
Submitted:
18 May 2023
Posted:
19 May 2023
You are already at the latest version
Abstract
: Cnidium officinale Makino, a perennial crop in the Umbeliperae family, is harvested in the summer for medicinal purposes. The average temperature in Korea has recently increased, and consequently, the area used to cultivate C. officinale has been reduced, and plant disease and soil sickness have increased. In this study, rhizosphere bacteria were isolated from C. officinale, and their antagonistic activity evaluated against Fusarium solani, which is a major issue during repeated cultivation. Twenty-eight strains were isolated from the root and rhizosphere soil, of which seven showed antagonistic activity against F. solani and were further investigated. An in planta test found that the dead rate of shoots was significantly lower in the group inoculated with PT1 isolated from the root tissue and that the fresh and dry weights were also higher, when compared with the other groups. PT1 was thus considered to have antagonistic activity against F. solani. The 16S rRNA analysis identified the PT1 as Leclercia adecarboxylata, and downstream studies confirmed the production of enzymes related to antagonism, such as siderophore and N-acetyl-β-glucosaminidase. The phosphorous solubilizing ability and secretion of related enzymes were also confirmed. The results indicate that PT1 could be utilized as a PGPR to help control fusarium wilt.
Keywords:
PGPR
; Cnidium officinale
; Leclercia adecarboxylata
; Bacillus
; Fusarium solani
; Fusarium wilt
1. Introduction
Cnidium officinale Makino is a perennial crop in the Umbeliperae family. It is harvested during the summer, and then dried and thinned for a variety of uses such as cosmetics, functional foods, and medicine. C. officinale is a representative medicinal plant in Korea, where 80% of its cultivation occurs in the inland Yeongyang, county which is in the north-eastern area of North Gyeongsang Province. Overall, C. officinale is the 8th most cultivated medicinal plant in Korea [1]. The active ingredients of C. officinale have been extensively researched. For example, in 2010, an extract from the above-ground part of C. officinale was evaluated for its antioxidant properties [1], and in 2011, the anti-inflammatory and anti-cancer effects of its rhizome extract were investigated in vitro conditions [2].
Currently, there are two major problems with the cultivation of C. officinale in Korea, the first of which relates to temperature. Over the past 100 years, the average annual temperature in Korea has risen by 1.5℃. Furthermore, the Intergovernmental Panel on Climate Change predicts that if humans continue to emit greenhouse gases in line with predictions, the average annual temperature will rise by 5.7℃ in 2100 [3]. However, the optimal temperature for growing C. officinale is 8.1℃–14.0℃. The production of rhizomes is usually also better at lower temperatures (< 14℃) than higher temperatures [4]. Thus, if temperature continue to rise, the areas suitable for C. officinale cultivation are predicted to dramatically decrease, and it is expected that only some alpine regions in Gangwon province will remain after 2090 [5].
The second issue is that C. officinale generally has low resistance to infection and can readily be infected by fungi. Major diseases caused by fungi include anthracnose, leaf blight, brown leaf spot, and fusarium wilt. Among them, Fusarium sp., which causes fusarium wilt, can continuously increase with repeated cultivation. However, cultivation techniques to prevent soil sickness that occur with the repeated cultivation of C. officinale have not yet been widely investigated. Therefore, to improve the productivity and sustainability of C. officinale, the development of cultivation technologies that can reduce soil sickness and countermeasures that can antagonize pathogens are required [6].
Microorganisms can be used to address these problems. Through interaction with various microorganisms in the rhizosphere, plants resist pathogens and environmental stress. These plant growth -promoting rhizobacteria (PGPR) compete for nutrients or secrete antagonists against pathogens and thus function as biocontrol agents (BCAs) [7]. In this study, various rhizobacteria were isolated from C. officinale, and their antagonistic activity against Fusarium solani, which is a major cause of soil sickness, was evaluated.
2. Materials and Methods
2.1. Isolation of bacteria from the rhizosphere and roots
C. officinale seedlings were collected from the fields of farmers in Cheongju, Korea, and transplanted to pots (60 × 45 × 23 cm; 6 seedlings per pot). After 30 d, three plants that were disease-free and showed good growth were selected, and their rhizosphere soil and roots were sampled. After washing the roots gently, they were then surface sterilized. First, they were washed with 70% ethanol for 10 s and then 0.5% NaClO for 5 min, and this process was repeated twice. Second, the roots with a sterilized surface were flushed and rinsed three times for 1 min with sterile water. After rinsing the roots, they were crushed with a mortar and pestle. Then, 2 g of crushed roots and 1 g of rhizosphere soil were aliquoted into 50 mL of sterile distilled water, respectively. The mixtures were incubated in a shaking incubator at 26℃ and, 150 rpm for 10 min. The mixtures were spread on tryptic soy agar (TSA) and potato dextrose agar (PDA) and further incubated at 26℃ for 2 days. Finally, strains with morphologically different characteristics were isolated in single cultures by streaking on the new same-type medium. The isolated strains were subjected to preliminary dual-culture with F. solani and finally selected based on antagonistic activity against F. solani.
2.2. Identification of rhizosphere and root bacteria
The DNA sequences of selected strains were analyzed by Macrogen (Seoul, Korea). To identify bacteria, a legion of 16s rRNA was used. DNA sequences were compared to standard databases in the nucleotide database of the National Center for Biotechnology Information (NCBI, Bethesda, MD, USA) using the BLASTn tool. For the phylogenetic tree, DNA sequences were compared to rRNA databases in NCBI using the BLASTn tool [8]. The phylogenetic tree of selected strains was illustrated using a neighbor-joining tree with MEGA 11 v.11, 1000 bootstrap replications, and a maximum composite likelihood substitution model [9].
2.3. In vitro antagonistic activity of the selected strains
To test the growth inhibition activity of the selected strains against pathogenic fungi, dual-cultures with F. solani (KACC No. 40384) were performed. The pathogenic fungal strain used in the experiment was obtained from the Korea Agricultural Collection (KACC). Dual-cultures were performed in 50 mm diameter Petri dishes with PDA. The pathogenic fungi, in the form of a 4 mm diameter agar plug, were raised on one side, and 3 μL of the overnight cultured strains were inoculated on 6 mm diameter paper disks at a distance of 30 mm from the pathogenic fungi. After incubation at 26℃ for 8 d, the inhibition rate was calculated by measuring the distance between the mycelia of the pathogenic fungi and the selected strains [10].
Where, Rc represents the average growth radius of the mycelia on the control plate and Rt represents the distance between the mycelia and bacteria on the treated plate.
