
Submitted:
05 May 2023
Posted:
10 May 2023
You are already at the latest version
Abstract
Synucleins are a family of small, soluble proteins mainly expressed in neural tissue and in certain tumors. Since their discovery, tens of thousands of scientific reports have been published about this family of proteins as they are associated to severe human diseases. Although the physiological function of these proteins still elusive, their relation with neurodegeneration and cancer was clearly described over the years. In this review, we summarize data connecting synucleins and cancer, going from the structural description of these molecules, to their involvement in tumor-related processes, and discuss the putative use of this proteins as cancer molecular biomarkers.
Keywords:
synucleins
; cancer
; aggregation
; pathways
; biomarkers
1. Introduction
Synucleins are small, highly conserved proteins implicated in neurodegenerative disorders and cancer. This family is composed of three members, alpha, beta and gamma synuclein (αS, βS and γS, respectively). Synucleins are commonly described as intrinsically disordered proteins (IDPs), as they lack of a fixed or ordered three-dimensional structure, and they contain intrinsically disordered regions that lack secondary structure and global topology [1].
The first member of this family to be discovered was αS, which was isolated from the electric ray Torpedo Californica in 1988 by the use of an antiserum against purified cholinergic synaptic vesicles [2]. The name synuclein was coined because this first study revealed a neuron-specific protein with nuclear and presynaptic terminal localization, which was proposed to be involved in coordinating nuclear and synaptic neuronal events [2]. Discovery of αS was rapidly followed by the identification of two close homologs, βS and γS. βS was first identified in 1990 by Nakajo et al. as a 14kDa phosphoneuroprotein present in bovine brain [3] and its complete sequence was soon published by the same group [4]. It was in 1994 when αS and βS were purified and sequenced from human brain, and their close homology established the existence of a family of human brain synucleins [5]. The last discovered member of the family was γS, identified as a differentially expressed gene in breast cancer and it was first named breast cancer-specific gene 1 (BCSG1) because it was abundant in advanced infiltrating breast carcinoma and almost undetectable in normal or benign breast lesions [6]. After cloning from brain genomic and cDNA libraries, the previously identified BCSG1, also called persyn [7,8], was named as SNCG and considered to be the third member of synuclein family [9].
2. Synucleins Structure and Homology
Synucleins are intrinsically disorder or unstructured proteins prone to aggregate, involved in severe human diseases. Around 30% of the eukaryotic proteins contain intrinsically disordered regions lacking of secondary structure and global topology, despite representing functional states [10]. This abundancy suggests their importance in key cellular processes as homeostasis and survival [11]. IDPs are characterized by containing few hydrophobic residues, a high net charge, low sequence complexity and structure-breaking residues (e.g., proline) that facilitate disorder [12,13].
The amino acid sequence of synucleins (127–140amino acids) is generally divided into three main regions: N-terminus, nonamyloid component (NAC) region, and C-terminus. Synucleins share significant sequence homology at the N-terminal region, while their C-termini are specific for each member of the family (Figure 1A).
The N-terminal part of synucleins is highly conserved among the three members and is responsible for their lipid-binding properties. αS N-terminal region is presumed to form one or two α-helices when interacting with the lipid bilayer of membranes [14]. Similar events were described for β and γS, as both proteins are also referred as IDPs, but upon binding to surfactants or lipids, they rearrange into predominantly a two α-helix conformation (Figure 1B). It was reported that αS N-terminal region binds synaptic vesicle membranes, showing a binding preference for highly curved membranes. Although βS and γS share this α-helical lipid-binding motif with αS, they reveal a reduced binding affinity towards membranes [15].
Synucleins central core is commonly named as NAC region, because amino acids positions 61-95 in αS were identified as the “non-amyloid β component” found in amyloid plaques associated with Alzheimer's disease. Several reports point to this region as the highly amyloidogenic part of the molecule, promoting the formation of β-amyloid plaques in vivo [16,17]. The absence of most of the NAC region in βS (Figure 1A,B) is the main determinant for its inability to form amyloid fibrils under physiological conditions. On the other hand, it was described that γS is prone to aggregate into small, soluble oligomers in solution and, upon oxidation of methionine 38, into larger aggregates [18,19].
C-terminal domain of synucleins does not form part of the amyloid core region or affect membrane binding ability of this family of proteins. The most remarkable characteristic of this region is its negative charge content, but the role of this protein domain is less understood and controversial. It was proposed that it could be involved in metal binding (calcium, copper, iron and possibly other metals), but the most putative function for this region is to mediate protein-protein interactions. Supporting this hypothesis, C-terminal region contains sites for posttranslational modifications in all members of this family of proteins, including serine and tyrosine phosphorylation which can modulate protein interactions [20]. In comparison, γS has a relatively shorter C-terminal domain with fewer acidic residues (Figure 1A,B).
A conserved feature for the three synucleins is the imperfect 11-mer repeat, with the predominant KTKEGV consensus sequence (Figure 1A). The 11-amino acid repeat spans seven times in αS and γS, and six times in βS, throughout the N-terminus and NAC region, with slight differences. These repeats were associated to reversible lipid binding, oligomer stabilization and aggregation [21].
Primary structural analysis of synucleins reveals that αS and βS are more closely related to each other than to γS, although three members of this family have been found in all vertebrates (Figure 1C,D) [22]. Up to date, no synuclein counterpart was identified in invertebrates, indicating that these proteins are vertebrate-specific. A more refined analysis indicates that the number of members for this family may be different among vertebrates. While all members are present in mammals and birds, this variates in fish depending on the species.
3. Synucleins Expression and Physiological Roles
Synucleins are abundant proteins that are mainly found in neural tissue (up to 0.1% of total brain proteins by some estimates) and, to a lesser degree, in red blood cells. In the brain, αS protein is mainly detected at cerebral cortex and cerebellum (Figure 1E), although according to “The Human Protein Atlas”, SNCA transcripts are found also in hippocampus, amygdala and thalamus among other parts (Figure 1F) [23,24]. Outside the brain, high expression of αS can be observed at bone marrow, kidney, skin, colon and other tissues. βS brain expression partially correlates with αS, although high levels for this protein are detected at hippocampus and caudate. Outside the brain, low levels of βS can be found in kidney and intestines, although SNCB transcripts can be also detected in retinal tissue (Figure 1E,F).
As it is the case at structural level, γS is the most divergent member regarding expression pattern. γS protein can be detected in brain at cerebral cortex, cerebellum and hippocampus, while expression of this member can be also found in adrenal gland, bladder, lung, breast, skin, colon and other organs (Figure 1E,F).
Although synucleins have been well studied in the context of neurodegeneration and cancer, a clear biological function for synuclein proteins remains poorly understood. As mentioned, the three members of this family bind curve lipid membranes and are involved in regulation of synaptic vesicle endocytosis [23,24,25]. αS maintains neurotransmitter release by regulating synaptic vesicle pools at the synapse [26] assisting SNARE-complex assembly. It was also suggested that αS maintains normal synaptic function during aging [26]. Nevertheless, almost no clear function was described for βS and γS in brain, despite their involvement in neurodegenerative diseases [27,28,29]. It was suggested for βS to modulate cell survival, metal levels and dopamine uptake, and to decrease αS aggregation [30,31,32]. γS physiological function still even more elusive, but it was proposed this protein influences neurofilament network integrity and to chaperone in retinal photoreceptor cells [33,34].
Beside synapses, αS participates in the physiology of other cellular organelles such as mitochondria, by interacting with mitochondrial proteins as respiratory chain complexes and ATP synthase and promoting the expression of Miro proteins which connect mitochondria to microtubules [35]. It was also described that αS interacts with cytoskeletal components and nuclear components [36,37]. αS also has physiological roles in nonneuronal cells as blood cells, having structural functions and metabolic activities [38].
4. Synucleins Are Cancer-Related Proteins
Enrichment analysis of genes associated with αS, βS and γS, clearly indicates that all members of this family are involved in cell signaling processes associated to disease development and particularly related to cancer (Figure 2A). This connection to cancer was clearly established for γS [6,39,40,41], but it was more recently when it was described a putative role in this pathology for the other two members.
The first report of the involvement of synucleins in cancer was in 1997, at the time the third member of this family was discovered [6] (Table 1). Two years after discovery, γS was proposed to stimulate breast cancer invasion and metastasis [41]. In 2000, Bruening et al. reported the expression of βS and γS in stage III/IV breast ductal carcinomas and αS, βS and γS in ovarian carcinomas [42]. At that time, they suggested γS as a putative target for cancer therapy. The same year, two different studies identified the expression of αS in brain tumors showing neuronal or mixed neuronal/glial differentiation [43,44]. In 2001, γS was described as a centrosome-associated protein in retinoblastoma, involved in signal transduction and cell cycle progression [45], and the next year it was described for this member to control cancer cells survival and chemotherapy resistance [46]. In contraposition with the pro-tumorigenic role proposed for γS, Zhou et al. suggested in 2003 a negative regulative role for this synuclein in the development of esophageal squamous cell carcinoma [47]. That same year Fung et al. determined expression of γS in high grade glial tumors and αS/βS in a high percent of medulloblastomas, but no association between synuclein expression and tumor aggressiveness was established [48]. Following these first reports, many studies pointed to explore the role of γS in different tumor types such as pancreatic adenocarcinoma [49], gastric cancer [50], bladder cancer [51], and cervical, colon, lung and prostate cancer [39]. During 2008 and 2009 Ye et al. reported the connection between γS expression and colorectal cancer progression, and explored also the expression of the other members of the family in this tumor type suggesting that co-expression of γS with αS or βS could increase sensitivity to predict advanced stage or lymph node invasion in this tumor type [52,53]. Almost at the same γS expression was associated with uterine papillary serous carcinoma [54]. In 2010, Matsuo and Kamitani described the expression of αS in melanoma, suggesting that αS may be the key to understand epidemiological studies reporting the co-occurrence of melanoma and Parkinson’s disease [55]. αS and βS were proposed as expression markers for specific leukemias by Maitta et al. in 2011 [56]. More recently (2012-2015) new roles for γS were described in endometrial adenocarcinoma [57], gallbladder cancer [58], and oral squamous cell carcinoma [59].
It is clear that many reports link synucleins expression with different types of tumors. Exploring databases recruiting RNA sequencing expression data of tumors and normal samples [60], it is possible to extend these observations. An increased expression of αS in melanoma compared to normal tissue (Figure 2B) correlates perfectly with previous reports [55,61]. This is also the case for pancreatic adenocarcinoma, where it was recently described an incremental expression for this protein [62]. However, there is a significant decrease in expression for this member of the family in colon adenocarcinoma in contrast with the increased protein level reported in colorectal cancer [53]. Interestingly, SNCA expression in lung adenocarcinoma is lower than in normal tissue, and high αS expression is related to immune infiltration and a better prognosis [63], supporting the negative association reported between Parkinson’s disease and lung cancer [64].
There is no RNA sequencing expression data for the status of βS in medulloblastoma, although protein expression was reported [48], nevertheless transcriptional levels of βS significantly decrease in gliomas and glioblastomas compared to normal tissue (Figure 2B).
