
Submitted:
19 May 2023
Posted:
23 May 2023
You are already at the latest version
Abstract
The phenolics concentrations of rapeseed are important because they are antinutritional com-pound or directly positive related to antioxidant actives. In this study, single nucleotide poly-morphisms (SNPs) identified from genotyping-by-sequencing (GBS), and genome-wide associa-tion study of phenolic contents were investigated in 95 rapeseed mutant lines derived from gamma rays and their original cultivar. A total of 3,196,318 SNPs were detected, identifying a set of 70,208 union SNPs used to perform association studies. Six phenolics and glucosinolate (pro-goitrin) were identified, sinapine was the major phenolic compound (91.2–99.2%) in all geno-types. The association study was conducted for six phenolic compounds, progoitrin, total sinap-ine content (TSC), total phenolic content (TPC) and seed yield. The association study identified 241 SNPs that were significantly associated with six phenolic compounds, progoitrin, TSC and TPC. Based on the detected SNP markers, BnaA06g31740D, a gene associated progoitrin, BnaC-nng39930D, a gene associated trans-sinapine 1, BnaA07g31720D, a gene associated trans-sinapine 2, BnaC03g31950D and BnaA06g09180D, a gene associated TSC, and BnaAnng27700D, a gene associ-ated TPC, were selected as candidate genes. The SNP makers associated major phenolic com-pounds identified and could be used as targets for breeding programs aimed at improving anti-oxidant contents and reducing antinutritional components.
Keywords:
rapeseed
; glucosinolate
; phenolics
; GBS
; association study
1. Introduction
Rapeseed (Brassica napus L.) is one of the most widely cultivated crops that produces beautiful flowers and is extensively used in food, animal feed, and bio-industries [1,2]. Rapeseed seed contains high levels of proteins, minerals, carotenoids, and vitamins, which possess antioxidant properties studied in food and feed. Despite its high nutritional importance, narrow adaptations of rapeseed seed are used as food and feed because of the presence of high amounts of antinutritional compounds, such as phenolics and glucosinolates [3,4]. Understanding rapeseed breeding to reduce the amount of phenolic and glucosinolate compounds is important for developing high-quality rapeseed for food and feed purposes [4,5]. Likewise, genetic enhancement of seed yield is one of the main goals in rapeseed breeding programme [1]. As one of the important crops for the food and animal feed industries, rapeseed needs to improve its nutritional value and seed yield, as the current product is insufficient to meet Korea’s demand. Therefore, novel nutritional traits and high seed yield are necessary for rapeseed genetic resources.
Sinapine, a major phenolic compound in rapeseed seeds, is known for its antioxidative effects and is the most important natural source of cancer-preventive agents [6,7]. Phenylpropanoids are the main metabolic pathway of phenolic compounds and are involved in the synthesis of sinapine, the major (up to 80%) phenolic in rapeseed. Glucosinolates are nitrogen and sulphur containing compounds found in Brassicaceae, including rapeseed. Glucosinolate biosynthesis is the interaction between the biosynthetic pathways of various mechanisms in plants [8]. Over 100 different glucosinolate compounds are found in plants and are chemically categorised as aliphatic, indolic, and aromatic [8,9]. Depending on their precursor amino acids, glucosinolates can be categorised into indole glucosinolates derived from tryptophan; aliphatic glucosinolates from alanine, leucine, valine, and methionines; and aromatic glucosinolates from phenylalanine and tyrosine. Progoitrin is an aliphatic glucosinolate, also known as a precursor of sulforaphane [8,9,10,11]. Several studies have recently explored the connections between rapeseed phenolics and glucosinolate biosynthesis and metabolic pathways [4,8]. Although glucosinolates and phenylpropanoids are synthesised through distinct biosynthetic pathways and have unique functions, a study of Arabidopsis mutants showed an interaction between glucosinolate biosynthesis and phenylpropanoid pathways [11]. In addition, glucosinolates are of significant interest for their health benefits due to their anticancer effects [12]. However, these compounds determine the bitter taste and biologically active isothiocyanates that negatively affect vegetables and feedstocks [1,3]. Since natural variation in sinapine and glucosinolate content is limited in rapeseed, mutations targeting sinapine content are a reasonable approach to improve its nutritional value [6,7,13]. Radiation-induced mutation is one of the most effective methods for converting functional compound compositions for commercialisation. Mutant genotypes derived from radiation breeding techniques often exhibit altered phenotypic traits, including changes in secondary metabolite production [14]. Gamma radiation is ionising radiation that induces diverse mutational changes, from simple base substitutions to single-and/or double-strand DNA breaks. Additionally, mutation breeding technology has improved crop phenolic compound content, surpassing natural variation [14,15,16]. The phenolics and glucosinolate biosynthetic pathways have been extensively studied in rapeseed plants, the molecular networks that regulate phenolic and glucosinolate compound metabolism in mutant genotypes derived from radiation breeding techniques are not well understood.
Next-generation sequencing (NGS) technologies reveal efficient sequencing of the plant genome and can be used to directly detect single-nucleotide polymorphisms (SNPs) in plants [17]. Genotyping-by-sequencing (GBS) evaluates the scaled-down label of the target genome. These methods have been developed to enable the direct study of breeding goals using high-throughput SNP genotyping analysis, quantitative trait locus (QTL) mapping, and association mapping [18,19]. Association mapping is the action of SNPs used for the indirect selection of target traits in crops [20,21,22]. There is a non-random association between SNP markers and nutritional characteristics in mutations in genetically diverse crops. It has been widely applied for nutritional characteristics, such as protein, phenolics, oil content, fatty acid composition, and glucosinolates in Brassica breeding [5,7,13,14,22].
We developed novel rapeseed mutant lines from a domestic rapeseed cultivar using gamma radiation irradiation, and these lines have various phenolic and glucosinolate contents. Therefore, this study aimed to identify SNP in rapeseed mutant lines from the original cultivar. Meanwhile, an association study using GBS analysis found new SNP locations for phenolic and glucosinolate content in novel mutant lines.
2. Materials and Methods
2.1. Plant materials and seed yield
The mutant lines were generated by treating the seeds of the commercial cultivar ‘Youngsan’ and ‘Tammi’ with various doses of gamma-ray (60Co) irradiation as follows: 500 Gy (46 mutant lines from Tammi), 600 Gy (37 mutant lines; Y6 lines), 800 Gy (7 mutant lines; Y8 lines), and 1000 Gy (5 mutant lines; Y10 lines) at 2009. The treated seeds were sown to obtain the M1 generation, and seeds from one silique developed from the main stem of each M1 plant were harvested. In the M2 generation, all individuals from the original cultivar were investigated for agronomic characteristics (seed yield, flowering time and dwarfism) mutations. Ninety-five rapeseed mutants, selected based on their phenolic compound contents, were obtained from the M7 and M9 generations. The self-fertilisation procedure was continued until the M9 generation. We analysed the uniformity of the phenolic contents by UPLC (Ultra Performance Liquid Chromatography) for three generations (M7 to M9) to select stable lines. Finally, 95 mutants that varied in phenolic compound content and exhibited stable inheritance of the mutated characteristics from M7 to M9 generations were selected. Radiation-generated mutant genotypes were grown by the Radiation Breeding Research Team at the Advanced Radiation Technology Institute of the Korea Atomic Energy Research Institute (35°.5′09″ N, 126°83′41″ E). The seeds were planted in plots (3 × 20 m) and row spacing of 20 and 40 cm, respectively. Fertilizer (N:P:K 4:2:2 w/w/w) was applied to 600 kg∙ha−1 at pre-sowing and the plants were not fertilized after sowing. Seed yield was surveyed on 95 mutants and original cultivar during three generations (M7 to M9). Young leaves were sampled from the two-original cultivar (Youngsan and Tammi) and 95 rapeseed mutants. Genomic DNA was isolated using the DNeasy Plant Mini Kit (Qiagen, Hilden, Germany).
2.2. Library Construction and Genotyping-by-Sequencing
The construction of GBS libraries involved the use of the restriction enzyme ApeKI (5′-GCWGC-3′) following a modified protocol from a previously published study [18]. The oligonucleotides for the top and bottom strands of each barcode adapter and common adapter were diluted in TE (50 μM each) and annealed using a thermocycler. 100 ng/μL of DNA samples were added to wells containing the adapter. The samples were then digested with ApeKI (New England Biolabs, Ipswich, MA, USA) overnight at 75 °C. The digested DNA samples, each with a different barcode adapter, were combined (5 μL each) and purified using a commercial kit (QIAquick PCR Purification Kit; Qiagen, Valencia, CA, USA) according to the manufacturer’s instructions. Amplification of the restriction fragments from each library was performed in 50 μL volumes using HerculaseII Fusion DNA Polymerase (Agilent, CA, USA) and primers (A) 5′-AAT GAT ACG GCG ACC ACC GAG ATC TAC ACT CTT TCC CTA CAC GAC GCT CTT CCG ATC T-3′ and (B) 5′-CAA GCA GAA GAC GGC ATA CGA GAT CGG TCT CGG CAT TCC TGC TGA ACC GCT CTT CCG ATC T-3′. The amplified sample pools were then used as the sequencing “library” and were sequenced on the Illumina HiSeq 2000 platform by SEEDERS Co. (Daejeon, Korea).
2.3. Sequence Preprocessing and Alignment to Reference Genome Sequence
The demultiplexing process involved the use of the barcode sequence, followed by adapter removal and sequence quality trimming. Adapter trimming was carried out using Cutadapt (Version 1.8.3) [23] and sequence quality trimming was performed using the DynamicTrim and LengthSort features of the SolexaQA program (V.1.13) [17]. The DynamicTrim tool trimmed low-quality bases at the ends of short reads based on the Phred score and produced high-quality cleaned reads. LengthSort removed excessive base cuts made by DynamicTrim and kept only reads with a Phred score of ≥ 20 and a length of ≥ 25 bp. The cleaned reads were then mapped to the reference genome sequence of Brassica napus V5.1 using BWA (0.6.1-r104) [24]. The mapping process was a preliminary step for detecting raw SNPs (In/Del) in the B. napus Ver. 5.1 (http://www.genoscope.cns.fr/brassicanapus accessed on April 2023) and sequenced samples. A SAM file was generated using default values, with the exception of the following options: seed length (-l) of 30, maximum differences in the seed (-k) of 1, number of threads (-t) of 16, mismatch penalty (-M) of 6, gap opening penalty (-O) of 15, and gap extension penalty (-E) of 8. The experiment was repeated to ensure accuracy.
2.4. Raw SNP Detection and Consensus Sequence Extraction
The cleaned reads were mapped to the standard genome sequence, and the resulting SAM files were used to detect raw single nucleotide polymorphisms (SNPs) using SAMtools (0.1.16) [25] and to extract consensus sequences. Before SNP detection, the SNPs were validated using an in-house script developed by SEEDERS [26]. Raw SNP detection was performed with default values, with the exception of the following options: minimum mapping quality for SNPs (-Q) of 30, minimum mapping quality for gaps (-q) of 15, minimum read depth (-d) of 3, minimum indel score for nearby SNP filtering (-G) of 30, SNPs within 15 bp of a gap to be filtered (-w) of 15, window size for filtering dense SNPs (-W) of 30, and maximum read depth (-D) of 489.
2.5. Generate SNP Matrix
An integrated SNP matrix was generated from the samples to analyze the SNPs between the objects of interest. The process involved the creation of a list of unions using raw SNP positions obtained by comparing each sample with a reference genome (Brassica napus V. 5.1). Non-SNP loci were filled in from the consensus sequence of each sample. The final SNP matrix was generated by filtering out miscalled SNP positions through comparisons among the samples. SNPs were then divided into homozygous (read depth ≥ 90%), heterozygous (40% to 60%), and other (undistinguishable by type) groups based on their positions [21]. The SNP positions were then classified into intergenic or genic regions of the reference genome, with the genic region further classified into CDS or intron regions. To identify the SNPs that were unique to each mutant line, the common SNP in the original cultivar ‘Youngsan’ and ‘Tammi’was first selected in the integrated SNP matrix, followed by the selection of polymorphic SNPs by comparing the common SNP of the original cultivar with the base sequence of each mutant. Finally, the SNP loci of each mutant line were integrated to secure the SNP locus of the union for subsequent gene ontology and functional analysis.