2.4. In planta tests
C. officinale rhizomes were divided into 3–4 pieces (8 ~ 12 g) based on the buds, and then planted in seedbeds (53 × 37 × 9 cm) and germinated for 11 days. Each strain and the pathogenic fungus, F. solani, were used to inoculate the treated groups. To check the effects of the infection and the effects of the PGPR for the four strains, untreated seedlings were used for the first control group, and the second control group was inoculated with only F. solani. For each group, 16 of the C. officinales were used, and 2.5 mL of the selected strains were inoculated at an optical density (OD) of 0.1 at 600 nm, respectively. After incubating for 24 h, the spore suspension of F. solani was inoculated into plants at a concentration of 2 × 105 spores/mL, except for the second control group. After 1 week, the selected strains were reinoculated using the same method. Since the damage by the pathogen progressed rapidly and there was a risk that the dead shoots would rot and be lost, the test was terminated after 17 d and the total number of shoots, dead shoots, and dry weights of shoots and rhizomes were measured. The statistical analysis of the results was performed using one-way analysis of variance and the Statistical Package for the Social Sciences version 25.0, and the post hoc test was performed using the Duncan method.
2.5. Temperature tolerance assay
To test the growth ability of the strains in relation to temperature, a temperature tolerance assay was conducted [11]. The strains were adjusted to OD 0.01 at 600 nm in 10 mL of TSB medium. Then, these were incubated in a shaking incubator for 150 rpm at 0℃–40℃ at intervals of 5℃. For each temperature, the OD value was measured at 600 nm using a UV-Viz spectrophotometer (UBI-490 model, Korea) at 24 h intervals for 5 d, respectively.
2.6. In Vitro Test of PGP Traits
2.6.1. Protease production assay
Protease was produced using the PDA medium with 2% skim-milk [12]. Paper disks with 6 mm diameters were placed on the medium, and 3 μL of overnight cultured strains were inoculated. The plate was cultured at 27℃ for 4 d. Clear zones around the paper disk represented the production of protease against protein in the Skim-Milk PDA.
2.6.2. Cellulase production assay
Cellulase production was performed using PDA with 1% carboxymethylcellulose (CMC) and 0.01% trypan blue [13]. Paper disks, 6 mm in diameter, were placed on the medium, and 3 μL of overnight cultured strains were inoculated. The plate was cultured at 27℃ for 4 d, and cellulase production was confirmed by a clear zone around the paper disk due to the degradation of CMC combined with trypan blue.
2.6.3. Chitinase production assay
The production of chitinase was assessed using PDA with 2% colloidal chitin [14,15]. To make colloidal chitin, 20 g of chitin from shrimp shells was dissolved in HCL by stirring for 2 h. The chitin-HCL solution was filtered to remove undissolved particles using glass wool. After adding 500 mL of distilled water to the solution, the solution was incubated for 24 h at 5℃ and then the supernatant was removed. To wash the colloidal chitin, 3L of 5℃ tap-water was added and filtered using Whatman filter paper grade 2. The washed colloidal chitin was suspended in 500 mL of 5℃ distilled water, and the pH was measured. This washing process was repeated until the pH was > 7.0. The washed colloidal chitin was stored at 5℃ and used in further experiments. Paper disks with a 6 mm diameter were placed on the medium, and 3 μL of overnight cultured strains were inoculated. After incubating at 27℃ for 4 d. Clear zones around the paper disk represented the production of chitinase against chitin in the colloidal chitin PDA.
2.6.4. Hydrogen cyanide (HCN) production
HCN production was assessed using TSA with glycine (4.4 g/L) [16]. The strains were cultured for 24 h and then streaked on medium. The filter papers were immersed in picric acid solution (0.5% picric acid; 1% Na2Co3), and attached to the upper lids of the Petri dishes and sealed with parafilm. The plates were then incubated at 27℃ for 7 d and HCN production was confirmed based on the color of the filter paper (changes from yellow to red or brown).
2.6.5. Phosphate solubilization
Phosphate solubilization was assessed in the selected strains using a National Botanical Research Institute phosphate (NBRIP) protocol. The NBRIP medium contained l−1: glucose, 10 g; Ca3(PO4)2, 5 g; MgCl2ㆍ6H2O, 5 g; MgSO4, 0.14 g; KCl, 0.2 g; (NH4)2SO4, 0.1 g [17]. Paper disks that were 6 mm in diameter were placed on the medium, and 3 μL of the overnight cultured strain was inoculated. The plate was cultured at 27℃ for 4 d, and phosphate solubilizing activity was confirmed by the occurrence of a halo zone around the paper disk.
2.6.6. Siderophore production
Siderophore production by the selected strains was assayed using a Blue Agar CAS (Chrome azurol S) protocol [18]. Paper disks that were 6 mm in diameter were placed on the medium, and 3 μL of the overnight cultured strains were inoculated. The plates were cultured at 27℃ for 4 days, and siderophore production was confirmed by a halo zone around the paper disk.
2.6.7. Indole-3-acetic acid production
The capacity for indole-3-acetic acid (IAA) production by the selected strains was tested using Salkowski’s reaction [19]. Overnight-cultured strains were adjusted to an OD of 0.1 at 600 nm in 10 mL of tryptophan broth (gelatin pancreatic peptone, 10 g; sodium chloride, 5 g; L-tryptophan, 1g; pH 7.5). The inoculated broths were cultured in a shaking incubator at 27℃ and 150 rpm for 4 d. Then, 1.5 mL of the cultured broth was centrifuged for 10 min at 7300 х g at 4℃, and the supernatant was added to 3 mL of Salkowski reagent (35% HCLO4, 98ml; 0.5M FeCl3, 2ml) and incubated for 30 min at 25℃ in the dark to allow the reaction to develop. The concentration of IAA was measured using a UV-Vis spectrometer at 530 nm. The standard curve of the IAA to allow quantitative measurements was determined using the following IAA concentrations: 50, 25, 12.5, 6.25, 3.125, 1.563 and 0 mg/mL.
2.7. Exoenzyme activity
Exoenzyme activities were determined using the APIZYM® test system (bioMérieux, Marcy-l’Étoile, France). Overnight cultured strains were adjusted to the turbidity of 6 McFarland in sterile distilled water, and then 65 μL was pipetted to each cupule and incubated at 37℃ for 4 h. Then, ZYM A and ZYM B were added, and the data were recorded and interpreted for 10 min.
3. Results
3.1. Isolation of bacteria from the rhizosphere soil and roots
After isolation using the PDA and TSA, 7 bacterial strains were isolated from the disease-free and well-growing roots of C. officinales, while 21 bacterial strains were isolated from the rhizosphere soil. After dual-culture with F. solani, 4 of the bacterial strains showed high levels of inhibition activity and were selected for further investigation. The selected strains were identified using their 16S rRNA genes, and the BLASTn results for the sequences from the GenBank database are summarized in Table 1 and Figure 1.
3.2. In vitro antagonistic activity of the selected strains
The dual culture test to assess the antagonistic activity of the selected strains against F. solani, showed that all 4 strains were confirmed to have inhibitory activity, as judged by the occurrence of the inhibition zone. Among them, the PT1 showed the highest inhibition rate at 21.1 ± 2.4 %, and this was followed by ST7 at 16.3 ± 4.9%, ST8 at 11.5 ± 1.4%, and SP4 at 10.9 ± 3.1% (Figure 2).