The connection between γS and cancer was the first described for a member of this family. As mentioned, γS was initially named as BCSG1 as a result of differential cDNA sequencing studies to identify genes differentially expressed in normal breast compared breast cancer [6]. Similar results were observed at protein level by immunostaining of normal and breast cancer tissues [65]. However, RNA sequencing expression data of breast tumors and normal samples suggest a reduction in transcripts for SNCG in this type of cancer (Figure 2B). A similar result can be observed for colon adenocarcinoma, in spite of the reports describing increased γS protein levels in colorectal cancer compared to normal tissue [66,67]. These disparities may reflex the stabilization of this protein in these types of tumors (maybe by posttranslational modifications or accumulation of stable high molecular species) or a more efficient translation of RNA. It is also important to note that transcriptional levels in these studies are not related to any specific cell type within the tumor which can account for discrepancies. As reported for protein levels, γS transcripts were significantly increased in ovarian and pancreatic carcinomas (Figure 2B).
5. Synucleins Regulation and Posttranslational Modifications
Expression of synucleins is regulated at different levels. αS expression is regulated by various transcription factors such as GATA-1/2, TRIM32, p21 and p27 by direct binding to the promotor region of SNCA [68]. αS expression is modulated by growth factors (nerve growth factor and basic fibroblast growth factor) via MAPK/ERK and PI3K pathways [69], β2-adrenoreceptor [70] and by dopamine [71].
βS transcriptional regulation was not studied in detail, but tissue distribution data indicate a close similarity of expression and regulation patterns with αS [72]. It was reported that βS expression could be controlled at transcriptional level by binding of MTF-1 (Metal Transcription Factor-1) to metal response elements at the promoter [73].
γS is overexpressed in a variety of invasive and metastatic cancers and is regulated by multiple transcriptional mechanisms. Overexpression of SNCG in cancer cells may be due to aberrant demethylation of CpG islands within the promoter, AP1 transactivation, and insulin-like growth factor signaling [50,74,75]. It was also reported that TGF-β induces SNCG expression by Smad-Twist1 axis [76].
Synucleins are not only regulated at transcriptional level, but also, they are substantially posttranslational modified. Synucleins posttranslational modifications (PTMs) may be critical to modulate proteins normal and pathophysiological functions, and to direct them to different cellular compartments. Several PTMs were described to modulate αS propensity to aggregate by triggering conformational changes, such as phosphorylation, ubiquitination (mono, di and tri ubiquitination), acetylation, nitration (all four tyrosine residues) and SUMOylation. Particularly, phosphorylation at serine 129 can be detected in blood and it was suggested as a potentially useful biomarker for Parkinson’s disease [77]. Several kinases were demonstrated to be responsible for phosphorylation at this position, including casein kinases I and II, G protein-coupled receptor kinase 2 (GRK2) LRRK2 and PLK, but phosphorylation at other αS amino acids was also reported (S87, Y125, Y133, and Y136) [78].
βS is modified by β-N-acetylglucosamine linked to hydroxyl groups on serine and threonine [79], but this PTM is specific for βS and not αS. βS is also phosphorylated/dephosphorylated at serine residues by polo-like kinase 1 and 3 and PP2A respectively [31].
Surprisingly, the most studied PTM for γS is its oxidation at methionine 38, which facilitates formation of aggregates and deposits and was detected in aberrant inclusions in amygdala of dementia with Lewy bodies patients, colocalizing with serine 29-phosphorylated αS [80]. It was proposed that γS oxidation at methionine 38 and tyrosine 39, two of the most easily oxidized residues, allows γS to seed the aggregation of αS [19].
6. Synucleins Aggregation and Cancer
αS oligomerization and aggregation were strongly studied in the context of neurogenerative diseases [28,72,80]. As mentioned, γS is also able to form high molecular weight fibrils and aggregates and it was also proposed for γS that could be secreted by exosomes, transmitted to other cells and promote aggregation of intracellular proteins in a prion-like manner, as described for αS [19]. Although those events were described in the context of neurodegeneration, several studies suggest a link between protein aggregation and cancer. For example, it was described that both wild type and mutant p53 proteins show kinetics of aggregation and fibrillar morphology that resemble those of classical amyloidogenic proteins, as αS [81,82], suggesting that p53-mutant cancers may be a class of protein aggregation diseases.
As αS and γS, p53 was described to be transmitted between cells in a prion-like mechanistic fashion [83]. After discovery of p53 aggregation, other potentially cancer-related proteins were shown to aggregate, as PTEN, p63 and p71 [84,85]. The question of the impact of protein aggregation in cancer is a research field currently growing, as new reports are connecting aggregation patterns with tumor treatment-resistance, tumor progression, and metastasis development [86,87,88].
It was also recently described that αS expression in melanoma is associated with the presence of high molecular weight species of this protein, and that treatment with aggregation-inhibiting compounds prevents tumor growth [61], suggesting a key role for this synuclein in melanoma progression, mainly related to autophagy. Additionally, knocking out αS gene in SKMel28 melanoma cells suppressed tumor growth and promoted dysregulation of cellular iron metabolism [89]. Finally, Dean and Lee demonstrated last year that αS localizes at melanosomes, where it modulates Pmel17 aggregation affecting melanosome maturation and melanin production [90].
Up to date, no reports up addressed the status of oligomerization/aggregation of γS in tumors. However, it is well described for this protein that is secreted from tumor cells, and elevated γS levels were especially reported in advanced stages of the pathology.
Although not in cancer, it was proposed in glaucoma that gS oligomeric/aggregated forms could enter in the bloodstream, generating autoantibodies [91]. The dynamic intracellular localization of γS and its ability to be transmitted from one cell to another suggest that more implications for this protein in cancer may appear soon.
7. Synucleins Controlled Pathways in Cancer
Many efforts have been directed to understand the role of synucleins in cancer during the last decades. From these studies it looks clear that synucleins play roles at different steps leading to tumor development and progression (Figure 3).
αS levels were reported to affect cell cycle progression and proliferation in osteosarcoma models, affecting tumor differentiation by down-regulating proteasome and PKC, and up-regulating lysosomal activity [92]. It was also described that melanoma and mammary carcinoma cells can uptake exogenously added αS, which promote in vitro proliferation of those cells [93]. Up-regulation of αS was also proposed to contribute to phenotype aggressiveness in meningiomas, affecting proliferation, apoptosis, migration and invasion of cells by modulating AKT/mTOR pathway [94]. In melanoma cells, αS was also proposed to modulate cell proliferation by interfering with iron metabolism [89]. Same study also showed in a mouse model that depletion of αS in tumors promotes apoptosis, linking this increased in cell death to high levels of ferric iron. However, Turriani et al. [61] suggested that proliferation of melanoma cells is modulated by αS aggregation, as treatment with oligomers modulators inhibited melanoma cells proliferation and increased apoptosis through dysregulation of cell autophagy. Interestingly, both excess and deficiency of iron can lead to cellular stress, affecting autophagic pathways. It was also demonstrated in neuron cells that iron promotes αS aggregation and transmission by inhibiting autophagosome-lysosome fusion, affecting AKT/mTORC1 signaling [95].
Involvement of AKT/mTOR pathway control by αS was also described for lung adenocarcinoma cells. However, in this context increased expression of αS inhibited proliferation of pulmonary cells, decreased PI3K levels and prevented AKT and mTOR phosphorylation [63]. To support the idea that αS proliferative control highly depends on the cellular context, it was reported that αS over-expression on PC12 cells enhanced proliferation by increasing cyclin B levels and ERK1/2 phosphorylation, and down-regulating retinoblastoma [96].
EMT, migration and invasion are pro-tumoral processes modulated by actin and tubulin cytoskeleton. Several reports described αS interaction with actin and tubulin in the context of neurodegeneration [37,97], therefore it is possible that more insights regarding the role of αS in cytoskeleton dynamics will be achieved in the near future.
Other process related to cancer reported for αS in neurodegenerative models that could play key roles in cancer include its ability to go to the nucleus. Studies based on in vitro and in vivo models suggest αS interacts with DNA and histones, and regulates transcription and DNA repair [98]. Still, the role of nuclear αS in tumor cells needs to be further addressed. Also, the impact of αS in mitochondrial energetics and dynamics in tumors needs to be explored in detail, as many reports proposed roles for this protein in mitochondrial oxidative phosphorylation, membrane potential, and homeostasis [99].
So far, there are few reports regarding mechanisms of action for βS in cancer. As protective roles have been attributed for this protein in neurodegeneration (mainly preventing αS aggregation), it would be possible to think that βS could directly or indirectly interfere with pathways such as ERK, PI3K-AKT described for αS both in cancer and neurodegenerative scenarios.
By contrast, several molecular implications were assigned for γS in cancer (Figure 3). It was reported in breast and ovarian cancers that γS over-expression leads to constitutive activation of ERK1/2 and down-regulation of JNK1, and that γS promotes resistance the chemotherapeutic drugs paclitaxel and vinblastine [46]. Also, the interaction of γS with the mitotic checkpoint protein Bub1R in breast cancer is well documented, leading to mitotic checkpoint compromise through Bub1R inactivation [100]. Furthermore, γS confers cellular resistance to anti-microtubule drugs by interfering with mitotic checkpoint control.
γS was also described to act as an androgen receptor co-activator in prostate cancer, modulating cell cycle progression, proliferation, migration and invasion [101].
The relation of synucleins with AKT/mTOR pathways is not only restricted to αS, as it was reported that γS binds to AKT kinase domain promoting its phosphorylation in non-small cell lung cancer models. In this tumor type, γS promotes cell survival and proliferation by AKT activation, playing a leading role in this pathogenesis [102]. Contrary to what we mentioned for αS, all the reports connecting γS with ERK or AKT pathways indicate that this family member promotes activation of these pathways including gastric [103], cervical [104], ovarian [105] and endometrial cancers [106]. Also, in all the tumor types in which was explored, γS down-modulates JNK, leading to increase survival and evading apoptosis.
γS expression is directly related to EMT, invasion and development of metastasis. It is reported that γS favors the expression of metalloproteinases 2 and 9 in retinoblastoma and breast, bladder, cervical and liver cancers [76,107,108], and it promotes cell motility by regulating Rho GTPases in breast and ovarian cancers [76,109].
Other interesting mechanistic fact associated to γS expression in cancer is related to its secretion. It is documented for colon and breast cancer that γS secretion promotes aggressive phenotype of cancer cells, favoring invasion [110].
8. Synucleins as Cancer Biomarkers
Understanding the characteristics of a tumor allows to personalize treatments for that particular cancer. The search for cancer biomarkers has grown in the last years as they are used to reflect incidence and outcome of cancer, but also the effects of treatments or interventions. Thus, cancer biomarkers are normally molecular indicators of cancer susceptibility/risk, occurrence/monitoring of cancer or patient outcome, and they can be detected in biopsy samples or, more interestingly, through non-invasive methods analyzing blood, saliva, urine, etc. The most relevant property for a biomarker is its usefulness to optimize decisions in the clinical practice.
Undoubtedly, αS has been suggested as a biomarker for Parkinson’s disease diagnosis by many studies [77]. But, can synucleins be used as cancer biomarkers? In fact, although more studies should be addressed, they can potentially be good candidates.
