Submitted:
11 May 2023
Posted:
12 May 2023
Read the latest preprint version here
Preprints on COVID-19 and SARS-CoV-2
Abstract
The novel coronavirus disease 2019 (COVID-19) known as severe acute respiratory syndrome - coronavirus 2 (SARS-CoV-2) has emerged in China in 2019, and caused an outbreak of unusual viral pneumonia. The olfactory dysfunction following the infection of different variants of SARS-CoV-2 is now accepted as a hallmark symptom in patients. Recent studies have pointed out the relationship between COVID-19 and altered or loss of smell in infected patients. This mini review provides an overview of the role of SARS-CoV-2 and the other acute respiratory viruses in the development the human olfactory pathophysiology. We highlight the importance of deciphering the molecular mechanisms underlying the olfactory dysfunction caused by SARS-CoV-2 to help design new drugs to restore the altered or loss of smell in affected patients.
Keywords:
Respiratory viruses
; Anosmia
; Olfaction Disorders
; loss of smell
; COVID-19
1. Introduction
In December 2019, a novel coronavirus designated as SARS-CoV-2 (severe acute respiratory syndrome - coronavirus 2) emerged in the city of Wuhan, China, and caused an outbreak of unusual viral pneumonia. Being highly transmissible, this novel coronavirus disease, also known as coronavirus disease 2019 (COVID-19), has spread fast all over the world [1,2]. The new emerging virus has overwhelmingly surpassed SARS and MERS in terms of both the number of infected people and the spatial range of epidemic areas [3]. On 11 March 2020, the WHO officially characterized the global COVID-19 outbreak as a pandemic [4]. As of 12 April 2023, 763 million confirmed cases and 6,8 million deaths have been reported globally [5]. SARS-CoV-2 is transmitted to humans through buccal and nasal cavities mainly through respiratory droplets from sneezes or coughs of infected people and spread to those nearby [6]. A key factor in the transmissibility of COVID-19 is the active virus replication in upper respiratory tract tissues and therefore its massive excretion [7]. Regarding symptoms reported in patients with COVID-19 infection the most common are fever, dry cough, shortness of breath (dyspnoea), myalgia, malaise, chills, confusion, headache, sore throat, rhinorrhea, chest pain, diarrhea, conjunctival congestion, nasal congestion, sputum production, hemoptysis, runny nose, fatigue and sneezing [8,9,10,11]. Several studies have reported olfactory dysfunction and hypogeusia in COVID-19 patients [12,13]. Interestingly, a clustering clinical data have shown that neurological symptoms like persistent headache was noticed in 50% of patients who suffered from olfactory dysfunction after months of recovery from SARS-CoV-2 infection suggesting in part a strongly link between the pathophysiological substrate of both cognition and olfaction related disorders [14]. A recent study has documented the effect of the use of personal protective equipment, especially by healthcare workers in preventing the spread of COVID-19. Their findings demonstrated that wearing a face mask significantly improves the daily health issues related to the disease symptoms and the working performance of those workers who have adhered to safety guideline rules [15]. Humans have developed five senses, traditionally vision, hearing, smell, taste and touch, to gather information from their surroundings and to benefit from where they live. Even though several factors, such as ageing, drug, and respiratory pathogens, are shown to lead to neurological invasiveness, different variants of SARS-CoV-2 have been considered a leading cause of anosmia in humans [14,16,17,18] . The respiratory coronavirus transmission throughout the nasal cavity and its consequence in impairing odorant detection have gained attention recently [19,20,21]. The first step of molecular detection occurs when a chemical odorant is recognized by one or several olfactory receptors (ORs) in the olfactory sensory neurons (OSN). The process of this odour recognition takes place in the nasal cavity before reaching the upper part of the human brain [22,23,24]. The importance of sensing and tasting is often shed in light only when a subject is facing difficulties in discriminating odours via food intake or the environment [25]. As smell is somehow not vital daily for humans and its loss may not be directly life-threatening, the loss of functional olfactory responses, in the long term, could be accounted for the poor quality of life, at worst, lead to death [14,26]. Since the beginning of this pandemic, a lot of reports have shown the issues of olfactory disorders in patients affected by COVID-19 [16,17,18,19]. This review mainly focuses on the loss of smell and particularly on understanding the molecular signaling underlying the olfactory pathophysiology in human patients from COVID-19 infection and the importance of using such mechanisms to find potential targets to overcome the loss of smell.
2. Methods
The present study synthesizes the current knowledge regarding the relationship of the respiratory viral pathogenesis of the olfactory system and the mechanisms underlying the loss of smell in patients infected by respiratory viruses in general and COVID-19 in particular in the last three years. A deep search has been performed using mostly the database PubMed, PMC and ScienceDirect to parse both original and review articles that have tackled the animals and human respiratory viruses having a negative impact on the olfactory system functionality. For our search, we used the combination of the following keywords: respiratory virus, coronavirus, rhinovirus, influenza viruses, parainfluenza viruses, respiratory syncytial virus (RSV), coxsackievirus, adenoviruses, poliovirus, enteroviruses (EVs), herpesviruses, anosmia, olfactory epithelium, human, mouse, hamster, loss of smell or olfactory dysfunction were considered in this review. Only the papers that have met the keyword criteria listed above were considered in this work.
3. Olfactory Receptor and Odorant Detection
Odorant perception is fundamental for survival in most animals. In humans, the sense of smell is important to appreciate our environment. Therefore, a functional olfactory system detects and discriminates among diverse chemical stimuli. Odours are important for behaviours such as feeding, mating, and avoiding dangerous smells, such as smoke, leaking propane gas, and spoiled food [25,27,28]. Processes of learning and memory are associated with these behaviours. This leads to a strong belief that the loss of olfactory function is indirectly related to human life-threatening [29,30] . The sense of smell, which is fundamental for their survival, is developed in most animals. Mostly, two different olfactory systems have been developed in mammals such as rodents: the main olfactory epithelium (MOE or OE), also called olfactory mucosa, connected to the main olfactory bulb , and the accessory system called vomeronasal organ (VNO) connected to the accessory olfactory bulb [22,31,32,33,34]. Here, the VNO will not be discussed. The configuration of the olfactory epithelium (OE) presents unique cytological characteristics as it contains different cell types such as the ciliated olfactory receptor neurons (ORNs), the sustentacular supporting cells, and the cells of Bowman’s glands (Figure 1). The ORNs reside in the intermediate layer of the olfactory mucosa while the sustentacular cells and sensory cilia are located in the apical layer where the dendrites of olfactory neurons are extended [35,36].
A deep understanding of the molecular signaling of the smelling recognition is required to understand the basis of the olfactory system and, consequently, the loss of olfactory function. In the last decade, pioneers have developed and studied the physiology of the olfactory system based on molecular biology, biochemistry, anatomy, and bioinformatics [23,25]. At first glance, getting insight into the molecular mechanisms of the perception of odours has emerged from several disciplines such as chemistry, biology and professional odour detectors [23,25].
In the human genome, around 857 olfactory receptors (OR) genes were identified, but 60% of them are pseudogenes. This high prevalence may be due to internal deletions during evolution, leaving around 390 putative functional ORs while around 465 are pseudogenes [23,37,38,39]. Although 1300 OR genes have been identified in the mouse genome, only 20% of these genes are pseudogenes [40,41]. This difference between humans and mice in the amount and percentage of functional OR genes may originate from the fact that olfaction is more solicited in rodents than in higher primates during evolution. The ORs of mammals are part of the superfamily of GPCRs [42]. They are considered one of the most critical groups in number and are known as rhodopsin-like owing to their structural similarity to rhodopsin. GPCRs in family A share many features, such as an Asp-Arg-Try sequence (DRY motif) in the second intracellular loop and a disulfide bridge from the two cysteines in the first (EC1) and second (EC2) extracellular [42]. In fact, olfaction is the perception of a combination of a myriad of odorant molecules. Odorants are generally known as small components (less than 400 Da) and are mostly hydrophobic volatile molecules. They are commonly aliphatic or aromatic molecules, with various functional groups including aldehydes, alcohols, carboxylic acids, ketones and esters [43]. Odorants can be cyclic, linear, or branched. The detection occurs when the odorants penetrate into the nasal cavity and reach the olfactory mucosa. The odorants then interact with specific OR in the olfactory mucosa. Once an OR is activated by an odorant, a nervous influx is sent to the cortex via the olfactory bulb. In addition, while acting as agonists for certain ORs, odorant stimuli can act as antagonists for other ORs [44,45], by preventing the activation of the receptors. This could be a plausible argument to explain why certain odours can be masked within some mixtures of odorants [46]. Surprisingly, the perception of an odour can also depend on its concentration. The olfactory response is efficient when odorant concentration is at a moderate level. Instead of activating, the high concentration of odorant inhibits the OR functionality. The OR functional response adaptation strongly depends on the concentration of odorant and has been a subject of debate in the sensing field [47,48,49,50]. Readers interested in the mammalian olfactory epithelium and the perception of odour coding are invited to view an excellent review by Kurian and colleagues published in 2020 [51].
