
Submitted:
11 May 2023
Posted:
12 May 2023
You are already at the latest version
Abstract
Cerebral ischemic stroke is one of the leading causes of death and disability worldwide. 2’-fucosyllactose (2’-FL), a human milk oligosaccharide, has anti-inflammatory effects and plays a protective role in arterial thrombosis, but its role in ischemic stroke remains unclear. This study aimed to investigate the neuroprotective effects of 2’-FL and its potential mechanisms in a mouse model of ischemic stroke. The administration of 2’-FL led to a reduction in the size of cerebral infarct and reactive oxygen species (ROS)-related products in the brain of MCAO mice. Further, 2’-FL promotes the recovery of neurological deficits and motor function in neurological scores and behavior tests. Biochemical studies showed that 2’-FL reduced upregulated IL-10 and downregu-lated TNF-α. Additionally,2’-FL enhanced M2-type microglial polar2’-FL upregulated IL-10 and downregulated TNF-α ization at 7 days after MCAO, with an increase in IL-4 levels and STAT6 activation at 3 days after MCAO. Our data showed that 2’-FL reduced the neurological symptoms of ischemic stroke and ROS accumulation in the brain through IL-4/STAT6-dependent M2-type mi-croglial polarization in MCAO mice. These results demonstrate that 2’-FL is a potentially effective therapeutic agent for ischemic stroke.
Keywords:
2’-fucosyllactose
; microglia polarization
; anti-inflammation
; reactive oxygen species
; ischemic injury
1. Introduction
Cerebral ischemic stroke is one of the leading causes of disability and mortality worldwide and represents a significant health burden in both developed and developing countries [1]. The initial injury in a stroke occurs within minutes when blood flow to the injured focal core of the brain is limited, resulting in a significant reduction in oxygen and glucose delivery to neurons [2]. The ischemia-reperfusion injury process accelerates neuronal cell death due to energy depletion, inducing various postischemic responses that exacerbate brain damage and trigger pathological events. These include an increase in reactive oxygen species (ROS), disruption of the blood–brain barrier (BBB), activation of apo-necrotic cell death, excitotoxicity, and excessive production of inflammatory mediators [3]. The brain responds to neuronal damage caused by ischemia-reperfusion injury by activating various anti-inflammatory responses to confer neuroprotection. Accordingly, the use of anti-inflammatory agents may be a promising therapeutic strategy for cerebral ischemic injury [4,5].
Glial cells, particularly microglia, play a major role in the immunological response following an ischemic stroke [6]. Microglia undergo rapid morphological changes, polarizing into pro- or anti-inflammatory phenotypes and thus steering the course of degeneration or eventual recovery following stroke injury [7]. M1 activation produces the inflammatory cytokines tumor necrosis factor-alpha (TNF-α) and interleukin (IL)-6 as well as ROS. M2 activation produces the anti-inflammatory cytokines IL-10 and transforming growth factor (TGF)-β, along with other growth factors [8,9]. Microglial polarization and modulation of microglial phenotypes from M1 to M2 have been found to promote brain repair [10] and treat ischemic stroke [11].
Human breast milk is an excellent source of nutrition and contains various essential bioactive ingredients, such as human milk oligosaccharides (HMOs), which are its third most abundant solid component [12]. Studies have shown that HMOs, particularly 2’-FL, are more than a prebiotic and have various functions, including immune system modulation, anti-inflammation, and cognition benefits [13,14,15,16]. However, evidence of these promising anti-inflammatory and neuroprotective effects has not been demonstrated in ischemia-reperfusion injury. Hence, this study aimed to investigate the protective effects of 2’-FL against ischemic stroke via modulation of microglial polarization.
2. Materials and Methods
2.1. Reagents
2’-FL was obtained from Advanced Protein Technologies Corp. (Suwon, Korea). Antibodies against-β-actin, NQO1, STAT6, and phospho-STAT6 (pSTAT6) were obtained from Santa Cruz Biotechnology (Dallas, TX, USA) and antibody against-PGC1-α was obtained from Novus Biologicals (Centennial, CO, USA). The antibodies against inducible nitric oxide synthase (iNOS) and phospho-AMPKα (pAMPKα) was obtained from Cell Signaling (Danvers, MA, USA), and antibodies against CD206 and Iba1 were obtained from Abcam (Cambridge, UK), and , . The antibody against-CD16/32; enzyme-linked immunosorbent assay (ELISA) kit for interleukin (IL)-10, IL-4, and TNF-α; and radioimmunoprecipitation assay (RIPA) lysis buffer were obtained from Thermo Fisher (Waltham, MA, USA). Horseradish peroxidase (HRP)-conjugated secondary antibodies were obtained from Bethyl (Farmingdale, NY, USA), and enhanced chemiluminescence (ECL) solution was purchased from Pierce (Rockford, IL, USA). ELISA kits for catalase and superoxide dismutase (SOD) were obtained from Enzo Life Sciences, Inc. (Farmingdale, NY, USA).
2.2. Animals
Male mice (C57BL/6, 6 weeks old, 17–18 g, BW) were obtained from DooYeol Biotech (Seoul, Korea) and housed at a constant temperature (21°C ± 2°C) and humidity with a 12-h light/dark cycle. Mice were then divided randomly into 5 groups of 44 animals each: (1) control group (water [orally administered], n = 8), (2) vehicle group (water [orally administered], n = 18), and (3) 2’-FL treated group (1g/kg BW) in water [orally administered], n = 18). Focal cerebral ischemic injury was induced in mice in the vehicle and 2’-FL treated groups through surgery. The Institute of Animal Care and Use Committee of the Korea Institute of Oriental Medicine approved all experiments, which were performed according to their guidelines (approval number. KIOM-D-20-077).
2.3. Middle Cerebral Artery Occlusion (MCAO) Model
To induce focal cerebral ischemia, an MCAO model was established using the intraluminal filament technique. Mice were anesthetized with 2.0% isoflurane to minimize their suffering. A silicon-coated 7-0 monofilament (L.M.S. Korea Inc., Gyeonggi, Korea) was inserted through the left internal carotid artery to occlude the middle cerebral artery. The inserted filament was withdrawn 45 min after MCAO, followed by reperfusion. The mice were kept warm to maintain their body temperature at 37ºC throughout the procedure. Subsequently, they were monitored until they woke up from anesthesia, after which they were returned to their cages. The survival rate of this MCAO surgery was approximately 85%.
2.4. Drug Administration
2’-FL (1 g/kg BW) was administered orally for 7 days prior to surgery, and then for 3 or 7 days beginning from the first day after MCAO. Control and vehicle groups received only water (administered orally).
2.5. Behavioral Experiments
2.5.1. Neurological Function Evaluation
Twenty-four hours after ischemic injury, neurological deficits were evaluated on a five-point scale [17] as follows: 0—no deficit; 1—failure to fully extend left forepaw; 2—resistance to contralateral pressure without turning and with forepaw buckling; 3—resistance to lateral pressure with left turning and forepaw flexion; and 4—no spontaneous walking, with depressed level of consciousness. The inclusion criterion of the model was a neurological function score of 1–3, and the exclusion criterion was a neurological function score of 0 or 4.
2.5.2. Corner Test
To test for sensorimotor dysfunction, we performed the corner test on days 3 and 7 after MCAO surgery. Mice suffering from MCAO-induced brain damage have functional deficits on the contralateral side [18] and therefore use their ipsilateral paws more often when turning corners. To assess this, two boards (30 cm × 20 cm) were attached at an angle of 30°, with one side left open. The direction in which the body of the mouse was raised at the inside corner was assessed. This procedure was repeated 10 times for each animal, and its number of turns to the ipsilateral side was calculated as a percentage.