It was established that γS has predictive and prognostic values in various types of cancer and it can be used as a stage-specific marker in several tumors [111]. Abnormal γS expression has been related to tumor development, promoting tumor progression and metastasis. The use of γS as a tumor progression biomarker arises from studies detecting serum γS in a high percentage of pancreatic adenocarcinomas, while no presence of this protein was detected in healthy controls [49,51].
As mentioned before, Ye et al. corroborated synucleins expression in colorectal cancer by IHC, predicting that γS is a good marker for cancer progression, but simultaneous detection of αS/γS or βS/γS predicted advanced stage and lymph node invasion [53]. Accordingly, analysis of transcriptional levels of αS, βS and γS in colon adenocarcinoma indicates a clear correlation between high transcriptional levels for genes coding these proteins with a poor prognosis (Figure 4). Similarly, it was described that γS in breast cancer is related to poor prognosis [65] and analysis of SNCG transcriptional levels in breast cancer are significantly associated to poor outcome.
Increased expression of αS in lung adenocarcinoma was proposed as a good prognostic biomarker as it directly correlates with increase immune infiltration and better prognosis [63]. In line with this report, high SNCA expression correlates with increased survival, and the inverse association is observed in melanoma, in agreement with the pro-tumoral role observed for αS in this tumor type.
βS expression could be also used as a prognostic marker in glioma and pheochromocytoma/paraganglioma as SNCB expression is associated with good and worst prognosis respectively.
Nevertheless, it is important to have in mind that tumor mRNA levels not always correlate with tumor protein levels or even with the abundancy of protein in fluids. As an example, expression of γS in bladder carcinoma was proposed to be a good marker to predict recurrence, but not a reliable marker for staging or prediction of survival rate [112]. Indeed, high transcriptional SNCG levels are associated with good prognosis in bladder cancer (data not shown).
During the last decades, several studies associated γS levels with poor outcome in endometrial adenocarcinoma [57,106], which correlates with a worst prognosis for uterine corpus endometrial carcinoma patients with high level of SNCG transcripts (Figure 4). It was also proposed that γS could be a prognostic marker for tumor cell migration in biliary carcinomas [113].
As synucleins are detectable in fluids such as blood, saliva, urine and others, further comprehension of the potential role of these proteins as biomarkers stands as a very promising field to improve diagnosis, progression and monitoring of patients through reliable non-invasive methods.
9. Conclusions
Cancer is one of the leading causes of death worldwide. On the other hand, populational longevity associated with the increase in life expectancy brings increasing risk for neurodegenerative disorders. Both pathologies affect millions of people, severely compromising quality and expectative of life and representing one of the most chronic diseases.
Cancer and neurodegeneration are associated to opposite ends as one is related to cell proliferation and cell death resistance, while the other is directly linked to premature cell death. Nevertheless, these two diseases are not so distant.
In this work, we focused on synucleins, a family of small proteins that could represent a link between cancer and neurodegeneration. Reports describing the involvement of synucleins in cancer increased rapidly during the last decades suggesting that these proteins, initially associated to neurodegeneration, play crucial roles in cancer progression and many studies provided conclusive evidence to support the idea that this family of proteins is involved in cell signaling processes related to cancer development. The importance of synucleins in cancer is such that they have been proposed as relevant biomarkers for several tumor types, showing a potential in enhancing the accuracy of diagnosis, tracking the progression of disease, and monitoring patients using non-invasive techniques.
Interestingly, as the physiological function of these proteins is not fully understood, knowledge gained about synucleins in one field could lead to advances in the other, and thus feed off each other.
Author Contributions
Conceptualization, L.C.Z., F.M. and M.M-M. Bioinformatics and data curation, A.A., L.E.A., F.M. and L.C.Z. Writing—original draft preparation, L.C.Z. and F.M.; writing—review and editing, M.M-M. Visualization and figures design, N.C.L., M.M., F.M and L.C.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants to M.M-M. from Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT, PICT2018-0900 and PICT2019-0801).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank all CIPReB personnel for help and collaboration, and Facultad de Ciencias Médicas for support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dunker, A.K.; Oldfield, C.J.; Meng, J.; Romero, P.; Yang, J.Y.; Chen, J.W.; Vacic, V.; Obradovic, Z.; Uversky, V.N. The Unfoldomics Decade: An Update on Intrinsically Disordered Proteins. BMC Genomics, 2008, 9. [Google Scholar] [CrossRef] [PubMed]
- Maroteaux, L.; Campanelli, J.T.; Scheller, R.H. Synuclein: A Neuron-Specific Protein Localized to the Nucleus and Presynaptic Nerve Terminal. J Neurosci. 1988, 8, 2804–2815. [Google Scholar] [CrossRef] [PubMed]
- Nakajo, S.; Omata, K.; Aiuchi, T.; Shibayama, T.; Okahashi, I.; Ochiai, H.; Nakai, Y.; Nakaya, K.; Nakamura, Y. Purification and Characterization of a Novel Brain-Specific 14-kDa Protein. J Neurochem 1990, 55, 2031–2038. [Google Scholar] [CrossRef] [PubMed]
- Nakajo, S.; Tsukada, K.; Omata, K.; Nakamura, Y.; Nakaya, K. A New Brain-specific 14-kDa Protein Is a Phosphoprotein: Its Complete Amino Acid Sequence and Evidence for Phosphorylation. Eur J Biochem 1993, 217, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Jakes, R.; Spillantini, M.G.; Goedert, M. Identification of Two Distinct Synucleins from Human Brain. FEBS Lett 1994, 345, 27–32. [Google Scholar] [CrossRef]
- Ji, H.; Liu, Y.E.; Jia, T.; Wang, M.; Liu, J.; Xiao, G.; Joseph, B.K.; Rosen, C.; Shi, Y.E. Identification of a Breast Cancer-Specific Gene, BCSG1, by Direct Differential CDNA Sequencing. Cancer Res 1997, 57, 759–764. [Google Scholar]
- Buchman, V.L. Persyn, a Member of the Synuclein Family, Has a Distinct Pattern of Expression in the Developing Nervous System. J Neurosci. 1998, 18, 9335–9341. [Google Scholar] [CrossRef]
- Ninkina, N.N.; Alimova-Kost, M. V.; Paterson, J.W.E.; Delaney, L.; Cohen, B.B.; Imreh, S.; Gnuchev, N. V.; Davies, A.M.; Buchman, V.L. Organization, Expression and Polymorphism of the Human Persyn Gene. Hum Mol Genet. 1998, 7, 1417–1424. [Google Scholar] [CrossRef]
- Lavedan, C.; Leroy, E.; Dehejia, A.; Buchholtz, S.; Dutra, A.; Nussbaum, R.L.; Polymeropoulos, M.H. Identification, Localization and Characterization of the Human γ-Synuclein Gene. Hum Genet 1998, 103, 106–112. [Google Scholar] [CrossRef]
- Alberti, S.; Hyman, A.A. Are Aberrant Phase Transitions a Driver of Cellular Aging? Bioessays 2016, 38, 959–968. [Google Scholar] [CrossRef]
- Berlow, R.B.; Dyson, H.J.; Wright, P.E. Functional Advantages of Dynamic Protein Disorder. FEBS Lett. 2015, 589, 2433–2440. [Google Scholar] [CrossRef] [PubMed]
- Wright, P.E.; Dyson, H.J. Intrinsically Disordered Proteins in Cellular Signalling and Regulation. Nat Rev Mol Cell Biol. 2015, 16, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Uversky, V.N.; Oldfield, C.J.; Dunker, A.K. Intrinsically Disordered Proteins in Human Diseases: Introducing the D 2 Concept. Annu Rev Biophys 2008, 37, 215–246. [Google Scholar] [CrossRef] [PubMed]
- Pirman, N.L.; Milshteyn, E.; Galiano, L.; Hwelett, J.C.; Fanucci, G.E. Characterization of the disordered-to-α-helical transition of IA₃ by SDSL-EPR spectroscopy. Protein Sci. 2011, 20(1), 150–159. [Google Scholar] [CrossRef] [PubMed]
- Carnazza, K.E.; Komer, L.E.; Xie, Y.X.; Pineda, A.; Briano, J.A.; Gao, V.; Na, Y.; Ramlall, T.; Buchman, V.L.; Eliezer, D. ; Sharma, M,.; Burré, J. Synaptic vesicle binding of α-synuclein is modulated by β- and γ-synucleins. Cell Rep 2022, 39, 110675. [Google Scholar] [CrossRef]
- Li, H.T.; Du, H.N.; Tang, L.; Hu, J.; Hu, H.Y. Structural transformation and aggregation of human α-synuclein in trifluoroethanol: Non-amyloid component sequence is essential and β-sheet formation is prerequisite to aggregation. Biopolymers 2002, 64(4), 221–226. [Google Scholar] [CrossRef]
- Uéda, K.; Fukushima, H.; Masliah, E.; Xia, Y.; Iwai, A.; Yoshimoto, M.; Otero, D.A.; Kondo, J.; Ihara, Y.; Saitoh, T. Molecular cloning of cDNA encoding an unrecognized component of amyloid in Alzheimer disease. Proc Natl Acad Sci U S A 1993, 90, 11282–11286. [Google Scholar] [CrossRef]
