
Submitted:
04 May 2023
Posted:
05 May 2023
You are already at the latest version
Abstract
Olfaction plays a crucial role in many important behavioral activities of insects, such as finding sexual partners, locating hosts, and selecting oviposition sites. Ionotropic receptors (IRs) play a central role in detecting chemosensory information from the environment and guiding insect behaviors and are potential target genes for pest control. Empoasca onukii Matsuda is a major pest of the tea plant Camellia sinensis (L.) O. Ktze, and seriously influences tea yields and quality. In this study, the ionotropic receptor gene EnouIR25a in E. onukii was cloned, and the expression pattern of EnouIR25a was detected in various tissues. Behavioral responses of E. onukii to volatile compounds emitted by tea plants were determined by the olfactometer bioassay and field trials. To further explore the function of EnouIR25a in olfactory recognition of compounds, RNAi (RNA interference) of EnouIR25a was carried out by ingestion of in vitro synthesized dsRNAs. The CDS length of EnouIR25a was 1266 bp and it encoded a 48.87 kD protein. EnouIR25a was enriched in the antennae of E. onukii. E. onukii were more significantly attracted by 1-phenylethanol at concentration of 100 µl/ml. Feeding with dsRNA-IR25a significantly downregulated the expression level of EonuIR25a, after 3 h of treatment, which disturbed the behavioral responses of E. onukii to 1-phenylethanol at concentration of 100µl/ml. The response rate of E. onukii to 1-phenylethanol was significantly decreased after dsRNA-IR25a treatment for 12 h. In summary, the ionotropic receptor gene EnouIR25a was highly expressed in the antennae of E. onukii and was involved in olfactory recognition of the tea plant volatile 1-phenylethanol. The present study may help us to use the ionotropic receptor gene as a target for the behavioral manipulation of E. onukii in the future.
Keywords:
Empoasca onukii Matsuda
; EonuIR25a
; plant volatiles
; RNA interference
; olfactory system
1. Introduction
The tea plant Camellia sinensis (L.) O. Kuntze, is a significant cash crop in Asian countries, such as China, India, and Sri Lanka. The tender tea buds and leaves are usually plucked to produce high-grade tea as a beverage[1]. However, C. sinensis suffers heavily from attacks by many herbivorous insects in their life cycles. The tea green leafhopper, Empoasca (Matsumurasca) onukii Matsuda, an extremely harmful piercing pest with ten generations per year, is by far the most grave threat to tea plant cultivation[2]. Both nymphs and adults of E. onukii attack tea plants by using their piercing mouthparts (stylet), which ultimately results in the plant’s yellowing, browning, and drying[3]. The most common methods used to control E. onukii are regularly applying chemical insecticides. However, the excessive use of pesticides is jeopardizing both human health and the environment. Organisms have genetically evolved to diversify to accommodate their environment[4].
Insect chemoreception systems play a crucial role in many aspects of insect behaviors, such as foraging, mate recognition, oviposition, and predator avoidance[5,6]. The chemosensory system of insects plays an important role in insect and ecological niche adaptation and population evolution[7]. Chemosensory receptor proteins are normally expressed in the dendrites of chemosensory neurons and are specifically responsible for the interaction with chemosensory signals[8,9]. Olfaction is a highly specific, intricate, and extraordinarily sensitive system, and in the insect olfactory perception process, various olfactory proteins containing odorant binding proteins (OBPs), chemosensory proteins (CSPs), odorant receptors (OR), ionotropic receptors (IR), and odorant degrading enzymes are involved[10,11]. Most insect olfactory receptors belong to two distinct protein families, ionotropic receptors (IRs), which are associated with the ionotropic glutamate receptor family, and odorant receptors (ORs), which evolved from the gustatory receptor family. A large and highly divergent family of ionotropic glutamate receptor (iGluR)-related genes, called ionotropic receptors (IRs), was screened and identified[12]. Four protein structural domains are covered in the structure of iGluR, including the amino-terminal domain (ATD), ligand binding domain (LBD), transmembrane domain (TMD), and carboxy-terminal domain (CTD) [13]. The extracellular ATD structural domain is followed by the LBD structural domain, which contains two semistructural domains, S1 and S2[14]. In the primary structure, S1 and S2 are separated by ion channel pores. The ion channel pore is formed by two transmembrane segments (TM1 and TM2) and a folded-in pore loop[15]. S2 is followed by a third transmembrane segment (TM3) and the cytoplasmic carboxy-terminal structural domain (CTD). The ATD, LBD, TMD, CTD, and coreceptor extra loop (CREL) are contained in coreceptors IR8a and IR25a[12,16].
IR25a is a highly conserved gene that evolved from a bilaterian non-NMDA receptor gene[16]. IR25a is expressed in different insect tissues, such as adult tentacles[17], proboscis[18,19], legs, wings and abdomen[20]. IR is involved in processes such as olfaction, taste sensation, hygrosensation and cold sensation in insects[21,22]. Drosophila IR93a, IR25a and IR40a are coexpressed in adult tentacles and are able to sense humidity in the environment[23]. In the arbovirus vector Aedes aegypti, AeIr8a null mutants lost neuronal and behavioral responses to acids[24]. RNA interference (RNAi) knockdown of AcIr76b specifically impacts larval responses to butylamine in Anopheles coluzzii[25]. In Mythimna separata, MsepIr8a is a possible acid coreceptor among the putative acid-sensing IRs characterized[26]. In Manduca sexta, IR8a is essential for hexanoic acid and 3-methylpentanoic acid-mediated fecal avoidance[27]. IR41a is sufficient to confer sensitivity to amine/imine compounds in Anopheles gambiae[28]. IR25a and IR76b are able to synergistically mediate the response to flavoring agents, including acetic acid, citric acid, tartaric acid, and hydrochloric acid therein[29]. In addition, IR25a is expressed in neurons of Drosophila larvae and adults to support thermosensation and wet sensation[23,30,31]. The study of the molecular mechanisms and functions of multiple sensory modalities of IR in insects will thus contribute to the use of olfactory mechanisms for efficient control of pests.
In this study, the ionotropic receptor gene EnouIR25a in E. onukii was cloned and bioinformatically analyzed. To characterize the EnouIR25a gene, sequence alignment and phylogenetic analysis were investigated. The expression levels of the EnouIR25a gene in different tissues of E. onukii were determined by quantitative real-time PCR (qRT-PCR). Behavioral responses of E. onukii to volatile compounds emitted by tea plants were determined by the olfactometer bioassay and field trials. To further explore the function of EnouIR25a in olfactory recognition of these compounds, RNAi of EnouIR25a was carried out by feeding with in vitro synthesized dsRNAs. This study provides the first comprehensive characterization of the EnouIR25a gene from E. onukii. Our findings will provide valuable insights into the design and implementation of novel strategies to control the damage caused by this tea plant pest.
2. Results
2.1. Sequence analysis of the EonuIR25a gene
The sequence obtained was identified by NCBI as the EonuIR25a sequence with accession number OQ064772. The full-length ORF consisted of 1266 nucleotides and encoded 422 amino acid residues; the molecular weight of the protein was 48.87 kD. The specificity of the gene was confirmed by agarose gel electrophoresis (Figure 1). The theoretical isoelectric point was 6.50, the extinction coefficient was 98695, the lipid coefficient was 89.92, the instability coefficient was 55.03, the mean hydrophobicity coefficient was -0.372, and it is a hydrophilic protein (Figure 2a). The EonuIR25a protein was composed of 20 amino acids, with leucine as the main component, accounting for 10.2% of the total amino acids, and histidine accounting for only 1.0% of the total (Table 2). The amino acid sequence of the EonuIR25a protein has four transmembrane domains located at amino acids 7-24, 44-62, 119-141, and 342-364 (Figure 2b). The results of phosphorylation site prediction showed that EonuIR25a contains 46 phosphorylation sites, mainly in serine and threonine structures (Figure 2c), and no signal peptide in the amino acid sequence (Figure 2d). In the secondary structure of EonuIR25a, the α-helix is the main component of the amino acid sequence, accounting for 47.51% of the total structural proportion and corresponding to the transmembrane region, while the β-turn is lacking, accounting for only 3.09% of the total structural proportion (Figure 2e). The EonuIR25a contains the conserved structural domains of Lig_chan (positions 41-350) and SBP_bac_3 (positions 1-317), and the tertiary structure of EonuIR25a has similarity to the secondary structure, including the ligand-binding domain (LBD) (S1: 7-24, 119- 141; S2: 44-62) and the Ion channel domain, and the N-terminus on the outside of the cell membrane and the C-terminus on the inside (Figure 2f).