2.5.3. Wire Grip Test
The wire grip test was performed on days 3 and 7 after MCAO surgery to monitor motor balance and grip strength. Mice were suspended, allowing them to hold on to a single wire 50 cm above the ground [19]. Performance was measured as follows: 0—not able to grip the wire; 1—hangs on to wire with one or two forepaws; 2—attempts to climb on the wire while holding it with a forepaw; 3—grasps the wire with one or both hind paws, in addition to forepaws; 4—wraps the tail around the wire while holding it with all four paws; and 5—reaches one end of the wire and escapes from the apparatus. Time to fall off the wire was also scored: 0—unable to hold on, falls in under 1 s; 1—holds for 1–9 s; 2—holds for 10–19 s; 3—holds for 20–29 s; and 4—holds wire for >30 s. The total score was presented as the sum of the performance and hanging time scores.
2.6. Nissl Staining
Mouse brains were removed, and tissue was fixed and frozen in Optimal cutting temperature compound. Subsequently, 30-μm-thick sections were cut and mounted on slides. To evaluate brain edema, the sections were stained with 0.1% cresyl violet for 5 min. The sizes of subareas including the striatum, cortex, and hippocampus on both contralateral and ipsilateral sides were measured using ImageJ 1.52v software. Brain damage volume was expressed as a percentage: atrophy volume = subarea volume of the ipsilateral hemisphere/total volume of brain × 100.
2.7. Reverse Transcription–Polymerase Chain Reaction (RT-PCR)
Total RNA from the ipsilateral side of brain tissues was extracted using the RNeasy Mini RNA isolation kit (Qiagen, Chatsworth, CA) according to the manufacturer's instructions and then 1 µg RNA was used to synthesize cDNA by using Omniscript Reverse Transcriptase (Qiagen). SYBR green-based quantitative PCR amplification was performed using the QuantStudio 6 Flex Real-time PCR System (Thermo Scientific). The expression levels were calculated with the 2–ΔΔCt method and normalized by gapdh expression level. Primers used to amplify the genes of interest are listed in Table 1.
2.8. Enzyme-Linked Immunoassay (ELISA)
Contralateral and ipsilateral cerebral tissues were homogenized and protein contents were collected. We evaluated catalase (ADI-907-027), SOD (ADI-SOD-157), IL-10 (#88-7105-88), TNF-α (#88-7323-24), and IL-4 (#BMS613) levels using ELISA kits according to the manufacturer’s protocol.
2.9. Western Blot
Total brain tissue was homogenized in RIPA lysis buffer containing protease and phosphatase inhibitors. An equal amount of protein (20 ug) was loaded into each well of a sodium dodecyl sulfate–polyacrylamide gel electrophoresis gel and then transferred to a PVDF polyvinylidene fluoride membrane. The membranes were blocked and incubated with the following primary antibodies: anti-β-actin (1:1000, sc-47778), NQO1 (1:1000, sc-376023), HO-1 (1:1000, #82206, 1:1000), iNOS (1:1000, #2982), Iba1 (1:1000, ab178846), CD16/32 (1:1000, #14-0161-82), CD206 (1:1000, ab64693), pAMPKα (1:1000, #2531), PGC1-α (1:1000, NBP1-04676), pSTAT6 (1:1000, sc-136019), and STAT6 (1:1000, sc-374021) at 4°C overnight. The next day, the membranes were washed and incubated with HRP-conjugated antibodies (A90-137P or A120-108P, 1:4000) for 1 h and developed with an ECL solution. Immunoreactivity was recorded using a digital imaging system (Q9 Alliance; UVITEC Ltd., England, UK). The results were quantified using ImageJ 1.52v software and normalized to β-actin.
2.10. Immunofluorescence (IF) Staining
Frozen sections were incubated with a blocking buffer for 1 h. Sections were incubated with primary antibodies overnight in a dilution buffer at 4°C. The following antibodies were used: Iba1 (1:500, #019-19741), CD206 (1:500), and pSTAT6 (1:500). The sections were then incubated with a fluorescent secondary antibody (Vector Laboratories, Inc., Burlingame, CA, USA) for 2 h in the dark. After mounting tissue on slides with a mounting medium (Vector Laboratories, Inc.), images were captured using a fluorescence microscope (Lion Heart FX; Agilent, Santa Clara, CA, USA). Iba1 density was measured using ImageJ 1.52v software, and CD206+/Iba1+ cells in the infarction region of the ipsilateral side were counted.
2.11. Statistical Analysis
Data analysis was performed using GraphPad Prism 5. Statistical significance was assessed using Bonferroni’s test (for one-way ANOVA) for comparisons of multiple groups. p-value of < 0.05 was considered statistically significant.
3. Results
3.1. 2'-FL Modulates the Polarization of Microglia in Ischemic Stroke
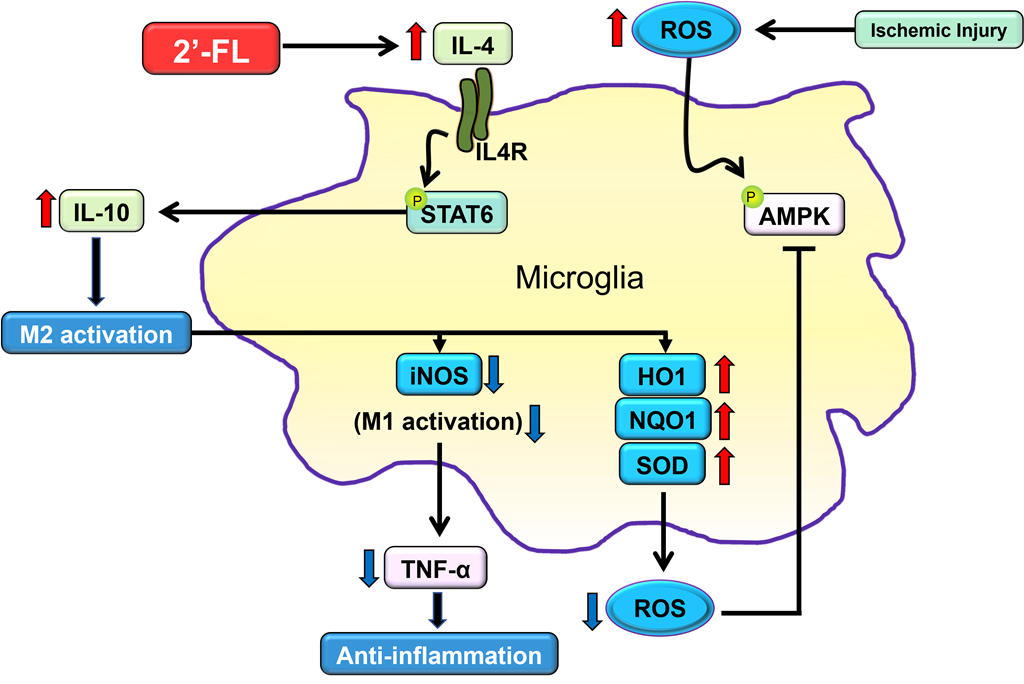
A study showed that M1 microglia markers exhibited an increasing trend during 14 days after ischemic stroke, and M2 markers increased from day 1 to day 7 and decreased after that time [10]. To investigate the possible modulation of M1/M2-type microglia by 2’-FL, we examined marker lists for genes associated with M1-type (CD32, iNOS and CD11b) and M2-type (Arg1, CCL-22, and TGF-β) microglia and then quantified these genes in our model using RT-PCR (Figure 1). We found that gene expression of the M1-type in MCAO mice increased on day 3 compared to 2’-FL-treated mice (Figure 1b). 2’-FL treatment increased M2-type gene expression for 1 to 7 days compared with untreated MCAO mice (vehicle) (Figure 1c). These data indicate that 2-’FL increases the expression of M2-type genes and maintains M2 activation for 3 to 7 days after ischemic stroke.