- Golebiewska, U.; Zurawsky, C.; Scarlata, S. Defining the oligomerization state of γ-synuclein in solution and in cells. Biochemistry 2014, 53(2), 293–299. [Google Scholar] [CrossRef]
- Surgucheva, I.; Sharov, V.S.; Surguchov, A. γ-Synuclein: Seeding of α-synuclein aggregation and transmission between cells. Biochemistry 2012, 51(23), 4743–4754. [Google Scholar] [CrossRef]
- Eliezer, D. The mysterious C-terminal tail of alpha-synuclein: Nanobody’s guess. J Mol Biol. 2013, 425(14), 2393–2396. [Google Scholar] [CrossRef]
- Bussell, R.; Eliezer, D. A structural and functional role for 11-mer repeats in α-synuclein and other exchangeable lipid binding proteins. J Mol Biol. 2003, 329(4), 763–778. [Google Scholar] [CrossRef] [PubMed]
- George, J.M. The Synucleins. Genome Biol. 2002, 3. [Google Scholar]
- Vargas, K.J.; Makani, S.; Davis, T.; Westphal, C.H.; Castillo, P.E.; Chandra, S.S. Synucleins regulate the kinetics of synaptic vesicle endocytosis. J Neurosci. 2014, 34(28), 9364–9376. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.H.; Eliezer, D. Secondary structure and dynamics of micelle bound β- and γ-synuclein. Protein Sci. 2006, 15(5), 1162–1174. [Google Scholar] [CrossRef] [PubMed]
- Sjöstedt, E.; Zhong, W.; Fagerberg, L.; Karlsson, M.; Mitsios, N.; Adori, C.; Oksvold, P.; Edfors, F.; Limiszewska, A.; Hikmet, F.; et al. An atlas of the protein-coding genes in the human, pig, and mouse brain. Science 2020, 367, eaay5947. [Google Scholar] [CrossRef] [PubMed]
- Cabin, D.E.; Shimazu, K.; Murphy, D.; Cole, N.B.; Gottschalk, W.; McIlwain, K.L.; Orrison, B.; Chen, A.; Ellis, C.E.; Paylor, R.; Lu, B.; Nussbaum, R.L. Synaptic vesicle depletion correlates with attenuated synaptic α-synuclein. J Neurosci. 2002, 22, 8797–8807. [Google Scholar] [CrossRef] [PubMed]
- Peters, O.M.; Millership, S.; Shelkovnikova, T.A.; Soto, I.; Keeling, L.; Hann, A.; Marsh-Armstrong, N.; Buchman, V.L.; Ninkina, N. Selective pattern of motor system damage in gamma-synuclein transgenic mice mirrors the respective pathology in amyotrophic lateral sclerosis. Neurobiol Dis. 2012, 48(1), 124–131. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, K.; Wider, C.; Vilariño-Güell, C.; Soto-Ortolaza, A.I.; Lincoln, S.J.; Kachergus, J.M.; Jasinska-Myga, B.; Ross, O.A.; Rajput, A.; Robinson, C.A.; Ferman, T.J.; Wszolek, K.; Dickson, D.W.; Farrer, M.J. Association of alpha-, beta-, and gamma-Synuclein with diffuse lewy body disease. Arch Neurol 2010, 67, 970–975. [Google Scholar] [CrossRef]
- Galvin, J.E.; Uryu, K.; Lee, V.M.; Trojanowski, J.Q. Axon pathology in Parkinson’s disease and Lewy body dementia hippocampus contains alpha-, beta-, and gamma-synuclein. Proc Natl Acad Sci U S A 1999, 96(23), 13450–13455. [Google Scholar] [CrossRef]
- Brown, J.W.; Buell, A.K.; Michaels, T.C.; Meisl, G.; Carozza, J.; Flagmeier, P.; Vendruscolo, M.; Knowles, T.P.; Dobson, C.M.; Galvagnion, C. β-Synuclein suppresses both the initiation and amplification steps of α-synuclein aggregation via competitive binding to surfaces. Sci Rep. 2016, 6, 36010. [Google Scholar] [CrossRef]
- Hayashi, J.; Carver, J.A. β-Synuclein: An Enigmatic Protein with Diverse Functionality. Biomolecules 2022, 12, 142. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Bar-On, P.; Ho, G.; Takenouchi, T.; Rockenstein, E.; Crews, L.; Masliah, E. β-Synuclein Regulates Akt Activity in Neuronal Cells: A Possible Mechanism for Neuroprotection in Parkinson’s Disease. J Biol Chem 2004, 279, 23622–23629. [Google Scholar] [CrossRef] [PubMed]
- Surgucheva, I.; Ninkina, N.; Buchman, V.L.; Grasing, K.; Surguchov, A. Protein Aggregation in Retinal Cells and Approaches to Cell Protection. Cell Mol Neurobiol 2005, 25, 1051–1066. [Google Scholar] [CrossRef] [PubMed]
- Buchman, V.L.; Adu, J.; Pinõn, L.G.P.; Ninkina, N.N.; Davies, A.M. Persyn, a Member of the Synuclein Family, Influences Neurofilament Network Integrity. Nat Neurosci 1998, 1, 101–103. [Google Scholar] [CrossRef] [PubMed]
- Barba, L.; Paolini Paoletti, F.; Bellomo, G.; Gaetani, L.; Halbgebauer, S.; Oeckl, P.; Otto, M.; Parnetti, L. Alpha and Beta Synucleins: From Pathophysiology to Clinical Application as Biomarkers. Mov Disord. 2022, 37, 669–683. [Google Scholar] [CrossRef] [PubMed]
- Pinho, R.; Paiva, I.; Jerčić, K.G.; Fonseca-Ornelas, L.; Gerhardt, E.; Fahlbusch, C.; Garcia-Esparcia, P.; Kerimoglu, C.; Pavlou, M.A.S.; Villar-Piqué, A.; Szego, É.; Lopes da Fonseca, T.; Odoardi, F.; Soeroes, S.; Rego, A.C.; Fischle, W.; Schwamborn, J.C.; Meyer, T.; Kügler, S.; Ferrer, I.; Attems, J.; Fischer, A.; Becker, S.; Zweckstetter, M.; Borovecki. F.; Outeiro, T.F. Nuclear Localization and Phosphorylation Modulate Pathological Effects of Alpha-Synuclein. Hum Mol Genet 2019, 28, 31–50. [Google Scholar] [CrossRef] [PubMed]
- Sousa, V.L.; Bellani, S.; Giannandrea, M.; Yousuf, M.; Valtorta, F.; Meldolesi, J.; Chieregatti, E. α-Synuclein and Its A30P Mutant Affect Actin Cytoskeletal Structure and Dynamics. Mol Biol Cell 2009, 20, 3725–3739. [Google Scholar] [CrossRef]
- Araki, K.; Sugawara, K.; Hayakawa, E.H.; Ubukawa, K.; Kobayashi, I.; Wakui, H.; Takahashi, N.; Sawada, K.; Mochizuki, H.; Nunomura, W. The Localization of α-Synuclein in the Process of Differentiation of Human Erythroid Cells. Int J Hematol 2018, 108, 130–138. [Google Scholar] [CrossRef]
- Liu, H.; Liu, W.; Wu, Y.; Zhou, Y.; Xue, R.; Luo, C.; Wang, L.; Zhao, W.; Jiang, J.D.; Liu, J. Loss of Epigenetic Control of Synuclein-γ Gene as a Molecular Indicator of Metastasis in a Wide Range of Human Cancers. Cancer Res 2005, 65, 7635–7643. [Google Scholar] [CrossRef]
- Goh, K.W.; Say, Y.H. γ-Synuclein Confers Both pro-Invasive and Doxorubicin-Mediated pro-Apoptotic Properties to the Colon Adenocarcinoma LS 174T Cell Line. Tumor Biol. 2015, 36, 7947–7960. [Google Scholar] [CrossRef]
- Jia, T.; Liu, Y.E.; Liu, J.; Shi, Y.E. Stimulation of Breast Cancer Invasion and Metastasis by Synuclein gamma. Cancer Res 1999, 59, 742–747. [Google Scholar] [PubMed]
- Bruening, W.; Giasson, B.I.; Klein-Szanto, A.J.P.; Lee, V.M.Y.; Trojanowski, J.Q.; Godwin, A.K. Synucleins Are Expressed in the Majority of Breast and Ovarian Carcinomas and in Preneoplastic Lesions of the Ovary. Cancer 2000, 88, 2154–2163. [Google Scholar] [CrossRef]
- Raghavan, R.; White, C.L.; Rogers, B.; Coimbra, C.; Rushing, E.J. Alpha-Synuclein Expression in Central Nervous System Tumors Showing Neuronal or Mixed Neuronal/Glial Differentiation. J Neuropathol Exp Neurol 2000, 59, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, M.; Suzuki, S.O.; Doh-Ura, K.; Iwaki, T. α-Synuclein Is Expressed in a Variety of Brain Tumors Showing Neuronal Differentiation. Acta Neuropathol 2000, 99, 154–160. [Google Scholar] [CrossRef]
- Surguchov, A.; Palazzo, R.E.; Surgucheva, I. Gamma Synuclein: Subcellular Localization in Neuronal and Non-Neuronal Cells and Effect on Signal Transduction. Cell Motil Cytoskeleton 2001, 49, 218–228. [Google Scholar] [CrossRef]
- Pan, Z.Z.; Bruening, W.; Giasson, B.I.; Lee, V.M.Y.; Godwin, A.K. γ-Synuclein Promotes Cancer Cell Survival and Inhibits Stress- and Chemotherapy Drug-Induced Apoptosis by Modulating MAPK Pathways. J Biol Chem. 2002, 277, 35050–35060. [Google Scholar] [CrossRef]
- Zhou, C.Q.; Liu, S.; Xue, L.Y.; Wang, Y.H.; Zhu, H.X.; Lu, N.; Xu, N.Z. Down-Regulation of γ-Synuclein in Human Esophageal Squamous Cell Carcinoma. World J Gastroenterol 2003, 9, 1900–1903. [Google Scholar] [CrossRef]
- Fung, K.M.; Rorke, L.B.; Giasson, B.; Lee, V.M.Y.; Trojanowski, J.Q. Expression of alpha-, beta-, and gamma-Synuclein in Glial Tumors and Medulloblastomas. Acta Neuropathol. 2003, 106, 167–175. [Google Scholar] [CrossRef]
- Li, Z.; Sclabas, G.M.; Peng, B.; Hess, K.R.; Abbruzzese, J.L.; Evans, D.B.; Chiao, P.J. Overexpression of Synuclein-γ in Pancreatic Adenocarcinoma. Cancer 2004, 101, 58–65. [Google Scholar] [CrossRef]
- Yanagawa, N.; Tamura, G.; Honda, T.; Endoh, M.; Nishizuka, S.; Motoyama, T. Demethylation of the Synuclein γ Gene CpG Island in Primary Gastric Cancers and Gastric Cancer Cell Lines. Clinical Cancer Research 2004, 10, 2447–2451. [Google Scholar] [CrossRef]
- Iwaki, H.; Kageyama, S.; Isono, T.; Wakabayashi, Y.; Okada, Y.; Yoshimura, K.; Terai, A.; Arai, Y.; Iwamura, H.; Kawakita, M.; Yoshiki, T. Diagnostic Potential in Bladder Cancer of a Panel of Tumor Markers (Calreticulin, γ-Synuclein, and Catechol-o-Methyltransferase) Identified by Proteomic Analysis. Cancer Sci 2004, 95, 955–956. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Zheng, M.H.; Cai, Q.; Feng, B.; Chen, X.H.; Yu, B.Q.; Gao, Y.B.; Ji, J.; Lu, A.G.; Li, J.W.; Wang, M.L.; Liu, B.Y. Aberrant Expression and Demethylation of γ-Synuclein in Colorectal Cancer, Correlated with Progression of the Disease. Cancer Sci 2008, 99, 1924–1932. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Wang, T.F.; Peng, Y.F.; Xie, J.; Feng, B.; Qiu, M.Y.; Li, L.H.; Lu, A.G.; Liu, B.Y.; Zheng, M.H. Expression of α-, β- and γ-Synuclein in Colorectal Cancer, and Potential Clinical Significance in Progression of the Disease. Oncol Rep 2010, 23, 429–436. [Google Scholar] [PubMed]