4. Viral infection Causing Loss of Smell
The fact that the olfactory receptor neurons (ORNs) are found in the nasal cavity and expressed in the olfactory epithelium (OE) makes them directly exposed to all kinds of air-bound and air-way pathogens that make the ORNs vulnerable. Whether the cause is physiological or pathological, the lifespan of ORNs is relatively short of few weeks in the OE. Moreover, the stem cell reprogramming ensures the continuous regeneration of new ORNs from OE basal cells in response to inflammation and OE severe damage mediated by neural injury [52,53,54]. Several airway pathogens, such as viruses, are causing damage to OE through particularly the sustentacular cells, triggering anosmia in mammals [55,56] [36,57,58]. Many respiratory tract infections due to viruses like rhinovirus, influenza viruses, parainfluenza viruses, respiratory syncytial virus (RSV), coxsackievirus, adenoviruses, poliovirus, enteroviruses (EVs), and herpesviruses, have been involved in the development of olfactory disorders such as partial or total loss of smell. Doty and others have termed this pathology as virus-induced olfactory dysfunction as post-viral olfactory disorder (PVOD) [59,60,61,62]. Chronic viral infection destroys the many cells within the apical layer of the OE that could lead to ORNs functional impairment in the nose. Interestingly, the OE basal cells can constantly replace damaged ORNs to new olfactory neurons allowing patients to recover functional olfactory response [53,54]. In the following sections, the most common viral infection of the upper respiratory tract leading to olfactory dysfunction in animal models and humans will be discussed.
5. Viruses Impacting Respiratory System
The respiratory system is exposed to the environment and is in permanent contact with air-way pathogens like viruses. Cytomegalovirus (CMV) belonging to the Herpesviridae family and, also called Human Herpes Virus 5 (HHV-5), is shown to impact mice’s olfactory system. The mouse neonatal exhibited an impaired olfactory function following a placental CMV infection. The virus preferentially impacts the sensing system before it deteriorates the hearing system in mice [63]. Moreover, the infection of the hippocampus and the olfactory bulb by CMV triggers a sensorineural handicap that can induce brain malformations at the late stages of gestation [64]. A recent investigation in humans, has identified 18 viruses in patients with post-viral olfactory dysfunction. Particularly, rhinovirus (RV) and coronavirus (CoV OC43) were more predominant suggesting that they are major causative agents of PVOD [65]. The Sendai virus (SeV), the murine counterpart of Parainfluenza virus, has been shown to directly infect the mouse brain via the olfactory neurons [66]. Another investigation demonstrated that SeV infection led to impairing mouse olfaction. Interestingly, the virus persists in OE and OB tissues for over two months, and reduces the regenerative power and the functionality of the ORNs [67]. The seasonal changes play a key role in influenza and parainfluenza type 3 infections to induce olfactory loss. This dysfunction occurs most frequently in winter and spring [68]. Preventing the influenza A virus from reaching and infecting the upper brain is crucial for maintaining the process of detecting odours. Mori et al. have shown that ORNs undergoing apoptosis after infection may be a preferential mechanism to provide protective effects against invasion of the neurovirulent virus from the peripheral to the CNS [69,70]. In mice infected with influenza virus, Bcl-2, bax, and iNOS may play a key role in apoptosis regulation of ORNs [71]. A recent complementary investigation demonstrated that the homologous vaccination significantly decreased the H5N1 virus replication in the olfactory mucosa, compared to prophylactic oseltamivir and thus hindered subsequent virus spread to the CNS [72]. Another recent study using transcriptomic analysis demonstrated that olfactory signaling is among the altered pathways in patients suffering from RSV infection. This finding further supports previous work that described RSV as a causative agent of post-viral olfactory dysfunction. Interestingly, the authors highlighted that this molecular signaling could be a promising future route to investigate drug targets against RSV infection [29,60].
6. Mechanisms of SARS-CoV-2 Mediating the Loss of Smell
The post-COVID and the long-term-COVID have both tremendously triggered a lot of complications in different human systems. The loss or reduction of smell, among other complications of the nervous system, is an associated symptom for patients affected by different variants of COVID-19 including omicron variant (Table 1) [73,74,75,76,77,78,79]. Moreover, studies reported that the prevalence of olfactory dysfunction differs greatly between populations and approaches [77,80,81]. The relationship between the loss of smell and the SARS-CoV-2 infection remains an enigma which could partly explain why the fight against this disorder is difficult. In the emergency context of the pandemic, many COVID-19 vaccines are authorized to help protect and eliminate the virus. Unfortunately, at least, two COVID-19 vaccines have been reported to cause the loss of smell one day after the administration. Also, it remains unclear what drive the occurrence of olfactory dysfunctions as the cases are relatively rare, although the development of a post-vaccine inflammatory reaction in the olfactory neuroepithelium is pointed to play a role in this process [82,83]. Therefore, this adds more complexity to compare the molecular signaling of the loss of smell driven by COVID-19 infected patients and individuals benefiting from COVID-19 vaccines and later contracting the disease. The COVID-19 pathology and the cellular mechanism by which the olfactory dysfunction occurs, remain unclear and is still under intensive investigation [18,84]. Earlier in the pandemic, reports hypothesized that five potential mechanisms were considered to get insights into the olfactory dysfunction in COVID-19 patients : (1) obstruction/congestion and rhinorrhea of the nasal airway, (2) damage and loss of ORNs, (3) Olfactory center damage in the brain, (4) damage of the olfactory supporting cells in the OE, and (5) Inflammation-related olfactory epithelium dysfunction [80,81]. Butowt et al, have recently reviewed that at least the following hypotheses (1)-(3) turned out to be implausible, for explaining the olfactory dysfunction in patients [85]. Here, we will particularly review the mechanisms related to the second and the fourth scenarios according the available findings. Healthy sensory cilia of ORNs in the olfactory epithelium are crucial in perceiving odorant molecules before sending the information to the olfactory bulbs and then to the upper parts of the brain [52]. It has been reported in humans that the SARS-CoV-2 may inderectly affects the olfactory cilia, hindering the smelling system's efficacy [86]. Reports suggested that ORNs lack to express the entry proteins of SARS-CoV-2 in the OE. The virus seems to establish a first contact in human nasal epithelia by binding its spike S protein to specific cells in the OE [87]. These reports are confirmed by study based on in-silico data, predicting the that mature ORNs do not express the virus entry protein, the angiotensin-converting enzyme 2 (ACE2), and therefore are not likely to be infected by SARS-CoV-2 [88]. Furthermore, supporting data by Bryche et al., showed that SARS-CoV-2 was not detected in the ORNs of golden Siryan hamsters [89]. However, in few cases, authors suggested that SARS-CoV-2 could infect ORNs in hamsters [90]. Based on the fact that COVID-19-related loss of smell disappeared within 1-2 weeks, while the regeneration of dead ORNs needs more than 2 weeks, many data tend to conclude that COVID-19-related olfactory dysfunction (OD) is not directly associated with the impairment of the ORNs [13,80,81,87,91]. Consequently, studying the entry proteins expression within the cells in the OE will help to understand the sensitivity of the OE to SARS-CoV-2 infection-related to the high prevalence of ODs in patients. Many groups are now interested to the organization of the sustentacular cells in the OE and thought that they might play central role in leading to OD. A high level of expression of ACE2 and the transmembrane serine protease 2 (TMPRSS2) is particularly found on the sustentacular cells suggesting a path to the neurotropism of SARS-CoV-2 in the OE. The ACE2 and TMPRSS2 are respectively known as the SARS-CoV-2 receptor and the SARS-CoV-2 cell entry-priming protease. ACE2 is found mainly on different parts of the sustentacular cells both in human and mouse. The ACE2 and TMPRSS2 genes tend to be co-regulated [81,92,93,94,95,96]. Different approaches using tissues, cells and organ systems in human, golden Syrian hamster, and hACE2 transgenic mouse have been employed to study the pathological impact of the SARS-CoV-2. Here, we discussed findings related particularly to the OE in inducing ODs in human. The spike protein (S protein) of SARS-CoV-2 mediates the passage of the virus into the host cell by fusing the viral and host cell membranes. In fact, via his spike S, SARS-CoV-2 employs the ACE2 as host functional receptor and TMPRSS2 as the cellular priming protease facilitating viral uptake, both signaling being confirmed by Single-cell RNA sequencing (scRNA-seq) datasets from the Human Cell Atlas consortium [97,98,99]. Another study showed that SARS-CoV-2 Nucleocapsid protein (NP), was observed in human OE through the neuronal marker Tuj1, 9 hours post infection. This data further supported the enrichment of ACE2 in human olfactory sustentacular cells [93,100]. Earlier in the pandemic, the golden Syrian hamster was used as a model to document the pathology of SARS-CoV-2 in the OE post infection. Reports showed that the sustentacular cells are rapidly infected by SARS-CoV-2. This viral neurtropism is associated with a massive recruitment of immune cells in the OE and lamina propria, which could drive the disorganization of the OE structure [89]. This study is consistent with high level of TNFα observed in OE samples from COVID-19 suffering patients [101]. Furthermore, the inflammation induced by SARS-CoV-2 infected supporting cells may play an important role in the onset and persistence of loss of smell in patients. This SARS-CoV-2-associated inflammation status was confirmed by analyzing the expression of selected targets in the olfactory bulb using RNA-seq and RT-qPCR tools. Interestingly, this study showed that the proinflammatory markers Cxcl10, Il-1β, Ccl5 and Irf7 overexpression continued up to 14 dpi, when animals had recovered from ageusia/anosmia (77). These findings are in line with a very recent study showing the implication of immune cell infiltration and altered gene expression in OE in driving persistent smell loss in a subset of patients with SARS-CoV-2. Moreover, this study particularly, demonstrates that T cell–mediated inflammation lasts longer in the OE after the acute SARS-CoV-2 infection has been eliminated from the tissue, suggesting a mechanistic insights into the long-term post–COVID-19 smell loss (78). The OE disorganization is followed by a drastic deterioration of the cilia layer of the ORNs that leads to the impairment of the olfactory capacity of the animal [89]. Investigations in humans and hamsters using respectively, Transmission Electron Microscopy (TEM) studies and Scanning Electron Microscopy (SEM) analysis showed various levels of cilia height that undergo regeneration in the course of patient recovery, including smell restoration. Data using the golden Syrian hamster showed that the regenerated cilia in the epithelium is accompanied by a decreased expression of FOXJ1+ highlighting the importance of this marker in the respiratory ciliogenesis. This later finding by Schreiner et al., could in part shed light on the inquiry of how could we regenerate cilia during patient recovery, although a lot needs to be done in the roadmap of treating loss of smell related to nasal cilia deterioration by SARS-CoV-2 (75, 76) (Table 1 ; Figure 2).
