Submitted:
12 May 2023
Posted:
15 May 2023
You are already at the latest version
Abstract
Porphyromonas gingivalis is a periodontopathogenic bacterium that can adhere and colonize periodontal tissues leading to inflammatory process, and consequently tissue destruction. New therapies using flavonoids such as hesperidin are being studied, and its promising properties have been highlighted. The aim of this study was to evaluate the effect of hesperidin on epithelial barrier function, reactive oxygen species (ROS) production and on inflammatory response caused by P. gingivalis in in vitro models. Integrity of the epithelial tight junctions challenged by P. gingivalis was determined by monitoring the transepithelial electrical resistance (TER). P. gingivalis adherence to a gingival keratinocyte monolayer and a basement membrane model were evaluated by a fluorescence assay. A fluorometric assay was used to determine the ROS production in gingival keratinocytes. The level of pro-inflammatory cytokines and matrix metalloproteinases (MMPs) secretion was evaluated by ELISA, while to assess NF-κB activation, the U937-3xjB-LUC monocyte cell line transfected with a luciferase reporter gene was used. Hesperidin protected against gingival epithelial barrier dysfunction caused by P. gingivalis and reduced the adherence of P. gingivalis to the basement membrane model. Hesperidin dose-dependently inhibited P. gingivalis-mediated ROS production by oral epithelial cells as well as the secretion of IL-1β, TNF- IL-8, MMP-2 and MMP-9 by macrophages challenged with P. gingivalis. Additionally, it was able to attenuate NF-κB activation in macrophages stimulated with P. gingivalis. These findings suggested that hesperidin has a protective effect on the epithelial barrier function, in addition to reduce ROS production and attenuate the inflammatory response associated with periodontal disease.
Keywords:
hesperidin
; inflammation
; oxidative stress
; macrophague
; keratynocite
; Porphyromonas gingivalis
1. Introduction
A large diversity of microorganisms resides in the oral cavity and colonize teeth, tongue, cheeks, gingival sulcus, tonsils, hard palate and soft palate [1]. In periodontal disease, an alteration in the microbial composition and/or pathogenicity occurred, resulting in microbial dysbiosis [2] that can induce an exacerbated inflammatory response associated with the secretion of cytokines and matrix metalloproteinases (MMPs), which promote destruction of the tooth supporting structures, including the periodontal ligament and alveolar bone [3,4]. Periodontal tissue damage is also caused by an excessive production of reactive oxygen species (ROS) which contribute to cellular apoptosis and cause oxidative stresses [5]. As described by Socransky et al. [6], “the red complex” composed by Porphyromonas gingivalis, Tannerella forsythia, and Treponema denticola has been strongly associated with the severity of periodontitis. Among these periodontopathogenic bacteria, the Gram-negative bacterium P. gingivalis is thought to cause the dysbiosis condition and initiate the inflammatory process of periodontal disease [7].
The oral epithelium is the main structure of the oral cavity that provides physical, chemical, and immunological barriers against chemical or microbial challenge [8]. During periodontal disease, colonization of the gingival sulcus by periodontopathogenic bacteria leads to the release of microbial virulence factors and toxins, including lipopolysaccharide (LPS) that stimulate the inflammatory process and tissue destruction [9]. P. gingivalis, known as a key periodontopathogenic bacterium, can express adhesive components [10] and damage the epithelial barrier through proteolytic degradation of the junction proteins [11]. Gingipains, including Arg-gingipains A and B, and Lys-gingipain, are the major extracellular and cell-bound proteolytic activities produced by P. gingivalis [12]. More specifically, gingipains mediate the collagen degradation, mainly type I collagen, which is particularly abundant in periodontal tissues [12]. In addition, gingipains induce host defense perturbation through degradation of cellular signaling molecules and inactivation of cellular functions [12,13].
To enhance the outcome of periodontal disease treatments, several adjuncts to non-surgical periodontal therapy have been proposed, including local delivery drugs such as antibiotics and antimicrobials agents. In recent years, natural products have been extensively studied due to their high safety, cost-effectiveness and beneficial properties as anti-inflammatory, antioxidant, and antimicrobial. In this context, hesperidin is a flavonoid glycoside found in high amounts in citrus fruits such as sweet oranges, lemons, limes, and tangerines [14]. Hesperidin has been reported to exert beneficial effects on various diseases, including neurological, cardiovascular and other disorders, due to their anti-inflammatory, anti-oxidative and anti-cancer properties [15]. With respect to periodontal disease, in vivo studies using rodent models showed that hesperidin can promote an ameliorative effect on the alveolar bone loss and attenuate the intensity of the inflammatory process [16,17].
Despite evidence suggesting a promising effect of hesperidin for periodontal disease, additional studies are necessary to elucidate the underlying mechanisms of action of this flavonoid. Therefore, the purpose of this study was to investigate the effects of hesperidin on P. gingivalis-mediated barrier function damage and ROS production in an oral epithelial model. Moreover, the ability of hesperidin to attenuate the expression of inflammatory cytokines and MMPs by P. gingivalis-stimulated macrophages was studied.
2. Results
2.1. Effect of hesperidin on the disruption of ephitelial barrier integrity
The epithelial barrier is a complex and critical structure that provides protection against tissue invasion by periodontopathogenic bacteria. In this context, we evaluated the ability of hesperidin to protect the integrity of the oral epithelial barrier withouth stimulus or against P. gingivalis by monitoring TER values over a 24-h period. Initially, we performed an MTT assay to determine non-cytotoxic concentrations of hesperidin. As shown in Figure 1, concentrations ≤ 1280 µM of hesperidin did not significantly decrease the cell viability after 48 h of exposition.
The statistical analysis showed that hesperidin was able to increase TER values following a 2-, 4- and 8-h exposure compared with control group (Figure 2A). In detail, hesperidin at 640 µM showed an increase of TER by 80.61% and 28.31% after 2- and 4-h, respectively. Also, a high TER values were observed after the treatment with hesperidin at 1280 µM resulted in an increase of 114.38%, 41.54% and 16.11% after 2-, -4 and 8-h. In general, hesperidin showed a dose-dependently effect. Treating the oral epithelial barrier model simultaneously with hesperidin and P. gingivalis was associated with significant higher TER values after 8- and 24-h treatments compared to treating the model with P. gingivalis alone (Figure 2B). More specifically, after an 8-h treatment, hesperidin at 640 µM showed an increase of TER by 15.36% while hesperidin at 1280 µM resulted in an increase of 79.81% after 8 h. Extending the incubation to 24 h showed an increase of TER of 13.22% and 61.13% for hesperidin at 640 µM and 1280 µM, respectively. Treating the oral epithelial barrier model for 24 h with P. gingivalis in the absence of hesperidin showed a significant decrease of TER values (187.90%) compared to the control group (without P. gingivalis stimulus).
2.2. Effect of hesperidin on the adherence of P. gingivalis and a basement membrane model
Figure 3 reports the effect of hesperidin on the adherence of P. gingivalis to human oral epithelial cells and to a basement membrane model. On the one hand, hesperidin did not show any capacity to reduce the adherence of P. gingivalis to human oral ephitelial cells (Figure 3A). On the other hand, the flavonoid dose-dependently inhibited the adherence of P. gingivalis to the basement membrane model (Figure 3B). Hesperidin at 640 µM and 1280 µM reduced bacterial adhesion by 21.11% and 43.06%, respectively.
2.3. Effect of hesperidin on ROS production
The excessive production of ROS by human oral epithelial cells may contribute to destruction of the tooth-supporting structures [18]. As reported in Figure 4, exposure of oral epithelial cells to P. gingivalis (MOI = 1000) time-dependently mediated ROS production. More specifically, a 3-fold increase was observed following a 5-h contact time. The presence of hesperidin (320, 640, and 1280 µM) reduced P. gingivalis-induced ROS production between 11.0% to 15.9% (5-h contact).
The effect of hesperidin on ROS production by human oral epithelial cells co-stimulated with P. gingivalis and H2O2 was also evaluated. Figure 5 shows high levels of ROS production by oral epithelial cells treated with P. gingivalis and H2O2; a 102.6-fold increase compared to the control group after 5-h contact time. Epithelial cells stimulated with P. gingivalis and H2O2 separately showed only a 11.4- and 3.3-fold increase in the production of ROS compared to the control group thus suggesting a synergistic effect between P. gingivalis and H2O2. However, the presence of hesperidin did not significantly attenuated ROS production in this synergistic model.
2.4. Effect of hesperidin on cytokines and MMPs expression
To assess the effect of hesperidin on the inflammatory response associated with periodontitis, macrophages were pre-treated for 2 h with non-cytotoxic concentrations of hesperidin and stimulated with P. gingivalis (MOI of 100) for 24 h. First, hesperidin at concentrations of 20, 40 and 80 µM were determined as being non-cytotoxic for macrophages in an MTT colorimetric assay (Figure 6).
Subsequent experiments using ELISA assay were performed to assess the levels of selected cytokines and MMPs secreted by macrophages. As shown in Figure 7, an increased secretion of IL-6, IL-8, TNF-α, and IL-1β by macrophages was observed when exposed to P. gingivalis, compared with un-stimulated controls. An increase of 17.6-, 8.5-, 138.5-, and 4.2-fold was found for IL-6, IL-8, TNF-α, and IL-1β, respectively. Hesperidin dose-dependently down-regulated the secretion of IL-8, TNF-α, and IL-1β. However, this effect was not observed for IL-6 secretion. More specifically, hesperidin at a concentration of 80 µM was able to inhibit 29.9, 42.0 and 30.2% of IL-8, TNF-α, and IL-1β secretion, respectively.