2.6. Ultra-high-performance liquid chromatography (UPLC) and Total phenolic content analysis
The phenolic compounds present in the fully mature seeds were analyzed using UPLC [7]. Rapeseed seed (10 g) were extracted using a mixture of methanol and water (80:20 v/v) and filtered through a polyvinylidene fluoride syringe filter. The UPLC system was equipped with an XR-ODS column and a C18 guard column, and the mobile phase consisted of solvent A (0.1% trifluoroacetic acid in distilled water) and solvent B (0.1% trifluoroacetic acid in acetonitrile). The phenolic compounds were separated using a gradient of solvent B, and detection was performed at 280 nm. The peaks in the HPLC chromatogram were identified based on spectra, standards, and literature data. Seed was collected from the fully mature periods for each mutant line. The samples were quantified using sinapine, and the results were based on three biological replicates for each sample.
The total phenolic content (TPC) was determined by the Folin–Ciocalteu colorimetric method [27]. A small quantity (0.2 mL) of each extract and 1.5 mL of Folin–Ciocalteu reagent (20% v/v) were mixed thoroughly. Four mL of Na2CO3 (7%) were added, then made up to 10 mL with water. The mixture was kept in the dark at room temperature for 90 min. The absorbance was then measured at 760 nm using a UV-spectrophotometer (UV-1800, Shimadzu, Kyoto, Japan). TPC was calculated using a calibration curve of sinapic acid (Sigma, USA).
2.7. Association and networking study
In the association analysis, the 70,208 union SNP dataset was used and the FarmCPU model was utilized in the genomic association and prediction integrated tool (GAPIT) R package [28]. The Q general linear model (GLM) was performed on the chosen Q-matrix derived from STRUCTURE to determine the experiment-wise P value for each marker’s significance. The next step is to create a Q-mixed linear model (Q-MLM) method by kinship matrix. Significance of SNPs was determined based on a p-value < 0.0001 with corresponding -log10 (p) vales of 4.0 for the significant threshold. The annotated genes that were selected as significant SNPs were evaluated for their probable interactions, co-expression, predictions, co-localization, and shared proteins to define gene functions using GeneMANIA (https://genemania.org/).
3. Results
3.1. Genotyping-by-sequencing of rapeseed genotypes
The study performed GBS analyzing on 95 mutant lines and the two-original cultivar of rapeseed using the Illumina HiSeq 2000 platform. A summary of the sequence data generated from the 95 rapeseed mutant lines and the two original cultivar is shown in Table 1 and Table S1. A total of 1,088 million reads were generated, with an average of 11 million reads per sample. After removing low-quality sequences, there were 967,244,884 clean reads, with an average of 9.9 million reads per plant sample. The total length range of the clean reads was between 140,097,724 bp and 3,013,306,105 bp, with an average read length of 1,121,971,559 bp. The total number of mapped reads was 740,184,076 for all lines, with an average of 7,630,764 reads per sample. The mapped reads to the reference genome sequence ranged from 27.79% to 91.10% with an average of 75.14%. The total length of the mapped region was 1,765,446,134 bp, with an average of 18,200,476 bp per sample. The mapped region covered approximately 2.14% of the reference genome sequence with an average depth of 15.82.
3.2. Identification of SNPs
The SNPs for each line were first selected from the union of SNPs in the matrix position between the samples and the reference genome sequence (Table S2). A total of 3,196,318 SNPs were identified, with the majority being homozygous (2,025,206), of which the largest number (51,816) of SNPs was recorded in the Tm8-10 mutant line, and the lowest number (10,717) was observed in the Y6-203 mutant line.
To identify gamma-irradiated mutant lines, polymorphic SNPs were collected by comparing the common SNPs in the original cultivar with the base sequences of the mutant lines were considered mutational changes (Table S3; Supplementary Material S1). The total polymorphic SNPs ranged from 417 (Y6-322) to 11,256 (Y10-32) with averaging 6,654. The homozygous SNP distribution in the rapeseed mutant lines ranged from 210 (Y6-322) to 10,357 (Y10-12), with a mean of 5,318 per mutant line. A union of 70,208 SNPs without overlap was constructed (Table S4), with 37,932 of them located in the genic region and 32,276 in the intergenic region, encompassing 16,550 genes. Among the genic region SNPs, 25,807 were located in coding sequences (CDS), 900 in promoters, 10,031 in introns, and 1,194 in untranslated regions (UTR).
The distribution of union SNP on the rapeseed chromosomes is shown in Figure 1. The union SNP distribution in rapeseed chromosomes ranged from 1850 (chromosomes A02) to 4638 (chromosome C01). Chromosomes C01, C02, C03 and C08 had more SNPs than other chromosomes. Meanwhile, chromosome A09 had the highest number (2,887) of SNPs on genic regions.
3.3. Phenolic compound content and seed yield
The phenolic compounds of the 95 mutant lines and the two original cultivars are shown in Table 2 and Figure 2. Seven compounds were identified: dihexose (peak 1, tR 4.6), progoitrin (2R-2-hydroxy 3-butenyl glucosinolates) (peak 2, tR 6.5), trans-sinapine 1 (peak 3, tR 16.1), trans-sinapine 2 (peak 4, tR 16.4), sinapine(4-O-8′)guaiacyl (peak 5, tR 17.7), sinapoyl malate (peak 6, tR 20.3), and methyl sinapate (peak 7, tR 25.4) (Table S5). The phenolic compound contents of the ‘Youngsan’ cultivar were 0.70 ± 0.29, 0.15 ± 0.06, 16.03 ± 0.04, 17.65 ± 2.00, 0.14 ± 0.08, 0.94 ± 0.11 and 0.05 ± 0.01 mg∙g−1 for dihexose, progoitrin, trans-sinapine1, trans-sinapine2, sinapine(4-O-8′) guaiacyl, sinapoyl malate, and methyl sinapate content, respectively. The ‘Tammi’ cultivar were 0.21 ± 0.04, 0.08 ± 0.01, 16.03 ± 0.29, 7.64 ± 1.63, 22.22 ± 1.48, 0.12 ± 0.03, 1.08 ± 0.27 and 0.09 ± 0.03 mg∙g−1 for dihexose, progoitrin, trans-sinapine1, trans-sinapine2, sinapine(4-O-8′) guaiacols, sinapoyl malate, and methyl sinapate content, respectively. Sinapine was the major (91.2–99.2%) phenolic compounds in all genotypes. Significant differences in sinapine content were observed between mutant lines. trans-Sinapine1 content of the mutant lines ranged from 1.86 ± 0.36 mg∙g−1 to 25.85 ± 1.32 mg∙g−1. The Tm10-3 mutant line had the highest trans-sinapine1 content, whereas the Tm10-5EF mutant line had the lowest. The highest trans-sinapine2 content was recorded at 25.67 ± 1.22 mg∙g−1 in the Tm7M-1 mutant line, and the lowest value of 1.11 ± 0.25 mg∙g−1 was found in the Tm10-3 line. A total of seven compound contents were ranged from 46.03 mg∙g−1 (Y6-81a) to 8.31 mg∙g−1 (Y6-29-30). Dihexose, progoitrin, sinapine(4-O-8) guaiacyl, sinapoyl malate, and methyl sinapate were present in relatively small amounts in all genotyes. The progotrin (glucosinolates) of the mutant lines ranged from 0.03 ± 1.22 mg∙g−1 for Tm10-9EF to 0.50 ± 0.05 mg∙g−1 for Tm6-7. Y-6-81a mutant line showed higher levels of dihexose (1.48 mg∙g−1), progoitrin (0.40 mg∙g−1), sinapine(4-O-8′) guaiacyl (0.21 mg∙g−1), cyclic spermidine-alkaloid (0.62 mg∙g−1) and methyl sinapate (0.29 mg∙g−1).
The seed yield of the 95 mutant lines and the two-original cultivars are shown in Figure 3. The seed yield of the ‘Youngsan’ and ‘Tammi’ cultivar were 2856.7 ± 27.5 kg∙ha−1 and 3420.0 ± 33.5 kg∙ha−1, respectively. Significant differences in seed yield was observed between mutant lines. The seed yield of the mutant lines ranged from 2333.3 ± 20.2 kg∙ha−1 (Y6-21 line) to 3602.7 ± 15.7 kg∙ha−1 (TM6-3 line) with average of 2943.6 ± 19.4 kg∙ha−1.
The total phenolic content (TPC) of the 95 mutant lines and the two original cultivars are shown in Figure 4. The total phenolic content of the Tammi and Youngsan cultivar were 57.29±0.78 mg∙g−1 and 66.04 mg∙g−1, respectively. The highest total phenolic content was found in the Y6-81a line (78.31 mg∙g−1), whereas the lowest was detected in the Y6-29-30 line (15.86 mg∙g−1). Fourteen mutant lines (Y8-18-23, Y10-3-21, Y6-81b, Y6-201, Y6-202, Y6-291, Y6172, Y6-29-30, Tm8-14, Tm10-1, Tm10-2, TM10-3, Tm10-4EF and Tm10-5EF) had lower TPC (<30 mg∙g−1). The Y6-11 and Y6-81a mutants showed significantly higher TPC than the original cultivar.
3.4. Association Analysis
All SNPs significantly associated with phenolic compounds and glucosinolate (progoitrin) are described in Table S6, Figures S1 and S2. Among the 70,208 union SNP dataset associations identified, 241 SNPs were significantly associated with phenolic compounds and glucosinolate. Of the 241 selected union SNPs, 119 were located in genic regions, and 122 were detected in intergenic regions. A total of nine SNP loci were significantly associated with dihexose compound, including 5 genes (BnaC02g20420D, BnaA03g40510D, BnaA03g18260D, BnaC03g32440D and BnaC03g20990D) were annotated from mutant lines. For progoitrin compound, 15 SNP loci were identified, including BnaC08g30570D, BnaC06g02290D, BnaC06g02290D, BnaA06g31740D, BnaC01g10570D, BnaA01g13400D, BnaC04g51220D, BnaA08g28090D, BnaC07g03420D, BnaC01g13560D, BnaC01g04810D, BnaC05g24360D, BnaAnng10510D, BnaA08g24650D and BnaC09g27750D. A total of 27 SNP loci were significantly associated with trans-sinapine1 compound, including BnaCnng39930D, BnaC02g35370D, BnaC06g07520D, BnaA09g13790D, BnaAnng26190D, BnaC05g11590D, BnaC08g35930D, BnaA09g19450D, BnaA06g07300D, BnaA06g08830D, BnaCnng18490D, BnaC06g38030D, BnaA08g09430D, BnaC06g08200D, BnaCnng23620D, BnaA10g22100D, BnaCnng58110D, BnaA01g20360D, BnaA07g37760D, BnaC04g10570D, BnaA09g24030D, BnaA06g09240D, BnaC04g10570D, BnaA08g25950D, BnaC05g46690D, BnaA07g31720D and BnaC03g31950D. Fifteen SNP loci were significantly associated with sinapine(4-O-8′)guaiacyl, including BnaA09g16810D, BnaA05g23210D, BnaC07g23750D, BnaCnng31150D, BnaC09g23720D, BnaC09g22600D, BnaC08g16340D, BnaC03g49370D, BnaC01g17020D, BnaA08g06400D, BnaC02g26880D, BnaC09g54330D, BnaC07g23750D. A total of 42 SNP loci were significantly associated with methyl sinapate compound, the most significant SNP with the lowest p value recorded in BnaA01g27460D. Three SNP loci were significantly associated with TPC content, including BnaAnng27700D and BnaC01g06630D. The candidate genes associated with the strongest SNP variant for two majors phenolics (trans-sinapine1 and trans-sinapine) were BnaCnng39930D for trans-sinapine1 and BnaA07g31720D for trans-sinapine2, corresponding to the RING/U-box superfamily protein and Pentatricopeptide repeat (PPR) superfamily protein respectively. Five SNP loci were significantly associated with total sinapine content (TSC) content, including BnaAnng09880D (no annotated function), BnaC06g38030D (transcription coactivators), BnaC03g31950D (S-adenosyl-L-methionine-dependent methyltransferases superfamily protein), BnaC05g08990D (RNA-binding; RRM/RBD/RNP motifs family protein) and BnaA06g09180D (protein kinase 2A). The C03_19651313 SNP marker on chromosome C03 was as annotated BnaC03g31950D, which is consistently associated with both TSC and sinapoyl malate. The candidate genes associated with the most variant for progoitrin were BnaC08g30570D (ARABIDILLO-2) (Table 3). No suggestive or significant SNPs were detected for the seed yield.
