
Submitted:
22 May 2023
Posted:
23 May 2023
You are already at the latest version
Abstract
Skin pigmentation ensures efficient photoprotection and relies on the pigment melanin, which is produced by epidermal melanocytes and transferred to surrounding keratinocytes. While the molecular mechanisms of melanin synthesis and transport in melanocytes are now well characterized, much less is known about melanin transfer and processing within keratinocytes. Over the past few decades, distinct models have been proposed to explain how melanin transfer occurs at the cellular and molecular levels. However, this remains a debated topic as up to four different models have been proposed, with evidence presented supporting each. Here, we review the current knowledge on the regulation of melanin exocytosis, internalization, processing and polarization. Regarding the different transfer models, we discuss how these might co-exist to regulate skin pigmentation under different conditions, i.e. constitutive and facultative skin pigmentation or physiological and pathological conditions. Moreover, we discuss recent evidence that sheds light on the regulation of melanin exocytosis by melanocytes and internalization by keratinocytes, as well as how melanin is stored within these cells in a compartment that we proposed to be named melanokerasome. Finally, we review the state of the art on the molecular mechanisms that lead to melanokerasome positioning above the nuclei of keratinocytes, forming supranuclear caps that shield the nuclear DNA from UV radiation. Thus, we provide a comprehensive overview of the current knowledge on the molecular mechanisms regulating skin pigmentation, from melanin exocytosis by melanocytes and internalization by keratinocytes, to processing and polarization within keratinocytes. A better knowledge of these molecular mechanisms will clarify long lasting questions in the field that are crucial for the understanding of skin pigmentation and can shed light on fundamental aspects of organelle biology. Ultimately, this knowledge can lead to novel therapeutic strategies to treat hypo- or hyper-pigmentation disorders, which have a high socio-economic burden to patients and healthcare systems worldwide, as well as having cosmetic applications.
Keywords:
Melanin
; Melanocyte
; Keratinocyte
; Melanosome
; Melanocore
; Melanokerasome
; Mlelanin transfer
; Melanin processing
; Melanin polarization
Introduction
The skin is the largest organ of the body and plays fundamental roles in water balance, regulation of body temperature, and protection against foreign agents and ultraviolet radiation (UVr)[1]. The upper layer of the skin – the epidermis – is mainly comprised of keratinocytes, which account for ~ 95% of the cells in this layer. Keratinocytes stratify into five different strata, according to their differentiation status: stratum basale, stratum spinosum, stratum granulosum, stratum lucidum (only present in thick skin) and stratum corneum[2]. The epidermis also contains melanocytes, which are specialized cells derived from the neural crest during embryonic development. Melanocytes are dispersed throughout the stratum basale and can extend dendrites to contact up to 40 keratinocytes, forming “epidermal-melanin units” (Figure 1A)[3,4]. The function of melanocytes is to synthesize the pigment melanin, subsequently transferring it to keratinocytes[5]. There are two types of melanin: black/brown eumelanin and yellow/red pheomelanin[6,7]. These are synthesized in melanocytes within melanosomes, which are lysosome-related organelles (LROs), as they are endosomal-derived, contain lysosomal markers, and are acidic in early stages of their biogenesis. Melanosomes display four distinct stages of maturation: In stages I and II, they are non-pigmented, initially resembling multivesicular endosomes. While pheomelanosomes maintain a spherical shape, eumelanosomes acquire an elliptic shape due to internal fibril nucleation of the premelanosomal protein (PMEL)[8]. Melanogenic enzymes required for eumelanin synthesis, including tyrosinase and tyrosinase-related proteins (TYRP1/2), are recruited to stage III melanosomes and melanin starts to progressively deposit onto PMEL fibrils, until melanosomes achieve full pigmentation (stage IV). Mature melanosomes accumulate at melanocyte dendrites and are then transferred to keratinocytes. After being internalized by keratinocytes, melanin is processed, i.e., trafficked before accumulating in the supranuclear area, where it exerts its function of protecting the DNA of these cells from UVr. Thus, skin pigmentation results from several processes that involve melanin synthesis in melanocytes, transfer from melanocytes to keratinocytes, and processing within keratinocytes. The molecular machinery involved in melanin synthesis and transport within melanocytes is now well characterized, but the mechanisms of melanin transfer remain controversial and poorly characterized. Moreover, only a few studies have addressed how melanin processing occurs within keratinocytes and much remains to be uncovered. Here, we discuss the current knowledge and research priorities on the mechanisms of melanin transfer and processing. This knowledge is fundamental to understand how skin pigmentation is sustained and can provide therapeutic strategies for pigmentary disorders, as well as having cosmetic applications.
Melanin transfer models
Melanin must be transferred to keratinocytes to exert its photoprotective function in these cells. The process by which melanin transfer occurs has been a controversial topic since as far back as the 1960’s, when melanin was first observed being transferred from a melanocyte to an epithelial cell (keratinocyte)[9]. As many as four models have been proposed to explain how melanin is transferred from melanocytes to keratinocytes, even though some could be variations of the same: (I) Cytophagocytosis of melanocyte dendrite tips, containing melanosomes, by keratinocytes; (II) Membrane fusion between keratinocytes and melanocytes and direct transfer of melanosomes; (III) Shedding of melanosome-laden globules by melanocytes and subsequent phagocytosis by keratinocytes; and (IV) Exocytosis of the melanin core by melanocytes, followed by phagocytosis by keratinocytes (Figure 2). These are subsequently summarized:
- 1.
- Cytophagocytosis of melanocyte dendrites by keratinocytes.
This transfer mode was first proposed after observations made by electron microscopy (EM) of melanocyte dendrite tips within keratinocytes in vitro [10]. The process can be divided in four steps: Firstly, a melanocyte dendrite tip comes in contact with the keratinocyte plasma membrane and is subsequently engulfed by it; secondly, the melanocyte dendrite tip is pinched off, leading to the formation of a cytoplasmic vesicle filled with melanosomes; thirdly, the compartment where melanin resides, which is theoretically limited by three membranes of different origins – the innermost one corresponding to the membrane of the melanosome and the outer ones to the plasma membranes of the melanocyte and the keratinocyte – fuses with lysosomes to form a phagolysosome. This leads to the degradation of the luminal membranes[11]. Lastly, the phagolysosome fragments into smaller vesicles containing aggregates or single melanin granules dispersed in the cytoplasm[12].
- 2.
- Fusion of melanocyte and keratinocyte membranes.
This model postulates that the plasma membranes of the melanocyte and keratinocyte fuse, allowing the formation of a conduit (e.g., filopodia or nanotubes) between the cytoplasm of the neighboring cells, through which melanosomes are transported[13]. As in the cytophagocytosis model, keratinocytes receive membrane-bound melanosomes. However, melanin within keratinocytes was shown by us and the group of Graça Raposo to be devoid of melanosome markers[14,15]. A possible explanation for this discrepancy is that the melanosome membrane is degraded inside keratinocytes, by the endolysosomal system. It should be noted that this model and the cytophagocytosis one could represent variations of the same. Indeed, the group of Desmond Tobin has proposed a filopodia-cytophagocytosis model, in which melanocytes extend filopodia that bind to the keratinocyte plasma membrane, and are then pulled and subsequently phagocytosed by the keratinocyte[16,17].
- 3.
- Shedding of melanosome-laden globules.
This model proposes that the blebbing of the melanocyte membrane leads to the formation and release of globules (1-3 µm in size) filled with melanosomes, which are then phagocytosed by keratinocytes[18,19]. Recently, other studies showed that globules filled with melanosomes are secreted from primary human melanocyte dendrites to the cell culture medium[20]. These melanosome-laden globules, when isolated from the medium and incubated with primary human keratinocytes, are internalized and processed in a similar manner as postulated by the cytophagocytosis model[21]. Indeed, according to this model, upon internalization, melanin is surrounded by three membranes. After processing, single membrane vesicles containing melanin are dispersed inside keratinocytes[21].
- 4.
- Coupled exo/phagocytosis of the melanin core.
This transfer model predicts that melanosomes fuse with the melanocyte plasma membrane, which leads to the exocytosis of the naked melanin core, termed melanocore, to the extracellular space and its subsequent internalization by keratinocytes. Consequently, melanocores become surrounded by a single membrane derived from the keratinocyte plasma membrane upon internalization. The first evidence supporting this model came with the observation of extracellular melanin in human hair and skin, which was hypothesized to be internalized by keratinocytes in aggregates or individual granules[22]. More recently, we and others provided further evidence supporting this model in which melanocores were observed in the interstitial space between melanocytes and keratinocytes[14,15]. We also showed that melanocores within keratinocytes are surrounded by a single membrane lacking the melanosome marker TYRP1, implying that the original melanosome membrane is lost during transfer[15,23].
The different models are supported by evidence obtained in various experimental systems, namely primary cells, cell lines and tissue explants from different species, including mouse, chicken, frog and human. Indeed, the differences between the models might reflect the specificities of each system/species. Importantly, the model of exo/phagocytosis is the one that collected more supporting evidence from the analysis of human skin sections and reconstructed human skin/epidermis in vitro models[15,24]. The shed globules is the other model that has been more explored. Furthermore, the four models can be essentially divided in the one that postulates the transfer of naked melanin (melanocore) and the ones that predict the transfer of membrane-bound melanosomes. Importantly, it cannot be excluded that several mechanisms of melanin transfer co-exist, even in the same organism, as an adaptation to different physiological or pathological conditions. To solve these long-lasting questions, a better understanding at the cellular and molecular levels of the pathways and regulators of melanin exocytosis from melanocytes and internalization by keratinocytes is essential.
Melanosome transport and exocytosis
Melanosome transport to the periphery of melanocytes precedes their transfer to neighboring keratinocytes[25,26,27]. Several regulators of melanosome transport on the cytoskeleton of melanocytes have been identified. The Ras-related protein in brain (Rab) 1, which mediates endoplasmic reticulum (ER)-Golgi trafficking, interacts with the SifA and kinesin-interacting protein (SKIP), as well as the anterograde microtubule-dependent motor kinesin 1, to regulate melanosome anterograde transport[28,29] (Figure 3A). Retrograde transport on microtubules is mediated by the intra-Golgi and endosomal trafficking regulator Rab36 and the product of dilute suppressor gene, melanoregulin, which form a complex with Rab-interacting lysosomal protein (RILP) and the dynein-interactor dynactin subunit 1 (DCTN1)[30,31] (Figure 3A). Similarly, the secretory protein Rab44 was reported to form a tripartite complex with dynactin and the microtubule-dependent motor dynein, to regulate melanosome retrograde transport[32,31] (Figure 3A). At the cell periphery, melanosomes are anchored to the cortical actin network and unidirectionally transported to the plasma membrane by the tripartite complex composed of the secretory protein Rab27a, the adaptor protein melanophilin, and the actin-dependent motor protein myosin Va[33,34] (Figure 3A). Despite the increasing knowledge about the molecular mechanisms that regulate melanosome positioning within melanocytes, it is not yet fully understood what is their relative importance, and of the position itself, for the efficiency of melanin transfer.
Interestingly, another actin-dependent motor – myosin X – was shown to regulate melanin transfer to keratinocytes through filopodia/nanotube assembly[16]. Moreover, N-methyl-D-aspartate receptor, the cell-cell adhesion molecule E-cadherin and the recycling endosome protein Rab17 were also implicated in melanin transfer through filopodia formation[17,35,36]. Furthermore, the plasma membrane protein caveolin-1 and cavin-1, which are required for plasma membrane invaginations called caveolae, were shown to be required for melanocyte-keratinocyte contacts, melanocyte dendricity and melanin transfer[37]. However, it is not clear in what transfer model(s) this process could be involved.
Our group showed a key role for the recycling endosome protein Rab11b and two subunits of the exocyst tethering complex (Exo70 and Sec8) in melanin exocytosis and transfer, supporting the exo/phagocytosis model[15,38] (Figure 3A). Recently, we showed that soluble factors derived from differentiated spinous/granular keratinocytes increase melanin exocytosis from melanocytes, when compared with soluble factors derived from undifferentiated keratinocytes. We also found that the secretory Rab protein Rab3a enhances melanin exocytosis and transfer to keratinocytes under stimulation from differentiated spinous/granular keratinocytes[39] (Figure 3A). Therefore, these studies support the existence of a Rab11b-dependent basal route of melanin transfer and a Rab3a-dependent one, stimulated by keratinocytes. Importantly, we observed that Rab3a-stimulated melanin exocytosis is triggered by keratinocyte-derived soluble factors, rather than extracellular vesicles[39]. Nevertheless, previous studies reported that keratinocyte-derived extracellular vesicles stimulate melanin synthesis by melanocytes[40]. Thus, we hypothesize that the Rab3a-stimulated melanin exocytosis pathway is required when melanin transfer needs to be upregulated, for instance, upon UVr exposure, also called facultative pigmentation. Additionally, the recycling endosome protein Rab11a was found to regulate melanin exocytosis upon Toll-like receptor-2 (TLR-2) activation[41]. Finally, the actin remodeling small GTPase Ras homolog gene family A (RhoA) was shown to be required for membrane blebbing and release of melanosome-laden globules in chicken embryonic skins[42].