According to the literature, different variants of SARS-CoV-2 do not directly target particularly the ORNs in the OE, instead they are found in majority expressed in the sustentacular cells [36]. Recent study by Seehusen et al, showed that K18-hACE2 transgenic mouse expressing the human ACE2 is highly sensitive to at least five variants of SARS-CoV-2 that infected not only the supportive cells in OE and the respiratory epithelium but invaded the CNS of the animal five days post infection. Interestingly, the expression of hACE2 seems to convey higher binding affinity when compared to the wild-type mouse [113]. Using this transgenic mouse reveals to be a serious option for therapy development against loss of smell as these animals exhibited low mortality when treated with COVID-19 convalescent antisera [113,114].
It is now accepted that ACE2 is not the only obligate entry for SARS-CoV-2 as it has been suggested that molecules including PIKfyve or neuropilin-1 (NRP-1) may participate in SARS-CoV-2 entry [115,116,117]. Like ACE2, NRP-1 is highly expressed in the respiratory and olfactory epithelium which further support the infectivity and entry of SARS-CoV-2 in the human OE. NRP-1 is not only found in supportive cells but is expressed in nearly every cell type in the nasal passages including the ORN, therefore giving SARS-CoV-2 a route to access those cells and impair the olfactory response. Interestingly, Daly et al., demonstrated that the selective inhibition of the S1-NRP-1 interaction reduces SARS-CoV-2 infection [117,118,119].
Taken together, the highly expression of ACE2, TMPRSS2 and NRP-1 in supportive and other olfactory cells and their impact in olfactory neurophysiology maintenance and in the development of human olfactory pathophysiology supports them as potential targets for signaling-based therapeutics of anosmia. Finally, for further insight into the molecular signaling and treatments in COVID-19 patients with impaired smelling, readers are invited to explore the following reviews. They have explored many different potential therapeutic agents against olfactory and gustatory dysfunctions [73].
7. Conclusions and Perspective
Our literature review further confirms the previous extended investigations showing that loss of smell and taste are among the key associated symptoms with most COVID-19 variants, including the omicron variant which causes runny nose, headache, fatigue, sneezing, and sore throat [11]. The last three years have been an important rush towards deciphering the underlying mechanisms the SARS-CoV-2 deploys to impair the olfaction in infected patients. Still few works have addressed the molecular signaling mediated by SARS-CoV-2-induced loss of smell in human patients. This work pointed the urge and necessity of finding an adequate therapeutic solution against the COVID-19 pathogen in general and the loss of smell affecting patients in particular. In addition, the mechanisms of taste dysfunction due to COVID-19 infection is not discussed in this review. But it would be interesting to decipher the possible pathogenesis between ageusia and anosmia in COVID-19 patients in the future.
Author Contributions
All authors contributed to the literature review, writing the article, revising the manuscript, have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wu JT, Leung K, Leung GM. Nowcasting and forecasting the potential domestic and international spread of the 2019-nCoV outbreak originating in Wuhan, China: a modelling study. Lancet. 2020;395(10225):689-97. Epub 2020/02/06. PubMed PMID: 32014114; PubMed Central PMCID: PMCPMC7159271. [CrossRef]
- Hui DS, E IA, Madani TA, Ntoumi F, Kock R, Dar O, et al. The continuing 2019-nCoV epidemic threat of novel coronaviruses to global health - The latest 2019 novel coronavirus outbreak in Wuhan, China. Int J Infect Dis. 2020;91:264-6. Epub 2020/01/19. PubMed PMID: 31953166; PubMed Central PMCID: PMCPMC7128332. [CrossRef]
- Fisher D, Heymann D. Q&A: The novel coronavirus outbreak causing COVID-19. BMC Med. 2020;18(1):57. Epub 2020/02/29. PubMed PMID: 32106852; PubMed Central PMCID: PMCPMC7047369. [CrossRef]
- World Heath Organization. Situation report – 51. Available online at : https://wwwwhoint/docs/default- source/coronaviruse/situation- reports/20200311-sitrep-51-covid-19pdf?sfvrsn=1ba62e57_10 (2020). (accessed on April 15, 2023).
- World Health Organization. Coronavirus disease (COVID-2019) situation reports. Available online at: www.who.int/emergencies/diseases/novelcoronavirus-2019/situation-reports/. (accessed on April 15, 2023).