3.3. In planta tests
Each of the strains and F. solani were inoculated into C. officinales and monitored for 18 d. After 17 d, the number of shoots, number of dead shoots, and the shoot fresh and dry weights were measured. An average of 17.8 new shoots was found to be sprouted from each plant, and the average shoot death rate in the first control group was 28.4%. In the treated and second control groups, there was an average of 47.6% shoot death rate, except with PT1 treated groups, which had an average death rate of 27% that similar to the first control group (Figure 3b). Furthermore, the dry weights of PT1 group were highest among all groups (Figure 3c,d). However, the standard deviation seemed to be largest in the PT1 group, as some plants were suppressed by the surrounding well-grown plants or had minor responses to the inoculation.
3.4. Growth traits in response to temperature
OD measurements were taken every 24 hours for 5days. Above 35℃, all strains showed slow growth rates and PT1 did not grow (Figure 4d). However, at 10–30, PT1 and ST7 showed the highest growth rates among all strains and, at temperatures <5℃, only PT1 and ST7 were shown to have growth (Figure 4a–c). All strains achieved their highest growth rate at 15℃ (Figure 4b).
3.5. In Vitro Test of PGP Traits
As a result of the in vitro test of PGP traits according to each assay, The ST8 and SP4 showed production of protease and cellulase related to antifungal ability by degrading the cell membranes and cell walls of fungi, respectively. The PT1 and ST7 showed production of siderophore related to antifungal ability and Fe uptake in plants, and capacity of phosphate solubilizing related to uptake of P in plants. In the test of IAA production, a plant growth promoting hormone, only PT1 secreted 13.19 ± 4.22 ppm of IAA. But there were no strains producing chitinase and HCN (Figure 5 and Table 2).
Each IAA value represents the mean of three replicates and the standard deviation (SD).
3.6. Exoenzyme activity
The exoenzyme characterization of strain enzymes using APIZYM confirmed that 7 types of enzymes (alkaline phosphatase, esterase, esterase lipase, acid phosphatase, naphthol-AS-BI-phosphohydrolase, α-glucosidase, β-glucosidase) were mainly produced in all strains. α-glucosidase was produced by SP4 and ST8. β-glucosidase was produced by ST7 and ST8. However, PT1 produced N-acetyl-β-glucosaminidase instead of glucosidase (Table 3).
4. Discussion
In this study, we isolated and evaluated bacteria from the roots and rhizosphere of C. officinale, and then further selected strains based on their antagonistic ability against F. solani, which causes fusarium wilt [20,21]. Four strains were identified as antagonistic bacteria. Among these, only the groups with PT1 treatment showed a significant effect with the in planta test. In the other groups, the death rate of the shoots increased after being treated with pathogenic fungi when compared with the first control group. However, in the group treated with PT1, the death rate of the shoots was not significantly different when compared with the first control group, while the fresh and dry weights increased. In addition, the group treated with PT1 had the highest inhibition rate in the in vitro antagonistic activity test. The PT1 strain was thus considered effective at controlling fusarium wilt in C. officinale.
Temperature is an important factor for the growth of plants and bacteria. C. officinale grows well in cool climates in the semi-highlands, which are approximately 400 m above sea level and have a wide diurnal range [4]. Yeongyang county is the largest production region for C. officinale, and its annual average temperature is 9℃–10℃, the average temperature from April–October during the cultivation period is 15–17℃, and the maximum does not exceed 28℃ [22]. The results of this study have shown that the PT1 strain can grow in temperatures ranging from 5℃ to 30℃, which may be broader than the range required for C. officinale. Therefore, the application of PT1 to the actual cultivation region is considered feasible.
In the APIZYM test, the PT1 was confirmed to produce acid phosphatase and naphthol-AS-BI-phosphohydrolase related to phosphate solubilization [23,24], in addition, N-acetyl-β-glucosaminidase, and several hydrolases were produced, and high siderophore production ability was confirmed using the CAS medium test. It is thus considered that PT1 has the ability to solubilize and convert insoluble phosphorous and iron in the soil into forms usable by plants, respectively [25,26,27]. In particular, in the phosphorus solubilizing assay, PT1 showed outstanding phosphorus solubilizing activity when compared with the other selected strains. In addition to solubilizing iron in the soil, siderophores are also related to the bacterial ability to antagonize pathogenic fungi [28,29,30], and N-acetyl-β-glucosaminidase can damage the cell wall of fungi and inhibit their growth [31,32].
The PT1 strain was specifically identified as Leclercia adecarboxylata. It is a gram-negative anaerobic Bacillus [33,34] and was first reported by Leclerc in 1962 [35]. This species has previously been isolated from herbaceous crops and investigated due to its ability to secrete IAA and biomolecules to increase resistance to abiotic stress or promote plant growth [36,37]. Moreover, the existence of trait and gene related PGPR and root colonization capacity has been studied [36,38]. Fusarium wilt occurs after the penetration of mycelium or germinating spores into the plant root tissues, from which they enter the xylem, and produce microconidia [39,40]. Therefore, to control fusarium wilt, it is important for bacteria to penetrate the plant root tissues and antagonize pathogenic fungi through plant-microbe interactions. The PT1 strain (L. adecarboxylata) was isolated from the root tissues, and was previously found to be capable of root colonization. Thus, it is expected that the PT1 strain can penetrate into the root tissue and inhibit the growth of pathogenic fungi.
The other three strains identified were Bacillus subtilis, Bacillus inaquosorum, and Bacillus vallismortis [41,42,43]. Although these strains did not show significant effects in the planta test, they were confirmed to produce glucosidase and protease unlike the PT1. Glucosidase catalyzes the hydrolysis of starch into a simple form [44], and recent studies have reported that it can protect plants against phatogen [45]. Protease can also antagonize pathogens and is an important enzyme in the nitrogen cycle [46,47].
The Bacillus sp. is the major bacteria in the rhizosphere, and they can survive in the soil for a long period under harsh environmental conditions via spore formation [48]. In particular, the possibility of Bacillus subtilis had been studied broadly for its potential as a PGPR [49], and it has been reported that root length and dry weight significantly increased when inoculated into the rhizosphere of maize [50]. Various microorganisms are involved in plant growth, and isolating and utilizing various microbial resources is also beneficial to the stability of the soil ecosystem [51]. Previous studies have reported that inoculations with two or more strain mixtures had better growth -promoting effects than inoculations with only one strain [52]. Therefore, this study isolated various PGPRs involved in promoting the growth of C. officinale and suggest the possibility of using them as biocontrol agents in the future.
5. Conclusions
The results of the in planta test showed that the group inoculated with both PT1 and pathogenic fungi had a higher rate of growth than the untreated group. The PT1 strain was thus considered to be a potential PGPR as well as have antagonism. However, as a result of PT1 inoculation, the effect was very different for each plant. Consequently, optimized inoculating techniques such as methods, concentrations, inoculation frequency, and timings should be investigated further. As plantation plants are grown outdoors, it is difficult to control their surrounding environment. A field test should thus also be conducted to investigate the effectiveness and safety of the method in a real-world environment [53].