The effect of hesperidin on MMP secretion is shown in Figure 8. The results indicate that levels of MMP-2, MMP-8, and MMP-9 increased by 2.4-, 1.6-, and 1.8-fold after P. gingivalis exposition when compared to un-stimulated controls. Hesperidin at a concentration of 80 µM in P. gingivalis-stimulated macrophages decreased the secretion of MMP-2 and MMP-9 by 74.4 and 30.4%, respectively. This effect was also dose-dependent. However, hesperidin did not show any significant inhibition on MMP-8 secretion by macrophages stimulated with P. gingivalis.
2.5. Effect of hesperidin on NF-κB activation
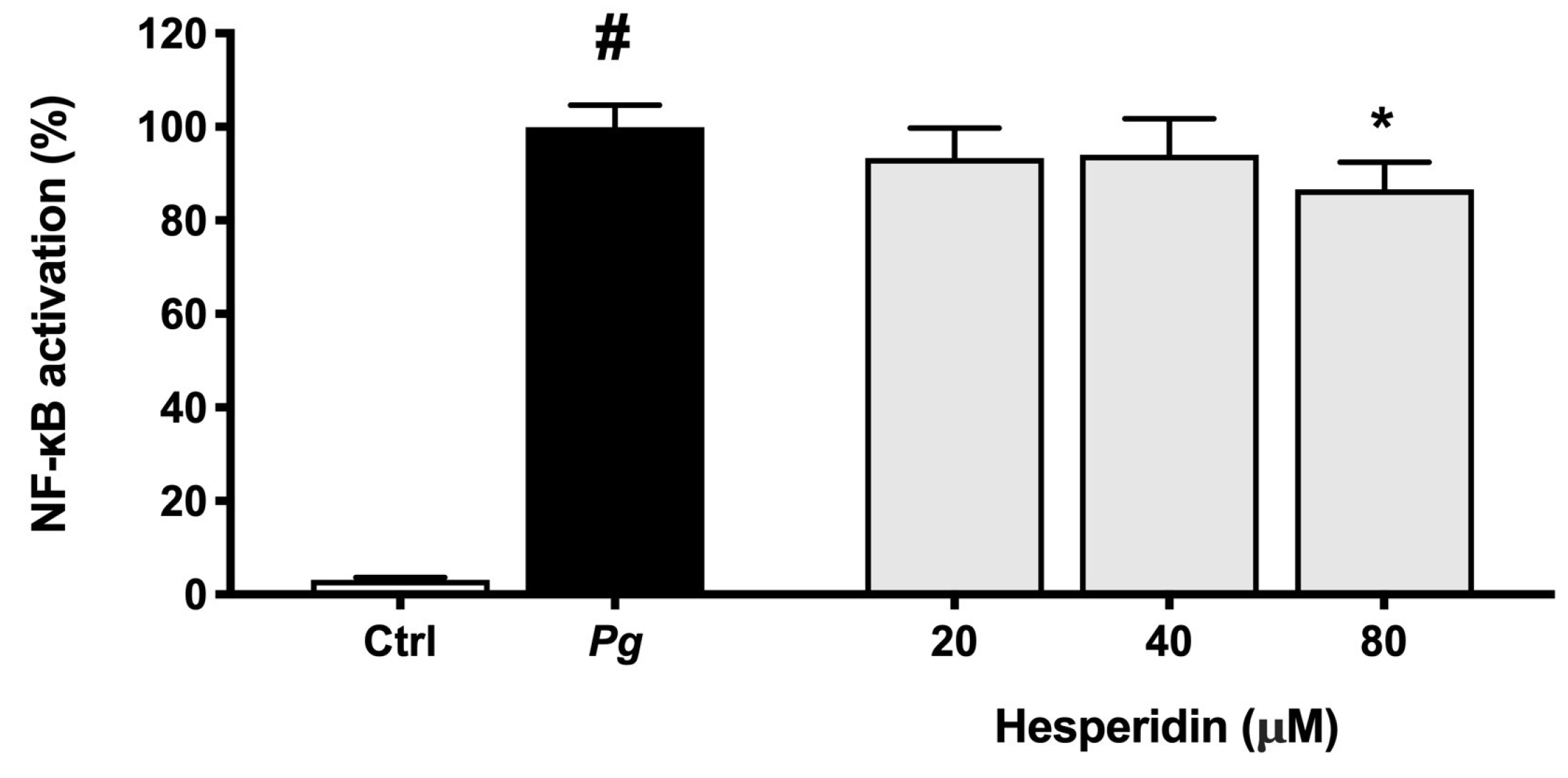
Lastly, to investigate the mechanism responsible for the inhibitory effect of hesperidin on the inflammatory response of P. gingivalis-stimulated macrophages, we verified the NF-κB activation after treatment with hesperidin. To this end, an alternative model using the U937 3xκ B-LUC monocytic cell line was used. Figure 9 showed a significant inhibition (13.4%) of NF-κB activation after exposure to hesperidin at a concentration of 80 µM.
3. Discussion
In periodontal disease, damage and destruction of periodontal tissues are caused mainly by an inappropriate host response to microorganisms and their products. P. gingivalis can disturb the epithelial integrity and invade into the deeper periodontal tissues triggering an inflammatory response [19]. In this context, therapies involving agents that enhance the oral mucosa barrier and exhibit anti-inflammatory and antioxidant properties are promising for periodontal disease treatment. Among them, hesperidin is an attractive flavonoid with excellent biological properties that can modulate the epithelial cellular immune response [15]. Then, the aim of the present study was to investigate the effect of hesperidin on P. gingivalis-mediated damage to the gingival epithelial barrier function, ROS production and inflammatory response using in vitro models.
The gingival epithelium is an important barrier against bacterial invasion and for this reason becomes an important early line of defense in the oral cavity [20]. Considering this concept, diverse flavonoids with capacity to enhance the oral epithelial barrier function are interesting candidates to the treatment or prevention of periodontal disease. In the present in vitro study, we reported interested results of TER after the treatment with hesperidin. More specifically, hesperidin showed a significant increase in TER following a short exposure (2,4 and 8 h). Although the protect capacity of hesperidin on oral epithelial barrier has not been investigated, Guo et al. [21] showed the capacity of hesperidin of enhance the barrier integrity as well as epithelial permeability in Caco-2 cells.
The ability of P. gingivalis to invade epithelial cells is triggered via the binding of fimbriae to cellular adhesion molecules thereby disrupting the epithelial integrity [22,23]. These cell-cell interactions are critical for the innate immune response against microbial and toxic challenge [24]. In the present study, using an in vitro epithelial barrier model stimulated with P. gingivalis, cells revealed a progressive increase of the TER after 2 and 8 h of incubation. After 24 h, the TER values began to decrease indicating a damage on the function of the epithelial barrier. These data are in agreement with a previous in vitro study that showed the same tendency in TER values [19]. On the other hand, our results showed that cells infected with P. gingivalis and treated with hesperidin showed a positive effect after 24 h by increasing the TER values when compared to the cells infected with bacteria alone. It remains unclear what is the exact mechanism of the protective effect of hesperidin on the epithelial barrier integrity in our model.
P. gingvalis is able to attach to the periodontal tissues and several investigations has been reported regarding its adhesion to epithelial cells [25]. Using an in vitro study, we showed that P. gingivalis was able to adhere to oral epithelial cells. However, hesperidin did not show a capacity to decrease this adhesion. In contrast, using a basement membrane model, hesperidin was able to decrease the adherence of P. gingivalis. Then, our data suggest that hesperidin may alter the ability of P. gingivalis to adhere to several extracellular matrix proteins, including laminin and type IV collagen, which are constituents of the basement membrane model. To the best of our knowledge, no study previously evaluated the effect of hesperidin on adherence of periodontopathogens to epithelial cells or periodontal tissues/cells.
During periodontal disease, the exacerbated production of ROS is responsible for cell apoptosis and dysfunction, alveolar bone loss, and periodontal inflammation [26]. In this way, the attenuation or inhibition of ROS production is a relevant and important event to regulate several signaling pathways, and consequently inflammatory response [27]. In the present study, we proposed two ROS production models using P. gingilvalis alone and P. gingivalis + H2O2 (exacerbated ROS production model) as stimulus. H2O2 is known as a major component of ROS and is widely used as an inducer of oxidative stress. Based in our results, hesperidin can inhibit the ROS production in response to P. gingivalis. However, hesperidin did not show down-regulating effects in an event with exacerbate levels of ROS production. Previous in vitro studies showed that hesperidin treatment was able to reduce the by-products of lipid peroxidation in the human erythrocyte membrane [28,29]. Moreover, hesperidin attenuates the production of intracellular ROS in macrophages stimulated with lipopolysaccharide [30]. On the other hand, in vivo studies using models of supplementation in rats showed that hesperidin reduces the expression of levels of ROS and thiobarbituric acid reactive substances (TBARS) and increased activity of antioxidants [31,32,33]. Additionally, Lim et al. [34] demonstrated the marked ability of hesperidin to inhibit high glucose-induced intracellular ROS production in SH−SY5Y neuronal cells. Despite the fact that the literature shows its promising antioxidant property, our results using P. gingivalis bacteria associated or not with H2O2 are not in accordance with them. This may be because P. gingivalis is able to invade and survive into epithelial cells and then accumulate a hemin layer on the cell surface that have a function of provide oxidative stress protection [35]. Therefore, more studies are necessary to understand the antioxidant mechanism and effect of hesperidin in the ROS production.
