
Submitted:
19 May 2023
Posted:
23 May 2023
You are already at the latest version
Abstract
During the last few decades bat lyssaviruses have become the topic of intensive molecular and epidemiological investigations. Since ancient times rhabdoviruses have caused fatal encephalitis in humans which has led to research into effective strategies for their eradication. Modelling of potential future cross-species virus transmissions forms a substantial component of the recent infection biology of rabies. In this article we summarise the available data on the phylogeography of both bats and lyssaviruses in Europe and the adjacent regions, especially in the contact zone between the Palearctic and Ethiopian realms. Within these zones three bat families are present with high potential for cross-species transmission and the spread of lyssaviruses in Phylogroup II to Europe (part of the western Palearctic). The lack of effective therapies for rabies viruses in Phylogroups II and III generates impetus for additional phylogenetic and virological research within this geographical region.
Keywords:
lyssavirus phylogroups
; Chiroptera
; evolution
; transition routes
; health
1. Introduction
The Order Chiroptera has a Laurasiatherian origin (“laurasian beasts”), evolved between 50 and 70 million years ago (MYA), and has undergone rapid diversification ([1,2]. Due to their capabilities of self- powered flight and echolocation, bats [3] comprise over 20%, or more than 1400 species, of all modern mammals and are globally distributed, with the exception of the extreme polar regions [4]. They have many characteristics that differentiate them from other mammalian species, such as their unique physiology [5,6], metabolism [7], and immune system [2,8,9]. These features make them a suitable reservoir for viral zoonoses [4,10,11] and more than 200 viruses have been isolated from or detected in bats (see [4,12,13,14]). The order comprises 45 species in Europe [15] from two superfamilies the Rhinolophoidea and Vespertilionoidea [16], representing a natural reservoir of RNA-viruses (Table 1). Viruses from 11 families have been isolated on the continent [17], and bat lyssaviruses in Europe (family Rhabdoviridae) have been the subject of detailed reviews by [18,19,20,21].
Lyssaviruses are a genus of negative-sense single-strand RNA viruses in the family Rhabdoviridae, subfamily Alpharhabdovirinae. Notably, they are members of the order Mononegavirales, which includes other prominent zoonotic pathogens such as filoviruses (Ebola, Marburg, etc.) and the neurotropic Bornaviridae [22]. Based on genetic divergence lyssaviruses are classified into 17 different viral species according to the latest ICTV Taxonomy release [23]. Recently, two putative new lyssaviruses were published [24,25,26]. Apart from the Mokola virus (MOKV) and Ikoma lyssavirus (IKOV), which have rodents and African civets as a reservoir, respectively [25,27,28], the rest of the lyssaviruses are transmitted by Chiroptera [27,29].
The genus includes the classical rabies virus (RABV), Lyssavirus Aravan (ARAV), Lyssavirus australis (ABLV), Lyssavirus Bokeloh (BBLV), Lyssavirus Duvenhage (DUVV), Lyssavirus Hamburg (EBLV-1), Lyssavirus Helsinki (EBLV-2), Lyssavirus Gannoruwa (GBLV), Lyssavirus Ikoma (IKOV), Lyssavirus Irkut (IRKV), Lyssavirus Khujand (KHUV), Lyssavirus Lagos (LBV), Lyssavirus Lleida (LLEBV), Lyssavirus Mokola (MOKV), Lyssavirus Shimoni (SHIBV), Kotalahti bat lyssavirus (KBVL), Lyssavirus Caucasicus (WCBV) and Lyssavirus Formosa, which includes Taiwan bat lyssavirus -1 (TWBLV-1) and Taiwan bat lyssavirus-2 (TWBLV-2) [21,24,30,31,32,33]. The current study aims to review the evolution, phylogeography and transmission routes of bat lyssaviruses in Europe.
2. Origin, Evolution and Geographic Distribution of Bat Lyssaviruses
Despite the greater diversity of African lyssaviruses [34], Hayman et al. [35] assumed that they have a Palearctic origin and highlighted “—Out of Africa” hypothesis. Lyssaviruses most recent common ancestor (MRCA) evolved from an insect rhabdovirus before 7,000–11,000 years ago [30,36] which was transmitted to representatives of the order Chiroptera and spread globally [36,37]. According to Rupprecht et al. [30] Africa is the most likely home to the ancestors of taxa within the Genus Lyssavirus, Family Rhabdoviridae. According to this review a large number of different lyssaviruses co-evolved with bats, as ultimate reservoirs, over millions of years. It is assumed that all mammals be susceptible to infection with the rabies virus. Apart from bats as reservoirs of infection the so-called mesocarnivores (canids, raccoons, skunks etc.) also play a significant role in the spread and transmission of the rabies virus. The fact that the biological localization of the virus is the central nervous system poses a real challenge for its eradication.
Lyssaviruses have undergone purifying selection followed by neutral evolution of the viral genomes, [38]. The low rate of nonsynonymous evolution of lyssaviruses is probably the result of constraints imposed by the need to replicate in multiple cell types (muscle, peripheral and central nervous systems, and salivary glands) within the host, which in turn boosts cross-species transmission (e. g. different groups of mammals), or because viral proteins are not subject to immune selection, which means existing lyssaviruses are well adapted to their reservoir [39,40]. The host switching of the classic rabies lyssavirus—RABV from bats to other mammals is estimated to have occurred 800 to 1,400 years ago, which does not explain the timing of the oldest putative human rabies cases, estimated to have circulated 4,000 years ago in ancient Mesopotamia [41]. A possible explanation is that the Mesopotamian RABV lineage disappeared as a consequence of genetic drift (loss of polymorphism) or its high fatality rates [41]. According to Rupprecht el al and Badrane et al. [42,43], bats are the primary evolutionary host of rabies viruses as a reservoir of all existing lyssaviruses except MOKV and IKOV, whereas other mammals and humans only maintain several lineages of RABV, including the extinct Mesopotamian strain [30,41,43].
In Europe bat lyssaviruses (Figure 1) were detected in the United Kingdom, the Netherlands, Finland, Denmark, Poland, Czech Republic, Germany, Switzerland, France, Spain, Hungary, Italy, Croatia, Bulgaria, Ukraine and Russia [19,21,44,45,46,47,48,49]. During the last two decades, previously unknown lyssaviruses were isolated as follows: WCBV in 2002 on the European side of the Caucasus Mts. [50], BBLV in 2010 from Germany [51], LLEBV in 2011 from Spain [52], and KBLV in 2017 from Finland [24].
The most frequent lineages are EBLV-1, first reported in 1955 from Germany, and EBLV-2, isolated in 1985 in Switzerland [44,45]. EBLV-1 is exclusively detected in Serotine bats (Eptesicus serotinus) while EBLV-2 is mainly found in Daubenton’s bats (Myotis daubentonii). EBLV-1 is present in two forms: EBLV-1a and EBLV-1b. EBLV-1a displays a wide geographical distribution between France and Russia with phylogenetic homogeneity—an indication of extensive dispersal by bats [20,53]. Resent research has shown that EBLV-1 is associated with the bat E. serotinus of the mountainous parts of Southern Europe, such as the French Alps or the Iberian Peninsula [54]. EBLV-1 showed the risk of spillover because of its close phylogenetic relation with a different bat, the E. isabellinus. The phylogenetic analysis of nine EBLV-1 strains of E. serotinus distributed in south of the Pyrenees revealed that two of them closely related with EBLV-1a sequences from Southern France i.e., this group expanded to Northern Spain. The results of the conducted research give the authors reason to assume the expansion of EBLV-1a subtype across southern France, with very recent arrival to the Iberian Peninsula i.e., current southwards dissemination [54]. In contrast, EBLV-1b is distributed between Spain and Poland with a well-defined geographic structure, indicating restricted contact between bat populations [20,53]. Therefore EBLV-1b had the potential to spread southwards according to the E. isabellinus distribution. The lineage of EBLV-1 is presumed to have arisen 500 to 750 years ago and has a relatively recent origin [53]. Conversely, the lineage of EBLV-2 is dated to more than 8000 years ago, with current establishment in Europe within the last 2000 years. [55]. EBLV-2 has been reported in Western Europe and is also represented by two forms: EBLV-2a and EBLV-2b [46,56]. The first occurs in the United Kingdom, Netherlands, Germany, Switzerland and Denmark, while the second includes the Finnish EBLV-2 strains, and a strain from Switzerland [55], where the divergence of the Finnish strains from the Swiss strain occurred within the last 200 years [55].
3. Virion structure and genome
Rhabdoviruses (family Rhabdoviridae) have a characteristic bullet-shaped virion morphology, with an envelope derived from the plasma membrane of the infected host cell and approximate dimensions of ~75 nm and a length ~180 nm, which distinguishes them from other taxa in the order Mononegavirales: Bornaviridae, Filoviridae, and Paramyxoviridae. They include an approximately 12 kb long non-segmented, linear, single-strand RNA genome. The basic genome includes five genes that encode (from 3′ to 5′) the nucleoprotein (or nucleocapsid protein, N), phosphoproteins (P), matrix protein (M), glycoprotein (G) and large protein (L, RNA—dependent RNA polymerase) [57,58].