Author Contributions
Conceptualization, S.H.L. and J.W.K.; Methodology, S.H.L., S.H.J. and J.W.K.; Validation, S.H.L., D.S.K. and J.W.K.; Formal analysis, S.H.L.; Data curation, S.H.L.; Funding acquisition, J.W.K.; Investigation, S.H.L., S.H.J. and J.Y.P.; Project administration, J.W.K.; Resource, D.S.K., J.A.K., H.Y.J and J.W.K.; Software, S.H.L. and J.Y.P.; Supervision, J.W.K.; Visualization, S.H.L.; Writing–original draft, S.H.L., D.S.K. and J.W.K.; Writing–review & editing, S.H.L. and J.W.K.
Data Availability Statement
All data related to this manuscript are incorporated in the manuscript only.
Acknowledgments
This study was carried out with the support of ´R&D Program for Forest Science Technology (Project No. FTIS 2019149C10-2323-0301)’ provided by Korea Forest Service (Korea Forestry Promotion Institute) and Technology Development Program to Solve Climate Changes through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT (NRF-2020R1F1A1054290)
Conflicts of Interest
The authors declare no conflict of interest.
References
- OH, Y.J.; Seo, H.R.; Choi, Y.M.; Jung, D.S. Evaluation of antioxidant activity of the extracts from the aerial parts of Cnidium officinale Makino. Korean Journal of Medicinal Crop Science, 2010, 18, 373-378.
- Leem, H.H.; Kim, E.O.; Seo, M.J.; Choi, S.W. Anti-inflammatory effects of volatile flavor extract from herbal medicinal prescriptions including Cnidium officinale Makino and Angelica gigas Nakai. Journal of the society of cosmetic scientists of Korea, 2011, 37, 199-210. [CrossRef]
- Seo, D.J.; Kim, H.C.; Lee, H.S.; Lee, S.J.; Lee, W.Y.; Han, S.H.; Kang J.W. Review of long-term climate change research facilities for forests. Korean Journal of Agricultural and Forest Meteorology, 2016, 18, 274-286. [CrossRef]
- Kim, K.K.; Kim, S.N.; Yoo, S.O.; An, J.G.; Kim, C.S.; Lee, M.S.; Kim, S.Y. Agricultural Technology Guide 007, Medicinal crops.; Hwang, J.W.; Cho, E.H.; Lim, E.S.; Jang, J.G.; Park, C.G.; Rural Development Administration: Jeonju, Korea, 2019; pp. 245-252, ISBN 978-89-480-3525-4 95520.
- Kim, Y.I.; Yu, H.H.; Yu, D.Y.; Jung, J.T.; Eom, Y.R. Changes in major components of Angelica Gigantis Radix and Glycyrrhizae Radix by storage temperature and period. Korean Medicinal Crop Society Conference Papers, 2015, 23, 336-337.
- Kim, K.Y.; Han, K.M.; Kim, H.J.; Kim, C.W.; Jeon, K.S.; Jung, C.R. Effect of Soil Properties on Soil Fungal Community in First and Continuous Cultivation Fields of Cnidium officinale Makino. Korean Journal of Medicinal Crop Science, 2020, 28, 209-220. [CrossRef]
- Kim, H.S.; Kim, J.Y.; Lee, S.M.; Park, H.J.; Lee, S.H.; Jang, J.S.; Lee, M.H. Isolation and characterization of various strains of Bacillus sp. having antagonistic effect against phytopathogenic fungi. Microbiology and Biotechnology Letters, 2019, 47, 603-613. [CrossRef]
- National Center for Biotechnology Information (NCBI)[Internet]. Bethesda (MD): National Library of Medicine (US), National Center for Biotechnology Information; [1988] – [cited 2017 Apr 06]. Available from: https://www.ncbi.nlm.nih.gov/ (accessed on 1 December 2022.).
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Molecular biology and evolution, 2021, 38, 3022-3027. [CrossRef]
- Kim, Y.S.; Kim, S.W.; Lamsal, K.; Lee, Y.S. Evaluation of rhizobacterial isolates for their antagonistic effects against various phytopathogenic fungi. The Korean Journal of Mycology, 2016, 44, 36-47. [CrossRef]
- Mekonnen, E.; Kebede, A.; Nigussie, A.; Kebede, G.; Tafesse, M. Isolation and characterization of urease-producing soil bacteria. International Journal of Microbiology. 2021, 2021, 8888641. [CrossRef]
- Ahmad, M.S.; Noor, Z.M.; Ariffin, Z.Z. Isolation and identification fibrinolytic protease endophytic fungi from Hibiscus leaves in Shah Alam. International Journal of Biological, Veterinary, Agricultural and Food Engineering. 2014, 8,1027-1030.
- Gohel, H.; Contractor, C.N.; Ghosh, S.K.; Braganza, V.J. A comparative study of various staining techniques for determination of extra cellular cellulase activity on Carboxy Methyl Cellulose (CMC) agar plates. Int J Curr Microbiol App Sci. 2014, 3, 261-266.
- Ramirez, M.G.; Avelizapa, L.I.R.; Avelizapa, N.G.R.; Camarillo, R.C. Colloidal chitin stained with Remazol Brilliant Blue R®, a useful substrate to select chitinolytic microorganisms and to evaluate chitinases. Journal of Microbiological Methods. 2004, 56, 213-219. [CrossRef]
- Liu, K.; Ding, H.; Yu, Y.; Chen, B. A cold-adapted chitinase-producing bacterium from Antarctica and its potential in biocontrol of plant pathogenic fungi. Marine drugs. 2019, 17, 695. [CrossRef]
- Deshwal, V.K.; Kumar, P. Production of plant growth promoting substance by Pseudomonads. J Acad Ind Res. 2013, 2, 221-215.
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS microbiology Letters. 1999, 170, 265-270. [CrossRef]
- Louden, B.C.; Haarmann, D.; Lynne, A.M. Use of blue agar CAS assay for siderophore detection. Journal of microbiology & biology education. 2011, 12, 51-53. [CrossRef]
- Glickmann, E.; Dessaux, Y. A critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Applied and environmental microbiology. 1995, 61, 793-796. [CrossRef]
- Gordon, T.R.; Okamoto, D.; Jacobson D.J. Colonization of muskmelon and nonsusceptible crops by Fusarium oxysporum f. sp. melonis and other species of Fusarium. Phytopathology. 1989, 79, 1095-1100.
- Wang, B.; Brubaker, C.L.; Burdon, J.J. Fusarium species and Fusarium wilt pathogens associated with native Gossypium populations in Australia. Mycological Research. 2004, 108, 35-44. [CrossRef]
- National Institute of Horticultural and Herbal Science, Korean Society for Horticultural Science. History of Korean Horticulture; Korean Society for Horticultural Science: Suwon, Korea, 2013; pp. 399-402, ISBN 978-89-97776-67-2 93520.