Since P. gingivalis plays an important role in the induction of inflammatory processes, we evaluated the anti-inflammatory activity of hesperidin against this periodontopathogen. Using a monocyte-macrophage model stimulated with bacteria, we brought evidence that hesperidin was able to inhibit the secretion of IL-1β, TNF-α and IL-8. Previously, the anti-inflammatory potential of hesperidin was demonstrated using various cell models [36,37] and the data are in according with our results. In contrast to our results, Li et al. [38] among the cytokines evaluated, also showed an inhibition of IL-6 after the treatment with hesperidin in an in vivo protocol.
High levels of MMPs such as MMP-1, -2, -8, -9 and -13 are related with the severity and tissue destruction of periodontal disease [39,40]. In this context, the downregulation of MMP-2 and -9 by hesperidin as observed in our results may promote an anti-collagenase effect. These findings are in agreement with previous studies that reported the effect of hesperidin to inhibit the expression of these MMPs [41,42,43]. Additionally, we showed for the first time that hesperidin was not able to reduce the expression of MMP-8 level. There are not previous data exploring the mechanism of hesperidin on MMP-8 expression. Overall, our results showed a selective inhibition of IL-1β, TNF-α, IL-8, MMP-2 and MMP-9 associated with hesperidin could represent a promising novel approach to the treatment or prevention of periodontal disease.
NF-κB pathway is a main key component of the inflammatory response that can be triggered by bacteria and their toxic products resulting in cytokine expression [44]. Therefore, the capacity of compounds to inactivate this pathway become an important characteristic for modulation of inflammatory response. In this context, our results showed that hesperidin was able to inhibit the NF-κB activation in human monoblastic leukemia cell line transfected with a luciferase gene coupled to a promoter of three NF-κ B- binding sites. As expected, these findings suggest that the inhibitory effect of hesperidin on pro-inflammatory cytokine expression is significantly mediated through the inhibition of the NF-κB pathway. Similarly, Sato et al. [45] suggested that hesperidin has a significant effect on the regulation of NF-κB signaling pathway.
4. Materials and Methods
4.1. Hesperidin
Hesperidin was obtained from the USDA-ARS Horticultural Research Laboratory (Fort Pierce, Florida, USA). A stock solution at a concentration of 1 M was prepared in dimethylsulfoxide (DMSO) and stored at 4°C in the dark. In all assays described below, the final concentration of DMSO in the culture medium was ≤ 0.1% (w/v).
4.2. Bacteria and growth conditions
The reference strain P. gingivalis ATCC 33277 was used in this study. Bacteria were grown in an anaerobic chamber (80% N2, 10% CO2, 10% H2) at 37°C in Todd-Hewitt broth (THB; Becton Dickinson and Company, Sparks, MD, USA) containing 0.001% (w/v) hemin and 0.0001% (w/v) vitamin K.
4.3. Cell cultures
The B11 human oral epithelial cell line [46], which was kindly provided by S. Gröger (Justus Liebig University Giessen, Germany), was cultivated in keratinocyte serum-free medium (K-SFM; Life Technologies Inc., Burlington, ON, Canada) supplemented with 50 µg/mL of bovine pituitary extract, 5 ng/mL of human epidermal growth factor, 100 µg/mL of penicillin G-streptomycin, and 0.5 µg/mL of amphotericin B. The U937 human monocyte cell line (CRL-1593.2), which was purchased from the American Type Culture Collection (Manassas, VA, USA) was cultivated in Roswell Park Memorial Institute 1640 medium (RPMI; Life Technologies Inc.) supplemented with 10% heat-inactivated fetal bovine serum (FBS) and 100 µg/mL of penicillin G/streptomycin. The human monoblastic leukemia cell line U937 3xκ B-LUC, a subclone of the U937 cell line transfected with a luciferase gene coupled to a promoter of three NF-κ B- binding sites [47], was kindly provided by R. Blomhoff (University of Oslo, Norway). This cell line was cultured in RPMI-1640 supplemented with 10% FBS, 100 μg/mL of penicillin G/streptomycin and 75 μg/mL of hygromycin B. All cell cultures were incubated in a humidified incubator with a 5% CO2 atmosphere at 37ºC.
4.4. Oral epithelial barrier integrity
To investigate the effect of hesperidin on the epithelial barrier integrity challenged or not with P. gingivalis, the B11 human oral epithelial cell line [46] was used. First, non-cytotoxic concentrations of hesperidin were determined using a MTT (3-[4,5-diethylthiazol-2-yl]-2,5diphenyltetrazolium bromide) colorimetric assay (Roche Diagnostics, Laval, QC, Canada) according to the manufacturer’s instructions following a 48-h treatment. The epithelial cells were seeded onto TranswellTM clear polyester membrane insert (6.5 mm diameter, 0.4 μm pore size; Corning Co., Cambridge, MA, USA) (3 x 105 cells per insert). Basolateral and apical compartments were filled with 0.6 mL and 0.1 mL of culture medium, respectively. Then, the plates were incubated at 37ºC in a 5% CO2 atmosphere for 72 h. After this period, the culture medium was replaced with fresh antibiotic-free K-SFM, and the plates were further incubated for 16 h. Then, in the first setup, non-cytotoxic concentrations of hesperidin (640 and 1280 µM) were added to the apical compartments, and in the second setup, P. gingivalis at a multiplicity of infection (MOI) of 104 prepared in antibiotic-free K-SFM was associated or not with non-cytotoxic concentrations and added to the apical compartments. Finally, the integrity of the epithelial tight junctions was determined by monitoring the transepithelial electrical resistance (TER) using an ohmmeter (EVOM2, World Precision Instruments, Sarasota, FL, USA). TER was determined after 0, 2, 4, 8, and 24 h of incubation at 37ºC in a 5% CO2 atmosphere. Resistance values were calculated in Ohms (Ω)/cm2 by multiplying the resistance values by the surface area of the membrane filter. Results were expressed as a percentage of the basal control value measured at time 0 (100% value). Assays were carried out in triplicate in two independent experiments and the means ± standard deviations were calculated.
4.5. Adherence of P. gingivalis to human oral epithelial cells and a basement membrane model
The effect of hesperidin on the adherence of P. gingivalis to human oral epithelial cells and a basement membrane model (Cultrex Basement Membrane Extract [BME]; R&D Systems, Minneapolis, MN, USA) was evaluated, as previously described [48]. First, a 24-h culture of P. gingivalis was harvested by centrifugation (9,000 x g for 5 min), washed in 10 mm phosphate-buffered saline (PBS; pH 7.2), and suspended in K-SFM medium or 50 mM PBS (pH 8) containing 0.03 mg/mL of fluorescein isothiocyanate (FITC), to assess adherence to B11 human oral epithelial cell or the basement membrane model, respectively. Then, the P. gingivalis suspension was incubated in the dark at 37°C for 30 min with constant shaking. Thereafter, the labeled bacteria were washed three times by centrifugation (9,000 x g for 5 min). In a first analysis, oral epithelial cells (1 × 106 cells/mL) were seeded in sterile black wall, clear flat bottom 96-well microplates (Greiner Bio-One North America Inc.) and were incubated overnight at 37°C in a 5% CO2 atmosphere to allow cell adherence. Then, the medium was removed and the cell monolayers were pre-incubated with hesperidin (320, 640 or 1280 µM; in K-SFM) for 30 min prior to adding FITC-labeled P. gingivalis at an MOI of 1000. The cells were incubated at 37°C in a 5% CO2 atmosphere. After 4 h, unbound bacteria were removed, the wells were washed with PBS and the number of bacteria adhered was determined by relative fluorescence units (RFU) using a Synergy 2 microplate reader (BioTek Instruments, Winooski, VT, USA) (excitation wavelength of 495 nm and an emission wavelength of 525 nm). In a second analysis, the BME containing laminin, collagen IV, entactin, and heparin sulfate proteoglycan was diluted 1:10 in ice-cold PBS and 100 μL were added to the wells of 96-well clear bottom black microplates. The microplates were maintained for 2 h at room temperature to allow gelification. Then, the wells were washed with PBS, and two-fold serial dilutions of hesperidin (320, 640 or 1280 µM in PBS) were added for 30 min. Thereafter, FITC-labeled P. gingivalis (OD660 = 0.5) were added to each well (100 μL), and the microplate was maintained at 37°C for 4 h. The number of bacteria adhered to the basement membrane model was determined as described above. Wells without P. gingivalis were used as controls to measure basal auto-fluorescence and wells without hesperidin were used to determine 100% adherence values. Adherence assays were carried out in triplicate in three independent experiments, and the means ± standard deviations were calculated.