A better understanding of the putative feedback loops linking melanin synthesis, melanosome transport and exocytosis will help clarify why different pathways can operate in different contexts to achieve the transfer of melanin. Moreover, it will be crucial to understand if the molecular regulators of melanin exocytosis can act together or in distinct pathways. Finally, future studies should explore if distinct stimuli require different proteins to regulate melanin exocytosis and if the different molecular regulators are related with different melanin transfer modes and pigmentation levels in human skin.
Melanin internalization
Efficient skin pigmentation and photoprotection relies on the capacity of keratinocytes to internalize melanin. However, the mechanisms regulating melanin internalization by keratinocytes remain poorly understood. Recent studies using mouse and human cells reported that the internalization of secreted melanin (i.e., melanocores or melanosome-laden globules) by keratinocytes is likely to be the main mode of melanin transfer in the skin[43,44]. Given the physiological size of melanosomes (0.3 - 0.5 μm in diameter), only macropinocytosis and phagocytosis are predicted to allow the internalization of such large cargo[45,46,47,48]. Although keratinocytes are not professional phagocytes, they were shown to efficiently phagocytose latex beads and bacteria[49,50]. Additionally, the stimulation of keratinocyte phagocytic capacity results in the increase of melanin transfer, strongly suggesting that phagocytosis is involved in melanin internalization by keratinocytes[44,50,51].
Both macropinocytosis and phagocytosis are highly dependent on actin cytoskeleton remodeling, which is regulated by Rho small GTPases[52]. Accordingly, melanin internalization was shown by us and others to require dynamic actin remodeling and depend on Rho GTPases[13,43,44]. Interestingly, melanocores and melanosomes require different Rho GTPases to be internalized: while melanocores are internalized in a Rac1- and Cdc42-dependent manner, melanosome internalization follow a RhoA-dependent route[43] (Figure3B). These observations are in line with a previous study showing that the internalization of melanosome-rich globules upon TLR3 stimulation is dependent on the activity of RhoA and requires Cdc42[44]. Furthermore, we observed that melanosome but not melanocore internalization is impaired by the inhibition of macropinocytosis[43]. Altogether, these observations suggest that different mechanisms are involved in melanin recognition and internalization by keratinocytes, and that melanocores are internalized by phagocytosis and melanosomes by macropinocytosis. Importantly, there is no evidence for the transfer of single melanosomes, although it represents an excellent control since the only difference between melanocores and melanosomes is the presence of a surrounding membrane on melanosomes.
The specific recognition of melanocores by a membrane receptor is likely required for their phagocytosis by keratinocytes. The best known receptor involved in melanin uptake by keratinocytes is the protease-activated receptor-2 (PAR-2)[53]. In the epidermis, PAR-2 is highly expressed in the granular layer (stratum granulosum), and involved in epidermal barrier maintenance, keratinocyte differentiation and pigmentation[54]. This seven transmembrane G-protein coupled receptor (GPCR) is activated by serine-proteases that cleave the N-terminal domain, resulting in the exposure of a new domain and receptor autoactivation[55,56]. Interestingly, the levels of PAR-2 expression correlate with skin phototype, i.e., dark skins exhibit higher expression levels and activity throughout the epidermis when compared to light skins, where PAR-2 expression and activity are concentrated at the lower layers[57]. Importantly, in vitro and in vivo PAR-2 stimulation increases keratinocyte melanin content and its activation was shown to enhance keratinocyte phagocytic capacity of melanin, beads and Escherichia coli K-12[50,53,58,59,60]. Moreover, PAR-2 activation in keratinocytes signals through Rho GTPases[43,61], which are key players in the phagocytic process, and are required for melanin internalization, as referred. Furthermore, our group showed that melanocores but not melanosomes are dependent on PAR-2 for their uptake by keratinocytes, supporting that melanocores are the form of melanin transferred to keratinocytes and the model of exo/phagocytosis[23,43] (Figure 3B). Recently, it was also shown that the cation channel transient receptor potential ankyrin 1 (TRPA1) is required for the activity of PAR-2 in regulating melanocore phagocytosis by keratinocytes[62]. Apart from PAR-2, the stimulation of other receptors, including TLR-3 and fibroblast growth factor receptor 2/keratinocyte growth factor receptor (FGFR-2/KGFR), were described to be involved in melanin phagocytosis by keratinocytes [44,51,63] (Figure 3B). Interestingly, while our group showed evidence for the specific activation of PAR-2 by melanocores, others showed that TLR-3 can regulate melanocore and melanosome internalization by keratinocytes[23,43,44]. Therefore, these reports reinforce the notion that the way melanin is presented to keratinocytes, i.e., naked melanin in melanocores or membrane-bound melanosomes, results in the activation of different receptors and possibly distinct downstream signaling pathways[23,43,44]. Of note, FGFR-2/KGFR expression inversely correlates with skin color, i.e., it is more expressed in light skin keratinocytes. How to reconcile this with the pattern of PAR-2 expression, which is higher in dark skins, would be interesting to explore in future studies to better understand the contribution of different forms of internalized melanin (membrane-bound melanosome-laden globules or melanocores), for the skin phototype. Finally, the role of UVr in the stimulation of melanin internalization is also worth mentioning. PAR-2 expression and activity, as well as FGFR-2/KGFR activity can be enhanced by exposure to UVr, increasing melanin transfer[64,65]. Moreover, a recent publication showed that the α7 nicotinic acetylcholine receptor (nAChR) responds to UVr exposure by increasing melanosome phagocytosis by keratinocytes[66].
Although phagocytosis has been widely assumed to be the internalization route followed by melanin, clear evidence for this has been lacking until recently. Since phagocytosis is a receptor-mediated process, it requires the specific recognition of the cargo to be internalized[67,68]. The evidence showing that melanocores and melanosomes require different regulators for their internalization[43], further supports that melanin is recognized differently by keratinocytes, depending on the presence or absence of a surrounding membrane. Whether melanosome-laden globules behave as melanosomes would be important to address in future studies. Additionally, the molecular determinants present on melanocores that are required for their specific recognition are not known and should be investigated, to allow the identification of the phagocytic receptor involved. Importantly, beads, which have been widely used as a control for melanin internalization, most likely lack the molecular determinants required for the specific recognition of melanin. Therefore, despite the effort to use beads with sizes similar to melanosomes, they are likely not a suitable model to study melanin internalization by keratinocytes.
Even though there is abundant evidence for the important role of PAR-2 and other receptors (e.g., FGFR2/KGFR, TLR-3) in melanin internalization, their function in this process is likely to be indirect, i.e., increasing the overall keratinocyte uptake efficiency, rather than being directly involved in melanin recognition[44,50,51,63]. In fact, GPCRs like PAR-2 were shown to recognize chemoattractants secreted by microorganisms, enhancing the phagocytic capacity through the remodeling of the actin cytoskeleton[69,70,71]. Thus, PAR-2 can be required for chemotaxis, i.e., directing keratinocytes towards melanin and facilitating its phagocytosis. Similarly, although TLRs can recognize molecular patterns on different cargoes, they are not phagocytic receptors, cooperating instead to prime phagocytosis[72,73]. Additionally, PAR-2 is highly expressed in the granular layer[54], whereas melanin transfer mostly occurs in the basal layer[74,75,76], which suggests a paracrine regulation of melanin internalization by suprabasal keratinocytes. This further implies that PAR-2 is likely not involved directly in melanin recognition, but instead in the regulation of keratinocyte phagocytic activity. Thus, how melanocores are recognized by keratinocytes to trigger their internalization through phagocytosis, remains to be addressed. Among the possible candidates are the C-type lectin receptors, especially considering the recent study showing that MelLec receptors, expressed in endothelial and myeloid cells, specifically recognize fungal melanin[77]. Interestingly, keratinocytes express the C-type lectin receptor Dectin-1, which is known to be involved in wound re-epithelialization and keratinocyte-mediated innate immune responses[78,79]. Despite the differences between fungal and mammalian melanin, the involvement of C-type lectin receptors such as Dectin-1 in human melanin recognition is worth testing. Remarkably, keratinocytes express Fc-γ receptors, as described in the context of inflammatory skin diseases such as psoriasis[80,81]. To the best of our knowledge, the involvement of Fc-γ receptors in melanin internalization by keratinocytes was not studied so far, but might be interesting to explore in future studies. Fc-γ receptors require the binding of an opsonin (e.g., antibody) to the particle internalized. This raises the question of whether melanin also requires an opsonin to be recognized and internalized by keratinocytes.
The identification of the phagocytic receptor(s) involved in melanin internalization can provide an appealing target to modulate this process and, consequently, skin color. Furthermore, the characterization of the receptor(s) and downstream signaling pathways involved in melanin internalization can also shed light on the mechanism regulating melanin processing and positioning within keratinocytes, as discussed below.
Melanin processing
Melanin processing refers to the steps that occur within keratinocytes after melanin internalization. Remarkably, the mechanisms involved remain elusive, despite keratinocytes being the melanin recipient cells, where melanin exerts a photoprotective effect, and differences in melanin processing linked to skin phototype. Indeed, while in light skins, melanin mainly accumulates within keratinocytes of the basal layer of the epidermis, in dark skins, melanin is also present in keratinocytes of the suprabasal layers[74,75,76] (Figure 1A). Importantly, it is well established that melanin organization within keratinocytes differs between phototypes [14,82,83,84,85]. In light skins, melanin is present in clusters, whereas dark skins predominantly contain single melanin granules that are usually larger, even though melanin-containing organelles display a similar size in light and dark skins[14,82,83,84,85] (Figure 1B). The mechanisms involved in the establishment of clusters or single granules are essentially unknown, except that they depend on the skin phototype of the keratinocyte and are independent on the type of melanin transferred (i.e., eumelanin vs. pheomelanin)[86].
Melanin turnover in melanocytes and keratinocytes has been suggested to be controlled by autophagy[87,88,89]. In keratinocytes, the autophagic/lysosomal activities were described to inversely correlate with skin color, i.e., higher in keratinocytes derived from light skins than in those from dark skins, independently of the keratinocyte differentiation status[87,90,91,92,93,94] (Figure 4B). Moreover, the pharmacological or genetical modulation of autophagy in distinct experimental conditions (i.e., 2D or 3D cultures and skin explants) was shown to alter keratinocyte melanin content[87,89,95,96,97] (see Table 1). Indeed, autophagy induction was found to lead to a reduction in the levels of melanin within keratinocytes, whereas autophagy inhibitors lead to an increase in these levels. This was attributed to melanin degradation vs. persistence, respectively[87,89,95,96,97]. Remarkably, the decline in autophagic activity in the epidermis with aging results in the accumulation of melanin, namely within keratinocytes, forming hyperpigmented lesions such as those seen in senile lentigo[95]. Interestingly, autophagy induction in aged skin was suggested to restore pigmentation, reinforcing the role of autophagy in melanin turnover[95].
Notwithstanding these observations, melanin is not found in double membrane organelles, i.e., autophagosomes, within keratinocytes, and melanin containing-compartments lack the classical autophagosomal protein LC3[14,98]. Indeed, melanin is found in single membrane compartments inside keratinocytes, supporting the exo/phagocytosis or the fusion of melanocyte and keratinocyte membranes transfer models[13,14,15] (Figure 4A). Despite the lack of clear evidence, it has been proposed that in the cytophagocytosis and shed globules models, the surrounding membrane(s) is(are) degraded during processing within keratinocytes[14,46,99]. For instance, it was suggested that during melanosome internalization, the melanosome membrane protein TYRP1 is quickly removed, possibly through degradation, but the mechanisms involved were not investigated[100]. This results in the formation of single membrane melanin-containing organelles within keratinocytes. The characterization of the molecular processes involved in the formation of these organelles is important to better understand the fate of the pigment in keratinocytes, and only recently this question started to be addressed. Graça Raposo’s group showed that, independently of the skin phototype, the lysosomal-associated membrane protein 1 (LAMP1) and the tetraspanin cell surface antigen 63 (CD63) – markers of late endosomes (LEs)/lysosomes – are present in the membrane of single and clustered melanocores[14] (Figure 4A). Additionally, we found that in mouse keratinocytes, melanin-containing compartments are positive for the early endocytic markers early endosome antigen 1 (EEA1) and Rab5, as well as for LAMP2 and CD63[23] (Figure 4A). Noteworthy, these experiments were performed at a fixed timepoint and, therefore, the maturation and dynamics of the compartment were not assessed. Accordingly, several Rab small GTPases, including the LE/lysosomal Rab7b, were found to localize to melanin-containing compartments inside mouse keratinocytes[100].