- Vaughan, A.; Guilbault, G.G.; Hackney, D. Fluorometric methods for analysis of acid and alkaline phosphatase. Analytical chemistry, 1971, 43, 721-724. [CrossRef]
- Janckila, A.J.; Takahashi, K.; Sun, S.Z.; Yam, L.T. Naphthol-ASBI phosphate as a preferred substrate for tartrate-resistant acid phosphatase isoform 5b. Journal of Bone and Mineral Research, 2001, 16, 788-793. [CrossRef]
- Ahmed, E.; Holmström, S.J. Siderophores in environmental research: Roles and applications. Microbial biotechnology. 2014, 7, 196-208. [CrossRef]
- Saha, M.; Sarkar, S.; Sarkar, B.; Sharma, B.K.; Bhattacharjee, S.; Tribedi, P. Microbial siderophores and their potential applications: A review. Environmental Science and Pollution Research. 2016, 23, 3984-3999. [CrossRef]
- Cochard, B.; Giroud, B.; Crovadore, J.; Chablais, R.; Arminjon, L.; Lefort, F. Endophytic PGPR from Tomato Roots: Isolation, In Vitro Characterization and In Vivo Evaluation of Treated Tomatoes (Solanum lycopersicum L.). Microorganisms 2022, 10, 765. [CrossRef]
- Nabila, N.; Kasiamdari, R.S. Antagonistic activity of siderophore-producing bacteria from black rice rhizosphere against rice blast fungus Pyricularia oryzae. Microbiology and Biotechnology Letters. 2021, 49, 217-224. [CrossRef]
- Woo, S.M.; Woo, J.W.; Kim, S.D. Purification and Characterization of the Siderophore from Bacillus licheniformis K11, a Multi-functional Plant Growth Promoting Rhizobacterium. Microbiology and Biotechnology Letters. 2007, 35, 128-34.
- Jeong, T.K.; Kim, J.H.; Song, H.K. Antifungal activity and plant growth promotion by rhizobacteria inhibiting growth of plant pathogenic fungi. Korean Journal of Microbiology. 2012, 48, 16-21. [CrossRef]
- Ohtakara, A. Chitinase and β-N-acetylhexosaminidase from Pycnoporus cinnabarinus. InMethods in Enzymology. 1988, 161, 462-470. [CrossRef]
- Mamarabadi, M.; Jensen, D.F.; Lübeck, M. An N-acetyl-β-d-glucosaminidase gene, cr-nag1, from the biocontrol agent Clonostachys rosea is up-regulated in antagonistic interactions with Fusarium culmorum. Mycological Research. 2009, 3, 33-43. [CrossRef]
- Tamura, K.; Sakazaki, R.; Kosako, Y.; Yoshizaki, E. Leclercia adecarboxylata Gen. Nov., Comb. Nov., formerly known asEscherichia adecarboxylata. Current Microbiology. 1986, 13, 79-84. [CrossRef]
- Richard, C. New Enterobacteriaceae found in medical bacteriology Moellerella wisconsensis, Koserella trabulsii, Leclercia adecarboxylata, Escherichia fergusonii, Enterobacter asbutiae, Rahnella aquatilis. InAnnales de biologie Clinique. 1989, 47, 231-236.
- Leclerc, H. Etude biochimique d'Enterobacteriaceae pigmentees. Ann Inst Pasteur 1962, 102, 726-741.
- Ahmed, B.; Shahid, M.; Syed, A.; Rajput, V.D.; Elgorban, A.M.; Minkina, T.; Bahkali, A.H.; Lee, J. Drought Tolerant Enterobacter sp./Leclercia adecarboxylata Secretes Indole-3-acetic Acid and Other Biomolecules and Enhances the Biological Attributes of Vigna radiata (L.) R. Wilczek in Water Deficit Conditions. Biology 2021, 10, 1149. [CrossRef]
- Danish, S.; Zafer-ul-Hye, M. Co-application of ACC-deaminase producing PGPR and timber-waste biochar improves pigments formation, growth and yield of wheat under drought stress. Scientific reports. 2019 Apr 12;9(1):1-3. [CrossRef]
- Bendaha, M.E.A.; Belaouni, H.A. Effect of the endophytic plant growth promoting EB4B on tomato growth. Hellenic Plant Protection Journal. 2020, 13, 54-65. [CrossRef]
- Steinkellner, S.; Mammerler, R.; Vierheilig, H. Microconidia germination of the tomato pathogen Fusarium oxysporum in the presence of root exudates. Journal of plant interactions. 2005, 1, 23-30. [CrossRef]
- Farda, B.; Djebaili, R.; Bernardi, M.; Pace, L.; Del Gallo, M.; Pellegrini, M. Bacterial Microbiota and Soil Fertility of Crocus sativus L. Rhizosphere in the Presence and Absence of Fusarium spp. Land. 2022, 11, 2048. [CrossRef]
- Kabisch, J.; Thürmer, A.; Hübel, T.; Popper, L.; Daniel, R.; Schweder, T. Characterization and optimization of Bacillus subtilis ATCC 6051 as an expression host. Journal of biotechnology. 2013, 163, 97-104. [CrossRef]
- Goto, K.; Omura, T.; Sadaie, Y. Application of the partial 16S rDNA sequence as an index for rapid identification of species in the genus Bacillus. The Journal of General and Applied Microbiology. 2000, 46, 1-8. [CrossRef]
- Yarza P.; Spröer C.; Swiderski J.; Mrotzek N.; Spring S.; Tindall B.J.; Gronow S.; Pukall R.; Klenk H.P.; Lang E.; et al. Sequencing orphan species initiative (SOS): Filling the gaps in the 16S rRNA gene sequence database for all species with validly published names. Systematic and Applied Microbiology. 2013, 36, 69-73. [CrossRef]
- Takahashi, N.; Shimomura, T. Action of rice α-glucosidase on maltose and starch. Agricultural and Biological Chemistry. 1973, 37, 67-74. [CrossRef]
- Morant, A.V.; Jørgensen, K.; Jørgensen, C.; Paquette, S.M.; Sánchez-Pérez, R.; Møller, B.L.; Bak, S. β-Glucosidases as detonators of plant chemical defense. Phytochemistry. 2008, 69, 1795-1813. [CrossRef]
- Zhang, D.; Spadaro, D.; Valente, S.; Garibaldi, A.l Gullino, M.L. Cloning, characterization, expression and antifungal activity of an alkaline serine protease of Aureobasidium pullulans PL5 involved in the biological control of postharvest pathogens. International journal of food microbiology. 2012, 153, 453-464. [CrossRef]
- Martinez, M.; Gómez-Cabellos, S.; Giménez, M.J.; Barro, F.; Diaz, I.; Diaz-Mendoza, M. Plant proteases: From key enzymes in germination to allies for fighting human gluten-related disorders. Frontiers in plant science. 2019, 10, 721. [CrossRef]
- Hashem, A.; Tabassum, B.; Abd_Allah, E.F. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi journal of biological sciences. 2019, 26, 1291-1297. [CrossRef]
- Sivasakthi, S.; Usharani, G.; Saranraj, P. Biocontrol potentiality of plant growth promoting bacteria (PGPR)-Pseudomonas fluorescens and Bacillus subtilis: A review. African journal of agricultural research. 2014, 9, 1265-1277.
