. Note: Use media containing 5% ATCC FBS for cell culture. The stimulus is α-MSH and the positive control is Kojic acid (a whitening functional ingredient). Inhibition of melanogenesis was calculated as CTL (100 %, α-MSH alone (0 %).Significance is indicated only when there is whitening effect. ***; p < 0.001 / **; p < 0.01. (B) Experimental conditions: DPPH test B16/ 8ⅹ 103 cells/ 96 well plate. Incubation 15 mins after sample treatment. Note: The experimental concentrations were specified considering non-toxic concentrations in both Raw 262.47, HS68, and B16 cells. Significance is shown only when there is an antioxidant advantage over CTL ***; p < 0.001 / **; p < 0.01.
Figure 8.
Whitening and antioxidant test of raw resveratrol and resveratrol-O-glucoside. (A) (i) Extracellular melanin contents (%) and (ii) Intracellular melanin contents (%). Experimental conditions; B16/ 25 ⅹ 103 cells/ 48 well plate/ 72h incubation period after sample treatment. Note: Use media containing 5% ATCC FBS for cell culture. The stimulus is α-MSH and the positive control is Kojic acid (a whitening functional ingredient). Inhibition of melanogenesis was calculated as CTL (100 %, α-MSH alone (0 %).Significance is indicated only when there is whitening effect. ***; p < 0.001 / **; p < 0.01. (B) Experimental conditions: DPPH test B16/ 8ⅹ 103 cells/ 96 well plate. Incubation 15 mins after sample treatment. Note: The experimental concentrations were specified considering non-toxic concentrations in both Raw 262.47, HS68, and B16 cells. Significance is shown only when there is an antioxidant advantage over CTL ***; p < 0.001 / **; p < 0.01.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