- Morgan, J.; Hoekstra, A. V.; Chapman-Davis, E.; Hardt, J.L.; Kim, J.J.; Buttin, B.M. Synuclein-γ (SNCG) May Be a Novel Prognostic Biomarker in Uterine Papillary Serous Carcinoma. Gynecol Oncol 2009, 114, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Kamitani, T. Parkinson’s Disease-Related Protein, α-Synuclein, in Malignant Melanoma. PLoS One 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Maitta, R.W.; Wolgast, L.R.; Wang, Q.; Zhang, H.; Bhattacharyya, P.; Gong, J.Z.; Sunkara, J.; Albanese, J.M.; Pizzolo, J.G.; A. Cannizzaro, L.; Ramesh, K.H.; Ratech, H. Alpha- and Beta-Synucleins Are New Diagnostic Tools for Acute Erythroid Leukemia and Acute Megakaryoblastic Leukemia. Am J Hematol. 2011, 86, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Mhawech-Fauceglia, P.; Wang, D.; Syriac, S.; Godoy, H.; Dupont, N.; Liu, S.; Odunsi, K. Synuclein-γ (SNCG) Protein Expression Is Associated with Poor Outcome in Endometrial Adenocarcinoma. Gynecol Oncol. 2012, 124, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; She, F.; Wang, D.; Yao, X.; Jiang, L.; Chen, Y. SNCG Gene Silencing in Gallbladder Cancer Cells Inhibits Key Tumorigenic Activities. Front Biosci. 2012, 17, 1589–1598. [Google Scholar] [CrossRef]
- Cheng, J.C.; Chiang, M.T.; Lee, C.H.; Liu, S.Y.; Chiu, K.C.; Chou, Y.T.; Huang, R.Y.; Huang, S.M.; Shieh, Y.S. γ-Synuclein Expression Is a Malignant Index in Oral Squamous Cell Carcinoma. J Dent Res 2016, 95, 439–445. [Google Scholar] [CrossRef]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An Enhanced Web Server for Large-Scale Expression Profiling and Interactive Analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef]
- Turriani, E.; Lázaro, D.F.; Ryazanov, S.; Leonov, A.; Giese, A.; Schön, M.; Schön, M.P.; Griesinger, C.; Outeiro, T.F.; Arndt-Jovin, D.J.; Becker, D. Treatment with Diphenyl-Pyrazole Compound Anle138b/c Reveals That α-Synuclein Protects Melanoma Cells from Autophagic Cell Death. Proc Natl Acad Sci U S A 2017, 114, E4971–E4977. [Google Scholar] [CrossRef] [PubMed]
- Bianchini, M.; Giambelluca, M.; Scavuzzo, M.C.; Di Franco, G.; Guadagni, S.; Palmeri, M.; Furbetta, N.; Gianardi, D.; Costa, A.; Gentiluomo, M.; Gaeta, R.; Pollina, L.E.; Falcone. A.; Vivaldi, C.; Di Candio, G.; Biagioni, F.; Busceti, C.L.; Soldani, P.; Puglisi-Allegra, S.; Morelli, L.; Fornai, F. In Pancreatic Adenocarcinoma Alpha-Synuclein Increases and Marks Peri-Neural Infiltration. Int J Mol Sci. 2022, 23, 3775. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wu, Z.; Ma, K. SNCA Correlates with Immune Infiltration and Serves as a Prognostic Biomarker in Lung Adenocarcinoma. BMC Cancer 2022, 22, 406. [Google Scholar]
- Becker, C.; Brobert, G.P.; Johansson, S.; Jick, S.S.; Meier, C.R. Cancer Risk in Association with Parkinson Disease: A Population-Based Study. Parkinsonism Relat Disord. 2010, 16, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Shou, C.; Meng, L.; Jiang, B.; Dong, B.; Yao, L.; Xie, Y.; Zhang, J.; Chen, Y.; Budman, D.R.; Shi, Y.E. Neuronal Protein Synuclein γ Predicts Poor Clinical Outcome in Breast Cancer. Int J Cancer 2007, 121, 1296–1305. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Dong, B.; Lu, A.; Qu, L.; Xing, X.; Meng, L.; Wu, J.; Eric Shi, Y.; Shou, C. Synuclein Gamma Predicts Poor Clinical Outcome in Colon Cancer with Normal Levels of Carcinoembryonic Antigen. BMC Cancer 2010, 10, 359. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Qu, L.; Dong, B.; Xing, X.; Ren, T.; Zeng, Y.; Jiang, B.; Meng, L.; Wu, J.; Shou, C. Combined Phenotype of 4 Markers Improves Prognostic Value of Patients with Colon Cancer. Am J Med Sci. 2012, 343, 295–302. [Google Scholar] [CrossRef]
- Taguchi, K.; Watanabe, Y.; Tsujimura, A.; Tanaka, M. Expression of α-Synuclein Is Regulated in a Neuronal Cell Type-Dependent Manner. Anat Sci Int. 2019, 94, 11–22. [Google Scholar] [CrossRef]
- Lee Clough, R.; Stefanis, L. A Novel Pathway for Transcriptional Regulation of alpha-synuclein. FASEB J. 2007, 21, 596–607. [Google Scholar] [CrossRef]
- Mittal, S.; Bjørnevik, K.; Im, D.S.; Flierl, A.; Dong, X.; Locascio, J.J.; Abo, K.M.; Long, E.; Jin, M.; Xu, B.; Xiang, Y.K.; Rochet, J.C.; Engeland, A.; Rizzu, P.; Heutink, P.; Bartels, T.; Selkoe, D.J.; Caldarone, B.J.; Glicksman, M.A.; Khurana, V.; Schüle, B.; Park, D.S.; Riise, T.; Scherzer, C.R. Β2-Adrenoreceptor Is a Regulator of the α-Synuclein Gene Driving Risk of Parkinson’s Disease. Science (1979) 2017, 357, 891–898. [Google Scholar] [CrossRef]
- Gómez-Santos, C.; Barrachina, M.; Giménez-Xavier, P.; Dalfó, E.; Ferrer, I.; Ambrosio, S. Induction of C/EBPβ and GADD153 Expression by Dopamine in Human Neuroblastoma Cells: Relationship with α-Synuclein Increase and Cell Damage. Brain Res Bull. 2005, 65, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Surguchov, A. Molecular and Cellular Biology of Synucleins. Int Rev Cell Mol Biol. 2008, 270, 225–317. [Google Scholar] [PubMed]
- McHugh, P.C.; Wright, J.A.; Brown, D.R. Transcriptional Regulation of the Beta-Synuclein 5′-Promoter Metal Response Element by Metal Transcription Factor-1. PLoS One 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yin, Y.; Hua, H.; Sun, X.; Luo, T.; Wang, J.; Jiang, Y. The Reciprocal Regulation of γ-Synuclein and IGF-I Receptor Expression Creates a Circuit That Modulates IGF-I Signaling. J Biol Chem. 2010, 285, 30480–30488. [Google Scholar] [CrossRef]
- Lu, A.; Zhang, F.; Gupta, A.; Liu, J. Blockade of AP1 Transactivation Abrogates the Abnormal Expression of Breast Cancer-Specific Gene 1 in Breast Cancer Cells. J Biol Chem. 2002, 277, 31364–31372. [Google Scholar] [CrossRef]
- Shao, T.; Song, P.; Hua, H.; Zhang, H.; Sun, X.; Kong, Q.; Wang, J.; Luo, T.; Jiang, Y. Gamma Synuclein Is a Novel Twist1 Target That Promotes TGF-β-Induced Cancer Cell Migration and Invasion Article. Cell Death Dis. 2018, 9, 625. [Google Scholar] [CrossRef]
- Foulds, P.G.; Mitchell, J.D.; Parker, A.; Turner, R.; Green, G.; Diggle, P.; Hasegawa, M.; Taylor, M.; Mann, D.; Allsop, D. Phosphorylated A-synuclein Can Be Detected in Blood Plasma and Is Potentially a Useful Biomarker for Parkinson’s Disease. FASEB J. 2011, 25, 4127–4137. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.; Li, J. D. The Roles of Post-Translational Modifications on α-Synuclein in the Pathogenesis of Parkinson’s Diseases. Front Neurosci. 2019, 13, 381. [Google Scholar] [CrossRef]
- Cole, R.N.; Hart, G.W. Cytosolic O-Glycosylation Is Abundant in Nerve Terminals. J Neurochem. 2001, 79, 1080–1089. [Google Scholar] [CrossRef]
- Surgucheva, I.; Newell, K.L.; Burns, J.; Surguchov, A. New α- and γ-Synuclein Immunopathological Lesions in Human Brain. Acta Neuropathol Commun. 2013, 2, 132. [Google Scholar] [CrossRef]
- Gong, H.; Yang, X.; Zhao, Y.; Petersen, R.; Liu, X.; Liu, Y.; Huang, K. Amyloidogenicity of P53: A Hidden Link Between Protein Misfolding and Cancer. Curr Protein Pept Sci. 2014, 16, 135–146. [Google Scholar] [CrossRef]
- Ano Bom, A.P.D.; Rangel, L.P.; Costa, D.C.F.; de Oliveira, G.A.P.; Sanches, D.; Braga, C.A.; Gava, L.M.; Ramos, C.H.I.; Cepeda, A.O.T.; Stumbo, A.C.; De Moura Gallo, C.V.; Cordeiro, Y.; Silva, J.L. Mutant P53 Aggregates into Prion-like Amyloid Oligomers and Fibrils. Journal of Biological Chemistry 2012, 287, 28152–28162. [Google Scholar] [CrossRef] [PubMed]
- Iwahashi, N.; Ikezaki, M.; Nishikawa, T.; Namba, N.; Ohgita, T.; Saito, H.; Ihara, Y.; Shimanouchi, T.; Ino, K.; Uchimura, K.; Nishitsuji, K. Sulfated Glycosaminoglycans Mediate Prion-like Behavior of P53 Aggregates. Proc Natl Acad Sci U S A 2020, 117, 33225–33234. [Google Scholar] [CrossRef] [PubMed]
- Claes, F.; Maritschnegg, E.; Baets, G. De; Siekierska, A.; Rubio, M.S.; Ramakers, M.; Michiels, E.; Smet, F. De; Depreeuw, J.; Vergote, I.; Vanderstichele, A.; van den Broeck, A.; Olbrecht, S.; Hermans, E.; Amant, F.; Lambrechts, D.; Nilsson, P. R.; Rousseau, F.; Schymkowitz, J. The Tumor Suppressor Protein PTEN Undergoes Amyloid-like Aggregation in Tumor Cells. bioRxiv. 2020, 11, 402115. [Google Scholar]
- Kehrloesser, S.; Osterburg, C.; Tuppi, M.; Schäfer, B.; Vousden, K.H.; Dötsch, V. Intrinsic Aggregation Propensity of the P63 and P73 TI Domains Correlates with P53R175H Interaction and Suggests Further Significance of Aggregation Events in the P53 Family. Cell Death Differ. 2016, 23, 1952–1960. [Google Scholar] [CrossRef]
- Direito, I.; Monteiro, L.; Melo, T.; Figueira, D.; Lobo, J.; Enes, V.; Moura, G.; Henrique, R.; Santos, M.A.S.; Jerónimo, C.; Amado F, Fardilha M, Helguero LA. Protein Aggregation Patterns Inform about Breast Cancer Response to Antiestrogens and Reveal the Rna Ligase Rtcb as Mediator of Acquired Tamoxifen Resistance. Cancers (Basel) 2021, 13, 3195. [Google Scholar] [CrossRef]
- Matafora, V.; Farris, F.; Restuccia, U.; Tamburri, S.; Martano, G.; Bernardelli, C.; Sofia, A.; Pisati, F.; Casagrande, F.; Lazzari, L.; Marsoni, S.; Bonoldi, E.; Bachi, A. Amyloid Aggregates Accumulate in Melanoma Metastasis Modulating YAP Activity. EMBO Rep. 2020, 21. [Google Scholar] [CrossRef]
