
Submitted:
23 May 2023
Posted:
24 May 2023
You are already at the latest version
Abstract
The name “Spergularia hanoverensis Simon” has been misapplied for an endemic taxon confined to inland semi-desert ecosystems in central-western South Africa. It is commonly accepted to be a small annual species occurring in saline habitats in a wide elevation range, but its real identity still remains obscure. In the context of a taxonomic and phylogenetic research on the African species of Spergularia, we found that the name apparently was never validly published still. After revision of herbarium material housed in the South African herbaria, a voucher collected from Hanover was found at PRE bearing some labels handwritten by E. Simon that suggest it might be an intended type for the name. Additional herbarium material and wild populations from Karoo region were identified that matched the samples in that voucher, and taxonomic research was conducted to clarify their identity. Those Karoo plants show a woody dense compact habit, woody perennial at base; stems prostrate to ascendent; leaves entirely glabrous, somewhat glaucous; with large white-hyaline conspicuous stipules; inflorescence glanduliferous, many-flowered subdichasial cyme, with minute bracts; flowers small, with white petals about equalling sepals, stamens 78, and styles free from base; capsule small, with seeds dimorphic, unwinged to broadly winged, with testa always densely tuberculate, among other characters. Molecular analyses of plastid (trnL-trnF region) and nuclear ribosomal (5.8S-ITS2 region) DNA sequence data support morphological differentiation of the Karoo plants, for which the name S. hanoverensis is here effectively published. A full morphological description, as well as data on ecology, habitat, distribution, and taxonomic and phylogenetic relationships of S. hanoverensis are compared to other members of the “South African group”, namely S. glandulosa. S. namaquensis, and S. quartzicola, from which the new species considerably differs. Adaptative significance of dimorphic seeds of S. hanoverensis is briefly commented in context to habitat preference of the species. An identification key is presented for the South African related taxa.
Keywords:
ITS phylogeny
; plant endemics
; plant morphology
; South African flora
; Spergularia
; Sperguleae
; taxonomy
; trnL-trnF phylogeny.
1. Introduction
Spergularia (Pers.) J.Presl & C.Presl, nom. cons., comprises ca 60 species of both annual and perennial plants with worldwide distribution, but mainly occurring in the Mediterranean Basin and temperate South America [1]. Some members of the genus are narrow endemics, while others are subcosmopolitan synanthropic plants [2]. In most cases, they often occur on soils rich in mineral salts, such as chlorides, sulphates, and nitrates, in both natural and human-disturbed habitats.
In recent times, molecular phylogenies recovered a consistent clade including Spergularia, Spergula L. and Rhodalsine Gay. Some authors [3,4] considered this group to be a distinct tribe to which the name Sperguleae Dumort. was applied, thus disregarding the traditional tripartite subfamilial arrangement of Caryophyllaceae Juss. [5], in which Spergularia was placed in Tribe Polycarpaeae DC., subfam. Paronychioideae (Juss.) Meissn. [2,5].
In the last decades, the relationships of Spergularia to other caryophyllid genera, such as Spergula, have been a matter of controversy [6, and references therein]. However, recent phylogenies by Kool and Thulin [7], based on plastid DNA sequences, recovered samples of Spergula and Spergularia (including Sanctambrosia Skottsb. and Spergularia flaccida (Madden) I.M.Turner = S. fallax Lowe) forming monophyletic sister clades, supported by the different number of styles and fruit valves, in both cases 5 vs. 3 respectively [8,9]. In this context, maintaining separate identities of both genera appears to be a good choice that is followed here.
Regarding the Spergularia representatives in southern part of Africa, Alonso et al. [8] have recently reinstated or validated two neglected species from western South Africa, i.e., S. glandulosa (Jacq.) Heynh. and S. namaquensis Schltr. ex M.Á.Alonso et al., and have also described [9] a rare endemic restricted to the quartz outcrops from Knersvlakte (SW Namaqualand), i.e., S. quartzicola M.Á.Alonso et al. All three species are perennials that were usually misidentified as the subcosmopolitan halophytic S. media (L.) C.Presl or as the nitrophilous alien S. bocconei (Scheele) Graebn. However, sound morphological vegetative and reproductive differences allow easy distinction from the rest of members of the genus [9]. Those three outstanding newly recovered or described taxa are to be added to the other species of Spergularia usually referred to in South Africa [10,11,12]. One of those is the enigmatic S. hanoverensis E.Simon, a name that apparently was never validly published [13]. This name has been applied to annual plants occurring in broad elevation and geographical ranges through Northern and Western Cape [14]. However, that concept often does not fit with the scarce herbarium material identified as S. hanoverensis, a plant whose identity remains confusing.
As part of an investigation project of “H2020 Research and Innovation Staff Exchange (RISE) Programme” of the European Commission (project num. 645636), intensive field work was carried out in western South Africa between 2015 and 2018, which has been complemented in subsequent years with new field explorations. During that research, some populations of an outstanding subshrubby and strongly woody at base, glabrous, white-flowered member of Spergularia were observed in inland saline and subsaline habitats of the Nama-Karoo Biome, mostly in the Northern Cape Province. A close examination of both living material and vouchers from South African herbaria revealed that those Karoo populations exhibited a combination of macro- and micromorphological characters and ecology absent in any described Southern African taxa of the genus. However, they are a perfect match with samples in a voucher labelled “Spergularia hanoverensis” made by the French botanist Eugène E. Simon and that is conserved in the Pretoria herbarium (PRE). This material is clearly akin to other members in the so-called group of “South African taxa” of Spergularia [sensu 8], which sometimes were misidentified as S. media. Phylogenetic studies of the Karoo plants, using nuclear (5.8S-ITS2 region) and plastid (trnL-trnF region) DNA sequence data, revealed that they could not be identified either with the Northern Hemisphere taxa of the S. media group, nor with other South African members of the genus.
In the present contribution, Spergularia hanoverensis is validly described for a plant endemic to the Nama-Karoo (NK) Biome [15] in western South Africa, according to the original concept “in schedis” of E.E. Simon. A brief story on that name is presented, and data on its morphology, ecology, distribution, and phylogenetic relationships are reported that support recognition of S. hanoverensis at specific rank in the “South African taxa” group.
2. Materials and Methods
2.1. Morphological and habitat studies
Detailed morphological studies were undertaken on both living plants from wild populations and herbarium specimens sourced from the herbaria ABH, BOL, GRA, K, M, HBG, NBG, P, and PRE (acronyms according to Thiers [16]), using an OLYMPUS SZX7 binocular. A personal collection of one of the authors (LM) that is currently deposited at ABH, including numerous vouchers of Spergularia, was studied. Digital images of Spergularia from iNaturalist (https://www.inaturalist.org/observations/?place_id=any&taxon_id=58170) were also checked and some found meeting the distribution of the studied species. Scanning Electron Microscope (SEM) micrographs of seeds were taken with a JEOL JSM-IT500HR operating at 15 kV. No special treatment of the material was required prior to observation. At least 5 mature seeds from different individuals of S. hanoverensis were observed in detail when available. Samples were glued directly on metallic stubs and then coated with 10 nm platinum in a QUORUM Q150T ES Plus sputter coater. The ImageJ software [17] was used for measurements on SEM micrographs. Authors of the taxa cited in the text follow IPNI [18]. Nomenclatural issues follow Turland et al. [19]; orthography of geographical names accord with Leistner and Morris [20], and the grid-number system is in accordance with the National Geospatial Information [21]. Bioclimate, bioregion, and vegetation classification agrees with Mucina and Rutherford [15].

Table 1.
Seed samples of Spergularia hanoverensis for SEM studies, with provenance and vouchers.
| Locality | Voucher |
|---|---|
| ZA: Northern Cape, Rietfontein to Brandvlei, G.Germishuizen 4022 | PRE0694503 |
| ZA: Northern Cape, Farm Grootfontein, G.Germishuizen 6524 | PRE0791338 |
| ZA: Northern Cape, Calvinia, A.A.Schmidt 408 | PRE0405924 |
| ZA: Northern Cape, Hanover, J.P.H.Acocks 18797 | PRE0405795 |
2.2. Molecular analyses
Herbarium vouchers and silica gel-dried material were used for total DNA extraction employing a modified 2 × cetyltrimethylammonium bromide (CTAB) protocol [22]. Total DNA was purified using MOBIO minicolumns and mostly kept in 0.1 × TE buffer [10 mM Tris-HCl, 1 mM ethylenediaminetetraacetic acid (EDTA), pH 8.0]. The trnL-trnF region (hereafter trnL-F) of chloroplast DNA (cpDNA) was amplified using specific primers trnL-BOC and trnL-BOF-R as described in Oxelman et al. [23], whereas the whole internal transcribed spacer –ITS– region (ITS1 spacer, 5.8S gene, ITS2 spacer) of nuclear ribosomal DNA (nrDNA) was amplified using the ITS1 (forward) and ITS4 (reverse) primers [24] and then adjusted to match the length of the 5.8S-ITS2 (hereafter ITS2) sequences retrieved from GenBank for alignment. Amplifications were performed on a reaction volume of 25 μL containing 22 μL of ABGene 1.1 × Master Mix, 2.5 mM MgCl2 (Thermo Scientific, Waltham, MA, USA), 0.5 μL of 0.4% bovine serum albumin (BSA), 0.5 μL of dimethyl sulfoxide (DMSO), 0.5 μL of each primer (10 pmol/μL) and 1 μL of template DNA on a 9700 GeneAmpl thermocycler (Applied Biosystems). The PCR programme for trnL-F was: 2 min at 97 °C, followed by 35 cycles of 97 °C for 20 s, 55 °C for 50 s, 72 °C for 1.5 min and a final extension of 72 °C for 8 min. The PCR programme for ITS2 was: 2 min at 95 °C, followed by 30 cycles of 95 °C for 1 min, 53 °C for 1 min, 72 °C for 2 min and a final extension of 72 °C for 5 min. In all cases, Sequencer 4.1 (Gene Codes Corp., Ann Arbor, MI, USA) was used to assemble complementary strands and to verify software base-calling.