3.2. 2’FL Abrogates Neurological Deficits and Motor Function in Ischemic Stroke
We next investigated how functional recovery in the mouse brain was affected by 2’-FL administration on days 3 and 7 after MCAO. Mice were given a neurological score and subjected to corner and wire grip tests to analyze their functional recovery (Figure 2a). 2’-FL improved neurological score at 3 and 7 days after MCAO (Figure 2b). In the corner test, mice in the vehicle group demonstrated significantly increased ipsilateral turning at 3 and 7 days post-MCAO, while 2-’FL treatment significantly reduced ipsilateral turning on day 7 (Figure 2c). In the wire grip test, the vehicle group showed a reduced time to fall at both 3 and 7 days post-MCAO compared to the control group. However, the 2’-FL-treated group demonstrated a significantly longer wire grip test time compared with the vehicle group on both days 3 and 7 (Figure 2d). The qualitative score by wire grip test was also significantly improved in the 2’-FL-treated group at 7 days after MCAO compared with the vehicle group (Figure 2e). These data indicate that 2’-FL improves neurological and behavioral function after ischemic stroke.
3.3. 2’-FL Reduces Brain Damage in Ischemic Stroke
To investigate the effects of 2’-FL on histological changes in the brain after MCAO, we examined atrophy and damage in the frontal cortex, striatum, and hippocampus using nissl staining. We observed edema in the brains of MCAO mice treated with either vehicle or 2’-FL at 3 days after MCAO. However, we found that 2’-FL markedly reduced atrophy and damage in the brain on day 7 after MCAO (Figure 2f). These data indicate that the administration of 2’-FL reduced brain damage at 7 days after ischemic stroke.
3.4. 2’-FL Reduces ROS Generation in Ischemic Stroke
To examine the effect of 2’-FL on ROS generation, we investigated the levels of catalase and SOD in the brain after MCAO. We observed that catalase activity was induced by ischemic injury, and 2’-FL treatment significantly increased catalase levels compared with the vehicle group at 7 days after MCAO (Figure 3a). On the other hand, the level of SOD was reduced 3 days after ischemic injury, and 2’-FL treatment increased this level. At 7 days after MCAO, 2’-FL treatment markedly increased SOD levels compared with the control and vehicle groups (Figure 3b). To further test antioxidant protein expression modulation by 2’-FL treatment, we determined the expression levels of HO-1, NQO1, and iNOS using western blot analysis. We found increased expression levels of HO-1 and decreased levels of iNOS at 3 days after MCAO in both the vehicle and 2’-FL-treated groups. At 7 days, the 2’-FL-treated group showed markedly increased HO-1 and further decreased iNOS expression compared with the vehicle-treated group (Figure 3c–f). Interestingly, the expression of NQO1 was markedly increased in the 2’-FL-treated group at 7 days after MCAO, but not at 3 days (Figure 3e). These data indicate that 2’-FL administration reduced ROS generation in the brain at 7 days after ischemic stroke.
3.5. 2’-FL Regulates Cytokines Related to Inflammation and Microglial Activation in Ischemic Stroke
Because microglial polarization to the M2 type promotes brain repair and confers a beneficial effect after ischemic stroke [10,20], we investigated whether 2’-FL regulates IL-10 and TNF-α secretion using ELISA. IL-10 levels were decreased by MCAO, but 2’-FL-treated MCAO mice demonstrated significantly higher IL-10 levels than vehicle-treated MCAO mice at 7 days (Figure 4a). TNF-α levels in MCAO mice treated with 2’-FL at 3 and 7 days were reduced compared with those in vehicle-treated mice (Figure 4b). Further, we investigated the expression levels of Iba1, CD32, and CD206 proteins using western blot analysis to confirm M1/M2 microglial polarization. The expression level of Iba1 at 7 days post-MCAO in the vehicle and 2’-FL groups was significantly increased compared with the control, but there was no significant increase at 3 days (Figure 4d). We found that ischemic injury induced an increased level of CD32 expression at 3 days. At 7 days, 2’-FL treatment decreased the CD32 expression level compared with the vehicle (Figure 4e). Furthermore, CD206 expression in the 2’-FL-treated group increased compared with the vehicle group at 7 days after MCAO but was unchanged at 3 days (Figure 4f). To confirm whether 2’-FL activates M2-type microglia in the damaged brain region, we explored M2 activation in the infarct region (striatum and cortex, ipsilateral side) after MCAO using IF staining (Figure 4g). We found that ischemic injury induced an increase in the intensity of density (IOD) in Iba1+ cells on the ipsilateral side, and after 7 days, the IOD of Iba1+ cells in the vehicle- and 2’-FL-treated groups was significantly increased compared with control (Figure 4h). The number of Iba1+/CD206+ cells in both the vehicle- and 2’-FL-treated groups increased 3 days after MCAO; however, at 7 days, the number of Iba1+/CD206+ cells in the 2’-FL-treated group was significantly higher than that in the vehicle group (Figure 4i). These data suggest that 2’-FL administration maintains M2 microglial activation for 7 days after ischemic stroke.
3.6. 2’-FL Enhances M2 Type Microglia via IL-4 Induction and STAT6 Activation after MCAO
Microglial polarization to the M2 type is associated with the STAT6 pathway and PGC-1α expression. AMPK phosphorylates PGC-1α directly after ischemic injury [10,21]. We explored the expression of pSTAT6, PGC-1α, and pAMPK proteins. The phosphorylation level of STAT6 was increased in the 2’-FL-treated group compared with the vehicle group at 3 days after MCAO but nullified at 7 days (Figure 5b). Expression of PGC-1α did not change among groups, but the expression of AMPK was markedly decreased by 2’-FL at 7 days after MCAO (Figure 5c,d). To confirm pSTAT6 modulation by 2’-FL in the brain, we performed the fluorescence microscopic image in the ipsilateral striatum after MCAO. We found that the 2’-FL-treated group had a significantly increased number of Iba1+/pSTAT6+ cells compared with the vehicle group on the ipsilateral side (Figure 5e,f). IL-4 switches modulate the STAT6 pathway [22]; therefore, we assessed IL-4 levels in the brain by ELISA. We found that 2’-FL treatment significantly increased IL-4 levels 3 days after MCAO compared with the vehicle group but not at 7 days (Figure 5g). These data suggest that 2’-FL administration increased IL-4 levels and STAT6 phosphorylation at 3 days after ischemic stroke and led to a decrease in AMPK activation at 7 days.
This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
4. Discussion
Stroke is one of the leading causes of death and disability worldwide. The main cause of stroke is the occlusion or rupture of the brain's blood vessels, which causes oxygen deprivation in brain cells [23]. The progression of cerebral ischemic disease is greatly influenced by inflammation. Moreover, cerebral ischemia is accompanied by increased serum concentrations of oxidative stress markers [24]. Hence, anti-inflammatory and antioxidant agents are promising therapeutic targets for reducing secondary damage [25,26]. At present, the thrombolytic recombinant tissue plasminogen activator (rtPA) is the only FDA-approved therapy for acute ischemic stroke that can effectively diminish infarct size and improve functional recovery. However, rtPA has several disadvantages, including the potential risk of hemorrhagic transformation, a limited therapeutic time window after stroke onset, and limited efficacy. Consequently, it is still difficult to provide safe and effective therapies for ischemic stroke, especially at an early stage [27]. Therefore, there is a need to research and identify therapeutic medications or alternatives that are beneficial in promoting functional recovery or quality of life following a stroke.
With an improved understanding of the mechanism underlying stroke, current research has focused on the duality of post-stroke inflammation, which is a significant contributing factor to the pathogenic process of stroke [28] and plays an important role in brain tissue damage and repair [29,30]. The enhancement of microglial activation is a potential target for repairing ischemia, and studies on the modification of the microglial phenotype to promote healing processes may help identify new therapeutic targets [31,32]. Particularly, M2 microglia produce anti-inflammatory cytokines, such as IL-10, thereby promoting phagocytosis of cell debris, tissue repair, and neuron survival [32,33]. A previous study demonstrated that HMO treatment elevated the production of IL-10 and decreased that of TNF-α in mouse mast cells, which alleviated the symptoms of food allergy [34]. Additionally, our previous study showed that 2’-FL attenuates platelet activation and reduces stroke-induced infarct volume [35]. Based on these studies, we investigated whether 2’-FL could reduce ischemic brain injury-induced inflammation by increasing M2-type microglial activation. We found that 2’-FL significantly diminished brain infarct size and improved functional recovery. We also demonstrated that treatment with 2'-FL significantly increased M2 microglial activation and the expression of markers such as CD206 in a mouse model of ischemic damage (Figure 4).