The lyssaviruses archetypes are design by variations of long non-coding region in the G gene and the small alternative open reading frames known as ORFs in the P gene [59]. ORFs present an ancestral pseudogene which are used for studying virus–host interactions in WCBV [37] due to the outstanding size which is 40% larger than in other bat lyssaviruses [60]. It has been found that in some rhabdoviruses very long non-coding regions (up to 749 nt) were present either within or between transcriptional units [37]. This region seems to serve as a resource to de novo emergence of genes which may be related to elucidating the taxonomy, phylogeny and evolution of lyssaviruses. This is most likely to occur when ORFs are present in transcribed non-coding regions (UTRs) such as the in the so-called ‘pseudogene ψ region’ of WCBV which is unique to an ORF of 180 nt has been identified lyssaviruses The de novo creation of genes in non-transcribed intergenic regions (IGRs), as well as those present in the G-L gene junctions of various hapavirus, is associated with prior or simultaneous evolution of new or modified transcriptional control sequences. In the trend towards increasing genome size and complexity in rhabdoviruses, the loss of a gene and/or genes is also likely to have occurred periodically, which may also be evolutionarily determined in family Rhabdoviridae [37].
4. Phylogeny of bat lyssaviruses
Based on the sequence analysis of lyssavirus N gene, serologic cross-reactivity and pathogenicity bat lyssaviruses are divided into three phylogroups [35,61,62,63] (Figure 2, S1). For simplicity, we used 19 reference available in GenBank (https://www.ncbi.nlm.nih.gov/genbank/) for our phylogenetic analysis. European viruses are included in Phylogroups I and III. Due to this fact, Phylogroup II is discussed only as a potential scenario for cross-species bat transmission.
Phylogroup I includes species RABV, ARAV, ABLV, BBLV, DUVV, EBLV-1, EBLV-2, GBLV, IRKV, KBLV, KHUV, TWBLV -1 and TWBLV -2 whereas LBV, MOKV, and SHIBV form Phylogroup II [24,31,43,44,62,66,67]. Phylogenetically, species LLEBV, IKOV and WCBV appear related and represent phylogroup III [27,50,68,69]. Phylogroup I is divided into two major groups: the first includes the Palearctic species IRKV, EBLV-1, TWBLV-1, TWBLV-2 and African DUVV lyssaviruses and the second ARAV, BBLV, KHUV and EBLV-2 which are also rabies viruses with Palearctic distribution, as well as Australian—ABLV, Oriental—GBLV and American—RABV lyssaviruses [35]. Interestingly, EBLV-1 is most closely related to DUVV, and IRKV, while EBLV-2 to KBLV, KHUV and BBLV [30,32]. Based on the close phylogenetic relation between EBLV-1 and DUVV species [70] hypothesized that EBLV-1 originated in North Africa and spread to Europe (Iberian Peninsula) via the Strait of Gibraltar. However, [35] present phylogenetic evidence, based on the rabies N gene sequences, that EBLV-1 and DUVV share a common ancestor with IRKV (isolate from Russia), and both have been transferred to Africa from the Palearctic region, and Europe in particular. Phylogenetic relationships in Phylogroup III demonstrate close phylogenetic relatedness between the LLEBV virus from Spain with the Eurasian WCBV and the African non-bat related IKOV lyssavirus [35,71]. Genetically LLEBV is more closely related to IKOV than to WCBV.
For a better understanding of lyssavirus phylogeny and their current distributions, a closer look at their bat species reservoirs is required. Genomic and mitochondrial analyses have placed bats into two suborders: Yinpterochiroptera—including the five families in the superfamily Rhinolophoidea plus the flying foxes—Pteropodidae, and Yangochiroptera—including the three superfamilies: Emballonuroidea, Vespertilionoidea, and Noctilionoidae, comprising a total of 13 families. Two superfamilies (Rhinolophoidea and Vespertilionoidea) are of particular interest in Europe because their representatives are the main reservoir of lyssaviruses. The greater horseshoe bat (Rhinolophus ferrumequinum) (Rhinolophidae, Rhinolophoidea), and the Vespertilionoidea species Greater mouse-eared bat (Myotis myotis), Lesser mouse-eared bat (M. blythii), Natterer’s bat (M. nattereri), Serotine bat (Eptesicus serotinus), Meridional serotine (E. isabellinus), Common pipistrelle (Pipistrellus pipistrellus), Nathusius’s pipistrelle (P. nathusii), Brown long-eared bat (Plecotus auritus), Common noctule (Nyctalus noctula), Parti-coloured bat (Vesperilio murinus) (Vespertilionidae), Common bent-wing bat (Miniopterus schreibersii) (Miniopteridae), European free-tailed bat (Tadarida teniotis) (Molossidae) have all been documented as being infected by EBLV-1 [47,48,72,73,74]. Regardless of the high number of bat hosts recorded for EBLV-1, EBVL-2 is restricted to Myotis daubentonii and M. dasycneme [44,46]. KBLV was found only in Myotis brandtii [24] and BBLV only in M. nattereri [74,75]. However, the phylogeny of the Natterer’s bat group is more complex. In Europe, M. nattereri is composed of M. escalerai (Iberia), M. species A (Italy and parts of the Pyrenees), M. species B (Northwest Africa), M. species C (Corsica), and the nominal form, M. nattereri present across the rest of Europe [76,77,78]. According to [79] BBLV-positive bats in Germany and France were of the nominal form. [80] provided a detailed phylogenetic analysis and reported signs of repeated hybridization between the Natterer’s bat lineages [80], with southern France a probable contact zone between M. species and M. nattereri. Additional research could reveal the potential of BBLV to spread to other Natterer’s bat lineages.
The Common bent-wing bat (Miniopterus schreibersii) has been proven to be a host to IRKV and DUVV from Phylogroup I, and WCBV and LLEBV from Phylogroup III. The species seems to be a universal reservoir for both phylogroups not only in Europe but also in Asia and Africa [35,81], due to the fact that M. schreibersii is a strictly cave-dwelling species [76,82] capable of long-distance migration [83]. The most abundant European lyssavirus group—EBLV-1 has still not been detected in the Common bent-wing bat despite its close phylogenetic relationship to DUVV.
5. Transmission routes of bat lyssaviruses
5.1. Bat intra- and cross-species transmission
The main transmission route of rabies viruses is via a bite from the host and the virions released into the saliva during the clinical period of rabies or/and during the end of the incubation period [84,85,86]. All lineages from Phylogroup I are transmitted by bats whereas the classical rabies virus—RABV has adapted to spread via carnivores and through bat species restricted to the Americas [21]. In Europe, EBLV-1 is the most prevalent lyssavirus among the Chiroptera, due to bat colony sizes, species richness, and the presence of migratory species. [87], suggested that in the roost the main factor for virus spillover is the large number of species and their individual mobility. However, roost sizes less than approximately 200 individuals are not enough for efficient maintenance of the lyssavirus infection [88]. On the other hand, migrant species such as Pipistrellus nathusii, Nyctalus noctula, N. leisleri and Miniopterus schreibersi can play a key role for the dispersal of EBLV-1 in Europe, and M. schreibersi also for WCBV and LLEBV [83,89,90,91]. These species are capable of flying long distances between summer and winter roosts and increasing the lyssavirus geographical range via cross-species transition is an expected scenario. Consequently, seasonal bat movements between hibernacula, breeding and mating sites are an important prerequisite for successful virus cross-species spillover. For instance, hibernation is characterized by decreased activity in the bat immune system and metabolism [92,93], thereby extending incubation periods and allowing virus persistence between transmission periods. Based on the transcriptomic responses of bat cells to EBVL-1 [94], concluded that the lack of bat cell reaction to infection in conditions simulating hibernation may contribute to the virus tolerance or persistence in bats. In addition, long hibernation roosts could facilitate transmission of EBLV-1 between geographically separated breeding populations [19]. This statement is also supported by a study focused on RABV in the Americas, where Myotis species have been found to harbor a diverse range of RABV variants, suggesting that increased contact between species increases viral transmission [68]. Breeding colonies are a good site to understand intraspecies transmission of antibodies via intra-uterine transfer, or viral transmissions as a result of biting or daily communal grooming via antibodies intra-uterine transfer [95] or biting during daily grooming [19,96]. Nerveless, characterizing EBLV-1 dynamics in juvenile bats is difficult due to unequal equilibration between them and adults and the chance that mothers can transfer antibodies to them via the placenta or during lactation [90,97]. The most efficient intra- and interspecific lyssavirus transmission route is during the mating period, due to aggressive male behavior when defending territory against other males and during mating with females [19]. In support of this assertion most cases of bat rabies in Europe peak in August and September [19,85,98].
Bats naturally infected with RABV have the virus in their nasal mucosa, leading to the proposition that airborne transmission of RABV between bats living in enclosed areas is possible [99,100,101]. Laboratory experiments showed that aerosols of the RABV virus were successfully overcome by bats but were fatal for the majority of experimental mice [102]. Johnson et al. [103] conducted a similar experiment with EBLV-2, where mice were intranasally inoculated and two of them developed the disease between 16- and 19-days post-infection. In addition, four cases of human rabies infection via aerosols were reported between 1956 and 1977 in a USA cave [102]. Li et al. [104] discovered that within caves bat guano exhibited a great diversity of viral species.