- Gandhi M, Yokoe DS, Havlir DV. Asymptomatic Transmission, the Achilles' Heel of Current Strategies to Control Covid-19. N Engl J Med. 2020;382(22):2158-60. Epub 2020/04/25. PubMed PMID: 32329972; PubMed Central PMCID: PMCPMC7200054. [CrossRef]
- Wolfel R, Corman VM, Guggemos W, Seilmaier M, Zange S, Muller MA, et al. Virological assessment of hospitalized patients with COVID-2019. Nature. 2020;581(7809):465-9. Epub 2020/04/03. [CrossRef] [PubMed]
- Yang X, Yu Y, Xu J, Shu H, Xia J, Liu H, et al. Clinical course and outcomes of critically ill patients with SARS-CoV-2 pneumonia in Wuhan, China: a single-centered, retrospective, observational study. Lancet Respir Med. 2020;8(5):475-81. Epub 2020/02/28. PubMed PMID: 32105632; PubMed Central PMCID: PMCPMC7102538. [CrossRef]
- Guan WJ, Ni ZY, Hu Y, Liang WH, Ou CQ, He JX, et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N Engl J Med. 2020;382(18):1708-20. Epub 2020/02/29. PubMed PMID: 32109013; PubMed Central PMCID: PMCPMC7092819. [CrossRef]
- Huang C, Wang Y, Li X, Ren L, Zhao J, Hu Y, et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet. 2020;395(10223):497-506. Epub 2020/01/28. PubMed PMID: 31986264; PubMed Central PMCID: PMCPMC7159299. [CrossRef]
- Iacobucci, G. Covid-19: Runny nose, headache, and fatigue are commonest symptoms of omicron, early data show. BMJ. 2021;375:n3103. Epub 2021/12/18. [CrossRef] [PubMed]
- Bagheri SH, Asghari A, Farhadi M, Shamshiri AR, Kabir A, Kamrava SK, et al. Coincidence of COVID-19 epidemic and olfactory dysfunction outbreak in Iran. Med J Islam Repub Iran. 2020;34:62. Epub 2020/09/26. PubMed PMID: 32974228; PubMed Central PMCID: PMCPMC7500422. [CrossRef]
- Lechien JR, Chiesa-Estomba CM, De Siati DR, Horoi M, Le Bon SD, Rodriguez A, et al. Olfactory and gustatory dysfunctions as a clinical presentation of mild-to-moderate forms of the coronavirus disease (COVID-19): a multicenter European study. Eur Arch Otorhinolaryngol. 2020;277(8):2251-61. Epub 2020/04/08. PubMed PMID: 32253535; PubMed Central PMCID: PMCPMC7134551. [CrossRef]
- Rebholz H, Braun RJ, Ladage D, Knoll W, Kleber C, Hassel AW. Loss of Olfactory Function-Early Indicator for Covid-19, Other Viral Infections and Neurodegenerative Disorders. Front Neurol. 2020;11:569333. Epub 2020/11/17. PubMed PMID: 33193009; PubMed Central PMCID: PMCPMC7649754. [CrossRef]
- Maniaci A, Ferlito S, Bubbico L, Ledda C, Rapisarda V, Iannella G, et al. Comfort rules for face masks among healthcare workers during COVID-19 spread. Ann Ig. 2021;33(6):615-27. Epub 2021/04/03. [CrossRef] [PubMed]
- de Melo GD, Lazarini F, Levallois S, Hautefort C, Michel V, Larrous F, et al. COVID-19-related anosmia is associated with viral persistence and inflammation in human olfactory epithelium and brain infection in hamsters. Sci Transl Med. 2021;13(596). Epub 2021/05/05. PubMed PMID: 33941622; PubMed Central PMCID: PMCPMC8158965. [CrossRef]
- Zazhytska M, Kodra A, Hoagland DA, Frere J, Fullard JF, Shayya H, et al. Non-cell-autonomous disruption of nuclear architecture as a potential cause of COVID-19-induced anosmia. Cell. 2022;185(6):1052-64 e12. Epub 2022/02/19. PubMed PMID: 35180380; PubMed Central PMCID: PMCPMC8808699. [CrossRef]
- Bilinska K, Butowt R. Anosmia in COVID-19: A Bumpy Road to Establishing a Cellular Mechanism. ACS Chem Neurosci. 2020;11(15):2152-5. Epub 2020/07/17. PubMed PMID: 32673476; PubMed Central PMCID: PMCPMC7467568. [CrossRef]
- Samaranayake LP, Fakhruddin KS, Panduwawala C. Sudden onset, acute loss of taste and smell in coronavirus disease 2019 (COVID-19): a systematic review. Acta Odontol Scand. 2020;78(6):467-73. Epub 2020/08/09. [CrossRef] [PubMed]
- Kirchdoerfer RN, Wang N, Pallesen J, Wrapp D, Turner HL, Cottrell CA, et al. Stabilized coronavirus spikes are resistant to conformational changes induced by receptor recognition or proteolysis. Sci Rep. 2018;8(1):15701. Epub 2018/10/26. PubMed PMID: 30356097; PubMed Central PMCID: PMCPMC6200764. [CrossRef]
- Othman BA, Maulud SQ, Jalal PJ, Abdulkareem SM, Ahmed JQ, Dhawan M, et al. Olfactory dysfunction as a post-infectious symptom of SARS-CoV-2 infection. Ann Med Surg (Lond). 2022;75:103352. Epub 2022/02/17. PubMed PMID: 35169465; PubMed Central PMCID: PMCPMC8830927. [CrossRef]
- Firestein, S. How the olfactory system makes sense of scents. Nature. 2001;413(6852):211-8. Epub 2001/09/15. [CrossRef] [PubMed]
- Olender T, Lancet D, Nebert DW. Update on the olfactory receptor (OR) gene superfamily. Hum Genomics. 2008;3(1):87-97. Epub 2009/01/09. PubMed PMID: 19129093; PubMed Central PMCID: PMCPMC2752031. [CrossRef]
- Reed, RR. After the holy grail: establishing a molecular basis for Mammalian olfaction. Cell. 2004;116(2):329-36. Epub 2004/01/28. [CrossRef] [PubMed]
- Dammalli M, Dey G, Madugundu AK, Kumar M, Rodrigues B, Gowda H, et al. Proteomic Analysis of the Human Olfactory Bulb. OMICS. 2017;21(8):440-53. Epub 2017/08/18. [CrossRef] [PubMed]
- Huttenbrink KB, Hummel T, Berg D, Gasser T, Hahner A. Olfactory dysfunction: common in later life and early warning of neurodegenerative disease. Dtsch Arztebl Int. 2013;110(1-2):1-7, e1. Epub 2013/03/02. PubMed PMID: 23450985; PubMed Central PMCID: PMCPMC3561743. [CrossRef]
- Dammalli M, Dey G, Kumar M, Madugundu AK, Gopalakrishnan L, Gowrishankar BS, et al. Proteomics of the Human Olfactory Tract. OMICS. 2018;22(1):77-87. Epub 2018/01/23. [CrossRef] [PubMed]
- Oboti L, Peretto P, Marchis SD, Fasolo A. From chemical neuroanatomy to an understanding of the olfactory system. Eur J Histochem. 2011;55(4):e35. Epub 2012/02/03. PubMed PMID: 22297441; PubMed Central PMCID: PMCPMC3284237. [CrossRef]
- Barral-Arca R, Gomez-Carballa A, Cebey-Lopez M, Bello X, Martinon-Torres F, Salas A. A Meta-Analysis of Multiple Whole Blood Gene Expression Data Unveils a Diagnostic Host-Response Transcript Signature for Respiratory Syncytial Virus. Int J Mol Sci. 2020;21(5). Epub 2020/03/12. PubMed PMID: 32155831; PubMed Central PMCID: PMCPMC7084441. [CrossRef]
- Boesveldt S, Postma EM, Boak D, Welge-Luessen A, Schopf V, Mainland JD, et al. Anosmia-A Clinical Review. Chem Senses. 2017;42(7):513-23. Epub 2017/05/23. PubMed PMID: 28531300; PubMed Central PMCID: PMCPMC5863566. [CrossRef]
- Brennan PA, Keverne EB. Something in the air? New insights into mammalian pheromones. Curr Biol. 2004;14(2):R81-9. Epub 2004/01/24. [CrossRef] [PubMed]
- Buck, LB. The molecular architecture of odor and pheromone sensing in mammals. Cell. 2000;100(6):611-8. Epub 2000/04/13. [CrossRef] [PubMed]
- Mombaerts, P. Genes and ligands for odorant, vomeronasal and taste receptors. Nat Rev Neurosci. 2004;5(4):263-78. Epub 2004/03/23. [CrossRef] [PubMed]
- Restrepo D, Arellano J, Oliva AM, Schaefer ML, Lin W. Emerging views on the distinct but related roles of the main and accessory olfactory systems in responsiveness to chemosensory signals in mice. Horm Behav. 2004;46(3):247-56. Epub 2004/08/25. [CrossRef] [PubMed]
- Lavoie J, Gasso Astorga P, Segal-Gavish H, Wu YC, Chung Y, Cascella NG, et al. The Olfactory Neural Epithelium As a Tool in Neuroscience. Trends Mol Med. 2017;23(2):100-3. Epub 2017/01/22. PubMed PMID: 28108112; PubMed Central PMCID: PMCPMC5399677. [CrossRef]