All identified genes were analysed in association networks using GeneMANIA (Figure 5). The associated gene network results were linked through these interacting networks: co-expression 96.05%, co-localisation 2.14%, predicted 1.67%, and shared protein domains 0.14%. Co-expression is predicted to affect the phenolic compound content in rapeseed. No direct network between the identified Defensin-like (DEFL) family protein (AT2G04045; BnaC02g35370D), Double Clp-N motif-containing P-loop nucleoside triphosphate hydrolases superfamily protein (AT4G29920; BnaAnng26190D), AT59 (BnaA06g09240D) and MEI1 (BnaC06g38030D) genes was detected by GeneMANIA. The program extended the network by 20 associating genes as CFM3A (No. 1), PDE312 (No. 2), PUB17 (No. 3), ATEXO70E1 (No. 4), FTSH12 (No. 5), CLASP (No. 6), AT3G59040 (No. 7), AT2G32170 (No. 8), emb1703 (No. 9), SKIP8 (No. 10), CITRX (No. 11), RH39 (No. 12), LECRK55 (No. 13), EMB2654 (No. 14), AT5G58330 (No. 15), GTE2 (No. 16), RIBA1 (No. 17), ACT8 (No. 18), MEE40 (No. 19) and PUB13 (No. 20) used to rank the genes. Interestingly, the TPR13 (Tetratricopeptide repeat-like superfamily protein) was consistently associated with among sinapine1, sinapine(4-O-8’)guaiacyl and progoitrin, while AT4G00740 (Probable methyltransferase PMT13) was significantly associated with total sinapine content and the sinapoyl malate.
4. Discussion
Rapeseed is a widely cultivated oil crop whose seeds are also a valuable food, feed and bio-industry source [1,2]. Phenolic and glucosinolates compounds are widely distributed in rapeseed and are known for their antioxidant properties, such as 2, 2′-diphenyl-1-picrylhydrazyl radical (DPPH), ferric reducing antioxidant power, peroxyl radical scavenging, ferric tripyridyl triazine complex reduction, and organic radical scavenging [1,2,3,4,5,6,7]. They have been associated with a number of health benefits, including reducing the risk of chronic diseases such as cancer, cardiovascular disease, and diabetes [6,7]. However, phenolic and glucosinolates compounds in rapeseed can contribute to a bitter taste and decreased palatability, making them less desirable for use in food and feed [5,6]. Despite its nutritional and medicinal value, the amount of phenolic and glucisinolates information available regarding rapeseed genetic resources and identifying genetic markers associated with these traits to facilitate breeding programs is limited.
Normally, the wide variability of phenolic compounds in rapeseed depends mainly on the genotype, while small variations have been attributed to cultivation conditions [29,30]. Typically, new genetic resources in rapeseed are developed through hybridisation. However, the narrow range of phenolic compounds in rapeseed genotypes has limited the accurate characterisation of new genetic resources [6,7,29,30]. Mutagenesis using gamma radiation is effective in inducing variability in phenolic compounds in various plants. Gamma-ray mutagens have already been used in rapeseed to improve many useful traits, such as seed yield, morphology, resistance to pathogens, and variability of fatty acid composition [14,15]. Our research group recently developed new cultivars for modifying phenolic compounds by gamma irradiation in Chrysanthemum, Rubus, and Dendrobium [16,31,32]. In this study, we analysed the phenolic compound content in the two original cultivar and 95 mutant lines of rapeseed obtained through gamma-ray irradiation. The original cultivar ‘Youngsan’ and ‘Tammi has major chemical characteristics such as mid-oleic acid (63.0 ~ 66.0%), low glucosinolate (0.43%), mid-oil content (43.9%), and non-erucic acid [33]. Several studies have investigated rapeseed oil content, fatty acid composition and glucosinolates; however, there are limited studies on phenolic compounds in rapeseed genetic resources [5,34,35]. The major phenolic compounds found in the original cultivar and mutant lines are sinapate esters with sinapoylcholine (sinapine) [33]. Sinapine is the most abundant phenolic compound in rapeseed, accounting for more than 65% of the total amount [29,33,34,35,36]. There have been no studies on the phenolic compound content of Korean breeding cultivar and mutant-induced genotypes. The sinapine concentrations in the original cultivar (29.86 and 33.68 mg∙g−1) were higher than those reported for 575 winter rapeseed genotypes (3.2 to 12.7 mg∙g−1) [37]. Meanwhile, the sinapine contents of the original cultivar in the present study were similar to those reported for two Korean domestic cultivars and one imported cultivar from India (29.74 to 52.24 mg∙g−1) [36]. Hence, reducing sinapine content has been one of the major goals in rapeseed breeding to improve the nutritional value of rapeseed meals. The phenolic compound content of the original cultivar ‘Youngsan’ and ‘Tammi’ seed is not highly valuable for the food or feed industry. The total phenolic content in the eight mutant lines was two times lower than that found in the original cultivar, whereas the Y6-81a mutant line had a significantly higher total phenolic content than the other genotypes. Previous study reported that the sinapine was the chief contributor to antioxidant activity in the Australian canola meal [7]. This ability to control phenolic compound content through radiation breeding may have applications in rapeseed breeding programs for the food, feed, and medicinal industries. Additionally, the seed coat colour of rapeseed was determined based on its phenolic compound content [6]. The original cultivar has a black seed coat. Notably, these mutants had increased or decreased phenolic compound content without any change in seed coat colour, except for the Y6-29-30 line (brown). Mutation-breeding technology can change a few characteristics of a pre-existing cultivar and improve nutrient quality [10]. High seed yield is an important trait for rapeseed breeding [37]. The original cultivar ‘Youngsan’ and ‘Tammi’ has major agronomic characteristics, such as early flowering and mid-seed yield (2,800 to 3500 kg/ha). The result of evaluating the seed yield of all mutant lines for three yields showed that two mutant lines (Y6-351 and Y8-18-13) were 10% higher (over 3,200 kg/ha) than the original cultivar. Unfortunately, the SNP detected in GBS analysis are not associated with seed yields. These mutant lines may be useful for the development of new cultivars.
NGS methods have been used in molecular research on rapeseed [13,21]. However, information about mutations in the phenolic compounds of rapeseed genotypes produced by mutation breeding is lacking. In this study, we explored novel SNPs in rapeseed mutant lines derived from gamma rays. The SNPs ranged from 417 to 11,256, averaging 6,649 SNPs per mutant line. In rapeseed mutant lines, many SNPs were detected on chromosomes C01, C02, and C03. Similarly, our previous study, many SNPs were detected on chromosomes C01, C03, A09 in fatty acid mutant lines generated by gamma rays [21]. The biological effect of gamma rays depends primarily on the genotype, and chromosome aberration is the most important target for mutation breeding [14]. Of the 70,208 union SNPs, 241 were significantly associated with seven phenolic compounds. Association studies based on SNP markers are currently widely used to analyse complex quality traits in crops. Previous studies have identified candidate genes associated with oil content, fatty acids, glucosinolate, sinapine, phytic acid, tannin, and crude fibre in rapeseed [13,15,21,37,38]. However, studies on the identification of significant SNPs associated with phenolics (sinapine, sinapoyl malate, and methyl sinapate) and glucosinolates (progoitrin) in various types of mutant lines by association analysis have been reported in our studies of networks involved in the genetic control of these traits. Phenolics and glucosinolates and are the typical bitter taste of vegetables and induce hypothyroidism. However, these compounds are of interest because of their potential as cancer preventive agents [39].
The glucosinolate content in rapeseed appears to be influenced by genetic and environmental factors, as well as nitrogen supply management. Nitrogen supply is required for developing rapeseed with low glucosinolates, possibly negatively affecting the cost [29,40]. The development of novel rapeseed cultivars with low glucosinolate content can solve this problem. A total of 21 genes were annotated from progoitrin, BnaC08g30570D (ARABIDILLO-2; ARD-2) gene was selected as a strong candidate gene. However, further research is needed to fully understand the role of BnaC08g30570D in glucosinolates biosynthesis. Tetratricopeptide repeat (TPR)-like superfamily protein were identified using networking analysis. TPR-like superfamily protein is no known direct connection glucosinolates. However, TPR domain is often found in conjunction with F-box domain or the WD40 domain, and these proteins are involved in DNA repair processes [41,42]. Therefore, it is possible that TPR could also play a role in regulating glucosinolate biosynthesis in plants.
Sinapine biosynthesis is a complex biochemical pathway in plants that leads to the production of the phenolic compound sinapine, also known as sinapoylcholine [4,5,7]. The gene BnaCnng39930D containing the trans-sinapine1- associated SNP was annotated as RING/U-box superfamily protein. The RING/U-box superfamily of proteins is a large and diverse group of E3 ubiquitin ligases that play important roles in regulating various cellular processes, including protein degradation, transcriptional regulation, and signal transduction [43]. While there is no direct evidence that RING/U-box proteins are related to phenolic biosynthesis in plants. BnaCnng39930D gene is involved in the biosynthesis of erucic acid, which is a fatty acid found in the seed oil of rapeseed [43]. Interestingly, previous our study revaled that oleic acid were the principal fatty acids represented in the rapeseed lines from ‘Tammi’ except theTm8-15 and Tm10-1St mutant lines. Oleic linoleic and erucic acid were the major fatty acids in Tm8-15 and Tm10-1St [21]. These mutants are significantly lower in sinapine content. However, further research is needed to fully understand the role of RING/U-box proteins in phenolic biosynthesis. The gene BnaA07g31720D containing trans-sinapine2-related SNP was annotated as pentatricopeptide repeat (PPR) superfamily protein. PPR proteins are a large family of RNA-binding proteins that are involved in various aspects of RNA metabolism, including RNA editing, splicing, and translation. PPR proteins may indirectly affect the biosynthesis of sinapine through their role in regulating the expression of genes involved in the Arabidopsis [44]. The PPR protein has been shown to regulate the expression of genes involved in the phenylpropanoid pathway, including cinnamate 4-hydroxylase (C4H), which is a key enzyme in the biosynthesis of sinapine. Similarly, the PPR protein OTP70 has been shown to regulate the expression of genes involved in the phenylpropanoid pathway in rice [45]. Thus, PPR superfamily protein from the BnaA07g31720D gene can be considered to affect sinapine compound content. Gene BnaA01g06890D (Heavy metal ATPase 2; HMA2), which contains methyl sinapate related SNP, was annotated that plays a role in heavy metal detoxification in plants by transporting heavy metal ions such as copper, zinc, cadmium, and lead out of the cytosol and into intracellular compartments or out of the cell [46]. It was found that HMA2 knockout mutants in Arabidopsis thaliana exhibited reduced levels of total phenolic compounds, including flavonoids and lignin, compared to wild-type plants [47]. However, the exact mechanism by which HMA2 influences phenolic biosynthesis in rapeseed is not yet investigation.
The associated studies showed that BnaC06g38030D (transcription coactivators) BnaC03g31950D (S-adenosyl-L-methionine-dependent methyltransferases superfamily protein), BnaC05g08990D (RNA-binding; RRM/RBD/RNP motifs family protein) and BnaA06g09180D (protein kinase 2A) genes showed significant associations with TSC. The biosynthesis of sinapine is mediated by a number of enzymes, including protein kinases. Protein kinases are enzymes that catalyze the transfer of a phosphate group from ATP to a target protein, typically another enzyme or a receptor protein [48]. In the context of sinapine biosynthesis, protein kinases are involved in the regulation of the pathway, controlling the activity of other enzymes involved in the process. A protein kinase involved in sinapine biosynthesis is a serine/threonine protein kinase from rapeseed [4,5,48]. Protein kinase genes are a family of genes that encode for enzymes known as protein kinases, which play a critical role in the regulation of cellular processes such as signal transduction, cell division, and metabolism. There is evidence to suggest that phenolic compounds can interact with protein kinases and modulate their activity [49,50]. Phenolic compounds have been shown to inhibit specific protein kinases, while others can activate or enhance the activity of certain kinases. In addition, some studies have suggested that certain phenolic compounds can alter the expression of protein kinase genes, potentially leading to changes in cellular signaling pathways [48,50]. The biosynthesis of sinapine involves several steps, one of which is catalyzed by S-adenosyl-L-methionine-dependent methyltransferases superfamily proteins (SAMs). SAMs are enzymes that transfer a methyl group from S-adenosyl-L-methionine (SAM) to a target molecule, usually a protein, DNA, RNA, or small molecule [49,50,51,52]. SAM-dependent methyltransferases play a crucial role in many biological processes, including gene expression, signal transduction, and metabolism [22,52]. In sinapine biosynthesis, SAM is used as a methyl donor by SAM-dependent methyltransferases to transfer a methyl group to a precursor molecule called sinapoylglucose, forming sinapoylmalate. This reaction is catalyzed by the enzyme malate sinapoyltransferase (SMT), which belongs to the SAM-dependent methyltransferases superfamily [5,22,52]. SMT is a key enzyme in the biosynthesis of sinapine and is expressed in several plant tissues, including leaves, stems, and seeds [51,53]. Its activity is regulated by various factors, including developmental stage, environmental conditions, and genetic background. SAM-dependent methyltransferases play an essential role in the biosynthesis of sinapine and other phenolic compounds in Brassica [50,51,52,53]. However, these candidate genes are characterised by multiple gene family members. A clear understanding of the relationships among them is necessary to interpret the functional mechanism of phenolic compound biosynthesis in rapeseed, and these closely associated SNPs may be used to develop a breeding strategy to improve glucosinolate and phenolic content.
