
Submitted:
03 May 2023
Posted:
08 May 2023
You are already at the latest version
Abstract
The paper summarizes the current knowledge about the structure, species, and content differ-ences of β-D-glucans as a storage and functional component, especially of cereal grains. The pa-per also provides comprehensive information on the biosynthesis of this plant cell wall polysac-charide, genetic and non-genetic factors influencing the content variability and indicates the pos-sibilities of breeding modern varieties with a defined content of this metabolite. The final part of the text is also devoted to the influence of significant technological processes applied to cere-als containing β-D-glucans. The chapter discusses the influence of these processes on the content and structure of the polysaccharide and the resulting changes in its physicochemical properties. Part of the discussed issue are the possibilities of effective application of β-D-glucans in different food products.
Keywords:
β-D-glucans
; cereals
; biosynthesis
; food application
1. Basic Characterization and Localization of β-D-Glucans in the Plant
The (1,3-1,4)-β-D-glucans (hereafter referred as β-D-glucans) are relatively small part components of the cell wall of vegetative tissues of cereals and grasses [1,2]. Mainly cereal grains such as barley (Hordeum vulgare L.), oat (Avena sativa L.), and rye (Secale cereale L.) are rich sources of β-D-glucans, while wheat (Triticum aestivum L.), rice (Oryza sativa L.), and maize (Zea mays L.) dispose of lower concentrations of this polysaccharide [3,4,5]. The highest concentrations of β-D-glucans are found in the outer layers of the grain [6,7].
Generally, the accumulation of β-D-glucans is observed in the cell wall of endosperm cells of the developing grains and in the surrounding maternal tissues, the aleuronal and subaleuronal layer [6,8,9]. This polysaccharide has also been found in vegetative organs of the plant, namely, in the root, coleoptile, stem, and leaf [10,11,12] and variability in the content of this polysaccharide was found during plant and tissues development [12,13]. β-D-glucans are not uniformly distributed in the grain, their localization varies among plant species and different plant tissues [9].
The β-D-glucans are unsubstituted, unbranched polysaccharides composed of β-D-glucopyranosyl monomers polymerized by both β-(1,3)- and β-(1,4)-linkages [14,15] and therefore the cereal β-D-glucans are also called as “mixed-linkage glucans - MLG”. From a functional point of view, the most important feature of this molecule is the arrangement of β-(1,3)- and β-(1,4)-linkages along the polysaccharide chain [1,2,16]. The bonds are not arranged in regularly repeating sequences, but they are also not arranged randomly [16,17,18]. (1,4)-β-bonds are usually more often presented in the polysaccharide than (1,3). (1,3)-β-D-glucosyl residues always occur in linear β-D-glucans chain as individual parts between (1,4)-β-D-oligoglucosyl units, which are mostly found in sequences of two or three [19].
The cereal β-D-glucan chain consists of glucopyranosyl monomers linked by a β-(1-4)-glycosidic bond in blocks of three or four monomers, which are called cellotriosyl and cellotetraosyl units. These units are separated by a single β-(1-3) bond, giving the chain a “staircase” like structure [20,21]. Adjacent (1,3)-β-D-glucosyl residues are not present, at least not in cereal β-D-glucans [1]. Adjacent (1,4)-β-D-oligoglucosyl units located between individual (1,3)-β-D-glucosyl residues can be considered as cellodextrin units, which usually consist of two or three adjacent (1,4)-β-D-glucosyl residues [1]. Most β-D-glucans in grasses have longer cellodextrin units, which consist of five to 20 adjacent (1,4)-β-D-oligoglucosyl units, which together make up to 10% polysaccharide chain [1]. Therefore, β-D-glucans in grasses can be considered as (1,3)-β-linked copolymers of cellotriosyl units, cellotetraosyl units, and longer (1,4)-β-D-oligoglucosyl units in which the ratio of cellotriosyl (DP3) to cellotetraosyl (DP4) units (the ratio of β-(1-3) to β-(1-4) units) ranges from 1.5 to 4.5 depending on the source of β-D-glucans [18] with an exception in sorghum endosperm having the ratio 1.15: 1 [3]. In barley, the ratio is 2.2 to 2.6: 1 [1] or 1.8 -3.5:1 [22], in wheat 3.0 to 4.5: 1, rye (1.9-3.0), and in oats 1.5 to 2.3: 1 [3,22].
The degree of polymerization (DP) of common β-D-glucans in grasses is about 1000 or more [23]. It is a unique feature of the β-D-glucans of each cereal and affects the solubility and viscosity and thus the physicochemical properties and applications of the polysaccharide in solution. For example, oat β-D-glucans with a lower molar ratio (1.5-2.3:1) are more soluble than barley and wheat β-D-glucans with a higher molar ratio (2.6:1 and 3.2: 1) [24]. Differences can be observed in the same genera as genotypic variability [2]. Environmental factors such as the conditions of cultivation also affect the degree of polymerization, whereas oat varieties disposing of higher content of β-D-glucans and cultivated in drier environment showed lower degree of polymerization [25].
The β-D-glucans found in cereals share the same molecular structure regardless of which source they are isolated from, but certain characteristics are specific to the source of this molecule. These characteristics are, for example, the presence and amount of long cellulose fragments, the ratio between β-(1-4) and β-(1-3) linkages, molecular size, and the ratio of cellotriosyl and cellotetraosyl units [26]. The molecular weight is approximately 31-2700 x 103 g/mol for barley, 65-3100 x 103 g/mol for oat, 21-1100 x 103 g/mol for rye, and 43-758 x 103 g/mol for wheat [22]. For sorghum it is 36 x 103 g/mol [27]. In the case of DP3: DP4 ratio, narrow range is for example observed in domestic cultivars of Avena sativa L. (2.05–2.11) compared to other cultivars of the genus Avena (1.81–2.33) [28]. The relative amount of the trisaccharide (DP3) in β-D-glucans decreases from wheat (67-72%) to barley (52 –69%) and oats (53-61%), while the relative amount of tetrasaccharide (DP4) has the opposite effect trend, the growth from wheat (21–24%), through barley (25–33%) and oats (34–41%) [22]. Some structural differences between soluble and insoluble β-D-glucans shows the DP3:DP4 ratio being higher for insoluble than for soluble β-D-glucans [14,20]. For example, water-soluble β-D-glucans from barley endosperm consist of about 72% of (1,4)-β-glucosyl residues and 28% of (1,3)-β-glucosyl residues [2]. However, comparing the results is difficult because the concept of insoluble β-D-glucans differs from study to study. This ratio defines in any way the "fingerprint" of the structure of cereal β-D-glucans [24,29].
A clear and understandable result of the structural features in β-D-glucans from Poaceae is that polysaccharides have (1,3)-β-bonds embedded at irregular intervals along the whole β-D-glucan chain. These bonds cause irregularly distributed molecular kinks in the polysaccharide, which not only prevent the extensive intermolecular arrangement of chains into well-structured microfibrils, but also lead to the formation of polysaccharides that are able to form a gel-like matrix in cell walls and are capable of solubility in water, despite its relatively high molecular weight [1,16]. Barley β-D-glucans assume an extended conformation with an axial ratio (length-width) of about 100 in aqueous media [23]. The gel-like structure allows the polysaccharide to provide some degree of structural support to the cell wall, but remains flexible, resilient, and porous enough to allow the transfer of water, nutrients, and other small molecules through the wall during plant growth and development [1].
β-D-glucans containing blocks of adjacent (1,4)-β-bonds may tend to aggregate between chains (and thus lower solubility) through strong hydrogen bonds along the cellulose segments. On the other hand, (1,3)-β-bonds divide the regularity of the (1,4)-β-binding sequence, making the polysaccharide more soluble and flexible. The random distribution of these oligosaccharides ensures that the molecular “kinks” caused by (1,3)-β-glucosyl bonds are also randomly arranged in the chain [2,18]. The ability of a cell to change the ratio of β-cellotriosyl and β-cellotetraosyl residues provides a mechanism by which the solubility of a polysaccharide can be fine-tuned and adapted to biological requirements [30]. On the other hand, it is stated that a helix consisting of at least three cellotriosyl residues would represent a stable crystal structure in β-D-glucan molecules; it is therefore possible that a higher content of cellotriosyl fragments could cause some conformational regularity in the chain of β-D-glucans, and thus a higher degree of organization of these polymers (i.e., low solubility) [14].
The heterogeneity of the fine structure of β-D-glucans - the ratio of β-(1,4) to β-(1,3) bonds and special distribution of bonds along the chain - obtained by chemical analysis has important implications for the physicochemical properties such as rheological behavior. The most important rheological properties of β-D-glucans include solubility in aqueous solutions and the ability to form a viscous environment [31]. Thus, moderate cellotriosyl: cellotetraosyl ratios (e.g., 1.5 to 2.5: 1) would meet functional requirements on a wall like a porous matrix, while much higher or lower ratios would characterize conformationally more regular, less soluble β-D-glucans, which would have an increased capacity for aggregation with other molecules of β-D-glucans or with cellulose and other cell wall polysaccharides, such as heteroxylans and others [2]. The solubility of β-D-glucans, an important parameter of their functional activities, is associated also with a higher content of OH- groups in the structure and so high affinity to water molecules and ability to dissolve in the medium [31].
2. Content of β-D-Glucans in Grains of Poales
Several studies have been focused on the content of β-D-glucans in cereals such as barley and oat grains as a good natural source of this polysaccharide. Generally, barley varieties contain higher amounts of β-D-glucans compared to oat varieties, however quality and properties of both β-D-glucans are different. However, wheat is not considered to be a good source of β-D-glucans because it has a much lower content, usually <1% on a dry basis.
Barley is an important cereal grain consumed throughout the world that can be used to develop functional food products rich in β-D-glucans [32,33]. The content of β-D-glucans in barley is on average 3–4% to 8% [33,34], although barley cultivars with the content 2–11% are observed. The polysaccharide is mainly distributed in the grain endosperm (75%) and aleurone layer (25%).
Current research of barley β-D-glucans attempts to identify suitable barley genotypes for use in a number of breeding applications such as human nutrition, livestock feed, malting, and brewing [35]. Varietal variability in the β-D-glucans content and their degradation during germination of barley plays a significant role in their application for malting and brewing. Residual malt β-D-glucan from incomplete degradation of endosperm cell walls during the malting process is associated with increased worth viscosity that can slow filtration and reduce brew house efficiency [36].
In addition to the genetic resource pools of cultivated barley, wild barley (Hordeum spontaneum L.) offers considerable potential as a genetic resource for barley β-D-glucans improvement. A comparative study of β-D-glucans content between cultivated and wild barley confirmed the higher range and variability of this parameter in wild species. The β-D-glucans contents of studied wild barley accessions ranged from 3.26% to 7.67% while cultivated barley varieties ranged from 2.68% to 4.74% [35].
Another recent large-scale analysis of 117 accessions of wild barley (Hordeum vulgare subsp. spontaneum L.) which were selected from ICARDA’s gene bank to represent 21 countries scattered along the natural geographic distribution of the species were carried out by Elouadi et al. [37]. The contents of β-D-glucans ranged from 1.44% to 11.30% in the Hordeum spontaneum accessions compared to 36 cultivated barley lines with contents from 1.62% to 7.81%. On the other hands a similar range (3.6–7.4%) of β-D-glucans contents have been already detected by Austrian researchers in 86 hull-less form of cultivated barely [38].
Generally, naked barley has higher starch and β-D-glucans levels than hulled barley. Additionally, mutations at the Lys3 and Lys5 loci can affect beta-glucan content, as well as other health-promoting compounds [39]. While varieties with high β-D-glucans content are preferred in human nutrition, varieties with low concentration are preferred for malting and for broiler fattening. The reliable production of low β-D-glucans malt based on a suitable barley genotype could replace viscosity control by changing the brewing processes or adding exogenous enzymes. Therefore, novel barley β-D-glucans endohydrolase (β-glucanase) alleles with increased thermostability e.g., from Hordeum spontaneum would be perspective to identify [36].
Oat is another major source of β-D-glucans among cereals. The oat usually contains 3 to 5% of this viscous and soluble fiber component [40,41]. Unlike barley grain, a main part of β-D-glucans is located in the thick cell walls in the region of the subaleural outer endosperm [42]. Therefore, core fractions consisting of subaleural layers are particularly high in this polysaccharide. The content of β-D-glucans in oat grain ranges from 2.3% to 8.5% [4,43] depending on the cultivar and other factors [4,44]. The content of β-D-glucans in diploid oat ranges from 2.85% to 6.77%, in tetraploid 3.58% to 5.12%, and in hexaploid oat species 2.88–5.90% [45].
A significantly lower variability in β-D-glucans contents was confirmed among four oat species A. sativa (3.60%), A. byzantina (3.40%), A. abyssinica (2.46%), and A. strigosa (2.97%) by the recent study of VIR oat collection by Popov et al. [46)]. These results indicated that the hexaploid cultivated oat species A. sativa and A. byzantina showed generally higher content of β-D-glucans than tetraploid and diploid wild accessions. At the same time, Loskutov and Polonkiy [47] reported a slightly higher β-D-glucans content in naked forms of A. sativa compared to hulled oat varieties. Thus, it seems that compared to barley species, the variability of β-D-glucans content in oats is not as high and probably not sufficiently mapped.
Also, the requirements for the specific beta-glucan content of oat grain are not as clearly defined as in the case of malting barley processing or poultry fattening. A recent study [48] even summarized that 1,3-β-D-glucan can be added to broiler feed to improve the development and integrity of the gut and enhance the immune status of birds without affecting their growth rate. However, barley β-glucans do not seem to have this potential and their negative effect on poultry performance is further being studied [49].