Moreover, we and Graça Raposo’s group showed that melanin accumulates in moderately acidic compartments (presenting low levels of Lysotracker and DAMP [3-(2,4-dinitroanilino)-3'-amino-N-methyldipropylamine] accumulation) with low degradative capacity, as shown by the low levels of dye quenched-bovine serum albumin (DQ-BSA) fluorescence, despite the abundance of these types of organelles within keratinocytes[14,23] (Figure 4A). Indeed, we reported that in cultured mouse keratinocytes, melanin is long-lived and persists for long periods[23], which suggests that melanin-containing compartments are specialized for long-term melanin storage inside keratinocytes. We termed these melanin-containing compartments inside keratinocytes melanokerasomes (MKSs)[46] to distinguish them from melanosomes in melanocytes. MKSs are likely unique melanin-storage compartments presenting low acidity and degradative capacity but maintaining characteristics of LEs/lysosomes, including the membrane markers LAMP1/2 and CD63[14,23] (Figure 4A). Given that CD63 is frequently associated with LROs in specialized cell types, and that LROs also present moderate luminal acidity and degrative capacity when compared to lysosomes, it has been suggested that MKSs could be a new type of LRO[14,101]. Although this is an interesting hypothesis that gives MKSs additional relevance as functional organelles, MKSs start as a phagosome and are thus not endosomal-derived or known to receive cargo from the Golgi. Additionally, there is no evidence that MKSs can undergo regulated exocytosis, the most notable distinguishing feature of LROs. Another possibility is that MKSs are lysosome-derived organelles or represent a dysfunctional lysosome, which has lost its degradative ability. Indeed, this could explain melanin persistence inside keratinocytes. However, it cannot be discarded that melanin is resistant to degradation (see below) . Thus, future studies should characterize MKS formation in a dynamic way, i.e., considering its spatiotemporal maturation and interaction with other organelles, as well as how they enable melanin persistence. This will help understand the biogenesis and lifecycle of these organelles specialized for melanin storage.
Another relevant question is whether melanin internalization is linked to its fate within keratinocytes. This applies not only to the influence of the transfer mode on the way melanin is processed (i.e., the transfer of melanosomes, melanosome-laden globules or melanocores), but also the contribution of the signaling pathways activated downstream of membrane receptors, such as PAR-2, FGFR-2/KGFR and TLR-3 (Figure 4A). For instance, PAR-2 was shown to decrease autophagy through the phosphatidylinositol 3-kinase (PI3K)/protein kinase B (Akt)/mammalian target of rapamycin (mTOR) pathway in kidney cells[102,103]. Moreover, melanin degradation was found to be promoted when this pathway is inhibited using the dietary flavonoid isoliquiritigenin, and, consequently, autophagy is activated[96] (Table 1). Indeed, the contribution of different melanin internalization routes for processing is unknown[45]. Interestingly, previous work from our lab showed that melanin clusters can be internalized by keratinocytes in the skin[15]. This raises several questions, including whether the clusters found in light skins are formed during internalization or after processing inside keratinocytes. It is also unclear whether melanin processing is different among phototypes and how this contributes to the distinct organization patterns observed in light vs. dark skins. In particular, the increased autophagic/lysosomal activities in light skins, when compared with dark ones, suggests the existence of distinct melanin degradation capacities by keratinocytes from different phototypes[87,90,91,92,93,94] (Figure 4B). This might also explain the detection of suprabasal melanin in dark skins, possibly resulting from decreased or delayed melanin degradation (Figure 4B). Thus, it is crucial to clarify the contribution of melanin internalization and autophagy for melanin organization within keratinocytes. It is likely that MKS formation and processing, starting with melanin internalization and culminating in the establishment of a unique storage compartment, the MKS, are determined by several factors.
Strikingly, there is contradictory evidence regarding melanin degradation within keratinocytes and it is not clear if melanin can be degraded at all. Given its protective properties, melanin should be preserved and the characteristics of the MKS, namely moderate acidity and degradative capacity, point towards that[14,23]. In fact, melanin solubility increases in alkaline conditions, whereas it remains insoluble and precipitates in acidic milieu[104,105]. Moreover, attempts to destroy melanin hydrolytically in vitro result in protein and lipid degradation but preservation of the melanin moiety [104,105]. Therefore, whether the formation of MKSs is required for melanin preservation and/or degradation within keratinocytes remains to be established. Interestingly, fungal melanin was shown to inhibit LC3-associated phagocytosis (LAP) and promote Aspergillus fumigatus pathogenicity[106,107]. Moreover, we proposed that melanin shares with microorganisms the capacity to subvert intracellular trafficking pathways to resist degradation[108]. Hence, the potential role of LAP in melanin processing should be tested. Although melanin is long-lived inside keratinocytes in vitro, it localizes mostly to the basal layer and its accumulation throughout the epidermis is only evident in darker phototypes, where suprabasal keratinocytes retain degraded melanin fragments known as “melanin dust”. Of note, the detection of melanin in the corneal layer (i.e., “melanin dust”), observed when using Fontana Masson staining, can hardly be found by EM, and is therefore attributed to an artifact of this staining[23,76,86,109,110]. This raises once more the question of whether melanin can be degraded during keratinocyte differentiation and if the right tools are available to perform this analysis accurately[110]. Recently, melanin accumulation in the basal layer has been attributed to the asymmetric division of keratinocytes, with basal keratinocytes retaining the pigment[98]. Importantly, the same study suggested that in stress conditions (e.g., regenerating skin), the symmetric division is prioritized, leading to broader melanin distribution between the daughter and progenitor keratinocytes[98]. Therefore, if melanin concentration in the basal layer results from the absence of degradation, asymmetric cell division, or both, should be evaluated. It should also be noted that melanin processing was studied using different models, including human skin explants, primary human keratinocytes and mouse/human keratinocyte cell lines. Thus, despite the relevant observations made, it is urgent to develop new strategies to study the dynamics of melanin processing in more physiologically relevant models. Moreover, only the combination of different approaches will allow the reliable analysis of the fate of melanin inside keratinocytes.
Melanin polarization
Melanin processing ends with its polarization to form supranuclear caps or “parasols” that protect the genetic material present in the nuclei of keratinocytes against the genotoxic aggressions caused by UVr exposure[111] (Figure 5). Indeed, previous studies showed a significant decrease in the formation of both cyclobutane pyrimidine dimers (CPDs) and (6–4) photoproducts (6–4PP) in cells with supranuclear melanin caps, comparing with cells without these[112]. Although the importance of the supranuclear melanin caps in the protection against UVr has been clearly established, the mechanisms behind melanin polarization in the skin remain to be uncovered. The formation of supra-nuclear melanin caps has been suggested to be stimulated by UVr[113]. Indeed, it was recently reported that UVA radiation induces melanin polarization, through the G-protein-coupled receptor opsin 3[114]. Nevertheless, our results suggest that supra-nuclear melanin caps can form in reconstructed human epidermises without UVr exposure[24]. Therefore, future studies should determine what drives melanin polarization and supra-nuclear cap formation. Interestingly, we have preliminary unpublished data suggesting that keratinocyte stratification, which involves proliferation, differentiation and migration can lead to melanin organization in supra-nuclear caps, akin of organelle polarization towards the leading edge of migrating cells[115].
Regarding the molecular players involved in melanin polarization, prior studies have shown that the intermediate chain of cytoplasmic dynein co-localizes with melanin both in cultured human keratinocytes and human skin explants[116]. The same study also reported that the treatment with anti-sense DNA for cytoplasmic dynein heavy chain leads to a significant dispersion of perinuclear melanin in human keratinocytes. Subsequent studies demonstrated that the silencing of dynactin p150Glued subunit impairs the perinuclear clustering of microspheres in human keratinocytes[117], which suggests that melanin polarization is dependent on the cytoplasmatic dynein-dynactin motor complex (Figure 5). These results were further corroborated by recent studies, which reported a significant impairment in melanin polarization upon the disruption of the actin cytoskeleton and the microtubule network[118]. The same study also reported the co-localization between melanin, the centrosome and nuclear satellites at the apical region of stratified keratinocytes, suggesting a potential role for centrosome-related proteins in the regulation of melanin polarization. Thus, very little is still known about the molecular machinery involved in melanin polarization within keratinocytes. In particular, how the retrograde dynein/dynactin-dependent transport is regulated and if the same machinery used by LEs/lysosomes is also employed by MKSs remains to be determined. Given the association of Rab7b with MKSs[100] and the role that Rab7 plays in retrograde transport of LEs/lysosomes[119], it should be confirmed if Rab7 is also involved in melanin polarization.
Conclusions and future perspectives
In contrast with the mechanisms of melanin synthesis, melanosome biogenesis, maturation and transport, those of melanin exocytosis, transfer and processing are much less understood. Indeed, one of the topics most debated in the field remains that of melanin transfer between melanocytes and keratinocytes. So far, this issue has been studied mainly in cultured primary cells and cell lines, including from melanoma and diverse animal models. However, these present clear limitations for the study of pigmentation, as they cannot fully recapitulate the complexity of the human skin architecture and crosstalk that occurs in vivo. Also, they can present changes in the mechanisms involved due to their transformation. Tissue explants have also been used, but despite recapitulating the physiology of skin pigmentation, they suffer changes when in culture. The advent of reconstructed pigmented skin/epidermises and, more recently, skin-on-a-chip models[120] has brought the possibility of manipulating melanocytes and/or keratinocytes and understand the mechanism of skin pigmentation in a setting very close to the physiological one. Therefore, these models should be more explored to determine if distinct melanin transfer modes can occur in constitutive and facultative pigmentation, as well as in pathological conditions.
Regarding melanin exocytosis from melanocytes, several molecules have now been identified to regulate this process. However, we have just started to unveil the molecular mechanisms involved in basal and stimulated pathways. Thus, the receptors, signals and molecules that can trigger stimulated melanin release should be addressed in future studies. Moreover, whether these signaling pathways and molecular regulators are the same or not under physiological (i.e., basal or facultative pigmentation) or pathological conditions (i.e., pigmentary disorders) remains to be determined. Undoubtedly, understanding better melanocyte-keratinocyte crosstalk mediated by both soluble factors, extracellular vesicles and cell-cell contacts will be a key to solve this riddle. Indeed, melanocores were found very close to melanocyte and keratinocyte membranes in human skin explants[15,24], indicative of a tight connection between these two cell types, albeit without membrane fusion. This architecture resembles the well-known immunological and neuronal synapses[121,122], suggesting the existence of a pigmentary synapse through which keratinocytes and melanocytes can communicate to control the production and transfer of melanin in the epidermis. More studies are needed to characterize this putative pigmentary synapse.
There is now evidence that melanocores are internalized by keratinocytes through phagocytosis but the phagocytic receptor(s) is(are) unknown. Therefore, the identification of the molecular determinants present in melanocores and the receptor(s) that specifically recognizes them will be crucial to clarify the transfer pathway(s) at the molecular level. Another fundamental question that remains to be addressed is whether the internalization route and downstream signaling pathways affect the fate of melanin, regarding its processing within keratinocytes. The mechanism(s) behind the differences in melanin organization inside keratinocytes from distinct phototypes is also an intriguing question that remains to be solved[14,82,83,84,85]. Along this, the understanding of how MKS, which are unique melanin-storage organelles, are established and their further characterization will be essential to unveil the mechanisms involved in fine-tuning melanin persistence within keratinocytes. Moreover, this can provide new insights into the adaptation of intracellular organelles to specialized functions. Unsolved questions also include the mechanisms through which autophagy regulates melanin turnover within keratinocytes, despite several studies showing that the modulation of this pathway alters the melanin content of keratinocytes[87,88,89]. Importantly, it must be determined if melanin can be degraded at all and if the turnover involves changes in melanin organization, i.e., single melanin granules in dark phototypes or melanin clusters more characteristic of light phototypes. These studies will have a great impact on the characterization of melanin dispersion throughout the epidermis in different phototypes, as well as the accumulation of melanin within keratinocytes in conditions such as post-inflammatory hyperpigmentation and senile lentigo[123]. Finally, it should be determined what is(are) the cue(s) for the polarization of melanin in supranuclear caps and how are these maintained throughout the life span of keratinocytes. The answers to these questions will shed light on fundamental mechanisms that are essential for skin pigmentation that have remained elusive until now, and will provide novel targets to modulate skin pigmentation to treat pigmentary disorders, as well as for cosmetic purposes.