4.6. Reactive oxygen species production by human oral epithelial cells
The B11 human oral epithelial cell line (3 x 105 cells/well) were seeded in black wall, clear flat bottom 96-well microplates (Greiner Bio-One North America, Monroe, NC, USA) and were incubated overnight at 37°C in a 5% CO2 atmosphere. A fluorometric assay to monitor the oxidation of 2’,7’-dichlorofluorescein-diacetate (DCF-DA; Sigma-Aldrich Canada Co., Oakville, ON, Canada) into a fluorescent compound was used to measure ROS production. A 40-mM stock solution of DCF-DA was freshly prepared in DMSO. The cells were washed with Hank’s balanced salt solution (HBSS; HyClone Laboratories, Logan, UT, USA) and were incubated for a further 30 min in the presence of 100 µM DCF-DA in HBSS. Keratinocytes were washed with HBSS to remove the excess of DCF-DA. Then, the cells were treated with P. gingivalis at an MOI of 100 either in the presence or absence of 1 mM hydrogen peroxide (H2O2) and in the absence or presence of hesperidin (320, 640 or 1280 µM) previously prepared in HBSS. ROS production by monitoring fluorescence emission was recorded every 20 min for 5 h using a Synergy 2 microplate reader (BioTek Instruments) (485 nm excitation filter and a 528 nm emission filter). The assays were carried out in triplicate in three independent experiments, and the means ± standard deviations were calculated.
4.7. Cytokine and matrix metalloproteinases (MMPs) secretion by macrophages
To evaluate the effect of hesperidin on the secretion of pro-inflammatory cytokines (IL-1β, IL-6, IL-8, TNF-α) and MMPs (MMP-2, MMP-8, MMP-9), U937 human monocytes (CRL-1593.2; American Type Culture Collection, Manassas, VA, USA) were used. Non-cytotoxic concentrations of hesperidin were first determined using a MTT (3-[4,5-diethylthiazol-2-yl]-2,5diphenyltetrazolium bromide) colorimetric assay (Roche Diagnostics, Laval, QC, Canada) according to the manufacturer’s instructions following a 24-h treatment. The monocytes (1 × 106 cells/mL) were differentiated into adherent macrophage-like cells by incubation in RPMI-10% FBS containing 100 ng/mL of phorbol myristic acid (PMA; Sigma-Aldrich Canada Co.) for 48 h into 12-well microplates. The adherent macrophage-like cells were washed to remove remaining PMA and non-adherent cells and were maintained in RPMI-10% FBS without PMA for 24 h. Thereafter, the medium was changed and the cells were maintained in RPMI-1% FBS and incubated overnight at 37°C in a 5% CO2 atmosphere. The macrophage-like cells were pre-treated for 2 h with hesperidin (20, 40, or 80 µM) previously prepared in RPMI-1% FBS and were stimulated with P. gingivalis cells at an MOI of 100 for 24 h. Then, the culture medium supernatants were collected and were kept at −20 °C until used. Culture medium collected from cells without treatment with hesperidin and bacteria were used as controls. Enzyme-linked immunosorbent assay (ELISA) kits (R&D Systems, Minneapolis, MN, USA; Invitrogen, Thermo Fisher Scientific Inc., Waltham, MA, USA) were used to determine cytokines and MMPs concentrations according to the manufacturers’ protocols. Assays were performed in triplicate in three independent experiments, and the means ± standard deviations were calculated.
4.8. Activation of NF-κB in P. gingivalis-stimulated monocytes
To evaluate the effect of hesperidin on P. gingivalis-induced NF-κB activation, the human monoblastic leukemia cell line U937 3xκ B-LUC was used. The monocytes were seeded (105 cells/well) into wells of black wall, black bottom, 96-well microplates (Greiner Bio-One North America) and were pre-incubated with hesperidin (20, 40, or 80 µM) for 30 min. Thereafter, the monocytes were stimulated with P. gingivalis at an MOI of 100 for 6 h. Wells containing monocytes without P. gingivalis or hesperidin were used as controls. Bright-Glo reagent (Promega Corporation, Durham, NC, USA) was used according to the manufacturer’s protocol to measure luciferase activity and determine NF-κB activation. Luminescence was monitored using a Synergy 2 microplate reader (BioTek Instruments). Assays were performed in triplicate in three independent experiments, and the means ± standard deviations were calculated.
4.9. Statistical analysis
A one-way ANOVA with a post hoc Bonferroni multiple comparison test was used to analyze the data. Results were considered statistically significant at p < 0.05, p < 0.01 or p < 0.001.
5. Conclusions
In summary, our results showed that the flavonoid hesperidin attenuates the gingival epithelial barrier dysfunction caused by P. gingivalis, and exerts a protective effect against P. gingivalis adhesion to extracellular matrix proteins. Furthermore, hesperidin provides oxidative stress protection by reducing ROS production by P. gingivalis-stimulated oral epithelial cells. This flavonoid attenuated the secretion of pro-inflammatory cytokines and NF-κB activation in macrophages stimulated with P. gingivalis. Within the limitations of this study, our promising results suggest that hesperidin may be an excellent candidate for adjuvant therapy for the treatment or prevention of periodontal disease. However, future studies are necessary to assess the exact mechanisms of action of hesperidin on epithelial barrier and inflammatory response during periodontal disease.
Author Contributions
Conceptualization, D.G and P.M.M.H.; methodology, P.M.M.H.; software, P.M.M.H.; validation, P.M.M.H.; formal analysis, P.M.M.H.; investigation, P.M.M.H.; resources, D.G.; data curation, P.M.M.H.; writing—original draft preparation, P.M.M.H.; writing—review and editing, P.M.M.H., D.M.P.S., J.M. and D.G.; visualization, P.M.M.H.; supervision, D.G.; project administration, D.G.; funding acquisition, D.G. All authors have read and agreed to the published version of the manuscript.
Funding
Maquera-Huacho: P.M. was supported by a scholarship provided by Sao Paulo Research Foundation (FAPESP; Grant #2018/16540-8 and #2019/15343-7). This research was funded by the Laboratoire de Contrôle microbiologique of Université Laval (2021-12).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We wish to thank S. Groeger and J. Meyle (Justus-Liebig-University Giessen, Germany), for providing the B11 keratinocyte cell line, and R. Blomhoff and H. Carlsen (University of Oslo, Norway) for their generous gift of the U937-3xκB-LUC cell line.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, Y.; Wang, X.; Li, H.; Ni, C.; Du, Z.; Yan, F. Human oral microbiota and its modulation for oral health. Biomed Pharmacother. 2018, 99, 883–893. [Google Scholar] [CrossRef] [PubMed]
- Roberts, F.A.; Darveau, R.P. Microbial protection and virulence in periodontal tissue as a function of polymicrobial communities: symbiosis and dysbiosis. Periodontol 2000. 2015, 69, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Ebersole, J.L.; Dawson, D.R. 3rd; Morford, L.A.; Peyyala, R.; Miller, C.S.; Gonzaléz, O.A. Periodontal disease immunology: 'double indemnity' in protecting the host. Periodontol 2000. 2013, 62, 163–202. [Google Scholar] [CrossRef] [PubMed]
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: dynamic communities and host interactions. Nat Rev Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cai, W.; Zhao, S.; Shi, L.; Chen, Y.; Li, X.; Sun, X.; Mao, Y.; He, B.; Hou, Y.; et al. Oxidative stress-related biomarkers in saliva and gingival crevicular fluid associated with chronic periodontitis: A systematic review and meta-analysis. J Clin Periodontol. 2019, 46, 608–622. [Google Scholar] [CrossRef] [PubMed]
- Socransky, S.S.; Haffajee, A.D.; Cugini, M.A.; Smith, C.; Kent, R.L. Jr. Microbial complexes in subgingival plaque. J Clin Periodontol. 1998, 25, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, D.; Liu, S.; Zhang, S.; Pan, Y. The Role of Porphyromonas gingivalis Outer Membrane Vesicles in Periodontal Disease and Related Systemic Diseases. Front Cell Infect Microbiol. 2021, 28, 10:585917. [CrossRef]
- Fujita, T.; Yoshimoto, T.; Kajiya, M.; Ouhara, K.; Matsuda, S.; Takemura, T.; Akutagawa, K.; Takeda, K.; Mizuno, N.; Kurihara, H. Regulation of defensive function on gingival epithelial cells can prevent periodontal disease. Jpn Dent Sci Rev. 2018, 54, 66–75. [Google Scholar] [CrossRef]
- Ji, S.; Choi, Y.S.; Choi, Y. Bacterial invasion and persistence: critical events in the pathogenesis of periodontitis? J Periodontal Res. 2015, 50, 570–585. [Google Scholar] [CrossRef]
- Holt, S.C.; Ebersole, J.L. Porphyromonas gingivalis, Treponema denticola, and Tannerella forsythia: the "red complex", a prototype polybacterial pathogenic consortium in periodontitis. Periodontol 2000. 2005, 38, 72–122. [Google Scholar] [CrossRef]
- Groeger, S.E.; Meyle, J. Epithelial barrier and oral bacterial infection. Periodontol 2000. 2015, 69, 46–67. [Google Scholar] [CrossRef]
- Grenier, D.; La, V.D. Proteases of Porphyromonas gingivalis as important virulence factors in periodontal disease and potential targets for plant-derived compounds: a review article. Curr Drug Targets. 2011, 12, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Amano, A. Disruption of epithelial barrier and impairment of cellular function by Porphyromonas gingivalis. Front Biosci. 2007, 12, 3965–3974. [Google Scholar] [CrossRef] [PubMed]
- Miles, E.A; Calder, P.C. Effects of citrus fruit juices and their bioactive components on inflammation and immunity: A narrative review. Front Immunol. 2021, 12:712608. [CrossRef]
- Li, C.; Schluesener, H. Health-promoting effects of the citrus flavanone hesperidin. Crit Rev Food Sci Nutr. 2017, 57, 613–631. [Google Scholar] [CrossRef] [PubMed]
- Kuo, P.J.; Fu, E.; Lin, C.Y.; Ku, C.T. Chiang, C.Y.; Fu, M.M.; Fu, M.W.; Tu, H.P.; Chiu, H.C. Ameliorative effect of hesperidin on ligation-induced periodontitis in rats. J Periodontol. 2019, 90, 271-280.