- Almaghrabi, O.A.; Abdelmoneim, T.S.; Albishri, H.M.; Moussa, T.A. Enhancement of maize growth using some plant growth promoting rhizobacteria (PGPR) under laboratory conditions. Life Sci J. 2014, 11, 764-772.
- Baldrian, P. Forest microbiome: Diversity, complexity and dynamics. FEMS Microbiology reviews. 2017, 41, 109-130. [CrossRef]
- Wang, H.; Liu, R.; You, M.P.; Barbetti, M.J.; Chen, Y. Pathogen Biocontrol Using Plant Growth-Promoting Bacteria (PGPR): Role of Bacterial Diversity. Microorganisms 2021, 9, 1988. [CrossRef]
- Doty, S.L.; Freeman, J.L.; Cohu, C.M.; Burken, J.G.; Firrincieli, A.; Simon, A.; Khan, Z.; Isebrands, J.G.; Lukas, J.; Blaylock, M.J. Enhanced degradation of TCE on a superfund site using endophyte-assisted poplar tree phytoremediation. Environmental science & technology. 2017, 51, 10050-10058. [CrossRef]
Figure 1.
Neighbor-joining phylogenetic tree based on the 16S rRNA genes of each selected strain. The trees were compared to the NCBI 16s rRNA database. The numbers at the nodes indicate bootstrap values >50% with 1000 bootstrap replications. ST7, ST8, and SP4 were isolated from rhizosphere soil and PT1 was isolated from the root tissue. Lactiplantibacillus sp., Paenibacillus sp., shewanella sp., and Paraferrimonas sp. are used as the out-group.
Figure 1.
Neighbor-joining phylogenetic tree based on the 16S rRNA genes of each selected strain. The trees were compared to the NCBI 16s rRNA database. The numbers at the nodes indicate bootstrap values >50% with 1000 bootstrap replications. ST7, ST8, and SP4 were isolated from rhizosphere soil and PT1 was isolated from the root tissue. Lactiplantibacillus sp., Paenibacillus sp., shewanella sp., and Paraferrimonas sp. are used as the out-group.
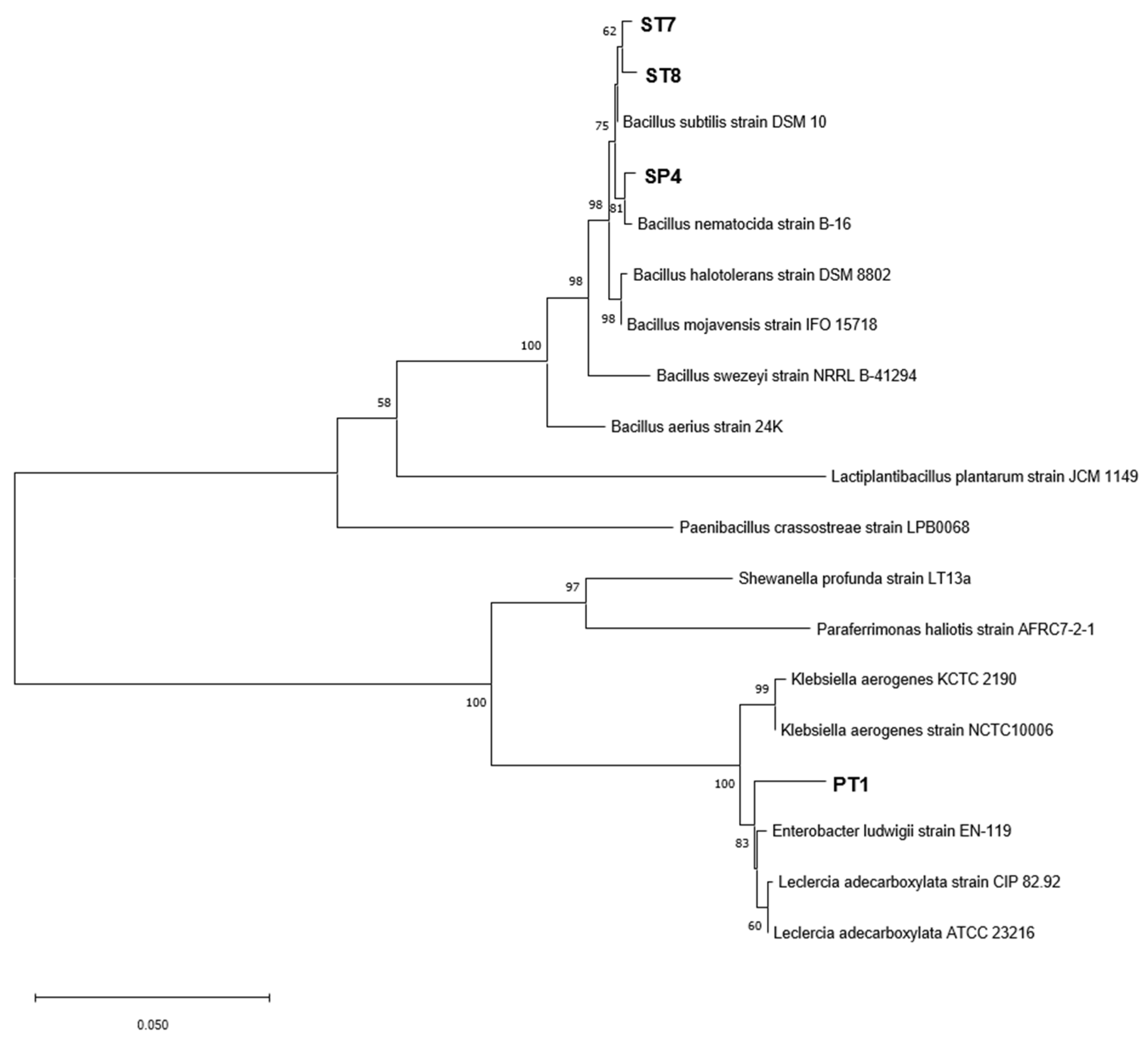
Figure 2.
Dual culture test for each of the selected strains to show antagonistic activity against F. solani (pathogenic fungi). The top half of each plate is inoculated with the selected strain on a paper disk, and the lower half is inoculated with F. solani in the form of an agar plug.
Figure 2.
Dual culture test for each of the selected strains to show antagonistic activity against F. solani (pathogenic fungi). The top half of each plate is inoculated with the selected strain on a paper disk, and the lower half is inoculated with F. solani in the form of an agar plug.