Sequencer 4.1 (Gene Codes Corp., Ann Arbor, MI, USA) was used to assemble complementary strands and verify software base-calling. Sequence alignment was performed using MUSCLE [25] conducted in MEGA X v.10.2.6 [26] with minor manual adjustments to get the final aligned matrix. Forty-five samples belonging to 24 species of Spergularia were used for phylogenetic reconstructions, using Rhodalsine geniculata (Poir.) F.N.Williams, Rh. platyphylla Gay in Christ, Spergula arvensis L., Spergula morisonii Boreau and Spergula pentandra L., as outgroups. Two datasets were built: one for the trnL-F region of cpDNA (matrix with 33 sequences and 990 positions) and another for the ITS2 region of nuclear nrDNA (matrix with 25 sequences and 282 positions). Sequences of each region were retrieved from different plant sources depending on GenBank availability [8,9], except for four of them gathered in South Africa and Spain, which were generated specifically for the present study (Table 2). For that reason, most accessions in the trnL-F and ITS2 data sets do not come from a unique plant source.
Phylogenetic analyses of both regions were obtained using Maximum Parsimony (MP), Maximum Likelihood (ML) and Neighbour Joining (NJ) methods. MP analysis was conducted in both PAUP (using Heuristic search options with the tree searching strategy based on Nearest Neighbour Interchange, NNI) and MEGA (using Heuristic search options with the tree searching strategy based on Subtree-Pruning-Regrafting –SPR– with search level 1; [27]) for result comparison, with 10,000 replicates. ML [28] and NJ [29] analyses were also performed in MEGA, as well as the selection of the best model of DNA substitutions for each method using the Akaike Information Criterion (AIC; [30]); models with the lowest BIC (Bayesian Information Criterion) scores were considered to best describe the substitution pattern for the ML and NJ analyses. Moreover, jModelTest 2.1.10 [31] was performed to determine the best model of DNA substitutions for the BI analyses, also using AIC. Phylogenetic reconstructions for ML and evolutionary distances for NJ for the ITS2 matrix were estimated using the K2 model (2-parameter method of Kimura [32]) with the rate variation model allowed for some sites to be evolutionarily invariable (+I, 30.26% sites), whereas for the trnL-F matrix the T92 model (3-parameter method of Tamura [33]) was applied with a discrete gamma distribution (G = 0.677) modelling the rate variation among sites. In every case, all sites in the matrixes were considered. For comparison purposes, remotion of all ambiguous positions for each sequence pair (Pairwise Deletion option) was also performed and no significant differences (only affecting BS values in a few branches) were observed in the obtained phylogenies.
Furthermore, Bayesian inference (BI) analyses were conducted with MrBayes 3.2 [34], in which the Markov Chain Monte Carlo (MCMC) algorithm was run for 10 million generations and sampled every 1000 generations. Two runs were performed. The general time reversible (GTR) + proportion of invariant sites (I) + gamma distribution (G) model was used in the analyses (set nst = 6 rates = invgamma), according to the results obtained with jModelTest under AIC. The first 25% generations (burninfrac = 0.25) were excluded, and the remaining trees were used to compile a posterior probability (PP) distribution using a 50% majority-rule consensus.
For all methods, support was assessed by the bootstrap [35] with 10,000 replicates but holding only 10 trees per replicate. Clades showing bootstrap percentage (BP) values of 50–74% were considered as weakly supported, 75–89% moderately supported and 90–100% strongly supported.
3. Results and discussion
3.1. On the trail of Spergularia hanoverensis: a brief story of the name and its application
The name “Spergularia hanoverensis Simon” was reported for the first time by Olivier and Germishuizen [36] in the list of South African Spergularia together with other seven taxa (six species plus and additional variety), to which no additional data were supplied. The species was not mentioned by Goldblatt and Manning [10], who only accepted three species in that genus, namely S. media, S. pallida G.Don (erroneously cited as Spergularia pallida (Dumort.) Piré, the authorship that corresponds to Stellaria pallida), and S. rubra. The name S. pallida is currently treated in synonymy of S. media. Later, Nkonki [14] in her compilation of the South African Caryophyllaceae, recovered S. hanoverensis and listed it together with three representatives of that genus: S. bocconei, S. media, and S. rubra. There, S. hanoverensis was catalogued as “Annual. Herb. Ht ?–0.03 m. Alt 300–1370 m. NC, WC”, this indicating that it was considered to be an annual herb, rather small in size, and occurring within a broad elevation range through the Northern Cape and Western Cape provinces of South Africa. However, that name was not listed by Snijman [12], who again only recognised three species (S. bocconei S. media, and S. rubra) in the Extra Cape Subregion (Greater Cape Floristic Region) of South Africa.
The recovery of an almost forgotten name such as S. hanoverensis most probably followed Nkonki’s revision of the Spergularia vouchers conserved in the National Herbarium at Pretoria. Among them, the specimen PRE12453 (Figure 1), which was part of E.E. Galpin’s personal collection carrying the num. 5967, bears six plant fragments and three labels with relevant information. Firstly, the Galpin’s herbarium label with the original collection data handwritten in ink: “Spergularia / Hanover C.C. / Coll: T.R. Sim, Jan 1902”. The collector, Dr. Thomas R. Sim (1858–1838), was an English botanist specialised in forestry, who worked from September 1894 to September 1902 in the Forestry Service of the Cape Colony as superintendent of plantations, stationed at Fort Cunnynghame, just north of Stutterheim (Eastern Cape Region), and then he moved to Natal. By that time, he should have had collected abundant plant material in the region that he distributed to contemporary botanists [37], including surely Galpin’s specimen. Secondly, a label handwritten in pencil and signed by E. Simon reads: “Species mihi adhuc ignota / An Sp. pallida G.Don.?? / Ulterius / nomen dabo” [A species still unknown to me. Perhaps Sp. pallida G.Don.?? / Later I will name it]. Finally, a third label with an unidentified handwriting in ink reads: “Scrap removed for / Dr. E. Simon, / Vice President of the / Soc. Bot. du Centre-Ouest / France, / (sent through the / S. A. Museum C.T. 5.1.39)”. This latter label is glued exactly in the place left by the fragment forwarded to Dr. Simon via Cape Town. Undoubtedly, PRE12453 is the material from which the name Spergularia hanoverensis was invented, and hence it might be considered as the intended type. This fact is crucial for a correct interpretation and further application of the name in its original concept, as shown below.
In this regard, it is worth mentioning that some confusion exists around the true authorship of S. hanoverensis. In IPNI [18] and POWO [13] that species name is connected to “C.Simon”, which is the standard abbreviation of Charles Simon (1908–1987). On the contrary, most of the South African literature as well as TROPICOS [38] attribute it to “Simon”, the standardisation of Eugène Simon (1848–1924), also known as “E.Simon primus”, which is partly congruent with the text annotated on the pencil label on the voucher PRE12453. However, a detail in that text allows a different but more accurate interpretation, since the fragment missing in that voucher was indeed sent in 1939 to E. Simon, who was then the Vice-President of the Societé Botanique du Centre-Ouest (France). According to Guédès [39], Eugène Ernest Simon (1871–1967), whose standard form is “E.Simon” (sometimes also cited as “E.Simon secundus”, cf. [40]), was the botanist who held the vice-presidency of the regional French botanical society since 1908, and who was specifically working on an unfinished monograph on the genus Spergularia. Therefore, he was the real proposer of the name S. hanoverensis, irrespectively the taxon name was never validly published and remains as “nomen nudum” according to Art. 38 Ex. 1 of the ICN [19].
In this regard, it is worth mentioning that some confusion exists around the true authorship of S. hanoverensis. In IPNI [18] and POWO [13] that species name is connected to “C.Simon”, which is the standard abbreviation of Charles Simon (1908–1987). On the contrary, most of the South African literature as well as TROPICOS [38] attribute it to “Simon”, the standardisation of Eugène Simon (1848–1924), also known as “E.Simon primus”, which is partly congruent with the text annotated on the pencil label on the voucher PRE12453. However, a detail in that text allows a different but more accurate interpretation, since the fragment missing in that voucher was indeed sent in 1939 to E. Simon, who was then the Vice-President of the Societé Botanique du Centre-Ouest (France). According to Guédès [39], Eugène Ernest Simon (1871–1967), whose standard form is “E.Simon” (sometimes also cited as “E.Simon secundus”, cf. [40]), was the botanist who held the vice-presidency of the regional French botanical society since 1908, and who was specifically working on an unfinished monograph on the genus Spergularia. Therefore, he was the real proposer of the name S. hanoverensis, irrespectively the taxon name was never validly published and remains as “nomen nudum” according to Art. 38 Ex. 1 of the ICN [19].