- Gonçalves, V.P.; Musskopf, M.L.; Rivera-Concepcion, A.; Yu, C.; Wong, S.W.; Tuin, S.A.; Jiao, Y.; Susin, C.; Spolidorio, L.C.; Miguez, P.A. Systemic dietary hesperidin modulation of osteoclastogenesis, bone homeostasis and periodontal disease in mice. Int J Mol Sci. 2022, 13, 7100. [Google Scholar] [CrossRef] [PubMed]
- Canakçi, C.F.; Ciçek, Y.; Canakçi, V. Reactive oxygen species and human inflammatory periodontal diseases. Biochemistry (Mosc). 2005, 70, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Groeger, S.; Doman, E.; Chakraborty, T.; Meyle, J. Effects of Porphyromonas gingivalis infection on human gingival epithelial barrier function in vitro. Eur J Oral Sci. 2010, 118, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, R.J. Bacterial adhesion to oral tissues: a model for infectious diseases. J Dent Res. 1989, 68, 750–760. [Google Scholar] [CrossRef]
- Guo, K.; Ren. J.; Gu, G.; Wang, G.; Gong, W.; Wu, X.; Ren, H.; Hong, Z.; Li, J. Hesperidin protects against intestinal inflammation by restoring intestinal barrier function and up-regulating Treg cells. Mol Nutr Food Res. 2019, 63, e1800975. [CrossRef]
- Bostanci, N.; Belibasakis, G.N. Porphyromonas gingivalis: an invasive and evasive opportunistic oral pathogen. FEMS Microbiol Lett. 2012, 333, 1–9. [Google Scholar] [CrossRef]
- Amano, A. Disruption of epithelial barrier and impairment of cellular function by Porphyromonas gingivalis. Front Biosci. 2007, 12, 3965–3974. [Google Scholar] [CrossRef]
- Groeger, S.; Meyle, J. Oral mucosal epithelial cells. Front Immunol. 2019, 10, 208. [Google Scholar] [CrossRef]
- Andrian, E.; Grenier, D.; Rouabhia, M. In vitro models of tissue penetration and destruction by Porphyromonas gingivalis. Infect Immun. 2004, 72, 4689–4698. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Mo, L.; Niu, Y.; Li, X.; Zhou, X.; Xu, X. The role of reactive oxygen species and autophagy in periodontitis and their potential linkage. Front Physiol. 2017, 8, 439. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T. Signal transduction by reactive oxygen species. J Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Kalpana, K.B.; Srinivasan, M.; Menon, V.P. Evaluation of antioxidant activity of hesperidin and its protective effect on H2O2 induced oxidative damage on pBR322 DNA and RBC cellular membrane. Mol Cell Biochem. 2009, 323, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Gu, H.; Ye, Y.; Lin, B.; Sun, L.; Deng, W.; Zhang, J.; Liu, J. Protective effects of hesperidin against oxidative stress of tert-butyl hydroperoxide in human hepatocytes. Food Chem Toxicol. 2010, 48, 2980–2987. [Google Scholar] [CrossRef] [PubMed]
- de Souza, A.B.F.; de Matos, N.A.; Castro, T.F.; Costa, G.P.; Oliveira, L.A.M.; Nogueira, K.O.P.C.; Ribeiro, I.M.L.; Talvani, A.; Cangussú, S.D.; de Menezes, R.C.A.; et al. Effects in vitro and in vivo of hesperidin administration in an experimental model of acute lung inflammation. Free Radic Biol Med. 2022, 180, 253–262. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, el-S.M.; Abo-Salem, O.M.; Abd-Ellah, M.F.; Abd-Alla, G.M. Hesperidin, an antioxidant flavonoid, prevents acrylonitrile-induced oxidative stress in rat brain. J Biochem Mol Toxicol. 2008, 22, 268-273.
- Sahu, B.D.; Kuncha, M.; Sindhura, G.J.; Sistla, R. Hesperidin attenuates cisplatin-induced acute renal injury by decreasing oxidative stress, inflammation and DNA damage. Phytomedicine. 2013, 20, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Simitzis, P.E.; Ilias-Dimopoulos, V.; Charismiadou, M.A.; Biniari, E.E.; Deligeorgis, S.G. Effects of dietary hesperidin supplementation on lamb performance and meat characteristics. Anim Sci J. 2013, 84, 136–143. [Google Scholar] [CrossRef]
- Lim, C.; Zhen, A.X.; Ok, S.; Fernando, P.D.S.M.; Herath, H.M.U.L.; Piao, M.J.; Kang, K.A.; Hyun, J.W. Hesperidin protects SH-SY5Y neuronal cells against high glucose-induced apoptosis via regulation of MAPK signaling. Antioxidants (Basel). 2022, 11, 1707. [Google Scholar] [CrossRef]
- Smalley, J.W.; Birss, A.J.; Silver, J. The periodontal pathogen Porphyromonas gingivalis harnesses the chemistry of the mu-oxo bishaem of iron protoporphyrin IX to protect against hydrogen peroxide. FEMS Microbiol Lett. 2000, 183, 159–164. [Google Scholar]
- Choi, I.Y.; Kim, S.J.; Jeong, H.J.; Park, S.H.; Song, Y.S.; Lee, J.H.; Kang, T.H.; Park, J.H.; Hwang, G.S.; Lee, E.J.; et al. Hesperidin inhibits expression of hypoxia inducible factor-1 alpha and inflammatory cytokine production from mast cells. Mol Cell Biochem. 2007, 305, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Nizamutdinova, I.T.; Jeong, J.J.; Xu, G.H.; Lee, S.H.; Kang, S.S.; Kim, Y.S.; Chang, K.C.; Kim, H.J. Hesperidin, hesperidin methyl chalone and phellopterin from Poncirus trifoliata (Rutaceae) differentially regulate the expression of adhesion molecules in tumor necrosis factor-alpha-stimulated human umbilical vein endothelial cells. Int Immunopharmacol. 2008, 8, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, J.; Cai, L.; Hu, C.M.; Zhang, L. Suppression of adjuvant arthritis by hesperidin in rats and its mechanisms. J Pharm Pharmacol. 2008, 60, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Tervahartiala, T.; Pirilä, E.; Ceponis, A.; Maisi, P.; Salo, T.; Tuter, G.; Kallio, P.; Törnwall, J.; Srinivas, R.; Konttinen, Y.T.; et al. The in vivo expression of the collagenolytic matrix metalloproteinases (MMP-2, -8, -13, and -14) and matrilysin (MMP-7) in adult and localized juvenile periodontitis. J Dent Res. 2000, 79, 1969–1977. [Google Scholar] [CrossRef] [PubMed]
- Uitto, V.J.; Overall, C.M.; McCulloch, C. Proteolytic host cell enzymes in gingival crevice fluid. Periodontol 2000. 2003, 31, 77–104. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, A.; Menon, V.P. Effect of hesperidin on matrix metalloproteinases and antioxidant status during nicotine-induced toxicity. Toxicology. 2007, 238, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Kamaraj, S.; Anandakumar, P.; Jagan, S.; Ramakrishnan, G.; Devaki, T. Modulatory effect of hesperidin on benzo(a)pyrene induced experimental lung carcinogenesis with reference to COX-2, MMP-2 and MMP-9. Eur J Pharmacol. 2010, 649, 320–327. [Google Scholar] [CrossRef]
- Qin, Z.; Chen, L.; Liu, M.; Tan, H.; Zheng, L. Hesperidin reduces adverse symptomatic intracerebral hemorrhage by promoting TGF-β1 for treating ischemic stroke using tissue plasminogen activator. Neurol Sci. 2020, 41, 139–147. [Google Scholar] [CrossRef]
- Milward, M.R.; Chapple, I.L.; Wright, H.J.; Millard, J.L.; Matthews, J.B.; Cooper, P.R. Differential activation of NF-kappaB and gene expression in oral epithelial cells by periodontal pathogens. Clin Exp Immunol. 2007, 148, 307–324. [Google Scholar] [CrossRef]
- Sato, K.; Sato, T.; Ohno-Oishi, M.; Ozawa, M.; Maekawa, S.; Shiga, Y.; Yabana, T.; Yasuda, M.; Himori, N.; Omodaka, K.; et al. CHOP deletion and anti-neuroinflammation treatment with hesperidin synergistically attenuate NMDA retinal injury in mice. Exp Eye Res. 2021, 213, 108826. [Google Scholar] [CrossRef]
- Gröger, S.; Michel, J.; Meyle, J. Establishment and characterization of immortalized human gingival keratinocyte cell lines. J Periodontal Res. 2008, 43, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Carlsen, H.; Moskaug, J.Ø.; Fromm, S.H.; Blomhoff, R. In vivo imaging of NF-kappa B activity. J Immunol. 2002, 168, 1441–1446. [Google Scholar] [CrossRef] [PubMed]
- Ben Lagha, A.; Maquera Huacho, P.; Grenier, D. A cocoa (Theobroma cacao L.) extract impairs the growth, virulence properties, and inflammatory potential of Fusobacterium nucleatum and improves oral epithelial barrier function. PLoS One. 2021, 16, e0252029. [CrossRef]
Figure 1.