- Genovese, I.; Fornetti, E.; Ruocco, G. Mitochondria Inter-Organelle Relationships in Cancer Protein Aggregation. Front Cell Dev Biol. 2022, 10. [Google Scholar] [CrossRef]
- Shekoohi, S.; Rajasekaran, S.; Patel, D.; Yang, S.; Liu, W.; Huang, S.; Yu, X.; Witt, S.N. Knocking out Alpha-Synuclein in Melanoma Cells Dysregulates Cellular Iron Metabolism and Suppresses Tumor Growth. Sci Rep. 2021, 11, 5267. [Google Scholar] [CrossRef]
- Dean, D.N.; Lee, J.C. Linking Parkinson’s Disease and Melanoma: Interplay Between α-Synuclein and Pmel17 Amyloid Formation. Mov Disord. 2021, 36, 1489–1498. [Google Scholar] [CrossRef]
- Pavlenko, T.A.; Roman, A.Y.; Lytkina, O.A.; Pukaeva, N.E.; Everett, M.W.; Sukhanova, I.S.; Soldatov, V.O.; Davidova, N.G.; Chesnokova, N.B.; Ovchinnikov, R.K.; et al. Gamma-Synuclein Dysfunction Causes Autoantibody Formation in Glaucoma Patients and Dysregulation of Intraocular Pressure in Mice. Biomedicines 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Sugama, S.; Nakai, M.; Takenouchi, T.; Wei, J.; Urano, T.; Inoue, S.; Hashimoto, M. α-Synuclein Stimulates Differentiation of Osteosarcoma Cells: Relevance to down-Regulation of Proteasome Activity. J Biol Chem. 2007, 282, 5736–5748. [Google Scholar] [CrossRef] [PubMed]
- Israeli, E.; Yakunin, E.; Zarbiv, Y.; Hacohen-Solovich, A.; Kisos, H.; Loeb, V.; Lichtenstein, M.; Ben-Gedalya, T.; Sabag, O.; Pikarsky, E.; et al. α-Synuclein Expression Selectively Affects Tumorigenesis in Mice Modeling Parkinson’s Disease. PLoS One 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Xu, K. Alpha-Synuclein Contributes to Malignant Progression of Human Meningioma via the Akt/MTOR Pathway. Cancer Cell Int. 2016, 16. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Chen, X.; Huang, S.; Li, G.; Mo, M.; Zhang, L.; Chen, C.; Guo, W.; Zhou, M.; Wu, Z.; Cen, L.; Long, S.; Li, S.; Yang, X.; Qu, S.; Pei, Z.; Xu, P. Iron Promotes α-Synuclein Aggregation and Transmission by Inhibiting TFEB-Mediated Autophagosome-Lysosome Fusion. J Neurochem 2018, 145, 34–50. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.S.; Kim, Y.M.; Junn, E.; Lee, G.; Park, K.-H.; Tanaka, M.; Ronchetti, R.D.; Quezado, M.M.; Mouradian, M.M. Cell Cycle Aberrations by α-Synuclein over-Expression and Cyclin B Immunoreactivity in Lewy Bodies. Neurobiol Aging 2003, 24, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.M.; Huang, Y.X.; Li, X.L.; Chen, C.; Shi, Q.; Wang, G.R.; Tian, C.; Wang, Z.Y.; Jing, Y.Y.; Gao, C.; Dong, X.P. Molecular Interaction of α-Synuclein with Tubulin Influences on the Polymerization of Microtubule in Vitro and Structure of Microtubule in Cells. Mol Biol Rep 2010, 37, 3183–3192. [Google Scholar] [CrossRef]
- Koss, D.J.; Erskine, D.; Porter, A.; Palmoski, P.; Menon, H.; Todd, O.G.J.; Leite, M.; Attems, J.; Outeiro, T.F. Nuclear Alpha-Synuclein Is Present in the Human Brain and Is Modified in Dementia with Lewy Bodies. Acta Neuropathol Commun. 2022, 10. [Google Scholar] [CrossRef]
- Thorne, N.J.; Tumbarello, D.A. The Relationship of Alpha-Synuclein to Mitochondrial Dynamics and Quality Control. Front Mol Neurosci. 2022, 15. [Google Scholar] [CrossRef]
- Gupta, A.; Inaba, S.; Wong, O.K.; Fang, G.; Liu, J. Breast Cancer-Specific Gene 1 Interacts with the Mitotic Checkpoint Kinase BubR1. Oncogene 2003, 22, 7593–7599. [Google Scholar] [CrossRef]
- Chen, J.; Jiao, L.; Xu, C.; Yu, Y.; Zhang, Z.; Chang, Z.; Deng, Z.; Sun, Y. Neural Protein Gamma-Synuclein Interacting with Androgen Receptor Promotes Human Prostate Cancer Progression. BMC Cancer 2012, 12, 593. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Niu, J.; Sun, E.; Rong, X.; Zhang, X.; Ju, Y. Gamma-Synuclein Binds to AKT and Promotes Cancer Cell Survival and Proliferation. Tumor Biol. 2016, 37, 14999–15005. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Liu, J.; Tian, J.; Zhang, Y.; Yan, M.; Zhu, C. SiRNA Targeting of the SNCG Gene Inhibits the Growth of Gastric Carcinoma SGC7901 Cells in Vitro and in Vivo by Downregulating the Phosphorylation of AKT/ERK. Cytogenet Genome Res. 2018, 154, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Gu, L.; Li, X.; Wang, J. Silencing of Synuclein-γ Inhibits Human Cervical Cancer through the AKT Signaling Pathway. Cell Mol Biol Lett. 2019, 24, 49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, X.H.; Li, C.; Wu, X.X.; Chen, Y.L.; Li, W.W.; Li, X.; Gong, F.; Tang, Q.; Jiang, D. SNCG Promotes the Progression and Metastasis of High-Grade Serous Ovarian Cancer via Targeting the PI3K/AKT Signaling Pathway. J Exp and Clin Cancer Res. 2020, 39, 79. [Google Scholar] [CrossRef]
- Sun, D.; Li, W.Y.; Chen, S.H.; Zhi, Z.F.; Lin, H.S.; Fan, J.T.; Fan, Y.J. ShRNA-Mediated Suppression of γ-Synuclein Leading to Downregulation of P38/ERK/JNK Phosphorylation and Cell Cycle Arrest in Endometrial Cancer Cells. Mol Biol. 2020, 54, 1006–1017. [Google Scholar] [CrossRef]
- Chen, J.; Huang, S.; Wu, K. jin; Wang, Y. kun; Jia, Y. jun; Lu, Y. shu; Weng, Z. yi [The Correlation of Synuclein-γ and Matrix Metalloproteinase 9 in Breast Cancer]. Zhonghua Wai Ke Za Zhi 2013, 51, 641–644. [Google Scholar]
- Surgucheva, I.G.; Sivak, J.M.; Fini, M.E.; Palazzo, R.E.; Surguchov, A.P. Effect of γ-Synuclein Overexpression on Matrix Metalloproteinases in Retinoblastoma Y79 Cells. Arch Biochem Biophys. 2003, 410, 167–176. [Google Scholar] [CrossRef]
- Pan, Z.Z.; Bruening, W.; Godwin, A.K. Involvement of RHO GTPases and ERK in Synuclein-γ Enhanced Cancer Cell Motility. Int J Oncol. 2006, 29, 1201–1205. [Google Scholar] [CrossRef]
- Liu, C.; Qu, L.; Lian, S.; Tian, Z.; Zhao, C.; Meng, L.; Shou, C. Unconventional Secretion of Synuclein-γ Promotes Tumor Cell Invasion. FEBS J. 2014, 281, 5159–5171. [Google Scholar] [CrossRef]
- Zhao, J.; Xing, N. Identification of γ-Synuclein as a Stage-Specific Marker in Bladder Cancer by Immunohistochemistry. Med Sci Monit. 2014, 20, 2550–2555. [Google Scholar] [PubMed]
- Chen, Z.; Ji, Z.; Wang, Q.; Shi, B.; Shou, C.; Liu, C.; Fan, H.; Li, H.; Davidson, K.T.; Wakefield, M.R.; Ball, T.W.; Fang, Y. Expression of γ-Synuclein in Bladder Carcinoma: A Possible Marker for Prognosis. Anticancer Res. 2016, 36, 951–956. [Google Scholar] [PubMed]
- Takemura, Y.; Ojima, H.; Oshima, G.; Shinoda, M.; Hasegawa, Y.; Kitago, M.; Yagi, H.; Abe, Y.; Hori, S.; Fujii-Nishimura, Y.; Kubota, N.; Masuda, Y.; Hibi, T.; Sakamoto, M.; Kitagawa, Y. Gamma-synuclein is a Novel Prognostic Marker that Promotes Tumor Cell Migration in Biliary Tract Carcinoma. Cancer Med. 2021, 10, 5599–5613. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structure of synucleins, phylogeny and expression in human tissues. (A) Multiple sequence alignment of human synucleins by Clustal W. Sequences were obtained from NCBI (accession numbers CAG33339.1; CAG33308.1 and CAG46587.1). “*” indicates identical amino acids; “:” and “.” indicate conserved and semi-conserved residues, respectively. Each synuclein is organized in a tripartite arrangement, with the N-terminal region (light violet), central NAC region (dark blue), and the C-terminal region (light green). Amino acids involved in aggregation are marked in light blue. 11-mer repeats are underlined. Amino acid numbers are displayed at right. (B) Human synucleins predicted structure by Pymol. αS full-length protein structure was obtained from PDB (Protein Data Bank) with the accession code: 1XQ8 (https://doi.org/10.2210/pdb1XQ8/pdb). The UniProt accession codes Q16143 and Q6FHG5 were used to predict βS and γS full-length protein structure using AlphaFold, and obtained in PDB format. Different regions of the proteins are highlighted in colors, according to (A). (C) Evolutionary tree of synucleins. αS, βS, and γS found in different branches of jawed vertebrates are shown. As synucleins were not reported in Caudata (amphibians) and Sphenodon (reptilia) genders, they are not represented. (D) Mammal synucleins genomic tree. The common node between αS (pink) and βS (green) is shown in blue. γS (violet) clade is shown in black. Homo Sapiens taxa are highlighted in red. FASTA files were downloaded from NCBI. Sequence alignment was generated with Molecular Evolutionary Genetics Analysis software (https://www.megasoftware.net/). The 311 sequences aligned were uploaded to the IQTREE WEB SERVER (http://iqtree.cibiv.univie.ac.at/), the best fit model according to AICc (Second-order Akaike's information criterion) JTT+G4 was used. The tree was generated using FigTree software (http://tree.bio.ed.ac.uk/software/figtree/). (E,F) Protein (E) and mRNA (F) synucleins expression in human tissues. Expression information was obtained from The Human Protein Atlas (https://www.proteinatlas.org/) and eighteen tissues were selected for plotting. Neuronal tissues are highlighted in violet. H: high; M: medium; L: low; ND: not detected; nTPM: normalized transcript per million.
Figure 1.