2.2. Multiple sequence alignment and phylogenetic tree analysis
The results of multiple sequence comparison showed that the sequence similarity between EonuIR25a and IR25a of other hemiptera ranged from 30-40%, and the sequence similarity with HvitIR25a (KAG8282323.1) of Homalodisca vitripennis was relatively high, with a sequence similarity of 36.52%, and it has a low sequence similarity of 31.63% with AlucIR25a (QFU27937.1) of Apolygus lucorum. In addition, the similar sequences were mainly concentrated in the central and posterior parts (Figure 3). Phylogenetic tree analysis showed that EonuIR25a clustered into clearly separated branches, and it clustered with the IR25a of Adelphocoris lineolatus, Apolygus lucorum, Halyomorpha halys, Nilaparvata lugens, Aphis gossypii, and Subpsaltria yangi. EonuIR25a was clustered into the smallest branch with HvitIR25a and HvitIR25a-like of Homalodisca vitripennis.Within the IR25a branch, the sequences cluster in an order-specific manner, reflecting that the highest similarity exists between sequences of insects belonging to the same type of insect gene. Considering the highly conserved nature of IR25a, it is speculated that it performs similar functions in different species (Figure 4).
2.3. Tissue expression analysis
The expression of EonuIR25a gene was 0.97 ± 0.01 and 1.00 ± 0.03 in female and male adult antennae, respectively, which was significantly higher than other tissues, and the expression of gene in male legs was only 0.16 ± 0.00. The expression of the EonuIR25a in male antennae was 6.16-fold and 4.5-fold higher than in male legs and male thorax, respectively. The distribution of EonuIR25a gene expression in the head and abdomen was higher in male adults than in females, but the distribution of expression in the legs was higher in female adults than in males. In addition, therewas significant difference between differfent tissues of EonuIR25a (d.f.=9, F=236.064, p<0.01), and there was no difference in antennae and thorax between male and female adults of EonuIR25a (P>0.05) (Figure 5).
note: A: antenna; H: head; T: thorax; Ab: abdomen; L: leg. Data in the figure are mean ± SE. Different lowercase letters on bars of the same color indicate significant differences in the relative expression levels of EonuIR25a among different tissues of male and female adults by Duncan’s test (P<0.05).
2.4. Correspondence of the compounds by E. onukii
After 3 days of treatment, phenylacetaldehyde was attractive to E. onukii at a concentration of 100 µl/ml, and the insect population of its sticky boards was 110.67 ± 10.68, with significant differences between the different treatments and the control (d. f.=9, F=7.392, P<0.01). After 7 days of treatment, phenylacetaldehyde and 1-phenylethanol were attractive to E. onukii at a concentration of 100 µl/ml, with insect populations of 175.67 ± 16.17 and 173 ± 5.86, respectively, there was no differrence in the attraction effect of the different treatments on E. onukii (d. f.=9, F=2.215, P>0.05). After 10 days of treatment, 1-phenylethanol was the best attractive effects of E. onukii , the number of E. onukii trapped by 1-phenylethanol at a concentration of 100, 10 and 0.1µl/ml were 200 ± 7.02, 180 ± 24.58 and 184 ± 6.56, respectively, there was significant differrence in the attraction effect of the different treatments on E. onukii (d. f.=9, F=2.42, P=0.048) (Figure 6a). On the Y-tube olfactometer tested, the E.onukii adults were more attracted to the volatiles including phenylacetaldehyde at a concentration of 10 µl/ml(χ2=7.251, d. f.=1, P<0.01), acetophenoneat at a concentration of 100 µl/ml(χ2=10.221, d. f.=1, P<0.01) , and 1-phenylethanol at a concentration of 100µl/ml (χ2=14.583, d. f.=1, P<0.01), there was a highly significant difference between these treatments and the control of attraction effect on E.onukii. 1-phenylethanol at a concentration of 0.1 µl/ml (χ2=0.724, d. f.=1, P>0.05), and acetophenoneat a concentration of 10µl/ml (χ2=0.724, d. f.=1, P>0.05) have a low attractive effects on E.onukii, there was no difference between these treatments and the control of attraction effect on E.onukii(Figure 6b).
2.5. RNA synthesis and analysis of interference efficiency
dsEonuIR25a was synthesized, and its accuracy was verified by 1% agarose gel electrophoresis, the product concentrations were tested to be above 2500ng/µl, meeting the criteria for dsRNA to be used in subsequent experiments (Figure 7a). RNA interference in E. onukii was performed by feeding tea seedlings after root soaking and foliar spraying, and the interference efficiency of RNAi was measured by qRT-PCR to detect the expression of the EonuIR25a gene of E. onukii after different lengths of time. The qRT-PCR results showed that expression of EonuIR25a significantly decreased after 3 hours of feeding on foliar spray treatment, and the gene expression decreased to 0.24 ± 0.01at 3h and 0.80 ± 0.35 at 48h.The expression decreased by 75.67%, 48.04%, 41.07% and 19.93% at 3, 12, 24 and 48 h after the foliar spray treatment, respectively ,there was significant difference in expression between different duration of RNAi treatments and control (P=0.03).The gene expression of EonuIR25a by root soaking was 0.24 ± 0.00 for 3 h and 0.76 ± 0.02 for 48 h.The expression of EonuIR25a decreased by 75.63%, 69.46%, 56.27% and 24.21% at four time points after root soaking treatment, respectively, there was significant difference in expression between different duration of RNAi treatments and control(P=0.03)(Figure 7b).