A previous study has shown that an increase in the harmful accumulation of ROS caused by hypoxia activates AMPK owing to an increase in the AMP/ATP ratio [36]. Similarly, we observed that treatment with 2'-FL resulted in sustained M2 polarization after 3 days, which significantly decreased ROS formation. In turn, AMPK activation was markedly reduced at 7 days. Further, the expression of the antioxidant proteins HO-1 and NQO-1 was substantially increased at 3 and 7 days after ischemic injury, whereas the expression of iNOS, a marker for M1 microglia, did not (Figure 3). Therefore, we anticipated that 2'-FL treatment would reduce M1 microglial activation at 7 days as a result of its continued support of M2 activation under ischemic conditions. Further, we found that 2’-FL induced STAT6 activation and IL-10 levels in the brain under ischemic injury (Figure 4, 5). Many bioactive components possess neuroprotective and anti-inflammatory properties, and most of these substances can alter the phenotype of microglia by activating them in a STAT6-dependent manner. For example, PGC-1α interacts with PPAR-γ, which is involved in IL-4-induced M2 polarization [9]. It has been demonstrated that resveratrol, a Sirt1 agonist, can change microglia from M1 to M2 by upregulating PGC-1α expression and activating the STAT6 pathway. Furthermore, PGC-1α is directly phosphorylated by AMPK [21]. Interestingly, our findings demonstrated that 2’-FL only elevated phosphorylation of STAT6 in the early stage (3 days after ischemic injury), without associated changes in PGC-1α (Figure 5b,c), suggesting that STAT6 activation induced by 2’-FL treatment was followed by a decrease in AMPK activity.
M2 macrophage activation is subclassified into M2a, M2b, and M2c. The M2a contributes to the repair of damaged tissue by expressing anti-inflammatory and neurotrophic factors [37,38]. The M2a state can be induced by IL-4 and is associated with tissue repair and phagocytosis. IL-4 activates STAT6 leading to the transcription of M2a-associated genes, including CD206. Interestingly, microglia cell morphology within the ischemic injury showed larger cell and soma area, with shorter branching [37]. However, 2’-FL treatment resulted in cells with a smaller soma area and higher branching (Figure 6). We, therefore, propose that 2’-FL treatment activates M2a microglia via IL-4–STAT6 signaling.
One limitation of our study is the inability to determine whether 2’-FL promotes IL-4 production directly or indirectly in the brain after ischemia. A previous study demonstrated that HMO treatment increased the levels of IL-10 and IL-6, but not TNF-α, in dendritic cells, which modulate the immune system via the promotion of T-cells [39]. This suggests that T-cells activated by 2’-FL may penetrate the brain and secrete IL-4. Additionally, our data showed that 2’-FL upregulated antioxidant defenses such as SOD and HO-1 at an early time point after ischemic injury and later induced an increase in catalase and NQO-1 levels (Figure 3e). This indicates that 2’-FL may have anti-inflammatory and antioxidant effects in the brain independent of IL-4 induction. The exact signaling mechanism and action of 2’-FL in the ischemic injury of the adult brain will be investigated in further studies.
5. Conclusions
Our study showed that 2'-FL reduces ROS levels to protect against inflammation and oxidative stress caused by ischemic damage. Further, treatment with 2'-FL maintained M2 microglial polarization through IL-4-induced STAT6 activation and upregulation of IL-10, which is crucial for the resolution of brain edema and recovery of motor behavior. Taken together, these data suggest that 2’-FL has the potential to be an effective therapeutic agent for ischemic stroke patients. Additionally, daily supplements of 2’-FL may show potential as a viable therapeutic agent for such diseases.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, methodology, K.K. and M.E.P.; investigation, M.E.P., Y.K., and H.K.; software, validation, formal analysis and visualization, M.E.P. and Y.K.; writing—original draft preparation, M.E.P, S.J. and K.K.; supervision, C.S.S., J.Y., S.J., and Y.S.; project administration, C.S.S., J.Y., S.J., and Y.S.; funding acquisition, K.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Korea Institute of Oriental Medicine Grant (ERT2011170) from Advanced Protein Technologies Corp. and the National Research Foundation of Korea Grant (NRF-2021R1I1A2040301) from the Korean Government.
Institutional Review Board Statement
All animal experimental procedures were approved by the Korea Institute of Oriental Medicine Institutional Animal Care and Use Committee (KIOM-D-20-077) and were conducted in accordance with the guidelines of the National Institutes of Health (NIH publication #83-23, revised in 1985).
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data supporting the results were shown in the paper, and can be applicable from the corresponding author.
Acknowledgments
We would like to thank Enago (http://www.enago.co.kr) for the English language review.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Peng, T.; Jiang, Y.; Farhan, M.; Lazarovici, P.; Chen, L.; Zheng, W. Anti-inflammatory Effects of Traditional Chinese Medicines on Preclinical in vivo Models of Brain Ischemia-Reperfusion-Injury: Prospects for Neuroprotective Drug Discovery and Therapy. Front. Pharmacol. 2019, 10, 204. [Google Scholar] [CrossRef] [PubMed]
- Doyle, K.P.; Simon, R.P.; Stenzel-Poore, M.P. Mechanisms of ischemic brain damage. Neuropharmacology 2008, 55, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Lahiani, A.; Brand-Yavin, A.; Yavin, E.; Lazarovici, P. Neuroprotective Effects of Bioactive Compounds and MAPK Pathway Modulation in "Ischemia"-Stressed PC12 Pheochromocytoma Cells. Brain Sci. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Reekmans, K.; Praet, J.; Daans, J.; Reumers, V.; Pauwels, P.; Van der Linden, A.; Berneman, Z.N.; Ponsaerts, P. Current challenges for the advancement of neural stem cell biology and transplantation research. Stem Cell Rev. Rep. 2012, 8, 262–278. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, M.; Shiba, T.; Hasebe, S.; Ogita, K. Adult neurogenesis is regulated by endogenous factors produced during neurodegeneration. J. Pharmacol. Sci. 2011, 115, 425–432. [Google Scholar] [CrossRef]
- Iadecola, C.; Anrather, J. Stroke research at a crossroad: asking the brain for directions. Nat. Neurosci. 2011, 14, 1363–1368. [Google Scholar] [CrossRef]
- Lyu, J.; Xie, D.; Bhatia, T.N.; Leak, R.K.; Hu, X.; Jiang, X. Microglial/Macrophage polarization and function in brain injury and repair after stroke. CNS Neurosci. Ther. 2021, 27, 515–527. [Google Scholar] [CrossRef]
- Colonna, M.; Butovsky, O. Microglia Function in the Central Nervous System During Health and Neurodegeneration. Annu. Rev. Immunol. 2017, 35, 441–468. [Google Scholar] [CrossRef]
- Guo, S.; Wang, H.; Yin, Y. Microglia Polarization From M1 to M2 in Neurodegenerative Diseases. Front. Aging Neurosci. 2022, 14, 815347. [Google Scholar] [CrossRef]
- Jiang, C.T.; Wu, W.F.; Deng, Y.H.; Ge, J.W. Modulators of microglia activation and polarization in ischemic stroke (Review). Mol. Med. Rep. 2020, 21, 2006–2018. [Google Scholar] [CrossRef]