5. Conclusions
In the present study, we obtained abundant information on phenolics and glucosinolates (progoitrin) in the seeds of novel rapeseed mutant genotypes. Significant differences in phenolic and progoitrin compositions were observed between the two-original cultivar (Youngsan and Tammi) and their γ-irradiated mutants. These compound quality improvements are the focus of rapeseed breeding programs. Our study helped identify mutant genotypes and 241 candidate genes significantly associated with these seed phenolic quality traits of rapeseed which may offer helpful insights to promote breeding in rapeseed. This is the first study describing the SNPs generated from GBS and association studies for the large variability of phenolics in rapeseed mutants derived from gamma irradiation. BnaCnng39930D, BnaA07g31720D, BnaC03g31950D, BnaA06g09180D and BnaA06g31740D were selected as potential candidate genes for sinapine and the progoitrin. The study of the molecular networks that regulate glucosinolates and phenolic compound metabolism in mutant genotypes derived from radiation breeding techniques can provide valuable insights into the regulation of plant secondary metabolism and help in the development of novel rapeseed cultivar with improved phenolic and glucosinolates compound profiles in mutant derived from gamma-ray. Our results may help researchers select suitable SNPs, associations, and networks among rapeseed mutants which may be useful for the identification of optimal genotypes for the food and medicinal industries.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Results of genotyping-by-sequencing analysis in rapeseed genotypes. Table S2: Summary of total SNP numbers and alignment to the reference genome sequence. Table S3: Summary of polymorphic SNPs collected by comparing the common SNPs in the original cultivar with the base sequences of the mutant lines. Table S4: List of union SNP matrix loci that were generated for 95 rapeseed mutant lines. Table S5: Peak assignments of the aqueous methanol extract from the seed of rapeseed Table S6: List of 241 significant associated SNPs with phenolic and glucosinolates by association mapping in rapeseed. Supplement material 1: List of polymorphic SNPs in 95 mutant lines. Figure S1. Manhattan plot for phenolic compound in the rapeseed mutant lines using four GLM models. Figure S2. Manhattan plot for phenolic compound in the rapeseed mutant lines using four MLM models.
Author Contributions
Conceptualization: C.-H.B., J.-W.A. and S.-Y.K.; formal analysis: J.R., J.I.L., D.-G.K., Y.-J.L., B.Y., J.H.K. and S.H.K.; investigation: J.I.L., W.J.K., J.-B.K. and S.-J.K.; writing—original draft preparation: J.R., J.I.L., B.Y. and J.K.; writing—review and editing: J.-W.A. and C.-H.B.; project administration: S.H.K.; funding acquisition: S.-J.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (RS-2022-00156231) and the research program of KAERI, Republic of Korea (Project No. 523320-23)
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hu, Q.; Hua, W.; Yin, Y.; Zhang, X.; Liu, L.; Shi, J.; Zhao, Y.; Qin, L.; Chen, C.; Wang, H. Rapeseed research and production in Chaina. Crop J. 2017, 127–135. [Google Scholar] [CrossRef]
- Delgado, M.; Felix, M.; Bengoechea, C. Development of bioplastic materials: From rapeseed oil industry by products to added-value biodegradable biocomposite materials. Ind. Crops and Prod. 2018, 125, 401–407. [Google Scholar] [CrossRef]
- Nega, T.; Woldes, Y. Review on nutritional limitations and opportunities of using rapeseed meal and other rapeseed by-products in animal feeding. J. Nutr. Health Food Eng. 2018, 8, 43–48. [Google Scholar] [CrossRef]
- Wolfram, K.; Schmidt, J.; Wray, V.; Milkowski, C.; Schliemann, W.; Strack, D. Profiling of phenylpropanoids in transgenic low-sinapine oilseed rapeseed (Brassica napus). Phytochemistry 2010, 71, 1076–1084. [Google Scholar] [CrossRef]
- Cartea, M.E; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules. 2011, 16, 251–280. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Meng, G.; Chen, S.; Chen, Y.; Jiang, J.; Wang, Y.P. Correlation analysis of phenolic contents and antioxidation in Yellow- and black-seeded Brassica napus. Molecules 2018, 23, 1815. [Google Scholar] [CrossRef] [PubMed]
- Obied, H.K.; Song, Y.; Foley, S.; Loughlin, M.; Reman, A.; Mailer, R.; Masud, T.; Agboola, S. Biophenols and antioxidant properties of Australian canola meal. J. Agric. Food Chem. 2013, 61, 9176–9184. [Google Scholar] [CrossRef] [PubMed]
- Tolra, R.P.; Alonso, R.; Poschenrieder, C.; Barcelo, D.; Barcelo, J. Determination of glucosinolates in rapeseed and Thlaspi caerulescens plants by liquid chromatography–atmospheric pressure chemical ionization mass spectrometry. J. Chromatogr. A., 2000, 889, 75–81 (18). [Google Scholar] [CrossRef]
- Bischoff, K.L. Glucosinolates. Nutraceuticals, 2016. 551-554 (19).
- Miklavcic Visnjevec, A.; Tamayo Tenorio, A.; Steenkjaer Hastrup, A.C.; Hansen, N.M.L.; Peeters, K.; Schwarzkopf, M. Glucosinolates and isothiocyantes in processed rapeseed determined by HPLC-DAD-qTOF. Plants, 2021, 10, 2548. [Google Scholar] [CrossRef]
- Kim, J.I.; Zhang, X.; Pascuzzi, P.E.; Liu, C.J.; Chapple, C. Glucosinolate and phenylpropanoid biosynthesis are linked by proteasome-dependent degradation of PAL. New Phytologist 2020, 225, 154–168. [Google Scholar] [CrossRef]
- Melim, C.; Lauro, M.R.; Pires, I.M.; Oliveira, P.J.; Cabral, C. The role of glucosinolates from cruciferous vegetables (Brassicaceae) in gastrointestinal cancers: From prevention to therapeutics. Pharmaceutics, 2022, 14, 190. [Google Scholar] [CrossRef] [PubMed]
- Bhinder, G.; Sharma, S.; Kaur, H.; Akhater, J.M.; Mittal Sandhu, S. Genomic regions associated with seed meal quality traits in Brassica napus germplasm. Front. Plant Sci. 2022, 13, 882766. [Google Scholar] [CrossRef] [PubMed]
- Spencer-Lopes, M.M.; Forster, B.P.; Jankuloski, L. Manual on mutation. In Food and Agriculture Organization of the United Nations, 3rd ed.; FAO/IAEA: Rome, Italy, 2018; pp. 187–189. [Google Scholar]
- Singh, S.; Verma, A.K. 2015. A review on efforts of induced mutagenesis for qualitative and quantitative improvement of oilseed brassicas. J. Pharm. Phytochem. 2015, 4, 298–302. [Google Scholar]
- Ryu, J.; Nam, B.; Kim, B.R.; Kim, S.H.; Jo, Y.D.; Ahn, J.W.; Kim, J.B.; Jin, C.H.; Han, A.R. Comparative analysis of phytochemical composition of gamma-irradiated mutant cultivars of Chrysanthemum morifolium. Molecules 2019, 24, 3003. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Banks, T.W.; Cloutier, S. SNP discovery through next generation sequencing and its applications. Int. J. Plant Genomics. 2012, 831460. [Google Scholar] [CrossRef] [PubMed]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef]
- Zhu, M.C.; Ran, H.; Zhao, H.Y.; Tang, Y.S.; Shi, X.T.; Jiang, H.Y.; Zhang, Z.Y.; Fu, F.Y.; Xu, X.F. Tang, Z.L. Liu, L.Z.; Lu, K.; Li, J.N. Qu, C.M. Identification of quantitative trait loci and candidate genes controlling seed pigments of rapeseed. J. Interg. Agri. 2021, 20, 2862–2879. [Google Scholar] [CrossRef]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative usage of fixed and random effect models for powerful and efficient genome-wide association studies. PLoS Genet. 2016, 12, e1005767. [Google Scholar] [CrossRef]
- Ryu, J.; Lyu, J.I.; Kim, D.G.; Koo, K.M.; Yang, B.; Jo, Y.D.; Kim, S.H.; Kwon, S.J.; Ha, B.K.; Kang, S.Y.; Kim, J.B.; Ahn, J.W. Single Nucleotide polymorphism (SNP) discovery and Association Study of Rapeseed (Brassica napus L.) Mutant Lines Using Genotyping-by-Sequencing (GBS). Agronomy, 2021, 11, 508. [Google Scholar] [CrossRef]
- Khalifa, A.M.; Abd-Elshafy, E.; Abu-Khudir, R.; Gaafar, R.M. Infuence of gamma radiation and phenylalanine on secondary metabolites in callus cultures of milk thistle (Silybum marianum L.). J. Genetic Eng. Biotech. 2022, 20, 166. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–13. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence align-ment/map format and SAM tools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Oh, S.K.; Lee, J.H.; Lee, B.M.; Jo, S.H. Genome-wide SNP calling using next generation sequencing data in tomato. Mol. Cells 2014, 37, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Diosady, L.L. Rapid method for total phenolic acid determination in rapeseed/canola meals. Food Resear. Inter. 1997, 30, 571–574. [Google Scholar] [CrossRef]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative usage of fixed and random effect models for powerful and efficient genome-wide association studies. PLoS Genet. 2016, 12, e1005767. [Google Scholar] [CrossRef] [PubMed]
- Raboanatahiry, N.; Li, H.; Yu, L.; Li, M. Rapeseed (Brassica napus): Processing, utilization, and genetic improvement. Agronomy 2021, 11, 1776. [Google Scholar] [CrossRef]
- Guirrou, I.; El Harrak, A.; El Antari, A.; Hssaini, L.; Hanine, H.; El Fechtali, M.; Nabloussi, A. Bioactive compounds assessment in six Moroccan rapeseed (Brassica napus L.) varieties grown in two contrasting environments. Agronomy, 2023, 13, 460. [Google Scholar] [CrossRef]
- Ryu, J.; Kim, W.J.; Im, J.; Kim, S.H; Lee, K.S; Jo, H.J; Kim, E.Y.; Kang, S.Y.; Lee, J.H.; Ha, H.K. Genotyping-by-sequencing based single nucleotide polymorphisms enabled kompetitive allele specific PCR marker development in mutant Rubus genotypes. Electronic J. Biotechnology, 2018, 35, 57–62. [Google Scholar] [CrossRef]
- Lim, G.H.; Kim, S.W.; Ryu, J.; Kang, S.Y.; Kim, J.B.; Kim, S.H. Upregulation of the MYB2 transcription factor is associated with increased accumulation of anthocyanin in the leaves of Dendrobium bigibbum. Int. J. Mol. Sci., 2020, 21, 5653. [Google Scholar] [CrossRef]
- Lee, T.S.; Lee, Y.H.; Kim, K.S.; Lee, H.K.; Jang, Y.S.; Choi, I.H.; Kim, K.S. Effect of sowing time on oil content and fatty acid composition characteristics in rapeseed cultivars. Kor. J. Plant Res. 2014, 27, 202–208. [Google Scholar] [CrossRef]
- Naczk, M.; Amarowicz, R.; Sullivan, A.; Shahidi, F. Current research development on polyphenolics of rapeseed/canola: a review. Food Chem. 1998, 62, 489–502. [Google Scholar] [CrossRef]
- Felde, T.Z.; Baumert, A.; Strack, D.; Becker, H.C.; Mollers, C. Genetic variation for sinapate ester content in winter rapeseed (Brassica napus L.) and development of NIRS calibration equations. Plant Breed 2007, 291–296. [Google Scholar] [CrossRef]
- Yang, S.C.; Arasu, M.V.; Chun, J.H.; Jang, Y.S.; Lee, Y.H.; Kim, I.H.; Lee, K.T.; Hong, S.T.; Kim, S.J. Identification and determination of phenolic compounds in rapeseed meals (Brassica napus L.). J. Agri. Chem. Envi. 2015, 4, 14–23. [Google Scholar] [CrossRef]
- Chen, G.; Geng, J.F.; Rahman, M.; Liu, X.P.; Tu, J.X.; Fu, T.D.; Li, G.Y.; McVetty, P.B.E.; Tahir, M. Identification of QTL for oil content, seed yield, and flowering time in oilseed rape (Brassica napus). Euphytica 2010, 175, 161–174. [Google Scholar] [CrossRef]
- Lu, K.; Wei, L.; Li, X.; Wang, Y.; Wu, M.; Liu, M.; Zhang, C.; Chen, Z.; Xiao, Z.; Jian, H.; et al. Whole-genome resequencing reveals Brassica napus origin and genetic loci involved in its improvement. Nat. Commun. 2019, 10, 1154. [Google Scholar] [CrossRef] [PubMed]
- Soundararajan, P.; Kim, J.S. Anti-carcinogenic glucosinolates in cruciferous vegetables and their antagonistic effects on prevention of cancers. Molecules, 2018, 23, 2983. [Google Scholar] [CrossRef]
- Jeschke, V.; Weber, K.; Moore, S.S.; Burow, M. Coordination of glucosinolate biosynthesis and turnover under different nutrient conditions. Front. Plant Sci. 10, 1560.