Structure of synucleins, phylogeny and expression in human tissues. (A) Multiple sequence alignment of human synucleins by Clustal W. Sequences were obtained from NCBI (accession numbers CAG33339.1; CAG33308.1 and CAG46587.1). “*” indicates identical amino acids; “:” and “.” indicate conserved and semi-conserved residues, respectively. Each synuclein is organized in a tripartite arrangement, with the N-terminal region (light violet), central NAC region (dark blue), and the C-terminal region (light green). Amino acids involved in aggregation are marked in light blue. 11-mer repeats are underlined. Amino acid numbers are displayed at right. (B) Human synucleins predicted structure by Pymol. αS full-length protein structure was obtained from PDB (Protein Data Bank) with the accession code: 1XQ8 (https://doi.org/10.2210/pdb1XQ8/pdb). The UniProt accession codes Q16143 and Q6FHG5 were used to predict βS and γS full-length protein structure using AlphaFold, and obtained in PDB format. Different regions of the proteins are highlighted in colors, according to (A). (C) Evolutionary tree of synucleins. αS, βS, and γS found in different branches of jawed vertebrates are shown. As synucleins were not reported in Caudata (amphibians) and Sphenodon (reptilia) genders, they are not represented. (D) Mammal synucleins genomic tree. The common node between αS (pink) and βS (green) is shown in blue. γS (violet) clade is shown in black. Homo Sapiens taxa are highlighted in red. FASTA files were downloaded from NCBI. Sequence alignment was generated with Molecular Evolutionary Genetics Analysis software (https://www.megasoftware.net/). The 311 sequences aligned were uploaded to the IQTREE WEB SERVER (http://iqtree.cibiv.univie.ac.at/), the best fit model according to AICc (Second-order Akaike's information criterion) JTT+G4 was used. The tree was generated using FigTree software (http://tree.bio.ed.ac.uk/software/figtree/). (E,F) Protein (E) and mRNA (F) synucleins expression in human tissues. Expression information was obtained from The Human Protein Atlas (https://www.proteinatlas.org/) and eighteen tissues were selected for plotting. Neuronal tissues are highlighted in violet. H: high; M: medium; L: low; ND: not detected; nTPM: normalized transcript per million.
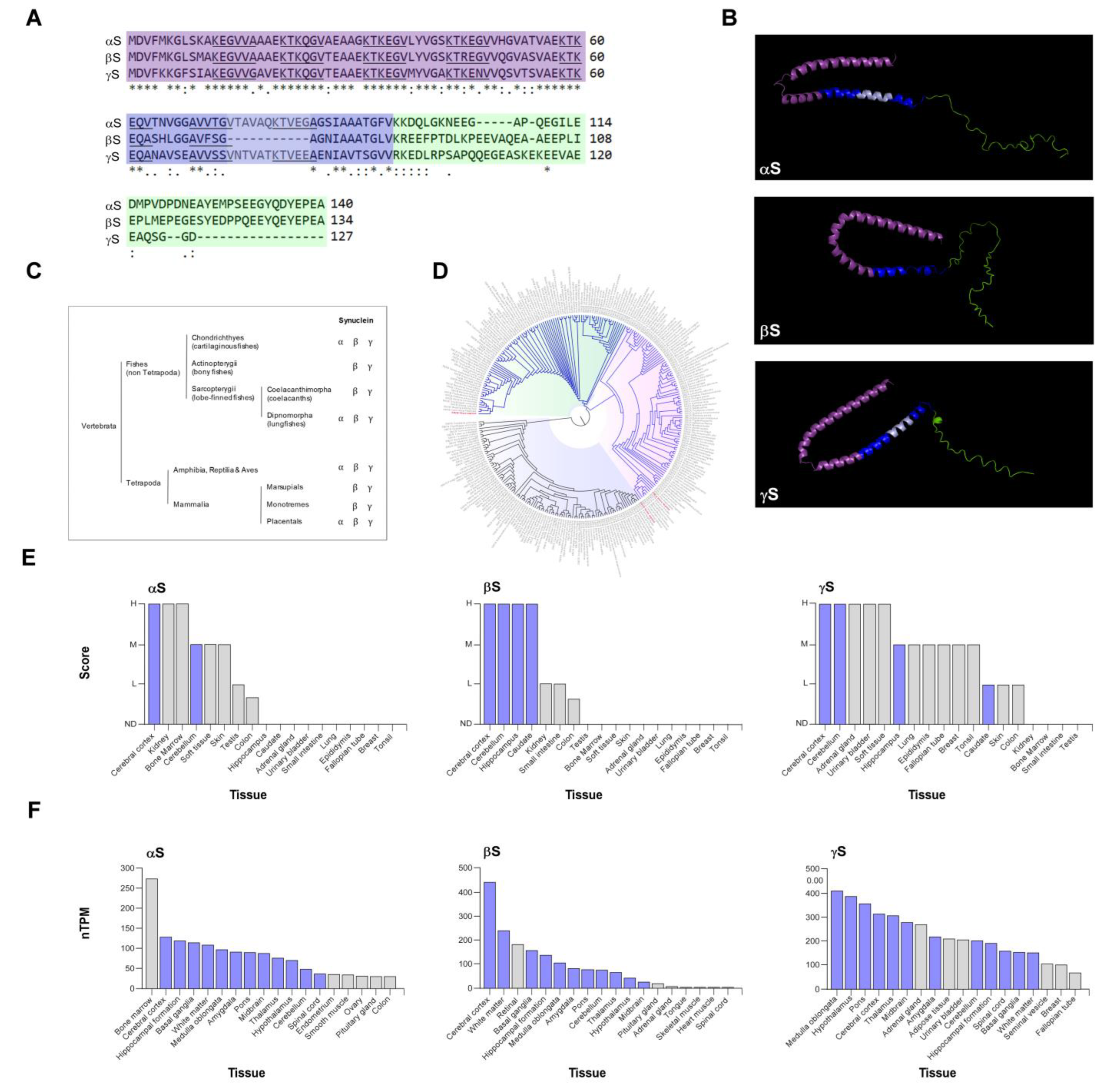
Figure 2.
Synucleins expression in cancer. (A) To identify synuclein-associated gene clusters, a search was conducted in the NCBI database, limited to Homo sapiens genes. A total of 323, 22, and 49 genes related to αS, βS, and γS, respectively, available in the NCBI database till April 27, 2023 were downloaded. Using these gene clusters, an enrichment analysis was performed, and a word cloud was generated to display the most common terms found in the results. Word size in the clouds is proportional to the frequency of occurrence in the over-representation analysis (p>0.05), and the thickness of the lines connecting clouds represents the number of shared terms. (B-D) Expression level of synucleins in tumor samples and noncancerous (normal) samples through GEPIA2 database (http://gepia2.cancer-pku.cn/). Light gray represents normal tissues (N) and light pink tumor tissues (T). The expression levels on the Y axis are expressed as log2(TPM + 1), TPM: transcript per million. The statistical analysis performed is T-Test. *p<0.01. (COAD: Colon adenocarcinoma; LUAD: Lung adenocarcinoma; PAAD: Pancreatic adenocarcinoma; SKCM: Skin Cutaneous Melanoma; GBM: Glioblastoma multiforme; LGG: Brain Lower Grade Glioma; BRCA: Breast invasive carcinoma; and OV: Ovarian serous cystadenocarcinoma). (n= number).
Figure 2.
Synucleins expression in cancer. (A) To identify synuclein-associated gene clusters, a search was conducted in the NCBI database, limited to Homo sapiens genes. A total of 323, 22, and 49 genes related to αS, βS, and γS, respectively, available in the NCBI database till April 27, 2023 were downloaded. Using these gene clusters, an enrichment analysis was performed, and a word cloud was generated to display the most common terms found in the results. Word size in the clouds is proportional to the frequency of occurrence in the over-representation analysis (p>0.05), and the thickness of the lines connecting clouds represents the number of shared terms. (B-D) Expression level of synucleins in tumor samples and noncancerous (normal) samples through GEPIA2 database (http://gepia2.cancer-pku.cn/). Light gray represents normal tissues (N) and light pink tumor tissues (T). The expression levels on the Y axis are expressed as log2(TPM + 1), TPM: transcript per million. The statistical analysis performed is T-Test. *p<0.01. (COAD: Colon adenocarcinoma; LUAD: Lung adenocarcinoma; PAAD: Pancreatic adenocarcinoma; SKCM: Skin Cutaneous Melanoma; GBM: Glioblastoma multiforme; LGG: Brain Lower Grade Glioma; BRCA: Breast invasive carcinoma; and OV: Ovarian serous cystadenocarcinoma). (n= number).
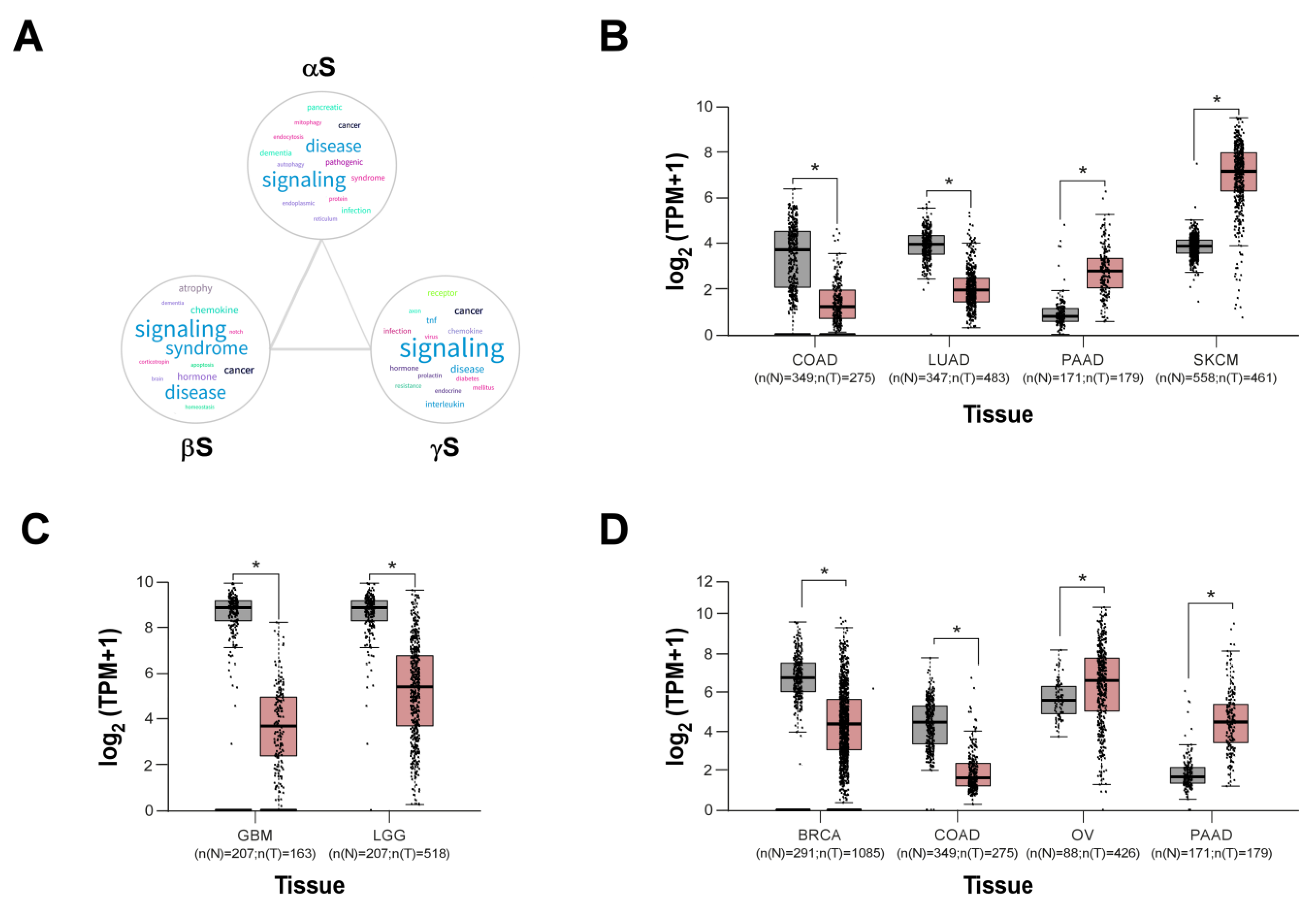
Figure 3.