- Wang, J.; Xing, H.; Wan, L.; Jiang, X.; Wang, C.; Wu, Y. Treatment targets for M2 microglia polarization in ischemic stroke. Biomed. Pharmacother. 2018, 105, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Mosca, F.; Giannì, M.L. Human milk: composition and health benefits. Pediatr. Med. Chir. 2017, 39, 155. [Google Scholar] [CrossRef] [PubMed]
- Weichert, S.; Koromyslova, A.; Singh, B.K.; Hansman, S.; Jennewein, S.; Schroten, H.; Hansman, G.S. Structural Basis for Norovirus Inhibition by Human Milk Oligosaccharides. J. Virol. 2016, 90, 4843–4848. [Google Scholar] [CrossRef] [PubMed]
- Oliveros, E.; Ramirez, M.; Vazquez, E.; Barranco, A.; Gruart, A.; Delgado-Garcia, J.M.; Buck, R.; Rueda, R.; Martin, M.J. Oral supplementation of 2'-fucosyllactose during lactation improves memory and learning in rats. J. Nutr. Biochem. 2016, 31, 20–27. [Google Scholar] [CrossRef]
- He, Y.; Liu, S.; Kling, D.E.; Leone, S.; Lawlor, N.T.; Huang, Y.; Feinberg, S.B.; Hill, D.R.; Newburg, D.S. The human milk oligosaccharide 2'-fucosyllactose modulates CD14 expression in human enterocytes, thereby attenuating LPS-induced inflammation. Gut 2016, 65, 33–46. [Google Scholar] [CrossRef]
- Mezoff, E.A.; Hawkins, J.A.; Ollberding, N.J.; Karns, R.; Morrow, A.L.; Helmrath, M.A. The human milk oligosaccharide 2'-fucosyllactose augments the adaptive response to extensive intestinal. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G427–438. [Google Scholar] [CrossRef]
- Bederson, J.B.; Pitts, L.H.; Tsuji, M.; Nishimura, M.C.; Davis, R.L.; Bartkowski, H. Rat middle cerebral artery occlusion: evaluation of the model and development of a neurologic examination. Stroke 1986, 17, 472–476. [Google Scholar] [CrossRef]
- Kim, Y.R.; Kim, H.N.; Ahn, S.M.; Choi, Y.H.; Shin, H.K.; Choi, B.T. Electroacupuncture promotes post-stroke functional recovery via enhancing endogenous neurogenesis in mouse focal cerebral ischemia. PLoS One 2014, 9, e90000. [Google Scholar] [CrossRef]
- Balkaya, M.; Krober, J.M.; Rex, A.; Endres, M. Assessing post-stroke behavior in mouse models of focal ischemia. J. Cereb. Blood Flow Metab. 2013, 33, 330–338. [Google Scholar] [CrossRef]
- Liu, X.; Liu, J.; Zhao, S.; Zhang, H.; Cai, W.; Cai, M.; Ji, X.; Leak, R.K.; Gao, Y.; Chen, J.; et al. Interleukin-4 Is Essential for Microglia/Macrophage M2 Polarization and Long-Term Recovery After Cerebral Ischemia. Stroke 2016, 47, 498–504. [Google Scholar] [CrossRef]
- Yang, W.C.; Hwang, Y.S.; Chen, Y.Y.; Liu, C.L.; Shen, C.N.; Hong, W.H.; Lo, S.M.; Shen, C.R. Interleukin-4 Supports the Suppressive Immune Responses Elicited by Regulatory T Cells. Front. Immunol. 2017, 8, 1508. [Google Scholar] [CrossRef] [PubMed]
- Collaborators, G.B.D.S. Global, regional, and national burden of stroke and its risk factors, 1990-2019: a systematic analysis for the Global Burden of Disease Study 2019. Lancet Neurol. 2021, 20, 795–820. [Google Scholar] [CrossRef]
- Polidori, M.C.; Cherubini, A.; Stahl, W.; Senin, U.; Sies, H.; Mecocci, P. Plasma carotenoid and malondialdehyde levels in ischemic stroke patients: relationship to early outcome. Free Radic. Res. 2002, 36, 265–268. [Google Scholar] [CrossRef]
- McColl, B.W.; Rothwell, N.J.; Allan, S.M. Systemic inflammatory stimulus potentiates the acute phase and CXC chemokine responses to experimental stroke and exacerbates brain damage via interleukin-1- and neutrophil-dependent mechanisms. J. Neurosci. 2007, 27, 4403–4412. [Google Scholar] [CrossRef] [PubMed]
- Kawabori, M.; Yenari, M.A. Inflammatory responses in brain ischemia. Curr. Med. Chem. 2015, 22, 1258–1277. [Google Scholar] [CrossRef]
- Lakhan, S.E.; Kirchgessner, A.; Hofer, M. Inflammatory mechanisms in ischemic stroke: therapeutic approaches. J. Transl. Med. 2009, 7, 97. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Stetler, R.A.; Leak, R.K.; Shi, Y.; Li, Y.; Yu, W.; Bennett, M.V.L.; Chen, J. Oxidative stress and DNA damage after cerebral ischemia: Potential therapeutic targets to repair the genome and improve stroke recovery. Neuropharmacology 2018, 134, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Yong, H.Y.F.; Rawji, K.S.; Ghorbani, S.; Xue, M.; Yong, V.W. The benefits of neuroinflammation for the repair of the injured central nervous system. Cell Mol. Immunol. 2019, 16, 540–546. [Google Scholar] [CrossRef]
- Tobin, M.K.; Bonds, J.A.; Minshall, R.D.; Pelligrino, D.A.; Testai, F.D.; Lazarov, O. Neurogenesis and inflammation after ischemic stroke: what is known and where we go from here. J. Cereb. Blood Flow Metab. 2014, 34, 1573–1584. [Google Scholar] [CrossRef]
- Narantuya, D.; Nagai, A.; Sheikh, A.M.; Wakabayashi, K.; Shiota, Y.; Watanabe, T.; Masuda, J.; Kobayashi, S.; Kim, S.U.; Yamaguchi, S. Microglia transplantation attenuates white matter injury in rat chronic ischemia model via matrix metalloproteinase-2 inhibition. Brain Res. 2010, 1316, 145–152. [Google Scholar] [CrossRef]
- Taylor, R.A.; Sansing, L.H. Microglial responses after ischemic stroke and intracerebral hemorrhage. Clin. Dev. Immunol. 2013, 2013, 746068. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Li, T.; Li, J.; Yang, J.; Liu, H.; Zhang, X.J.; Le, W. Jmjd3 is essential for the epigenetic modulation of microglia phenotypes in the immune pathogenesis of Parkinson's disease. Cell Death Differ. 2014, 21, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Li, P.; Guo, Y.; Wang, H.; Leak, R.K.; Chen, S.; Gao, Y.; Chen, J. Microglia/macrophage polarization dynamics reveal novel mechanism of injury expansion after focal cerebral ischemia. Stroke 2012, 43, 3063–3070. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Courtade, L.; Han, S.; Lee, S.; Mian, F.M.; Buck, R.; Forsythe, P. Attenuation of food allergy symptoms following treatment with human milk oligosaccharides in a mouse model. Allergy 2015, 70, 1091–1102. [Google Scholar] [CrossRef]
- Pak, M.E.; Kim, Y.; Park, Y.J.; Go, Y.; Shin, C.S.; Yoon, J.; Jeon, S.; Song, Y.; Kim, K. Human milk oligosaccharide, 2′-Fucosyllactose, attenuates platelet activation in arterial thrombosis. J. Funct. Foods 2022, 94, 105138–105146. [Google Scholar] [CrossRef]
- Hinchy, E.C.; Gruszczyk, A.V.; Willows, R.; Navaratnam, N.; Hall, A.R.; Bates, G.; Bright, T.P.; Krieg, T.; Carling, D.; Murphy, M.P. Mitochondria-derived ROS activate AMP-activated protein kinase (AMPK) indirectly. J. Biol. Chem. 2018, 293, 17208–17217. [Google Scholar] [CrossRef]
- Lan, X.; Han, X.; Li, Q.; Yang, Q.W.; Wang, J. Modulators of microglial activation and polarization after intracerebral haemorrhage. Nat. Rev. Neurol. 2017, 13, 420–433. [Google Scholar] [CrossRef]
- Franco, R.; Fernandez-Suarez, D. Alternatively activated microglia and macrophages in the central nervous system. Prog. Neurobiol. 2015, 131, 65–86. [Google Scholar] [CrossRef]
- Xiao, L.; van De Worp, W.R.; Stassen, R.; van Maastrigt, C.; Kettelarij, N.; Stahl, B.; Blijenberg, B.; Overbeek, S.A.; Folkerts, G.; Garssen, J.; et al. Human milk oligosaccharides promote immune tolerance via direct interactions with human dendritic cells. Eur. J. Immunol. 2019, 49, 1001–1014. [Google Scholar] [CrossRef]
Figure 1.