3. Biosynthesis of β-D-Glucans in Poales
The starting point for molecular-genetic approaches to the study of cell walls of Poales was the sequencing of the rice genome and the subsequent identification of the superfamily of cellulose synthase genes (Cellullose Synthase A, CesA), while these genes are responsible for the synthesis of the hexose polysaccharide framework and cellulose itself. Cellulose Synthase Like genes (Csl) were gradually discovered [50]. This family is further divided into eight subclasses: CslA, CslB, CslC, CslD, CslE, CslF, CslG, and CslH, each containing multiple genes [51]. Later, the Csl superfamily was supplemented by the CslJ family [52]. In general, CesA genes encode cellulose synthesizing enzymes and Csl genes are responsible for the biosynthesis of hemicellulosic polysaccharides [50]. The CslC gene group encodes an enzyme that controls the synthesis of the xyloglucan backbone [53], while the CslF and CslH subfamily genes mediate the synthesis of β-D-glucans in Poaceae [54,55], with the CslF subfamily being key [54]. The ClsJ subfamily is also involved in the biosynthesis of β-D-glucans [1,52], but its phylogenetic significance is still unclear [56]. Not all Csl subfamilies are represented in higher plants. The CslB and CslG subfamilies are only found in dicots and gymnosperms, and CslF and CslH are only found in monocots [52], with the CslF branch diverging from the CslD subfamily, which evolved by diversification of CslD and CesA [57].
The biosynthesis of β-D-glucans, a unique polysaccharide of grass cell walls, is encoded by three groups of Csl genes [58], namely CslF, CslH and CslJ [57], while research has shown that selection pressure on CslF subfamily genes is related to the ratio DP3 : DP4 in the molecule of polysaccharide [59]. The involvement of the CslH subfamily was demonstrated by genetic transformation of Arabidopsis thaliana, in which CslH genes from rice were inserted and the plant produced β-D-glucans. Similar research was also carried out with the CslF subfamily [60]. Likewise, research suggests that the CslJ subfamily encodes the synthesis of β-D-glucans in barley, and this subfamily has been observed in barley, wheat, sorghum, and maize, but not in rice [52]. All genes from the CslF, CslH, and CslJ subfamilies mediate β-D-glucan synthesis in heterologous expression systems [54,55,61], although it is unclear whether each the gene in these groups controls the synthesis of β-D-glucans in vivo [30]. Phylogenetically, the CslJ and CslH subfamilies existed before the monocot-dicot split and are now present only in monocots [57].
In barley tissues, the CslF6 gene is the most transcribed in terms of β-D-glucans content [62,63,64], while it fulfils the same function in wheat, oats, and rice [62,65]. Knocking out the CslF6 gene from its function in rice plants caused not only a significant decrease in the content of β-D-glucans in the coleoptile, but also an increased activation of mechanisms related to plant protection against bacterial infections [65].
The expression levels of CslF subfamily genes intensively studied in different barley tissues and at different developmental stages of the grain showed significant differences in expression between samples. The HvCslF6 gene is expressed in almost all tissues and during the entire grain development [66], but e.g. the HvCslF3 gene is predominantly expressed in the coleoptile and root hairs [66,67], HvCslF9 in the coleoptile, roots, and developing seed 8-10 days after pollination, or HvCslF7 in the stem and stalk. Based on these results, it can be concluded that the expression of CslF genes is regulated by tissue-specific factors. The strict regulation of CslF gene expression is particularly visible during seed development, where at least four genes are expressed at very specific stages. These conclusions suggest a multistep process of β-D-glucan biosynthesis, with genes early in the biosynthetic pathway expressed first, followed by other members [66].
Most of the research on the genes coding the synthesis of β-D-glucans used barley and rice as plant material. For oat, 5 genes from the CslF family are available in the GenBank database: AsCslF3, AsCslF4, AsCslF6, AsCslF8, and AsCslF9, with AsCslF6, AsCslF8, and AsCslF9 showing higher similarity to barley, while AsCslF3 and AsCslF4 showing less conservatism. Expression analysis of genes involved in the biosynthesis of β-D-glucans in grain, leaves, and panicle of oat revealed a high level of expression of several genes in panicles, while neither AsCslF4 nor AsCslF9 gene was expressed in leaves. All genes of the CslF and CslH subfamilies were expressed during grain development. Further results revealed that all genes showed increased expression at later stages of grain development, when β-D-glucans are predicted to accumulate more prominently as grain storage polysaccharides [68]. Comparisons with published results from analysis of barley showed that barley, in contrast to oats, shows increased expression of CslF6 the 1st to 4th [66] or 6th day after pollination [68] with a significant decrease on the 8th day after pollination [66], while the expression of CslF4 and CslF8 remains low [67].
The allohexaploid (2 n = 6 x = 42; AA,CC, and DD are subgenomes) character of oat with a large complex genome is the cause of more complex and complicated molecular-genetic experiments related to the biosynthesis of β-D-glucans [7]. In the work, Coon [68] focused on the variability of expression of CslF6 gene homologues in species of the genus Avena. The genomic sequence of CslF6 was approximately 5.2 kb, and due to the large differences between individual homologues in the intron regions, the length of the genes may vary. The A-genome ortholog was 5,268 bp, the C-genome was 5,162 bp, and the D-genome sequence was 5,162 bp in length. The CslF6 gene contained two introns, the first with a length of 1600 bp and the second with a size of 748 bp. The CslF6 splice site was determined by comparison of A. sativa genomic sequences with CslF6 coding sequences from H. vulgare. The coding sequences were highly conserved among the homologs for the analyzed samples of roots, young plants, and mature embryos at the stage 1-3 days, 4-6, and 7-9 days after pollination. CslF6 expression was significantly lower in mature embryos than at any other developmental stage of the embryo. An exception was the species A. strigosa, which had intermediate expression of CslF6 in mature embryos [68].
Regarding the localization of β-D-glucans synthesis, the current dogma for cell wall polysaccharide biosynthesis is that cellulose (and callose) is synthesized at the plasma membrane, whereas matrix phase polysaccharides are assembled in the Golgi apparatus [8]. Novel model indicates that β-D-glucan does not conform to this paradigm and in various Poaceae species the CslF6 specific antibody labelling is present in the endoplasmic reticulum, Golgi, secretory vesicles, and the plasmatic membrane and the CslH1 to the same locations apart from the membrane [69]. Updated model of the β-D-glucans synthesis shows that CslF6 is the major synthase and is proposed to act similarly to CesAs, synthesizing glucan chains de novo (scenario 1). A less likely but possible alternative is that CslF6 produces cello-dextrins that are joined together at the plasma membrane by yet an unidentified protein via a single β-(1,3)-glycosidic linkage, creating the β-D-glucan chains that are then recognized by the β-D-glucan-specific antibody (scenario 2) [69].
4. Environmental Factors Influencing the β-D-Glucans Content
Differences in β-D-glucans content among species such as oats, barley, wheat, and sorghum are influenced by genetic and environmental factors [70]. Also, a mathematical model was developed that is able to pertinently assess the importance of several factors such as genetics, agronomic, and environmental factors to content of β-D-glucans during cultivation of cereals [44]. Data collected from five locations for two years (2008, 2009) analyzing the content of β-D-glucans in 11 oat genotypes identified locality, genotype, and environmental interaction to be factors influencing the content of this polysaccharide [45]. According to some authors, the main environmental factor that predisposed the content of β-D-glucans is water during maturation of the seed [71]. Heat stress and lack of water before harvests increases the amount of β-D-glucans in the grain [32,72]. However, the irrigation during oat growth is responsible for the degradation of β-D-glucans content in the grain [73]. The access to water during maturation of seeds affects the content of β-D-glucans as well as the dry conditions during harvesting. β-D-glucans was found to be affected by a reduced availability of water [74] or positively correlated to the amount of precipitation in July and August and to warmer temperatures [75].
Nitrogen fertilization has proved as a factor influencing the level of biochemical compounds in the grain and leads to an increase in the content of β-D-glucans [76]. Higher levels of nitrogen in the soil and the use of nitrogen fertilizers greatly increase the total content of β-D-glucans in oat and barley grains [32]. On the other hand, [76,77] also confirm the influence of environment on β-D-glucans variability, but in their results, the effect of genotype dominates over environmental factors. It also was confirmed by the work of Dvořáček et al. [78] comparing 11 selected yield and nutritional parameters in 5 oat varieties. The results showed the significantly highest influence of genotype on the variability of β-glucans (over 40%), even in comparison with all other parameters evaluated (e.g. content of crude protein, fat, TGW, yield). Furthermore, only year was significant for β-D-glucans variability with an effect of about 21%. On the other hand, the influence of locality and conventional or organic farming management was negligible and not significant (Figure 1).
It can be further assumed that the major influence of environmental conditions on β-D-glucans level will be related to the influence of the processes of synthesis transport and deposition of β-D-glucans during ontogenesis. Tiwari and Cummins [32] reported the highest levels of β-D-glucans after anthesis. During the first 15 days after flowering was accumulated 70–90% of total content of β-D-glucans at various nitrogen levels [13].
Comparison of the content of β-D-glucans in the panicle of hulled and naked oat samples during plant ontogenesis also described Hozlár et al. [12]. The accumulation of β-D-glucans in panicles during oat ontogenesis showed an increasing trend, while in all oat varieties the content increased from 0.69% and 0.34% to 2.23% and 2.22% in both hulled and naked oat, respectively. β-D-Glucans content in developing barley kernel was monitored by [79]. Authors confirmed individual time trend of β-D-glucans accumulation during grain maturation in tested seven barley cultivars.
It can thus be deduced that the effect of unfavorable weather conditions on the content of β-D-glucans in grain may be variety-specific, depending on the timing of these effects and the period of maximum accumulation of β-D-glucans.
5. Function of β-D-Glucans in the Plant Organism
Interest in cereal β-D-glucans has increased after their acceptance as bioactive and functional ingredients in a healthy diet [80,81] in humans and animals. Nevertheless, β-D-glucans play an important role in the structure and functionality of cereal cell walls [24]. The architectonical function in the cell wall and the storage function in the plant seeds as a source of energy for developing sprouts has been suggested [62], also the protective role of this cell wall polysaccharide against biotic or abiotic stresses is discussed in the literature [82].
Initially, it was thought that β-D-glucans serve to store energy in elongated plant cells and seeds, because content of this glucose cell wall polysaccharide is increased in young tissues [2,16,18]. This functionality of β-D-glucans is based on the fact, that the breakdown of β-D-glucans into glucose is relatively simple and involves only two enzymatic steps that allow rapid mobilization of glucose reserves compared to a longer mobilization process of starch reserves [16]. Also, the localization of this polysaccharide nearby starchy endosperm is an advantage for this molecule to serve as a source of energy for developing seedlings. It has also been shown that β-D-glucans can be metabolized as an energy source in vegetative tissues during periods of glucose deficiency. This seems to be confirmed by the fact that barley sprouts, when moved from light to dark, show increased expression of β-glucan endohydrolases and glucolases [24,83], suggesting that the plant mobilizes glucose reserves stored in β-D-glucans to compensate the decrease in photosynthetic activity in the dark [84]. In this case, plant tissues, which are not traditionally associated with photosynthetic activity, use β-D-glucans as a storage vector for glucose other than starch (or instead of the starch polysaccharide). An example is Brachypodium distachon, which grains contain up to 45% of β-D-glucans and only 6% of starch [85] compared to cereals and most wild grasses, which have 30–70% of starch as the main storage carbohydrate in the grain and generally less as 6% of β-D-glucans [86]. However, growing the plant to produce seeds is very difficult.
The architectonical role of β-D-glucans appears from the localization of β-D-glucans microfibrils in the cell wall and from the physicochemical properties of this molecule. In type I cell walls found in dicots and some monocots, the cellulose is encased in a gel-like layer of pectin to provide elasticity and stability to the cell wall. Type II cell walls are found in Poales species and contain much less pectin, its function being taken over by β-D-glucans together with arabinoxylans [87], which are highly accumulated during cell elongation, when they can make up to 20% of dry weight of the cell wall [88].The gel-like structure allows the polysaccharide to provide structural support for the cell wall, but at the same time remain flexible, elastic, and porous enough for the transport of water, nutrients and other small molecules across the cell wall during plant tissue development [1]. In young developing plant tissues, it is important that cell walls are porous so that water molecules, nutrients and low molecular weight hormones can be freely transported between cells and individual tissues. In the specialized conductive tissues necessary for the long-distance transport of water molecules and nutrients through the plant, the walls, on the contrary, must be impermeable.
In most plant tissues, cell walls are also important for intercellular adhesion. These different functional requirements for cell walls are maintained by the formation of reinforced gel-like structures consisting of cellulose microfibrils that have high tensile strength and are embedded in a gel matrix phase that consists predominantly of non-cellulosic polysaccharides [24]. This matrix provides the cell with flexibility and a certain mechanical support for maintaining the functional properties of the cell wall.