Viability of human oral epithelial cell line B11 after treatment (48 h) with hesperidin. *Significant difference in comparison with the control group (p < 0.05).
Figure 1.
Viability of human oral epithelial cell line B11 after treatment (48 h) with hesperidin. *Significant difference in comparison with the control group (p < 0.05).
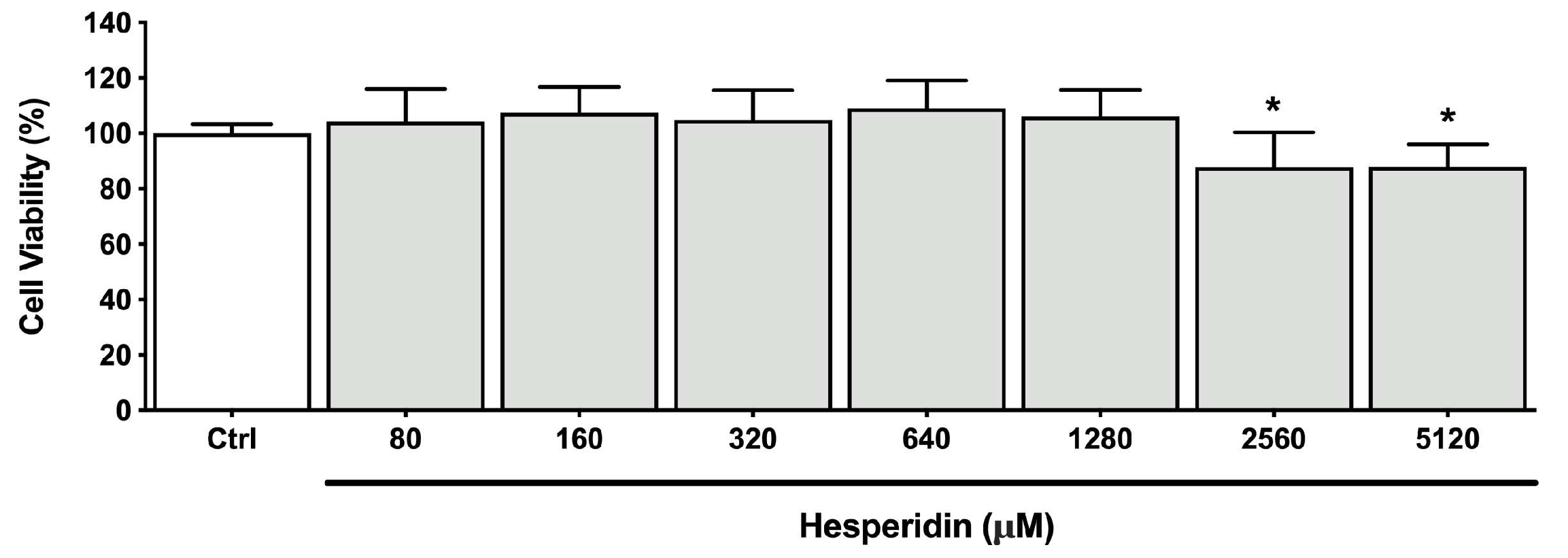
Figure 2.
Time- and dose-dependent effects of hesperidin on oral epithelial barrier model (A) and challenged with P. gingivalis (B) as determined by monitoring TER. A 100% value was assigned to the TER values recorded at time 0. Results are expressed as the means ± SD of triplicate assays. *, significant increase (p < 0.05) compared to control cells (A) or P. gingivalis-stimulated cells (B). §, significant decrease (p < 0.05) compared to non-stimulated control cells (B).
Figure 2.
Time- and dose-dependent effects of hesperidin on oral epithelial barrier model (A) and challenged with P. gingivalis (B) as determined by monitoring TER. A 100% value was assigned to the TER values recorded at time 0. Results are expressed as the means ± SD of triplicate assays. *, significant increase (p < 0.05) compared to control cells (A) or P. gingivalis-stimulated cells (B). §, significant decrease (p < 0.05) compared to non-stimulated control cells (B).
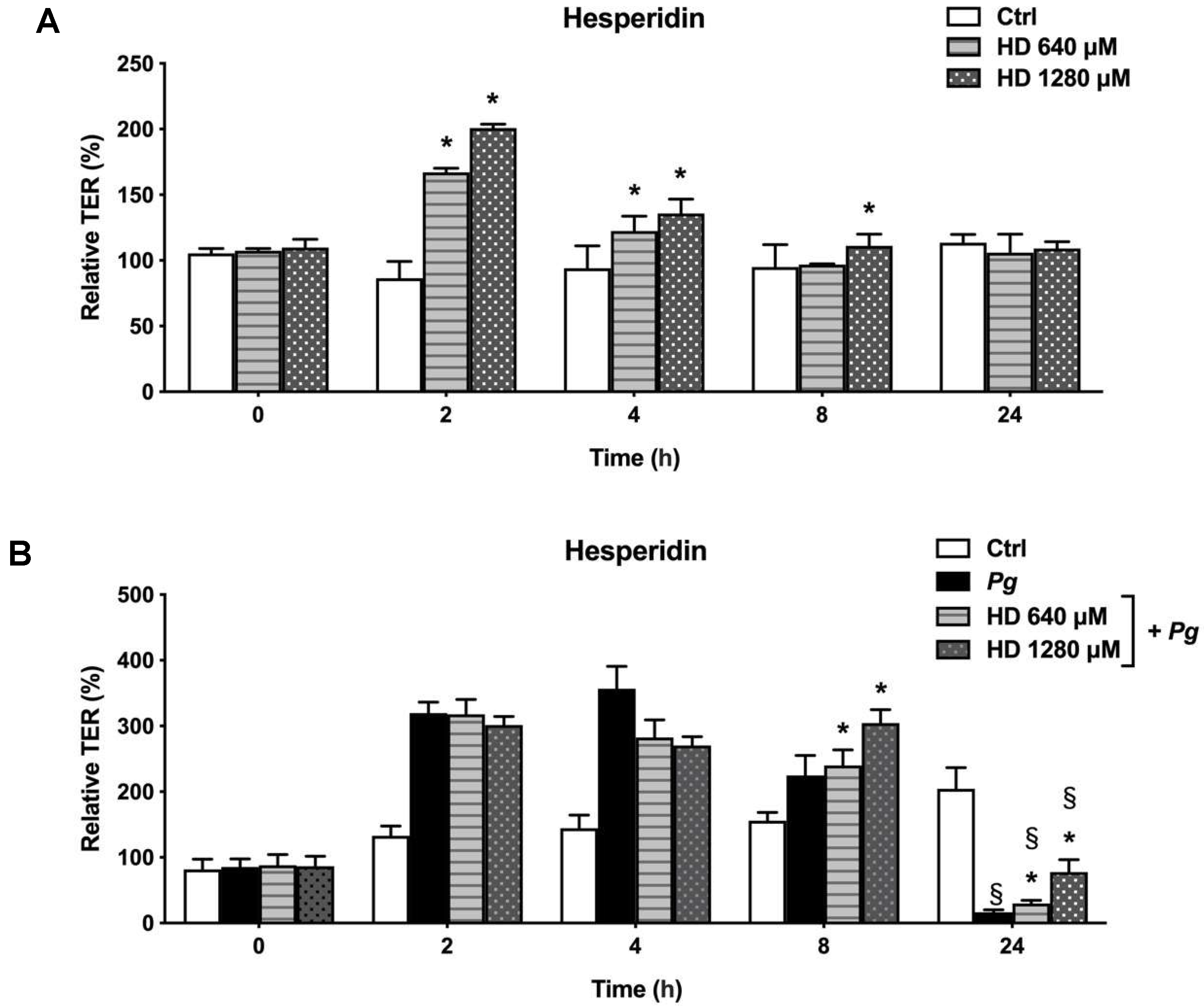
Figure 3.
Effect of hesperidin on the adherence of P. gingivalis to human oral epithelial cells (A) and Cultrex® Basement Membrane Extract (B). Results are expressed as the means ± SD to triplicate assays. *, significantly different from the control (p < 0.05).
Figure 3.
Effect of hesperidin on the adherence of P. gingivalis to human oral epithelial cells (A) and Cultrex® Basement Membrane Extract (B). Results are expressed as the means ± SD to triplicate assays. *, significantly different from the control (p < 0.05).