2’-FL induces a change in mRNA expression of M1 and M2 polarization markers in ischemic stroke. (a) Reverse-transcription polymerase chain reaction (RT-PCR) was performed using total RNA extracted from ischemic brains at 1, 3, 7, and 14 days after middle cerebral artery occlusion (MCAO) (n=3). (b) Expression of mRNA for M1 markers (CD32, iNOS and CD11b) and (c) for M2 markers (Arg1, CCL-22 and TGF-β). Data are expressed as Mean ± SEM. * < 0.05, ** < 0.01 and ### < 0.001 vs the Con group, * < 0.05, ** < 0.01 vs the vehicle group.
Figure 1.
2’-FL induces a change in mRNA expression of M1 and M2 polarization markers in ischemic stroke. (a) Reverse-transcription polymerase chain reaction (RT-PCR) was performed using total RNA extracted from ischemic brains at 1, 3, 7, and 14 days after middle cerebral artery occlusion (MCAO) (n=3). (b) Expression of mRNA for M1 markers (CD32, iNOS and CD11b) and (c) for M2 markers (Arg1, CCL-22 and TGF-β). Data are expressed as Mean ± SEM. * < 0.05, ** < 0.01 and ### < 0.001 vs the Con group, * < 0.05, ** < 0.01 vs the vehicle group.
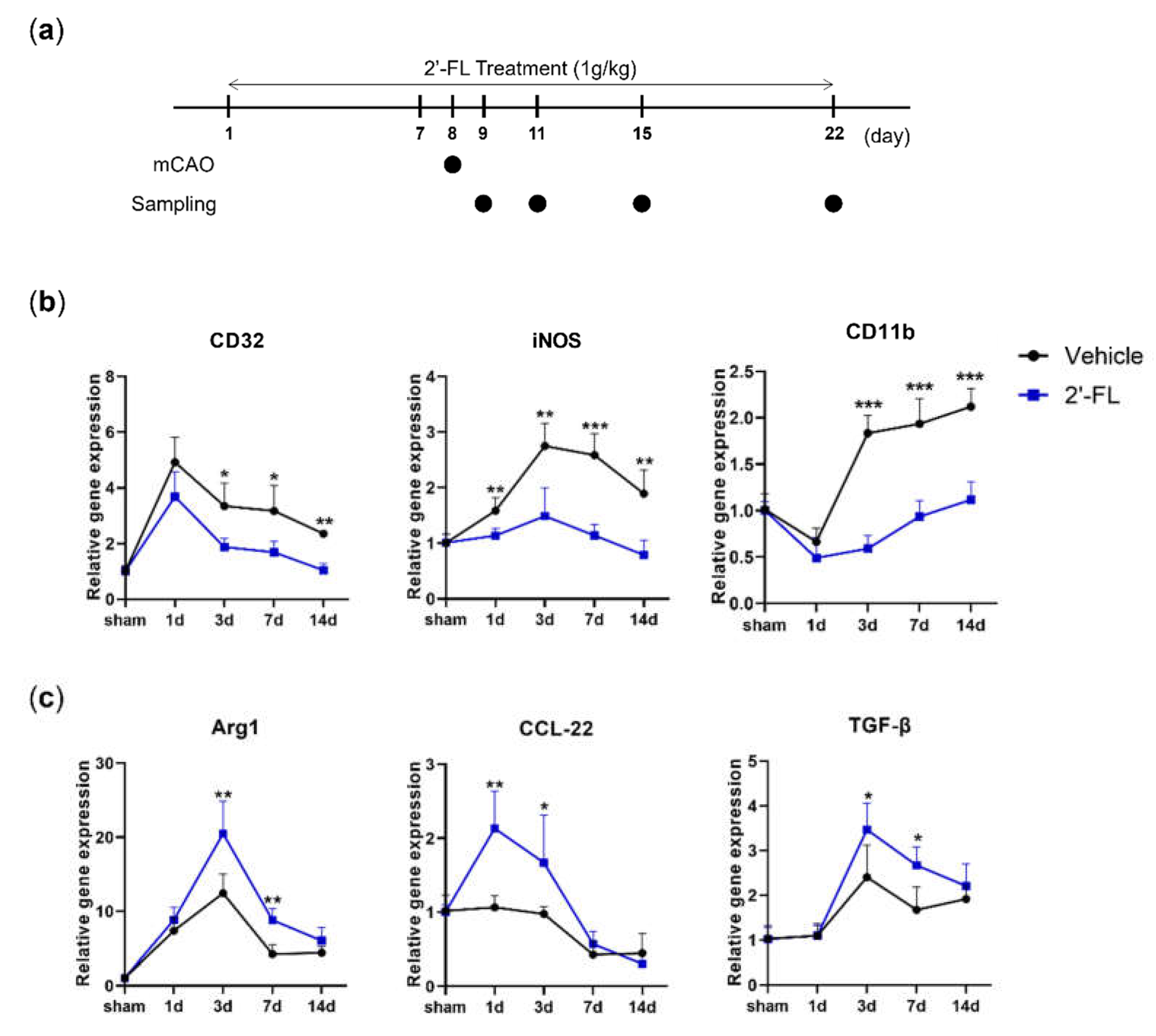
Figure 2.
2’-FL improves neurological and motor function and reduced brain edema after ischemic brain injury. (a) Schedule of an experimental study. (b) Score of neurological deficits, (c) corner test and (d) time to fall down, and (e) Score in wire grip test were evaluated 3 and 7 days after MCAO surgery (n=6). (f) Nissle staining and brain tissue edema volume were evaluated at 3 and 7 days after MCAO surgery (n=3). Data are expressed as Mean ± SEM. ## < 0.01, ### < 0.001 vs the Con group, * < 0.05, ** < 0.01 vs the vehicle group.
Figure 2.
2’-FL improves neurological and motor function and reduced brain edema after ischemic brain injury. (a) Schedule of an experimental study. (b) Score of neurological deficits, (c) corner test and (d) time to fall down, and (e) Score in wire grip test were evaluated 3 and 7 days after MCAO surgery (n=6). (f) Nissle staining and brain tissue edema volume were evaluated at 3 and 7 days after MCAO surgery (n=3). Data are expressed as Mean ± SEM. ## < 0.01, ### < 0.001 vs the Con group, * < 0.05, ** < 0.01 vs the vehicle group.