That name, however, is currently available in several checklists and web pages featuring the African and/or South African floras, although no relevant new information has been added other than Nkonki’s [14]. In particular, S. hanoverensis was assessed as LC (Least Concern) by Cholo [41], according to the IUCN red list categories [42], mainly on the basis of the already published information that included mapping of four sites far apart based on vouchers at PRE. Both morphological and distributional data of Nkonki [14] are also shown in APD [43] and in GBIF [44].
The extant published information on S. hanoverensis is still very scarce and rather confusing, particularly after reviewing herbarium material of Spergularia at PRE. The fragments affixed to the voucher PRE12453 (Figure 1) show some outstanding characteristics allowing safe identification:
- i)
- a subshrubby perennial plant, mostly glabrous, with a compact, often many-branched woody underground base;
- ii)
- stems often prostrate-ascendent;
- iii)
- leaves somewhat glaucous, with large whitish stipules, very apparent and showy, long-lasting, giving a Paronychia-like aspect;
- iv)
- inflorescence glanduliferous, broadly subdichasial, with inconspicuous bracts;
- v)
- flowers numerous, small, with white petals;
- vi)
- capsule small, many-seeded;
- vii)
- seeds small, triangular, blackish-brown, matte, densely papillate, unwinged or with a discolorous, vestigial to poorly developed wing.
At first glance, they show some resemblance with other South African taxa of Spergularia, often misidentified as S. media. This might justify the scarcity of references to S. hanoverensis in the literature prior to Nkonki [14], and also neglection of that name in more recent accounts [12]. Other specimens at PRE first identified as S. hanoverensis indeed correspond to S. bocconei (collection M.B.Bayer 6006, PRE762662) or S. namaquensis (collection H. M.Steyn 23, PRE583500).
Despite the contrasting interpretations of S. hanoverensis, the Karoo plants matching Simon’s concept of that species do not fit the current application of the name in recent checklists and databases of the African flora. Further, the unique combination of characters found in Simon’s taxon is missing in the remaining members of the “South African taxa” group of Spergularia, as defined by Alonso et al. [8,9]. The newly obtained morphological and molecular evidence support acceptance of the taxon at specific rank.
3.2. Taxonomic treatment
3.2.1. Spergularia hanoverensis E.Simon ex M.Á.Alonso, M.B.Crespo, Mart.-Azorín & Mucina, sp. nov. (Figure 1)
Type: [South Africa. Northern Cape, 3124 (Hanover)]. Herbarium-E.E. Galpin, nº 5967 – Hanover C[ape]. C[olony]., January 1902, T.R. Sim s.n. (holotype: PRE12453!). Note: A fragment of this gathering (an intended isotype) was apparently sent to E. Simon in 1939, but no such material is currently conserved among Simon’s collection at MPU (Montpellier). Maybe it was lost during shipping or as a result of the start of the World War II.
Diagnosis: Planta speciosa ab Spergularia namaquensi et S. glandulosa ob characteribus vegetativis ex parte simillima; a priore caulibus compactis a basi valde lignosis et inflorescentiis terminalibus fere ebracteatis (bracteis minutissimis) accedit, et a posteriore floribus parvis imprimis petalis quam sepalis aequilongis vel parce brevioribus et stylis e basi omnino liberis congruit. Sed plane ab eas distinctissima et bene distinguenda caulibus foliisque glaberrimis; foliis longiore mucronatis (ad 1 mm), stipulis magnis, acuminatis, hyalinis, speciosis, longe persistentibus aspectu Paronychiae; staminibus 7–8; et praecipue seminibus dimorphis, aliis minoribus, 0.5–0.75 × 0.3–0.6 mm, exalatis obovato-cunetatis, et al.iis majoribus, 0.4–1.1(–1.4) × 0.6–1.2(–1.4) mm, ala vestigialibus vel perfecta (sed formis aliis in alias transeuntes), omnibus disco densiore et uniformiter tuberculato-papilloso.
Description: Subshrub, mostly glabrous excepting the glanduliferous inflorescence, with a compact, often many-branched woody underground base. Stems up to 30 cm high, but commonly smaller, usually prostrate, subcespitose, slightly nodose, with ascending branches. Leaves 4–10(–17) × 0.5–0.7 mm, narrowly linear, semicylindrical and subcanaliculate, green to glaucous-green, caducous when withering, ending in a whitish to yellowish mucro up to 1 mm long; stipules 4–6 × 1–2 mm, whitish-scarious, glabrous, triangular-acuminate, showy and apparent, often reaching at least half the leaf length (namely in the axillary leaf fascicles, where stipules are longer than leaves themselves), those on the young shoots fused up to the basal third and finally only at the base, long-lasting after leaf abscission and giving a Paronychia-like aspect. Inflorescence a sub-dichasial cyme, broadly branched, many-flowered (5–7 flowers per branch) and densely covered with glanduliferous short hairs; bracts 1–2.5 mm long, slightly longer than stipules, inconspicuous, much shorter than leaves. Flowers pentamerous, on erect-patent to patent slender pedicels 2.5–5 mm long at anthesis. Sepals 2–3.3 × 1–1.5 mm, slightly accrescent in fruit, oblong to elliptic, obtuse to subacute, with a central green band 0.5–0.7 mm broad, and scarious margins 0.3–0.4 mm broad (wider in the inner sepals), patent to slightly deflexed at anthesis. Petals 1.8–2.8 × 1–1.2 mm, about equalling to slightly shorter than sepals, white, elliptic, entire. Stamens 7–8, slightly shorter than petals, filament up to 3 mm long, filiform, slightly widened at the base, whitish, anther 0.3–0.4 mm long, yellow, dorsifixed. Ovary ca 1–2 mm, subglobose, yellowish; styles 3, ca 1 mm long, free from base, yellowish, with short apical stigmata. Capsule 3–4 × 2.5–3.5 mm, broadly ovate, slightly longer than sepals, glabrous, shinning, yellowish-green outside but reddish inside, opening by three slightly recurved valves, on patent to reflexed pedicels 4–10 mm long, up to 3 times longer than sepals. Seeds numerous, dimorphic; some of them unwinged, 0.5–0.75 × 0.3–0.6 mm, ovate-cuneate to subtriangular in outline, flattened, blackish-brown and matte, the others similar but larger, 0.7–1.1(–1.4) × 0.6–1.2(–1.4) mm, with disk 0.6–0.9 × 0.4–0.8 mm and a discolorous (whitish to greyish), vestigial to entirely developed, eroded wing (both types usually present in a single capsule, with intermediate stages); testa ornamented in all cases with minute irregular tubercles and densely covered all over with stalked globose and also minutely tuberculated papillae.
Etymology: The specific epithet (hanoverensis, –e) refers to Hanover, a small town in the Karoo region of the Northern Cape Province in central South Africa, from where the plant was collected and is native to. Provided that E.E. Simon was the first in recognising this taxon as new “in schedis”, we preserve the original name he chose later.
Phenology: Flowering in late October-early January (occasionally in July–August), fruiting in November-February (occasionally in August–September).
Habitat and distribution: Spergularia hanoverensis is a edaphophilous species usually found on seasonal stream banks, in riverbeds and depressions, often on saline calcareous-clayish or sandy soils but sometimes among rocks or stony saline substrates. The elevation of the localities ranges between 700 and 1400 m. The known distribution of the species extends through most of the southern part of the Karoo Region in central and western South Africa, ranging from Ceres and Calitzdorp in the Western Cape to Hanover in the Northern Cape Province (Figure 2), a territory mostly included in the Nama-Karoo (NK) Biome and reaching the southern Succulent-Karoo biomes (mostly the SKk, SKt and SKv Bioregions) sensu Mucina and Rutherford [15], where it occurs in the so-called “Bushmanland vloere” (code AZi 5). In those areas, the climate is subdesert arid and continental (only scarcely ameliorated by the ocean influences), with average temperatures ranging from –5°C in winter to 43°C in summer and frosts being usual at high elevations. The average annual precipitation varies between 100 and 500 mm, though rather differently distributed, the rainfall occurring mostly during late summer (December to April) with a peak in March [15].
Notes: We attribute authorship of the new species to Eugène E. Simon, standardised E.Simon, who was the first botanist to identify it “in schedis” as a new species. Wild populations of that species include numerous individuals covering a large territory in South Africa, and no special threats are known so far to occur that might lead to any inferred decline in either the number of populations or the number of individuals. Therefore, its conservation status is suggested here as Least Concern (LC) according to IUCN [42].