2.6. Changes in the response of E. onukii to compounds after silencing EonuIR25a
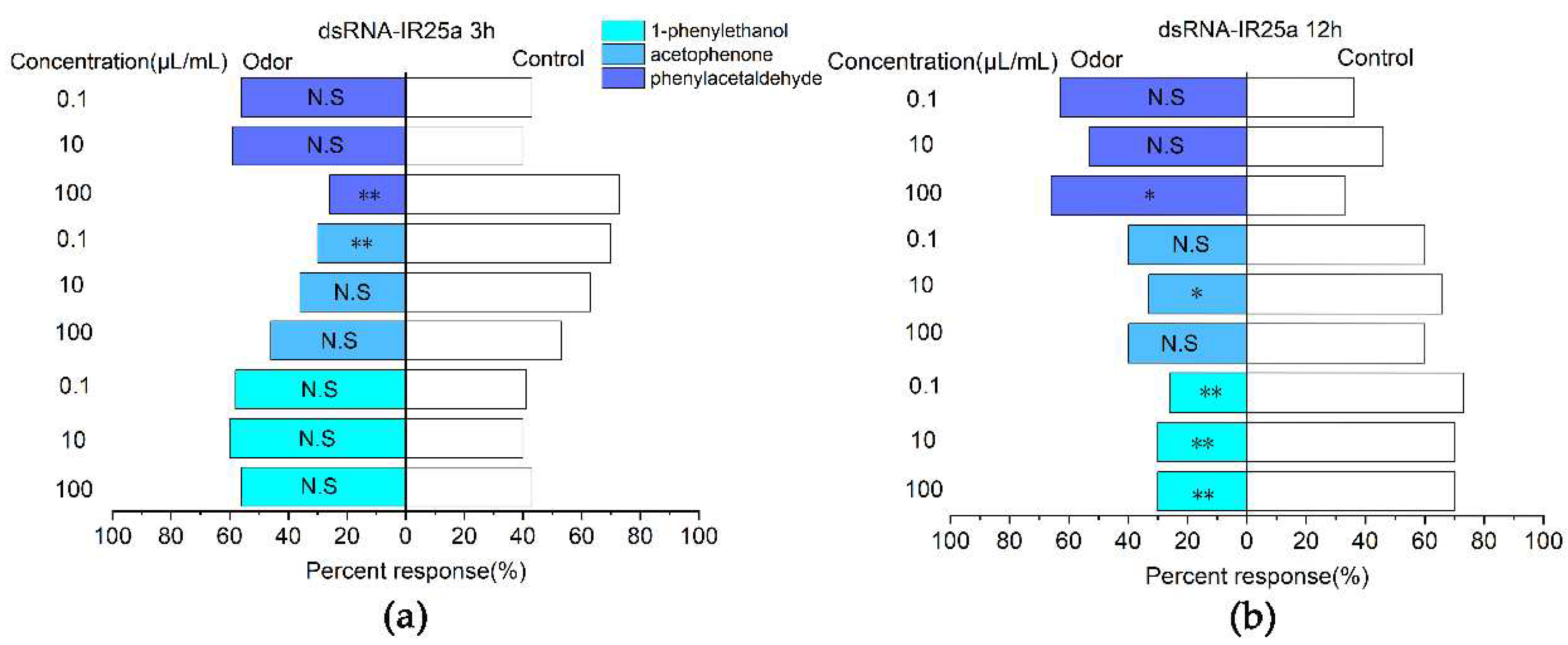
The analysis of the efficiency of RNA interference showed that the E. onukii fed tea seedlings treated by the root sock were more efficient in RNAi efficiency. Therefore, E. onukii was silenced with the EonuIR25a gene for 3 and 12 h, respectively, and was selected for Y-type olfactometer experiments. For 3 h, the responses to phenylethylaldehyde at a concentration of 100 µl/ml (χ2=10.221, d. f.=1, P<0.01) and acetophenone at a concentration of 0.1µl/ml (χ2=7.251, d. f.=1, P<0.01) were significantly downregulated, which was significantly different from the control compared to the non-RNAi E. onukii (Figure 8a). For 12 h, the response rates of E. onukii to 1-phenylethanol at concentrations of 0.1(χ2=10.221, d. f.=1, P<0.01), 10(χ2=0.08, d. f.=1, P>0.05) and 100 µl/ml(χ2=7.251, d. f.=1, P<0.01) were significantly reduced, and there were significant differences between the treatments and the control. There were no significant differences between phenylethylaldehyde at concentrations of 0.1 (χ2=2.93, d. f.=1, P>0.05), 10 µl/ml(χ2=1.636, d. f.=1, P>0.05), acetophenone at concentrations of 100 (χ2=1.636, d. f.=1, P>0.05) , 0.1 µl/ml (χ2=1.636, d. f.=1, P>0.05) and the control (Figure 8b).
3. Discussion
In this study, we cloned the gene encoding the EonuIR25a protein in E. onukii. The CDS length of EnouIR25a was 1266 bp and it encoded a 48.87 kD protein.After predictive analysis, EonuIR25a was found to have four transmembrane domains, mainly consisting of α-helices and irregularly coiled-winding folding, the sequence was found to contain the conserved structural domains of Lig_chan (positions 41-350) and SBP_bac_3 (positions 1-317), the Lig_chan family includes four transmembrane regions of ionotropic glutamate receptors and NMAD receptors, consistent with the typical structure of ionotropic receptors[32,33]. We found that there was similarity between EonuIR25a and IR sequences of some hemiptera, which sequence similarity ranging from 30-40%. EonuIR25a is closely related to HvitIR25a of H. vitripennis with a sequence similarity of 36.52%, and the sequence similarity between Drosophila IRs has been reported to be 10%-70%, implying that IRs are functionally diverse in insects[34]. Evolutionary tree analysis revealed that EonuIR25a clustered into the same branch with HvitIR25a and HvitIR25a-like, indicating that IR25a is highly conserved in different insects and presumably plays similar functions in insects[35]. This study showed that EonuIR25a was mainly distributed in the antennae and head, with a small amount of expression in the thorax, abdomen and legs, and it is speculated that EonuIR25a may belong to the olfactory IRs[22]. In addition, studies on Anopheles gambiae[28], Schistocerca gregaria[36] and Mythimna separata [26] also demonstrated that olfactory IRs were mainly expressed in the neurons of the tentacle cone sensor. In addition, previous studies have shown that IRs are widely expressed in organ tissues of insects, mainly in the antennae, labellum, anterior wing margin and legs, which is consistent with the results of this study[22,37] . Previous studies have shown that Drosophila IRs can sense odor substances, including acetic acid, propionic acid, butyric acid, hexanoic acid, 2-oxovaleric acid and phenylacetic acid[12,38,39,40]. It was found that Drosophila IR8a and IR64a are expressed in adult antennal vesicle neurons to perceive acidic tastes[41]. In Drosophila, Ir25a and Ir76b are necessary for odor-evoked electrophysiological responses to amines[42]. From the tissue distribution, it was speculated that EonuIR25a may be involved in the perception of odorant substances by leafhoppers.
Plant volatiles help herbivorous insects locate hosts, and therefore, they could be used to help develop pesticide-free pest management strategies[43]. Compared with phenylacetaldehyde and acetophenone, the results of field and indoor tests showed that 1-phenylethanol had more attractive effects on E. onukii, indicating that alcohols have an effect on the behavioral choices of insects. Research has indicated that a mixture of 1-phenylethanol has a significant attractive effect on 1st instar larvae of Helicoverpa armigera[44]. In addition, research has shown that alcohols can attract insects such as Dendroctonus frontalis [45], Helicoverpa assulta[46], and Dasychira baibarana [47], which is similar to the results of this study.
The results of Y-tube olfactometer showed that the response rate of E. onukii 1-phenylethanol at concentration of 100µl/ml was 56.67% after interfering with EonuIR25a for 3 h, which was not significantly different from the control. After 12h of interference with EonuIR25a, the response rate to 1-phenylethanol at concentration of 0.1, 10 and 100µl/ decreased to 26.67%, 30% and 30%, respectively. The results of RNAi showed that the tendency of behavioral selection of E. onukii to 1-phenylethanol was reduced after interference with EonuIR25a, and it was speculated that EonuIR25a could function in the recognition of tea plant volatile 1-phenylethanol by E. onukii. The current functions for ionotropic receptor recognition of olfaction are less available and are mainly concentrated in model-living Drosophila. The ionotropic receptors of Drosophila can recognize alcohols such as n-propyl alcohol and ethyl alcohol[12]. In Agrotis segetum, AsegIR75p.1 elicits electrophysiological signaling responses for alcohol ligand C6 unsaturated compounds[48], which is consistent with the results of the present research.