- Gangurde, S.S.; Nayak, S.N.; Joshi, P.; Purohit, S.; Sudini, H.K.; Chitikineni, A.; Hong, Y.; Guo, B.; Chen, X.; Pandey, M.K.; et al. Comparative transcriptome analysis identified candidate genes for late leaf spot resistance and cause of defoliation in groundnut. Int. J. Mol. Sci. 2021, 22, 4491. [Google Scholar] [CrossRef]
- Zhang, X.; Gou, M.; Liu, C.J. Arabidopsis Kelch repeat F-Box proteins regulate phenylpropanoid biosynthesis via controlling the turnover of phenylalanine ammonia-lyase. Plant Cell. 2013, 25, 4994–5010. [Google Scholar] [CrossRef]
- Qu, C.; Li, J.; Zhu, K.; Liang, D.; Liu, J.; Hu, Q. Metabolic engineering of fatty acids for breeding of new oilseed crops: Strategies, progress, and perspectives. Engineering. 2017, 3, 27–35. [Google Scholar]
- Zhu, Q.; Dugardeyn, J.; Zhang, C.; Takenaka, M.; Kristina Kuhn, K.; Craddock, C.; Smalle, J.; Karampelias, M.; Denecke, J.; Peters, J.; Gerats, T.; Brennicke, A.; Eastmond, P.; Meyer, E.H.; Van Der Straeten, D. SLO2, a mitochondrial pentatricopeptide repeat protein affecting several RNA editing sites, is required for energy metabolism. The Plant J. 2012. 71, 335–345. [CrossRef]
- Chateigner-Boutin, A.L.; des Francs-Small, C.C.; Delannoy, E.; Kahlau, S.; Tanz, S.K.; de Longevialle, A.F.; Sota Fujii, S.; Small, L. OTP70 is a pentatricopeptide repeat protein of the E subgroup involved in splicing of the plastid transcript rpoC1. The Plant J. 2012, 65, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Eren, E.; Arguello, J.M. Arabidopsis HMA2, a divalent heavy metal-transporting PIB-type ATPase, is involved in cytoplasmic Zn2+ homeostasis. Plant Physiol. 2004. 136, 3712–3723. [CrossRef]
- Yuan, Y.; Wu, H.; Wang, N.; Li, J.; Zhao, W.; Du, J.; Wang, D.; Ling, H.Q. FIT interacts with AtbHLH38 and AtbHLH39 in regulating iron uptake gene expression for iron homeostasis in Arabidopsis. Cell Res. 2008, 18, 385–397. [Google Scholar]
- Nguyen, V.P.T.; Stewart, J.D.; Ioannou, I.; Allais, F. Sinapic acid and sinapate esters in Brassica: Innate accumulation, biosynthesis, accessibility via chemical synthesis or recovery from biomass, and biological activities. Front. Chem. 2021, 9, 664602. [Google Scholar] [CrossRef]
- Strehmel, N.; Hoehenwarter, W.; Mönchgesang, S.; Majovsky, P.; Krüger, S.; Scheel, D.; Lee, J. Stress-related mitogen-activated protein kinases stimulate the accumulation of small molecules and proteins in Arabidopsis thaliana root exudates. Front. Plant Sci. 2017, 8, 1292. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Bao, L.L.; Zhao, F.Y.; .Tang, M.Q.; Chen, T.; Li, Y.; Wang, B.X.; Fu, B.; Fang, H.; Li, G.Y.; Cao, J.; Ding, L.N.; Zhu, K.M.; Liu, S.Y.; Tan, X.L. BnaMPK3 Is a key regulator of defense responses to the devastating plant pathogen Sclerotinia sclerotiorum in oilseed rape. Front. Plant Sci. 2019, 10, 91. [Google Scholar] [CrossRef]
- Tohge, T.; Watanabe, M.; Hoefgen, R.; Fernie, A.R. The evolution of phenylpropanoid metabolism in the green lineage. CRBMol. Biol. Rev. 2013, 2, 123–152. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant, 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Tohge, T.; Watanabe, M.; Hoefgen, R.; Fernie, A.R. The evolution of phenylpropanoid metabolism in the green lineage. CRBMol. Biol. Rev. 2013, 2, 123–152. [Google Scholar] [CrossRef]
Figure 1.
Chromosomal distribution of the 70,208 union SNPs from GBS in gamma-irradiated 95 mutant lines. Reference genome sequence: Brassica napus V.5.1 (http://www.genoscope.cns.fr/brassicanapus).
Figure 1.
Chromosomal distribution of the 70,208 union SNPs from GBS in gamma-irradiated 95 mutant lines. Reference genome sequence: Brassica napus V.5.1 (http://www.genoscope.cns.fr/brassicanapus).
Figure 2.
3D Chromatogram of UPLC analysis in different rapeseed genotypes. A: Youngsan, B: Tammi, C: Y6-11, D: Y6-81a, E: Tm7M-1, F: Tm10-3, G: Tm10-5EF, H: Y6-29-30.
Figure 2.
3D Chromatogram of UPLC analysis in different rapeseed genotypes. A: Youngsan, B: Tammi, C: Y6-11, D: Y6-81a, E: Tm7M-1, F: Tm10-3, G: Tm10-5EF, H: Y6-29-30.
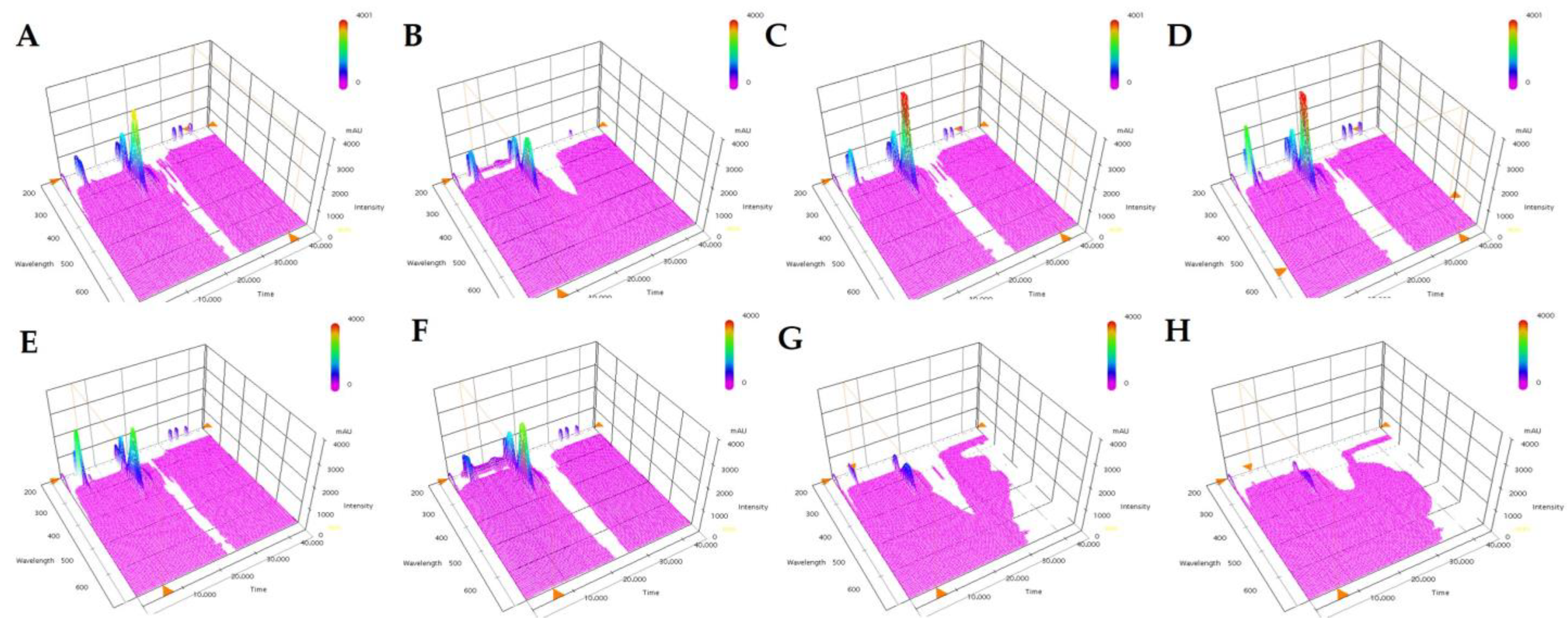
Figure 3.
The seed yield in the two original cultivar and 95 mutant lines.
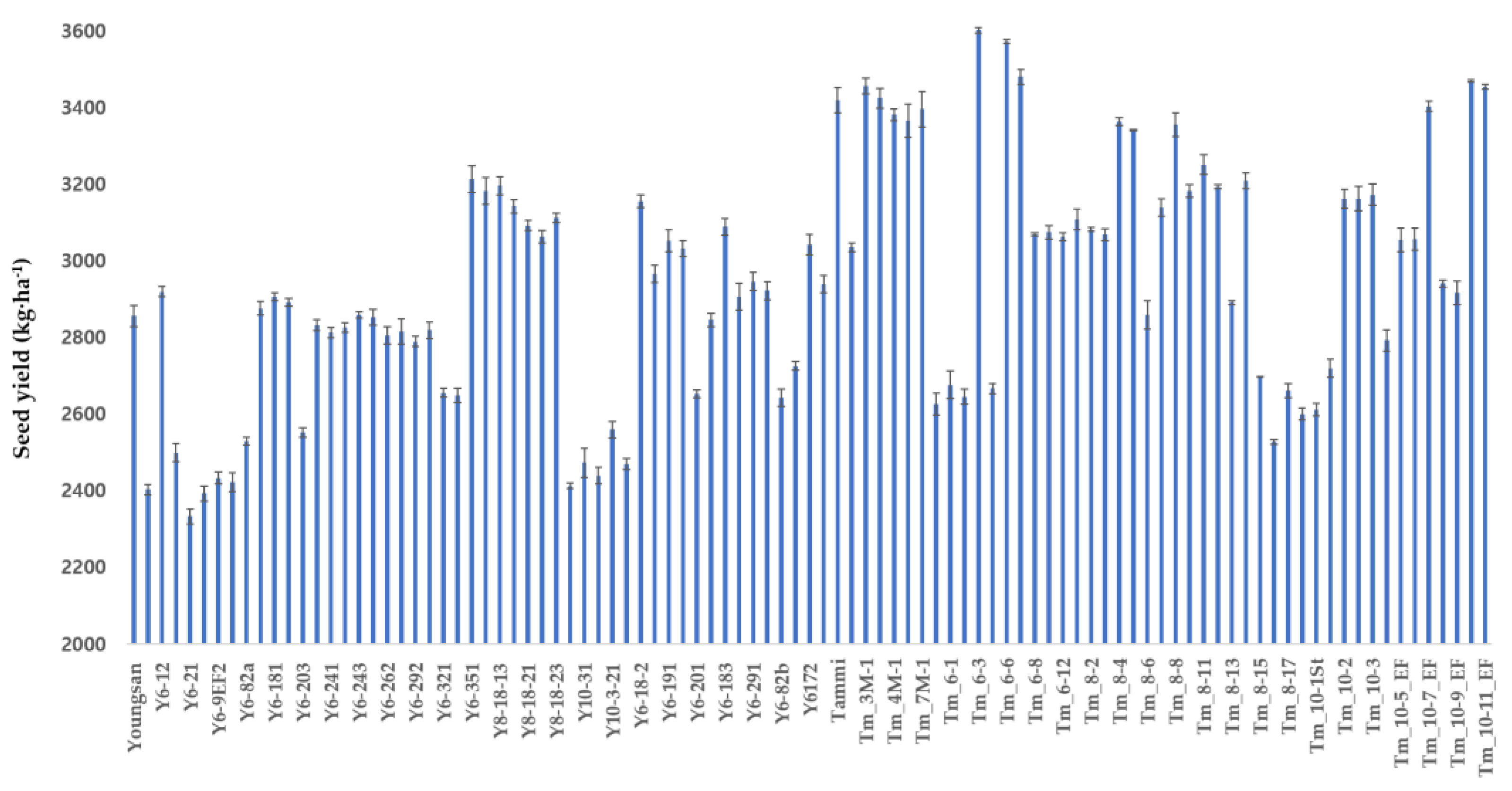
Figure 4.