Other studied materials: South Africa. Northern Cape Province: 3020 (Brandvlei): Between Brandvlei and Williston, Karreekop farm (-DC), 30°58’59”S 20°39’18”E, 987 m, P3624 August 2022, M.Mart.Azorín, M.B.Crespo, M.Á.Alonso, J.L.Villar & M.Pinter (ABH83276). 3119 (Calvinia): Calvinia C[ape]. P[rovince]. (-BD), 1936, ut Spergularia marginata Kit., A.A.Schmidt 408 (PRE0405924). 3119 (Calvinia): Banks of Kareehoutrivier, 24 km south of Bo-Downes (-DD), cushion along the seasonal stream banks, 20 November 1983, D.Snijman 772 (NBG). 3120 (Williston): Calvinia district, Rietfontein, on road to Brandvlei (-AC), karoo, sandy soil, 29 November 1986, G.Germishuizen 4022 (PRE0694503). 3121 (Fraseburgh): Williston District, Farm Grootfontein, about 42 km north Williston, on road to Rheebokhyer, about 7 km west of farmhouse (-AA), elev. 1207 m, karoo, among rocks, alongside dry, sandy riverbed, 2 April 1993, G.Germishuizen 6524 (PRE0791338). 3124 (Hanover): Outskirts of Hanover, on S. side (-AB), in white calcareous clay in vley, locally freq., elev. 4500 ft, 27 February 1956, J.P.H.Acocks 18797 (3 sheets) (PRE0405795). 3124 (Hanover): SW side of Hanover (-AB), kalk vlei, elev. 4500 ft, 19 February 1959, J.P.H.Acocks 20246 (3 sheets) (PRE0405797). 3220 (Sutherland): Roggeveld, Soekop, Onderste grasvlakte camp (-AA), 32°03′24.5”S 20°08′39.5”E, elev. 1193 m, 27 September 2006, H.Rosch 656 (NBG209250). 3220 (Sutherland): SW of Middelpos, road to Ganaga Pass, Zoekop Farm, Witdam (-AA), 32°03’16.5”S 20°08’39.2”E, 1190 m, 25 August 2022, M.Mart.Azorín, M.B.Crespo, M.Á.Alonso, J.L.Villar & M.Pinter (ABH83287; ABH83288). Western Cape Province: 3219 (Wuppertal): Swartruggens, Knolfontein (-DC), 32°51′17.2”S; 19°36′03”E, elev. 1190 m, 3 February 2011, I.Jardine 1522 (NBG276922). 3219 (Wuppertal): Swartruggens, Knolfontein (-DC), 32°51′17.2”S 19°37′05”E, elev. 1186 m, 12 December 2011, I.Jardine 1760 (NBG277559). 3319 (Worcester): Ceres, Bokkerivier Farm (-BD), 11 November 1963, L.J.Booysen 111 (NBG).
3.3. Taxonomic and phylogenetic relationships of Spergularia hanoverensis
In the context of an ongoing phylogenetic survey on the South African taxa of Spergularia based on trnL-F cpDNA and ITS2 nDNA sequences, we recently reported [8,9] the first preliminary phylogenetic trees including southern African taxa of that genus. Our results were congruent with the plastid phylogeny of Sperguleae (sensu [3]) obtained by Kool and Thulin [7], and also supported that trnL-F and ITS2 regions offer information useful enough for credible phylogenetic reconstructions of Spergularia.
The addition of samples of S. hanoverensis to our molecular matrix yields trees almost identical in general topology to those obtained in our previous contributions [8,9]. Further, the new species falls nested in the “South African taxa” subclade of Clade A [sensu 8, 9] in both the plastid and the nuclear phylogenies (Figure 3 and Figure 4), as recovered in our BI consensus trees (in which PP values are placed above branches and BP values below branches, respectively from the ML and MP analyses). However, the internal relationships among the four members of that southern lineage are not equally resolved, and often with low PP and BP values.
The aligned trnL-F database was 990 bp, 147 of which (14.85%) were potentially parsimony informative. Analyses of this dataset, using NJ, MP, ML, and BI methods yielded trees with similar topologies and similar bootstrap and branch length values. The obtained trnL-F BI phylogenetic tree (Figure 3) recovers all four South African taxa in the strongly supported Clade A (1.00 PP, 97/100 BP), in which both samples of S. hanoverensis are unresolved together with the remaining accessions of S. glandulosa, S. namaquensis and S. quartzicola to form the group of “South African taxa” (1.00 PP, 87/100 BP). They all fall together in a polytomic group (1.00 PP, 86/100 BP) with mostly Southern Hemisphere taxa, namely S. villosa (Pers.) Cambess., S. tasmanica (Kindb.) L.G.Adams, S. ramosa Cambess., S. pissisii I.M.Johnst. They all are sister to the well supported subclade (0.81 PP, 54/100 BP) formed by S. denticulata (Phil.) Phil. and the Pacific North American S. macrotheca (Hornem. ex Cham. & Schltdl.) Heynh.
The aligned ITS2 database was 282 bp, 41 of which (14.54%) were potentially parsimony informative. Also, analyses using NJ, MP, ML, and BI methods revealed trees with similar topologies and similar bootstrap and branch length values. The obtained ITS2 BI phylogenetic tree (Figure 4) places the samples of S. hanoverensis as well supported sister (0.99 PP, 88/94 BP) of the well supported subclade of S. glandulosa (0.93 PP, 80/79 BP), a subshrub from saline coastal areas of southwestern and southern South Africa. They both are well supported sister (0.99 PP, 88/94 BP) to the subclade including the Namaqualand woody subshrubs S. quartzicola and S. namaquensis, which constitute a strongly supported group (1.00 PP, 96/97 BP) not well resolved internally. All these are strongly supported sister (1.00 PP, 93/95 BP) to a third subclade formed by the reddish-flowered S. rubra and S. rupicola (1.00 PP, 88/92 BP) plus the Central European S. echinosperma (Čelak.) Asch. & Graebn. All those constitute the strongly supported (0.98 PP, 90/97 BP) Clade A.
Those molecular relationships of Spergularia hanoverensis to the remaining members in the “South African taxa” group are rather well correlated to some morphological traits (Table 3). The newly described species shares some floral resemblance with S. glandulosa, such as the small white flowers with petals equalling sepal in length, and the styles entirely free from base, features that might justify some phylogenetic closeness as recovered in our ITS2 tree (Figure 3B). However, S. hanoverensis shows a compact habit; leaves glabrous and long acuminate (ca 1 mm long), bearing at base long-lasting, broad stipules; and inflorescences multi-flowered, subdichasial, with minute bracts; all those allowing easy separation at first glance. In fact, S. hanoverensis exhibits a combination of characters missing as a whole in any of the known members of the South African group, though some of them occurring in any of the three other taxa. In particular, the compact and usually many-branched woody underground base and the multi-flowered subdichasial inflorescences with minute bracts are shared with S. namaquensis, and the entirely glabrous and broadly stipulate leaves are also present in S. quartzicola. But both latter taxa show larger flowers with 10 stamens and columnar, long-fused styles, instead of the smaller flowers with 7–8 stamens and free styles of S. hanoverensis. Finally, the capsule and seed features offer the most remarkable diagnostic differences among all those South African species. In fact, S. hanoverensis produces the smallest capsules (ca 3–4 × 2.5–3.5 mm), and also smaller seeds (0.4–0.5 mm long). These are markedly dimorphic, some of them being unwinged, ovate-cuneate to subtriangular in outline, flattened, blackish-brown and matte, whereas the others are similar but with a discolorous (whitish to greyish), vestigial to entirely developed, eroded wing, and testa surface ornamented with minute irregular tubercles and densely covered all over with stalked globose and also minutely tuberculated papillae. Both seed types are sometimes present in a single capsule (Figure 5a–c, f–g), with intermediate stages among individuals (Figure 5b) and among populations. This fact makes S. hanoverensis the only known member of the “South African taxa” group with such dimorphic seeds.
Seed dimorphism is rather frequent in Spergularia [40,45,46,47,48,49,50], and for a long time it was considered a good diagnostic character for species delimitation [51]. As it occurs in other dimorphic taxa such as the widespread annual pinkish-flowered S. marina (L.) Besser [52,53], heteromorphism also includes variable seed morphology as well as considerable variation in seed size in S. hanoverensis, the winged seeds being larger and their disk being also larger than the unwinged seeds. However, seeds of S. hanoverensis, although dimorphic in both size and gross morphology (winged and unwinged), are homogeneous in micromorphology, with traits quite distinct from S. marina (L.) Besser [54]. According to our observations from herbarium material, no correlation appears to exist between seed features and their geographical provenance, so that individuals from different sites apparently exhibit seeds with similar variation patterns of size and gross morphology. In other words, most of the variation in seed size occurs within rather than between populations, as also reported for S. salina [53].
Species with seed dimorphism combine different morphological syndromes for a more efficient dispersal in contrasting habitats, the wind being the vector for winged seeds whereas the water for unwinged ones, as suggested by Telenius and Torstensson [55]. Acquisition of dimorphism in S. hanoverensis might have brought a positive evolutionary effect for dispersal in the inland karroid saline ecosystems it inhabits in South Africa (i.e., seasonally wet depressions, intermittent riverbeds and ravines, etc.), by combining anemochory (winged seeds) and hydrochory (unwinged seeds). Its phylogenetically close relatives from Namaqualand (S. namaquensis and S. quatzicola) also occur in subdesert karroid ecosystems, mostly in dry sandy or quartz substrates, and therefore production of unwinged seeds would have no favourable adaptative value in sites not necessarily connected to marsh habitats.
4. Conclusion
Although Spergularia hanoverensis has been widely interpreted in a sense including several biological entities, the name is validated and circumscribed here as it was first outlined “in schedis” by E.E. Simon. Our molecular and morphological data are distinctive enough to accept it as a proper species, which falls together with taxa in the “South African taxa” group of Spergularia. It is endemic mostly to the inland saline habitats of the Nama-Karoo Biome (central-western South Africa).
New data from wild populations are needed to complete the distribution area of S. hanoverensis and to test the accuracy of our observations on herbarium vouchers regarding within- or between-population variation in seed size and gross morphology. The dichotomous identification key in Appendix A will surely help to that purpose.
Author Contributions
The following individual contributions were made: conceptualization, M.Á.A., M.B.C., M.M.A. and L.M; methodology, M.Á.A. M.B.C., and M.M.A.; formal analyses, M.B.C., M.Á.A. and M.M.A.; writing—original draft preparation, M.Á.A. and M.B.C.; writing—review and editing, M.B.C.; collecting permit acquisition, M.M.A. and L.M.; field work, M.Á.A., M.B.C., M.M.A. and L.M. All authors have contributed substantially to the general discussion of results and taxonomic treatment, and have also read and agreed to the published version of the manuscript.