Author Contributions
LBL, LCC, JC and MVN performed the literature search. All the authors were involved in ideation, manuscript drafting and revision.
Acknowledgments
We would like to thank Dr. Michael J. Hall for critical reading of the manuscript. This study was supported by Fundação para a Ciência e a Tecnologia (Portugal) through grant PTDC/BIA-CEL/29765/2017 and PhD fellowships to LBL, LCC, MVN and JC (SFRH/BD/131938/2017, 2020.08812.BD, PD/BD/137442/2018 and PD/BD/136905/2018, respectively). This work was also supported by iNOVA4Health – UIDB/04462/2020 and UIDP/04462/2020 and by the Associated Laboratory LS4FUTURE (LA/P/0087/2020), two programs financially supported by Fundação para a Ciência e a Tecnologia/Ministério da Ciência, Tecnologia e Ensino Superior (Portugal).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Grice EA, Segre JA. The skin microbiome. Nat Rev Microbiol 2011;9:244–253. [CrossRef]
- Solanas G, Benitah SA. Regenerating the skin: a task for the heterogeneous stem cell pool and surrounding niche. Nat Rev Mol Cell Biol 2013 1411 2013;14:737–748. [CrossRef]
- Cox NH. Fitzpatrick’s Dermatology in General Medicine. Br J Dermatol 2004;150:794–794. [CrossRef]
- Lin JY, Fisher DE. Melanocyte biology and skin pigmentation. Nature 2007;445:843–850. [CrossRef]
- Wasmeier, C.; Hume, A. N.; Bolasco, G. and Seabra MC, Wasmeier C, Hume AN, Bolasco G, Seabra MC, Wasmeier, C.; Hume, A. N.; Bolasco, G. and Seabra MC. Melanosomes at a glance. J Cell Sci 2008;121:3995–3999. [CrossRef]
- Ito S, Wakamatsu K. Chemistry of mixed melanogenesis--pivotal roles of dopaquinone. Photochem Photobiol 2008;84:582–592. [CrossRef]
- Ito S, Wakamatsu K. Human hair melanins: what we have learned and have not learned from mouse coat color pigmentation. Pigment Cell Melanoma Res 2011;24:63–74. [CrossRef]
- Raposo G, Tenza D, Murphy DM, Berson JF, Marks MS. Distinct protein sorting and localization to premelanosomes, melanosomes, and lysosomes in pigmented melanocytic cells. J Cell Biol 2001;152:809–823. [CrossRef]
- CRUICKSHANK CN, HARCOURT SA. PIGMENT DONATION IN VITRO. J Invest Dermatol 1964;42:183–184. [CrossRef]
- Okazaki K, Uzuka M, Morikawa F, Toda K, Seiji M. Transfer mechanism of melanosomes in epidermal cell culture. J Invest Dermatol 1976;67:541–547. [CrossRef]
- Wolff K. Melanocyte-keratinocyte interactions in vivo: the fate of melanosomes. Yale J Biol Med 1973;46:384.
- YAMAMOTO O, BHAWAN J. Three modes of melanosome transfers in Caucasian facial skin: hypothesis based on an ultrastructural study. Pigment Cell Res 1994;7:158–169. [CrossRef]
- Scott G, Leopardi S, Printup S, Madden BC. Filopodia are conduits for melanosome transfer to keratinocytes. J Cell Sci 2002;115:1441–1451. [CrossRef]
- Hurbain I, Romao M, Sextius P, Bourreau E, Marchal C, Bernerd F, et al. Melanosome Distribution in Keratinocytes in Different Skin Types: Melanosome Clusters Are Not Degradative Organelles. J Invest Dermatol 2018;138:647–656. [CrossRef]
- Tarafder AK, Bolasco G, Correia MS, Pereira FJCC, Iannone L, Hume AN, et al. Rab11b mediates melanin transfer between donor melanocytes and acceptor keratinocytes via coupled exo/endocytosis. J Invest Dermatol 2014;134:1056–1066. [CrossRef]
- Singh SK, Kurfurst R, Nizard C, Schnebert S, Perrier E, Tobin DJ. Melanin transfer in human skin cells is mediated by filopodia - A model for homotypic and heterotypic lysosome-related organelle transfer. FASEB J 2010;24:3756–3769. [CrossRef]
- Singh SK, Baker R, Sikkink SK, Nizard C, Schnebert S, Kurfurst R, et al. E-cadherin mediates ultraviolet radiation- and calcium-induced melanin transfer in human skin cells. Exp Dermatol 2017;26:1125–1133. [CrossRef]
- Cerdan D, Redziniak G, Bourgeois CA, Monsigny M, Kieda C. C32 human melanoma cell endogenous lectins: characterization and implication in vesicle-mediated melanin transfer to keratinocytes. Exp Cell Res 1992;203:164–173. [CrossRef]
- Aspengren S, Hedberg D, Wallin M. Studies of pigment transfer between Xenopus laevis melanophores and fibroblasts in vitro and in vivo. Pigment Cell Res 2006;19:136–145. [CrossRef]
- Ando H, Niki Y, Yoshida M, Ito M, Akiyama K, Kim J-H, et al. Involvement of pigment globules containing multiple melanosomes in the transfer of melanosomes from melanocytes to keratinocytes. Cell Logist 2011;1:12–20. [CrossRef]
- Ando H, Niki Y, Ito M, Akiyama K, Matsui MS, Yarosh DB, et al. Melanosomes are transferred from melanocytes to keratinocytes through the processes of packaging, release, uptake, and dispersion. J Invest Dermatol 2012;132:1222–1229. [CrossRef]
- Swift JA. TRANSFER OF MELANIN GRANULES FROM MELANOCYTES TO THE CORTICAL CELLS OF HUMAN HAIR. Nature 1964;203:976–977. [CrossRef]
- Correia MS, Moreiras H, Pereira FJCC, Neto M V., Festas TC, Tarafder AK, et al. Melanin Transferred to Keratinocytes Resides in Nondegradative Endocytic Compartments. J Invest Dermatol 2018;138:637–646. [CrossRef]
- Hall MJ, Lopes-Ventura S, Neto M V., Charneca J, Zoio P, Seabra MC, et al. Reconstructed human pigmented skin/epidermis models achieve epidermal pigmentation through melanocore transfer. Pigment Cell Melanoma Res 2022. [CrossRef]
- Jimenez M, Kameyama K, Maloy WL, Tomita Y, Hearing VJ. Mammalian tyrosinase: biosynthesis, processing, and modulation by melanocyte-stimulating hormone. Proc Natl Acad Sci U S A 1988;85:3830–3834. [CrossRef]
- Noguchi S, Kumazaki M, Yasui Y, Mori T, Yamada N, Akao Y. MicroRNA-203 regulates melanosome transport and tyrosinase expression in melanoma cells by targeting kinesin superfamily protein 5b. J Invest Dermatol 2014;134:461–469. [CrossRef]
- Hara M, Yaar M, Byers HR, Goukassian D, Fine RE, Gonsalves J, et al. Kinesin participates in melanosomal movement along melanocyte dendrites. J Invest Dermatol 2000;114:438–443. [CrossRef]
- Ishida M, Ohbayashi N, Maruta Y, Ebata Y, Fukuda M. Functional involvement of Rab1A in microtubule-dependent anterograde melanosome transport in melanocytes. J Cell Sci 2012;125:5177–5187. [CrossRef]
- Ishida M, Ohbayashi N, Fukuda M. Rab1A regulates anterograde melanosome transport by recruiting kinesin-1 to melanosomes through interaction with SKIP. Sci Reports 2015 51 2015;5:1–9. [CrossRef]
- Ohbayashi N, Maruta Y, Ishida M, Fukuda M. Melanoregulin regulates retrograde melanosome transport through interaction with the RILP-p150Glued complex in melanocytes. J Cell Sci 2012;125:1508–1518. [CrossRef]
- Matsui T, Ohbayashi N, Fukuda M. The Rab interacting lysosomal protein (RILP) homology domain functions as a novel effector domain for small GTPase Rab36: Rab36 regulates retrograde melanosome transport in melanocytes. J Biol Chem 2012;287:28619. [CrossRef]
- Maruta Y, Fukuda M. Large Rab GTPase Rab44 regulates microtubule-dependent retrograde melanosome transport in melanocytes. J Biol Chem 2022;298:102508. [CrossRef]
- Strom M, Hume AN, Tarafder AK, Barkagianni E, Seabra MC. A family of Rab27-binding proteins: Melanophilin links Rab27a and myosin Va function in melanosome transport. J Biol Chem 2002;277:25423–25430. [CrossRef]
- Hume AN, Ushakov DS, Tarafder AK, Ferenczi MA, Seabra MC. Rab27a and MyoVa are the primary Mlph interactors regulating melanosome transport in melanocytes. J Cell Sci 2007;120:3111–3122. [CrossRef]
- Beaumont KA, Hamilton NA, Moores MT, Brown DL, Ohbayashi N, Cairncross O, et al. The Recycling Endosome Protein Rab17 Regulates Melanocytic Filopodia Formation and Melanosome Trafficking. Traffic 2011;12:627–643. [CrossRef]
- Ni J, Wang N, Gao L, Li L, Zheng S, Liu Y, et al. The effect of the NMDA receptor-dependent signaling pathway on cell morphology and melanosome transfer in melanocytes. J Dermatol Sci 2016;84:296–304. [CrossRef]
- Domingues L, Hurbain I, Gilles-Marsens F, Sirés-Campos J, André N, Dewulf M, et al. Coupling of melanocyte signaling and mechanics by caveolae is required for human skin pigmentation. Nat Commun 2020;11. [CrossRef]
- Moreiras H, Pereira FJC, Neto M V, Bento-Lopes L, Festas TC, Seabra MC, et al. The exocyst is required for melanin exocytosis from melanocytes and transfer to keratinocytes. Pigment Cell Melanoma Res 2020;33:366–371. [CrossRef]
- Cabaço LC, Bento-Lopes L, Neto M V., Ferreira A, Staubli WBL, Ramalho JS, et al. Rab3a regulates melanin exocytosis and transfer induced by keratinocyte-conditioned medium. J Investig Dermatology Innov 2022;2:100139. [CrossRef]
- Lo Cicero A, Delevoye C, Gilles-Marsens F, Loew D, Dingli F, Guéré C, et al. Exosomes released by keratinocytes modulate melanocyte pigmentation. Nat Commun 2015;6. [CrossRef]
- Koike S, Yamasaki K, Yamauchi T, Shimada-Omori R, Tsuchiyama K, Aiba S. Toll-like receptor 2 utilizes RAB11A for melanosome transfer from melanocytes to keratinocytes. J Dermatol Sci 2019;94:310–312. [CrossRef]
- Tadokoro R, Murai H, Sakai KI, Okui T, Yokota Y, Takahashi Y. Melanosome transfer to keratinocyte in the chicken embryonic skin is mediated by vesicle release associated with Rho-regulated membrane blebbing. Sci Rep 2016;6:38277. [CrossRef]
- Moreiras, H, Bento-Lopes, L, Neto, MV, Escrevente, C, Cabaço, LC, Hall, MJ, Ramalho, JS, Seabra, MC, Barral D. Melanocore uptake by keratinocytes occurs through phagocytosis and involves protease-activated receptor-2 internalization. Traffic 2022;23:331–345. [CrossRef]
- Koike S, Yamasaki K, Yamauchi T, Shimada-Omori R, Tsuchiyama K, Ando H, et al. TLR3 stimulation induces melanosome endo/phagocytosis through RHOA and CDC42 in human epidermal keratinocyte. J Dermatol Sci 2019;96:168–177. [CrossRef]
- Benito-Martínez S, Salavessa L, Raposo GG, Marks MS, Delevoye C. Melanin Transfer and Fate within Keratinocytes in Human Skin Pigmentation. Integr Comp Biol 2021;61:1546–1555. [CrossRef]