In recent years, RNAi has attracted much interest as a pest control tool[49], and the stability of dsRNA and the ultimate effectiveness and specificity of RNAi are worth considering[50]. Currently, the most commonly applied dsRNA delivery methods in insects are microinjection and feeding[51,52,53,54]. It is noteworthy that most microinjections are time-consuming and require a variety of equipment from in-house manufactured devices to complex microprocessor-controlled syringes, and microinjections are obviously limited to the laboratory[52]. In this study, the feeding method was chosen for RNA interference due to the small size of E. onukii and the high mechanical damage and mortality of microinjection found in the preexperiment. E. onukii was found to exhibit olfactory defects in the recognition of 1-phenylethanol by feeding dsRNA, suggesting that this method could to some extent interfere with the target gene in E. onukii. The interference efficiency of root soaking was significantly more effective and stable than foliar spraying . It is assumed that dsRNA is absorbed into the vascular bundles of the plant through the roots and stays for a longer time, thus producing a stable interference effect.Meanwhile, considering the leaf area of tea seedlings, the dose of dsRNA and the number of sprays also had an effect on the inhibitory effect of target genes, and our study tentatively verified that feeding method was effective in interfering with E. onukii[55].
The method of feeding used in our study has the advantages of dsRNA being formulated into a sprayable form for application to large crops, thereby selectively targeting insects as they feed. In addition, feeding of dsRNA poses a moderate risk of resistance to gene silencing, and it can be targeted to appropriate transcripts with high fidelity without affecting nontarget species[53].In particular, it should be noted that dsRNA is effective against insects that suck or chew on their mouthparts[56,57,58]. Therefore, feeding of dsRNA is not laborious, is easy to perform and is applicable to insect pest management[59].
Olfactory signaling is a target for pest management, with attempts to interfere with and thus disrupt odorant- or pheromone-driven behaviors. Previous studies have shown that insect ionotropic receptors are involved in the olfactory recognition process[60,61]. The functions of a few IRs in insects have been identified[60,61,62,63], but the molecular mechanisms and functions of IRs in most insects, especially in nonmodel insects, have not been reported, therefore, there is an urgent need for functional studies of IRs in nonmodel insects. In summary, the field results indicate that 1-phenylethanol has an attractive effect on E. onukii, suggesting that the compound could produce behavioral modulation of E. onukii. The recognition ability of 1-phenylethanol in E. onukii was reduced after interference of EonuIR25a by RNAi, which indicates that EonuIR25a functions in the recognition of 1-phenylethanol by E. onukii, and it also reflects that there is certain plasticity in the olfactory mechanism of insects. In addition, the findings demonstrate that RNAi could be applied as an important tool for gene function verification. IRs could participate in the olfactory recognition process of insects. In the future, insect semiochemicals could be developed for pest management and control by using ionotropic receptors in combination with tea plant volatiles.
4. Materials and methods
4.1. Insect culture
Adults of E. onukii were collected from an organic tea garden at Shandong Taishan Chaxi Valley Agricultural Development Co., Ltd, Shandong Province, China, in 2022. The colonies were kept and maintained on fresh tea seedlings in an artificial climate chamber in the laboratory of Shandong Agricultural University. The colony was maintained at 25 ± 1 °C, 60 ± 5% relative humidity, and a 16 h:8 h (L:D) photoperiod.
4.2. Total RNA isolation and RT-PCR
Total RNA was isolated using the FastPure® Cell/Tissue Total RNA Isolation Kit (Vazyme, Nanjing, China) from E. onukii adults. RNA samples were prepared and stored at -70 °C. RNA concentration and purity were assessed spectrophotometrically by measuring their absorbances at 260 and 280 nm in a biophotometer (Eppendorf, Germany). Gene-specific primers were designed to clone the ORF of the EnouIR25a gene, and cDNAs were synthesized from 2 lg of female antennal RNA using the MonScriptTm RTIIl All-in-One Mix (Mona, Wuhan, China). PCRs were conducted using Phanta Max Super-Fidelity DNA Polymerase (Vazyme, Nanjing, China) under the following conditions: denaturation at 95 °C for 3 min, 35 cycles of denaturation at 95 °C for 15 s, annealing at 55 °C for 55 s and extension at 72 °C for 1 min. The final extension step was at 72 °C for 5 min.
4.3. Cloning and nucleotide sequencing
The PCR products were then purified using the FastPure EndoFree Plasmid Mini Kit (Vazyme, Nanjing, China) and ligated into a pET-30a vector (Abiowell, Changsha, China) according to the manufacturer’s instructions. Afterward, plasmids were transformed into competent Escherichia coli DH5α competent cells, 100 μL of nonresistant LB solution was added, and the cells were shaken for 2 h in a 37 °C incubator and coated. After incubation in an incubator at 37 °C for 12 h, 20 positive clones of single bacteria were selected and incubated overnight at 37 °C in LB liquid medium containing 100 mg/mL Kana for PCR identification of the bacterial solution. The bacterial solution was identified as a 10 μL PCR system: 2×Taq Master Mix 5 μL, 10 μmol/L forward and reverse primers 0.4 μL each, ddH2O 3.2 μL, and bacterial solution 1 μL. Reaction conditions: denaturation at 95 °C for 3 min, 35 cycles of denaturation at 95 °C for 15 s, annealing at 55 °C for 15 s and extension at 72 °C for 1 min. The final extension step was at 72 °C for 5 min. Several of these positive colonies were then purified using the FastPure® Plasmid Mini Kit (Vazyme, Nanjing, China) and sent to Sangon Biotech (Shanghai) Co., Ltd. (China) for sequencing.
4.4. Bioinformatics analysis
The putative signal peptide was predicted using SignalP 3.0 Server (https://services.healthtech.dtu.dk/service.php?SignalP-5.0). A calculated molecular weight, theoretical isoelectric point, and grand average of hydropathicity were predicted using the ProtParm Tool (https://web.expasy.org/protparam/). The potential protein subcellular localization and transmembrane domains were predicted using WoLF PSORT (https://wolfpsort.hgc.jp/) and TMHMM Server (version 2.0) (https://services.healthtech.dtu.dk/service.php?TMHMM-2.0), respectively. Sopma (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.plpage=npsa_sopma.html) and Swiss-Model (https://swissmodel.expasy.org/interactive) were used to predict the secondary and tertiary structures, respectively. Server (http://www.cbs.dtu.dk/services/NetPhos/) was used to predict the protein phosphorylation sites; Protscale (https://web.expasy.org/protscale/) was used for protein hydrophilicity or hydrophobicity analysis; three-dimensional structural modeling of the EonuIR25a protein was built by the homology-based method using the SWISS-MODEL program (https://swissmodel.expasy.org/).
4.5. Phylogenetic analysis
To explore the similarity of candidate IR25a sequences from different insect orders, a Hemiptera-based phylogenetic analysis was performed, including the EnouIR25a protein sequences and orthologs in other species. Sequence data were analyzed using DNAMAN (version 5.2) and the BLAST program (http://blast.ncbi.nlm.nih.gov/Blast.cgi). The neighbor-joining tree was constructed using the MEGA 5.0 program[64] with a p-distance model and pairwise deletion gaps. Bootstrapping was performed by resampling the amino acid positions of 1000 replicates.
4.5. Expression in Several Tissues Using qRT-PCR
The relative transcript abundance of the EnouIR25a gene in the female antennae, male antennae and body tissues (heads, thoraxes, abdomens, legs) was determined by qRT-PCR. The reference gene b-actin was also used for normalization. The expression of EnouIR25a in several tissues was estimated by qRT-PCR using an ABI Prism 7500 Fast Detection System and SYBR Green SuperReal PreMix Plus (TianGen, Beijing, China). Each reaction was performed in a 20 μL final volume containing 10 μL 2 × ChamQ SYBR qPCR Master Mix, 0.4 μL each 10 μmol/L forward and reverse primers, 0.4 μL 50 × ROX Reference Dye 1, 1 μL template cDNA, and 7.8 μL ddH2O. The reactions were performed under the following conditions: predenaturation for 30 s at 95 °C, followed by 40 cycles of 10 s at 95 °C and annealing at 60 °C for 30 s, with a melting curve at 95 °C for 15 s, as instructed by the manufacturer. Each qRT-PCR experiment was performed with three biological replicates, and each biological replicate was assessed three times. The comparative 2-ΔΔCt method[63] was used to calculate the relative transcript levels in each tissue.