Funding
This work was partly supported by H2020 Research and Innovation Staff Exchange Programme of the European Commission, project 645636: ‘Insect-plant relationships: insights into biodiversity and new applications’ (FlyHigh); and by the grants UAUSTI19-08, UAUSTI21-04, ACIE17-01, ACIE18-03 and ACIE21-01, from diverse programmes of Universidad de Alicante. L.M. acknowledges logistic support of the Iluka Chair in Vegetation Science and Biogeography at Murdoch University, Perth, Australia.
Data Availability Statement
DNA sequence data generated in the present research are available at GenBank (https://www.ncbi.nlm.nih.gov/genbank/).
Acknowledgments
We thank the curators and staff of the herbaria ABH, BOL, GRA, K, M, HBG, NBG, P and PRE for facilitating access or providing material and/or information on their collections of Spergularia. Especially, Cornelia Klak (BOL), Anthony Magee (NBG) and Erik van Wick (PRE) for their help during our visit to the South African herbaria. Further, Mashiane Mothogoane (PRE) facilitated permit for publication of the type voucher of S. hanoverensis. José Luis Villar (Universidad de Alicante) and Michael Pinter (Karl-Franzens University Graz) kindly helped with the field work, and Josy Zarur de Matos, Jordi Abad and Miguel Pascual (University of Alicante) with the laboratory work. All herbarium specimens were collected under permission provided by the Department of Environment and Nature Conservation of Northern Cape Province (permits FLORA0057/2017, FLORA0058/2017) and CapeNature of Western Cape Province of South Africa (permits AAA005-00080-0028, AAA008-00031-0028, 0027-AAA008-00699, 0028-AAA008-00203).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A. Identification key for taxa in the South African group of Spergularia.
- 1
- Petals 1.8–5 mm long, about equalling or slightly shorter than sepals; styles 0.5–0.7 mm long, free from the base; seed disk densely tuberculate or papillate ............................................................................................................................................................. 2
- -
- Petals 6–9 mm long, ca. 1.5 times longer than sepals; styles 1–2 mm long, fused up to the middle or beyond; seed disk smooth or with scattered conical tubercles or granular protuberances on edge .................................................................................................................................................................... 3
- 2
- Plant diffuse, suffruticose at base; leaves often densely hairy-glanduliferous (at least at base), with a short mucro 0.2–0.5 mm long; flowers with leaf-like bracts; stamens 10; seeds monomorphic, broadly winged, with disk loosely covered with conical tubercles ...................................................................................................................................... S. glandulosa
- -
- Plant compact, strongly woody at base; leaves entirely glabrous, with a long mucro up to 1 mm long; flowers with inconspicuous bracts, much shorter than leaves; stamens 7–8; seeds dimorphic, unwinged to broadly winged, with disk densely covered with stalked globose papillae ..................................................................................................................................... S. hanoverensis
- 3
- Branchlets and leaves densely glanduliferous at least when young (sometimes leaves glabrescent when withering); sepals 4–5 mm wide, patent to slightly deflexed at anthesis, with broad scarious margins; seed wing entire to slightly eroded ....................................................................................................................................... S. namaquensis
- -
- Branchlets and leaves entirely glabrous (sometimes branchlets sparsely glanduliferous when young); sepals 1.5–3 mm wide, strongly deflexed at anthesis, with narrow scarious margins; seed wing deeply and irregularly lacerate ............................................................................................................... S. quartzicola
References
- Hernández-Ledesma. , P, Berendsohn, W.G., Borsch, T., von Mering, S., Akhani, H., Arias, S., Castañeda-Noa, I., Eggli, U., Eriksson, R., Flores-Olvera, H., Fuentes-Bazán, S., Kadereit, G., Klak, C., Korotkova, N., Nyffeler, R., Ocampo, G., Ochoterena, H., Oxelman, B., Rabeler, R.K., Sánchez, A., Schlumpberger, B.O., Uotila, P. A taxonomic backbone for the global synthesis of species diversity in the angiosperm order Caryophyllales. Willdenowia 2015, 45, 281–383. [Google Scholar] [CrossRef]
- Bittrich, V. Caryophyllaceae. In The families and genera of vascular plants; Kubitzki, K., Rohwer, J.C., Bittrich, V., Eds.; Springer, Berlin, 1993; Vol. 2 (Magnoliid, Hamamelid, and Caryophyllid families). pp. 206–236.
- Harbaugh, D.T. , Nepokroeff, M., Rabeler, R.K., McNeill, J., Zimmer, E.A., Wagner, W.L. A new lineage-based tribal classification of the family Caryophyllaceae. Int. J. Pl. Sci. 2010, 171, 185–198. [Google Scholar] [CrossRef]
- Greenberg, A.K. , Donoghue, M.J. Molecular systematics and character evolution in Caryophyllaceae. Taxon 2011, 60, 1637–1652. [Google Scholar] [CrossRef]
- Pax, F. , Hoffmann, K. Caryophyllaceae. In Die naturlichen Pflanzenfamilien, 2nd edition; Engler, A., Harms, H., Eds.; Wilhelm Engelmann, Leipzig, 1934; Vol. 16c; pp. 275–364.
- López González, G. Sobre el género Spergula L. [incl. Spergularia (Pers.) Pers. ex J. Presl & C. Presl, nom. cons.] (Caryophyllaceae) y sus especies en la península ibérica e Islas Baleares. Lagascalia 2010, 30, 7–18. [Google Scholar]
- Kool, A. , Thulin, M. A giant spurrey on a tiny island: on the phylogenetic position of Sanctambrosia manicata (Caryophyllaceae) and the generic circumscriptions of Spergula, Spergularia and Rhodalsine. Taxon 2017, 66, 615–622. [Google Scholar] [CrossRef]
- Alonso, M.Á. , Crespo, M.B., Martínez-Azorín, M., Mucina, L. Taxonomic identity and evolutionary relationships of South African taxa related to the Spergularia media group (Caryophyllaceae). Plant Syst. Evol. 2021, 307, 24. [Google Scholar] [CrossRef]
- Alonso, M.Á. , Crespo, M.B., Martínez-Azorín, M., Mucina, L. Morphological and molecular data support recognition of Spergularia quartzicola (Caryophyllaceae) as a new species endemic to South Africa. Plant Biosyst. 2022, 156, 506–514. [Google Scholar] [CrossRef]
- Goldblatt, P. , Manning, J.C., Eds. Cape plants: a conspectus of the Cape Flora of South Africa; National botanical institute, Cape Town and Missouri Botanical Garden, St Louis, USA, 2000.
- Manning, J.C. , Goldblatt, P., Eds. Plants of the greater cape floristic region, 1: the core cape flora; South African National Biodiversity Institute, Pretoria, South Africa, 2012.
- Snijman, D.A. Caryophyllaceae (= Illicebraceae). In Plants of the greater cape floristic region, 2: the extra cape flora; Snijman, D.A.., Ed.; South African National Biodiversity Institute, Pretoria, South Africa, 2013, pp. 346–348.
- POWO. Plants of the World Online (continuously updated). Available online: https://powo.science.kew.org/ (accessed on 2 May 2023).
- Nkonki, T. Caryophyllacee. In Plants of southern Africa: an annotated checklist; Germishuizen, G., Meyer, N.L., Eds.; South African National Biodiversity Institute, Pretoria, South Africa, 2003 [Strelitzia 14].
- Mucina, L., Rutherford, M.C., Eds. The vegetation of South Africa, Lesotho and Swaziland. South African National Biodiversity Institute: Pretoria, South Africa, 2006. [Strelitzia 19].
- Thiers, B. Index Herbariorum: a global directory of public herbaria and associated staff (continuously updated). Available online: http://sweetgum.nybg.org/ih/ (accessed on 3 May 2023).
- Rasband, W.S.; ImageJ. U.S. National Institutes of Health, Bethesda, Maryland. Available online: https://imagej.nih.gov/ij/ (accessed on 3 May 2023).
- IPNI. The International Plant Names Index. Available online: http://www.ipni.org (accessed on 3 May 2023).
- Turland, N.J. , Wiersema, J.H., Barrie, F.R., Greuter, W., Hawksworth, D.L., Herendeen, P.S., Knapp, S., Kusber, W.-H., Li, D.-Z., Marhold, K., May, T.W., McNeill, J., Monro, A.M., Prado, J., Price, M.J., Smith G.F., Eds.; International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code) adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017. − Koeltz Scientific Books: Königstein, Germany, 2018. [Google Scholar] [CrossRef]
- Leistner, O.A. , Morris, J.W. Southern African place names. Ann. Cape Prov. Mus. Nat. Hist. 1976, 12, 1–565. [Google Scholar]
- National Geospatial Information. SA mapsheet referencing. Available online: http://www.ngi.gov.za/indexphp/what-we-do/maps-and-geospatial-information/41-sa-mapsheet-referencing (accessed on 3 May 2023).