5.2. Other vertebrates cross-species transmission
Seidlova et al. [48] reported that lyssavirus seroprevalence in Palearctic bats is 3.2%, with 2.0% in Europe. Due to the adequatly taken measures for terestrial mammals rabies eradication in Europe [105],bats remain potental lyssavirus reservoir for humans in Europe [75]. Even so transmission is rare, the most recent case of bat-human spillover was in southwest central France in 2019 when a human male died from EBLV-1 (https://www.zmescience.com/ecology/animals-ecology/extremely-rare-case-of-death-from-bat-rabies-in-france/), one human died from EBLV-2 in Finland in 1985, and a third in the UK in 2002 [46]. Due to growing urbanization, pets present an expected intermediate host for bat-to-human lyssavirus transmission, and both EBLV-1 and WCBV have been found in cats in France [106] and Italy [49,107]. Experimental data suggests that the infectious dose for lyssaviruses is low and certainly some human infections, especially those involving transmission from bats, support this finding [108].
5.3. Within the contact zone of the Palearctic and Ethiopian realms—potential scenarios
Heretofore, Phylogroup II was only known from the African continent in two bat families: Pteropodidae and Hipposideridae [28,109,110]. LBV was isolated from the Straw-coloured fruit bat (Eidolon helvum) and Egyptian fruit bat (Rousettus aegyptiacus), and SHIBV from the Striped leaf-nosed bat (Macronycteris vittatus).
Compared with other Pteropodids, Rousettus aegyptiacus has the northernmost distribution, reaching Cyprus and southern Turkey [111], and roosts in a variety of underground sites [112] with other Palearctic species [113]. As a rule, cave-dwelling bats form large summer and winter colonies [82], where cross-species pathogen transmission is common [114]. A recent study [115] has shown that the families Rhinolophidae and Pteropodidae were a common factor in cross-species transmission of β-coronaviruses between continents due to their close phylogenetic relationship. This might also be applicable across the contact zones of different zoogeographic realms, e. g., Africa (Ethiopian realm) and Europe (Palearctic realm). Considering that the distributions of Rousettus aegyptiacus and Rhinolophus ferrumequinum overlap in the Southeastern zone of the Palearctic, on the border with the Ethiopian realm (Figure 3), the successful transfer of lyssaviruses from Phylogroup II via the Egyptian fruit bat to other cave-dwelling species, e. g., R. ferrumequinum, could be a plausible scenario due to their close phylogenetic relationship within the suborder Yinpterochiroptera [16]. Furthermore, phylogeny can act as a biotic factor driving the occurrence of RNA virus cross-species transmission between closely related host species [116]. For example, R. aegyptiacus and various species in the genus Rhinolophus are known to be successful virus reservoirs of β-coronavirus, Marburg virus, Henipavirus, Orthorubulavirus, Pararubulavirus and Lyssaviruses [14,28,97].
Additional molecular investigations are needed to solve the relationship between Phylogroup II and the other phylogroups. This is an important consideration since Phylogroup II is spread within the families Pteropodidae and Hipposideridae, both of which are within the suborder Yinpterochiroptera and closely related to the family Rhinolophidae, which includes cave-dwelling species widely distributed across Europe. Taking into account the close phylogenetic relationship between these bat families, the spread of the lyssavirus Phylogroup II to the Palearctic realm is a possible scenario within the contact zone with the Ethiopian realm. From the epidemiological point of view, these bat families and the contact zone are significant for cross-species bat transmission routes of β-coronaviruses.
6. Conclusions
Bat lyssaviruses in Europe belong to two phylogroups: I and III, based on the molecular analyses of rabies G and N genes. The phylogeny of the lyssaviruses is closely related to the phylogeography of Palearctic bat species. EBLV-1 has been detected in species in the superfamilies Yangochiroptera and Yinpterochiroptera, which demonstrates the virus plasticity between highly divergent bat lineages. In contrast, EBLV-2, KBLV and BBLV have limited distributions in Europe, with their reservoirs restricted to Myotis spp. Phylogenetic relationships within Phylogroup I demonstrate that EBLV-1 and DUVV are closely related and spillover between different bat families, at a large distance from each other, is possible (Europe—Africa), while the second closest group, EBLV-2, KBLV and BBLV has a restricted distribution in just one bat genus. The existing rabies vaccines developed for RABV eradication are effective against lyssaviruses from Phylogroup I but offer little to no protection against phylogroup II and III viruses.
Living under the shadow of the recent COVID-19 pandemic, the detection and prevention of future pathogens is of crucial importance. The newly discovered bat lyssaviruses (BBLV, LLEBV and KBVL) and vesiculoviruses from the USA (SDRV1 and SDRV2) and China (YSBV, TYBV, and QZBV) warn us that the Rhabdoviridae represent an unexplored pathogen pool with new and yet to be described viruses potentially adverse to human health.
Supplementary Materials
Applicable.
Author Contributions
H. D. conceptualized and wrote the first draft of the review. P. H., J. Z., and J. P. reviewed and edited the manuscript. N. I-A., S. B., I. G., K. K., K. M., B. N., and P. O. contributed to developing and finalizing the review. H. D. and P. H. have equal contribution.
Funding
This research was funded by Bulgarian National Science Fund, Grant No. KП-06-H31/14.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Lee Knight for linguistic proofreading of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ruedi, M.; Stadelmann, B.; Gager, Y.; Douzery, E.J.P.; Francis, C.M.; Lin, L.-K.; Guillén-Servent, A.; Cibois, A. Molecular phylogenetic reconstructions identify East Asia as the cradle for the evolution of the cosmopolitan genus Myotis (Mammalia, Chiroptera). Mol. Phylogenetics Evol. 2013, 69, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Teeling, E.C.; Vernes, S.C.; Davalos, L.M.; Ray, D.A.; Gilbert, M.T.P.; Myers, E. ; K Consortium Bat Biology, Genomes, and the Bat1K Project: To Generate Chromosome-Level Genomes for All Living Bat Species. Annu. Rev. Anim. Biosci. 2018, 6, 23–46. [Google Scholar] [CrossRef]
- Teeling, E. C.; Jones, G.; Rossiter, S. J. Phylogeny, genes, and hearing: implications for the evolution of echolocation in bats. In Bat bioacoustics Springer, New York, 2016; pp. 25-54.
- Beltz, L. A. Bats and human health: Ebola, SARS, rabies and beyond. Jonh Wiley & Sons, Ltd. 2017; p. 371.
- Wackermannová, M.; Pinc, L.; Jebavý, L. Olfactory Sensitivity in Mammalian Species. Physiol. Res. 2016, 65, 369–390. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, G.S.; Adams, D.M.; Haghani, A.; Lu, A.T.; Zoller, J.; Breeze, C.E.; Arnold, B.D.; Ball, H.C.; Carter, G.G.; Cooper, L.N.; et al. DNA methylation predicts age and provides insight into exceptional longevity of bats. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.-Y.; Liang, L.; Zhu, Z.-H.; Zhou, W.-P.; Irwin, D.M.; Zhang, Y.-P. Adaptive evolution of energy metabolism genes and the origin of flight in bats. Proc. Natl. Acad. Sci. 2010, 107, 8666–8671. [Google Scholar] [CrossRef] [PubMed]
- Subudhi, S.; Rapin, N.; Misra, V. Immune System Modulation and Viral Persistence in Bats: Understanding Viral Spillover. Viruses 2019, 11, 192. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Baker, M.L.; Kulcsar, K.; Misra, V.; Plowright, R.; Mossman, K. Novel Insights Into Immune Systems of Bats. Front. Immunol. 2020, 11, 26. [Google Scholar] [CrossRef] [PubMed]
- Begeman, L.; Suu-Ire, R.; Banyard, A.C.; Drosten, C.; Eggerbauer, E.; Freuling, C.M.; Gibson, L.; Goharriz, H.; Horton, D.L.; Jennings, D.; et al. Experimental Lagos bat virus infection in straw-colored fruit bats: A suitable model for bat rabies in a natural reservoir species. PLOS Neglected Trop. Dis. 2020, 14, e0008898. [Google Scholar] [CrossRef]
- Irving, A.T.; Ahn, M.; Goh, G.; Anderson, D.E.; Wang, L.-F. Lessons from the host defences of bats, a unique viral reservoir. Nature 2021, 589, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Brook, C.E.; Dobson, A.P. Bats as ‘special’ reservoirs for emerging zoonotic pathogens. Trends Microbiol. 2015, 23, 172–180. [Google Scholar] [CrossRef]
- Hayman, D.T. Bats as Viral Reservoirs. Annu. Rev. Virol. 2016, 3, 77–99. [Google Scholar] [CrossRef] [PubMed]
- Latinne, A.; Hu, B.; Olival, K.J.; Zhu, G.; Zhang, L.; Li, H.; Chmura, A.A.; Field, H.E.; Zambrana-Torrelio, C.; Epstein, J.H.; et al. Origin and cross-species transmission of bat coronaviruses in China. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- EUROBATS. Action Plan for the Conservation of Bat Species in the European Union 2016–2021; Inf.EUROBATS.AC21. B: EUROBATS, 2006.