- Moreiras H, Seabra MC, Barral DC. Melanin Transfer in the Epidermis: The Pursuit of Skin Pigmentation Control Mechanisms. Int J Mol Sci 2021;22. [CrossRef]
- Kumari S, Mg S, Mayor S. Endocytosis unplugged: multiple ways to enter the cell. Cell Res 2010;20:256. [CrossRef]
- Thottacherry JJ, Sathe M, Prabhakara C, Mayor S. Spoiled for Choice: Diverse Endocytic Pathways Function at the Cell Surface. Annu Rev Cell Dev Biol 2019;35:55–84. [CrossRef]
- Wolff K, Konrad K. Phagocytosis of latex beads by epidermal keratinocytes in vivo. J Ultrastruct Res 1972;39:262–280. [CrossRef]
- Sharlow ER, Paine CS, Babiarz L, Eisinger M, Shapiro S, Seiberg M. The protease-activated receptor-2 upregulates keratinocyte phagocytosis. J Cell Sci 2000;113:3093–3101. [CrossRef]
- Belleudi F, Purpura V, Scrofani C, Persechino F, Leone L, Torrisi MR. Expression and signaling of the tyrosine kinase FGFR2b/KGFR regulates phagocytosis and melanosome uptake in human keratinocytes. FASEB J 2011;25:170–181. [CrossRef]
- Mylvaganam S, Freeman SA, Grinstein S. The cytoskeleton in phagocytosis and macropinocytosis. Curr Biol 2021;31:R619–R632. [CrossRef]
- Seiberg M, Paine C, Sharlow E, Andrade-Gordon P, Costanzo M, Eisinger M, et al. The protease-activated receptor 2 regulates pigmentation via keratinocyte-melanocyte interactions. Exp Cell Res 2000;254:25–32. [CrossRef]
- Henehan M, De Benedetto A. Update on protease-activated receptor 2 in cutaneous barrier, differentiation, tumorigenesis and pigmentation, and its role in related dermatologic diseases. Exp Dermatol 2019;28:877–885. [CrossRef]
- Adams MN, Ramachandran R, Yau MK, Suen JY, Fairlie DP, Hollenberg MD, et al. Structure, function and pathophysiology of protease activated receptors. Pharmacol Ther 2011;130:248–282. [CrossRef]
- Heuberger DM, Schuepbach RA. Protease-activated receptors (PARs): mechanisms of action and potential therapeutic modulators in PAR-driven inflammatory diseases. Thromb J 2019;17. [CrossRef]
- Babiarz-Magee L, Chen N, Seiberg M, Lin CB. The expression and activation of protease-activated receptor-2 correlate with skin color. Pigment Cell Res 2004;17:241–251. [CrossRef]
- Lin CB, Chen N, Scarpa R, Guan F, Babiarz-Magee L, Liebel F, et al. LIGR, a protease-activated receptor-2-derived peptide, enhances skin pigmentation without inducing inflammatory processes. Pigment Cell Melanoma Res 2008;21:172–183. [CrossRef]
- Seiberg M. Keratinocyte-Melanocyte Interactions During Melanosome Transfer. Pigment Cell Res 2001;14:236–242. [CrossRef]
- Seiberg M, Paine C, Sharlow E, Andrade-Gordon P, Costanzo M, Eisinger M, et al. Inhibition of melanosome transfer results in skin lightening. J Invest Dermatol 2000;115:162–167. [CrossRef]
- Scott G, Leopardi S, Parker L, Babiarz L, Seiberg M, Han R. The Proteinase-Activated Receptor-2 Mediates Phagocytosis in a Rho-Dependent Manner in Human Keratinocytes. J Invest Dermatol 2003;121:529–541. [CrossRef]
- Wang Y, Li Z, Wu W, Liu Y, Xiao Y, Qi D, et al. TRPA1 promotes melanosome phagocytosis in keratinocytes via PAR-2/CYLD axis. J Dermatol Sci 2022;106:181–188. [CrossRef]
- Cardinali G, Ceccarelli S, Kovacs D, Aspite N, Lotti LV, Torrisi MR, et al. Keratinocyte growth factor promotes melanosome transfer to keratinocytes. J Invest Dermatol 2005;125:1190–1199. [CrossRef]
- Scott G, Deng A, Rodriguez-Burford C, Seiberg M, Han R, Babiarz L, et al. Protease-activated receptor 2, a receptor involved in melanosome transfer, is upregulated in human skin by ultraviolet irradiation. J Invest Dermatol 2001;117:1412–1420. [CrossRef]
- Marchese C, Maresca V, Cardinali G, Belleudi F, Ceccarelli S, Bellocci M, et al. UVB-induced activation and internalization of keratinocyte growth factor receptor. Oncogene 2003;22:2422–2431. [CrossRef]
- Guo MS, Wu Q, Dong TT, Tsim KWK. The UV-induced uptake of melanosome by skin keratinocyte is triggered by α7 nicotinic acetylcholine receptor-mediated phagocytosis. FEBS J 2023;290:724–744. [CrossRef]
- Uribe-Querol E, Rosales C. Phagocytosis: Our Current Understanding of a Universal Biological Process. Front Immunol 2020;11:1066. [CrossRef]
- Freeman SA, Grinstein S. Phagocytosis: receptors, signal integration, and the cytoskeleton. Immunol Rev 2014;262:193–215. [CrossRef]
- Senoo H, Sesaki H, Iijima M. A GPCR Handles Bacterial Sensing in Chemotaxis and Phagocytosis. Dev Cell 2016;36:354. [CrossRef]
- Pan M, Neilson MP, Grunfeld AM, Cruz P, Wen X, Insall RH, et al. A G-protein-coupled chemoattractant receptor recognizes lipopolysaccharide for bacterial phagocytosis. PLoS Biol 2018;16. [CrossRef]
- Wen X, Xu X, Sun W, Chen K, Pan M, Wang JM, et al. G-protein–coupled formyl peptide receptors play a dual role in neutrophil chemotaxis and bacterial phagocytosis. Mol Biol Cell 2019;30:346–356. [CrossRef]
- Doyle SE, O’Connell RM, Miranda GA, Vaidya SA, Chow EK, Liu PT, et al. Toll-like receptors induce a phagocytic gene program through p38. J Exp Med 2004;199:81–90. [CrossRef]
- Deng T, Feng X, Liu P, Yan K, Chen Y, Han D. Toll-like receptor 3 activation differentially regulates phagocytosis of bacteria and apoptotic neutrophils by mouse peritoneal macrophages. Immunol Cell Biol 2013;91:52–59. [CrossRef]
- Del Bino S, Duval C, Bernerd F. Clinical and biological characterization of skin pigmentation diversity and its consequences on UV impact. Int J Mol Sci 2018;19. [CrossRef]
- Nordlund JJ. The Melanocyte and the Epidermal Melanin Unit: An Expanded Concept. Dermatol Clin 2007;25:271–281. [CrossRef]
- Alaluf S, Atkins D, Barrett K, Blount M, Carter N, Heath A. Ethnic variation in melanin content and composition in photoexposed and photoprotected human skin. Pigment Cell Res 2002;15:112–118. [CrossRef]
- Stappers MHT, Clark AE, Aimanianda V, Bidula S, Reid DM, Asamaphan P, et al. Recognition of DHN-melanin by MelLec, is required for protective immunity to Aspergillus. Nature 2018;555:382. [CrossRef]
- van den Berg LM, Zijlstra-Willems EM, Richters CD, Ulrich MMW, Geijtenbeek TBH. Dectin-1 activation induces proliferation and migration of human keratinocytes enhancing wound re-epithelialization. Cell Immunol 2014;289:49–54. [CrossRef]
- Lee HM, Shin DM, Choi DK, Lee ZW, Kim KH, Yuk JM, et al. Innate immune responses to Mycobacterium ulcerans via toll-like receptors and dectin-1 in human keratinocytes. Cell Microbiol 2009;11:678–692. [CrossRef]
- Livden JK. Fc gamma receptors on keratinocytes in psoriasis. Arch Dermatol Res 1988;280:12–17. [CrossRef]
- Cauza K, Grassauer A, Hinterhuber G, Horvat R, Rappersberger K, Wolff K, et al. FcgammaRIII expression on cultured human keratinocytes and upregulation by interferon-gamma. J Invest Dermatol 2002;119:1074–1079. [CrossRef]
- SzabÓ G, Gerald AB, Pathak MA, Fitzpatrick TB. Racial differences in the fate of melanosomes in human epidermis. Nature 1969;222:1081–1082. [CrossRef]
- Minwalla L, Zhao Y, Poole IC Le, Wickett RR, Boissy RE, Le Poole IC, et al. Keratinocytes Play a Role in Regulating Distribution Patterns of Recipient Melanosomes In Vitro. J Invest Dermatol 2001;117:341–347. [CrossRef]
- Yoshida Y, Hachiya A, Sriwiriyanont P, Ohuchi A, Kitahara T, Takema Y, et al. Functional analysis of keratinocytes in skin color using a human skin substitute model composed of cells derived from different skin pigmentation types. FASEB J 2007;21:2829–2839. [CrossRef]
- Thong HY, Jee SH, Sun CC, Boissy RE. The patterns of melanosome distribution in keratinocytes of human skin as one determining factor of skin colour. Br J Dermatol 2003;149:498–505. [CrossRef]
- Ebanks JP, Koshoffer A, Wickett RR, Schwemberger S, Babcock G, Hakozaki T, et al. Epidermal keratinocytes from light vs. dark skin exhibit differential degradation of melanosomes. J Invest Dermatol 2011;131:1226–1233. [CrossRef]
- Murase D, Hachiya A, Takano K, Hicks R, Visscher MO, Kitahara T, et al. Autophagy has a significant role in determining skin color by regulating melanosome degradation in keratinocytes. J Invest Dermatol 2013;133:2416–2424. [CrossRef]
- Ho H, Ganesan AK. The pleiotropic roles of autophagy regulators in melanogenesis. Pigment Cell Melanoma Res 2011;24:595–604. [CrossRef]
- Kim JY, Kim JY, Ahn Y, Lee EJ, Hwang S, Almurayshid A, et al. Autophagy induction can regulate skin pigmentation by causing melanosome degradation in keratinocytes and melanocytes. Pigment Cell Melanoma Res 2020;33:403–415. [CrossRef]
- Chen N, Seiberg M, Lin CB. Cathepsin L2 levels inversely correlate with skin color. J Invest Dermatol 2006;126:2345–2347. [CrossRef]
- Yi WJ, Su MY, Shi Y, Jiang S, Xu SZ, Lei TC. Degraded melanocores are incompetent to protect epidermal keratinocytes against UV damage. Cell Cycle 2018;17:844–857. [CrossRef]
- Ebanks JP, Koshoffer A, Wickett RR, Hakozaki T, Boissy RE. Hydrolytic enzymes of the interfollicular epidermis differ in expression and correlate with the phenotypic difference observed between light and dark skin. J Dermatol 2013;40:27–33. [CrossRef]
- Kim JY, Lee EJ, Ahn Y, Park S, Bae YJ, Kim TG, et al. Cathepsin L, a target of hypoxia-inducible factor-1-α, is involved in melanosome degradation in melanocytes. Int J Mol Sci 2021;22:1–13. [CrossRef]
- Homma T, Kageyama S, Nishikawa A, Nagata K. Melanosome degradation in epidermal keratinocytes related to lysosomal protease cathepsin V. Biochem Biophys Res Commun 2018;500:339–343. [CrossRef]
- Murase D, Kusaka-Kikushima A, Hachiya A, Fullenkamp R, Stepp A, Imai A, et al. Autophagy declines with premature skin aging resulting in dynamic alterations in skin pigmentation and epidermal differentiation. Int J Mol Sci 2020;21:1–17. [CrossRef]
- Yang Z, Zeng B, Pan Y, Huang P, Wang C. Autophagy participates in isoliquiritigenin-induced melanin degradation in human epidermal keratinocytes through PI3K/AKT/mTOR signaling. Biomed Pharmacother 2018;97:248–254. [CrossRef]