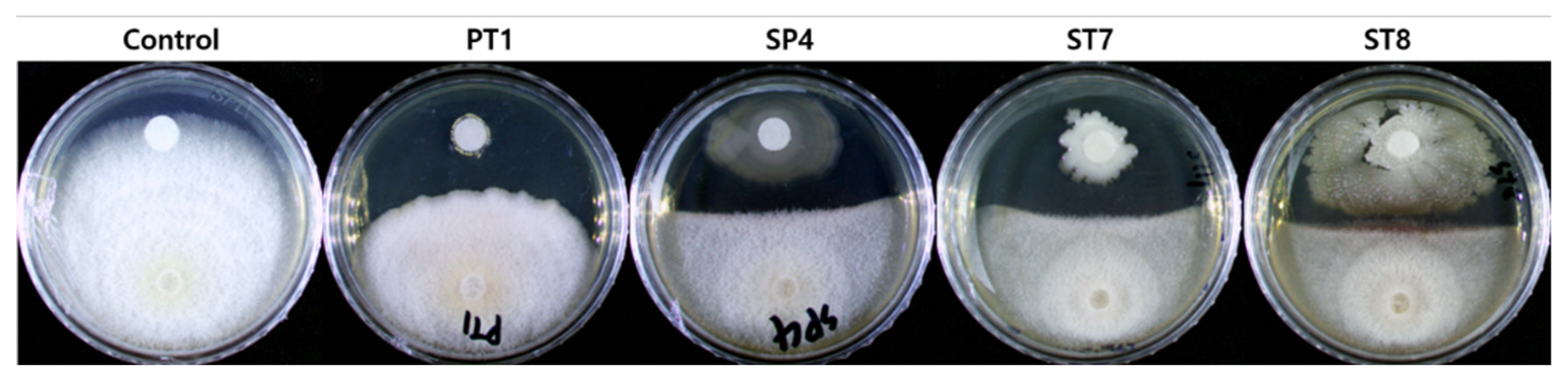
Figure 3.
In planta test for the antagonistic activity of each selected strains (a) C. officinales were maintained at 25℃ for 17 d after inoculation (after 28 d of growth); Boxplots showing rate of (b) dead shoots; (c) shoot dry weight; and (d) root + rhizome dry weight. Different letters indicate statistically significant differences (p < 0.05). Con, first control group that was not inoculated; Fu, a second control group was inoculated with only F. solani (pathogenic fungi); PT1, SP4, ST7, and ST8 groups were inoculated with each of the selected strains and F. solani, respectively. Line in the box represents median, the top of box is the 75th percentile, the bottom of the box is the 25th percentile, whiskers represent minimum and maximum values and points represent outliers.
Figure 3.
In planta test for the antagonistic activity of each selected strains (a) C. officinales were maintained at 25℃ for 17 d after inoculation (after 28 d of growth); Boxplots showing rate of (b) dead shoots; (c) shoot dry weight; and (d) root + rhizome dry weight. Different letters indicate statistically significant differences (p < 0.05). Con, first control group that was not inoculated; Fu, a second control group was inoculated with only F. solani (pathogenic fungi); PT1, SP4, ST7, and ST8 groups were inoculated with each of the selected strains and F. solani, respectively. Line in the box represents median, the top of box is the 75th percentile, the bottom of the box is the 25th percentile, whiskers represent minimum and maximum values and points represent outliers.
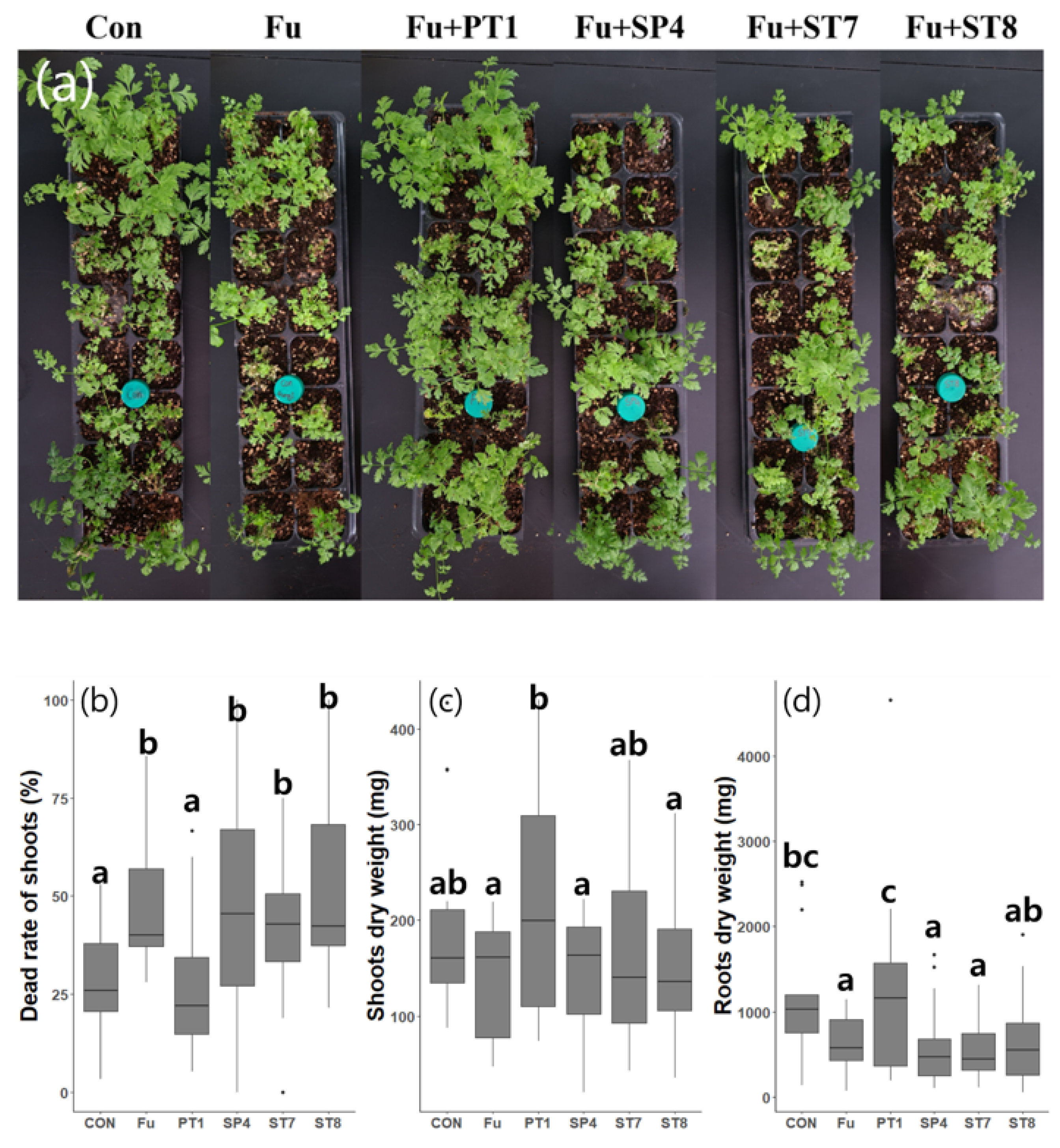
Figure 4.
Bacterial growth curve characterization for each selected strain under different temperatures (5℃–40℃). Each strain was inoculated in TSB broth at OD 0.01 at 600 nm and cultured for 120 h. Each point represents the mean of three replicates and stand error (SE). (a) 5℃; (b) 10℃; (c) 15℃; (d) 20℃; (e) 25℃; (f) 30℃; (g) 35℃ and (h) 40℃.
Figure 4.