- Teeling, E.C.; Springer, M.S.; Madsen, O.; Bates, P.; O’Brien, S.J.; Murphy, W.J. A Molecular Phylogeny for Bats Illuminates Biogeography and the Fossil Record. Science 2005, 307, 580–584. [Google Scholar] [CrossRef]
- Kohl, C.; Kurth, A. European Bats as Carriers of Viruses with Zoonotic Potential. Viruses 2014, 6, 3110–3128. [Google Scholar] [CrossRef]
- Smreczak, M.; Orłowska, A.; Marzec, A.; Trębas, P.; Müller, T.; Freuling, C. M.; Żmudziński, J. F. Bokeloh bat lyssavirus isolation in a Natterer’s bat, Poland. Zoonoses Public Health. 2018, 65(8), 1015–1019. [Google Scholar] [CrossRef]
- Vos, A.; Kaipf, I.; Denzinger, A.; Fooks, A.R.; Johnson, N.; Müller, T. European bat lyssaviruses — an ecological enigma. Acta Chiropterologica 2007, 9, 283–296. [Google Scholar] [CrossRef]
- Banyard, A. C.; Hayman, D. ; Johnson, McElhinney, N. L; Fooks, A. R. Bats and lyssaviruses. Adv. Virus Res. 2011, 79, 239–89. [Google Scholar] [CrossRef]
- Rupprecht, C.E.; Freuling, C.M.; Mani, R.S.; Palacios, C.; Sabeta, C.T.; Ward, M. A history of rabies—The foundation for global canine rabies elimination. In Rabies, Scientific Basis of the Disease and Its Management, 4th ed.; Fooks, A.R., Jackson, A.C., Eds.; Academic Press: Cambridge, MA, USA, 2020; ISBN 978-0-12-818705-0. [Google Scholar] [CrossRef]
- Afonso, C.L.; Amarasinghe, G.K.; Bányai, K.; Bào, Y.; Basler, C.F.; Bavari, S.; Bejerman, N.; Blasdell, K.R.; Briand, F.-X.; Briese, T.; et al. Taxonomy of the order Mononegavirales: update 2016. Arch. Virol. 2016, 161, 2351–2360. [Google Scholar] [CrossRef]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Alfenas-Zerbini, P.; Dempsey, D.M.; Dutilh, B.E.; García, M.L.; Hendrickson, R.C.; et al. Recent changes to virus taxonomy ratified by the International Committee on Taxonomy of Viruses (2022). Arch. Virol. 2022, 167, 2429–2440. [Google Scholar] [CrossRef]
- Nokireki, T.; Tammiranta, N.; Kokkonen, U.-M.; Kantala, T.; Gadd, T. Tentative novel lyssavirus in a bat in Finland. Transbound. Emerg. Dis. 2018, 65, 593–596. [Google Scholar] [CrossRef]
- Coertse, J.; Markotter, W.; le Roux, K.; Stewart, D.; Sabeta, C.T.; Nel, L.H. New isolations of the rabies-related Mokola virus from South Africa. BMC Veter- Res. 2016, 13, 1–8. [Google Scholar] [CrossRef]
- Biswas, M.R.; Alzubaidi, M.S.; Shah, U.; Abd-Alrazaq, A.A.; Shah, Z. A Scoping Review to Find out Worldwide COVID-19 Vaccine Hesitancy and Its Underlying Determinants. Vaccines 2021, 9, 1243. [Google Scholar] [CrossRef]
- Marston, D.A.; Horton, D.L.; Ngeleja, C.; Hampson, K.; McElhinney, L.M.; Banyard, A.C.; Haydon, D.; Cleaveland, S.; Rupprecht, C.E.; Bigambo, M.; et al. Ikoma lyssavirus, highly divergent novel lyssavirus in an African civet. Emerg. Infect. Dis. 2012, 18, 664–667. [Google Scholar] [CrossRef] [PubMed]
- Markotter, W.; Kgaladi, J.; Nel, L.H.; Marston, D.; Wright, N.; Coertse, J.; Müller, T.F.; Sabeta, C.T.; Fooks, A.R.; Freuling, C.M. Diversity and Epidemiology of Mokola Virus. PLoS Negl. Trop. Dis. 2013, 7, e2511. [Google Scholar]
- Sabeta, C.T.; Markotter, W.; Mohale, D.K.; Shumba, W.; Wandeler, A.I.; Nel, L.H. Mokola Virus in Domestic Mammals, South Africa. Emerg. Infect. Dis. 2007, 13, 1371–1373. [Google Scholar] [CrossRef] [PubMed]
- Rupprecht, C.; Kuzmin, I.; Meslin, F. Lyssaviruses and rabies: current conundrums, concerns, contradictions and controversies. F1000Research 2017, 6, 184. [Google Scholar] [CrossRef]
- Hu, S.-C.; Hsu, C.-L.; Lee, M.-S.; Tu, Y.-C.; Chang, J.-C.; Wu, C.-H.; Lee, S.-H.; Ting, L.-J.; Tsai, K.-R.; Cheng, M.-C.; et al. Lyssavirus in Japanese Pipistrelle, Taiwan. Emerg. Infect. Dis. 2018, 24, 782–785. [Google Scholar] [CrossRef] [PubMed]
- Calvelage, S.; Tammiranta, N.; Nokireki, T.; Gadd, T.; Eggerbauer, E.; Zaeck, L.M.; Potratz, M.; Wylezich, C.; Höper, D.; Müller, T.; et al. Genetic and Antigenetic Characterization of the Novel Kotalahti Bat Lyssavirus (KBLV). Viruses 2021, 13, 69. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.-C.; Hsu, C.-L.; Lee, F.; Tu, Y.-C.; Chen, Y.-W.; Chang, J.-C.; Hsu, W.-C. Novel Bat Lyssaviruses Identified by Nationwide Passive Surveillance in Taiwan, 2018–2021. Viruses 2022, 14, 1562. [Google Scholar] [CrossRef]
- Nel, L. H.; Rupprecht, C. E. Emergence of lyssaviruses in the Old World: the case of Africa. Curr Top Microbiol Immunol. 2007, 315, 161–93. [Google Scholar] [CrossRef]
- Hayman, D.T.S.; Fooks, A.R.; Marston, D.A.; Garcia-R, J.C. The Global Phylogeography of Lyssaviruses—Challenging the ‘Out of Africa’ Hypothesis. PLOS Neglected Trop. Dis. 2016, 10, e0005266. [Google Scholar] [CrossRef] [PubMed]
- Longdon, B.; Murray, G.G.R.; Palmer, W.J.; Day, J.P.; Parker, D.J.; Welch, J.J.; Obbard, D.J.; Jiggins, F.M. The evolution, diversity, and host associations of rhabdoviruses. Virus Evol. 2015, 1, vev014. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.J.; Firth, C.; Widen, S.G.; Blasdell, K.R.; Guzman, H.; Wood, T.G.; Paradkar, P.N.; Holmes, E.C.; Tesh, R.B.; Vasilakis, N. Evolution of Genome Size and Complexity in the Rhabdoviridae. PLOS Pathog. 2015, 11, e1004664–e1004664. [Google Scholar] [CrossRef]
- Caraballo, D.A.; Lema, C.; Novaro, L.; Gury-Dohmen, F.; Russo, S.; Beltrán, F.J.; Palacios, G.; Cisterna, D.M. A Novel Terrestrial Rabies Virus Lineage Occurring in South America: Origin, Diversification, and Evidence of Contact between Wild and Domestic Cycles. Viruses 2021, 13, 2484. [Google Scholar] [CrossRef]
- Singh, R.; Singh, K.P.; Cherian, S.; Saminathan, M.; Kapoor, S.; Reddy, G.M.; Panda, S.; Dhama, K. Rabies—epidemiology, pathogenesis, public health concerns and advances in diagnosis and control: a comprehensive review. Veter- Q. 2017, 37, 212–251. [Google Scholar] [CrossRef]
- Potratz, M.; Zaeck, L.M.; Weigel, C.; Klein, A.; Freuling, C.M.; Müller, T.; Finke, S. Neuroglia infection by rabies virus after anterograde virus spread in peripheral neurons. Acta Neuropathol. Commun. 2020, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Baghi, H.B.; Rupprecht, C.E. Notes on three periods of rabies focus in the Middle East: From progress during the cradle of civilization to neglected current history. Zoonoses Public Heal. 2020, 68, 697–703. [Google Scholar] [CrossRef]
- E Rupprecht, C.; Turmelle, A.; Kuzmin, I.V. A perspective on lyssavirus emergence and perpetuation. Curr. Opin. Virol. 2011, 1, 662–670. [Google Scholar] [CrossRef]
- Fooks, A.R.; Brookes, S.M.; Johnson, N.; McELHINNEY, L.M.; Hutson, A.M. European bat lyssaviruses: an emerging zoonosis. Epidemiology Infect. 2003, 131, 1029–1039. [Google Scholar] [CrossRef]
- Schatz, J.; Fooks, A.R.; McElhinney, L.; Horton, D.; Echevarria, J.; Vázquez-Moron, S.; Kooi, E.A.; Rasmussen, T.B.; Müller, T.; Freuling, C.M. Bat Rabies Surveillance in Europe. Zoonoses Public Heal. 2012, 60, 22–34. [Google Scholar] [CrossRef]
- McElhinney, L.M.; Marston, D.A.; Wise, E.L.; Freuling, C.M.; Bourhy, H.; Zanoni, R.; Moldal, T.; Kooi, E.A.; Neubauer-Juric, A.; Nokireki, T.; et al. Molecular Epidemiology and Evolution of European Bat Lyssavirus 2. Int. J. Mol. Sci. 2018, 19, 156. [Google Scholar] [CrossRef] [PubMed]
- Šimić, I.; Lojkić, I.; Krešić, N.; Cliquet, F.; Picard-Meyer, E.; Wasniewski, M.; Ćukušić, A.; Zrnčić, V.; Bedeković, T. Molecular and serological survey of lyssaviruses in Croatian bat populations. BMC Veter- Res. 2018, 14, 274. [Google Scholar] [CrossRef]
- Seidlova, V.; Zukal, J.; Brichta, J.; Anisimov, N.; Apoznański, G.; Bandouchova, H.; Bartonička, T.; Berková, H.; Botvinkin, A.D.; Heger, T.; et al. Active surveillance for antibodies confirms circulation of lyssaviruses in Palearctic bats. BMC Veter- Res. 2020, 16, 1–9. [Google Scholar] [CrossRef]
- Coxon, C.; McElhinney, L.; Pacey, A.; Gauntlett, F.; Holland, S. Preliminary Outbreak Assessment: Rabies in a Cat in Italy. Department for Environment, Food and Rural Affairs, Animal and Plant Health Agency, Advice Services—International Disease Monitoring. 2020 Available online at: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/897070/rabies-cat-italy-poa.