In Gramineae, the metabolism of β-D-glucans is responsible for plant responses to environmental signals within a moderate, physiological range [89]. For example, in the work of Takeda et al. [63], barley variety producing standard content of β-D-glucans (3.8%) was characterized by better wintering compared to a mutant with knocked enzyme responsible to produce this polysaccharide. Recent studies also point to the potential of β-D-glucans to be involved in defense mechanisms in the Poaceae family against selected forms of environmental stresses [82,90,91,92]. The gel-like layer of β-D-glucans in the cell wall can act as a defensive barrier that protects the cell from fungal invasion, but also provides a potential signaling system that indicates when such an attack is taking place. As fungi attack plant cells and release β-glucanases to digest the protective gel layer surrounding the cell, they slowly dissolve the protective layer of β-D-glucans and expose the cytoplasmic membrane itself. The antioxidant activity of β-D-glucans was also described [93], whereby antioxidant compounds are an accepted factor of the plant’s defense system to cope with biotic aggressors such as fungal pathogens [94,95]. The increase or decrease in the biosynthesis and associated content of β-D-glucans is conditioned by the expression of plant genes involved in both β-D-glucan synthesis and its degradation. After the exposure of plants to external factors such as pests or pathogens, the β-D-glucan metabolism as well as the entire β-D-glucans turnover are affected and become much more complicated. A study in rice mutants knocking out rice genes involved in the biosynthesis of β-D-glucans showed a phenotype that had a spontaneous response to the lesion, presumably suggesting that fibers of β-D-glucans work as a repressor of the signaling cascade targeting programmed cell death [65]. Also, reduced deoxynivalenol toxin (DON) content in barley genotypes infected with Fusarium graminearum was observed in grains with higher content of β-D-glucans [91] and in oat grains artificially infected with F. graminearum and F. culmorum, where grains containing higher amounts of β-D-glucans showed lover content of DON and pathogenic DNA [82].
The function of β-D-glucans in Poaceae as well as the evolutionary preservation of this polysaccharide in monocotyledons are still discussed, as this phenomenon of β-D-glucans has not been precisely explained [1,2,16]. The conformational regularity or irregularity of β-D-glucans defines the properties and thus the physicochemical activities in the matrix, in the cell wall. It is this conformational irregularity of β-D-glucans that appears to be a feature limited to Poaceae polysaccharides and raises the question whether this irregularity is a key feature that has led to widespread acceptance and preservation of β-D-glucans in the walls of Poaceae during evolution [57,61,64]. During the adaptation from an aquatic to a soil environment, plants had to develop a mechanism for their cells to withstand substantial expansion pressures, to be able to prevent the rupture of the cytoplasmic membrane, and at the same time contain a structure compatible with the growth of cells and cell tissues, while the construction of the cell wall itself of photosynthetically active embryophytes is crucial [96]. The plant cell wall is designed so that its architecture is strong, but at the same time flexible and able to withstand pressure, tension, and various adverse environmental conditions [97]. There is an assumption that the unique structure of β-D-glucans and the extensive presence of this polysaccharide in species of the genus Poales brings enormous evolutionary and adaptive advantages to the plant [24] and is the reason that this group of plants shows extraordinary dynamics of various pressures during evolution [56,64] and inhabits often extreme and inhospitable habitats.
6. Molecular Markers as Breeding Tools for β-D-Glucans Manipulations
In the case of β-D-glucans, the effectiveness of the standard breeding process is related to the identification of suitable genetic resources, knowledge of the responsible genes, the availability of genetic markers, and a good estimate of the heritability of the trait. Efficient phenotyping tools for screening estimation of the beta-glucan parameter in early generations are also an advantage.
There are many studies demonstrating the moderate to high heritability of β-D-glucans found in oats and barley. The average heritability level (h2b) of β-D-glucans was estimated to be 0.55 in individual oat plants [98]. An even higher range of (h2b = 0.75 – 0.84) in the case of β-D-glucans was found in barley [99]. Holthaus et al. [98] also mentioned that groat β-D-glucans content in oat polygenically controlled primarily by genes with additive effects. Genetic variation for β-glucan seemed adequate for effective selection and genotype x environment interaction was minor. Swanston [100] reported that β-D-glucans content is a quantitative trait with several associated QTLs; one QTL is located on chromosome 7H and is within 5 cM of the Nud gene in the case of barley.
Using current molecular technologies, putative genes responsible for β-D-glucans variation in the grain are continuously studied based on the still limited information on β-D-glucans synthesis. However, gene families involved in the synthesis of these polysaccharides have been identified and include the Cellulose-synthase-like (Csl) genes [101], which were described in detail in the previous chapter on β-D-glucans biosynthesis. Members of the CslF and CslH gene families and Glucan synthase-like (or Callose synthases), are prime candidates for β-D-glucans synthesis. Nevertheless, Csl gene families does not completely explain the variation of β-D-glucans content in cereal grains. There is accumulating evidence that several other genes, including those that hydrolyze β-D-glucans, contribute significantly to their content in mature grains [102].
New studies of genome wide association study [GWAS] associating genetic mutations with measures of β-D-glucans content in cultivated barley have become a powerful tool in identifying candidate genes for selective breeding. Seven putative candidate genes encoding some enzymes in glucose metabolism were found to be associated with β-D-glucans content. One of the putative genes, HORVU6Hr1G088380, could be an important gene controlling barley β-D-glucans content [103]. Marcotuli et al. [101] identified seven genomic regions associated with β-D-glucans in a tetraploid wheat collection, located on chromosomes 1A, 2A (two), 2B, 5B, and 7A (two). Analysis of marker trait associations (MTAs) in syntenic regions of several grass species revealed putative candidate genes that might influence β-D-glucans levels in the endosperm, possibly via their participation in carbon partitioning. Walling et al. [102] applied GWAS to the Wild Barley Diversity Collection (H. spontaneum) and identified a total of 13 quantitative traits loci (QTL) spread across the seven barley chromosomes that explained most of the variation in β-D-glucans content. Transcriptional dynamics of two barley genotypes differing in grain β-D-glucans content during grain development was investigated by authors [104]. 22 differentially expressed genes (DEGs) affecting β-D-glucans accumulation during late developmental stages were selected. Most of these DEGs (encoding alpha-amylase inhibitor, glucan endo-1,3-beta-glucosidase, and sugar transporter) showed different expression patterns in the two genotypes, which might explain the genotypic difference in changes in β-D-glucans content 21–28 days post-anthesis (DPA) (Figure 2).
The hexaploid structure of the oat genome further complicates the detection of β-D-glucans associated markers. However, Newell et al. [105] associated three DArT markers with β-D-glucans concentration in oats. These markers had sequence homology to rice; one of these DArT sequences, opt.0133, was located on rice chromosome seven and was, by our definition, adjacent to the CslF gene family. A recent large GWAS analysis carried out on an oat panel with 413 genotypes was evaluated for β-D-glucans content under subtropical conditions [106]. Seven quantitative trait loci (QTL) associated with β-D-glucans content were identified and located on Mrg02, Mrg06, Mrg11, Mrg12, Mrg19, and Mrg20. The QTL located on Mrg02, Mrg06, and Mrg11 seem to be genomic regions syntenic with barley.
Genome editing methods for influencing β-D-glucans content in barley were used by Garcia-Gimenez et al. [67] They used CRISPR/Cas9 to generate mutations in members of the Csl gene superfamily that encode known (HvCslF6andHvCslH1) and putative (HvCslF3 and HvCslF9) β-D-glucans synthases. Grains from CslF6-2 (homozygous) mutants almost completely lack β-D-glucans (0.11%), whereas grains from CslF6-2/+ (heterozygous lines) have intermediate levels of β-D-glucans (1.45%) compared with the wild type control (5.00%). Their data further indicated that only HvCslF6 from multiple members of the CslF/H family showed impact on the abundance of β-D-glucans in mature grain. Genome editing procedures in case of content decreasing of β-D-glucans overcome traditional breeding practices and could be a good breeding strategy for the new malting varieties of barley.
Molecular methods in combination with effective phenotyping procedures for β-D-glucans content (eg. modified McCleary enzymatic determination or NIRS) are key for the development of new varieties with defined content of the monitored polysaccharide in the breeding process [107,108]. NIRS models developed by Paudel et al. [108] declared high index of determination (h2 ≥ 0.93) and low standard error of cross-validation (SECV ≤ 0.23) for β-D-glucans quantification in ground and whole oat groats as well. The above findings thus hold great promise for obtaining new varieties of oats and barley with declared β-D-glucan content according to the requirements of processors.
7. Impact of Processing Technologies on β-D-Glucans in Cereals
The interest in possible content and structural changes of beta-glucans during technological processing is related to their confirmed positive health effect. In case of oat beta-glucans, the scientific opinion even confirmed their effect on lowering blood cholesterol and reducing the risk of (ischemic) heart disease in accordance with Article 14 of Regulation (EC) No. 1924/2006 (EFSA Journal 2010;8(12):1885).
β-D-Glucans are an important component of dietary fiber, whose bioavailability, nutritional quality, and physicochemical properties are fundamentally influenced by the respective technological processing.
The addition of dietary fiber affects the technology and consequently the sensory quality of the food. The role of fiber is to contribute to the structure, which is essential for the physicochemical properties of fiber-enriched foods [109]. High viscosity is a hallmark of β-D-glucans and is a function of the concentration of β-D-glucans in solution and their Mw [110]. In this context, there are a number of cereal processing processes that can significantly alter the content, solubility and physic-biological properties of β-D-glucans in the resulting products.
7.1. Effect of the Milling Process and Grain Peeling
The milling process has a major influence on the dietary fiber content. The milling process is influenced by different milling equipment, where targeted sieving and separation can influence the quality of the product, and fractions with high fiber, protein, or other substances can be obtained [26]. The fiber and mineral content will increase with a higher degree of milling. In naked barley, the β-D-glucans content after the different passages varies between 8, 11, and 5% [111,112].
The intensity of grain peeling before milling also has a significant effect. When the wheat grain is peeled to 3%, which represents the pericarp, fiber content of 62% was determined in this fraction. With a higher degree of peeling, from 6% or more, the fiber content of the final product decreases significantly. Higher levels of peeling increase the proportion of endosperm and aleurone layer in the final product. Peeling reduces the fiber content of the bran from an initial 62% in the unpeeled grain to 45% after peeling with 12% of the outer layers [113].
Bran separation significantly increased the beta-glucan content of oats. This increase was more than twofold compared to the whole grain content. Smaller but significant changes were also observed in this context for the separation technique used Izydorczyk et al. [114]. These authors also mapped the effect of different roll milling conditions during bran separation on total b-glucan content in oat bran. The studies showed that the β-D-glucans content in the bran fractions increased when the break rolls were operated under a dull-to-dull rather than sharp-to-sharp disposition.
In cultivars with lower β-D-glucans content and standard starch composition, most of the β-D-glucans are found in the subaleuronal layer and the surrounding endosperm. But, in cultivars with high β-D-glucans content (waxy, amylose starch composition), more than 80% were mainly in the endosperm. The central endosperm was found to contain less β-D-glucans than the middle layers [112]. A significant effect of growing conditions, location, and vintage on the β-D-glucans content in barley grain was observed, however, the distribution of this polysaccharide in the grain was not observed [111]. Specific products with certain characteristics such as high fiber, protein, or starch content could be obtained by using the most suitable milling and separation method for a certain cereal [115]. A significant effect and positive correlation between fiber content and milling technology and genotype have been found for Triticum spelta and common wheat [116]. Passages or specific types of high-fiber flours can be produced by appropriate milling and separation methods adapted to the cereal.
7.2. Effects and Changes of β-D-Glucans in the Process of Dough Preparation, Fermentation, and Baking
The addition of fiber-rich fractions into special high-fiber flours, or fiber preparations must consider the workability and formation of the dough and the sensory acceptability of the final bakery product. During the preparation of the dough and its subsequent heat treatment (baking, frying), there are greater and more varied changes in fiber content and quality than during the milling process. During dough preparation, the water binding capacity is influenced by the origin and particle size of the flour, the protein content and quality, and the type of starch. Higher fiber content results in higher water absorption, which can cause lower gas retention, lower specific volume of the final product, and lower crumb porosity [117,118,119,120]. The addition of barley flour or β-D-glucans-rich fractions to baker's strong wheat flour caused a reduction in specific bread volume [117,120].
The negative effects of the addition of dietary fiber on dough consistency can be influenced by proper dough preparation [121,122], for example by hydrating the dietary fiber before adding it to the dough, where better water utilization by other biopolymers is achieved, as well as by different baking methods [123]. The use of barley flour sourdough led to a decrease in water absorption, but the use of fermented oat bran with a high β-D-glucans content increased water absorption [124]. The addition of β-D-glucans leads to increased dough development time and reduced dough stability [117,120,124].
The maturation and fermentation of dough has a significant effect on the quantity and quality of fiber, including beta-glucans, but not all results are completely consistent (see below). During the long fermentation period, water is redistributed from non-starch polysaccharides back into the gluten structure, maintaining the specific bread volume[26,121]. Dough maturing and fermentation allows for longer enzyme action and the progression of hydrolytic processes. Fermentation at 30°C for 20 h with added lactobacilli reduced the dietary fiber and β-D-glucans content, but the molecular weight of this polysaccharide was not affected [125,126]. Heat treatment of oat bran prevented the decrease in the content and molecular weight of β-D-glucans during dough fermentation. Sourdough of oat bran led to an increase in soluble arabinoxylan content and a decrease in dietary fiber content [127]. Andersson et al. [128], Andersson et al. [122] and Rieder et al. [123] showed a decrease in the molecular weight of β-D-glucans and their content in barley and rye dough with a fermentation time of 0-60 min and in a sourdough from barley wholemeal flour and oat bran leaven under fermentation conditions of 30 °C, 20 h. Comino et al. [129] found that there were no significant changes in the content of arabinoxylans and β-D-glucans, the arabinose/xylose ratio for arabinoxylans, and the cellotriosyl/cellotetraosyl units for β-D-glucans during dough preparation and fermentation.