- Yun CY, Choi N, Lee JU, Lee EJ, Kim JY, Choi WJ, et al. Marliolide derivative induces melanosome degradation via nrf2/p62-mediated autophagy. Int J Mol Sci 2021;22. [CrossRef]
- Joly-Tonetti N, Wibawa JID, Bell M, Tobin DJ. An explanation for the mysterious distribution of melanin in human skin: a rare example of asymmetric (melanin) organelle distribution during mitosis of basal layer progenitor keratinocytes. Br J Dermatol 2018;179:1115–1126. [CrossRef]
- Mottaz JH, Zelickson AS. Melanin transfer: a possible phagocytic process. J Invest Dermatol 1967;49:605–610. [CrossRef]
- Marubashi S, Fukuda M. Rab7B/42 is functionally involved in protein degradation on melanosomes in keratinocytes. Cell Struct Funct 2020;45:45–55. [CrossRef]
- Delevoye C, Marks MS, Raposo G. Lysosome-related organelles as functional adaptations of the endolysosomal system. Curr Opin Cell Biol 2019;59:147–158. [CrossRef]
- Du C, Zhang T, Xiao X, Shi Y, Duan H, Ren Y. Protease-activated receptor-2 promotes kidney tubular epithelial inflammation by inhibiting autophagy via the PI3K/Akt/mTOR signalling pathway. Biochem J 2017;474:2733–2747. [CrossRef]
- Chandrabalan A, Ramachandran R. Molecular mechanisms regulating Proteinase-Activated Receptors (PARs). FEBS J 2021;288:2697–2726. [CrossRef]
- Borovanský J, Elleder M. Melanosome degradation: Fact or fiction. Pigment Cell Res 2003;16:280–286. [CrossRef]
- Ito S, Wakamatsu K. Chemical degradation of melanins: application to identification of dopamine-melanin. Pigment Cell Res 1998;11:120–126. [CrossRef]
- Akoumianaki T, Kyrmizi I, Valsecchi I, Beauvais A, Akoumianaki T, Kyrmizi I, et al. Aspergillus Cell Wall Melanin Blocks LC3-Associated Phagocytosis to Promote Pathogenicity. Cell Host Microbe 2016;19:79–90. [CrossRef]
- Chamilos G, Akoumianaki T, Kyrmizi I, Brakhage A, Beauvais A, Latge JP. Melanin targets LC3-associated phagocytosis (LAP): A novel pathogenetic mechanism in fungal disease. Autophagy 2016;12:888–889. [CrossRef]
- Moreiras H, Lopes-da-Silva M, Seabra MC, Barral DC. Melanin processing by keratinocytes: A non-microbial type of host-pathogen interaction? Traffic 2019;20:301–304. [CrossRef]
- Joly-Tonetti N, Wibawa JID, Bell M, Tobin D. Melanin fate in the human epidermis: a reassessment of how best to detect and analyse histologically. Exp Dermatol 2016;25:501–504. [CrossRef]
- d’Ischia M, Napolitano A, Michalczyk-Wetula D, Płonka PM. Melanin “dust” or “ghost”? Exp Dermatol 2016;25:505–506. [CrossRef]
- Brenner M, Hearing VJ. The protective role of melanin against UV damage in human skin. Photochem Photobiol n.d.;84:539–549. [CrossRef]
- Kobayashi N, Nakagawa A, Muramatsu T, Yamashina Y, Shirai T, Hashimoto MW, et al. Supranuclear melanin caps reduce ultraviolet induced DNA photoproducts in human epidermis. J Invest Dermatol 1998;110:806–810. [CrossRef]
- Gibbs S, Murli S, De Boer G, Mulder AAT, Mieke Mommaas A, Ponec M. Melanosome capping of keratinocytes in pigmented reconstructed epidermis--effect of ultraviolet radiation and 3-isobutyl-1-methyl-xanthine on melanogenesis. Pigment Cell Res 2000;13:458–466. [CrossRef]
- Lan Y, Zeng W, Wang Y, Dong X, Shen X, Gu Y, et al. Opsin 3 mediates UVA-induced keratinocyte supranuclear melanin cap formation. Commun Biol 2023;6. [CrossRef]
- Vaidžiulyte K, Coppey M, Schauer K. Intracellular organization in cell polarity – placing organelles into the polarity loop. J Cell Sci 2019;132. [CrossRef]
- Randolph Byers H, Maheshwary S, Amodeo DM, Dykstra SG. Role of Cytoplasmic Dynein in Perinuclear Aggregation of Phagocytosed Melanosomes and Supranuclear Melanin Cap Formation in Human Keratinocytes. J Invest Dermatol 2003;121:813–820. [CrossRef]
- Byers HR, Dykstra SG, Boissel SJS. Requirement of dynactin p150(Glued) subunit for the functional integrity of the keratinocyte microparasol. J Invest Dermatol 2007;127:1736–1744. [CrossRef]
- Castellano-Pellicena I, Morrison CG, Bell M, O’Connor C, Tobin DJ. Melanin Distribution in Human Skin: Influence of Cytoskeletal, Polarity, and Centrosome-Related Machinery of Stratum basale Keratinocytes. Int J Mol Sci 2021;22:1–19. [CrossRef]
- Jordens I, Fernandez-Borja M, Marsman M, Dusseljee S, Janssen L, Calafat J, et al. The Rab7 effector protein RILP controls lysosomal transport by inducing the recruitment of dynein-dynactin motors. Curr Biol 2001;11:1680–1685. [CrossRef]
- Risueño I, Valencia L, Jorcano JL, Velasco D. Skin-on-a-chip models: General overview and future perspectives. APL Bioeng 2021;5:1–12. [CrossRef]
- Dustin ML. The immunological synapse. Cancer Immunol Res 2014;2:1023–1033. [CrossRef]
- Südhof TC. The cell biology of synapse formation. J Cell Biol 2021;220. [CrossRef]
- Mizutani Y, Yamashita M, Hashimoto R, Atsugi T, Ryu A, Hayashi A, et al. Three-dimensional structure analysis of melanocytes and keratinocytes in senile lentigo. Reprod Syst Sex Disord 2021;70:224–231. [CrossRef]
Figure 1.
Melanin distribution throughout the epidermis and organization within keratinocytes from distinct phototypes. A. Melanin dispersion in the epidermis is achieved through the formation of epidermal-melanin units, where one melanocyte transfers melanin to 30-40 surrounding basal keratinocytes. The accumulation of melanin is higher in the basal layers regardless of the skin phototype. However, in lower phototypes (i.e., lighter skins), less melanin is concentrated in the basal layer, whereas in higher phototypes (i.e., darker skins), higher levels can be found in the basal layer and melanin is also present in the immediate suprabasal layers. B. Additionally, electron microscopy studies have shown that melanin organizes differently within keratinocytes derived from different phototypes. In lower phototypes, melanin is mainly stored in clusters of several melanin granules within a single membrane organelle, while higher phototypes prominently accumulate single granules in single membrane organelles. This was shown to be determined by the keratinocyte phototype and does not depend on the type of melanin transferred (i.e., eumelanin or pheomelanin).
Figure 1.
Melanin distribution throughout the epidermis and organization within keratinocytes from distinct phototypes. A. Melanin dispersion in the epidermis is achieved through the formation of epidermal-melanin units, where one melanocyte transfers melanin to 30-40 surrounding basal keratinocytes. The accumulation of melanin is higher in the basal layers regardless of the skin phototype. However, in lower phototypes (i.e., lighter skins), less melanin is concentrated in the basal layer, whereas in higher phototypes (i.e., darker skins), higher levels can be found in the basal layer and melanin is also present in the immediate suprabasal layers. B. Additionally, electron microscopy studies have shown that melanin organizes differently within keratinocytes derived from different phototypes. In lower phototypes, melanin is mainly stored in clusters of several melanin granules within a single membrane organelle, while higher phototypes prominently accumulate single granules in single membrane organelles. This was shown to be determined by the keratinocyte phototype and does not depend on the type of melanin transferred (i.e., eumelanin or pheomelanin).
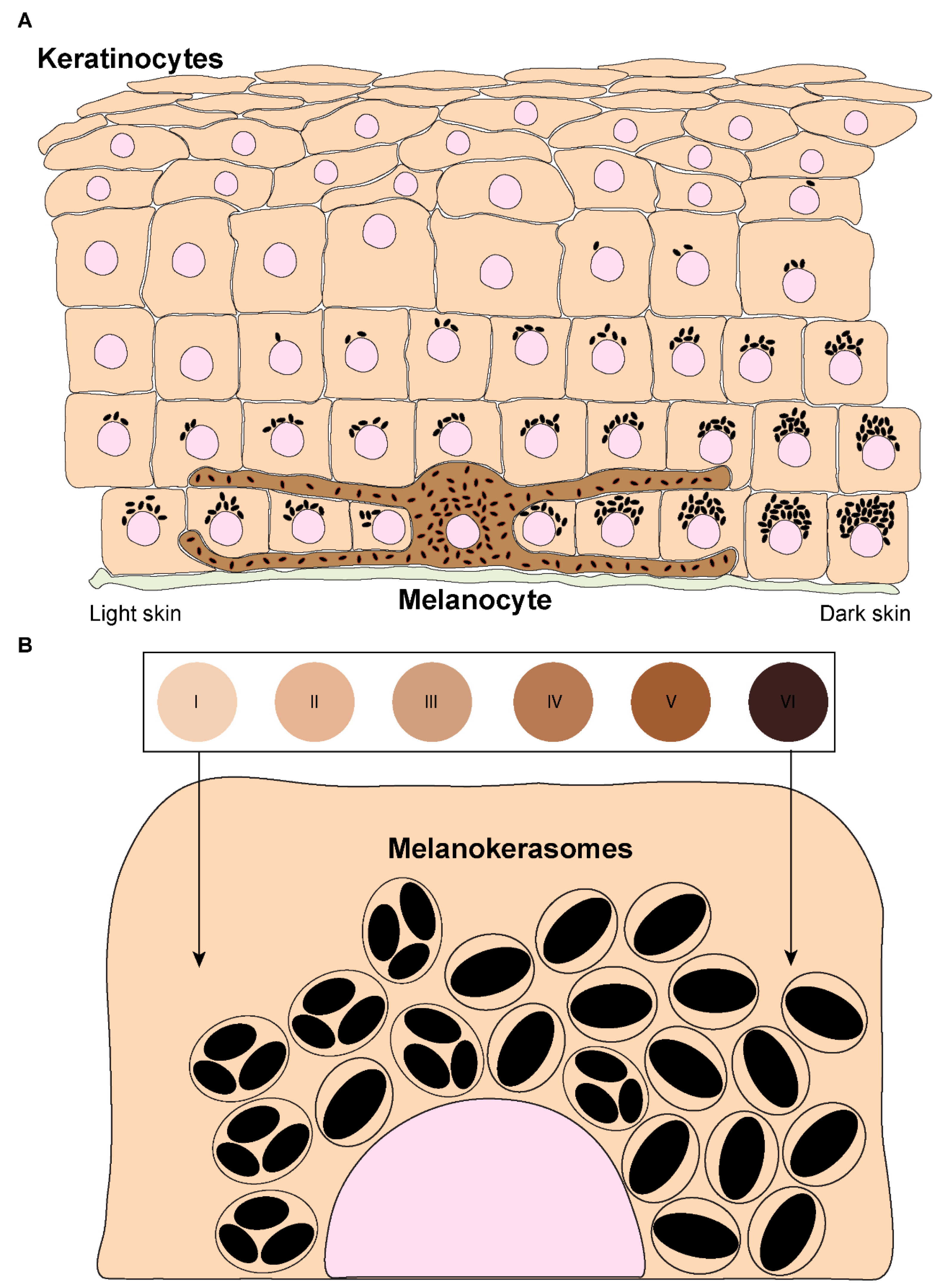
Figure 2.
Melanin transfer from melanocytes to keratinocytes. Four different models have been proposed to explain the mechanism(s) of melanin transfer in the skin, including the cytophagocytosis of melanocyte dendrites by keratinocytes, the direct fusion of melanocyte and keratinocyte membranes, the shedding of melanosome-laden globules by melanocytes and the coupled exo/phagocytosis of the melanin core. Despite the evidence published for all these models, using different models, recent studies using human and mouse cell lines, as well as more complex models such as reconstructed human skins/epidermises and skin biopsies, have supported the transfer of melanin in globules or as melanocores. These two models rely on the exocytosis of melanin by melanocytes, which is then phagocytosed by keratinocytes. Since melanin is mainly found in single membrane organelles, it has been proposed that the membranes derived form the melanosome and the melanocyte are degraded within keratinocytes after internalization, in the case of shed globules model. In the case of melanocore transfer, they accumulate within single membrane organelles right after their internalization.
Figure 2.
Melanin transfer from melanocytes to keratinocytes. Four different models have been proposed to explain the mechanism(s) of melanin transfer in the skin, including the cytophagocytosis of melanocyte dendrites by keratinocytes, the direct fusion of melanocyte and keratinocyte membranes, the shedding of melanosome-laden globules by melanocytes and the coupled exo/phagocytosis of the melanin core. Despite the evidence published for all these models, using different models, recent studies using human and mouse cell lines, as well as more complex models such as reconstructed human skins/epidermises and skin biopsies, have supported the transfer of melanin in globules or as melanocores. These two models rely on the exocytosis of melanin by melanocytes, which is then phagocytosed by keratinocytes. Since melanin is mainly found in single membrane organelles, it has been proposed that the membranes derived form the melanosome and the melanocyte are degraded within keratinocytes after internalization, in the case of shed globules model. In the case of melanocore transfer, they accumulate within single membrane organelles right after their internalization.