- Kuzmin, I.V.; Hughes, G.J.; Botvinkin, A.D.; Orciari, L.A.; Rupprecht, C.E. Phylogenetic relationships of Irkut and West Caucasian bat viruses within the Lyssavirus genus and suggested quantitative criteria based on the N gene sequence for lyssavirus genotype definition. Virus Res. 2005, 111, 28–43. [Google Scholar] [CrossRef]
- Freuling, C. Novel Lyssavirus in Natterer’s Bat, Germany. Emerg. Infect. Dis. 2011, 17, 1519–22. [Google Scholar] [CrossRef] [PubMed]
- Arechiga Ceballos, N.; Vazquez Moron, S.; Berciano, J.M.; Nicolas, O.; Aznar Lopez, C.; Juste, J.; Rodriguez Nevado, C.; Aguilar Setien, A.; Echevarria, J.E. Novel lyssavirus in bat, Spain. Emerg. Infect. Dis. 2013, 19, 793–795. [Google Scholar] [CrossRef]
- Davis, P.L.; Holmes, E.C.; Larrous, F.; Van der Poel, W.H.M.; Tjørnehøj, K.; Alonso, W.J.; Bourhy, H. Phylogeography, Population Dynamics, and Molecular Evolution of European Bat Lyssaviruses. J. Virol. 2005, 79, 10487–10497. [Google Scholar] [CrossRef]
- Mingo-Casas, P.; Sandonís, V.; Obón, E.; Berciano, J.M.; Vázquez-Morón, S.; Juste, J.; Echevarría, J.E. First cases of European bat lyssavirus type 1 in Iberian serotine bats: Implications for the molecular epidemiology of bat rabies in Europe. PLOS Neglected Trop. Dis. 2018, 12, e0006290. [Google Scholar] [CrossRef]
- Jakava-Viljanen, M.; Nokireki, T.; Sironen, T.; Vapalahti, O.; Sihvonen, L.; Huovilainen, A. Erratum to: Evolutionary trends of European bat lyssavirus type 2 including genetic characterization of Finnish strains of human and bat origin 24 years apart. Arch. Virol. 2015, 160, 1875–1875. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.L.; Aegerter, J.N.; Brookes, S.M.; McElhinney, L.M.; Jones, G.; Smith, G.C.; Fooks, A.R. TARGETED SURVEILLANCE FOR EUROPEAN BAT LYSSAVIRUSES IN ENGLISH BATS (2003–06). J. Wildl. Dis. 2009, 45, 1030–1041. [Google Scholar] [CrossRef]
- Dietzgen, R.G.; Kondo, H.; Goodin, M.M.; Kurath, G.; Vasilakis, N. The family Rhabdoviridae: mono- and bipartite negative-sense RNA viruses with diverse genome organization and common evolutionary origins. Virus Res. 2017, 227, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Lelli, D.; Prosperi, A.; Moreno, A.; Chiapponi, C.; Gibellini, A.M.; De Benedictis, P.; Leopardi, S.; Sozzi, E.; Lavazza, A. Isolation of a novel Rhabdovirus from an insectivorous bat (Pipistrellus kuhlii) in Italy. Virol. J. 2018, 15, 37. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.J.; Dietzgen, R.G.; Joubert, D.A.; Blasdell, K.R. Rhabdovirus accessory genes. Virus Res. 2011, 162, 110–125. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Wu, X.; Tordo, N.; Rupprecht, C.E. Complete genomes of Aravan, Khujand, Irkut and West Caucasian bat viruses, with special attention to the polymerase gene and non-coding regions. Virus Res. 2008, 136, 81–90. [Google Scholar] [CrossRef]
- Badrane, H.; Bahloul, C.; Perrin, P.; Tordo, N. Evidence of TwoLyssavirusPhylogroups with Distinct Pathogenicity and Immunogenicity. J. Virol. 2001, 75, 3268–3276. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Orciari, L.A.; Arai, Y.T.; Smith, J.S.; Hanlon, C.A.; Kameoka, Y.; Rupprecht, C.E. Bat lyssaviruses (Aravan and Khujand) from Central Asia: phylogenetic relationships according to N, P and G gene sequences. Virus Res. 2003, 97, 65–79. [Google Scholar] [CrossRef]
- Kuzmin, I.; Novella, I.; Dietzgen, R.; Padhi, A.; Rupprecht, C. The rhabdoviruses: Biodiversity, phylogenetics, and evolution. Infect. Genet. Evol. 2009, 9, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press (OUP): Oxford, Oxfordshire, United Kingdom, 2000. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Gunawardena, P.S.; Marston, D.A.; Ellis, R.J.; Wise, E.L.; Karawita, A.C.; Breed, A.C.; McElhinney, L.M.; Johnson, N.; Banyard, A.C.; Fooks, A.R. Lyssavirus in Indian Flying Foxes, Sri Lanka. Emerg. Infect. Dis. 2016, 22, 1456–1459. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, I.V.; Mayer, A.E.; Niezgoda, M.; Markotter, W.; Agwanda, B.; Breiman, R.F.; Rupprecht, C.E. Shimoni bat virus, a new representative of the Lyssavirus genus. Virus Res. 2010, 149, 197–210. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Bozick, B.; Guagliardo, S.A.; Kunkel, R.; Shak, J.R.; Tong, S.; Rupprecht, C.E. Bats, emerging infectious diseases, and the rabies paradigm revisited. Emerg. Heal. Threat. J. 2011, 4, 7159. [Google Scholar] [CrossRef]
- Aréchiga -Ceballos, N. A.; Morón, S. V.; Berciano, J. M.; Nicolás, O.; López, C. A.; Juste, J.; Nevado, C. R.; Setién, Á. A.; Echevarría, J. E. Novel lyssavirus in bat Spain. Emerg. Infect. Dis. 2013, 19(5), 793–795. [Google Scholar] [CrossRef]
- Amengual, B.; E Whitby, J.; Cobo, J.S.; King, A.; Bourhy, H. Evolution of European bat lyssaviruses. J. Gen. Virol. 1997, 78, 2319–2328. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, I. V.; Rupprecht, C.E. Bats and Viruses: A New Frontier of Emerging Infectious Diseases. Bat Lyssaviruses. Published by John Wiley and Sons, Inc., Hoboken, New Jersey Published simultaneously in Canada, 2015 pp. -97. [CrossRef]
- Echevarrı́a, J.E.; Avellón, A.; Juste, J.; Vera, M.; Ibáñez, C. Screening of Active Lyssavirus Infection in Wild Bat Populations by Viral RNA Detection on Oropharyngeal Swabs. J. Clin. Microbiol. 2001, 39, 3678–3683. [Google Scholar] [CrossRef] [PubMed]
- Serra-Cobo, J.; Amengual, B.; Abellan, C.; Bourhy, H. European bat Lyssavirus infection in Spanish bat populations. Emerg. Infect. Dis. 2002, 8, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Picard-Meyer, E.; Servat, A.; Robardet, E.; Moinet, M.; Borel, C.; Cliquet, F. Isolation of Bokeloh bat lyssavirus in Myotis nattereri in France. Arch. Virol. 2013, 158, 2333–2340. [Google Scholar] [CrossRef]
- Parize, P.; Robledo, I.C.T.; Cervantes-Gonzalez, M.; Kergoat, L.; Larrous, F.; Serra-Cobo, J.; Dacheux, L.; Bourhy, H. Circumstances of Human–Bat interactions and risk of lyssavirus transmission in metropolitan France. Zoonoses Public Heal. 2020, 67, 774–784. [Google Scholar] [CrossRef] [PubMed]