4.6. Field experiment
The experiment was conducted in September 2022 in the organic tea garden of Taishan Chaxi Valley Agricultural Co. The response of adult E. onukii to different doses of 3 volatile compounds was measured in the field using a combination of sticky boards and trap cores. The 3 volatile compounds were as follows: acetophenone (99%, Macklin, Shanghai, China), phenylacetaldehyde (95%, Macklin), and 1-phenylethanol (98%, Macklin). Solutions of each compound were prepared in paraffin at three concentrations (0.1, 10, and 100 µl/ml). A total of 750 µL of the volatile organic compound was added dropwise to a clean slow-release carrier, sealed in a bag, then placed in a refrigerator at 4 °C, and used after the volatile organic compound was completely adsorbed by the slow-release carrier. For the field experiment, one trap core was fixed in the center of the sticky cards using wire as the treatment, with three replicates for each treatment, and paraffin was used as a blank control. The sticky cards were placed on tea trees in the ‘Fuding Dabai’ variety park, and the treatments and replicates were distributed in a tessellated manner. The sticky cards were settled with an interval of 3 m. The numbers of E. onukii in each treatment were counted after 3, 7 and 10 d of the experiment.
4.7. Olfactometer Bioassays
The responses of E. onukii to tea plant volatiles were tested in a Y-tube olfactometer. Females and males of mixed ages used for olfactometer bioassays were collected from the artificial incubator, which contained recently emerged E. onukii adults. The glass Y-tube olfactometer consisted of a 60-mm-long base tube and two 60-mm-long arms separated from each other at an angle of 90 °C. The inner diameter of the base tube and arms was 10 mm. Air was pumped into the apparatus by an electromagnetic air pump (ACO serial, Sunsun Group Co., LTD, China), filtered through activated charcoal and split into two air streams, each of which was fed through a glass flask and into one arm of the olfactometer at a speed of 60 ml/s controlled by a float-type flowmeter (LZB-3 WB, Changzhou Shuanghuan Thermo Technical Instrument Co., LTD, China). The two glass flasks (100 ml) provided test and control odor sources. The connections between the components of the olfactometer were made of Teflon tubing. Olfactometer experiments were carried out in a dark room at 25 ± 2 °C and RH 60 ± 5 %, and the observation time was 17:00 h-21:00 h. Test E. onukii were transferred individually to the base tube of the Y-tube and observed separately for 5 min. The choice of E. onukii for one of the two odor sources was recorded when it crossed a half-length of either arm within 5 min. If the tested E. onukii did not cross half the length of either arm after 5 min, it was recorded as ‘No choice’. After five E. onukii were tested, the odor sources entering the arms of the Y-tube were swapped to avoid directional effects due to lighting in the apparatus. Before each test, the apparatus was cleaned by rinsing with pure ethanol and dried in an oven (120 °C). The responses of adult E. onukii to acetophenone, phenylacetaldehyde, and 1-phenylethanol at different doses were measured using a Y-tube olfactometer. Solutions of each compound were prepared in paraffin at three concentrations (0.1, 10, and 100 µl/ml). One hundred microliters of the test solution was applied to a piece of filter paper (1 × 1 cm), which was placed into the treatment flask. Filter paper with 100 µl paraffin in the treatment flask was used as a control. Olfactometer bioassays were conducted as detailed above. The filter papers with the test solution and paraffin were changed every 20 min. Each odor comparison was repeated thirty times with one E. onukii each time.
4.8. dsRNA Synthesis
Using the E. onukii cDNA template and specific primers containing the T7 promoter sequence at their 5’ ends (Table 1), regions of the E. onukii gene were amplified by qRT-PCR. The profile used in the reactions included denaturation at 95 °C for 3 min, 35 cycles of denaturation at 95 °C for 15 s, annealing at 55 °C for 55 s and extension at 72 °C for 1 min. The final extension step was at 72 °C for 5 min. PCR products were detected by 1% agarose gel electrophoresis and recovered using the Easy Pure Quick Gel Extraction Kit as a synthetic dsRNA template. Purified amplicons were transcribed in vitro to synthesize dsRNA using a T7 RNAi Transcription Kit (Vazyme, Nanjing, China). The integrity and quantity of dsRNAs were evaluated by spectroscopy analysis with a NanoDrop™ OneC Microvolume UV-Vis Spectrophotometer (Thermo Fisher Scientific) and by 1% agarose gel electrophoresis, and the purified products were stored at -80 °C.
4.9. Feeding method to interfere with EnouIR25a in E. onukii
The prepared RNAi interference products were thawed, and the concentration was adjusted to 1000 ng/μl. Then, 400 μl of interference product was aspirated into a 1.5 ml enzyme-free centrifuge tube. The roots of an annual one-bud, three-leaf ‘Longjing 43’ tea seedling were cleaned with sterile water. The tea seedlings were air-dried until the root surface was free of droplet-like liquid and then immersed in enzyme-free centrifuge tubes for subsequent root sock treatment. The prepared RNAi interference fragment products were thawed, and the concentration was adjusted to 1000 ng/μl. One annual one-bud-three-leaf ‘Longjing 43’ tea seedling using sterile water was immersed in an enzyme-free centrifuge tube. dsEonuIR25a was sprayed onto tea seedlings from four directions (approximately 400 μl) with a sprayer (2 ml) for subsequent foliar spray treatment. The tea seedlings were covered with 30*200 mm flat glass test tubes after root soaking and spray treatment. The mouth of the tube was sealed with a sealing film, and the sealing film was punctured with a 2 mm diameter hole by forceps. The test tubes were placed vertically upside down in a plant incubator, which was set to a temperature range of 25-28 °C, humidity of 45-50% rh, light intensity of 4000 LX, and photoperiod of 12 h. The samples were taken after 3 h, 12 h, 24 h and 48 h of feeding on tea seedlings of E. onukii. Each of the 10 E. onukii were used as one sample, and the experiment had three biological replicates, which were collected and frozen in liquid nitrogen for use.
4.10. Data Statistics and Analysis
All statistical analyses were conducted with IBM-SPSS statistical software (v.18.0; IBM, Armonk, NY, USA). All real-time qPCR assays were performed using three biological replicates. The data were analyzed using the threshold cycle number (CT) and the 2−∆∆Ct method, and significant differences in the expression patterns of the EnouIR25a gene in different tissues were analyzed using ANOVA followed by Duncan’s test. Significance was determined as p<0.05. The indoor behavioral responses of the adult E. onukii in the olfactometer were analyzed using a χ2 test, and significance was determined as *P<0.05; **P<0.01. The mean numbers of E. onukii in the field experiment were compared by Duncan's test in ANOVA, and significance was determined as p<0.05. Images were drawn by Origin64 software.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table 1 and Table 2 in the article have been added to the supplementary material.
Author Contributions
All authors conceived the ideas and designed methodology;contributed to the manuscripts and final submission.XYL and YZ conceived research, performed the experiment and analyzed the data. YHZ and RRZ performed the writing-original draft preparation. ZQZ and XXX secured funding and critically revised the manuscript. All authors read and approved the final manuscript. All authors read and approved the final manuscript.
Funding
This research received no external funding.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Acknowledgments
We acknowledge financial support from Natural Science Foundation of Shandong Province (ZR2020MC122;ZR2020QC132), National Natural Science Foundation of China(32102303), Project of Laoshan District Tea Innovation Group (LSCG2022000017), Key Research and Development Program of Shandong Province (2022LZGCQY020),Project for Technological Innovation and Development of Tai’an (2021NS082), Project for Special Commissioners in Agricultural Science and Technology of Tai’an (2021TPY021).