Total phenolic content in 97 rapeseed genotypes (n = 3).
Figure 5.
Networking analysis of phenolics and glucosinolates associated SNP genes by GeneMANIA. S1: trans-sinapine 1, S2: trans-sinapine 2, SG: sinapine(4-O-8′)guaiacyl, SM: sinapoyl malate, TPC: total phenolic content, TSC: total sinapine content, MS: methyl sinapate, Pro: progoitrin. 1~20: ranked number.
Figure 5.
Networking analysis of phenolics and glucosinolates associated SNP genes by GeneMANIA. S1: trans-sinapine 1, S2: trans-sinapine 2, SG: sinapine(4-O-8′)guaiacyl, SM: sinapoyl malate, TPC: total phenolic content, TSC: total sinapine content, MS: methyl sinapate, Pro: progoitrin. 1~20: ranked number.
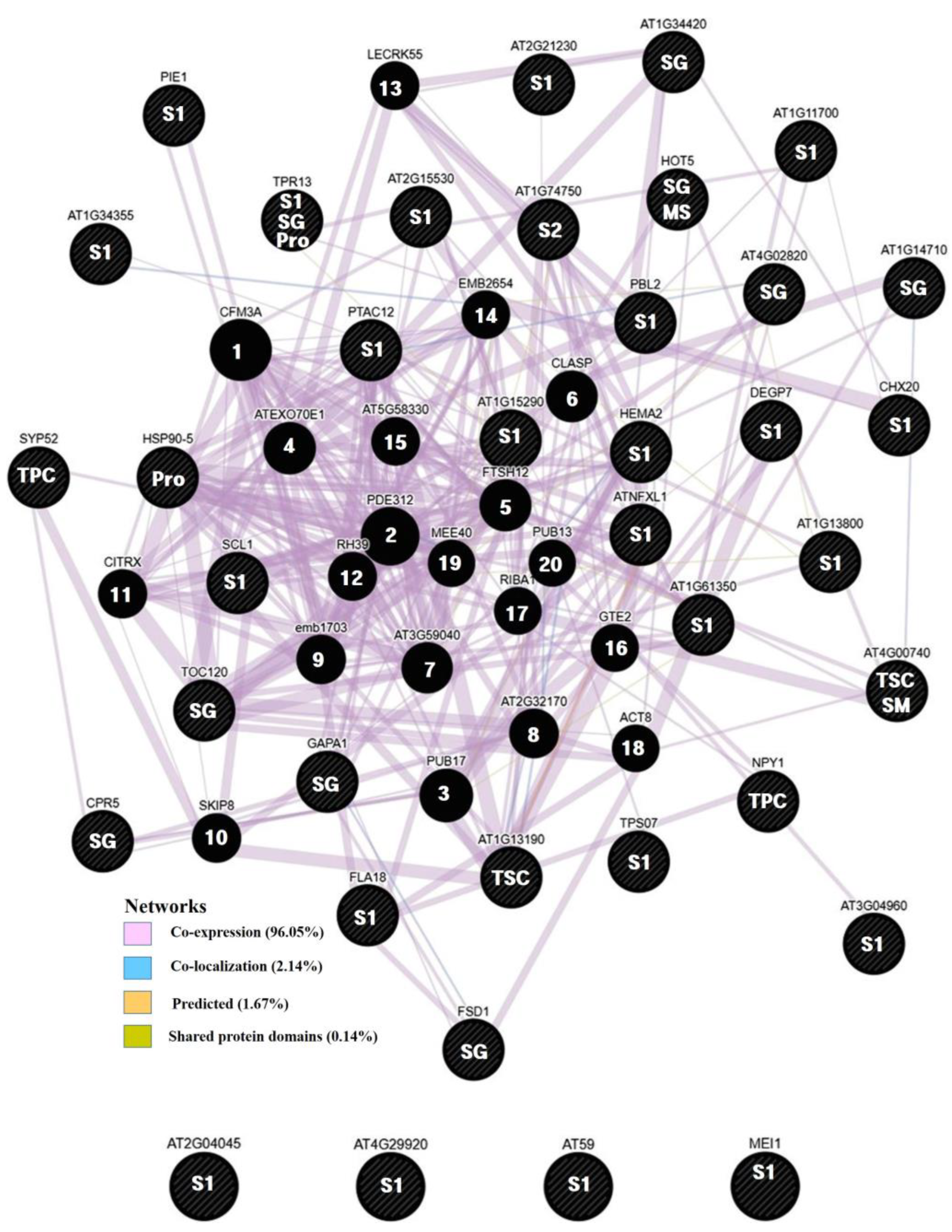

Table 1.
Summary of GBS data and alignment to the reference genome sequence.
| Total | Average/Plant | |
|---|---|---|
| Raw data | ||
| Reads | 1,088,590,054 | 11,222,578 |
| Bases (bp) | 164,377,098,154 | 1,694,609,259 |
| After trimming | ||
| Reads | 967,244,884 | 9,971,597 |
| Bases (bp) | 170,061,496,186 | 1,112,595,675 |
| Mapped reads on reference genome1 | ||
| Mapped reads | 740,184,076 | 7,630,764 |
| Bases (bp) | 1,765,446,134 | 18,200,476 |
| Reference genome coverage (%) | 2.14% | |
1 Reference genome sequence: Brassica napus V5.1 (http://www.genoscope.cns.fr/brassicanapus accessed on 10 April 2023).
Table 2.
Contents of phenolic compounds in seed extracts from 97 different rapeseed genotypes.
| No. | Line names | Dihexose | Progoitrin | trans-Sinapine1 | trans-Sinapine2 | Sinapine (4-O-8’) guaiacyl | Sinapoyl malate | Methyl sinapate | Total |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Youngsan | 0.70 ± 0.29 | 0.15 ± 0.06 | 16.03 ± 0.04 | 17.65 ± 2.00 | 0.14 ± 0.08 | 0.94 ± 0.11 | 0.05 ± 0.01 | 35.66 ± 2.69 |
| 2 | Y6-3 | 0.49 ± 0.21 | 0.24 ± 0.07 | 13.25 ± 2.00 | 11.77 ± 1.04 | 0.15 ± 0.02 | 0.22 ± 0.05 | 0.05 ± 0.01 | 26.18 ± 3.50 |
| 3 | Y6-12 | 0.56 ± 0.20 | 0.18 ± 0.02 | 14.00 ± 2.04 | 12.50 ± 1.13 | 0.16 ± 0.01 | 0.27 ± 0.06 | 0.06 ± 0.02 | 27.73 ± 3.58 |
| 4 | Y6-11 | 0.52 ± 0.23 | 0.13 ± 0.07 | 18.46 ± 0.30 | 22.89 ± 0.88 | 0.16 ± 0.06 | 0.30 ± 0.06 | 0.04 ± 0.01 | 42.50 ± 1.71 |
| 5 | Y6-21 | 0.38 ± 0.13 | 0.16 ± 0.06 | 12.41 ± 1.04 | 09.11 ± 0.55 | 0.21 ± 0.04 | 0.04 ± 0.01 | 0.18 ± 0.07 | 22.48 ± 1.99 |
| 6 | Y6-9EF1 | 0.60 ± 0.24 | 0.13 ± 0.07 | 14.74 ± 0.74 | 14.18 ± 1.47 | 0.20 ± 0.06 | 0.68 ± 0.13 | 0.04 ± 0.01 | 30.57 ± 2.82 |
| 7 | Y6-9EF2 | 0.67 ± 0.30 | 0.26 ± 0.05 | 14.87 ± 0.25 | 13.54 ± 1.40 | 0.18 ± 0.03 | 0.32 ± 0.06 | 0.04 ± 0.01 | 29.88 ± 2.21 |
| 8 | Y6-81a | 1.48 ± 0.66 | 0.40 ± 0.00 | 18.07 ± 1.13 | 24.77 ± 1.45 | 0.21 ± 0.03 | 0.81 ± 0.15 | 0.29 ± 0.08 | 46.03 ± 3.71 |
| 9 | Y6-82a | 1.23 ± 0.55 | 0.36 ± 0.01 | 15.70 ± 0.88 | 16.85 ± 1.65 | 0.23 ± 0.04 | 0.69 ± 0.12 | 0.25 ± 0.07 | 35.31 ± 3.43 |
| 10 | Y6-171 | 1.11 ± 0.48 | 0.34 ± 0.02 | 13.82 ± 0.24 | 14.19 ± 1.55 | 0.20 ± 0.02 | 0.63 ± 0.12 | 0.22 ± 0.07 | 30.51 ± 2.60 |
| 11 | Y6-181 | 0.40 ± 0.13 | 0.15 ± 0.05 | 13.72 ± 0.88 | 10.19 ± 0.82 | 0.08 ± 0.00 | 0.34 ± 0.10 | 0.12± 0.04 | 25.01 ± 2.01 |
| 12 | Y6-192 | 0.43 ± 0.16 | 0.15 ± 0.05 | 11.25 ± 1.12 | 09.12 ± 0.57 | 0.07 ± 0.00 | 0.34 ± 0.09 | 0.15 ± 0.05 | 21.51 ± 2.04 |
| 13 | Y6-203 | 0.29 ± 0.10 | 0.14 ± 0.06 | 12.06 ± 0.58 | 09.78 ± 0.72 | 0.17 ± 0.03 | 0.02 ± 0.01 | 0.12 ± 0.04 | 22.59 ± 1.54 |
| 14 | Y6-221 | 0.33 ± 0.12 | 0.15 ± 0.05 | 10.95 ± 0.55 | 08.72 ± 0.50 | 0.18 ± 0.05 | 0.02 ± 0.01 | 0.13 ± 0.04 | 20.49 ± 1.31 |
| 15 | Y6-241 | 0.32 ± 0.12 | 0.15 ± 0.05 | 10.81 ± 0.03 | 08.85 ± 0.53 | 0.19 ± 0.05 | 0.02 ± 0.00 | 0.12 ± 0.04 | 20.47 ± 0.83 |
| 16 | Y6-242 | 0.28 ± 0.10 | 0.14 ± 0.06 | 12.77 ± 0.23 | 10.14 ± 0.82 | 0.16 ± 0.03 | 0.02 ± 0.01 | 0.14 ± 0.04 | 23.64 ± 1.29 |
| 17 | Y6-243 | 0.29 ± 0.10 | 0.14 ± 0.05 | 11.63 ± 1.47 | 09.39 ± 0.64 | 0.14 ± 0.03 | 0.02 ± 0.00 | 0.11 ± 0.03 | 21.72 ± 2.34 |
| 18 | Y6-251 | 0.90 ± 0.40 | 0.27 ± 0.03 | 13.43 ± 1.24 | 10.93 ± 1.05 | 0.28 ± 0.08 | 0.01 ± 0.00 | 0.11 ± 0.03 | 25.93 ± 2.83 |
| 19 | Y6-262 | 0.40 ± 0.12 | 0.16 ± 0.04 | 11.94 ± 0.17 | 10.17 ± 0.86 | 0.19 ± 0.05 | 0.01 ± 0.00 | 0.14 ± 0.04 | 23.01 ± 1.28 |
| 20 | Y6-271 | 0.33 ± 0.11 | 0.10 ± 0.02 | 09.74 ± 1.40 | 08.46 ± 0.48 | 0.13 ± 0.03 | 0.02 ± 0.01 | 0.08 ± 0.03 | 18.87 ± 2.07 |
| 21 | Y6-292 | 0.25 ± 0.06 | 0.13 ± 0.05 | 10.97 ± 1.22 | 09.28 ± 0.77 | 0.07 ± 0.01 | 0.05 ± 0.01 | 0.14 ± 0.06 | 20.89 ± 2.20 |
| 22 | Y6-294 | 0.29 ± 0.09 | 0.13 ± 0.05 | 12.69 ± 0.08 | 10.62 ± 1.02 | 0.11 ± 0.04 | 0.01 ± 0.00 | 0.18 ± 0.07 | 24.03 ± 1.34 |
| 23 | Y6-321 | 0.30 ± 0.11 | 0.09 ± 0.02 | 12.08 ± 1.45 | 09.54 ± 0.88 | 0.13 ± 0.02 | 0.04 ± 0.01 | 0.10 ± 0.04 | 22.27 ± 2.54 |
| 24 | Y6-322 | 0.44 ± 0.17 | 0.10 ± 0.01 | 16.71 ± 1.53 | 13.38 ± 1.61 | 0.15 ± 0.03 | 0.04 ± 0.01 | 0.17 ± 0.06 | 30.99 ± 3.43 |
| 25 | Y6-351 | 0.46 ± 0.22 | 0.09 ± 0.04 | 12.26 ± 0.02 | 12.78 ± 1.35 | 0.11 ± 0.02 | 0.19 ± 0.04 | 0.04 ± 0.01 | 25.94 ± 1.70 |
| 26 | Y8-18-12 | 0.33 ± 0.13 | 0.11 ± 0.01 | 10.64 ± 1.65 | 09.07 ± 0.67 | 0.08 ± 0.02 | 0.03 ± 0.01 | 0.12 ± 0.04 | 20.37 ± 2.53 |
| 27 | Y8-18-13 | 0.31 ± 0.11 | 0.10 ± 0.02 | 09.82 ± 1.63 | 08.41 ± 0.48 | 0.06 ± 0.02 | 0.04 ± 0.01 | 0.08 ± 0.02 | 18.82 ± 2.28 |
| 28 | Y8-18-14 | 0.41 ± 0.17 | 0.13 ± 0.00 | 11.24 ± 0.22 | 09.69 ± 0.77 | 0.07 ± 0.02 | 0.04 ± 0.01 | 0.13 ± 0.06 | 21.72 ± 1.25 |
| 29 | Y8-18-21 | 0.31 ± 0.11 | 0.09 ± 0.02 | 10.01 ± 1.55 | 08.37 ± 0.50 | 0.07 ± 0.02 | 0.02 ± 0.01 | 0.10 ± 0.03 | 18.98 ± 2.24 |