Bacterial growth curve characterization for each selected strain under different temperatures (5℃–40℃). Each strain was inoculated in TSB broth at OD 0.01 at 600 nm and cultured for 120 h. Each point represents the mean of three replicates and stand error (SE). (a) 5℃; (b) 10℃; (c) 15℃; (d) 20℃; (e) 25℃; (f) 30℃; (g) 35℃ and (h) 40℃.
Figure 5.
Agar plates showing the production of various enzymes for each selected strain. For the protease production, cellulase production, and phosphate solubilizing tests, strains that formed a clear zone indicated a positive result. For the siderophore production test, a strain that formed a halo zone indicated a positive result.
Figure 5.
Agar plates showing the production of various enzymes for each selected strain. For the protease production, cellulase production, and phosphate solubilizing tests, strains that formed a clear zone indicated a positive result. For the siderophore production test, a strain that formed a halo zone indicated a positive result.
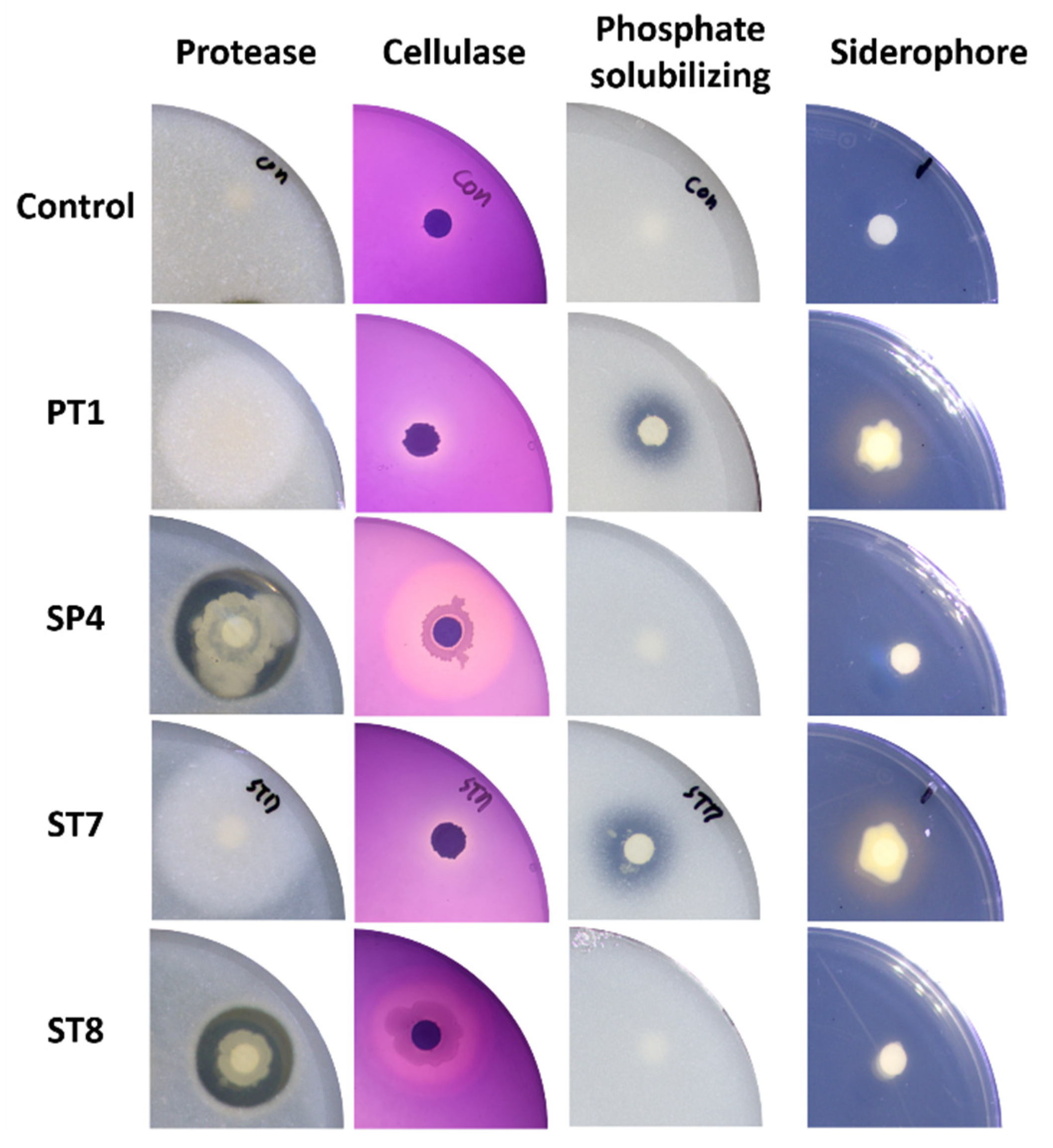

Table 1.
BLASTn results for the four selected strains in the standard NCBI database.
| Strains | 16S rRNA identification1 | GenBank Accession No.1 |
Similarity (%) | Isolated from |
| ST7 | Bacillus inaquosorum | NR_104873.1 | 99 | Rhizosphere soil |
| ST8 | Bacillus subtilis | CP019663.1 | 99 | Rhizosphere soil |
| SP4 | Bacillus vallismortis | NR_024696.1 | 99 | Rhizosphere soil |
| PT1 | Leclercia adecarboxylata | NR_104933.1 | 99 | Root tissue |
1 Species and strains from each database with the highest levels of similarity are indicated.
Table 2.
Enzyme production characterization to assess the PGP traits of each strain.
| Strain | Protease | Cellulase | Chitinase | HCN | PS | SP | IAA (ppm) |
| PT1 | - | - | - | - | + | + | 13.19 ± 4.22 |
| SP4 | + | + | - | - | - | - | NA |
| ST7 | - | - | - | - | + | + | NA |
| ST8 | + | + | - | - | - | - | NA |
+ indicates a positive result, and indicates a negative result. HCN, hydrogen cyanide; PS, phosphate solubilizing; SP, siderophore; IAA, indole-3-acetic acid.
Table 3.
Enzyme production characterization of each strain using APIZYM.
| Enzyme | PT1 | SP4 | ST7 | ST8 |
| Alkaline phosphatase | + | + | + | + |
| Esterase (C4) | + | + | + | + |
| Esterase lipase (C8) | + | + | + | + |
| Lipase (C14) | - | - | - | - |
| Leucine arylamidase | - | - | - | - |
| Valine arylamidase | - | - | - | - |
| Crystine arylamidase | - | - | - | - |
| Trypsin | - | - | - | - |
| a-chymotrypsin | - | - | - | - |
| Acid phospatase | + | - | + | + |
| Naphthol-AS-BI-phosphohydrolase | + | + | + | + |
| α-galactosidase | - | - | - | - |
| β-glucuronidase | - | - | - | - |
| β-glucosidase | - | - | - | - |
| α-glucosidase | - | + | - | + |
| β-glucosidase | - | - | + | + |
| N-acetyl-β-glucosaminidase | + | - | - | - |
| α-mannosidase | - | - | - | - |
| α-fucosidase | - | - | - | - |
+ indicates a positive result, and–indicates a negative result.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