The content of fiber components in the product after the baking process depends on the type of preparation and leavening of the bread (leavened with yeast, air, or sourdough). During the baking of wholemeal yeast-raised rye bread, there were no significant changes in the content of arabinoxylans, β-D-glucans, and arabinogalactans, while the content of resistant starch and cellulose increased and the content of fructans decreased. The molecular weight of the water-soluble high-molecular-weight arabinoxylans did not change during bread preparation but decreased with the addition of the sourdough. The molecular weight of β-D-glucans decreased only slightly during baking [123]. However, Tiwari et al. [127] and Rakha et al. [128] described a rapid decrease in the content and molecular weight of high molecular weight β-D-glucans with an increase in the content of medium and low molecular weight β-D-glucans because of baking time and temperature. Comino et al. [129] described in wheat, barley, and rye endosperm only insignificant changes during baking in the content of arabinoxylans and β-D-glucans and at the level of their structure, but there was a slight increase in the solubility of arabinoxylans and β-D-glucans.
7.3. Effect of Extrusion
Most extrusion analyses note changes in the properties of major components and major polymers (starches, proteins, and fiber). Extrusion causes irreversible changes in the flour components, especially the flour polymers, i.e., starch, protein, and fiber, by breaking the polymer chains. The disruption of polymer chains leads to a decrease in molecular weight, the formation of starch-lipid, protein-lipid, and protein-protein complexes, and an increase in the solubility of fiber and starch [132]. Information on the direct assessment of changes in the properties of β-D-glucans after extrusion is, however, limited. In the case of dietary fiber, of which β-D-glucans are an important component, a number of changes occurred during extrusion. During extrusion, thermomechanical changes, depolymerization, changes in solubility, and transglycosidation processes have the effect of reducing the insoluble dietary fiber and total dietary fiber content and on the other hand, increasing the soluble dietary fiber content [129,132,133,134,135]. As a result of extrusion, a higher loss in insoluble dietary fiber is observed compared the increase in soluble dietary fiber. This is explained by the conversion of cellulose to lower molar mass soluble dietary fiber fractions that could be further degraded into low molecular weight components and sugar derivatives that cannot be determined as dietary fiber [133]. During extrusion, total dietary fiber content can be increased by the formation of resistant starch or indigestible Maillard reaction products. Increasing the extruder screw speed increased the soluble dietary fiber content. The soluble dietary fiber content of the product and the molecular weight can be influenced by setting the temperature and the initial addition of water [132]. The molecular weight of soluble dietary fiber plays an important role in the expansion rate [132]. The results of the increase in water-soluble fiber components after extrusion may be related to the increase in water-soluble β-D-glucans and arabinoxylans found by [129,136,137,138] in barley, wheat, and rye. In contrast, no changes were observed by these authors in the ratio of basic building units.
8. The Range of Applications of β-D-Glucans in Food Products
Cereal β-D-glucans, mainly found in barley and oats, possess technological advantages and multiple health benefits. However, changes in their physicochemical properties and health-promoting effects can occur during food processing and storage, representing a major disadvantage [139]. To maximize the health benefits of β-D-glucans for a wide range of consumers, it is essential to consider the most salient aspects of buying and consuming food products. This includes ensuring a high textural and sensory quality of the product [140]. To maximize the potential of cereal β-D-glucans, it is essential to extend their applications beyond bread and cereal beverages. Doing so will enable the full spectrum of benefits associated with this polysaccharide to be realized [139].
β-D-glucans have potential applications in medicine, pharmacy, veterinary, cosmetic, chemical industries and in food and feed production. The biological activity and effects of β-D-glucans depend significantly on their source, the isolation method used, and their specific physicochemical properties, such as structure, viscosity, and molecular weight [141]. The following technological operations in the production of food and food supplements also have a significant influence. β-D-Glucans are valuable functional ingredients because they can influence the quality, structure, rheological properties, and stability of various food systems (Table 1). They have various physical properties; they are capable of thickening, stabilizing, emulsifying, and gelling [81]. The addition of β-D-glucans to foods has been shown to improve the physical properties of food products. β-D-glucans affect the consistency of the product and can be used as fillers, stabilizers, thickeners, and fat replacements in many dairy and bakery products, improving sensory quality or extending the shelf life of the product and slowing down the aging process. In terms of human nutrition, they reduce energy and increase the nutritional value of foods [142]. The naturally occurring β-D-glucans in cereals present an attractive opportunity for manufacturers to design foods with added health value and positive labelling for consumers [110].
Due to their emulsifying and thickening properties, low molecular weight β-D-glucans [159] are commonly used successfully in liquid food matrices such as beverages, sauces, and soups [160,161]. Low molecular weight β-D-glucans have less impact on the product’s final viscosity [159]. The addition of cereal β-D-glucans to chocolate-flavored milk has been demonstrated to be successful in improving the texture, mouthfeel, and taste of the chocolate, as well as increasing the viscosity of the product. At a concentration of 3%, the cereal β-D-glucans were able to enhance the overall quality of the beverage [152]. Angelov et al. [162] found that the production of a cereal-based fermented beverage did not result in any changes in β-D-glucans content during fermentation and storage. This suggests that the β-D-glucans present in the beverage were unaffected by the fermentation and storage process.
For solid food matrices, the addition of β-D-glucans is mainly investigated for cereal-based products. The addition of β-D-glucans to bakery products is known to lead to an increase in dough viscosity and crumb strength, as well as improved elasticity and coloration [163]. Bread with high β-D-glucans content had higher loaf volume and lower glycemic index [164]. Bread with added oat beta-glucans has a lower specific volume and porosity, darker color, higher hardness and lower elasticity and consistency than the control white bread. However, these negative effects can be effectively counteracted by optimizing the water content of the products [143]. The baking properties most affected by β-D-glucans were volume and color. Oat β-D-glucans softened the crust of gluten-free bread but had the opposite effect for wheat bread [144]. High-glucan barley flour has been used in the production of pasta at a rate of 30% [145] and to produce couscous [147].
The fortification of yellow alkaline noodles with barley β-D-glucans has been shown to reduce their sensory quality [146]. However, when a new sponge cake formulation was tested with 4,4% addition of β-D-glucans, sponge cakes had excellent consumer acceptability [148]. An even higher addition of 5.2% β-D-glucans in a cereal bar led to an improvement of taste and overall appearance ratings [149]. Additionally, barley β-D-glucans have been used as a fat substitute in yoghurt and have been able to improve the texture and sensory quality with just a 0.5% addition [150].
The use of β-D-glucans in cheese production is interesting. β-D-glucans can structure the mixtures during the fermentation of the lactic curd, actively bind the whey, and result in higher yields of cheese produced [165]. Cereal β-D-glucans have the ability to replace the texture and taste of milk fat in products but impair the sensory properties of protein-containing products [166]. β-D-glucan has been found to reduce the acidity of cheese and increase the yield of products by preventing whey separation and contributing to texture formation. However, grain β-D-glucans can significantly impair the sensory properties of cheese, be a source of flavor and change the color of the dairy product [153,167]. Due to the water retention ability of β-D-glucans, they have a significant ability to reduce losses, increase food product yields and reduce food process times [168].
9. Conclusions
New knowledge about the biosynthesis and genetic composition of β-D-glucans in combination with new molecular techniques should lead to the development of new varieties more resistant to stress conditions. Genetically defined modification of the content of β-D-glucans in cereal grain will enable targeted recommendations of barley or oat varieties suitable for individual processing categories (malt, feed, or food, cosmetic or pharmaceutical industry). Gentle technological processing will make it possible to stabilize or even increase the concentration of this cell wall polysaccharide in the final products with a positive impact both on the health status of the consumer and on the structural and sensory properties of the final product.
Author Contributions
Conceptualization, M.H. and V.D.; methodology, M.H.; validation, M.H. and V.G.; investigation, M.H., V.D., L.J. and V.G.; writing—original draft preparation, M.H., V.D. and L.J.; writing—review and editing, M.H., V.D. and V.G.; visualization, V.G.; supervision, M.H.; project administration, M.H. and V.D.; funding acquisition, M.H. and V.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Slovak Research and Development Agency under the Contract no. APVV-18-0154 and by the Ministry of Agriculture of the Czech Republic, institutional support MZE-RO0423.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fincher, G.B. Revolutionary Times in Our Understanding of Cell Wall Biosynthesis and Remodeling in the Grasses. Plant Physiology 2009, 149, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Burton, R.A.; Gidley, M.J.; Fincher, G.B. Heterogeneity in the Chemistry, Structure and Function of Plant Cell Walls. Nat Chem Biol 2010, 6, 724–732. [Google Scholar] [CrossRef]
- Fincher, G.B.; Stone, B.A. CEREALS|Chemistry of Nonstarch Polysaccharides. In Encyclopedia of Grain Science; Wrigley, C., Ed.; Elsevier: Oxford, 2004; pp. 206–223. [Google Scholar] [CrossRef]
- Havrlentova 2006 Content of β-D-Glucan in Cereal Grains. J. Food Nutr. Res. 2006, 45, 97–103.
- Redaelli, R.; Sgrulletta, D.; Scalfati, G.; De Stefanis, E.; Cacciatori, P. Naked Oats for Improving Human Nutrition: Genetic and Agronomic Variability of Grain Bioactive Components. Crop science 2009. https://agris.fao.org/agris-search/search.do?recordID=US201301657634.
- Gajdošová, A.; Petruláková, Z.; Havrlentová, M.; Červená, V.; Hozová, B.; Šturdík, E.; Kogan, G. The Content of Water-Soluble and Water-Insoluble β-d-Glucans in Selected Oats and Barley Varieties. Carbohydrate Polymers 2007, 70, 46–52. [Google Scholar] [CrossRef]
- Sikora, P.; Tosh, S.M.; Brummer, Y.; Olsson, O. Identification of High β-Glucan Oat Lines and Localization and Chemical Characterization of Their Seed Kernel β-Glucans. Food Chemistry 2013, 137, 83–91. [Google Scholar] [CrossRef]
- Buckeridge, M.S.; Rayon, C.; Urbanowicz, B.; Tiné, M.A.S.; Carpita, N.C. Mixed Linkage (1→3),(1→4)-β- d -Glucans of Grasses. Cereal Chemistry Journal 2004, 81, 115–127. [Google Scholar] [CrossRef]
- Sykut-Domańska, E.; Rzedzicki, Z.; Zarzycki, P.; Sobota, A.; Błaszczak, W. Distribution of (1,3)(1,4)-Beta-D-Glucans in Grains of Polish Oat Cultivars and Lines (Avena Sativa L.). Pol. J. Food Nutr. Sci. 2016, 66, 51–56. [Google Scholar] [CrossRef]
- Trethewey, J.A.K.; Campbell, L.M.; Harris, P.J. (1→3),(1→4)-ß-d-Glucans in the Cell Walls of the Poales (Sensu Lato): An Immunogold Labeling Study Using a Monoclonal Antibody. American Journal of Botany 2005, 92, 1660–1674. [Google Scholar] [CrossRef]
- Vega-Sanchez, M.; Verhertbruggen, Y.; Scheller, H.V.; Ronald, P. Abundance of Mixed Linkage Glucan in Mature Tissues and Secondary Cell Walls of Grasses. Plant Signaling & Behavior 2013, 8, e23143. [Google Scholar] [CrossRef]
- Hozlár, P.; Gregusová, V.; Nemeček, P.; Šliková, S.; Havrlentová, M. Study of Dynamic Accumulation in β-D-Glucan in Oat (Avena Sativa L.) during Plant Development. Polymers 2022, 14. [Google Scholar] [CrossRef]
- Fan, M.; Zhang, Z.; Wang, F.; Li, Z.; Hu, Y. Effect of Nitrogen Forms and Levels on β-Glucan Accumulation in Grains of Oat ( Avena Sativa L.) Plants. Z. Pflanzenernähr. Bodenk. 2009, 172, 861–866. [Google Scholar] [CrossRef]
- Izydorczyk, M.S.; Macri, L.J.; MacGregor, A.W. Structure and Physicochemical Properties of Barley Non-Starch Polysaccharides—II. Alkaliextractable β-Glucans and Arabinoxylans. Carbohydrate Polymers 1998, 35, 259–269. [Google Scholar] [CrossRef]
- Chang, S.-C.; Saldivar, R.K.; Liang, P.-H.; Hsieh, Y.S.Y. Structures, Biosynthesis, and Physiological Functions of (1,3;1,4)-β-d-Glucans. Cells 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Fincher, G.B. Exploring the Evolution of (1,3;1,4)-β-d-Glucans in Plant Cell Walls: Comparative Genomics Can Help! Current Opinion in Plant Biology 2009, 12, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Staudte, R.G.; Woodward, J.R.; Fincher, G.B.; Stone, B.A. Water-Soluble (1→3), (1→4)-β-d-Glucans from Barley (Hordeum Vulgare) Endosperm. III. Distribution of Cellotriosyl and Cellotetraosyl Residues. Carbohydrate Polymers 1983, 3, 299–312. [Google Scholar] [CrossRef]
- Burton, R.A.; Fincher, G.B. (1,3;1,4)-β-D-Glucans in Cell Walls of the Poaceae, Lower Plants, and Fungi: A Tale of Two Linkages. Molecular Plant 2009, 2, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Skendi, A.; Biliaderis, C.G.; Lazaridou, A.; Izydorczyk, M.S. Structure and Rheological Properties of Water Soluble β-Glucans from Oat Cultivars of Avena Sativa and Avena Bysantina. Journal of Cereal Science 2003, 38, 15–31. [Google Scholar] [CrossRef]
- Izydorczyk, M.S.; Biliaderis, C.G.; Macri, L.J.; MacGregor, A.W. Fractionation of Oat (1→3), (1→4)-β-D-Glucans and Characterisation of the Fractions. Journal of Cereal Science 1998, 27, 321–325. [Google Scholar] [CrossRef]
- Tosh, S.; Brummer, Y.; Wolever, T.; Wood, P. Glycemic Response to Oat Bran Muffins Treated to Vary Molecular Weight of β-Glucan. Cereal chemistry. 2008, 85, 211–217. [Google Scholar] [CrossRef]
- Lazaridou, A.; Biliaderis, C.G. Molecular Aspects of Cereal β-Glucan Functionality: Physical Properties, Technological Applications and Physiological Effects. Journal of Cereal Science 2007, 46, 101–118. [Google Scholar] [CrossRef]
- Woodward, J.R.; Fincher, G.B.; Stone, B.A. Water-Soluble (1→3), (1→4)-β-D-Glucans from Barley (Hordeum Vulgare) Endosperm. II. Fine Structure. Carbohydrate Polymers 1983, 3, 207–225. [Google Scholar] [CrossRef]
- Burton, R.; Fincher, G. Current Challenges in Cell Wall Biology in the Cereals and Grasses. Frontiers in plant science 2012, 3, 130. [Google Scholar] [CrossRef] [PubMed]
- Doehlert, D.C.; Simsek, S. Variation in Β-glucan Fine Structure, Extractability, and Flour Slurry Viscosity in Oats Due to Genotype and Environment. Cereal chemistry 2012, 89, 242–246. [Google Scholar] [CrossRef]
- Izydorczyk, M.S.; Dexter, J.E. Barley β-Glucans and Arabinoxylans: Molecular Structure, Physicochemical Properties, and Uses in Food Products–a Review. Food Research International 2008, 41, 850–868. [Google Scholar] [CrossRef]
- Ramesh, H.P.; Tharanathan, R.N. Carbohydrates—The Renewable Raw Materials of High Biotechnological Value. Critical Reviews in Biotechnology 2003, 23, 149–173. [Google Scholar] [CrossRef]
- Miller, S.S. Wood, P.J., Pietrzak, L.N., Fulcher, R.G. Mixed Linkage beta-Glucan, Protein Content, and Kernel Weight in Avena Species. Cereal Chem. 1993. Available online: https://www.cerealsgrains.org/publications/cc/backissues/1993/Documents/cc1993a47.html (accessed on 10 April 2023).