- Salicini, I.; Ibáñez, C.; Juste, J. Multilocus phylogeny and species delimitation within the Natterer’s bat species complex in the Western Palearctic. Mol. Phylogenetics Evol. 2011, 61, 888–898. [Google Scholar] [CrossRef] [PubMed]
- Salicini, I.; Ibáñez, C.; Juste, J. Deep differentiation between and within Mediterranean glacial refugia in a flying mammal, the Myotis nattereri bat complex. J. Biogeogr. 2012, 40, 1182–1193. [Google Scholar] [CrossRef]
- Puechmaille, S.J.; Allegrini, B.; Boston, E.S.; Dubourg-Savage, M.-J.; Evin, A.; Knochel, A.; Le Bris, Y.; Lecoq, V.; Lemaire, M.; Rist, D.; et al. Genetic analyses reveal further cryptic lineages within the Myotis nattereri species complex. Mamm. Biol. 2012, 77, 224–228. [Google Scholar] [CrossRef]
- Eggerbauer, E.; Pfaff, F.; Finke, S.; Höper, D.; Beer, M.; Mettenleiter, T.C.; Nolden, T.; Teifke, J.-P.; Müller, T.; Freuling, C.M. Comparative analysis of European bat lyssavirus 1 pathogenicity in the mouse model. PLOS Neglected Trop. Dis. 2017, 11, e0005668. [Google Scholar] [CrossRef]
- Çoraman, E.; Dietz, C.; Hempel, E.; Ghazaryan, A.; Levin, E.; Presetnik, P.; Zagmajster, M.; Mayer, F. Reticulate evolutionary history of a Western Palaearctic Bat Complex explained by multiple mtDNA introgressions in secondary contacts. J. Biogeogr. 2019, 46, 343–354. [Google Scholar] [CrossRef]
- Biswas, M.R.; Alzubaidi, M.S.; Shah, U.; Abd-Alrazaq, A.A.; Shah, Z. A Scoping Review to Find out Worldwide COVID-19 Vaccine Hesitancy and Its Underlying Determinants. Vaccines 2021, 9, 1243. [Google Scholar] [CrossRef] [PubMed]
- Dietz, C.; von Helversen, O.; Nill, D. Bats of Britain. Europe and Northwest Africa. A and C Black, London. 2009. [Google Scholar]
- Wright, P.G.R.; Newton, J.; Agnelli, P.; Budinski, I.; Di Salvo, I.; Flaquer, C.; Fulco, A.; Georgiakakis, P.; Martinoli, A.; Mas, M.; et al. Hydrogen isotopes reveal evidence of migration of Miniopterus schreibersii in Europe. BMC Ecol. 2020, 20, 1–7. [Google Scholar] [CrossRef]
- Freuling, C.; Vos, A.; Johnson, N.; Kaipf, I.; Denzinger, A.; Neubert, L.; Mansfield, K.; Hicks, D.; Nuñez, A.; Tordo, N.; et al. Experimental infection of serotine bats (Eptesicus serotinus) with European bat lyssavirus type 1a. J. Gen. Virol. 2009, 90, 2493–2502. [Google Scholar] [CrossRef]
- de Thoisy, B.; Bourhy, H.; Delaval, M.; Pontier, D.; Dacheux, L.; Darcissac, E.; Donato, D.; Guidez, A.; Larrous, F.; Lavenir, R.; et al. Bioecological Drivers of Rabies Virus Circulation in a Neotropical Bat Community. PLOS Neglected Trop. Dis. 2016, 10, e0004378–e0004378. [Google Scholar] [CrossRef]
- Bonnaud, E.M.; Troupin, C.; Dacheux, L.; Holmes, E.C.; Monchatre-Leroy, E.; Tanguy, M.; Bouchier, C.; Cliquet, F.; Barrat, J.; Bourhy, H. Comparison of intra- and inter-host genetic diversity in rabies virus during experimental cross-species transmission. PLOS Pathog. 2019, 15, e1007799. [Google Scholar] [CrossRef]
- Colombi, D.; Serra-Cobo, J.; Métras, R.; Apolloni, A.; Poletto, C.; López-Roig, M.; Bourhy, H.; Colizza, V. Mechanisms for lyssavirus persistence in non-synanthropic bats in Europe: insights from a modeling study. Sci. Rep. 2019, 9, 537. [Google Scholar] [CrossRef]
- Horton, D.L.; Breed, A.C.; Arnold, M.E.; Smith, G.C.; Aegerter, J.N.; McElhinney, L.M.; Johnson, N.; Banyard, A.C.; Raynor, R.; Mackie, I.; et al. Between roost contact is essential for maintenance of European bat lyssavirus type-2 in Myotis daubentonii bat reservoir: ‘The Swarming Hypothesis’. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tájek, P.; Tájková, P. A long distance migration in Nyctalus leisleri from the Czech Republic to southern France (Chiroptera: Vespertilionidae). Lynx new Ser. 2021, 51, 223–226. [Google Scholar] [CrossRef]
- Dundarova, H.; Michev, B.; Pandourski, I. Bats over the Western Black Sea open water area. Acta Zool. Bulg. 2021, 73(4), 543–546. [Google Scholar]
- Vega, S.; Lorenzo-Rebenaque, L.; Marin, C.; Domingo, R.; Fariñas, F. Tackling the Threat of Rabies Reintroduction in Europe. Front. Veter- Sci. 2021, 7. [Google Scholar] [CrossRef]
- Bouma, H.R.; Carey, H.V.; Kroese, F.G.M. Hibernation: the immune system at rest? J. Leukoc. Biol. 2010, 88, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Field, K.A.; Sewall, B.J.; Prokkola, J.M.; Turner, G.G.; Gagnon, M.F.; Lilley, T.M.; White, J.P.; Johnson, J.S.; Hauer, C.L.; Reeder, D.M. Effect of torpor on host transcriptomic responses to a fungal pathogen in hibernating bats. Mol. Ecol. 2018, 27, 3727–3743. [Google Scholar] [CrossRef]
- Constantine, D.G. Bat rabies and other lyssavirus infections. Reston, Va., U.S. Geological Survey Circular 1329. 2009, 68 p.
- Harazim, M.; Perrot, J.; Varet, H.; Bourhy, H.; Lannoy, J.; Pikula, J.; Seidlová, V.; Dacheux, L.; Martínková, N. Transcriptomic responses of bat cells to European bat lyssavirus 1 infection under conditions simulating euthermia and hibernation. BMC Immunol. 2023, 24, 1–13. [Google Scholar] [CrossRef]
- Gilbert, A. T.; McCracken, G. F.; Sheeler, L. L.; Muller, L. I.; O’Rourke, D.; Kelch, W. J.; New Jr., J. C. Rabies surveillance among bats in Tennessee, USA, 1996–2010. J. Wildl. Dis. 2015, 51(4), 821-832.
- Jin J, Lu Z, Li Y, Cowart LA, Lopes-Virella MF, Huang Y. Docosahexaenoic acid antagonizes the boosting effect of palmitic acid on LPS inflammatory signaling by inhibiting gene transcription and ceramide synthesis. PLoS One. 2018, 13: e0193343. [CrossRef]
- Shipley, R.; Wright, E.; Selden, D.; Wu, G.; Aegerter, J.; Fooks, A.R.; Banyard, A.C. Bats and Viruses: Emergence of Novel Lyssaviruses and Association of Bats with Viral Zoonoses in the EU. Trop. Med. Infect. Dis. 2019, 4, 31. [Google Scholar] [CrossRef]
- Constantine, D.G.; Emmons, R.W.; Woodie, J.D. Rabies Virus in Nasal Mucosa of Naturally Infected Bats. Science 1972, 175, 1255–1256. [Google Scholar] [CrossRef] [PubMed]