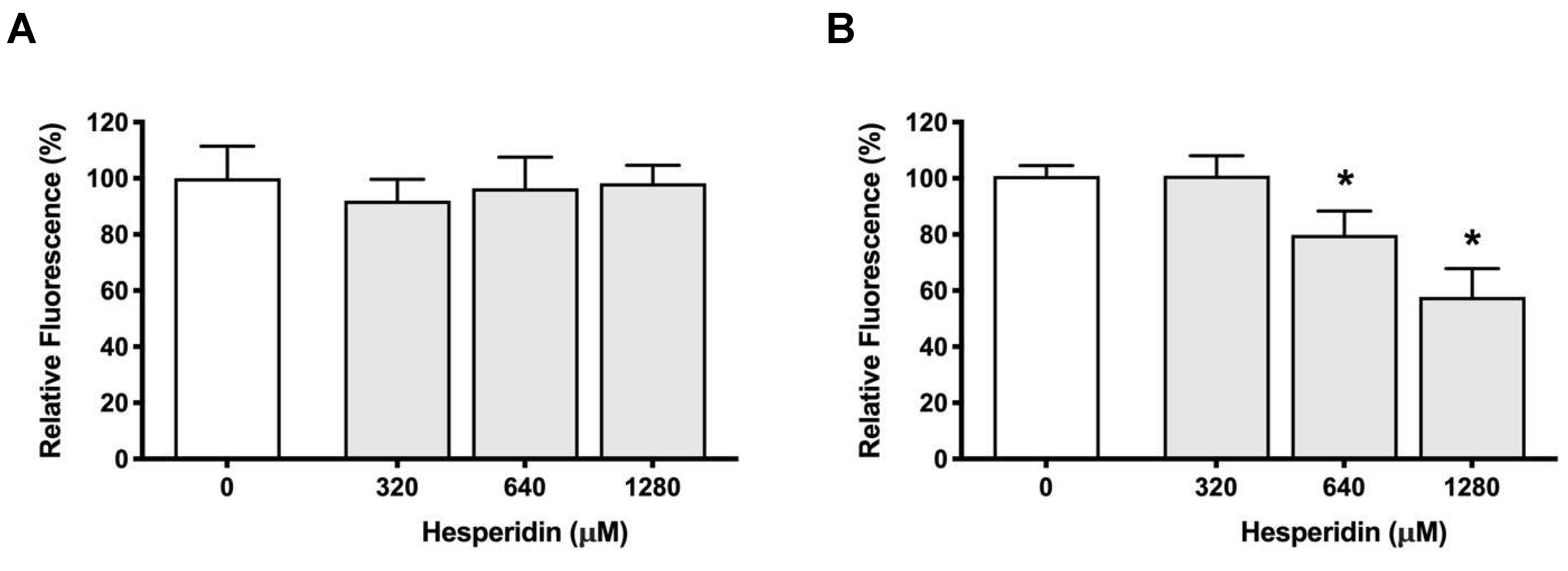
Figure 4.
Time- and dose-dependent effects of hesperidin on ROS production by P. gingivalis-stimulated epithelial cells. Results are expressed as the means ± SD of three independent experiments. Values are significantly different from P. gingivalis-stimulated epithelial cells (p < 0.05).
Figure 4.
Time- and dose-dependent effects of hesperidin on ROS production by P. gingivalis-stimulated epithelial cells. Results are expressed as the means ± SD of three independent experiments. Values are significantly different from P. gingivalis-stimulated epithelial cells (p < 0.05).
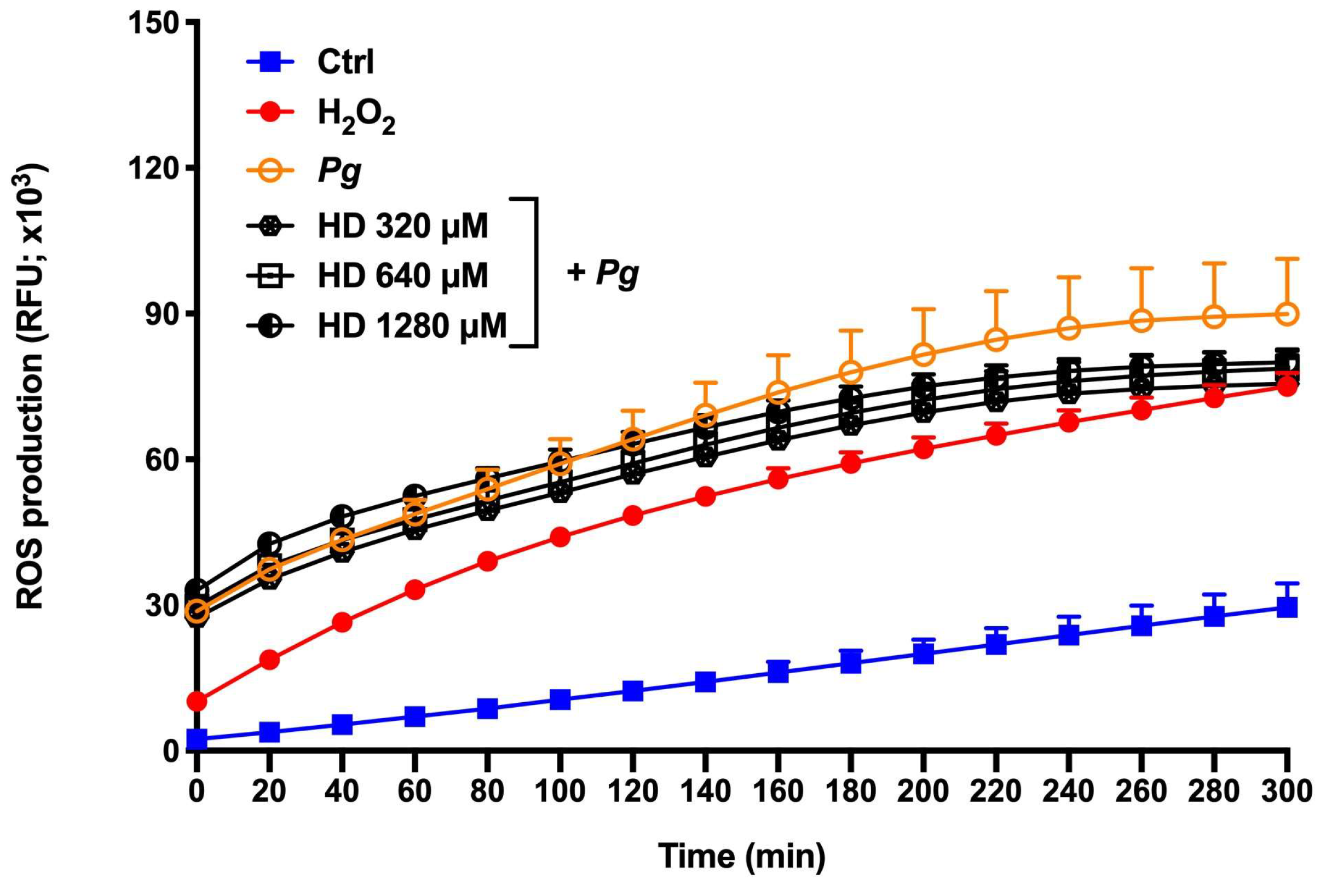
Figure 5.
Time- and dose-dependent effects of hesperidin on ROS production by human epithelial cells stimulated synergistically with hydrogen peroxide (H2O2) and P. gingivalis. Results are expressed as the means ± SD of three independent experiments. Values are significantly different from H2O2 and P. gingivalis-stimulated epithelial cells (p < 0.05).
Figure 5.
Time- and dose-dependent effects of hesperidin on ROS production by human epithelial cells stimulated synergistically with hydrogen peroxide (H2O2) and P. gingivalis. Results are expressed as the means ± SD of three independent experiments. Values are significantly different from H2O2 and P. gingivalis-stimulated epithelial cells (p < 0.05).

Figure 6.
Cell viability of macrophage-like cells after treatment (24 h) with hesperidin. *Significant difference in comparison with the control group (p < 0.05).
Figure 6.
Cell viability of macrophage-like cells after treatment (24 h) with hesperidin. *Significant difference in comparison with the control group (p < 0.05).