- Liang F, Wang Y. COVID-19 Anosmia: High Prevalence, Plural Neuropathogenic Mechanisms, and Scarce Neurotropism of SARS-CoV-2? Viruses. 2021;13(11). Epub 2021/11/28. PubMed PMID: 34835030; PubMed Central PMCID: PMCPMC8625547. [CrossRef]
- Malnic B, Godfrey PA, Buck LB. The human olfactory receptor gene family. Proc Natl Acad Sci U S A. 2004;101(8):2584-9. Epub 2004/02/26. PubMed PMID: 14983052; PubMed Central PMCID: PMCPMC356993. [CrossRef]
- Sosinsky A, Glusman G, Lancet D. The genomic structure of human olfactory receptor genes. Genomics. 2000;70(1):49-61. Epub 2000/11/23. [CrossRef] [PubMed]
- Zozulya S, Echeverri F, Nguyen T. The human olfactory receptor repertoire. Genome Biol. 2001;2(6):RESEARCH0018. Epub 2001/06/26. PubMed PMID: 11423007; PubMed Central PMCID: PMCPMC33394. [CrossRef]
- Young JM, Trask BJ. The sense of smell: genomics of vertebrate odorant receptors. Hum Mol Genet. 2002;11(10):1153-60. Epub 2002/05/17. [CrossRef] [PubMed]
- Zhang X, Firestein S. The olfactory receptor gene superfamily of the mouse. Nat Neurosci. 2002;5(2):124-33. Epub 2002/01/22. [CrossRef] [PubMed]
- Breer H, Raming K, Krieger J, Boekhoff I, Strotmann J. Towards an identification of odorant receptors. J Recept Res. 1993;13(1-4):527-40. Epub 1993/01/01. [CrossRef] [PubMed]
- Amoore, JE. Stereochemical and vibrational theories of odour. Nature. 1971;233(5317):270-1. Epub 1971/09/24. [CrossRef] [PubMed]
- Araneda RC, Peterlin Z, Zhang X, Chesler A, Firestein S. A pharmacological profile of the aldehyde receptor repertoire in rat olfactory epithelium. J Physiol. 2004;555(Pt 3):743-56. Epub 2004/01/16. PubMed PMID: 14724183; PubMed Central PMCID: PMCPMC1664868. [CrossRef]
- Sanz G, Schlegel C, Pernollet JC, Briand L. Comparison of odorant specificity of two human olfactory receptors from different phylogenetic classes and evidence for antagonism. Chem Senses. 2005;30(1):69-80. Epub 2005/01/14. [CrossRef] [PubMed]
- Ma M, Shepherd GM. Functional mosaic organization of mouse olfactory receptor neurons. Proc Natl Acad Sci U S A. 2000;97(23):12869-74. Epub 2000/10/26. PubMed PMID: 11050155; PubMed Central PMCID: PMCPMC18856. [CrossRef]
- Wade F, Espagne A, Persuy MA, Vidic J, Monnerie R, Merola F, et al. Relationship between homo-oligomerization of a mammalian olfactory receptor and its activation state demonstrated by bioluminescence resonance energy transfer. J Biol Chem. 2011;286(17):15252-9. Epub 2011/04/02. PubMed PMID: 21454689; PubMed Central PMCID: PMCPMC3083150. [CrossRef]
- Stewart WB, Kauer JS, Shepherd GM. Functional organization of rat olfactory bulb analysed by the 2-deoxyglucose method. J Comp Neurol. 1979;185(4):715-34. Epub 1979/06/15. [CrossRef] [PubMed]
- Minic J, Persuy MA, Godel E, Aioun J, Connerton I, Salesse R, et al. Functional expression of olfactory receptors in yeast and development of a bioassay for odorant screening. FEBS J. 2005;272(2):524-37. Epub 2005/01/19. [CrossRef] [PubMed]
- Johnson BA, Leon M. Odorant molecular length: one aspect of the olfactory code. J Comp Neurol. 2000;426(2):330-8. Epub 2000/09/12. [CrossRef] [PubMed]
- Kurian SM, Gordon S, Barrick B, Dadlani MN, Fanelli B, Cornell JB, et al. Feasibility and Comparison Study of Fecal Sample Collection Methods in Healthy Volunteers and Solid Organ Transplant Recipients Using 16S rRNA and Metagenomics Approaches. Biopreserv Biobank. 2020;18(5):425-40. Epub 2020/08/25. [CrossRef] [PubMed]
- Glezer I, Malnic B. Olfactory receptor function. Handb Clin Neurol. 2019;164:67-78. Epub 2019/10/13. [CrossRef] [PubMed]
- Schwob, JE. Neural regeneration and the peripheral olfactory system. Anat Rec. 2002;269(1):33-49. Epub 2002/03/14. [CrossRef] [PubMed]
- Graziadei PP, Graziadei GA. Neurogenesis and neuron regeneration in the olfactory system of mammals. I. Morphological aspects of differentiation and structural organization of the olfactory sensory neurons. J Neurocytol. 1979;8(1):1-18. Epub 1979/02/01. [CrossRef] [PubMed]
- Urata S, Maruyama J, Kishimoto-Urata M, Sattler RA, Cook R, Lin N, et al. Regeneration Profiles of Olfactory Epithelium after SARS-CoV-2 Infection in Golden Syrian Hamsters. ACS Chem Neurosci. 2021;12(4):589-95. Epub 2021/02/02. PubMed PMID: 33522795; PubMed Central PMCID: PMCPMC7874468. [CrossRef]
- Dicpinigaitis, PV. Post-viral Anosmia (Loss of Sensation of Smell) Did Not Begin with COVID-19! Lung. 2021;199(3):237-8. Epub 2021/04/25. PubMed PMID: 33893845; PubMed Central PMCID: PMCPMC8067782. [CrossRef]
- Suzuki M, Saito K, Min WP, Vladau C, Toida K, Itoh H, et al. Identification of viruses in patients with postviral olfactory dysfunction. Laryngoscope. 2007;117(2):272-7. Epub 2007/02/06. PubMed PMID: 17277621; PubMed Central PMCID: PMCPMC7165544. [CrossRef]
- Lee DY, Lee WH, Wee JH, Kim JW. Prognosis of postviral olfactory loss: follow-up study for longer than one year. Am J Rhinol Allergy. 2014;28(5):419-22. Epub 2014/09/10. [CrossRef] [PubMed]
- Welge-Lussen A, Wolfensberger M. Olfactory disorders following upper respiratory tract infections. Adv Otorhinolaryngol. 2006;63:125-32. Epub 2006/05/31. [CrossRef] [PubMed]
- Seiden, AM. Postviral olfactory loss. Otolaryngol Clin North Am. 2004;37(6):1159-66. Epub 2004/11/27. [CrossRef] [PubMed]
- Moran DT, Jafek BW, Eller PM, Rowley JC, 3rd. Ultrastructural histopathology of human olfactory dysfunction. Microsc Res Tech. 1992;23(2):103-10. Epub 1992/10/15. [CrossRef] [PubMed]
- Doty RL, Hawkes CH. Chemosensory dysfunction in neurodegenerative diseases. Handb Clin Neurol. 2019;164:325-60. Epub 2019/10/13. [CrossRef] [PubMed]
- Lazarini F, Katsimpardi L, Levivien S, Wagner S, Gressens P, Teissier N, et al. Congenital Cytomegalovirus Infection Alters Olfaction Before Hearing Deterioration In Mice. J Neurosci. 2018;38(49):10424-37. Epub 2018/10/21. PubMed PMID: 30341181; PubMed Central PMCID: PMCPMC6596252. [CrossRef]
- Teissier N, Fallet-Bianco C, Delezoide AL, Laquerriere A, Marcorelles P, Khung-Savatovsky S, et al. Cytomegalovirus-induced brain malformations in fetuses. J Neuropathol Exp Neurol. 2014;73(2):143-58. Epub 2014/01/16. [CrossRef] [PubMed]
- Tian J, Liu L, Zhou K, Hong Z, Chen Q, Jiang F, et al. Metal-organic tube or layered assembly: reversible sheet-to-tube transformation and adaptive recognition. Chem Sci. 2020;11(36):9818-26. Epub 2021/06/08. PubMed PMID: 34094242; PubMed Central PMCID: PMCPMC8162108. [CrossRef]
- Mori I, Komatsu T, Takeuchi K, Nakakuki K, Sudo M, Kimura Y. Parainfluenza virus type 1 infects olfactory neurons and establishes long-term persistence in the nerve tissue. J Gen Virol. 1995;76 ( Pt 5):1251-4. Epub 1995/05/01. [CrossRef] [PubMed]
- Tian J, Pinto JM, Cui X, Zhang H, Li L, Liu Y, et al. Sendai Virus Induces Persistent Olfactory Dysfunction in a Murine Model of PVOD via Effects on Apoptosis, Cell Proliferation, and Response to Odorants. PLoS One. 2016;11(7):e0159033. Epub 2016/07/20. PubMed PMID: 27428110; PubMed Central PMCID: PMCPMC4948916. [CrossRef]
- Nakashima T, Suzuki H, Teranishi M. Olfactory and gustatory dysfunction caused by SARS-CoV-2: Comparison with cases of infection with influenza and other viruses. Infect Control Hosp Epidemiol. 2021;42(1):113-4. Epub 2020/05/06. PubMed PMID: 32366336; PubMed Central PMCID: PMCPMC7298085. [CrossRef]
- van Riel D, Verdijk R, Kuiken T. The olfactory nerve: a shortcut for influenza and other viral diseases into the central nervous system. J Pathol. 2015;235(2):277-87. Epub 2014/10/09. [CrossRef] [PubMed]