- Cui, W.; Wood, P.J. Relationships between Structural Features, Molecular Weight and Rheological Properties of Cereal β-D-Glucans. In Hydrocolloids; Nishinari, K., Ed.; Elsevier Science: Amsterdam, 2000; pp. 159–168. [Google Scholar] [CrossRef]
- Bulone, V.; Schwerdt, J.G.; Fincher, G.B. Co-Evolution of Enzymes Involved in Plant Cell Wall Metabolism in the Grasses. Front. Plant Sci. 2019, 10, 1009. [Google Scholar] [CrossRef]
- Du, B.; Meenu, M.; Liu, H.; Xu, B. A Concise Review on the Molecular Structure and Function Relationship of β-Glucan. IJMS 2019, 20, 4032. [Google Scholar] [CrossRef]
- Tiwari, U.; Cummins, E. Factors Influencing β-Glucan Levels and Molecular Weight in Cereal-Based Products. Cereal Chemistry Journal 2009, 86, 290–301. [Google Scholar] [CrossRef]
- Markovic, S.; Djukic, N.; Knezevic, D.; Lekovic, S. Divergence of Barley and Oat Varieties According to Their Content of β-Glucan. J Serb Chem Soc 2017, 82, 379–388. [Google Scholar] [CrossRef]
- MacGregor, A.W. BARLEY. In Encyclopedia of Food Sciences and Nutrition (Second Edition); Caballero, B., Ed.; Academic Press: Oxford, UK, 2003; pp. 379–382. [Google Scholar] [CrossRef]
- Nishantha, M.D.L.C.; Zhao, X.; Jeewani, D.C.; Bian, J.; Nie, X.; Weining, S. Direct Comparison of β-Glucan Content in Wild and Cultivated Barley. International Journal of Food Properties 2018, 21, 2218–2228. [Google Scholar] [CrossRef]
- Lauer, J.C.; Cu, S.; Burton, R.A.; Eglinton, J.K. Variation in Barley (1 → 3, 1 → 4)-β-Glucan Endohydrolases Reveals Novel Allozymes with Increased Thermostability. Theor Appl Genet 2017, 130, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Elouadi, F.; Amri, A.; El-baouchi, A.; Kehel, Z.; Salih, G.; Jilal, A.; Kilian, B.; Ibriz, M. Evaluation of a Set of Hordeum Vulgare Subsp. Spontaneum Accessions for β-Glucans and Microelement Contents. Agriculture 2021, 11. [Google Scholar] [CrossRef]
- Eticha, F.; Grausgruber, H.; Berghoffer, E. Multivariate Analysis of Agronomic and Quality Traits of Hull-Less Spring Barley (Hordeum Vulgare L.). Journal of plant breeding and crop science 2010, 2, 81–95. [Google Scholar] [CrossRef]
- Meints, B.; Vallejos, C.; Hayes, P. Multi-Use Naked Barley: A New Frontier. Journal of Cereal Science 2021, 102, 103370. [Google Scholar] [CrossRef]
- Peterson, D.M. Oat Antioxidants. Journal of Cereal Science 2001, 33, 115–129. [Google Scholar] [CrossRef]
- Mälkki, Y. Trends in Dietary Fibre Research and Development. Acta Alimentaria 2004, 33, 39–62. [Google Scholar] [CrossRef]
- Fulcher, R.G.; Miller, S.S. Structure of Oat Bran and Distribution of Dietary Fiber Components. Oat bran 1993, 1–24. [Google Scholar]
- Welch, R.W.; Brown, J.C.W.; Leggett, J.M. Interspecific and Intraspecific Variation in Grain and Groat Characteristics of Wild Oat (Avena) Species: Very High Groat (1→3),(1→4)-β- -Glucan in an Avena Atlantica Genotype. Journal of Cereal Science 2000, 31, 273–279. [Google Scholar] [CrossRef]
- Tiwari, U.; Cummins, E. Simulation of the Factors Affecting β-Glucan Levels during the Cultivation of Oats. Journal of Cereal Science 2009, 50, 175–183. [Google Scholar] [CrossRef]
- Redaelli, R.; Del Frate, V.; Bellato, S.; Terracciano, G.; Ciccoritti, R.; Germeier, C.U.; De Stefanis, E.; Sgrulletta, D. Genetic and Environmental Variability in Total and Soluble β-Glucan in European Oat Genotypes. Journal of Cereal Science 2013, 57, 193–199. [Google Scholar] [CrossRef]
- Popov, V.S.; Khoreva, V.I.; Konarev, A.V.; Shelenga, T.V.; Blinova, E.V.; Malyshev, L.L.; Loskutov, I.G. Evaluating Germplasm of Cultivated Oat Species from the VIR Collection under the Russian Northwest Conditions. Plants 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Loskutov, I.G.; Polonskiy, V.I. Content of β-Glucans in Oat Grain as a Perspective Direction of Breeding for Health Products and Fodder.
- Amer, S.A.; Attia, G.A.; Aljahmany, A.A.; Mohamed, A.K.; Ali, A.A.; Gouda, A.; Alagmy, G.N.; Megahed, H.M.; Saber, T.; Farahat, M. Effect of 1,3-Beta Glucans Dietary Addition on the Growth, Intestinal Histology, Blood Biochemical Parameters, Immune Response, and Immune Expression of CD3 and CD20 in Broiler Chickens. Animals 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.P.; Pescatore, A.J. Barley β-Glucan in Poultry Diets. Annals of translational medicine 2014, 2, 20. [Google Scholar] [CrossRef] [PubMed]
- Richmond, T.A.; Somerville, C.R. The Cellulose Synthase Superfamily. 2000, 55(1), 44–52. [CrossRef]
- Hazen, S.P.; Scott-Craig, J.S.; Walton, J.D. Cellulose Synthase-Like Genes of Rice. Plant Physiol. 2002, 128, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Farrokhi, N.; Burton, R.A.; Brownfield, L.; Hrmova, M.; Wilson, S.M.; Bacic, A.; Fincher, G.B. Plant Cell Wall Biosynthesis: Genetic, Biochemical and Functional Genomics Approaches to the Identification of Key Genes. Plant Biotechnology Journal 2006, 4, 145–167. [Google Scholar] [CrossRef] [PubMed]
- Cocuron, J.-C.; Lerouxel, O.; Drakakaki, G.; Alonso, A.P.; Liepman, A.H.; Keegstra, K.; Raikhel, N.; Wilkerson, C.G. A Gene from the Cellulose Synthase-like C Family Encodes a -1,4 Glucan Synthase. PLANT BIOLOGY 6.
- Burton, R.A.; Wilson, S.M.; Hrmova, M.; Harvey, A.J.; Shirley, N.J.; Medhurst, A.; Stone, B.A.; Newbigin, E.J.; Bacic, A.; Fincher, G.B. Cellulose Synthase-like CslF Genes Mediate the Synthesis of Cell Wall (1, 3; 1, 4)-ß-D-Glucans. Science 2006, 311, 1940–1942. [Google Scholar] [CrossRef] [PubMed]
- Doblin, M.S.; Pettolino, F.A.; Wilson, S.M.; Campbell, R.; Burton, R.A.; Fincher, G.B.; Newbigin, E.; Bacic, A. A Barley Cellulose Synthase-like CSLH Gene Mediates (1,3;1,4)-β-d-Glucan Synthesis in Transgenic Arabidopsis. Proc Natl Acad Sci U S A 2009, 106, 5996–6001. [Google Scholar] [CrossRef]
- Yin, Y.; Huang, J.; Xu, Y. The Cellulose Synthase Superfamily in Fully Sequenced Plants and Algae. BMC Plant Biol 2009, 9, 99. [Google Scholar] [CrossRef]
- Little, A.; Lahnstein, J.; Jeffery, D.W.; Khor, S.F.; Schwerdt, J.G.; Shirley, N.J.; Hooi, M.; Xing, X.; Burton, R.A.; Bulone, V. A Novel (1,4)-β-Linked Glucoxylan Is Synthesized by Members of the Cellulose Synthase-Like F Gene Family in Land Plants. ACS Cent. Sci. 2019, 5, 73–84. [Google Scholar] [CrossRef]
- Zhang, J.; Yan, L.; Liu, M.; Guo, G.; Wu, B. Analysis of Beta-D-Glucan Biosynthetic Genes in Oat Reveals Glucan Synthesis Regulation by Light. Annals of Botany. [CrossRef]
- Dimitroff, G.; Little, A.; Lahnstein, J.; Schwerdt, J.G.; Srivastava, V.; Bulone, V.; Burton, R.A.; Fincher, G.B. (1,3;1,4)-β-Glucan Biosynthesis by the CSLF6 Enzyme: Position and Flexibility of Catalytic Residues Influence Product Fine Structure. Biochemistry 2016, 55, 2054–2061. [Google Scholar] [CrossRef]
- Nemeth, C.; Freeman, J.; Jones, H.D.; Sparks, C.; Pellny, T.K.; Wilkinson, M.D.; Dunwell, J.; Andersson, A.A.M.; Åman, P.; Guillon, F.; et al. Down-Regulation of the CSLF6 Gene Results in Decreased (1,3;1,4)- β - d -Glucan in Endosperm of Wheat. Plant Physiology 2010, 152, 1209–1218. [Google Scholar] [CrossRef] [PubMed]
- Little, A.; Schwerdt, J.G.; Shirley, N.J.; Khor, S.F.; Neumann, K.; O’Donovan, L.A.; Lahnstein, J.; Collins, H.M.; Henderson, M.; Fincher, G.B.; et al. Revised Phylogeny of the Cellulose Synthase Gene Superfamily: Insights into Cell Wall Evolution. Plant Physiology 2018, 177, 1124–1141. [Google Scholar] [CrossRef] [PubMed]
- Burton, R.A.; Collins, H.M.; Kibble, N.A.J.; Smith, J.A.; Shirley, N.J.; Jobling, S.A.; Henderson, M.; Singh, R.R.; Pettolino, F.; Wilson, S.M.; et al. Over-Expression of Specific HvCslF Cellulose Synthase-like Genes in Transgenic Barley Increases the Levels of Cell Wall (1,3;1,4)-β-d-Glucans and Alters Their Fine Structure: Over-Expression of CslF Genes in Barley. Plant Biotechnology Journal 2011, 9, 117–135. [Google Scholar] [CrossRef] [PubMed]
- Taketa, S.; Yuo, T.; Tonooka, T.; Tsumuraya, Y.; Inagaki, Y.; Haruyama, N.; Larroque, O.; Jobling, S.A. Functional Characterization of Barley Betaglucanless Mutants Demonstrates a Unique Role for CslF6 in (1,3;1,4)-β-D-Glucan Biosynthesis. Journal of Experimental Botany 2012, 63, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Schwerdt, J.G.; MacKenzie, K.; Wright, F.; Oehme, D.; Wagner, J.M.; Harvey, A.J.; Shirley, N.J.; Burton, R.A.; Schreiber, M.; Halpin, C.; et al. Evolutionary Dynamics of the Cellulose Synthase Gene Superfamily in Grasses. Plant Physiology 2015, 168, 968–983. [Google Scholar] [CrossRef] [PubMed]
- Vega-Sánchez, M.E.; Verhertbruggen, Y.; Christensen, U.; Chen, X.; Sharma, V.; Varanasi, P.; Jobling, S.A.; Talbot, M.; White, R.G.; Joo, M.; et al. Loss of Cellulose Synthase - Like F6 Function Affects Mixed-Linkage Glucan Deposition, Cell Wall Mechanical Properties, and Defense Responses in Vegetative Tissues of Rice. Plant Physiol. 2012, 159, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Burton, R.A.; Jobling, S.A.; Harvey, A.J.; Shirley, N.J.; Mather, D.E.; Bacic, A.; Fincher, G.B. The Genetics and Transcriptional Profiles of the Cellulose Synthase-Like HvCslF Gene Family in Barley. Plant Physiol. 2008, 146, 1821–1833. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Gimenez, G.; Barakate, A.; Smith, P.; Stephens, J.; Khor, S.F.; Doblin, M.S.; Hao, P.; Bacic, A.; Fincher, G.B.; Burton, R.A.; et al. Targeted Mutation of Barley (1,3;1,4)-β-glucan Synthases Reveals Complex Relationships between the Storage and Cell Wall Polysaccharide Content. Plant J. 2020, 104, 1009–1022. [Google Scholar] [CrossRef]
- Coon, M.A. Characterization and Variable Expression of the CslF6 Homologs in Oat (Avena Sp.). 2012. Master thesis. Brigham Young University – Provo. https://scholarsarchive.byu.edu/cgi/viewcontent.cgi?article=4749&context=etd.