- Allendorf, S.D.; Cortez, A.; Heinemann, M.B.; Harary, C.M.A.; Antunes, J.M.A.; Peres, M.G.; Vicente, A.F.; Sodré, M.M.; da Rosa, A.R.; Megid, J. Rabies virus distribution in tissues and molecular characterization of strains from naturally infected non-hematophagous bats. Virus Res. 2012, 165, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Fisher, C.R.; Streicker, D.G.; Schnell, M.J. The spread and evolution of rabies virus: conquering new frontiers. Nat. Rev. Microbiol. 2018, 16, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.D.; Rudd, R.J.; Bowen, R.A. Effects of Aerosolized Rabies Virus Exposure on Bats and Mice. J. Infect. Dis. 2007, 195, 1144–1150. [Google Scholar] [CrossRef]
- Johnson, N.; Phillpotts, R.; Fooks, A.R. Airborne transmission of lyssaviruses. J. Med Microbiol. 2006, 55, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Victoria, J.G.; Wang, C.; Jones, M.; Fellers, G.M.; Kunz, T.H.; Delwart, E. Bat Guano Virome: Predominance of Dietary Viruses from Insects and Plants plus Novel Mammalian Viruses. J. Virol. 2010, 84, 6955–6965. [Google Scholar] [CrossRef] [PubMed]
- Lojkić, I.; Šimić, I.; Bedeković, T.; Krešić, N. Current Status of Rabies and Its Eradication in Eastern and Southeastern Europe. Pathogens 2021, 10, 742. [Google Scholar] [CrossRef] [PubMed]
- Dacheux, L.; Larrous, F.; Mailles, A.; Boisseleau, D.; Delmas, O.; Biron, C.; Bouchier, C.; Capek, I.; Muller, M.; Ilari, F.; et al. European Bat Lyssavirus Transmission among Cats, Europe. Emerg. Infect. Dis. 2009, 15, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Leopardi, S.; Barneschi, E.; Manna, G.; Zecchin, B.; Priori, P.; Drzewnioková, P.; Festa, F.; Lombardo, A.; Parca, F.; Scaravelli, D.; et al. Spillover of West Caucasian Bat Lyssavirus (WCBV) in a Domestic Cat and Westward Expansion in the Palearctic Region. Viruses 2021, 13, 2064. [Google Scholar] [CrossRef]
- Banyard, A.C.; Evans, J.S.; Luo, T.R.; Fooks, A.R. Lyssaviruses and Bats: Emergence and Zoonotic Threat. Viruses 2014, 6, 2974–2990. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Niezgoda, M.; Franka, R.; Agwanda, B.; Markotter, W.; Beagley, J.C.; Urazova, O.Y.; Breiman, R.F.; Rupprecht, C.E. Lagos Bat Virus in Kenya. J. Clin. Microbiol. 2008, 46, 1451–1461. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Turmelle, A.S.; Agwanda, B.; Markotter, W.; Niezgoda, M.; Breiman, R.F.; Rupprecht, C.E.; Woldehanna, S.; Zimicki, S.; Nel, L.; et al. Commerson’s Leaf-Nosed Bat (Hipposideros commersoni) is the Likely Reservoir of Shimoni Bat Virus. Vector-Borne Zoonotic Dis. 2011, 11, 1465–1470. [Google Scholar] [CrossRef]
- Lučan, R.K.; Bartonička, T.; Benda, P.; Bilgin, R.; Jedlička, P.; Nicolaou, H.; Reiter, A.; Shohdi, W.M.; Šálek, M.; Řeřucha. ; et al. Reproductive seasonality of the Egyptian fruit bat (Rousettus aegyptiacus) at the northern limits of its distribution. J. Mammal. 2014, 95, 1036–1042. [Google Scholar] [CrossRef]
- Benda, P.; Abi-Said, M.; Bartonička, T.; Bilgin, R.; Faizolahi, K.; Lučan, R. K.; Nicolaou, H.; Raiter, A.; Shohdi, W. M.; Uhrin, M.; Horáček, I. Rousettus aegyptiacus (Pteropodidae) in the Palaearctic: list of records and revision of the distribution range. Vespertilio. 2011, 15, 3—36. [Google Scholar]
- Benda, P.; Abi Said, M. R.; Bou Jaoude, I.; Karanouh, R.; Lučan, R. K.; Sadek, R. ; Ševčik, Uhrin, M. ; Horáček, I. Bats (Mammalia: Chiroptera) of the Eastern Mediterranean and Middle East. Part 13. Review of distribution and ectoparasites of bats in Lebanon. Acta Soc. Zool. Bohem. 2016, 80, 207–316. [Google Scholar]
- Willoughby, A.R.; Phelps, K.L.; PREDICT Consortium; Olival, K. J. A Comparative Analysis of Viral Richness and Viral Sharing in Cave-Roosting Bats. Diversity 2017, 9, 35. [Google Scholar] [CrossRef]
- Wang, Z.; Huang, G.; Huang, M.; Dai, Q.; Hu, Y.; Zhou, J.; Wei, F. Global patterns of phylogenetic diversity and transmission of bat coronavirus. Sci. China Life Sci. 2022, 66, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Albery, G.F.; Becker, D.J.; Brierley, L.; Brook, C.E.; Christofferson, R.C.; Cohen, L.E.; Dallas, T.A.; Eskew, E.A.; Fagre, A.; Farrell, M.J.; et al. The science of the host–virus network. Nat. Microbiol. 2021, 6, 1483–1492. [Google Scholar] [CrossRef]
Figure 1.
Distribution of bat lyssaviruses in Europe. Abbreviations used: EBLV1—European bat-1 lyssavirus; EBLV2—European bat-2 lyssavirus; BBLV—Bokeloh bat lyssavirus; KBLV—Kotalahti bat lyssavirus; WCBV—West Caucasian bat lyssavirus; LLEBV—Lleida bat lyssavirus; Seropositive—Seropositive Blood samples.
Figure 1.
Distribution of bat lyssaviruses in Europe. Abbreviations used: EBLV1—European bat-1 lyssavirus; EBLV2—European bat-2 lyssavirus; BBLV—Bokeloh bat lyssavirus; KBLV—Kotalahti bat lyssavirus; WCBV—West Caucasian bat lyssavirus; LLEBV—Lleida bat lyssavirus; Seropositive—Seropositive Blood samples.
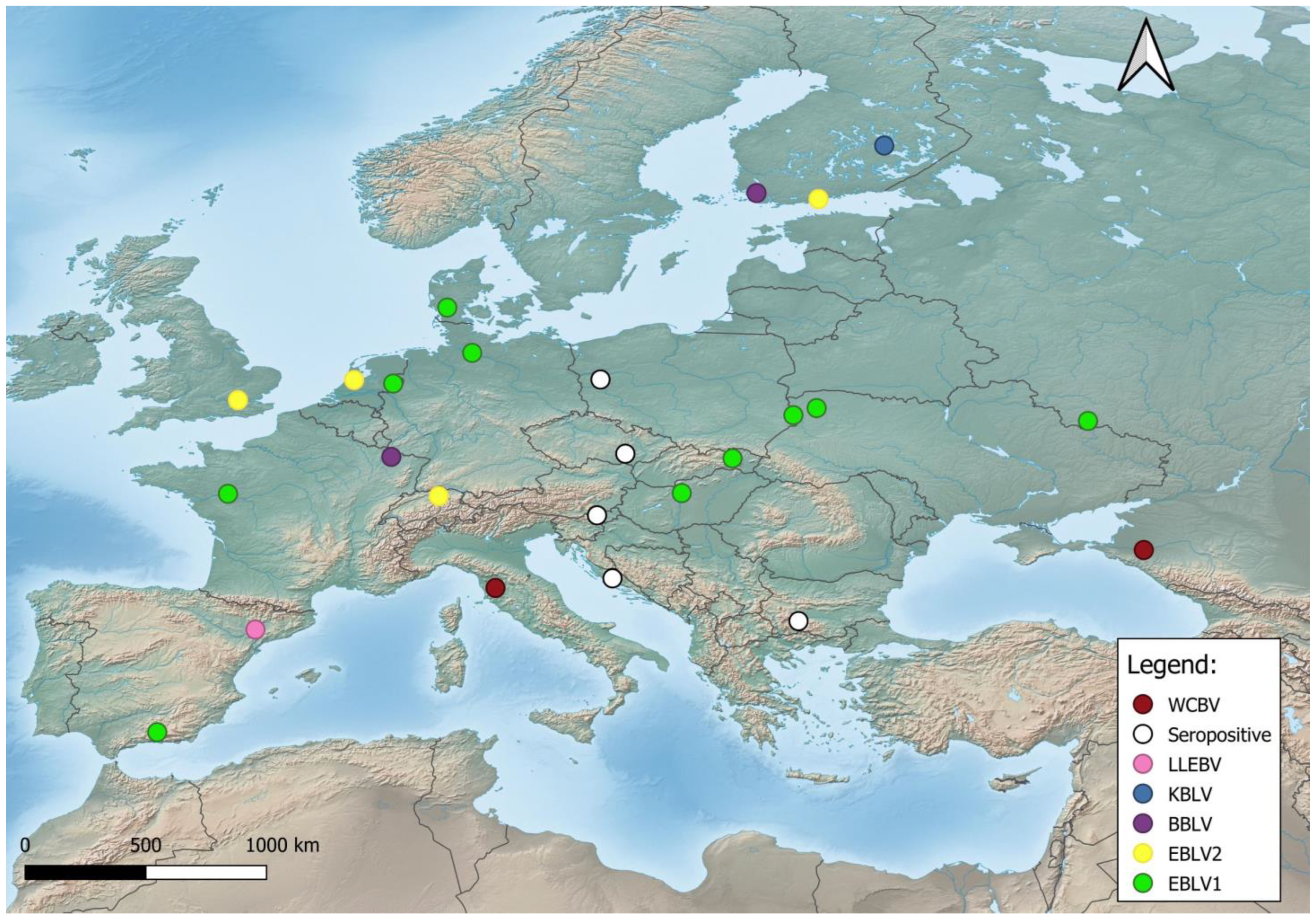
Figure 2.
Phylogeny of bat lyssaviruses. The sequences of the N gene of representative reference sequences of lyssaviruses used in the analysis were derived from Genbank. The evolutionary history was inferred by using the Maximum Likelihood method and General Time Reversible model [64]. There was a total of 568 positions in the final dataset. Evolutionary analyses were conducted in MEGA X [65]. Virus names are: classical rabies virus (RABV), Lyssavirus Aravan (ARAV), Lyssavirus australis (ABLV), Lyssavirus Bokeloh (BBLV), Lyssavirus Duvenhage (DUVV), Lyssavirus Hamburg (EBLV-1), Lyssavirus Helsinki (EBLV-2), Lyssavirus Gannoruwa (GBLV), Lyssavirus Ikoma (IKOV), Lyssavirus Irkut (IRKV), Lyssavirus Khujand (KHUV), Lys-savirus Lagos (LBV), Lyssavirus Lleida (LLEBV), Lyssavirus Mokola (MOKV), Lyssa-virus Shimoni (SHIBV), Kotalahti bat lyssavirus (KBVL), Lyssavirus Caucasicus (WCBV) and Lyssavirus Formosa, which includes Taiwan bat lyssavirus -1 (TWBLV-1) and Taiwan bat lyssavirus-2 (TWBLV-2).
Figure 2.