Conflicts of Interest
The authors declare no conflict of interests.
References
- da Silva Pinto, M. Tea: a new perspective on health benefits. Food. Res. Int. 2013, 53, 558-567. [CrossRef]
- Ye, G. Y.; Xiao, Q.; Chen, M.; Chen, X. X.; Yuan, Z. Y.; Stanley, D. W.; Hu, C. Tea: biological control of insect and mite pests in China. Biol. Control. 2014, 68, 73-91. [CrossRef]
- Xin, Z. J.; Li, X. W.; Bian, L.; Sun, X. L. Tea green leafhopper, Empoasca vitis, chooses suitable host plants by detecting the emission level of (3Z)-hexenyl acetate. B. Entomol. Res. 2017, 107, 77-84. [CrossRef]
- Silva, L. A brief history of biochemical genetics’ 50 years and a reflection about past and present research directions. Biochem. Genet. 2018, 56, 1-6. [CrossRef]
- Gadenne, C.; Barrozo, R. B.; Anton, S. Plasticity in insect olfaction: To smell or not to Smell? Annu. Rev. Entomol. 2016, 61, 317-333. [CrossRef]
- Wang, Z. H.; Yang, F.; Sun, A.; Song, J. Y .; Shan, S.; Zhang, Y. J.; Wang S. N. Expressional and functional comparisons of five clustered odorant binding proteins in the brown marmorated stink bug Halyomirpha halys. Int. J. Biol. Macromol. 2022, 209, 1352-1358. [CrossRef]
- Conchou, L.; Lucas, P.; Meslin, C.; Proffit, M.; Staudt, M.; Renou, M. Insect odorscapes: from plant volatiles to natural olfactory scenes. Front. Physiol. 2019, 10, 972. [CrossRef]
- Fleischer, J.; Pregitzer, P.; Breer, H.; Krieger. J. Access to the odor world: olfactory receptors and their role for signal transduction in insects. Cell. Mol. Life. Sci. 2018, 75, 485-508. [CrossRef]
- Robertson, H. M. Molecular evolution of the major arthropod chemoreceptor gene families. Annu. Rev. Entomol. 2019, 64, 227-242. [CrossRef]
- Leal, W. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373-391. [CrossRef]
- Pelosi, P .; Iovinella, I.; Zhu, J.; Wang, G. R.; Dani, F. R. Beyond chemoreception: Diverse tasks of soluble olfactory proteins in insects. Biol. Rev. 2018, 93, 184-200. [CrossRef]
- Benton, R.; Vannice, K. S; Gomez-Diaz, C.; Vosshall, L. B. Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila. Cell. 2009, 136, 149-162. [CrossRef]
- Mayer, M. L.; Armstrong, N. Structure and function of glutamate receptor ion channels. Annu. Rev. Physiol. 2004, 66, 161-181. [CrossRef]
- Armstrong, N.; Sun, Y.; Chen, G. Q.; Gouaux, E. Structure of a glutamate-receptor ligand-binding core in complex with kainate. Nature. 1998, 395, 913-917. [CrossRef]
- Kuner, T.; Seeburg, P. H.; Guy, H. R. A common architecture for K+ channels and ionotropic glutamate receptors?Trends. Neurosci. 2003, 26, 27-32. [CrossRef]
- Croset, V.; Rytz, R.; Cummins, S. F.; Budd, A.; Brawand, D.; Kaessmann, H.; Gibson, T. J.; Benton, R. Ancient protostome origin of chemosensory ionotropic glutamate receptors and the evolution of insect taste and olfaction. PLoS. Genet. 2010, 6, e1001064. [CrossRef]
- Chen, C. H.; Buhl, E.; Xu, M.; Croset, V.; Rees, J. S.; Lilley, K. S.; Benton, R.; Hodge, J. J. L.; Stanewsky, R. Drosophila Ionotropic Receptor 25a mediates circadian clock resetting by temperature.Nature. 2015, 527, 516-520. [CrossRef]
- Lee, Y.; Poudel, S.; Kim, Y.; Thakur, D.; Montell, C. Calcium taste avoidance in Drosophila. Neuron. 2018, 97, 67-74. [CrossRef]
- Steck, K.; Walker, S. J.; Itskov, P. M.; Baltazar, C.; Moreira, J. M.; Ribeiro, C. Internal amino acid state modulates yeast taste neurons to support protein homeostasis in Drosophila. Elife. 2018, 7, e31625. [CrossRef]
- Sánchez-Alcañiz, J. A.; Silbering, A. F.; Croset, V.; Zappia, G.; Sivasubramaniam, A. K.; Abuin, L.; Sahai, S. Y.; Münch, D.; Steck, K.; Auer, T. O.; Cruchet, S.; Neagu-Maier G. Larisa.; Sprecher, S. G.; Ribeiro, C.; Yapici, N.; Benton R. An expression atlas of variant ionotropic glutamate receptors identifies a molecular basis of carbonation sensing. Nat. Commun. 2018, 9(1), 4252. [CrossRef]
- Wang, S. N.; Peng, Y.; Lu, Z.Y.; Dhiloo, K. H.; Zheng, Y.; Shan, S.; Li, R. J.; Zhang, Y. J.; Guo, Y. Y. Cloning and expression profile of ionotropic receptors in the parasitoid wasp Microplitis mediator (Hymenoptera: Braconidae). J. Insect. Physiol. 2016, 90, 27-35. [CrossRef]
- Rimal, S.; Lee, Y. The multidimensional ionotropic receptors of Drosophila melanogaster. Insect. Mol. Biol. 2018, 27, 1-7. [CrossRef]
- Knecht, Z. A.; Silbering, A. F.; Ni, L.; Klein, M.; Budelli, G.; Abuin, L.; Ferrer, A. J.; Samuel, A. D. T. Distinct combinations of variant ionotropic glutamate receptors mediate thermosensation and hygrosensation in Drosophila. Elife. 2016, 5, e17879. [CrossRef]
- Raji, J. I.; Melo, N.; Castillo, J. S.; Gonzalez, S.; Saldana, V.; Stensmyr, M. C.; DeGennaro, M. Aedes aegypti mosquitoes detect acidic volatiles found in human odor using the IR8a pathway. Curr. Biol. 2019, 29, 1253-1262. [CrossRef]
- Liu, C.; Jason, Pitts, R., Bohbot, J. D., Jones, P. L., Wang, G., Zwiebel, L. J. Distinct olfactory signaling mechanisms in the malaria vector mosquito Anopheles gambiae. PLoS. Biol. 2010, 8, 27-28. [CrossRef]