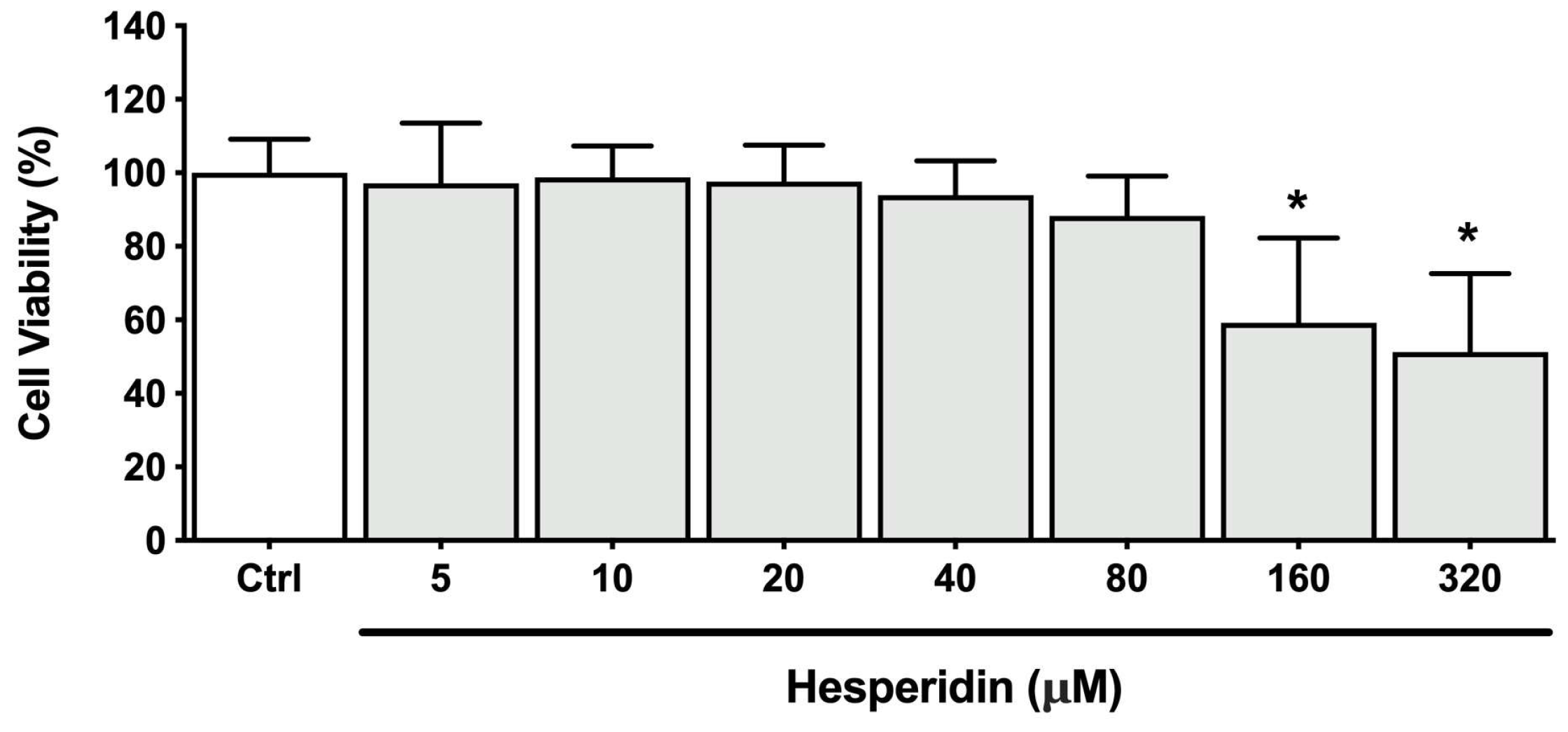
Figure 7.
Secretion of IL-6 (A), IL-8 (B), TNF-α (C), and IL-1β (D) by macrophage-like cells treated with hesperidin and stimulated with P. gingivalis (MOI = 100) for 24 h. Results are expressed as the means ± SD of triplicate assays for three independent experiments. #, significant increase (p < 0.001) compared to cells not stimulated with P. gingivalis. *, significant decrease (p < 0.001) compared to P. gingivalis-stimulated cells.
Figure 7.
Secretion of IL-6 (A), IL-8 (B), TNF-α (C), and IL-1β (D) by macrophage-like cells treated with hesperidin and stimulated with P. gingivalis (MOI = 100) for 24 h. Results are expressed as the means ± SD of triplicate assays for three independent experiments. #, significant increase (p < 0.001) compared to cells not stimulated with P. gingivalis. *, significant decrease (p < 0.001) compared to P. gingivalis-stimulated cells.

Figure 8.
Secretion of MMP-2 (A), MMP-8 (B), and MMP-9 (C) by macrophage-like cells treated with hesperidin and stimulated with P. gingivalis (MOI = 100) for 24 h. Results are expressed as the means ± SD of triplicate assays for three independent experiments. #, significant increase (p < 0.001) compared to cells not stimulated with P. gingivalis. *, significant decrease (p < 0.001) compared to P. gingivalis-stimulated cells.
Figure 8.
Secretion of MMP-2 (A), MMP-8 (B), and MMP-9 (C) by macrophage-like cells treated with hesperidin and stimulated with P. gingivalis (MOI = 100) for 24 h. Results are expressed as the means ± SD of triplicate assays for three independent experiments. #, significant increase (p < 0.001) compared to cells not stimulated with P. gingivalis. *, significant decrease (p < 0.001) compared to P. gingivalis-stimulated cells.
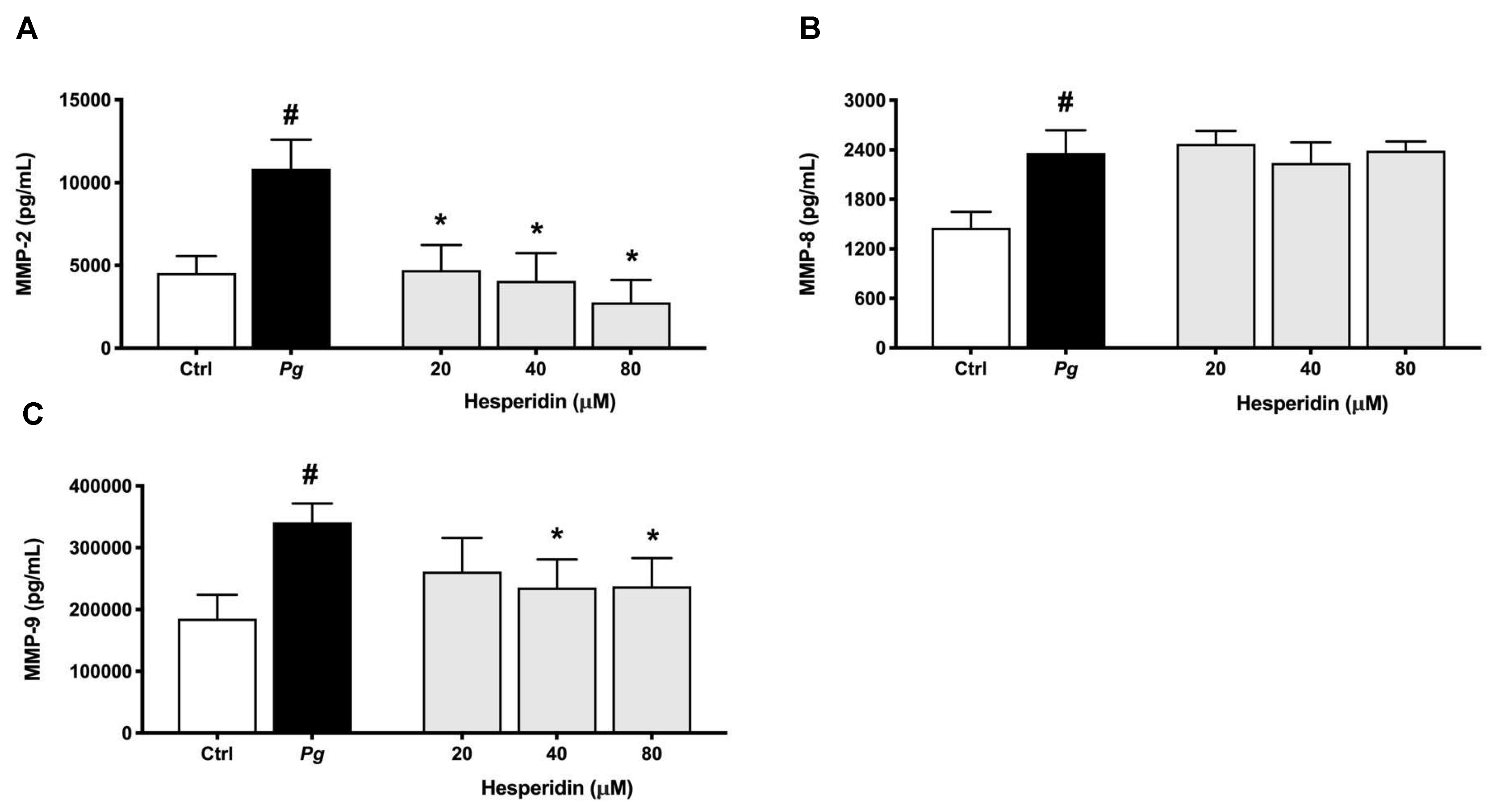
Figure 9.
NF-κB activation in U937-3xκB monocytic cell line induced by P. gingivalis (MOI = 100). A value of 100% was assigned to the activation obtained with P. gingivalis in the absence of hesperidin. Results are expressed as the means ± SD of triplicate assays for three independent experiments. #, significant increase (p < 0.01) compared to cells not stimulated with P. gingivalis. *, significant decrease (p < 0.01) compared to P. gingivalis-stimulated cells.
Figure 9.
NF-κB activation in U937-3xκB monocytic cell line induced by P. gingivalis (MOI = 100). A value of 100% was assigned to the activation obtained with P. gingivalis in the absence of hesperidin. Results are expressed as the means ± SD of triplicate assays for three independent experiments. #, significant increase (p < 0.01) compared to cells not stimulated with P. gingivalis. *, significant decrease (p < 0.01) compared to P. gingivalis-stimulated cells.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