- Doyle, J.J. , Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Oxelman B, Kornhall P, Olmstead RG, Bremer B Further disintegration of Scrophulariaceae. Taxon 2005, 54, 411–425. [CrossRef]
- White, T.J. , T.D., Lee, S.B., Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR protocols: a guide to methods and applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, USA, 1990. [Google Scholar]
- Edgar, R.C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucl. Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S. , Stecher, G., Li, M., Knyaz, C., Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. , Kumar, S. Molecular evolution and phylogenetics, Oxford University Press: New York, USA, 2000. [Google Scholar]
- Felsenstein, J. Evolutionary trees from DNA sequences: a maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Saitou, N. , Nei, M. The neighbour-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control. 1874, 19, 716–723. [Google Scholar] [CrossRef]
- Darriba, D. , Taboada, G.L., Doallo, R., Posada, D. jModelTest 2: more models, new heuristics and parallel computing. Nat. Method. 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G + C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef]
- Ronquist, F. , Teslenko, M., van der Mark, P., Ayres, D.L., Darling, A., Hohna, S., Larget, B., Liu, L., Suchard, M.A., Huelsenbeck, J.P. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Olivier, P.M. , Germishuizen, G. Caryophyllaceae. In List of species of southern African plants; Gibbs Russell, G.E., Germishuizen, G., Herman, P., Olivier, P.M., Perold, S.M., Reid, C. Retief,E., Smook, L., van Rooy, J, Welman, W.G., Eds.; Botanical Research Institute, Pretoria, South Africa, 1984, pp. 64–65. [Mem. Bot. Surv. South Africa 48].
- Plug, C. Sim, Dr Thomas Robertson (botany, forestry). S2A3 Biographical Database of Southern African Science. Available on line: https://www.s2a3.org.za/bio/Biograph_final.php?serial=2584 (accessed on 2 May 2023).
- TROPICOS. Tropicos.org. Missouri Botanical Garden. Available online: https://tropicos.org/ (accessed on 2 May 2023).
- Guédès, M. Eugène Ernest Simon (1871–1967). Bull. Soc. Bot. France, 1967, 114, 357–359. [Google Scholar] [CrossRef]
- Monnier, P. Ratter, J.A. Spergularia. In Flora europaea, 2nd edition; Tutin, T.G., Burges, N.A., Chater, A.O., Edmondson, J.R., Heywood, V.H., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds; Cambridge University Press, Cambridge, UK, 1993; Vol. 1, pp. 186–188.
- Cholo, F.; Spergularia hanoverensis Simon. National Assessment (2006/05/31): Red List of South African Plants, version 2020.1. Available online: http://redlist.sanbi.org/species.php?species=3967-6 (accessed on 3 May 2023).
- IUCN. IUCN Red List Categories and Criteria: Version 3.1, 2nd ed.; UCN Species Survival Commission: Gland, Switzerland, and Cambridge, UK, 2012. [Google Scholar]
- APD. 2008. The African Plant Database. Available on line: https://www.ville-ge.ch/musinfo/bd/cjb/africa/recherche.php (accessed 3 May 2023). 3 May.
- GBIF. The Global Biodiversity Information Facility. Available on line: https://www.gbif.org/species/8231915 (accessed 3 May 2023). 3 May.
- Rossbach, R.P. El género Spergularia (Caryophyllaceae) en Chile. Darwiniana 1943, 6, 211–256. [Google Scholar]
- Salisbury, E.J. Spergularia salina and Spergularia marginata and their heteromorphic seeds. Kew Bull. 1958, 1, 41–51. [Google Scholar]
- Maire, R. Flore de l’Afrique du Nord, vol. 9. Paul Lechevalier, Paris, France, 1963.
- Ratter, J.A. Spergularia. In Flora iberica; Castroviejo, S., Laínz, M., López González, G., Montserrat, P., Muñoz Garmendia, F., Paiva, J., Villar, L., Eds.; Real Jardín Botánico, CSIC, Madrid, Spain, 1990; vol, 2, pp. 149–161.
- Hartman, R.L., Rabeler, R.K. Spergularia. In Flora of North America north of Mexico; Flora of North America Editorial Committee, Eds.; Oxford University Press, Oxford, UK, 2005; vol. 5(2), pp. 16–23.
- Adams, L.G. , West, J.G., Cowley, K.J. Revision of Spergularia (Caryophyllaceae) in Australia. Austr. Syst. Bot. 2008, 21, 251–270. [Google Scholar] [CrossRef]
- Lebel, M.E. Morphologie du genre Spergularia. Bull. Soc. Bot. France 1868, 15, 50–64. [Google Scholar] [CrossRef]
- Sterk, A.A. Biosystematic studies on Spergularia media and S. marina in the Netherlands II. The morphological variation of S. marina. Acta. Bot. Neerl. 1969, 18, 467–476. [Google Scholar] [CrossRef]
- Telenius, A. , Torstensson, P. Seed type and seed size variation in the heteromorphic saltmarsh annual Spergularia salina along the coast of Sweden. Plant Biol. 1999, 1, 585–593. [Google Scholar] [CrossRef]
- Memon, R.A. , Bhatti, G.R., Khalid, S., Arshad, M.G., Mirbahar, A.A., Qureshi, A. Microstructural features of seeds of Spergularia marina (L.) Griseb. (Caryophyllaceae). Pak. J. Bot. 2010, 42, 1423–1427. [Google Scholar]
- Telenius, A. , Torstensson, P. Seed wings in relation to seeds size in the genus Spergularia. Oikos 1991, 61, 216–222. [Google Scholar] [CrossRef]
Figure 1.
Voucher designated here as holotype of Spergularia hanoverensis (PRE12453), which was studied and annotated by E.E. Simon as belonging to an unknown species (© SANBI, Herbarium, Pretoria). Ruler length = 15 cm (minimum scale = 0.5 mm).
Figure 1.
Voucher designated here as holotype of Spergularia hanoverensis (PRE12453), which was studied and annotated by E.E. Simon as belonging to an unknown species (© SANBI, Herbarium, Pretoria). Ruler length = 15 cm (minimum scale = 0.5 mm).

Figure 2.
Distribution map of the known species of the “South African taxa” group of Spergularia: S. hanoverensis (green hexagons); S. glandulosa (yellow circles); S. namaquensis (red squares); and S. quartzicola (greyish triangles).
Figure 2.
Distribution map of the known species of the “South African taxa” group of Spergularia: S. hanoverensis (green hexagons); S. glandulosa (yellow circles); S. namaquensis (red squares); and S. quartzicola (greyish triangles).
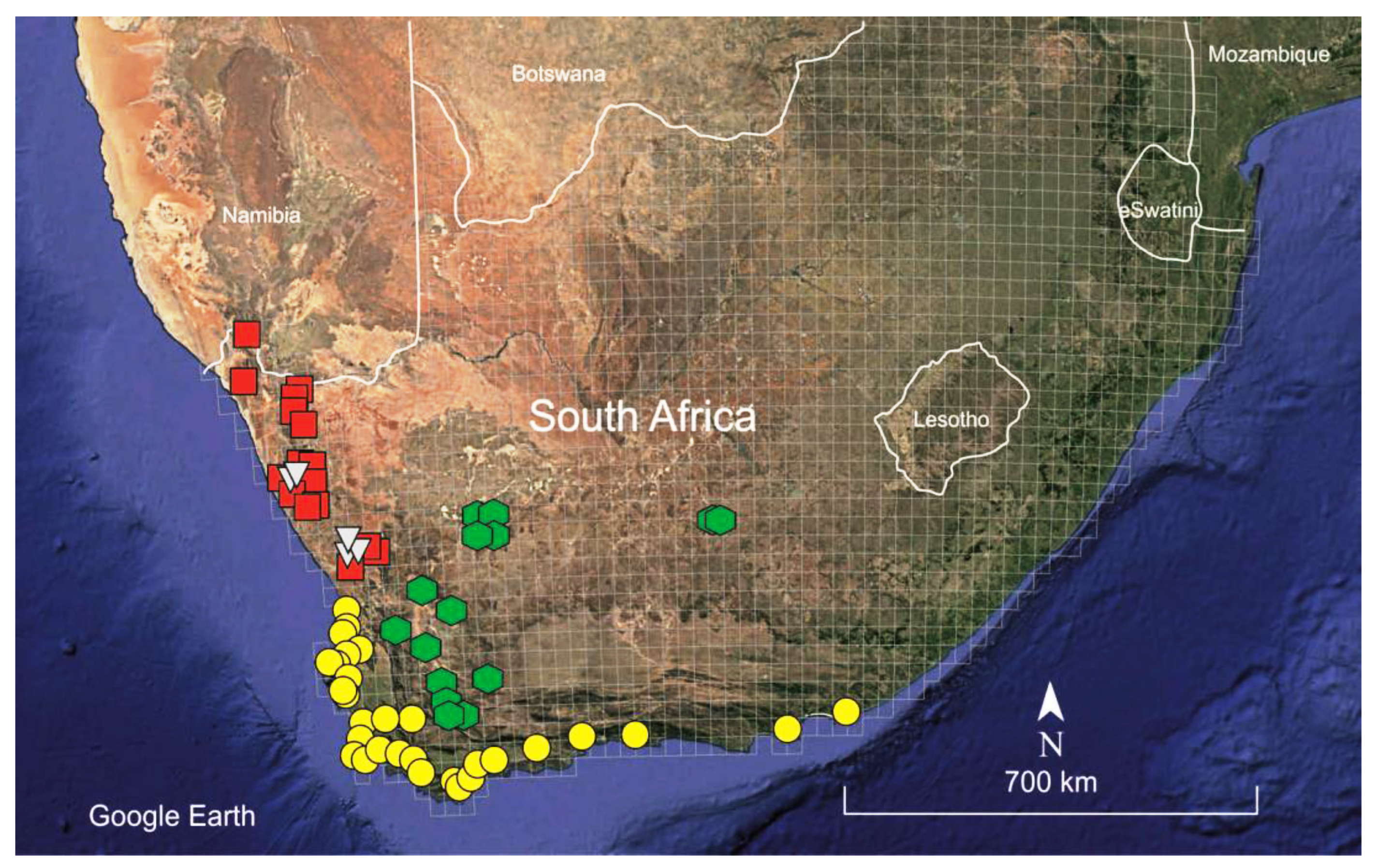
Figure 3.