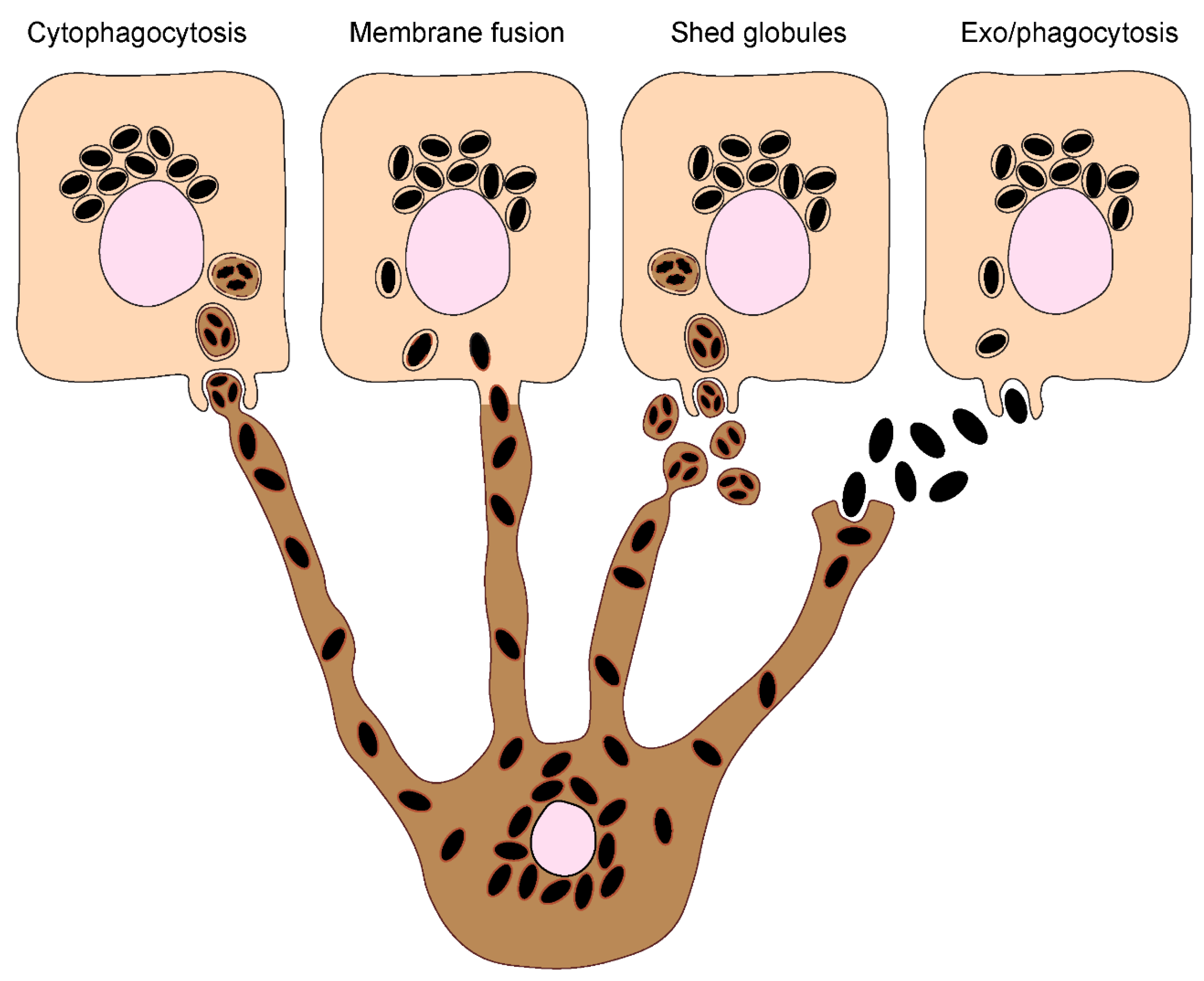
Figure 3.
Melanin exocytosis by melanocytes and internalization by keratinocytes. A. Melanin exocytosis in physiological conditions is thought to occur at the tips of melanocyte dendrites, requiring melanosome transport to the cell periphery. Anterograde long-range transport occurs via microtubules and is mediated by the complex Rab1-SKIP-kinesin 1, resulting in melanosome dispersion throughout the cytoplasm. On the other hand, retrograde transport results in perinuclear accumulation of melanosomes and is regulated by Rab36-melanoregulin-RILP-DCTN1 and/or Rab44-dynactin-dynein. Melanosome positioning results from the balance between anterograde and retrograde transport. Since retrograde transport overrides anterograde transport, melanosomes tend to accumulate at the perinuclear region. Melanosome peripheral positioning requires the switch from microtubule-based transport to actin-mediated transport, through the tripartite complex composed by Rab27a-melanophilin-Myosin Va. The mechanisms regulating the late steps of melanin exocytosis were only recently addressed. The small GTPase Rab11b and the exocyst complex subunits Exo70 and Sec8 were found to regulate basal melanin exocytosis and transfer. Furthermore, Rab3a enhances melanin exocytosis and transfer to keratinocytes under stimulation by soluble factors derived from differentiated keratinocytes. The existence of different regulators of melanin exocytosis suggests that this process can be fine-tuned to ensure efficient melanin transfer depending on the conditions. B. The transfer of secreted melanin requires its internalization by keratinocytes. Recent studies found key differences in the molecular players involved in melanin recognition and internalization by keratinocytes, according to the way melanin is presented to keratinocytes, i.e., “naked” melanocores or membrane-bound melanosomes. Melanocores were shown to be phagocytosed in a PAR-2-dependent manner by keratinocytes, requiring also Rac1 and Cdc42 for efficient internalization. These observations suggest the specific recognition of melanocores by keratinocytes, although the phagocytic receptor(s) involved remain(s) to be determined. In contrast, purified membrane-bound melanosomes are internalized through macropinocytosis in a PAR-2 independent manner, but requiring RhoA and CtBP1/BARS. Whether the internalization of melanosome-laden globules requires the same regulators as melanosomes remains to be determined. Additionally, FGFR2b/KGFR and TLR3 stimulation was shown to enhance the internalization of melanosomes and melanocores, although the mechanisms were not characterized. Whether the existence of different internalization routes affects melanokerasome processing is not known.
Figure 3.
Melanin exocytosis by melanocytes and internalization by keratinocytes. A. Melanin exocytosis in physiological conditions is thought to occur at the tips of melanocyte dendrites, requiring melanosome transport to the cell periphery. Anterograde long-range transport occurs via microtubules and is mediated by the complex Rab1-SKIP-kinesin 1, resulting in melanosome dispersion throughout the cytoplasm. On the other hand, retrograde transport results in perinuclear accumulation of melanosomes and is regulated by Rab36-melanoregulin-RILP-DCTN1 and/or Rab44-dynactin-dynein. Melanosome positioning results from the balance between anterograde and retrograde transport. Since retrograde transport overrides anterograde transport, melanosomes tend to accumulate at the perinuclear region. Melanosome peripheral positioning requires the switch from microtubule-based transport to actin-mediated transport, through the tripartite complex composed by Rab27a-melanophilin-Myosin Va. The mechanisms regulating the late steps of melanin exocytosis were only recently addressed. The small GTPase Rab11b and the exocyst complex subunits Exo70 and Sec8 were found to regulate basal melanin exocytosis and transfer. Furthermore, Rab3a enhances melanin exocytosis and transfer to keratinocytes under stimulation by soluble factors derived from differentiated keratinocytes. The existence of different regulators of melanin exocytosis suggests that this process can be fine-tuned to ensure efficient melanin transfer depending on the conditions. B. The transfer of secreted melanin requires its internalization by keratinocytes. Recent studies found key differences in the molecular players involved in melanin recognition and internalization by keratinocytes, according to the way melanin is presented to keratinocytes, i.e., “naked” melanocores or membrane-bound melanosomes. Melanocores were shown to be phagocytosed in a PAR-2-dependent manner by keratinocytes, requiring also Rac1 and Cdc42 for efficient internalization. These observations suggest the specific recognition of melanocores by keratinocytes, although the phagocytic receptor(s) involved remain(s) to be determined. In contrast, purified membrane-bound melanosomes are internalized through macropinocytosis in a PAR-2 independent manner, but requiring RhoA and CtBP1/BARS. Whether the internalization of melanosome-laden globules requires the same regulators as melanosomes remains to be determined. Additionally, FGFR2b/KGFR and TLR3 stimulation was shown to enhance the internalization of melanosomes and melanocores, although the mechanisms were not characterized. Whether the existence of different internalization routes affects melanokerasome processing is not known.
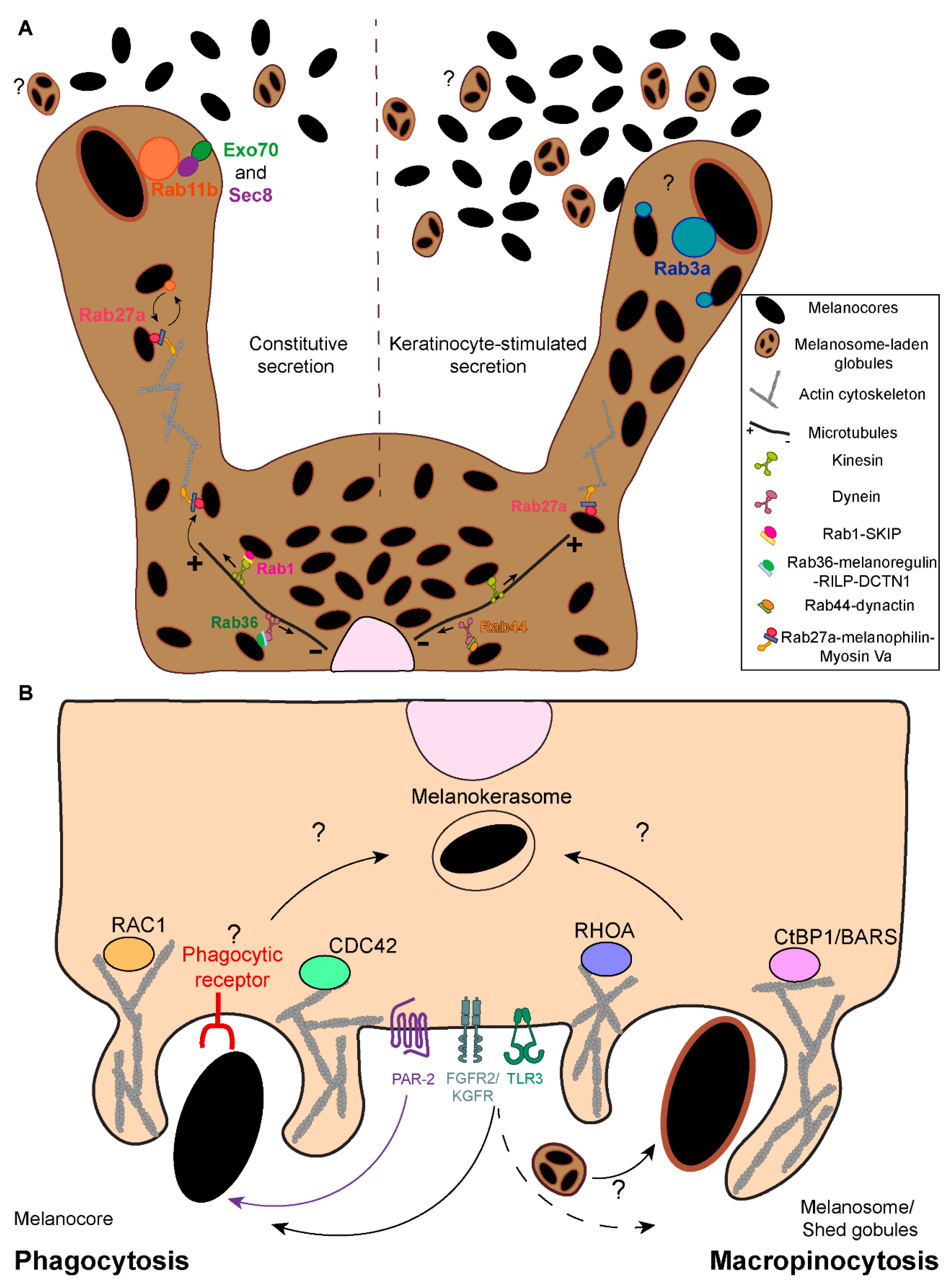
Figure 4.
Melanin processing within keratinocytes. A. Melanin processing comprises the steps between melanin internalization and accumulation in the supranuclear region. Recent studies proposed that transferred melanin inside keratinocytes is surrounded by early and late endocytic markers. Whether this organelle undergoes further maturation is not known, but our recent observations suggest that melanin is stored in a specialized organelle that we named melanokerasome (MKS).MKS have a poorly acidic lumen and non-degradative properties, suggesting that they are specialized for melanin persistence within keratinocytes. The biogenesis of MKS and the relevance of the interaction with other organelles is not fully understood, despite the possible involvement of early and late endocytic vesicles as well as autophagic vesicles. Additionally, the contribution of receptor signaling (e.g. PAR-2, KGFR/FGFR2 and TLR3 or phagocytic receptors) for the trafficking and biogenesis of MKSs is not known. B. Autophagy has been reported to regulate melanin degradation within keratinocytes, although the mechanisms are not understood. Moreover, autophagic and lysosomal activities are higher in lighter skins when compared to darker skins, suggesting differences in melanin processing among phototypes. Nevertheless, it is not fully understood whether MKSs from distinct phototypes present different properties regarding their ability to allow melanin persistence or degradation, i.e., in terms of their acidity and degradative capacity. Additionally, the contribution of autophagic/lysosomal activities from distinct phototypes for the establishment of clusters vs. single granules in the different phototypes has not been not explored. EEs - Early endosomes; LEs - Late endosomes; MVBs – Multivesicular bodies; AVs – Autophagic vesicles; Lys – Lysosomes.