| 30 | Y8-18-22 | 0.35 ± 0.13 | 0.11 ± 0.02 | 13.04 ± 1.33 | 10.94 ± 1.08 | 0.09 ± 0.02 | 0.03 ± 0.01 | 0.16 ± 0.07 | 24.71 ± 2.65 |
| 31 | Y8-18-23 | 0.25 ± 0.08 | 0.09 ± 0.02 | 07.66 ± 0.60 | 06.74 ± 0.18 | 0.05 ± 0.02 | 0.02 ± 0.01 | 0.11 ± 0.04 | 14.91 ± 0.95 |
| 32 | Y10-12 | 0.37 ± 0.13 | 0.16 ± 0.04 | 10.26 ± 0.82 | 08.99 ± 0.77 | 0.09 ± 0.03 | 0.06 ± 0.02 | 0.18 ± 0.07 | 20.11 ± 1.88 |
| 33 | Y10-31 | 0.23 ± 0.08 | 0.09 ± 0.02 | 09.57 ± 0.22 | 08.11 ± 0.51 | 0.07 ± 0.03 | 0.03 ± 0.01 | 0.14 ± 0.05 | 18.24 ± 0.93 |
| 34 | Y10-32 | 0.31 ± 0.11 | 0.09 ± 0.02 | 12.85 ± 0.69 | 10.45 ± 1.31 | 0.14 ± 0.04 | 0.05 ± 0.01 | 0.17 ± 0.08 | 24.06 ± 2.28 |
| 35 | Y10-3-21 | 0.33 ± 0.11 | 0.14 ± 0.04 | 07.20 ± 0.57 | 06.05 ± 0.07 | 0.06 ± 0.01 | 0.04 ± 0.01 | 0.07 ± 0.02 | 13.89 ± 0.84 |
| 36 | Y10-1b2 | 0.43 ± 0.18 | 0.12 ± 0.00 | 10.15 ± 0.12 | 08.20 ± 0.51 | 0.19 ± 0.05 | 0.02 ± 0.00 | 0.11 ± 0.04 | 19.22 ± 0.91 |
| 37 | Y6-18-2 | 0.27 ± 0.09 | 0.09 ± 0.03 | 09.79 ± 0.49 | 07.94 ± 0.49 | 0.04 ± 0.01 | 0.02 ± 0.00 | 0.11 ± 0.03 | 18.25 ± 1.14 |
| 38 | Y6-81b | 0.40 ± 0.14 | 0.15 ± 0.04 | 07.39 ± 0.72 | 06.18 ± 0.09 | 0.07 ± 0.01 | 0.03 ± 0.01 | 0.05 ± 0.01 | 14.27 ± 1.03 |
| 39 | Y6-191 | 0.37 ± 0.12 | 0.10 ± 0.02 | 13.00 ± 0.23 | 09.84 ± 0.95 | 0.07 ± 0.03 | 0.02 ± 0.01 | 0.14 ± 0.05 | 23.54 ± 1.41 |
| 40 | Y8-18-11 | 0.25 ± 0.10 | 0.09 ± 0.02 | 08.53 ± 0.57 | 07.26 ± 0.32 | 0.06 ± 0.01 | 0.02 ± 0.01 | 0.13 ± 0.05 | 16.35 ± 1.08 |
| 41 | Y6-201 | 0.51 ± 0.20 | 0.14 ± 0.01 | 07.18 ± 0.50 | 06.28 ± 0.13 | 0.15 ± 0.04 | 0.02 ± 0.01 | 0.08 ± 0.02 | 14.37 ± 0.91 |
| 42 | Y6-202 | 0.20 ± 0.07 | 0.08 ± 0.02 | 07.05 ± 0.07 | 05.78 ± 0.06 | 0.08 ± 0.02 | 0.02 ± 0.01 | 0.11 ± 0.03 | 13.32 ± 0.27 |
| 43 | Y6-183 | 0.46 ± 0.15 | 0.10 ± 0.02 | 09.16 ± 0.58 | 07.80 ± 0.50 | 0.04 ± 0.02 | 0.02 ± 0.01 | 0.14 ± 0.04 | 17.73 ± 1.32 |
| 44 | Y6-252 | 0.37 ± 0.12 | 0.10 ± 0.01 | 09.76 ± 0.53 | 08.12 ± 0.54 | 0.15 ± 0.04 | 0.01 ± 0.00 | 0.07 ± 0.02 | 18.59 ± 1.26 |
| 45 | Y6-291 | 0.29 ± 0.09 | 0.10 ± 0.02 | 06.85 ± 0.05 | 06.69 ± 0.26 | 0.04 ± 0.01 | 0.10 ± 0.02 | 0.11 ± 0.03 | 14.17 ± 0.49 |
| 46 | Y6-222 | 0.37 ± 0.15 | 0.10 ± 0.01 | 09.73 ± 0.47 | 08.30 ± 0.62 | 0.13 ± 0.03 | 0.02 ± 0.01 | 0.12 ± 0.05 | 18.77 ± 1.34 |
| 47 | Y6-82b | 1.41 ± 0.70 | 0.29 ± 0.08 | 13.32 ± 0.82 | 16.32 ± 2.64 | 0.09 ± 0.03 | 0.71 ± 0.16 | 0.25 ± 0.07 | 32.39 ± 4.50 |
| 48 | Y6-22 | 0.41 ± 0.17 | 0.11 ± 0.01 | 08.91 ± 0.35 | 07.69 ± 0.55 | 0.17 ± 0.03 | 0.02 ± 0.01 | 0.14 ± 0.06 | 17.44 ± 1.18 |
| 49 | Y6172 | 0.32 ± 0.11 | 0.10 ± 0.01 | 06.41 ± 0.54 | 05.50 ± 0.11 | 0.05 ± 0.01 | 0.02 ± 0.00 | 0.06 ± 0.02 | 12.47 ± 0.80 |
| 50 | Y6-29-30 | 0.39 ± 0.13 | 0.15 ± 0.03 | 04.31 ± 0.64 | 04.01 ± 0.16 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.04 ± 0.01 | 08.96 ± 0.99 |
| 51 | Tammi | 0.21 ± 0.04 | 0.08 ± 0.01 | 07.64 ± 1.63 | 22.22 ± 1.48 | 0.12 ± 0.03 | 1.08 ± 0.27 | 0.09 ± 0.03 | 31.45 ± 3.50 |
| 52 | Tm2M-1 | 0.30 ± 0.06 | 0.05 ± 0.01 | 03.67 ± 0.66 | 17.14 ± 1.32 | 0.05 ± 0.02 | 0.29 ± 0.07 | 0.04 ± 0.01 | 21.54 ± 2.15 |
| 53 | Tm3M-1 | 0.69 ± 0.13 | 0.45 ± 0.19 | 03.36 ± 0.55 | 17.96 ± 1.40 | 0.01 ± 0.00 | 0.35 ± 0.09 | 0.07 ± 0.03 | 22.89 ± 2.39 |
| 54 | Tm3M-2 | 0.07 ± 0.01 | 0.08 ± 0.01 | 18.85 ± 1.74 | 12.29 ± 2.72 | 0.05 ± 0.01 | 0.30 ± 0.08 | 0.02 ± 0.01 | 31.65 ± 4.58 |
| 55 | Tm4M-1 | 0.48 ± 0.04 | 0.35 ± 0.13 | 04.50 ± 0.77 | 19.64 ± 1.35 | 0.07 ± 0.03 | 0.86 ± 0.20 | 0.07 ± 0.02 | 25.97 ± 2.54 |
| 56 | Tm4M-2 | 0.34 ± 0.03 | 0.10 ± 0.01 | 04.15 ± 0.67 | 19.71 ± 1.55 | 0.07 ± 0.03 | 0.41 ± 0.10 | 0.06 ± 0.02 | 24.84 ± 2.41 |
| 57 | Tm7M-1 | 0.57 ± 0.12 | 0.41 ± 0.09 | 07.09 ± 0.97 | 25.67 ± 1.22 | 0.12 ± 0.04 | 1.01 ± 0.26 | 0.31 ± 0.09 | 35.18 ± 2.79 |
| 58 | Tm7M-2 | 0.52 ± 0.11 | 0.37 ± 0.07 | 04.39 ± 1.14 | 19.56 ± 1.30 | 0.09 ± 0.03 | 0.78 ± 0.20 | 0.24 ± 0.07 | 25.94 ± 2.91 |
| 59 | Tm6-1 | 0.03 ± 0.01 | 0.08 ± 0.01 | 11.41 ± 1.82 | 12.92 ± 2.77 | 0.07 ± 0.02 | 0.02 ± 0.01 | 0.02 ± 0.01 | 24.55 ± 4.65 |
| 60 | Tm6-2 | 0.06 ± 0.00 | 0.13 ± 0.02 | 06.34 ± 0.99 | 11.16 ± 2.59 | 0.04 ± 0.02 | 0.02 ± 0.01 | 0.02 ± 0.01 | 17.78 ± 3.63 |
| 61 | Tm6-3 | 0.05 ± 0.00 | 0.14 ± 0.01 | 03.80 ± 0.50 | 12.30 ± 2.88 | 0.05 ± 0.02 | 0.03 ± 0.01 | 0.02 ± 0.01 | 16.38 ± 3.43 |
| 62 | Tm6-4 | 0.54 ± 0.10 | 0.48 ± 0.00 | 04.82 ± 1.14 | 21.71 ± 0.90 | 0.10 ± 0.03 | 0.87 ± 0.22 | 0.27 ± 0.08 | 28.79 ± 2.47 |
| 63 | Tm6-6 | 0.21 ± 0.03 | 0.11 ± 0.00 | 04.79 ± 0.94 | 24.41 ± 1.38 | 0.07 ± 0.02 | 0.38 ± 0.09 | 0.01 ± 0.00 | 29.98 ± 2.47 |
| 64 | Tm6-7 | 0.43 ± 0.07 | 0.50 ± 0.05 | 04.54 ± 0.80 | 11.23 ± 2.65 | 0.05 ± 0.02 | 0.03 ± 0.01 | 0.02 ± 0.01 | 16.80 ± 3.61 |
| 65 | Tm6-8 | 0.39 ± 0.06 | 0.35 ± 0.04 | 03.10 ± 0.35 | 11.21 ± 2.65 | 0.04 ± 0.01 | 0.02 ± 0.01 | 0.02 ± 0.01 | 15.13 ± 3.12 |
| 66 | Tm6-10 | 0.03 ± 0.01 | 0.07 ± 0.00 | 02.77 ± 0.43 | 12.22 ± 2.71 | 0.04 ± 0.01 | 0.02 ± 0.01 | 0.02 ± 0.01 | 15.16 ± 3.18 |
| 67 | Tm6-12 | 0.55 ± 0.09 | 0.40 ± 0.03 | 04.86 ± 0.98 | 16.93 ± 1.01 | 0.07 ± 0.02 | 0.02 ± 0.01 | 0.02 ± 0.01 | 22.85 ± 2.15 |
| 68 | Tm6-13 | 0.13 ± 0.01 | 0.08 ± 0.00 | 04.06 ± 0.81 | 13.70 ± 2.66 | 0.06 ± 0.02 | 0.03 ± 0.01 | 0.03 ± 0.01 | 18.09 ± 3.52 |
| 69 | Tm8-2 | 0.10 ± 0.04 | 0.07 ± 0.01 | 02.85 ± 0.54 | 11.25 ± 2.53 | 0.04 ± 0.01 | 0.03 ± 0.01 | 0.02 ± 0.01 | 14.37 ± 3.14 |
| 70 | Tm8-3 | 0.04 ± 0.00 | 0.06 ± 0.00 | 09.94 ± 2.35 | 11.65 ± 2.66 | 0.05 ± 0.01 | 0.07 ± 0.02 | 0.03 ± 0.01 | 21.84 ± 5.06 |
| 71 | Tm8-4 | 0.13 ± 0.02 | 0.06 ± 0.01 | 13.21 ±3.34 | 13.43 ± 2.66 | 0.09 ± 0.02 | 0.05 ± 0.01 | 0.04 ± 0.01 | 27.01 ± 6.07 |