- Wilson, S.M.; Ho, Y.Y.; Lampugnani, E.R.; Van de Meene, A.M.L.; Bain, M.P.; Bacic, A.; Doblin, M.S. Determining the Subcellular Location of Synthesis and Assembly of the Cell Wall Polysaccharide (1,3; 1,4)-β- d -Glucan in Grasses. The Plant Cell 2015, 27, 754–771. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Vendrell, A.M.; Guasch, J.; Francesch, M.; Molina-Cano, J.L.; Brufau, J. Determination of β-(1–3), (1–4)-D-Glucans in Barley by Reversed-Phase High-Performance Liquid Chromatography. Journal of Chromatography A 1995, 718, 291–297. [Google Scholar] [CrossRef]
- Redaelli, R.; Scalfati, G.; Ciccoritti, R.; Cacciatori, P.; De Stefanis, E.; Sgrulletta, D. Effects of Genetic and Agronomic Factors on Grain Composition in Oats. Cereal Research Communications 2015, 43, 144–154. [Google Scholar] [CrossRef]
- Savin, R.; Molina-Cano, J. Changes in Malting Quality and Its Determinants in Response to Abiotic Stresses. Barley science-recent advances from molecular biology to agronomy of yield and quality, Food Products Press, New York, 2002, 523–550.
- Güler, M. Barley Grain β-Glucan Content as Affected by Nitrogen and Irrigation. Field Crops Research 2003, 84, 335–340. [Google Scholar] [CrossRef]
- Peterson, D.M. Genotype and Environment Effects on Oat Beta-Glucan Concentration. Crop Science, 1991; 31. [Google Scholar] [CrossRef]
- Doehlert, D.C.; McMullen, M.S.; Hammond, J.J. Genotypic and Environmental Effects on Grain Yield and Quality of Oat Grown in North Dakota. Crop Science 2001, 41, 1066–1072. [Google Scholar] [CrossRef]
- Dvoncova, D.; Havrlentova, M.; Hlinkova, A.; Hozlar, P. Effect of Fertilization and Variety on the β-Glucan Content in the Grain of Oats. ZNTJ, 2010. [Google Scholar] [CrossRef]
- Gorash, A.; Armonienė, R.; Mitchell Fetch, J.; Liatukas, Ž.; Danytė, V. Aspects in Oat Breeding: Nutrition Quality, Nakedness and Disease Resistance, Challenges and Perspectives: Aspects in Oat Breeding. Ann Appl Biol 2017, 171, 281–302. [Google Scholar] [CrossRef]
- Dvořáček, V.; Jágr, M.; Kotrbová Kozak, A.; Capouchová, I.; Konvalina, P.; Faměra, O.; Hlásná Čepková, P. Avenanthramides: Unique Bioactive Substances of Oat Grain in the Context of Cultivar, Cropping System, Weather Conditions and Other Grain Parameters. Plants 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- De Arcangelis, E.; Messia, M.C.; Marconi, E. Variation of Polysaccharides Profiles in Developing Kernels of Different Barley Cultivars. Journal of Cereal Science 2019, 85, 273–278. [Google Scholar] [CrossRef]
- Mathews, R.; Kamil, A.; Chu, Y. Global Review of Heart Health Claims for Oat Beta-Glucan Products. Nutrition Reviews 2020, 78 (Supplement_1), 78–97. [Google Scholar] [CrossRef]
- Ahmad, A.; Anjum, F.M.; Zahoor, T.; Nawaz, H.; Dilshad, S.M.R. Beta Glucan: A Valuable Functional Ingredient in Foods. Critical Reviews in Food Science and Nutrition 2012, 52, 201–212. [Google Scholar] [CrossRef]
- Havrlentová, M.; Gregusová, V.; Šliková, S.; Nemeček, P.; Hudcovicová, M.; Kuzmová, D. Relationship between the Content of β-D-Glucans and Infection with Fusarium Pathogens in Oat (Avena Sativa L.) Plants. Plants 2020, 9, 1776. [Google Scholar] [CrossRef]
- Hrmova, M.; Fincher, G.B. Structure-Function Relationships of β- D-Glucan Endo- and Exohydrolases from Higher Plants. Plant Molecular Biology 2001, 47, 73–91. [Google Scholar] [CrossRef]
- Roulin, S.; Buchala, A.J.; Fincher, G.B. Induction of (1→3,1→4)-β-D-Glucan Hydrolases in Leaves of Dark-Incubated Barley Seedlings. Planta 2002, 215, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Guillon, F.; Tranquet, O.; Quillien, L.; Utille, J.-P.; Ordaz Ortiz, J.J.; Saulnier, L. Generation of Polyclonal and Monoclonal Antibodies against Arabinoxylans and Their Use for Immunocytochemical Location of Arabinoxylans in Cell Walls of Endosperm of Wheat. Journal of Cereal Science 2004, 40, 167–182. [Google Scholar] [CrossRef]
- Trafford, K.; Haleux, P.; Henderson, M.; Parker, M.; Shirley, N.J.; Tucker, M.R.; Fincher, G.B.; Burton, R.A. Grain Development in Brachypodium and Other Grasses: Possible Interactions between Cell Expansion, Starch Deposition, and Cell-Wall Synthesis. Journal of Experimental Botany 2013, 64, 5033–5047. [Google Scholar] [CrossRef] [PubMed]
- Carpita, N.C. Structure and Biogenesis of the Cell Walls of Grasses. Annual review of plant biology 1996, 47, 445–476. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-B.; Olek, A.T.; Carpita, N.C. Cell Wall and Membrane-Associated Exo-β-d-Glucanases from Developing Maize Seedlings1. Plant Physiology 2000, 123, 471–486. [Google Scholar] [CrossRef] [PubMed]
- Hoson, T. Apoplast as the Site of Response to Environmental Signals. Journal of Plant Research 1998, 111, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Havrlentová, M.; Šliková, S.; Gregusová, V.; Kovácsová, B.; Lančaričová, A.; Nemeček, P.; Hendrichová, J.; Hozlár, P. The Influence of Artificial Fusarium Infection on Oat Grain Quality. Microorganisms 2021, 9, 2108. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Schöneberg, T.; Vogelgsang, S.; Morisoli, R.; Bertossa, M.; Mauch-Mani, B.; Mascher, F. Resistance against Fusarium Graminearum and the Relationship to β-Glucan Content in Barley Grains. Eur J Plant Pathol 2018, 152, 621–634. [Google Scholar] [CrossRef]
- Gregusová, V.; Kaňuková, Š.; Hudcovicová, M.; Bojnanská, K.; Ondreičková, K.; Piršelová, B.; Mészáros, P.; Lengyelová, L.; Galuščáková, Ľ.; Kubová, V.; et al. The Cell-Wall β-d-Glucan in Leaves of Oat (Avena Sativa L.) Affected by Fungal Pathogen Blumeria Graminis f. Sp. Avenae. Polymers, 2022; 14. [Google Scholar] [CrossRef]
- Kofuji, K.; Aoki, A.; Tsubaki, K.; Konishi, M.; Isobe, T.; Murata, Y. Antioxidant Activity of β-Glucan. ISRN Pharm 2012, 2012, 125864. [Google Scholar] [CrossRef]
- Lattanzio, V.; Lattanzio, V.M.T.; Cardinali, A. Role of Phenolics in the Resistance Mechanisms of Plants against Fungal Pathogens and Insects. Phytochemistry: Advance in research 2006, 661, 23–67. [Google Scholar]
- Bai, Y.-P.; Zhou, H.-M.; Zhu, K.-R.; Li, Q. Effect of Thermal Processing on the Molecular, Structural, and Antioxidant Characteristics of Highland Barley β-Glucan. Carbohydrate Polymers 2021, 271, 118416. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, I.; Willats, W.G. Plant Cell Walls: New Insights from Ancient Species. Plant signaling & behavior 2008, 3, 743–745. [Google Scholar] [CrossRef]
- Kerstens, S.; Decraemer, W.F.; Verbelen, J.-P. Cell Walls at the Plant Surface Behave Mechanically like Fiber-Reinforced Composite Materials. Plant Physiology 2001, 127, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Holthaus, J.; Holland, J.; White, P.; Frey, K. Inheritance of Β-glucan Content of Oat Grain. Crop Science 1996, 36, 567–572. [Google Scholar] [CrossRef]
- Eshghi, R.; Akhundova, E. Inheritance Pattern of β-Glucan and Protein Countents in Hulless Barley. International Journal of Agriculture and Biology 2010, 12, 68–72. [Google Scholar]
- Swanston, J. The Barley Husk: A Potential Barrier to Future Success. Barley: Physical properties, genetic factors and environmental impacts on growth. Nova Science Publishers, inc., New york 2014, 81–106.
- Marcotuli, I.; Houston, K.; Schwerdt, J.G.; Waugh, R.; Fincher, G.B.; Burton, R.A.; Blanco, A.; Gadaleta, A. Genetic Diversity and Genome Wide Association Study of β-Glucan Content in Tetraploid Wheat Grains. PLoS One 2016, 11, e0152590. [Google Scholar] [CrossRef] [PubMed]
- Walling, J.G.; Sallam, A.H.; Steffenson, B.J.; Henson, C.; Vinje, M.A.; Mahalingam, R. Quantitative Trait Loci Impacting Grain Β-glucan Content in Wild Barley (Hordeum Vulgare Ssp. Spontaneum) Reveals Genes Associated with Cell Wall Modification and Carbohydrate Metabolism. Crop Science 2022, 62, 1213–1227. [Google Scholar] [CrossRef]
- Geng, L.; Li, M.; Xie, S.; Wu, D.; Ye, L.; Zhang, G. Identification of Genetic Loci and Candidate Genes Related to β-Glucan Content in Barley Grain by Genome-Wide Association Study in International Barley Core Selected Collection. Molecular Breeding 2021, 41, 1–12. [Google Scholar] [CrossRef]
- Geng, L.; He, X.; Ye, L.; Zhang, G. Identification of the Genes Associated with β-Glucan Synthesis and Accumulation during Grain Development in Barley. Food Chemistry: Molecular Sciences 2022, 5, 100136. [Google Scholar] [CrossRef]
- Newell, M.A.; Asoro, F.G.; Scott, M.P.; White, P.J.; Beavis, W.D.; Jannink, J.-L. Genome-Wide Association Study for Oat (Avena Sativa L.) Beta-Glucan Concentration Using Germplasm of Worldwide Origin. Theor Appl Genet 2012, 125, 1687–1696. [Google Scholar] [CrossRef]
- Zimmer, C.M.; Oliveira, G.; Pacheco, M.T.; Federizzi, L.C. Characterization and Absolute Quantification of the Cellulose Synthase-like F6 Homoeologs in Oats. Euphytica 2020, 216, 173. [Google Scholar] [CrossRef]
- Motilva, M.-J.; Serra, A.; Borrás, X.; Romero, M.-P.; Domínguez, A.; Labrador, A.; Peiró, L. Adaptation of the Standard Enzymatic Protocol (Megazyme Method) to Microplaque Format for β-(1, 3)(1, 4)-d-Glucan Determination in Cereal Based Samples with a Wide Range of β-Glucan Content. Journal of Cereal Science 2014, 59, 224–227. [Google Scholar] [CrossRef]
- Paudel, D.; Caffe-Treml, M.; Krishnan, P. A Single Analytical Platform for the Rapid and Simultaneous Measurement of Protein, Oil, and Beta-Glucan Contents of Oats Using Near-Infrared Reflectance Spectroscopy. Cereal Foods World 2018, 63, 17–25. [Google Scholar] [CrossRef]
- van der Kamp, J.W.; Asp, N.-G.; Miller-Jones, J.; Schaafsma, G. Dietary Fibre: Bio-Active Carbohydrates for Food and Feed; Wageningen Academic Publishers, 2004.