Figure 4.
Melanin processing within keratinocytes. A. Melanin processing comprises the steps between melanin internalization and accumulation in the supranuclear region. Recent studies proposed that transferred melanin inside keratinocytes is surrounded by early and late endocytic markers. Whether this organelle undergoes further maturation is not known, but our recent observations suggest that melanin is stored in a specialized organelle that we named melanokerasome (MKS).MKS have a poorly acidic lumen and non-degradative properties, suggesting that they are specialized for melanin persistence within keratinocytes. The biogenesis of MKS and the relevance of the interaction with other organelles is not fully understood, despite the possible involvement of early and late endocytic vesicles as well as autophagic vesicles. Additionally, the contribution of receptor signaling (e.g. PAR-2, KGFR/FGFR2 and TLR3 or phagocytic receptors) for the trafficking and biogenesis of MKSs is not known. B. Autophagy has been reported to regulate melanin degradation within keratinocytes, although the mechanisms are not understood. Moreover, autophagic and lysosomal activities are higher in lighter skins when compared to darker skins, suggesting differences in melanin processing among phototypes. Nevertheless, it is not fully understood whether MKSs from distinct phototypes present different properties regarding their ability to allow melanin persistence or degradation, i.e., in terms of their acidity and degradative capacity. Additionally, the contribution of autophagic/lysosomal activities from distinct phototypes for the establishment of clusters vs. single granules in the different phototypes has not been not explored. EEs - Early endosomes; LEs - Late endosomes; MVBs – Multivesicular bodies; AVs – Autophagic vesicles; Lys – Lysosomes.
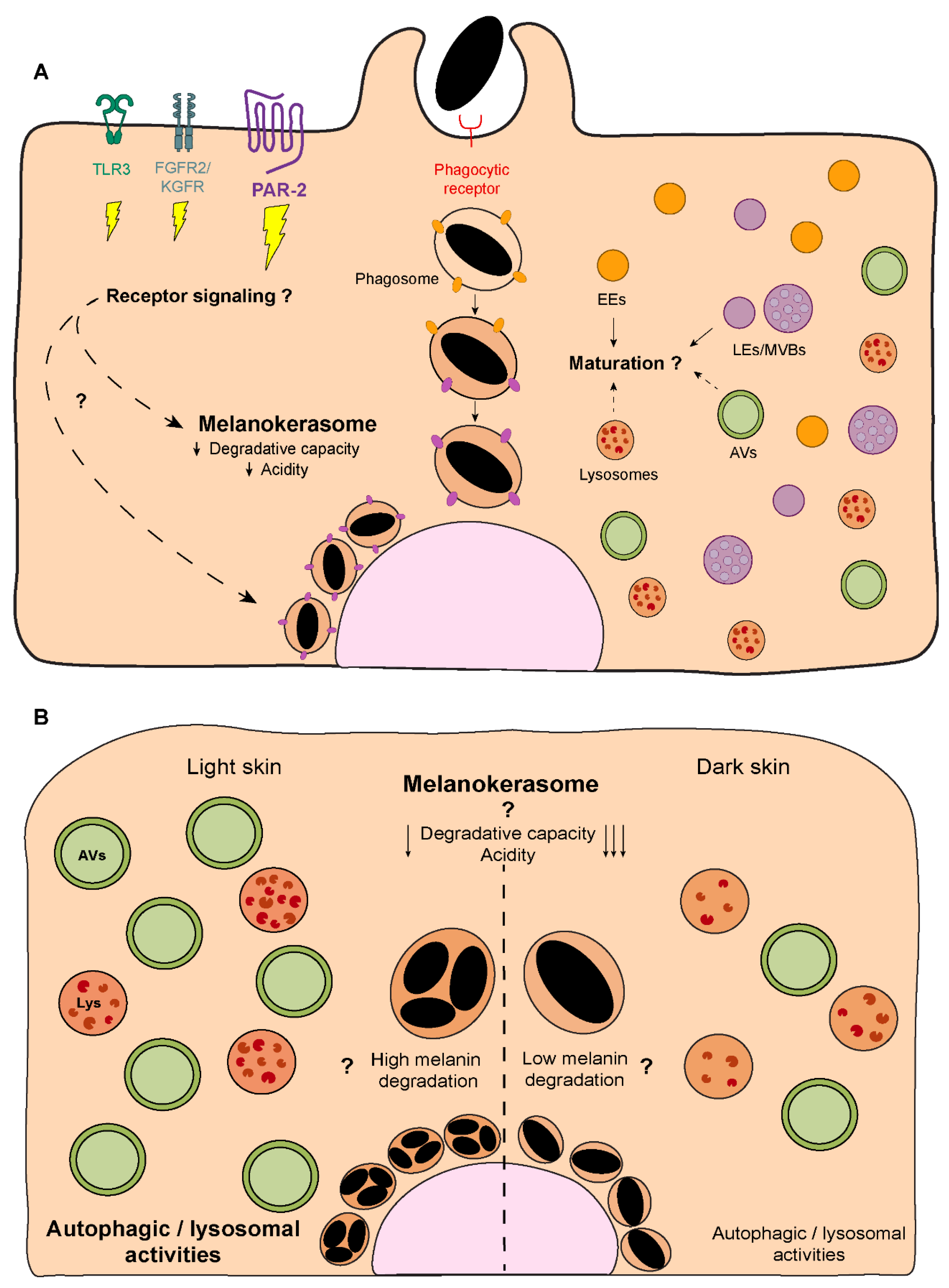
Figure 5.
Melanin trafficking and positioning within keratinocytes. The accumulation of melanin at the supranuclear region of keratinocytes is crucial for its photoprotective properties in human skin. Despite the importance of this process, the molecular mechanisms involved are still poorly understood. Melanin is known to be retrogradely transported along microtubules, in a process mediated by the dynein-dynactin motor complex. It is not known if an adaptor protein (or proteins) is(are) involved in this process and how the transport is regulated. Given the importance of this accumulation for the role of melanin in keratinocytes, further studies should be conducted in order to better characterize melanin positioning in the skin.
Figure 5.
Melanin trafficking and positioning within keratinocytes. The accumulation of melanin at the supranuclear region of keratinocytes is crucial for its photoprotective properties in human skin. Despite the importance of this process, the molecular mechanisms involved are still poorly understood. Melanin is known to be retrogradely transported along microtubules, in a process mediated by the dynein-dynactin motor complex. It is not known if an adaptor protein (or proteins) is(are) involved in this process and how the transport is regulated. Given the importance of this accumulation for the role of melanin in keratinocytes, further studies should be conducted in order to better characterize melanin positioning in the skin.
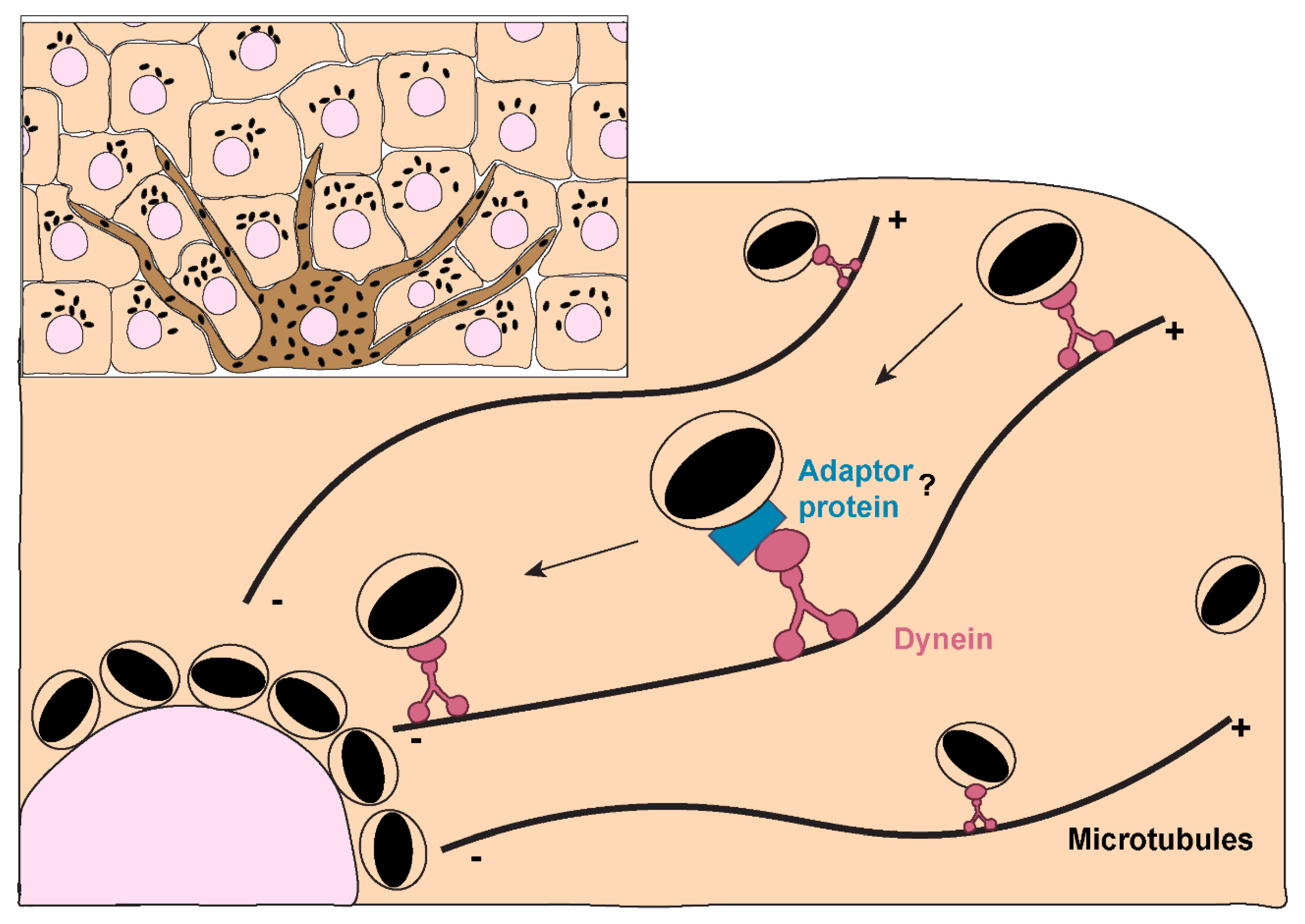

Table 1.
– List of compounds that increase or decrease autophagy in keratinocytes and lead to decreased or increased melanin content, respectively.
Table 1.
– List of compounds that increase or decrease autophagy in keratinocytes and lead to decreased or increased melanin content, respectively.
| Autophagy modulator | Mechanism proposed | Model | References |
|---|---|---|---|
| Isoliquiritigenin | Decrease in melanin content through the suppression of the PI3K/AKT/mTOR pathway | Human epidermal keratinocytes | 91 |
| ATG7 silencing | Increase in melanin content | HaCaT human keratinocytes | 91 |
| 3-Methyladenine (3-MA) | Increase in melanin content | HaCaT human keratinocytes | 91 |
| Pentasodium tetracarboxymethyl palmitoyl dipeptide-12 (PTPD-12) | Decrease in melanin content | Human epidermal keratinocytes | 84 |
| ATG7 silencing | Increase in melanin content | Human epidermal keratinocytes | 82 |
| ATG13 or UVRAG silencing | Increase in melanin content | Human epidermal keratinocytes | 82 |
| Lysosome inhibitors (E-64-D and pepstatin A) | Increase in melanin content | RHPE and skin explants | 82 |
| Verapamil | Decrease in melanin content | RHPE and skin explants | 82 |
| Rapamycin | Decrease in melanin content | RHPE and skin explants | 82 |
| Hydroxychloroquine | Increase in melanin content | RHPE and skin explants | 82 |
| Marliolide derivative (5-methyl-3-tetradecylidene-dihydro-furan-2-one; DMF02) | Decrease in melanin content through Nrf2-p62 activation | HaCaT and human epidermal keratinocytes | 92 |
| Torin 1 | Decrease in melanin content through mTOR inhibition | Skin explants | 90 |
ATG - Autophagy-related protein; UVRAG - UV radiation resistance associated; RHPE - reconstructed human pigmented epidermis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