Bayesian phylogenetic tree of Spergularia accessions from trnL–trnF plastid DNA sequences. The position of the South African taxa is marked in blue in Clade A, where S. hanoverensis is not full resolved. Numbers above branches indicate posterior probabilities (PP), whereas numbers below branches correspond to bootstrap percentages (BP) from both ML/MP (strict consensus) trees.
Figure 3.
Bayesian phylogenetic tree of Spergularia accessions from trnL–trnF plastid DNA sequences. The position of the South African taxa is marked in blue in Clade A, where S. hanoverensis is not full resolved. Numbers above branches indicate posterior probabilities (PP), whereas numbers below branches correspond to bootstrap percentages (BP) from both ML/MP (strict consensus) trees.
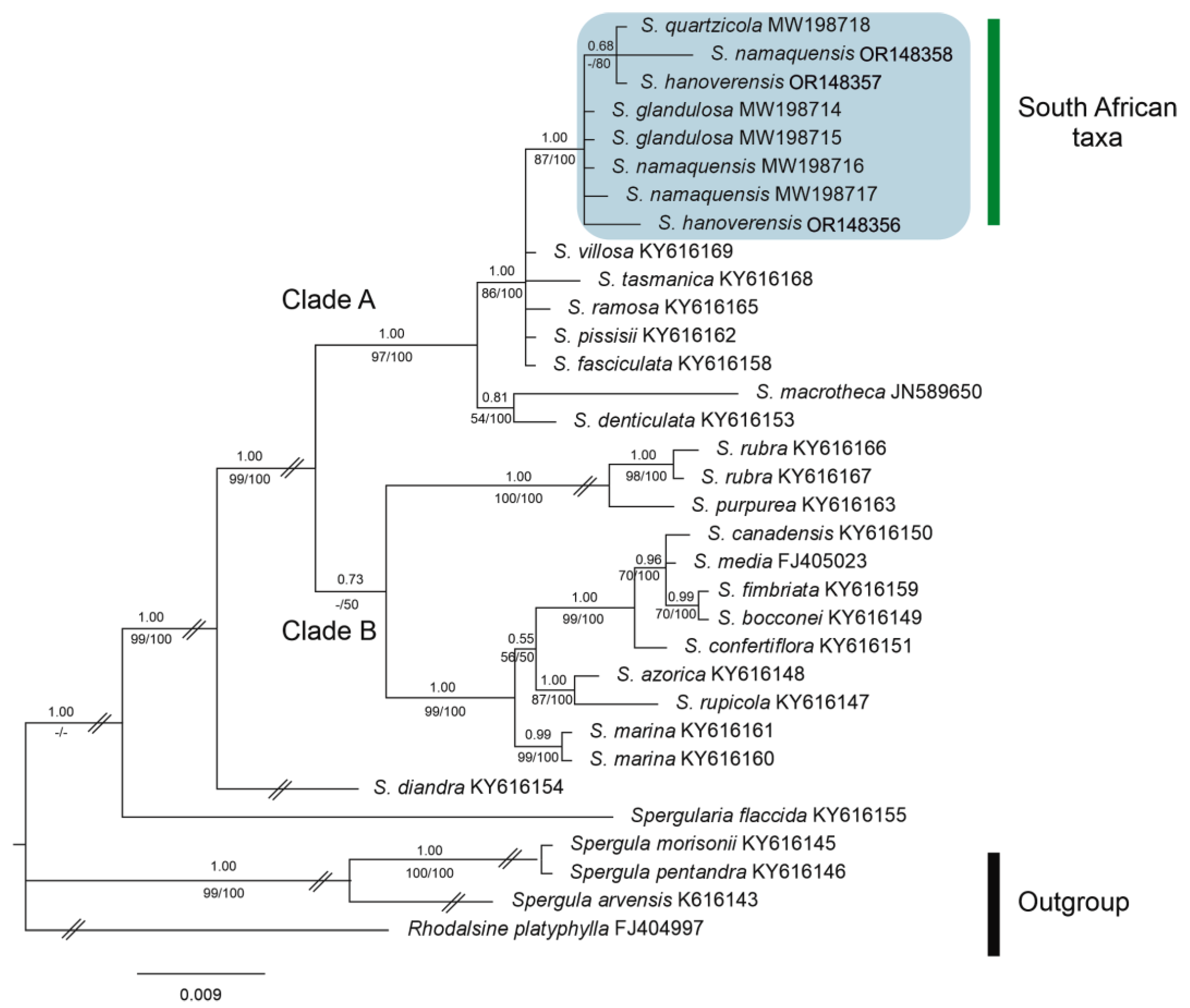
Figure 4.
Bayesian phylogenetic tree of Spergularia accessions from 5.8S-ITS2 nuclear DNA sequences. The position of the South African taxa is marked in blue in Clade A. Numbers above branches indicate posterior probabilities (PP) from the Bayesian analysis, whereas numbers below branches correspond to bootstrap percentages (BP) from both ML/MP (strict consensus) trees.
Figure 4.
Bayesian phylogenetic tree of Spergularia accessions from 5.8S-ITS2 nuclear DNA sequences. The position of the South African taxa is marked in blue in Clade A. Numbers above branches indicate posterior probabilities (PP) from the Bayesian analysis, whereas numbers below branches correspond to bootstrap percentages (BP) from both ML/MP (strict consensus) trees.
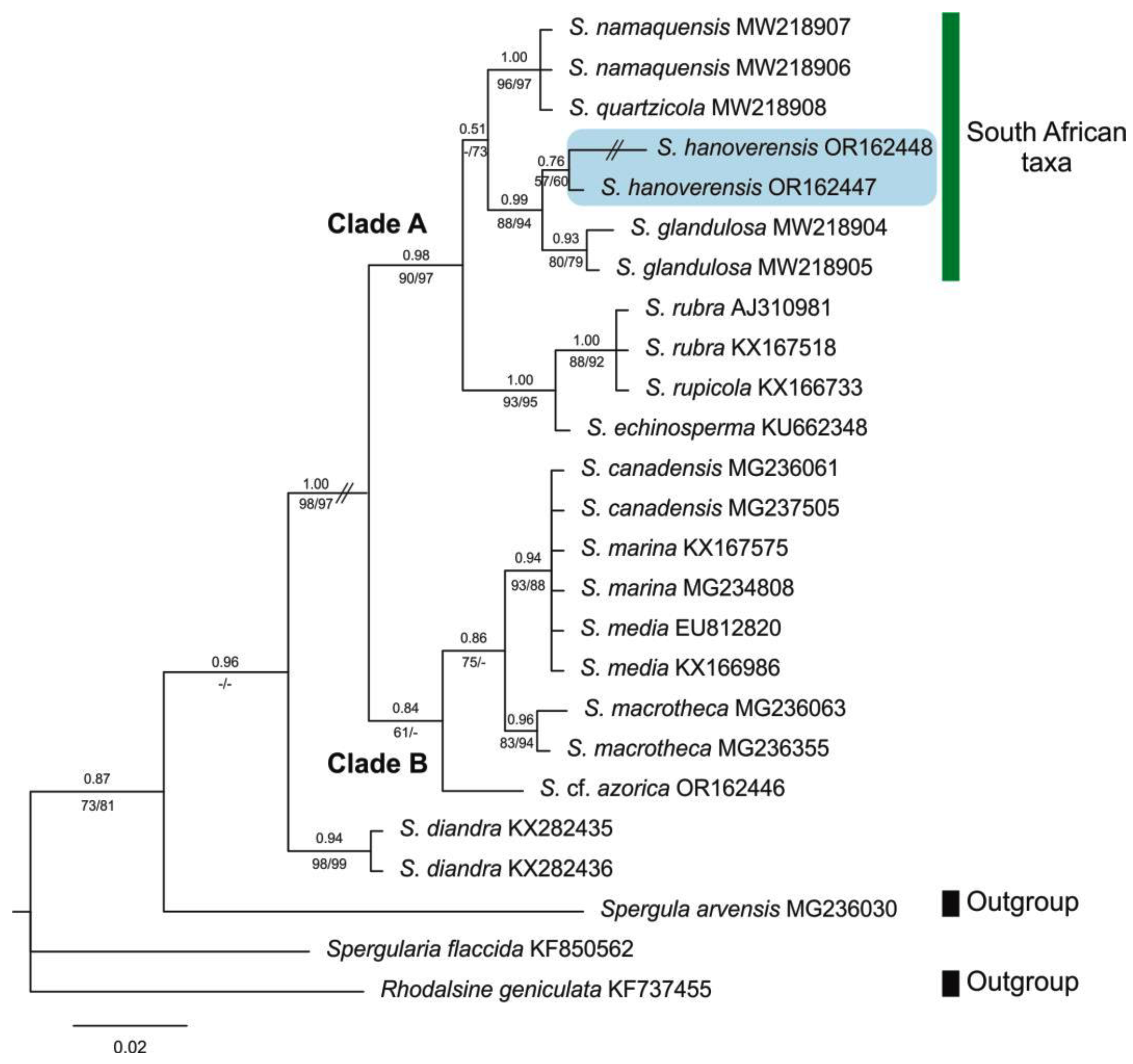
Figure 5.