- Mori I, Goshima F, Imai Y, Kohsaka S, Sugiyama T, Yoshida T, et al. Olfactory receptor neurons prevent dissemination of neurovirulent influenza A virus into the brain by undergoing virus-induced apoptosis. J Gen Virol. 2002;83(Pt 9):2109-16. Epub 2002/08/20. [CrossRef] [PubMed]
- Chen Z, Ni D, Gao Y, Lin J. [Apoptosis related genes--Bcl-2, bax and iNOS, expressed in the olfactory epithelium of mice infected with influenza virus]. Lin Chung Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 2007;21(11):510-2. Epub 2007/08/07. [PubMed]
- Siegers JY, van den Brand JM, Leijten LM, van de Bildt MM, van Run PR, van Amerongen G, et al. Vaccination Is More Effective Than Prophylactic Oseltamivir in Preventing CNS Invasion by H5N1 Virus via the Olfactory Nerve. J Infect Dis. 2016;214(4):516-24. Epub 2016/07/23. [CrossRef] [PubMed]
- Khani E, Khiali S, Beheshtirouy S, Entezari-Maleki T. Potential pharmacologic treatments for COVID-19 smell and taste loss: A comprehensive review. Eur J Pharmacol. 2021;912:174582. Epub 2021/10/23. PubMed PMID: 34678243; PubMed Central PMCID: PMCPMC8524700. [CrossRef]
- Silva Andrade B, Siqueira S, de Assis Soares WR, de Souza Rangel F, Santos NO, Dos Santos Freitas A, et al. Long-COVID and Post-COVID Health Complications: An Up-to-Date Review on Clinical Conditions and Their Possible Molecular Mechanisms. Viruses. 2021;13(4). Epub 2021/05/01. PubMed PMID: 33919537; PubMed Central PMCID: PMCPMC8072585. [CrossRef]
- Fernandez-de-Las-Penas C, Cancela-Cilleruelo I, Rodriguez-Jimenez J, Gomez-Mayordomo V, Pellicer-Valero OJ, Martin-Guerrero JD, et al. Associated-Onset Symptoms and Post-COVID-19 Symptoms in Hospitalized COVID-19 Survivors Infected with Wuhan, Alpha or Delta SARS-CoV-2 Variant. Pathogens. 2022;11(7). Epub 2022/07/28. PubMed PMID: 35889971; PubMed Central PMCID: PMCPMC9320021. [CrossRef]
- von Bartheld CS, Hagen MM, Butowt R. The D614G Virus Mutation Enhances Anosmia in COVID-19 Patients: Evidence from a Systematic Review and Meta-analysis of Studies from South Asia. ACS Chem Neurosci. 2021;12(19):3535-49. Epub 2021/09/18. PubMed PMID: 34533304; PubMed Central PMCID: PMCPMC8482322. [CrossRef]
- von Bartheld CS, Wang L. Prevalence of Olfactory Dysfunction with the Omicron Variant of SARS-CoV-2: A Systematic Review and Meta-Analysis. Cells. 2023;12(3). Epub 2023/02/12. PubMed PMID: 36766771; PubMed Central PMCID: PMCPMC9913864. [CrossRef]
- Rodriguez-Sevilla JJ, Guerri-Fernadez R, Bertran Recasens B. Is There Less Alteration of Smell Sensation in Patients With Omicron SARS-CoV-2 Variant Infection? Front Med (Lausanne). 2022;9:852998. Epub 2022/05/03. PubMed PMID: 35492353; PubMed Central PMCID: PMCPMC9039252. [CrossRef]
- Chee J, Chern B, Loh WS, Mullol J, Wang Y. Pathophysiology of SARS-CoV-2 Infection of Nasal Respiratory and Olfactory Epithelia and Its Clinical Impact. Curr Allergy Asthma Rep. 2023;23(2):121-31. Epub 2023/01/05. PubMed PMID: 36598732; PubMed Central PMCID: PMCPMC9811886. [CrossRef]
- Mutiawati E, Fahriani M, Mamada SS, Fajar JK, Frediansyah A, Maliga HA, et al. Anosmia and dysgeusia in SARS-CoV-2 infection: incidence and effects on COVID-19 severity and mortality, and the possible pathobiology mechanisms - a systematic review and meta-analysis. F1000Res. 2021;10:40. Epub 2021/04/08. PubMed PMID: 33824716; PubMed Central PMCID: PMCPMC7993408. [CrossRef]
- Butowt R, von Bartheld CS. Anosmia in COVID-19: Underlying Mechanisms and Assessment of an Olfactory Route to Brain Infection. Neuroscientist. 2021;27(6):582-603. Epub 2020/09/12. PubMed PMID: 32914699; PubMed Central PMCID: PMCPMC7488171. [CrossRef]
- Fantin F, Frosolini A, Tundo I, Inches I, Fabbris C, Spinato G, et al. A singular case of hyposmia and transient audiovestibular post-vaccine disorders: case report and literature review. Transl Neurosci. 2022;13(1):349-53. Epub 2022/10/29. PubMed PMID: 36304095; PubMed Central PMCID: PMCPMC9547348. [CrossRef]
- Lechien JR, Diallo AO, Dachy B, Le Bon SD, Maniaci A, Vaira LA, et al. COVID-19: Post-vaccine Smell and Taste Disorders: Report of 6 Cases. Ear Nose Throat J. 2021:1455613211033125. Epub 2021/09/02. [CrossRef] [PubMed]
- Mao L, Jin H, Wang M, Hu Y, Chen S, He Q, et al. Neurologic Manifestations of Hospitalized Patients With Coronavirus Disease 2019 in Wuhan, China. JAMA Neurol. 2020;77(6):683-90. Epub 2020/04/11. PubMed PMID: 32275288; PubMed Central PMCID: PMCPMC7149362. [CrossRef]
- Butowt R, Bilinska K, von Bartheld CS. Olfactory dysfunction in COVID-19: new insights into the underlying mechanisms. Trends Neurosci. 2023;46(1):75-90. Epub 2022/12/06. PubMed PMID: 36470705; PubMed Central PMCID: PMCPMC9666374. [CrossRef]
- Buqaileh R, Saternos H, Ley S, Aranda A, Forero K, AbouAlaiwi WA. Can cilia provide an entry gateway for SARS-CoV-2 to human ciliated cells? Physiol Genomics. 2021;53(6):249-58. Epub 2021/04/16. PubMed PMID: 33855870; PubMed Central PMCID: PMCPMC8213509. [CrossRef]
- Butowt R, Meunier N, Bryche B, von Bartheld CS. The olfactory nerve is not a likely route to brain infection in COVID-19: a critical review of data from humans and animal models. Acta Neuropathol. 2021;141(6):809-22. Epub 2021/04/28. PubMed PMID: 33903954; PubMed Central PMCID: PMCPMC8075028. [CrossRef]
- Bilinska K, Jakubowska P, Von Bartheld CS, Butowt R. Expression of the SARS-CoV-2 Entry Proteins, ACE2 and TMPRSS2, in Cells of the Olfactory Epithelium: Identification of Cell Types and Trends with Age. ACS Chem Neurosci. 2020;11(11):1555-62. Epub 2020/05/08. PubMed PMID: 32379417; PubMed Central PMCID: PMCPMC7241737. [CrossRef]
- Bryche B, St Albin A, Murri S, Lacote S, Pulido C, Ar Gouilh M, et al. Massive transient damage of the olfactory epithelium associated with infection of sustentacular cells by SARS-CoV-2 in golden Syrian hamsters. Brain Behav Immun. 2020;89:579-86. Epub 2020/07/07. PubMed PMID: 32629042; PubMed Central PMCID: PMCPMC7332942. [CrossRef]
- Sia SF, Yan LM, Chin AWH, Fung K, Choy KT, Wong AYL, et al. Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature. 2020;583(7818):834-8. Epub 2020/05/15. PubMed PMID: 32408338; PubMed Central PMCID: PMCPMC7394720. [CrossRef]
- Lee Y, Min P, Lee S, Kim SW. Prevalence and Duration of Acute Loss of Smell or Taste in COVID-19 Patients. J Korean Med Sci. 2020;35(18):e174. Epub 2020/05/10. PubMed PMID: 32383370; PubMed Central PMCID: PMCPMC7211515. [CrossRef]
- Fodoulian L, Tuberosa J, Rossier D, Boillat M, Kan C, Pauli V, et al. SARS-CoV-2 Receptors and Entry Genes Are Expressed in the Human Olfactory Neuroepithelium and Brain. iScience. 2020;23(12):101839. Epub 2020/12/01. PubMed PMID: 33251489; PubMed Central PMCID: PMCPMC7685946. [CrossRef]
- Chen M, Shen W, Rowan NR, Kulaga H, Hillel A, Ramanathan M, Jr., et al. Elevated ACE-2 expression in the olfactory neuroepithelium: implications for anosmia and upper respiratory SARS-CoV-2 entry and replication. Eur Respir J. 2020;56(3). Epub 2020/08/21. PubMed PMID: 32817004; PubMed Central PMCID: PMCPMC7439429. [CrossRef]
- Hoffmann M, Kleine-Weber H, Schroeder S, Kruger N, Herrler T, Erichsen S, et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell. 2020;181(2):271-80 e8. Epub 2020/03/07. PubMed PMID: 32142651; PubMed Central PMCID: PMCPMC7102627. [CrossRef]