- Tang, R.; Jiang, N. J.; Ning, C., Li, C.; Huang, L. Q.; Wang, C. Z. The olfactory reception of acetic acid and ionotropic receptors in the Oriental armyworm, Mythimna separata Walker. Insect. Biochem. Mol. Biol. 2020, 118, 103312. [CrossRef]
- Zhang, J.; Bisch-Knaden, S.; Fandino, R. A.; Yan, S.; Obiero, G. F.; Grosse-Wilde, E.; Hansson, B. S.; Knaden, M. The olfactory coreceptor IR8a coverns larval fecesmediated competition avoidance in a hawkmoth. PNAS. 2019, 116(43), 21828-21833. [CrossRef]
- Pitts, R. J.; Derryberry, S. L.; Zhang, Z. W.; Zwiebel, L. J. Variant ionotropic receptors in the malaria vector mosquito Anopheles gambiae tuned to amines and carboxvlic acids. Sci. Rep. 2017, 7, 40297. [CrossRef]
- Ahn, J. E.; Chen, Y.; Amrein, H. Molecular basis of fatty acid taste in Drosophila. Elife. 2017, 6, e30115. [CrossRef]
- Frank, D. D.; Enjin, A.; Jouandet, G. C.; Zaharieva, E. E.; Para, A., Stensmyr, M. C., Gallio, M. Early integration of temperature and humidity stimuli in the Drosophila brain. Curr. Biol. 2017, 27, 2381-2388. [CrossRef]
- Budelli, G., Ni, L., Berciu, C., van Giesen, L., Knecht, Z. A., Chang, E. C., Kaminski, B.; Silbering, A. F.; Samuel , A.; Klein, Mason.; Benton R.; Nicastro D.; Garrity P. A. Ionotropic receptors specify the morphogenesis of phasic sensors controlling rapid thermal preference in Drosophila. Neuron. 2019, 101, 738-747. [CrossRef]
- Ni, L. The structure and function of ionotropic receptors in Drosophila. Front. Mol. Neurosci. 2021, 13, 638839. [CrossRef]
- Rytz, R.; Croset, V.; Benton, R. Ionotropic receptors (IRs): chemosensory ionotropic glutamate receptors in Drosophila and beyond. Insect. Biochem. Molec. 2013, 43(9): 888-897. [CrossRef]
- Benton, R.; Vannice, K. S.; Gomez-Diaz C.; Vosshall, L. B. Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila. Cell, 2009, 136: 149-162. [CrossRef]
- Wang, T. T.; Hao, Y. J.; He, Z. B.; Met, T.; Chen, B. Characteristics and classification position of the ionotropic receptor genes IR8a and IR25a in four vector mosquito species of medical importance. Acta. Entomol. Sin. 2017, 60(4), 379-388. [CrossRef]
- Guo, M.; Krieger, J.; Grosse-Wilde, E.; Missbach, C.; Zhang, L.; Breer, H. Variant ionotropic receptors are expressed in olfactory sensory neurons of coeloconic sensilla on the antenna of the desert locust (Schistocerca gregaria). Int. J. Biol. Sci. 2014, 10(1), 1-14. [CrossRef]
- Ganguly, A.; Pang, L.; Duong, V. K.; Lee, A.; Schoniger, H.; Varady, E.; Dahanukar, A. A molecular and cellular context-dependent role for Ir76b in detection of amino acid taste. Cell. Rep. 2017, 18(3), 737-750. [CrossRef]
- Ai, M.; Min, S.; Grosjean, Y.; Leblanc, C.; Bell, R.; Benton, R.; Suh, G. S. B. Acid sensing by the Drosophila olfactory system. Nature. 2010, 468(7324), 691-695. [CrossRef]
- Min, S.; Ai, M.; Shin, S. A.; Suh, G. S. Dedicated olfactory neurons mediating attraction behavior to ammonia and amines in Drosophila. PNAS. 2013, 110(14), E1321-E1329. [CrossRef]
- Hussain, A.; Zhang, M.; Ucpunar, H. K.; Syensson, T.; Quillery, E.; Gompel, N.; Ignell, Rickard.; Grunwald Kadow, I. C. Ionotropic chemosensory receptors mediate the taste and smell of polyamines. PLoS. Biol. 2016, 14(5), e1002454. [CrossRef]
- Chen, Y.; Amrein, H. Ionotropic receptors mediate Drosophila oviposition preference through sour gustatory receptor neurons. Curr. Biol. 2017, 27(18), 2741-2750. [CrossRef]
- Ai, M.; Blais, S.; Park, J. Y.; Min, S.; Neubert, T. A.; Suh, G. S. Ionotropic glutamate receptors IR64a and IR8a form a functional odorant receptor complex in vivo in Drosophila. J. Neurosci. 2013, 33(26): 10741-10749. [CrossRef]
- Gregg, P. C.; Del Socorro, A. P.; Hawes, A. J.; Binns, M. R. Developing bisexual attract-and-kill for polyphagous insects: ecological rationale versus pragmatics. J. Chem. Ecol. 2016, 42, 666-675. [CrossRef]
- Di, C.; Ning, C.; Huang, L. Q.; Wang, C. Z. Design of larval chemical atractants based on odorant response spectra of odorantreceptors in the cotton bollworm. Insect . Biochem. Mol. Biol. 2017, 84, 48-62. [CrossRef]
- Shepherd, W. P.; Sullivan, B. T . Southern pine beetle, Dendroctonus frontalis, antennal and behavioral responses to nonhost leaf and bark volatiles. J. Chem. Ecol. 2013, 39(4), 481-493. [CrossRef]
- Wang, C.; Li, G. N.; Miao, C. J.; Zhao, M.; Wang, B.; Guo, X. R. Nonanal modulates oviposition preference in female Helicoverpa assulta (Lepidoptera: Noctuidae) via the activation of peripheral neurons. Pest. Manag. Sci. 2020, 76(9), 3159-3167. [CrossRef]
- Magsi, F. H.; Luo, Z. X.; Zhao, Y. J.; Li, Z. Q.; Cai, X. M.; Bian, L.; Chen Z. M. Electrophysiological and behavioral responses of Dasychira baibarana (Lepidoptera: Lymantriidae) to tea plant volatiles. Environ. Entomol. 2021, 50(3), 589-598. [CrossRef]
- Hou, X. Q.; Zhang, D. D.; Powell, D.; Wang, H. L.; Andersson, M. N.; Löfstedt, C. Ionotropic receptors in the turnip moth Agrotis segetum respond to repellent medium-chain fatty acids. BMC. Biol. 2022, 20(1), 1-19. [CrossRef]