Seed morphology variation in Spergularia hanoverensis. (a–c) Northern Cape: Rietfontein: dimorphic large seeds from a single capsule (PRE0694503); (d) Northern Cape: Williston, Farm Grootfontein: large unwinged seed (PRE0791338); (e) Northern Cape: Hanover: small unwinged seed (PRE0405795); (f–g) Northern Cape: Calvinia: dimorphic small seeds from a single capsule (PRE0405924); (h) Rietfontein: testa ornamentation of winged seed (PRE0694503); (i) Northern Cape: Calvinia: testa ornamentation of unwinged seed (PRE0405924); (j) Northern Cape: Hanover: detail of testa papillae (PRE0405795); (k) Northern Cape: Calvinia: testa ornamentation of winged seed (PRE0405924). Scale bars = 100 μm.
Figure 5.
Seed morphology variation in Spergularia hanoverensis. (a–c) Northern Cape: Rietfontein: dimorphic large seeds from a single capsule (PRE0694503); (d) Northern Cape: Williston, Farm Grootfontein: large unwinged seed (PRE0791338); (e) Northern Cape: Hanover: small unwinged seed (PRE0405795); (f–g) Northern Cape: Calvinia: dimorphic small seeds from a single capsule (PRE0405924); (h) Rietfontein: testa ornamentation of winged seed (PRE0694503); (i) Northern Cape: Calvinia: testa ornamentation of unwinged seed (PRE0405924); (j) Northern Cape: Hanover: detail of testa papillae (PRE0405795); (k) Northern Cape: Calvinia: testa ornamentation of winged seed (PRE0405924). Scale bars = 100 μm.

Table 2.
List of Spergularia accessions generated specifically for this study. See Alonso et al. [8] for additional details on the rest of accessions used in the analyses.
Table 2.
List of Spergularia accessions generated specifically for this study. See Alonso et al. [8] for additional details on the rest of accessions used in the analyses.
| Taxon | Provenance (herbarium voucher) | Source | GenBank accession code | |
|---|---|---|---|---|
| trnL-trnF | 5.8S-ITS2 | |||
| S. cf. azorica Lebel | Spain: Canary Isl., Tenerife (ABH79973) | This paper | ||
| S. hanoverensis E.Simon ex M.Á.Alonso et al. | South Africa: Zoekop Farm (ABH83276) | This paper | ||
| South Africa: Karreekop Farm (ABH83288) | This paper | |||
| S. namaquensis Schltr. Ex M.Á.Alonso et al. | South Africa: Skoverfontein (ABH83197) | This paper | ||
Table 3.
Comparison of morphological characters of Spergularia hanoverensis with its perennial relatives of the “South African taxa” group.
Table 3.
Comparison of morphological characters of Spergularia hanoverensis with its perennial relatives of the “South African taxa” group.
| S. glandulosa | S. namaquensis | S. media | S. quartzicola | S. hanoverensis | |
|---|---|---|---|---|---|
| Height (cm) | up to 15 | up to 20 | 5–50 (–65) | up to 30 | up to 30 |
| Habit | subshrub | subshrub | perennial herbs, weakly lignified at base | subshrub | subshrub |
| Stems | caespitose, strongly nodose, procumbent to prostrate | erect, nodose, with ascending branches | diffuse, procumbent to prostrate | erect, nodose below, with subfastigiate branches | compact, slightly nodose, with ascending branches |
| Young branches indumentum | densely covered all over with gladuliferous, pluricellular hairs | densely covered all over with glanduliferous, pluricellular hairs | glabrous | glabrous (occasionally sparsely glanduliferous when young) | glabrous |
| Leaves (mm) | 8–20 × 0.5–1 | 8–25 × 0.8–1.2 | 10–35 × 0.3–2.5 | 8–45 × 0.5–1.2 | 4–10 (–1.7) × 0.5–0.7 |
| Indumentum of leaves | covered with glanduliferous hairs, occasionally glabrescent | covered with glanduliferous hairs, occasionally glabrescent | glabrous | glabrous | glabrous |
| Leaf mucro (mm) | 0.2–0.4 | 0.4–0.5 | 0–0.2 | 0.2–0.5 | 0.5-1.0 |
| Stipules (mm) | 3–6 × 3–4 | 2–4 × 2–4 | 2.3–2.5 × 1.3–2 | 3–5 × 2–2.6 | 4–6 × 1–2 |
| Stipules shape and connation | triangular to broadly triangular, those of young stems fused up to 1/3 of their length | triangular to broadly triangular, those of young stems fused up to 1/3 of their length | triangular, not acuminate, those of young stems fused up to 1/2 of their length | narrowly triangular, long acuminate, those of young stems fused up to 1/3 of their length | triangular-acuminate, those of young stems fused up to 1/3 of their length |
| Stipules indumentum | whitish-scarious, with glanduliferous hairs in the basal part | whitish-scarious, with glanduliferous hairs in the basal part | whitish-scarious, glabrous | whitish-scarious, glabrous | whitish-scarious, glabrous |
| Floral bracts/ Stipules | bracts longer to slightly longer than stipules | bracts slightly longer than stipules | bracts slightly longer than stipules | bracts longer than stipules | bracts longer than stipules |
| Inflorescence | monochasial to dichasial cyme, narrowly branched | dichasial cyme, broadly branched | dichasial cyme, broadly branched | dichasial cyme, broadly branched. rarely monochasial | dichasial cyme, broadly branched |
| Bract length (mm) | (2–)6–10 | (2–)5–9 | 1.7–2 | (1.5–)3–7 | (1–)1.5–2.5 |
| Indumentum of bracts | glandular-pubescent | glandular-pubescent | glabrescent to glandular-pubescent | glandular-pubescent | glandular-pubescent |
| Sepals (mm) | 2–6 × 2–3, with narrowly scarious margins, erect-patent at anthesis | 5–7.5 × 4–5, with broadly scarious margins, patent to slightly deflexed at anthesis | 4–6 × 2, with broadly scarious margins, erect-patent at anthesis | 4.6–6 × 1.5–3, with narrowly scarious margins, strongly deflexed at anthesis | 2–3 × 1–1.5 with narrowly scarious margins, patent to slightly deflexed at anthesis |
| Indumentum of sepals | glandular-pubescent | glandular-pubescent | glandular-pubescent | glandular-pubescent | glandular-pubescent |
| Petal colour | white | white | white or sometimes pink at apex | white | white |
| Sepals/petals | slightly shorter than to equalling sepals | up to 1.5 times longer than sepals | up to 1.2 longer than sepals | up to 1.5 times longer than sepals | slightly shorter than to equalling sepals |
| Petals (mm) | 2–5 × 1.5–3 | 6–9 × 4–5 | 4–5 × 1.8–2 | 6–8 × 4–5 | 1.8–2.8 × 1–1.2 |
| Stamens | 10, widened at bade | 10, not widened at base | (7–9)10, widened at base | 10, dissimilar (5 widened at base) | 7–8, slightly widened at the base |
| Stamens/petals | shorter than petals | shorter than petals | shorter than petals | shorter than petals | shorter than petals |
| Anther | 0.5–0.8 | 0.6–1 | 0.9–1 | 0.8–1 | 0.3–0.4 |
| Styles | 3, free from base | 3, fused in column to half or beyond | 3, free from base | 3, fused in column to half or beyond | 3, free from base |
| Style length (mm) | 0.5–0.7 | 1–1.1 | 0.5–0.6 | 1.5–2 | 1 |
| Capsule (mm) | 4–6.5 × 1.5–3.5 | 6–9 × 3–4 | (5–)6–9 × (3.5–)4–6 | 5.5–6.5 × 3–3.5 | 3–4 × 2.5–3.5 |
| Capsule/sepals | slightly longer than sepals | equalling to r slightly longer than sepals | 1/3 longer than sepals | equalling to slightly longer than sepals | slightly longer than sepals |
| Seed | winged, discolour | winged, discolour | winged, discolour | winged, discolour | winged, discolour, to unwinged |
| Seed colour | scarious whitish wing and dull brown to blackish disk | scarious whitish wing and blackish disk | scarious whitish wing and blackish disk | scarious whitish wing and blackish disk | scarious whitish wing and blackish-brown and matte |
| Size of seed (mm) | 0.9–1.3 × 1.2–1.3 | 1.1–1.3 × 1.2–1.4 | 1.2–1.5 × 1.3–1.6 | 1.1–1.2 × 1.1–1.2 | 0.5–0.75 × 0.3–0.6 and 0.7–1.1(–1.4) × 0.6–1.2(–1.4) |
| Seed disk (mm) | 0.6–0.7 × 0.6–0.9, loosely covered with conical tubercles | 0.6–0.7 × 0.6–0.8, smooth or with conical tubercles only on edges | 0.6–0.8 × 0.5–0.7 smooth | 0.6–0.7 × 0.6–0.7, smooth or with granular protuberances, mostly on edges | 0.5–0.9 × 0.4–0.9 mm, densely covered with globose, stalked papillae |
| Width of seed wing (mm) | 0.3–0.4 | 0.4–0.6 | 0.3–0.5 | 0.5–0.6 | 0–0.5 |
| Wing edge | entire to slightly eroded | entire to slightly eroded | entire to slightly eroded | deeply and irregularly lacerate | eroded, when present |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