- Ziegler CGK, Allon SJ, Nyquist SK, Mbano IM, Miao VN, Tzouanas CN, et al. SARS-CoV-2 Receptor ACE2 Is an Interferon-Stimulated Gene in Human Airway Epithelial Cells and Is Detected in Specific Cell Subsets across Tissues. Cell. 2020;181(5):1016-35 e19. Epub 2020/05/16. PubMed PMID: 32413319; PubMed Central PMCID: PMCPMC7252096. [CrossRef]
- Gkogkou E, Barnasas G, Vougas K, Trougakos IP. Expression profiling meta-analysis of ACE2 and TMPRSS2, the putative anti-inflammatory receptor and priming protease of SARS-CoV-2 in human cells, and identification of putative modulators. Redox Biol. 2020;36:101615. Epub 2020/08/31. PubMed PMID: 32863223; PubMed Central PMCID: PMCPMC7311357. [CrossRef]
- Sungnak W, Huang N, Becavin C, Berg M, Queen R, Litvinukova M, et al. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat Med. 2020;26(5):681-7. Epub 2020/04/25. PubMed PMID: 32327758; PubMed Central PMCID: PMCPMC8637938. [CrossRef]
- Li W, Li M, Ou G. COVID-19, cilia, and smell. FEBS J. 2020;287(17):3672-6. Epub 2020/07/22. PubMed PMID: 32692465; PubMed Central PMCID: PMCPMC7426555. [CrossRef]
- Sternberg A, Naujokat C. Structural features of coronavirus SARS-CoV-2 spike protein: Targets for vaccination. Life Sci. 2020;257:118056. Epub 2020/07/10. PubMed PMID: 32645344; PubMed Central PMCID: PMCPMC7336130. [CrossRef]
- Chen M, Pekosz A, Villano JS, Shen W, Zhou R, Kulaga H, et al. Evolution of nasal and olfactory infection characteristics of SARS-CoV-2 variants. bioRxiv. 2022. Epub 2022/04/21. PubMed PMID: 35441175; PubMed Central PMCID: PMCPMC9016639. [CrossRef]
- Torabi A, Mohammadbagheri E, Akbari Dilmaghani N, Bayat AH, Fathi M, Vakili K, et al. Proinflammatory Cytokines in the Olfactory Mucosa Result in COVID-19 Induced Anosmia. ACS Chem Neurosci. 2020;11(13):1909-13. Epub 2020/06/12. PubMed PMID: 32525657; PubMed Central PMCID: PMCPMC7299394. [CrossRef]
- Seo JS, Yoon SW, Hwang SH, Nam SM, Nahm SS, Jeong JH, et al. The Microvillar and Solitary Chemosensory Cells as the Novel Targets of Infection of SARS-CoV-2 in Syrian Golden Hamsters. Viruses. 2021;13(8). Epub 2021/08/29. PubMed PMID: 34452517; PubMed Central PMCID: PMCPMC8402700. [CrossRef]
- Chen CR, Kachramanoglou C, Li D, Andrews P, Choi D. Anatomy and cellular constituents of the human olfactory mucosa: a review. J Neurol Surg B Skull Base. 2014;75(5):293-300. Epub 2014/10/11. PubMed PMID: 25302141; PubMed Central PMCID: PMCPMC4176544. [CrossRef]
- Klingenstein M, Klingenstein S, Neckel PH, Mack AF, Wagner AP, Kleger A, et al. Evidence of SARS-CoV2 Entry Protein ACE2 in the Human Nose and Olfactory Bulb. Cells Tissues Organs. 2020;209(4-6):155-64. Epub 2021/01/25. PubMed PMID: 33486479; PubMed Central PMCID: PMCPMC7900466. [CrossRef]
- Cooper KW, Brann DH, Farruggia MC, Bhutani S, Pellegrino R, Tsukahara T, et al. COVID-19 and the Chemical Senses: Supporting Players Take Center Stage. Neuron. 2020;107(2):219-33. Epub 2020/07/09. PubMed PMID: 32640192; PubMed Central PMCID: PMCPMC7328585. [CrossRef]
- Solbu TT, Holen T. Aquaporin pathways and mucin secretion of Bowman's glands might protect the olfactory mucosa. Chem Senses. 2012;37(1):35-46. Epub 2011/07/13. [CrossRef] [PubMed]
- Doty, RL. Olfactory dysfunction in COVID-19: pathology and long-term implications for brain health. Trends Mol Med. 2022;28(9):781-94. Epub 2022/07/10. PubMed PMID: 35810128; PubMed Central PMCID: PMCPMC9212891. [CrossRef]
- Villar PS, Delgado R, Vergara C, Reyes JG, Bacigalupo J. Energy Requirements of Odor Transduction in the Chemosensory Cilia of Olfactory Sensory Neurons Rely on Oxidative Phosphorylation and Glycolytic Processing of Extracellular Glucose. J Neurosci. 2017;37(23):5736-43. Epub 2017/05/14. PubMed PMID: 28500222; PubMed Central PMCID: PMCPMC6596473. [CrossRef]
- Acevedo C, Blanchard K, Bacigalupo J, Vergara C. Possible ATP trafficking by ATP-shuttles in the olfactory cilia and glucose transfer across the olfactory mucosa. FEBS Lett. 2019;593(6):601-10. Epub 2019/02/26. [CrossRef] [PubMed]
- Villar PS, Vergara C, Bacigalupo J. Energy sources that fuel metabolic processes in protruding finger-like organelles. FEBS J. 2021;288(12):3799-812. Epub 2020/11/04. [CrossRef] [PubMed]
- Krishnan S, Nordqvist H, Ambikan AT, Gupta S, Sperk M, Svensson-Akusjarvi S, et al. Metabolic Perturbation Associated With COVID-19 Disease Severity and SARS-CoV-2 Replication. Mol Cell Proteomics. 2021;20:100159. Epub 2021/10/08. PubMed PMID: 34619366; PubMed Central PMCID: PMCPMC8490130. [CrossRef]
- Khan M, Yoo SJ, Clijsters M, Backaert W, Vanstapel A, Speleman K, et al. Visualizing in deceased COVID-19 patients how SARS-CoV-2 attacks the respiratory and olfactory mucosae but spares the olfactory bulb. Cell. 2021;184(24):5932-49 e15. Epub 2021/11/20. PubMed PMID: 34798069; PubMed Central PMCID: PMCPMC8564600. [CrossRef]
- Seehusen F, Clark JJ, Sharma P, Bentley EG, Kirby A, Subramaniam K, et al. Neuroinvasion and Neurotropism by SARS-CoV-2 Variants in the K18-hACE2 Mouse. Viruses. 2022;14(5). Epub 2022/05/29. PubMed PMID: 35632761; PubMed Central PMCID: PMCPMC9146514. [CrossRef]
- Zheng J, Wong LR, Li K, Verma AK, Ortiz ME, Wohlford-Lenane C, et al. COVID-19 treatments and pathogenesis including anosmia in K18-hACE2 mice. Nature. 2021;589(7843):603-7. Epub 2020/11/10. PubMed PMID: 33166988; PubMed Central PMCID: PMCPMC7855185. [CrossRef]
- Davies J, Randeva HS, Chatha K, Hall M, Spandidos DA, Karteris E, et al. Neuropilin-1 as a new potential SARS-CoV-2 infection mediator implicated in the neurologic features and central nervous system involvement of COVID-19. Mol Med Rep. 2020;22(5):4221-6. Epub 2020/10/02. PubMed PMID: 33000221; PubMed Central PMCID: PMCPMC7533503. [CrossRef]
- Kang YL, Chou YY, Rothlauf PW, Liu Z, Soh TK, Cureton D, et al. Inhibition of PIKfyve kinase prevents infection by Zaire ebolavirus and SARS-CoV-2. Proc Natl Acad Sci U S A. 2020;117(34):20803-13. Epub 2020/08/09. PubMed PMID: 32764148; PubMed Central PMCID: PMCPMC7456157. [CrossRef]
- Mayi BS, Leibowitz JA, Woods AT, Ammon KA, Liu AE, Raja A. The role of Neuropilin-1 in COVID-19. PLoS Pathog. 2021;17(1):e1009153. Epub 2021/01/05. PubMed PMID: 33395426; PubMed Central PMCID: PMCPMC7781380. [CrossRef]
- Cantuti-Castelvetri L, Ojha R, Pedro LD, Djannatian M, Franz J, Kuivanen S, et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science. 2020;370(6518):856-60. Epub 2020/10/22. PubMed PMID: 33082293; PubMed Central PMCID: PMCPMC7857391. [CrossRef]
- Daly JL, Simonetti B, Klein K, Chen KE, Williamson MK, Anton-Plagaro C, et al. Neuropilin-1 is a host factor for SARS-CoV-2 infection. Science. 2020;370(6518):861-5. Epub 2020/10/22. PubMed PMID: 33082294; PubMed Central PMCID: PMCPMC7612957. [CrossRef]
Figure 1.
Functional anatomy and structure of the human olfactory system. (a) The human olfactory system contains multiple organs, including the olfactory epithelium (OE), the olfactory tract (OT). Odorants enter the nasal cavity (NC) and access the OE during inhalation, by being pumped through the sasal conchae. (b) Sensory neurons of the OE project to the olfactory bulb (OB) (taken and modified from https://sites.google.com/a/edmail.edcc.edu/savannatest/literature-research/pheromones-and-neurocircuits) (Accessed on November 18, 2022).
Figure 1.