| 72 | Tm8-5 | 0.03 ± 0.01 | 0.06 ± 0.01 | 20.81 ± 1.32 | 11.85 ± 2.86 | 0.07 ± 0.02 | 0.06 ± 0.02 | 0.02 ± 0.01 | 32.91 ± 4.23 |
| 73 | Tm8-6 | 0.09 ± 0.02 | 0.07 ± 0.00 | 22.15 ± 1.57 | 18.04 ± 0.30 | 0.10 ± 0.02 | 0.06 ± 0.02 | 0.04 ± 0.01 | 40.55 ± 1.94 |
| 74 | Tm8-7 | 0.14 ± 0.03 | 0.08 ± 0.01 | 02.74 ± 0.69 | 17.10 ± 1.03 | 0.09 ± 0.02 | 0.25 ± 0.06 | 0.01 ± 0.00 | 20.40 ± 1.84 |
| 75 | Tm8-8 | 0.09 ± 0.00 | 0.07 ± 0.00 | 04.85 ± 1.03 | 11.00 ± 2.70 | 0.07 ± 0.02 | 0.04 ± 0.01 | 0.03 ± 0.01 | 16.17 ± 3.77 |
| 76 | Tm8-10 | 0.07 ± 0.03 | 0.06 ± 0.00 | 04.34 ± 0.86 | 10.71 ± 2.47 | 0.05 ± 0.01 | 0.06 ± 0.02 | 0.02 ± 0.01 | 15.31 ± 3.39 |
| 77 | Tm8-11 | 0.17 ± 0.04 | 0.31 ± 0.04 | 05.54 ± 1.20 | 12.38 ± 2.43 | 0.07 ± 0.02 | 0.05 ± 0.01 | 0.04 ± 0.01 | 18.55 ± 3.75 |
| 78 | Tm8-12 | 0.08 ± 0.02 | 0.06 ± 0.00 | 04.74 ± 1.17 | 10.31 ± 2.38 | 0.05 ± 0.02 | 0.03 ± 0.01 | 0.03 ± 0.01 | 15.30 ± 3.60 |
| 79 | Tm8-13 | 0.18 ± 0.01 | 0.27 ± 0.03 | 07.81 ± 0.38 | 15.66 ± 0.69 | 0.08 ± 0.02 | 0.04 ± 0.01 | 0.05 ± 0.02 | 24.09 ±1.16 |
| 80 | Tm8-14 | 0.08 ± 0.00 | 0.05 ± 0.01 | 04.38 ± 1.09 | 07.92 ± 2.14 | 0.04 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | 12.53 ± 3.27 |
| 81 | Tm8-15 | 0.17 ± 0.01 | 0.28 ± 0.02 | 08.00 ± 1.97 | 10.37 ± 2.43 | 0.08 ± 0.02 | 0.08 ± 0.02 | 0.05 ± 0.02 | 19.03 ±4.49 |
| 82 | Tm8-16 | 0.09 ± 0.00 | 0.06 ± 0.00 | 03.30 ± 0.79 | 09.37 ± 2.33 | 0.12 ± 0.02 | 0.03 ± 0.01 | 0.04 ± 0.01 | 13.02 ± 3.16 |
| 83 | Tm8-17 | 0.17 ± 0.01 | 0.31 ± 0.00 | 08.42 ± 1.18 | 11.47 ± 2.51 | 0.26 ± 0.04 | 0.07 ± 0.02 | 0.06 ± 0.02 | 20.74 ± 3.78 |
| 84 | Tm10-1 | 0.08 ± 0.01 | 0.06 ± 0.01 | 03.28 ± 0.73 | 07.32 ± 2.03 | 0.10 ± 0.02 | 0.06 ± 0.01 | 0.02 ± 0.01 | 10.92 ± 2.81 |
| 85 | Tm10-1St | 0.16 ± 0.02 | 0.30 ± 0.03 | 05.39 ± 0.42 | 09.93 ± 2.28 | 0.34 ± 0.06 | 0.04 ± 0.00 | 0.03 ± 0.01 | 16.20 ± 2.82 |
| 86 | Tm10-1Lin | 0.13 ± 0.01 | 0.16 ± 0.03 | 07.08 ± 0.83 | 09.06 ± 2.15 | 0.03 ± 0.01 | 0.02 ± 0.01 | 0.03 ± 0.01 | 16.52 ± 3.05 |
| 87 | Tm10-2 | 0.09 ± 0.02 | 0.07 ± 0.00 | 02.12 ± 0.33 | 07.52 ± 2.04 | 0.12 ± 0.02 | 0.04 ± 0.01 | 0.02 ± 0.01 | 09.98 ± 2.43 |
| 88 | Tm10Oel | 0.14 ± 0.01 | 0.22 ± 0.03 | 11.30 ± 1.75 | 12.54 ± 2.19 | 0.13 ± 0.02 | 0.02 ± 0.01 | 0.05 ± 0.02 | 24.40 ± 4.03 |
| 89 | Tm10-3 | 0.08 ± 0.00 | 0.05 ± 0.00 | 25.85 ± 1.32 | 01.11 ± 0.25 | 0.05 ± 0.01 | 0.03 ± 0.01 | 0.04 ± 0.02 | 27.21 ± 1.60 |
| 90 | Tm10-4EF | 0.51 ± 0.08 | 0.20 ± 0.04 | 03.11 ± 0.58 | 07.95 ± 2.03 | 0.30 ± 0.06 | 0.03 ± 0.01 | 0.02 ± 0.01 | 12.13 ± 2.81 |
| 91 | Tm10-5EF | 0.10 ± 0.01 | 0.08 ± 0.03 | 01.86 ± 0.36 | 06.93 ± 1.92 | 0.14 ± 0.02 | 0.03 ± 0.01 | 0.03 ± 0.01 | 09.18 ± 2.35 |
| 92 | Tm10-6EF | 0.08 ± 0.01 | 0.04 ± 0.00 | 08.02 ± 1.80 | 10.28 ± 2.09 | 0.10 ± 0.03 | 0.03 ± 0.01 | 0.05 ± 0.02 | 18.60 ± 3.95 |
| 93 | Tm10-7EF | 0.09 ± 0.01 | 0.06 ± 0.01 | 04.51 ± 0.26 | 10.40 ± 2.13 | 0.31 ± 0.06 | 0.02 ± 0.00 | 0.02 ± 0.01 | 15.39 ± 2.47 |
| 94 | Tm10-8EF | 0.08 ± 0.01 | 0.04 ± 0.00 | 08.61 ± 0.54 | 08.03 ± 2.03 | 0.06 ± 0.01 | 0.13 ± 0.03 | 0.04 ± 0.01 | 16.99 ± 2.64 |
| 95 | Tm10-9EF | 0.28 ± 0.10 | 0.03 ± 0.01 | 04.36 ± 0.30 | 10.81 ± 2.00 | 0.27 ± 0.06 | 0.03 ± 0.01 | 0.04 ± 0.01 | 15.83 ± 2.49 |
| 96 | Tm10-10EF | 0.06 ± 0.01 | 0.22 ± 0.02 | 04.88 ± 0.27 | 20.93 ± 0.87 | 0.53 ± 0.12 | 0.80 ± 0.16 | 0.27 ± 0.07 | 27.68 ± 1.53 |
| 97 | Tm10-11EF | 0.31 ± 0.11 | 0.04 ± 0.01 | 11.82 ± 0.83 | 09.59 ± 2.16 | 0.33 ± 0.08 | 0.05 ± 0.01 | 0.05 ± 0.02 | 22.20 ± 3.21 |
Values (mean ± SD) of extracts analyzed individually in triplicate.
Table 3.
Cnadidate genes associated with phenolic compounds by association study in rapeseed mutant line.
Table 3.
Cnadidate genes associated with phenolic compounds by association study in rapeseed mutant line.
| Triats | Chr_position | LOG10(P) | Transcript:feature | Description | TAIR ID | Allele |
|---|---|---|---|---|---|---|
| DIa | C02_16859648 | 4.70 | BnaC02g20420D:CDS | Ulp1 protease family protein | AT5G45570 | T/G |
| MSb | A01_ 19200278 | 6.74 | BnaA01g06890D:CDS | Heavy metal atpase 2 | AT4G30110 | C/T |
| Proc | C08_30292042 | 5.66 | BnaC08g30570D:CDS | ARABIDILLO-2 | AT3G60350 | A/T |
| Pro | A06_21272025 | 4.68 | BnaA06g31740D:CDS | Tetratricopeptide repeat (TPR)-like superfamily protein | AT3G27960 | C/T |
| SGd | A09_10109576 | 6.52 | BnaA09g16810D:Intron | GroES-like zinc-binding dehydrogenase family protein | AT5G43940 | T/C |
| S1e | CNN_38500012 | 6.42 | BnaCnng39930D:CDS | RING/U-box superfamily protein | AT2G15530 | G/A |
| S2f | A07_22086512 | 4.59 | BnaA07g31720D:CDS | Pentatricopeptide repeat (PPR) superfamily protein | AT1G79590 | G/T |
| TSCg | ANN_10591701 | 4.47 | BnaAnng09880D:CDS | No annotated function | - | A/T |
| TSC | SC06_35793928 | 4.33 | BnaC06g38030D:CDS | Transcription coactivators | AT1G77320 | G/T |
| TSC | C03_19651313 | 4.25 | BnaC03g31950D:Intron | S-adenosyl-L-methionine-dependent methyltransferases superfamily protein | AT4G00740 | T/G |
| SMh | 4.59 | |||||
| TSC | C05_4843154 | 4.21 | BnaC05g08990D:promoter | RNA-binding (RRM/RBD/RNP motifs) family protein | G/A | |
| TSC | A06_4969999 | 4.07 | BnaA06g09180D:Intron | Protein kinase 2A | A/T | |
| TPCi | ANN_31705069 | 5.39 | BnaAnng27700D:promoter | Syntaxin of plants 52 | AT1G79590 | G/T |
a DI: dihexose, b MS: methyl sinapate, c Pro: progoitrin, d SG: sinapine(4-O-8’)guaiacyl, e S1: trans-sinapine1, f S2: trans-sinapine2, g TSC: total sinapine content, h SM: sinapoyl malate, i TPC: total phenolic content.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