Phylogeny of bat lyssaviruses. The sequences of the N gene of representative reference sequences of lyssaviruses used in the analysis were derived from Genbank. The evolutionary history was inferred by using the Maximum Likelihood method and General Time Reversible model [64]. There was a total of 568 positions in the final dataset. Evolutionary analyses were conducted in MEGA X [65]. Virus names are: classical rabies virus (RABV), Lyssavirus Aravan (ARAV), Lyssavirus australis (ABLV), Lyssavirus Bokeloh (BBLV), Lyssavirus Duvenhage (DUVV), Lyssavirus Hamburg (EBLV-1), Lyssavirus Helsinki (EBLV-2), Lyssavirus Gannoruwa (GBLV), Lyssavirus Ikoma (IKOV), Lyssavirus Irkut (IRKV), Lyssavirus Khujand (KHUV), Lys-savirus Lagos (LBV), Lyssavirus Lleida (LLEBV), Lyssavirus Mokola (MOKV), Lyssa-virus Shimoni (SHIBV), Kotalahti bat lyssavirus (KBVL), Lyssavirus Caucasicus (WCBV) and Lyssavirus Formosa, which includes Taiwan bat lyssavirus -1 (TWBLV-1) and Taiwan bat lyssavirus-2 (TWBLV-2).

Figure 3.
Zoogeographical distributions of Rousettus aegyptiacus and Rhinolophus ferrumequinum according to IUCN and their contact zone between the Ethiopian and Palearctic realms.
Figure 3.
Zoogeographical distributions of Rousettus aegyptiacus and Rhinolophus ferrumequinum according to IUCN and their contact zone between the Ethiopian and Palearctic realms.
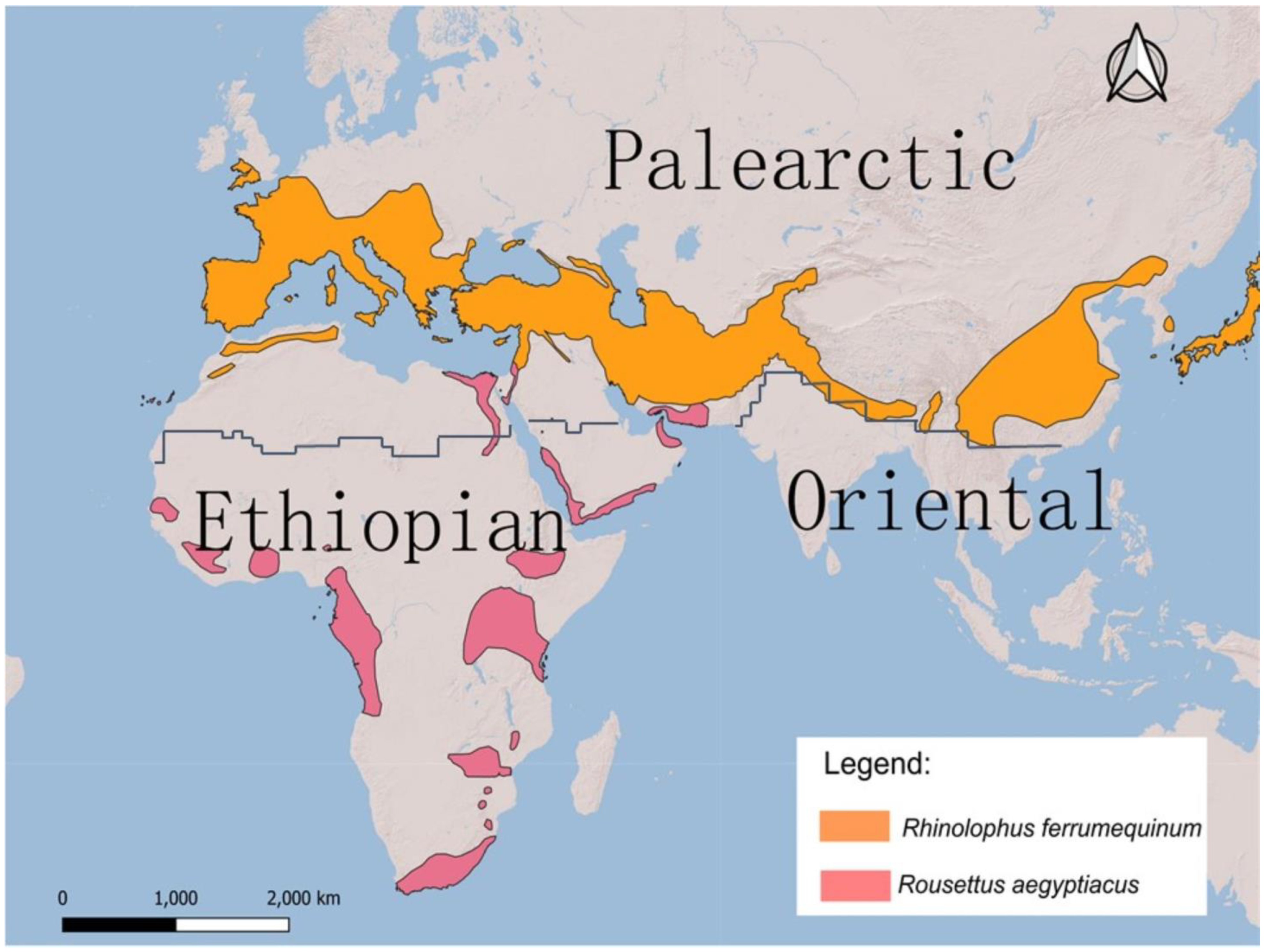

Table 1.
European bat species, their IUCN conservation status and lyssavirus presence.
| Species | IUCN status |
Bat lyssaviruses in Europe | ||||||
|---|---|---|---|---|---|---|---|---|
| Rhinolophidae | EBLV-1 | EBLV-2 | BBLV | KBLV | LLEBV | WCBV | ||
| Rhinolophus blasii | Blasius’s horseshoe bat | VU | ||||||
| Rhinolophus euryale | Mediterranean horseshoe bat | VU | ||||||
| Rhinolophus ferrumequinum | Greater horseshoe bat | NT | + | |||||
| Rhinolophus hipposideros | Lesser horseshoe bat | NT | ||||||
| Rhinolophus mehelyi | Mehely’s horseshoe bat | VU | ||||||
| Vespertilionidae | ||||||||
| Barbastella barbastellus | Western Barbastelle bat | VU | ||||||
| Eptesicus bottae | Botta’s Serotine | N/A | ||||||
| Eptesicus nilssonii | Northern bat | LC | ||||||
| Eptesicus isabellinus | Isabelline Serotine bat | N/A | + | |||||
| Eptesicus serotinus | Common Serotine | LC | + | |||||
| Hypsugo savii | Savi’s pipistrelle | LC | ||||||
| Myotis alcathoe | Alcathoe whiskered bat | DD | ||||||
| Myotis aurascens | Steppe whiskered bat | LC | ||||||
| Myotis bechsteinii | Myotis bechsteinii | VU | ||||||
| Myotis blythii | Lesser mouse-eared bat | NT | + | |||||
| Myotis brandtii | Brandt’s bat | LC | + | |||||
| Myotis capaccinii | Long-fingered bat | VU | ||||||
| Myotis dasycneme | Pond bat | NT | + | |||||
| Myotis daubentonii | Daubenton’s bat | LC | + | |||||
| Myotis escalerai | Escalerai bat | N/A | ||||||
| Myotis emarginatus | Geoffroy’s bat | LC | ||||||
| Myotis myotis | Greater mouse-eared bat | LC | + | |||||
| Myotis mystacinus | Whiskered bat | LC | ||||||
| Myotis nattereri | Natterer’s bat | LC | + | + | ||||
| Myotis punicus | Maghreb mouse-eared bat | NT | ||||||
| Nyctalus azoreum | Azorean bat | EN | ||||||
| Nyctalus lasiopterus | Greater noctule bat | DD | ||||||
| Nyctalus leisleri | Leisler’s bat | LC | ||||||
| Nyctalus noctula | Common noctule | LC | + | |||||
| Pipistrellus kuhlii | Kuhl’s pipistrelle | LC | ||||||
| Pipistrellus hanaki | Hanaki’s Dwarf Bat | N/A | ||||||
| Pipistrellus maderensis | Madeira pipistrelle | EN | ||||||
| Pipistrellus nathusii | Nathusius’s pipistrelle | LC | + | |||||
| Pipistrellus pipistrellus | Common pipistrelle | LC | + | |||||
| Pipistrellus pygmaeus | Pygmy pipistrelle | LC | ||||||
| Plecotus auritus | Brown long-eared bat | LC | + | |||||
| Plecotus austriacus | Grey long-eared bat | LC | ||||||
| Plecotus kolombatovici | Kolombatovic’s Long-eared bat | NT | ||||||
| Plecotus macrobullaris | Mountain long-eared bat | NT | ||||||
| Plecotus sardus | Sardinian long-eared bat | VU | ||||||
| Plecotus teneriffae | Tenerife long-eared bat | EN | ||||||
| Vespertilio murinus | Parti-coloured bat | LC | + | |||||
| Miniopteridae | ||||||||
| Miniopterus schreibersii | Schreibers’s long-fingered bat | NT | + | + | + | |||
| Molossidae | ||||||||
| Tadarida teniotis | European free-tailed bat | LC | + | |||||
| Pteropodidae | ||||||||
| Rousettus aegyptiacus | Egyptian fruit bat | N/A (EN?) | + | |||||
Abbreviations: IUCN red list categories: EN: Endangered – Very high risk of extinction in the wild; VU: Vulnerable – High risk of extinction in the wild; NT: Near Threatened – Likely to become threatened in the near future; LC: Least Concern – Does not qualify for a more at-risk category. Widespread and abundant taxa are included in this category; DD: Data Deficient – Inadequate information to make a direct, or indirect, assessment of its risk of extinction based on its distribution and/or population status; N/A: not assessed
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