Functional anatomy and structure of the human olfactory system. (a) The human olfactory system contains multiple organs, including the olfactory epithelium (OE), the olfactory tract (OT). Odorants enter the nasal cavity (NC) and access the OE during inhalation, by being pumped through the sasal conchae. (b) Sensory neurons of the OE project to the olfactory bulb (OB) (taken and modified from https://sites.google.com/a/edmail.edcc.edu/savannatest/literature-research/pheromones-and-neurocircuits) (Accessed on November 18, 2022).
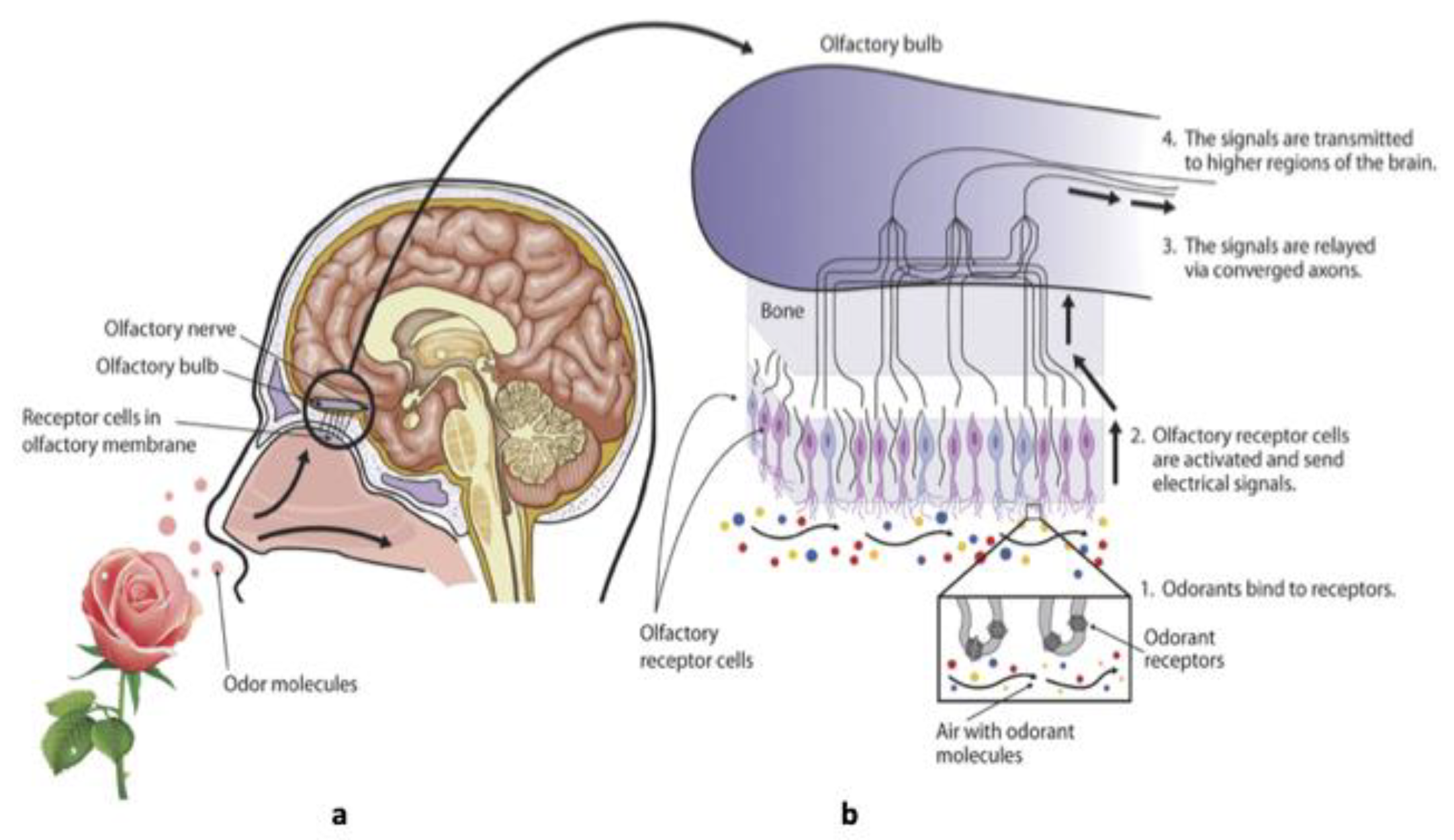
Figure 2.
Summary of how SARS-CoV-2 infected support cells induce damage to the mucus and cilia that prevent the olfactory receptor neurons to bind to their odorant molecules.
Figure 2.
Summary of how SARS-CoV-2 infected support cells induce damage to the mucus and cilia that prevent the olfactory receptor neurons to bind to their odorant molecules.
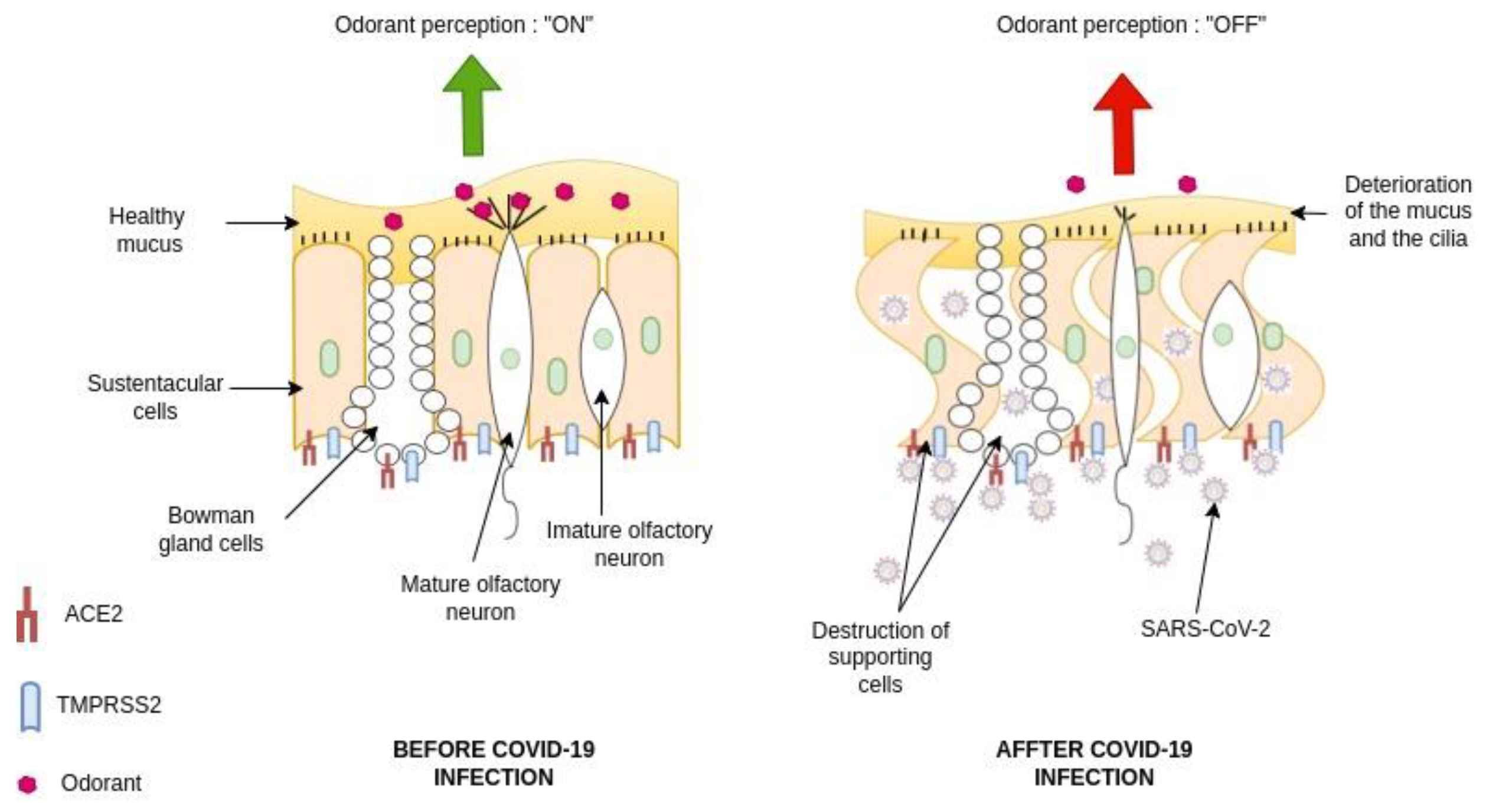

Table 1.
Summary of the mechanisms of SARS-CoV-2-induced olfactory dysfunction.
| Effect of SARS-CoV-2 Infection on Cells in the OE | References |
|
[102] [103] [88] [104] [105] [106] [89] [107] |
|
[108] [109] [110] [110] [85] [111] [112] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