Synucleins are involved in several cancer-related cellular processes. Graphic scheme illustrating key cancer-related cellular processes described for synucleins. Figure shows the main processes involved in a central cell and two neighbores. Light pink boxes represent processes involving αS; light violet boxes represent γS processes; light orange boxes represent processes in which both proteins are involved. αS involvement in autophagy, mitochondrial metabolism, and the generation of ROS is well established. Additionally, different aggregation states of αS play important roles in cancer. γS levels in tumor cells regulate microtubule dynamics, cell-cell adhesion, apoptosis, and influence signaling pathways involving ERK and AKT. Both synucleins were reported to be involved in cell proliferation and migration. In addition, both proteins can be secreted and transmitted to neighboring cells as monomer or oligomer through different mechanisms (by exosomes release, cross cell membrane by membrane diffusion, and/or attaching proteins that function as membrane receptors), inducing the aggregation and accumulation of cytosolic synuclein in the proximal cell via prion-like properties, which could have important implications in cancer progression. Depicted organelles and structures include mitochondria, autolysosomes, exosomes, membrane proteins, adhesion proteins, and the cytoskeleton (tubulin and actin filaments). (MMP: extracellular matrix metalloproteinases, ROS: mitochondrial reactive oxygen species).
Figure 3.
Synucleins are involved in several cancer-related cellular processes. Graphic scheme illustrating key cancer-related cellular processes described for synucleins. Figure shows the main processes involved in a central cell and two neighbores. Light pink boxes represent processes involving αS; light violet boxes represent γS processes; light orange boxes represent processes in which both proteins are involved. αS involvement in autophagy, mitochondrial metabolism, and the generation of ROS is well established. Additionally, different aggregation states of αS play important roles in cancer. γS levels in tumor cells regulate microtubule dynamics, cell-cell adhesion, apoptosis, and influence signaling pathways involving ERK and AKT. Both synucleins were reported to be involved in cell proliferation and migration. In addition, both proteins can be secreted and transmitted to neighboring cells as monomer or oligomer through different mechanisms (by exosomes release, cross cell membrane by membrane diffusion, and/or attaching proteins that function as membrane receptors), inducing the aggregation and accumulation of cytosolic synuclein in the proximal cell via prion-like properties, which could have important implications in cancer progression. Depicted organelles and structures include mitochondria, autolysosomes, exosomes, membrane proteins, adhesion proteins, and the cytoskeleton (tubulin and actin filaments). (MMP: extracellular matrix metalloproteinases, ROS: mitochondrial reactive oxygen species).
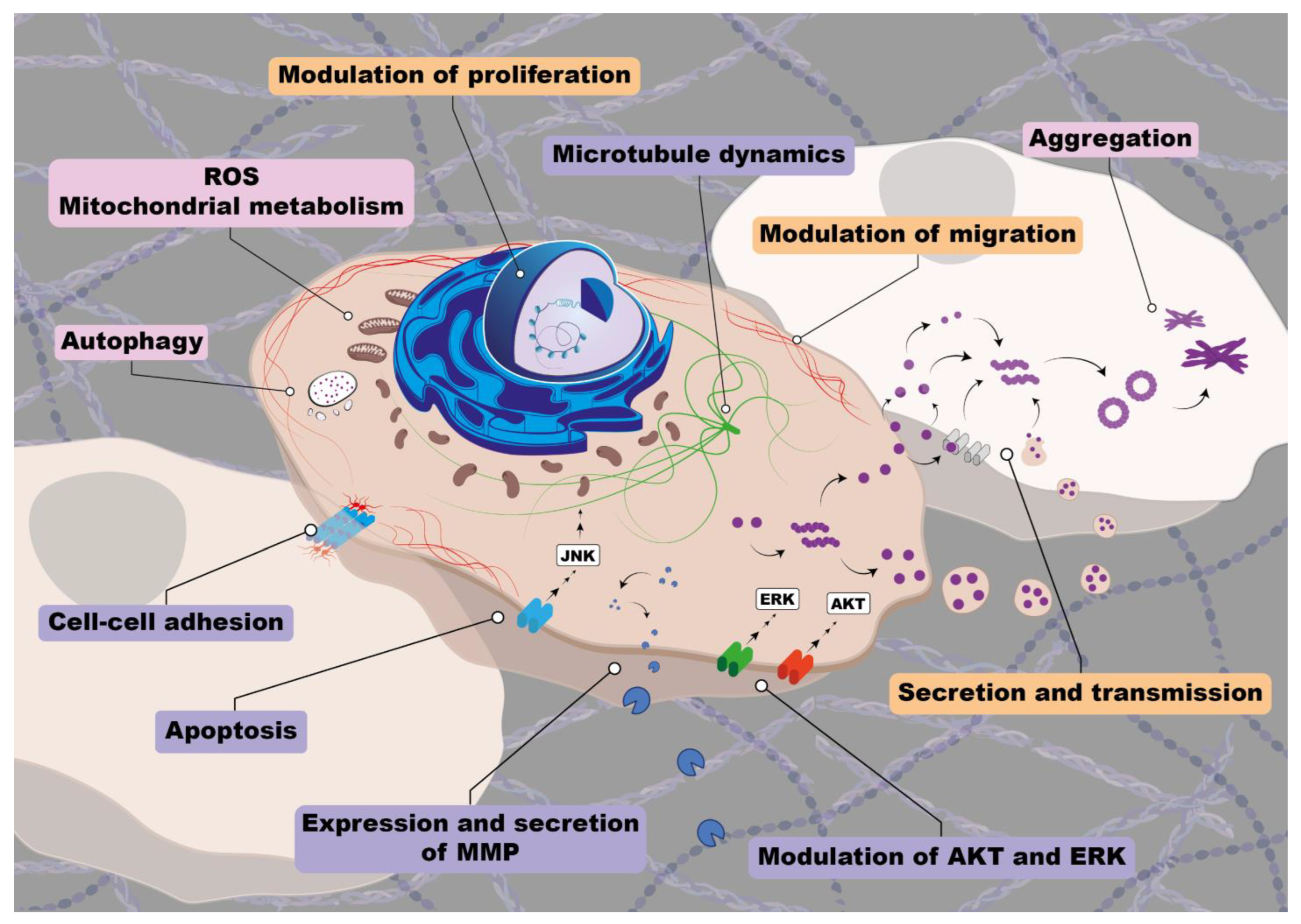
Figure 4.
Correlation between synucleins expression and overall survival. (A-C) Association between synucleins expression and overall survival (OS) using TCGA (The Cancer Genome Atlas) mRNA expression datasets for different tumors. Kaplan–Meier curves for OS of cancer patients with low (gray line) versus high (violet line) expression of αS (A), βS (B), and γS (C) were generated using Survminer R package (p<0.05) and compared by log-rank tests. The number of patients for each case is described in the figure (n). The x-axis depicts time in days. (COAD: Colon adenocarcinoma; LUAD: Lung adenocarcinoma; SKCM: Skin Cutaneous Melanoma; LGG: Brain Lower Grade Glioma; BRCA: Breast invasive carcinoma; and PCPG: Pheochromocytoma and Paraganglioma and UCEC: Cervical squamous cell carcinoma and endocervical adenocarcinoma).
Figure 4.
Correlation between synucleins expression and overall survival. (A-C) Association between synucleins expression and overall survival (OS) using TCGA (The Cancer Genome Atlas) mRNA expression datasets for different tumors. Kaplan–Meier curves for OS of cancer patients with low (gray line) versus high (violet line) expression of αS (A), βS (B), and γS (C) were generated using Survminer R package (p<0.05) and compared by log-rank tests. The number of patients for each case is described in the figure (n). The x-axis depicts time in days. (COAD: Colon adenocarcinoma; LUAD: Lung adenocarcinoma; SKCM: Skin Cutaneous Melanoma; LGG: Brain Lower Grade Glioma; BRCA: Breast invasive carcinoma; and PCPG: Pheochromocytoma and Paraganglioma and UCEC: Cervical squamous cell carcinoma and endocervical adenocarcinoma).

Table 1.
Selected reports describing the involvement of synucleins in cancer.
| Synucleins | Year | Tumor type | Type of study | Reference |
| γS | 1997 | Breast | Patients | [6] |
| γS | 1999 | Breast | Cell lines | [41] |
| βS, γS | 2000 | Breast, Ovarian | Cell lines and patients | [42] |
| αS | 2000 | Ganglioglioma, Ganglioneuroblastoma | Patients | [43] |
| αS | 2000 | Medulloblastoma, Pineocytoma, Pineoblastoma | Patients | [44] |
| γS | 2001 | Retinoblastoma | Cell lines | [45] |
| γS | 2001 | Ovarian | Cell lines | [46] |
| γS | 2003 | Esophageal | Cell lines and patients | [47] |
| αS, βS | 2003 | Glial, Medulloblastoma | Patients | [48] |
| γS | 2004 | Pancreatic | Cell lines and patients | [49] |
| γS | 2004 | Gastric | Cell lines and patients | [50] |
| γS | 2004 | Bladder | Patients | [51] |
| γS | 2005 | Prostate, Cervical, Colon, Lung | Patients | [39] |
| αS | 2007 | Osteosarcoma | Cell lines | [92] |
| γS | 2008 | Colon | Cell lines and patients | [52] |
| γS | 2009 | Uterine serous papillary carcinoma | Cell lines and patients | [54] |
| αS, βS, γS | 2010 | Colon | Cell lines and patients | [53] |
| αS | 2010 | Melanoma | Cell lines and patients | [55] |
| αS, βS | 2011 | Leukemia | Cell lines and patients | [56] |
| αS | 2011 | Melanoma, Breast, Lung | Cell lines and mice | [93] |
| γS | 2012 | Endometrium | Patients | [57] |
| γS | 2016 | Squamous cell carcinoma of the oral cavity | Cell lines and patients | [59] |
| αS | 2016 | Meningioma | Cell lines and patients | [94] |
| γS | 2021 | Biliary tract | Cell lines and patients | [113] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