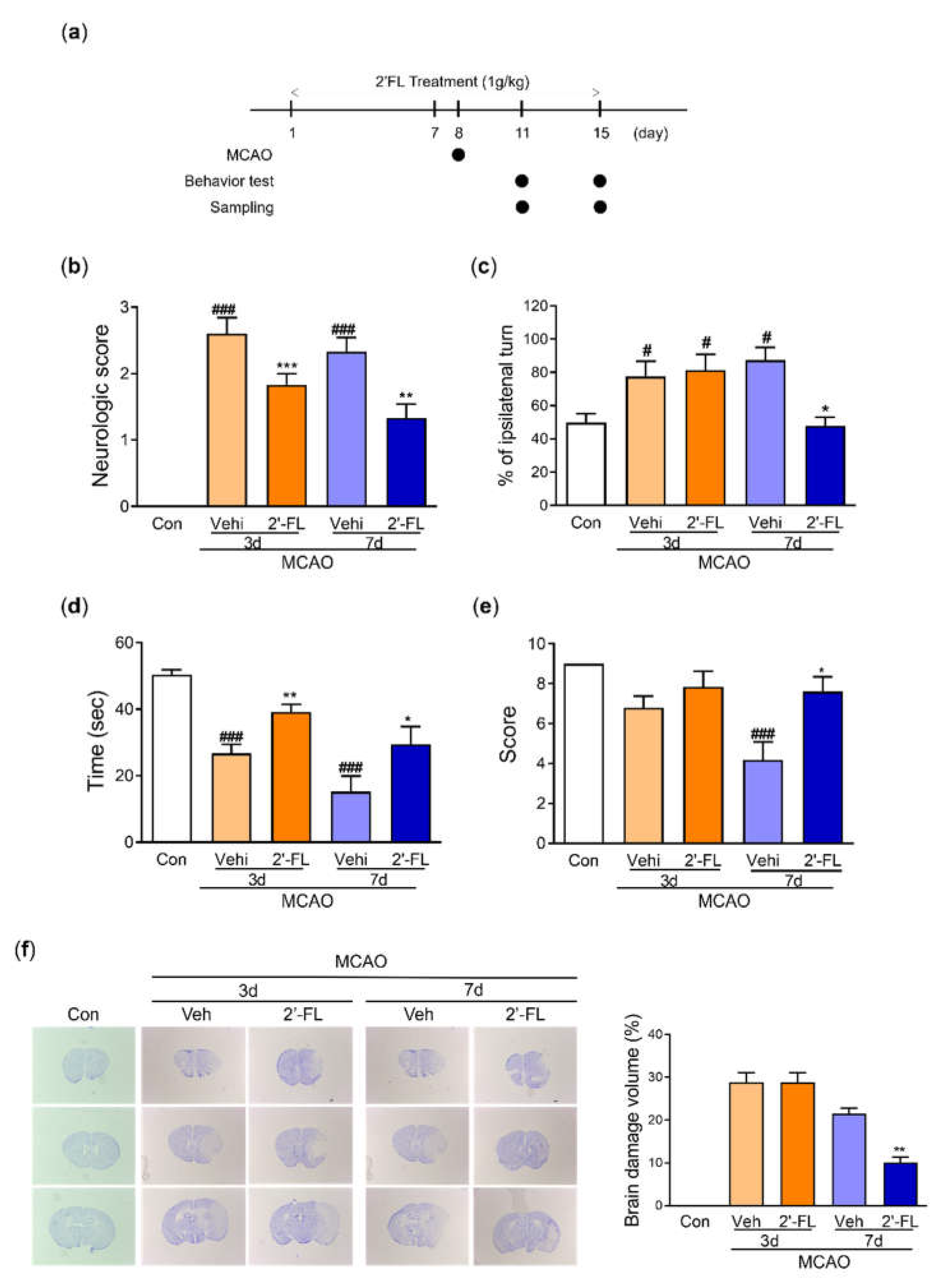
Figure 3.
2’-FL attenuates ROS production after ischemic brain injury. Level of (a) catalase and (b) SOD in the whole brain. (c) Representative western blot and its densitometric analysis of (d) HO-1, (e) NQO1, and (f) iNOS protein in whole brain on 3 and 7 days after MCAO. Data are expressed as Mean ± SEM (N = 3). # < 0.01, ## < 0.05, ### < 0.001 vs the Con group, * < 0.05, ** < 0.01 vs the vehicle group.
Figure 3.
2’-FL attenuates ROS production after ischemic brain injury. Level of (a) catalase and (b) SOD in the whole brain. (c) Representative western blot and its densitometric analysis of (d) HO-1, (e) NQO1, and (f) iNOS protein in whole brain on 3 and 7 days after MCAO. Data are expressed as Mean ± SEM (N = 3). # < 0.01, ## < 0.05, ### < 0.001 vs the Con group, * < 0.05, ** < 0.01 vs the vehicle group.
Figure 4.
2’-FL activates microglia by regulating cytokines after ischemic brain injury. Level of (a) IL-10 and (b) TNF-a in the whole brain. (c) Representative western blot and its densitometric analysis of (d) Iba1, (e) CD32, and (f) CD206 protein in the whole brain on 3 and 7 days after MCAO. (g) Photomicrograph for CD206+/Iba1+ in the striatum of ipsilateral side and its histogram for (h) IOD of Iba1+ cells and (i) the number of CD206+/Iba1+ cells. Data are expressed as Mean ± SEM (N = 3). # < 0.05 ## < 0.01 vs the Con group, * < 0.05, ** < 0.01, *** < 0.001 vs the vehicle group.
Figure 4.
2’-FL activates microglia by regulating cytokines after ischemic brain injury. Level of (a) IL-10 and (b) TNF-a in the whole brain. (c) Representative western blot and its densitometric analysis of (d) Iba1, (e) CD32, and (f) CD206 protein in the whole brain on 3 and 7 days after MCAO. (g) Photomicrograph for CD206+/Iba1+ in the striatum of ipsilateral side and its histogram for (h) IOD of Iba1+ cells and (i) the number of CD206+/Iba1+ cells. Data are expressed as Mean ± SEM (N = 3). # < 0.05 ## < 0.01 vs the Con group, * < 0.05, ** < 0.01, *** < 0.001 vs the vehicle group.
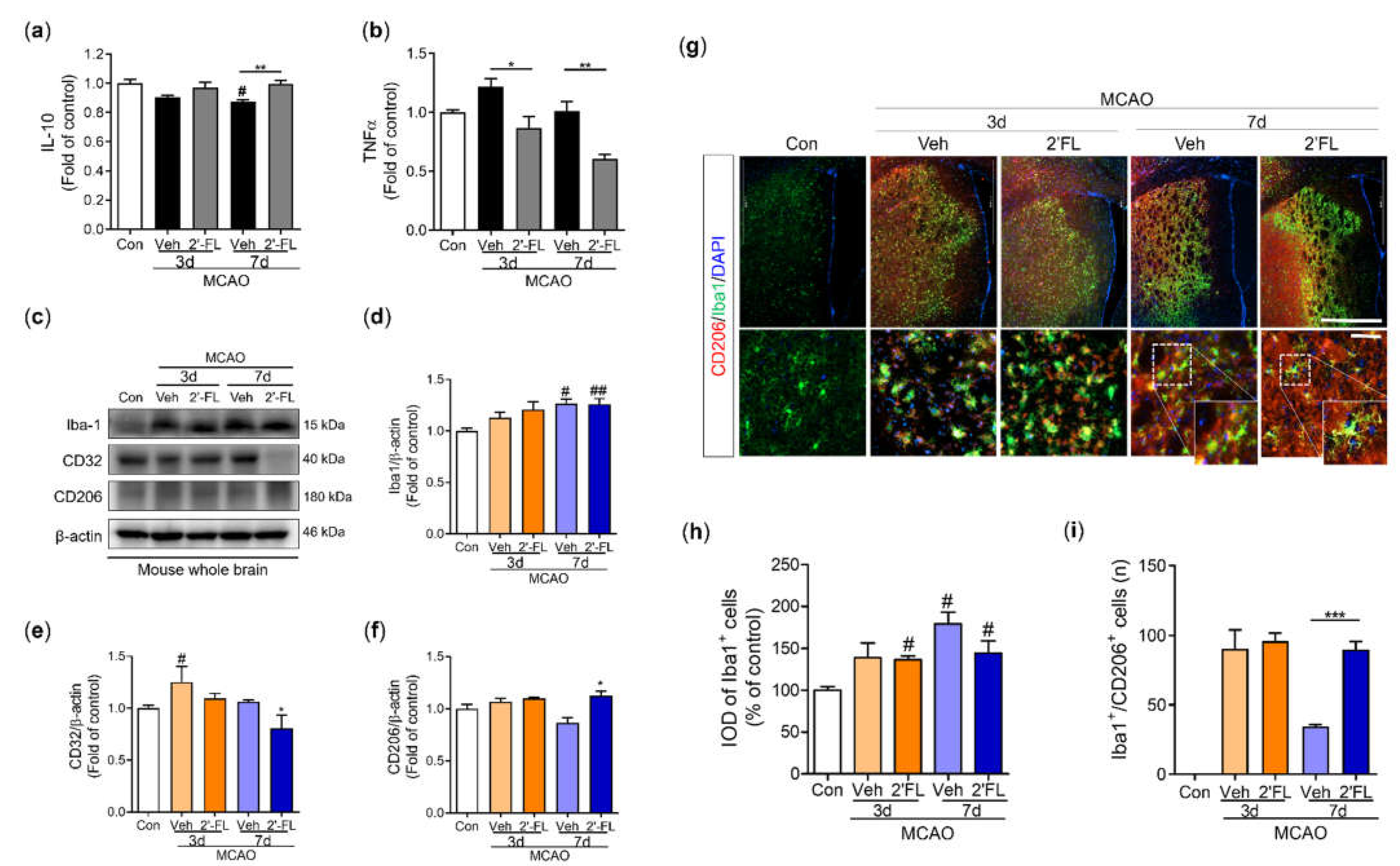
Figure 5.