- Henrion, M.; Francey, C.; Lê, K.-A.; Lamothe, L. Cereal B-Glucans: The Impact of Processing and How It Affects Physiological Responses. Nutrients 2019, 11, 1729. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Li, L.; Wang, X. Molecular Characterization of Arabinoxylans from Hull-Less Barley Milling Fractions. Molecules 2011, 16, 2743–2753. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Rossnagel, B.; Tyler, R.; Bhatty, R. Distribution of Β-glucan in the Grain of Hull-less Barley. Cereal Chemistry 2000, 77, 140–144. [Google Scholar] [CrossRef]
- De Brier, N.; Hemdane, S.; Dornez, E.; Gomand, S.V.; Delcour, J.A.; Courtin, C.M. Structure, Chemical Composition and Enzymatic Activities of Pearlings and Bran Obtained from Pearled Wheat (Triticum Aestivum L.) by Roller Milling. Journal of Cereal Science 2015, 62, 66–72. [Google Scholar] [CrossRef]
- Izydorczyk, M.; Cenkowski, S.; Dexter, J. Optimizing the Bioactive Potential of Oat Bran by Processing. Cereal Foods World 2014, 59, 127–136. [Google Scholar] [CrossRef]
- Tosi, E.A.; Ré, E.; Lucero, H.; Masciarelli, R. Dietary Fiber Obtained from Amaranth (Amaranthus Cruentus) Grain by Differential Milling. Food Chemistry 2001, 73, 441–443. [Google Scholar] [CrossRef]
- Escarnot, E.; Dornez, E.; Verspreet, J.; Agneessens, R.; Courtin, C.M. Quantification and Visualization of Dietary Fibre Components in Spelt and Wheat Kernels. Journal of Cereal Science 2015, 62, 124–133. [Google Scholar] [CrossRef]
- Cavallero, A.; Empilli, S.; Brighenti, F.; Stanca, A. High (1→ 3, 1→ 4)-β-Glucan Barley Fractions in Bread Making and Their Effects on Human Glycemic Response. Journal of cereal science 2002, 36, 59–66. [Google Scholar] [CrossRef]
- Holtekjølen, A.; Olsen, H.; Færgestad, E.; Uhlen, A.; Knutsen, S. Variations in Water Absorption Capacity and Baking Performance of Barley Varieties with Different Polysaccharide Content and Composition. LWT-Food Science and Technology 2008, 41, 2085–2091. [Google Scholar] [CrossRef]
- Izydorczyk, M.; Chornick, T.; Paulley, F.; Edwards, N.; Dexter, J. Physicochemical Properties of Hull-Less Barley Fibre-Rich Fractions Varying in Particle Size and Their Potential as Functional Ingredients in Two-Layer Flat Bread. Food Chemistry 2008, 108, 561–570. [Google Scholar] [CrossRef]
- Knuckles, B.; Hudson, C.; Chiu, M.; Sayre, R. Effect of Beta-Glucan Barley Fractions in High-Fiber Bread and Pasta. Cereal foods world (USA) 1997.
- Jacobs, M.S.; Izydorczyk, M.S.; Preston, K.R.; Dexter, J.E. Evaluation of Baking Procedures for Incorporation of Barley Roller Milling Fractions Containing High Levels of Dietary Fibre into Bread. Journal of the Science of Food and Agriculture 2008, 88, 558–568. [Google Scholar] [CrossRef]
- Kinner, M.; Nitschko, S.; Sommeregger, J.; Petrasch, A.; Linsberger-Martin, G.; Grausgruber, H.; Berghofer, E.; Siebenhandl-Ehn, S. Naked Barley—Optimized Recipe for Pure Barley Bread with Sufficient Beta-Glucan According to the EFSA Health Claims. Journal of cereal science 2011, 53, 225–230. [Google Scholar] [CrossRef]
- Andersson, R.; Fransson, G.; Tietjen, M.; Åman, P. Content and Molecular-Weight Distribution of Dietary Fiber Components in Whole-Grain Rye Flour and Bread. Journal of Agricultural and Food Chemistry 2009, 57, 2004–2008. [Google Scholar] [CrossRef]
- Rieder, A.; Holtekjølen, A.K.; Sahlstrøm, S.; Moldestad, A. Effect of Barley and Oat Flour Types and Sourdoughs on Dough Rheology and Bread Quality of Composite Wheat Bread. Journal of Cereal Science 2012, 55, 44–52. [Google Scholar] [CrossRef]
- Marklinder, I.; Johansson, L.; Haglund, Å.; Nagel-Held, B.; Seibel, W. Effects of Flour from Different Barley Varieties on Barley Sour Dough Bread. Food Quality and Preference 1996, 7, 275–284. [Google Scholar] [CrossRef]
- Lambo, A.M.; Öste, R.; Nyman, M.E.-L. Dietary Fibre in Fermented Oat and Barley β-Glucan Rich Concentrates. Food Chemistry 2005, 89, 283–293. [Google Scholar] [CrossRef]
- Salovaara, H.; Gates, F.; Tenkanen, M. Dietary Fibre Components and Functions; Wageningen Academic Publishers, 2007.
- Andersson, R.; Åman, P. 27 Cereal Arabinoxylan: Occurrence, Structure and Properties. Advanced dietary fibre technology 2008, 301. [Google Scholar]
- Comino, P.; Collins, H.; Lahnstein, J.; Gidley, M.J. Effects of Diverse Food Processing Conditions on the Structure and Solubility of Wheat, Barley and Rye Endosperm Dietary Fibre. Journal of Food Engineering 2016, 169, 228–237. [Google Scholar] [CrossRef]
- Tiwari, U.; Cummins, E.; Sullivan, P.; Flaherty, J.O.; Brunton, N.; Gallagher, E. Probabilistic Methodology for Assessing Changes in the Level and Molecular Weight of Barley β-Glucan during Bread Baking. Food Chemistry 2011, 124, 1567–1576. [Google Scholar] [CrossRef]
- Rakha, A.; Åman, P.; Andersson, R. Characterisation of Dietary Fibre Components in Rye Products. Food Chemistry 2010, 119, 859–867. [Google Scholar] [CrossRef]
- Sozer, N.; Poutanen, K. Fibre in Extruded Products. Fibre-Rich and Wholegrain Foods-Improving Quality 2013, 226–272. [CrossRef]
- Gajula, H.; Alavi, S.; Adhikari, K.; Herald, T. Precooked Bran-Enriched Wheat Flour Using Extrusion: Dietary Fiber Profile and Sensory Characteristics. Journal of Food Science 2008, 73, S173–S179. [Google Scholar] [CrossRef]
- Sobota, A.; Sykut-Domańska, E.; Rzedzicki, Z. Effect of Extrusion-Cooking Process on the Chemical Compositon of Corn-Wheat Extrudates, With Particular Emphasis on Dietary Fibre Fractions. Pol. J. Food Nutr. Sci. 2010, 60, 251–259. [Google Scholar]
- Vasanthan, T.; Gaosong, J.; Yeung, J.; Li, J. Dietary Fiber Profile of Barley Flour as Affected by Extrusion Cooking. Food Chemistry 2002, 77, 35–40. [Google Scholar] [CrossRef]
- Andersson, A.A.M.; Armö, E.; Grangeon, E.; Fredriksson, H.; Andersson, R.; Åman, P. Molecular Weight and Structure Units of (1→3, 1→4)-β-Glucans in Dough and Bread Made from Hull-Less Barley Milling Fractions. Journal of Cereal Science 2004, 40, 195–204. [Google Scholar] [CrossRef]
- Gujral, H.S.; Sharma, P.; Rachna, S. Effect of Sand Roasting on Beta Glucan Extractability, Physicochemical and Antioxidant Properties of Oats. LWT - Food Science and Technology 2011, 44, 2223–2230. [Google Scholar] [CrossRef]
- Sharma, P.; Gujral, H.S. Extrusion of Hulled Barley Affecting β-Glucan and Properties of Extrudates. Food and Bioprocess Technology 2013, 6, 1374–1389. [Google Scholar] [CrossRef]
- Schmidt, M. Cereal Beta-Glucans: An Underutilized Health Endorsing Food Ingredient. Critical Reviews in Food Science and Nutrition 2022, 62, 3281–3300. [Google Scholar] [CrossRef]
- Loebnitz, N.; Grunert, K.G. Impact of Self-Health Awareness and Perceived Product Benefits on Purchase Intentions for Hedonic and Utilitarian Foods with Nutrition Claims. Food Quality and Preference 2018, 64, 221–231. [Google Scholar] [CrossRef]
- Wood, P.J. Cereal β-Glucans in Diet and Health. Journal of Cereal Science 2007, 46, 230–238. [Google Scholar] [CrossRef]
- Zeković, D.B.; Kwiatkowski, S.; Vrvić, M.M.; Jakovljević, D.; Moran, C.A. Natural and Modified (1→3)-β-D-Glucans in Health Promotion and Disease Alleviation. Critical Reviews in Biotechnology 2005, 25, 205–230. [Google Scholar] [CrossRef]
- Ortiz de Erive, M.; He, F.; Wang, T.; Chen, G. Development of β-Glucan Enriched Wheat Bread Using Soluble Oat Fiber. Journal of Cereal Science 2020, 95, 103051. [Google Scholar] [CrossRef]
- Hager, A.-S.; Ryan, L.A.M.; Schwab, C.; Gänzle, M.G.; O’Doherty, J.V.; Arendt, E.K. Influence of the Soluble Fibres Inulin and Oat β-Glucan on Quality of Dough and Bread. European Food Research and Technology 2011, 232, 405–413. [Google Scholar] [CrossRef]
- De Paula, R.; Abdel-Aal, E.-S.M.; Messia, M.C.; Rabalski, I.; Marconi, E. Effect of Processing on the Beta-Glucan Physicochemical Properties in Barley and Semolina Pasta. Journal of Cereal Science 2017, 75, 124–131. [Google Scholar] [CrossRef]
- Choo, C.L.; Aziz, N.A.A. Effects of Banana Flour and β-Glucan on the Nutritional and Sensory Evaluation of Noodles. Food Chemistry 2010, 119, 34–40. [Google Scholar] [CrossRef]
- Messia, M.C.; Oriente, M.; Angelicola, M.; De Arcangelis, E.; Marconi, E. Development of Functional Couscous Enriched in Barley β-Glucans. Journal of Cereal Science 2019, 85, 137–142. [Google Scholar] [CrossRef]
- Moza, J.; Gujral, H.S. Influence of Barley Non-Starchy Polysaccharides on Selected Quality Attributes of Sponge Cakes. LWT - Food Science and Technology 2017, 85, 252–261. [Google Scholar] [CrossRef]
- Vitaglione, P.; Lumaga, R.B.; Montagnese, C.; Messia, M.C.; Marconi, E.; Scalfi, L. Satiating Effect of a Barley Beta-Glucan–Enriched Snack. Journal of the American College of Nutrition 2010, 29, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.S.; Tudorica, C.M. Carbohydrate-Based Fat Replacers in the Modification of the Rheological, Textural and Sensory Quality of Yoghurt: Comparative Study of the Utilisation of Barley Beta-Glucan, Guar Gum and Inulin. International Journal of Food Science & Technology 2008, 43, 824–833. [Google Scholar] [CrossRef]
- Raikos, V.; Grant, S.B.; Hayes, H.; Ranawana, V. Use of β-Glucan from Spent Brewer’s Yeast as a Thickener in Skimmed Yogurt: Physicochemical, Textural, and Structural Properties Related to Sensory Perception. Journal of Dairy Science 2018, 101, 5821–5831. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, B.; Patel, T. Increased Sensory Quality and Consumer Acceptability by Fortification of Chocolate Flavored Milk with Oat Beta Glucan. International Journal of Clinical and Biomedical Research 2016, 2, 25–28. [Google Scholar]
- Volikakis, P.; Biliaderis, C.G.; Vamvakas, C.; Zerfiridis, G.K. Effects of a Commercial Oat-β-Glucan Concentrate on the Chemical, Physico-Chemical and Sensory Attributes of a Low-Fat White-Brined Cheese Product. Food Research International 2004, 37, 83–94. [Google Scholar] [CrossRef]
- Santipanichwong, R.; Suphantharika, M. Carotenoids as Colorants in Reduced-Fat Mayonnaise Containing Spent Brewer’s Yeast β-Glucan as a Fat Replacer. Food Hydrocolloids 2007, 21, 565–574. [Google Scholar] [CrossRef]
- Bhaskar, D.; Khatkar, S.K.; Chawla, R.; Panwar, H.; Kapoor, S. Effect of β-Glucan Fortification on Physico-Chemical, Rheological, Textural, Colour and Organoleptic Characteristics of Low Fat Dahi. Journal of Food Science and Technology 2017, 54, 2684–2693. [Google Scholar] [CrossRef]
- Elsanhoty, R.; Zaghlol, A.; Hassanein, A. The Manufacture of Low Fat Labneh Containing Barley β-Glucan 1-Chemical Composition, Microbiological Evaluation and Sensory Properties. Current Research in Dairy Sciences 2009, 1, 1–12. [Google Scholar] [CrossRef]
- Vithanage, C.; Mishra, V.K.; Vasiljevic, T.; Shah, N.P. Use of β-Glucan in Development of Low-Fat Mozzarella Cheese. Milchwissenschaft 2008. [Google Scholar]
- Mykhalevych, A.; Polishchuk, G.; Nassar, K.; Osmak, T.; Buniowska-Olejnik, M. β-Glucan as a Techno-Functional Ingredient in Dairy and Milk-Based Products—A Review. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Lyly, M.; Salmenkallio-Marttila, M.; Suortti, T.; Autio, K.; Poutanen, K.; Lähteenmäki, L. Influence of Oat β-Glucan Preparations on the Perception of Mouthfeel and on Rheological Properties in Beverage Prototypes. Cereal Chemistry 2003, 80, 536–541. [Google Scholar] [CrossRef]
- Karp, S.; Wyrwisz, J.; Kurek, M.A. Comparative Analysis of the Physical Properties of o/w Emulsions Stabilised by Cereal β-Glucan and Other Stabilisers. International Journal of Biological Macromolecules 2019, 132, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, N.; Xiong, Z.; Wang, G.; Xia, Y.; Lai, P.; Ai, L. Structural Characterization and Rheological Properties of β-D-Glucan from Hull-Less Barley (Hordeum Vulgare L. Var. Nudum Hook. f.). Phytochemistry 2018, 155, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Angelov, A.; Gotcheva, V.; Kuncheva, R.; Hristozova, T. Development of a New Oat-Based Probiotic Drink. International Journal of Food Microbiology 2006, 112, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Harasym, J.; Suchecka, D.; Gromadzka-Ostrowska, J. Effect of Size Reduction by Freeze-Milling on Processing Properties of Beta-Glucan Oat Bran. Journal of Cereal Science 2015, 61, 119–125. [Google Scholar] [CrossRef]
- Jayachandran, M.; Chen, J.; Chung, S.S.M.; Xu, B. A Critical Review on the Impacts of β-Glucans on Gut Microbiota and Human Health. The Journal of Nutritional Biochemistry 2018, 61, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Avramia, I.; Amariei, S. Spent Brewer’s Yeast as a Source of Insoluble β-Glucans. International Journal of Molecular Sciences 2021, 22. [Google Scholar] [CrossRef]
- Singh, M.; Kim, S.; Liu, S.X. Effect of Purified Oat β-Glucan on Fermentation of Set-Style Yogurt Mix*. Journal of Food Science 2012, 77, E195–E201. [Google Scholar] [CrossRef]
- Sahan, N.; Yasar, K.; Hayaloglu, A.A.; Karaca, O.B.; Kaya, A. Influence of Fat Replacers on Chemical Composition, Proteolysis, Texture Profiles, Meltability and Sensory Properties of Low-Fat Kashar Cheese. Journal of Dairy Research 2008, 75, 1–7. [Google Scholar] [CrossRef]
- Tudorica, C.M.; Jones, T.E.R.; Kuri, V.; Brennan, C.S. The Effects of Refined Barley β-Glucan on the Physico-Structural Properties of Low-Fat Dairy Products: Curd Yield, Microstructure, Texture and Rheology. Journal of the Science of Food and Agriculture 2004, 84, 1159–1169. [Google Scholar] [CrossRef]
Figure 1.