2’-FL activates STAT6 after ischemic brain injury. (a) Representative western blot and its densitometric analysis of (b) pSTAT6, (c) PGC1α, and (d) pAMPK in whole brain at 3 and 7 days after MCAO. (e) Photomicrograph for pSTAT6+/Iba1+ in the striatum of ipsilateral side and (f) its histogram for IOD of pSTAT6+ cells. (g) Level of IL-4 in the whole brain. Data are expressed as Mean ± SEM (N = 3). # < 0.05 vs the Con group, * < 0.05, ** < 0.01 vs the vehicle group.
Figure 5.
2’-FL activates STAT6 after ischemic brain injury. (a) Representative western blot and its densitometric analysis of (b) pSTAT6, (c) PGC1α, and (d) pAMPK in whole brain at 3 and 7 days after MCAO. (e) Photomicrograph for pSTAT6+/Iba1+ in the striatum of ipsilateral side and (f) its histogram for IOD of pSTAT6+ cells. (g) Level of IL-4 in the whole brain. Data are expressed as Mean ± SEM (N = 3). # < 0.05 vs the Con group, * < 0.05, ** < 0.01 vs the vehicle group.
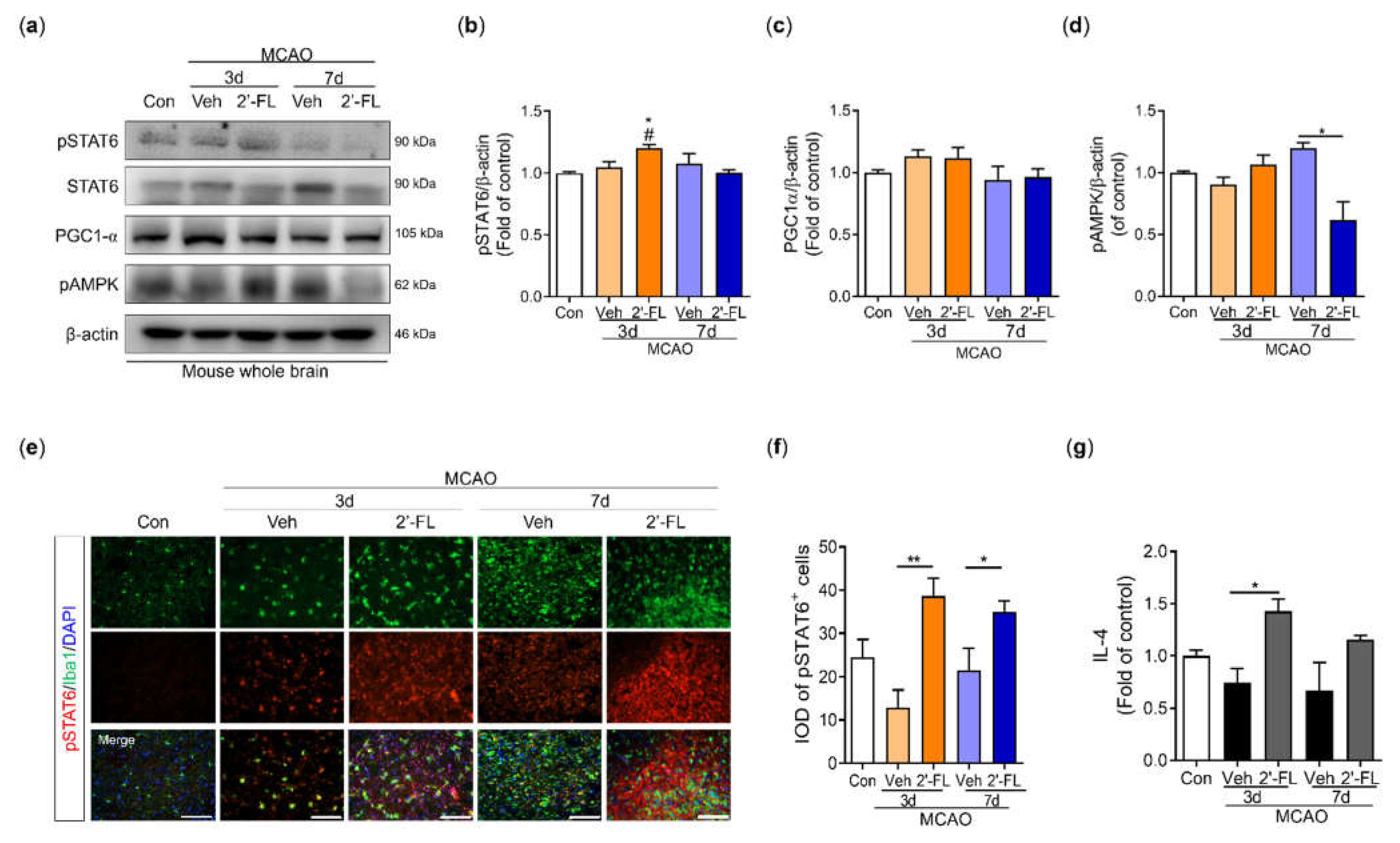
Figure 6.
The proposed neuroinflammation mechanism of 2’-FL in regulating microglial M1/M2 polarization following ischemic injury via IL-4/STAT6 signaling pathways.
Figure 6.
The proposed neuroinflammation mechanism of 2’-FL in regulating microglial M1/M2 polarization following ischemic injury via IL-4/STAT6 signaling pathways.

Table 1.
Primers for RT-PCR. CD32, low-affinity immunoglobulin gamma Fc region receptor III-b; iNOS, inducible nitric oxide synthase; CD11b, integrin alpha-M; Arg-1, arginase-1; CCL-22, C-C motif chemokine ligand-22; TGF-β, transforming growth factor-β.
Table 1.
Primers for RT-PCR. CD32, low-affinity immunoglobulin gamma Fc region receptor III-b; iNOS, inducible nitric oxide synthase; CD11b, integrin alpha-M; Arg-1, arginase-1; CCL-22, C-C motif chemokine ligand-22; TGF-β, transforming growth factor-β.
| Gene | Primer (forward) | Primer (reverse) |
|---|---|---|
| CD32 | AATCCTGCCGTTCCTACTGATC | GTGTCACCGTGTCTTCCTTGAG |
| iNOS | CAAGCACCTTGGAAGAGGAG | AAGGCCAAACACAGCATACC |
| CD11b | CCAAGACGATCTCAGCATCA | TTCTGGCTTGCTGAATCCTT |
| Arg1 | TCACCTGAGCTTTGATGTCG | CTGAAAGGAGCCCTGTCTTG |
| CCL-22 | CTGATGCAGGTCCCTATGGT | GCAGGATTTTGAGGTCCAGA |
| TGF-β | TGCGCTTGCAGAGATTAAAA | CGTCAAAAGACAGCCACTCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