The share of the main significant factors and interactions in the total sum of squares (%) according to analysis of variance (ANOVA) model for selected grain parameters of five oat cultivars grown at two localities and different growing systems (organic vs. conventional) over 3 years. Avenanthramide (AVN), total content of tested AVNs (Σ AVNs), crude protein (CP), starch (ST), fat (FT), ß-D-glucan (ß-GLU), avenin protein fraction (AVE), glutelin protein fraction (GLU), immunoreactive avenin peptides (G12), ash (ASH), thousand-grain weight (TGW), yield (YLD) [78].
Figure 1.
The share of the main significant factors and interactions in the total sum of squares (%) according to analysis of variance (ANOVA) model for selected grain parameters of five oat cultivars grown at two localities and different growing systems (organic vs. conventional) over 3 years. Avenanthramide (AVN), total content of tested AVNs (Σ AVNs), crude protein (CP), starch (ST), fat (FT), ß-D-glucan (ß-GLU), avenin protein fraction (AVE), glutelin protein fraction (GLU), immunoreactive avenin peptides (G12), ash (ASH), thousand-grain weight (TGW), yield (YLD) [78].
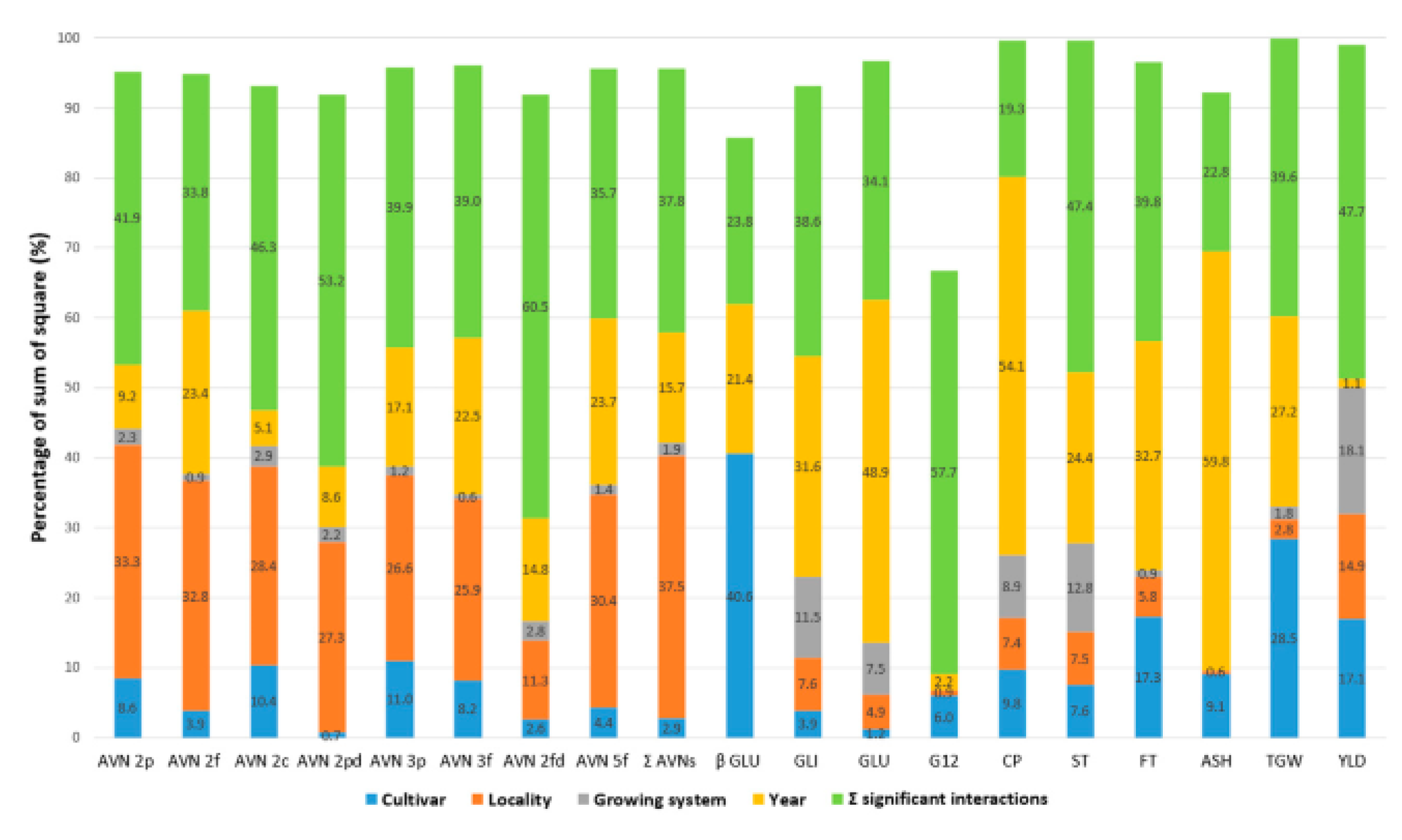
Figure 2.
The proposed biological process of β-D-glucans synthesis during grain development. A. Proposed model system of barley grain β-D-glucans biosynthesis. The model shows some key enzymes and DEGs that are involved in β-glucan biosynthesis during grain development [104].
Figure 2.
The proposed biological process of β-D-glucans synthesis during grain development. A. Proposed model system of barley grain β-D-glucans biosynthesis. The model shows some key enzymes and DEGs that are involved in β-glucan biosynthesis during grain development [104].
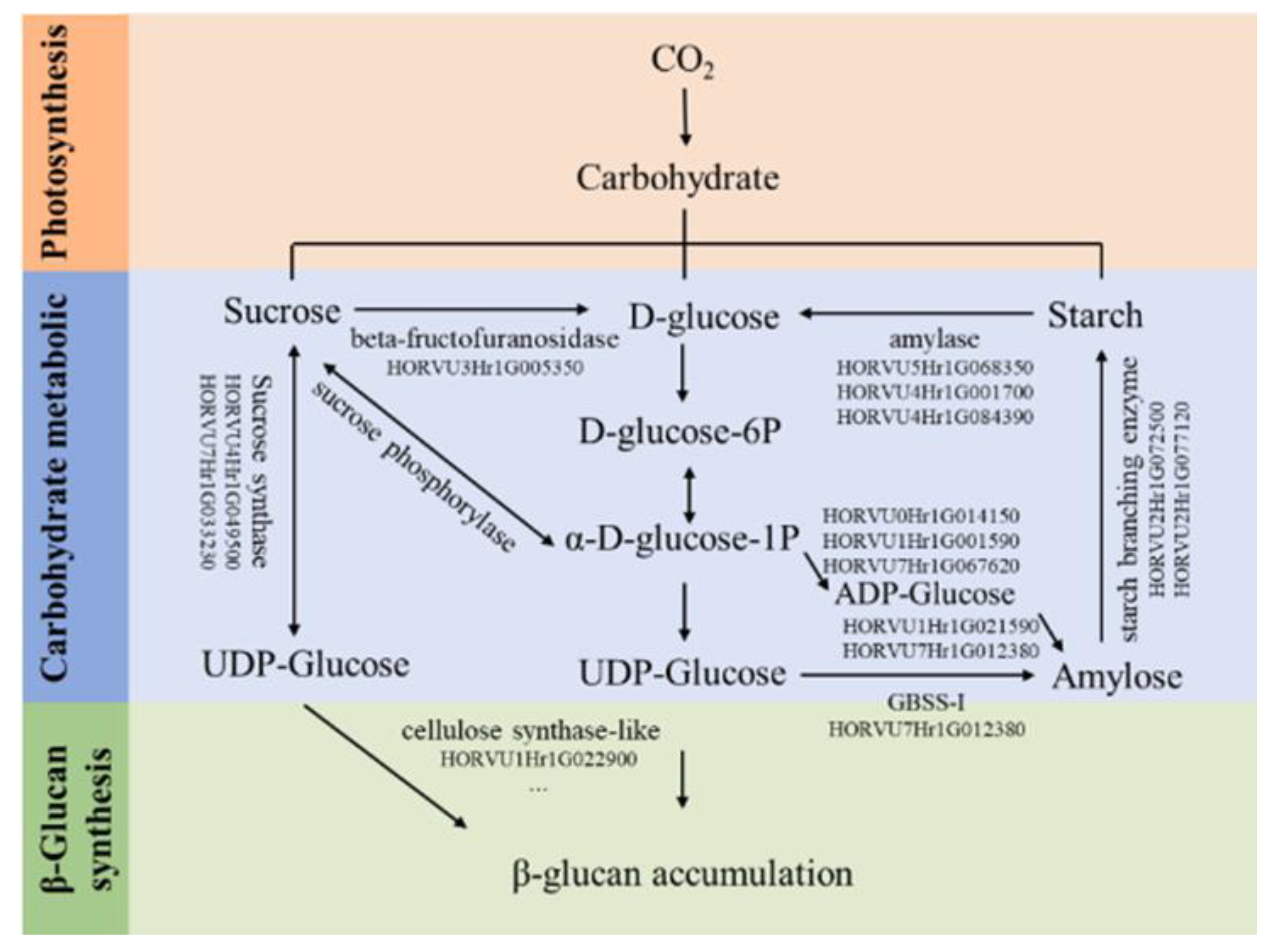

Table 1.
Examples of β-D-glucans applications in food processing .
| Application | Addition | Source | Product | References |
|---|---|---|---|---|
| Cereal processing | 2.5 and 5 % | barley | bread | [81] |
| Cereal processing | 0.2, 0.6, 1.0, and 1.4 % | barley | bread | [19] |
| Cereal processing | 10, 12, and 14 % soluble oat fiber (70% β-D-glucans) | oat | bread | [143] |
| Cereal processing | 2.6 and 5.6% | whey | bread | [144] |
| Cereal processing | 30% high-glucan barley flour | barley | pasta | [145] |
| Cereal processing | 10% | oat | yellow alkaline noodles | [146] |
| Cereal processing | 20 and 30% high-glucan barley flour | barley | couscous | [147] |
| Cereal processing | 100% barley flour (3.4-4.4% β-D-glucans) |
barley | sponge cake | [148] |
| Cereal processing | 5.2% | barley | biscuit bar | [149] |
| Milk processing | 0.5% | barley | yoghurt | [150] |
| Milk processing | 0.2-0.8% | yeast | yoghurt | [151] |
| Milk beverage production | 3% | oat | chocolate-flavored milk | [152] |
| Cheese production | 0.7 and 1.4% | oat | white-brined cheese | [153] |
| Cheese production | fat replacement 3.47% and 6.84% | yeast | cheddar cheese | [154] |
| Cheese production | 0.5% | barley | Dahi cheese | [155] |
| Cheese production | 5% | barley | Labneh | [156] |
| Cheese production | 0.2% | barley | mozzarella | [157] |
| Milk processing | 0.5% | cereal (not specified) | cottage cheese | [158] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