- Schutter, K. D.; Taning, C. N. T .; Daele, L. V.; Van Damme, E. J. M.; Dubruel, P.; Smagghe, G. RNAi-based biocontrol products: market status, regulatory aspects, and risk assessment. Front. Insect. Sci. 2022, 1, 818037. [CrossRef]
- Christiaens, O.; Whyard, S.; Vélez, A. M.; Smagghe, G. Double-stranded RNA technology to control insect pests: current status and challenges. Front. Plant. Sci. 2020, 11, 451. [CrossRef]
- Souza, D.; Christensen, S. A.; Wu, K.; Buss, L.; Kleckner, K.; Darrisaw, C.; Shirk, P. D.; Siegfried, B. D. RNAi-induced knockdown of white gene in the southern green stink bug (Nezara viridula L.). Sci. Rep. 2022, 12(1), 10396. [CrossRef]
- Joga, M. R.; Zotti, M.; Smagghe, G.; Christiaens, O. RNAi efficiency, systemic properties, and novel delivery methods for pest insect control: what we know so far. Front. Physiol. 2016, 7, 553. [CrossRef]
- Kunte, N.; Mcgraw, E.; Bell, S.; Held, D.; Avila, L. Prospects, challenges and current status of RNAi through insect feeding. Pest. Manag. Sci. 2020, 76(1), 26-41. [CrossRef]
- Andrade, E. C.; Hunter, W. B. RNAi feeding bioassay: development of a non-transgenic approach to control Asian citrus psyllid and other hemipterans. Entomol. Exp. Appl. 2017, 162(3), 389-396. [CrossRef]
- Yan, S.; Qian, J.; Cai, C.; Ma, Z. Z.; Li, J. H.; Yin, M. Z.; Ren, B.; Shen, J. Spray method application of transdermal dsRNA delivery system for efficient gene silencing and pest control on soybean aphid Aphis glycines. J. Pest .Sci. 2020, 93: 449-459. [CrossRef]
- Li, H.; Bowling, A.J.; Gandra, P.; Rangasamy, M.; Pence, H. E.; McEwan, R. E.; Khajuria, C.; Siegfried, B. D.; Narva, K. E. Narva Systemic RNAi in western corn rootworm, Diabrotica virgifera virgifera, does not involve transitive pathways. Insect. Sci. 2018, 25, 45-56. [CrossRef]
- Khajuria, C.; Velez, A. M.; Rangasamy, M.; Wang, H. C.; Fishilevich, E.; Frey, M. L. Parental RNA interference of genes involved in embryonic development of the western corn rootworm, Diabrotica virgifera virgifera LeConte. Insect. Biochem. Mol. Biol. 2015, 63, 54-62. [CrossRef]
- Ramaseshadri, P.; Segers, G.; Flannagan, R.; Wiggins, E.; Clinton, W.; Ilagan, O.; McNulty, B.; Clark, T.; Bolognesi, R. Physiological and cellular responses caused by RNAi-mediated suppression of Snf7 orthologue in western corn rootworm (Diabrotica virgifera virgifera) larvae. PLoS. One. 2013, 8(1), e54270. [CrossRef]
- Zhu, K.Y.; Palli, S. R. Mechanisms, applications, and challenges of insect RNA interference. Annu. Rev. Entomol. 2020, 65, 293-311. [CrossRef]
- Cui, Y. Y.; Wang, J. G.; Liu, Q. Y.; Li, D. M.; Zhang, W.; Liu, X. B.; Wang, J.; Song, X. P.;·Yao, F.; Wu, H. X.; Zhao, N. Identification and expression of potential olfactory-related genes related to Niemann–Pick C2 protein and ionotropic receptors in Haemaphysalis longicornis. Exp. Appl. Acarol. 2022, 87(4), 337-350. [CrossRef]
- Zhang, Y.; Yang, B. Y.; Yu, J.; Pang, B. P.; Wang, G. R. Expression profiles and functional prediction of ionotropic receptors in Asian corn borer, Ostrinia furnacalis (Lepidoptera: Crambidae). J. Integr. Agr. 2022, 21(2), 474-485. [CrossRef]
- Wicher, D.; Miazzi, F. Functional properties of insect olfactory receptors: ionotropic receptors and odorant receptors. Cell. Tissue. Res. 2021, 383: 7-19. [CrossRef]
- 61. Yin, N. N.; Nuo, S. M.; Xiao, H. Y.; Zhao, Y. J.; Zhu, J.Y.; Liu, N. Y. The ionotropic receptor gene family in Lepidoptera and Trichoptera: Annotation, evolutionary and functional perspectives. Genomics. 2021, 113(1), 601-612. [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731-2739. [CrossRef]
- Livak, K. J.; Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods. 2001, 25(4), 402-408. [CrossRef]
Figure 1.
Identification of PCR products of EonuIR25a.
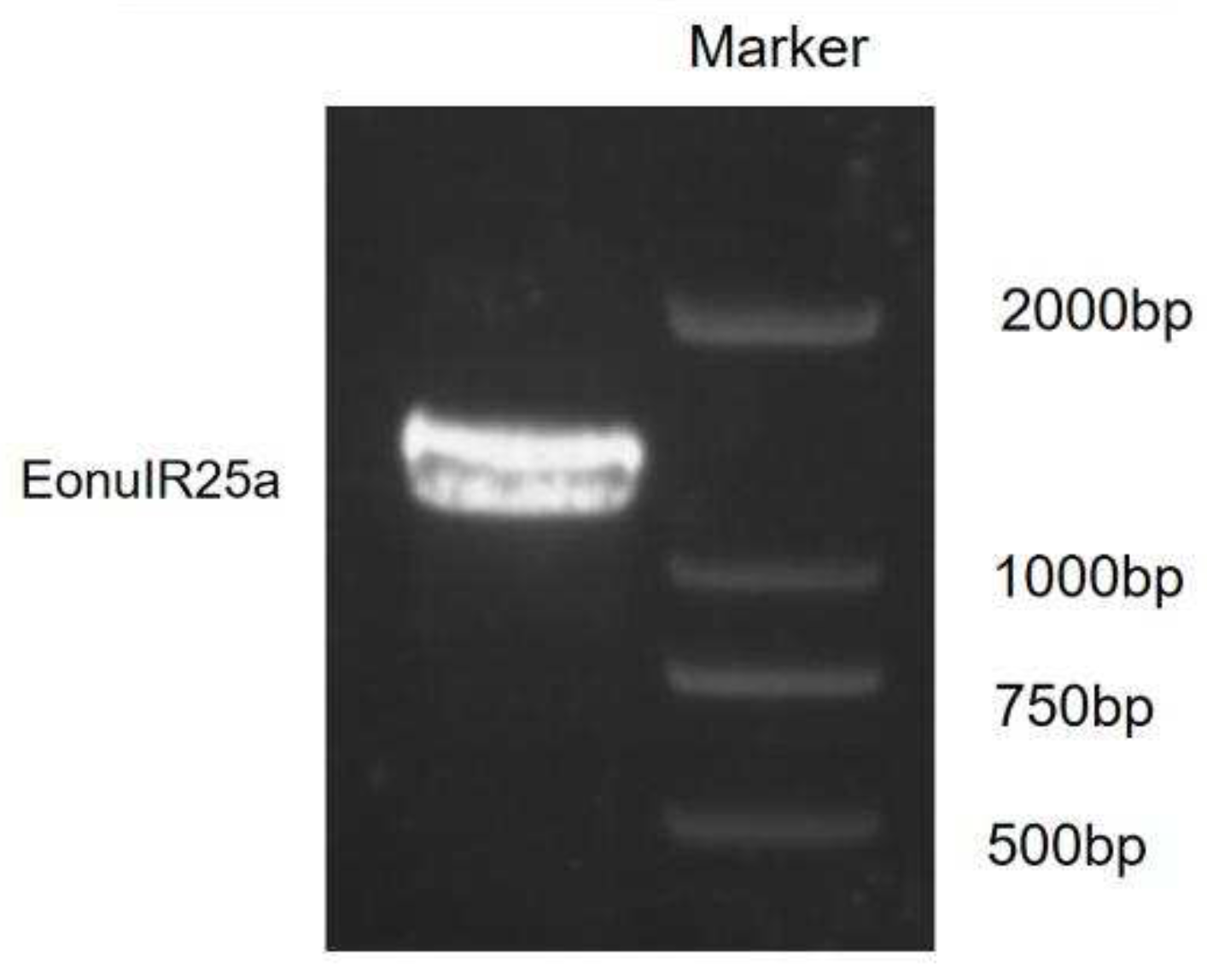
Figure 2.
Protein structure prediction of EonuIR25a in Empoasca onukii (Matsuda). (a): Hydrophilicity prediction of EonuIR25a protein. (b): Structure prediction of transmembrane proteins of EonuIR25a protein. (c): Phosphorylation site prediction of EonuIR25a protein; (d): Signal peptide prediction of EonuIR25a protein. (e): Secondary structure of IR25a protein in E.onukii. note: blue, red, green and purple masks indicate alpha helix, extended strand, beta turn and random coil, respectively. (f): Tertiary structure of IR25a in E.onukii. note: N and C indicate N-and C-termini of the protein.
Figure 2.
Protein structure prediction of EonuIR25a in Empoasca onukii (Matsuda). (a): Hydrophilicity prediction of EonuIR25a protein. (b): Structure prediction of transmembrane proteins of EonuIR25a protein. (c): Phosphorylation site prediction of EonuIR25a protein; (d): Signal peptide prediction of EonuIR25a protein. (e): Secondary structure of IR25a protein in E.onukii. note: blue, red, green and purple masks indicate alpha helix, extended strand, beta turn and random coil, respectively. (f): Tertiary structure of IR25a in E.onukii. note: N and C indicate N-and C-termini of the protein.
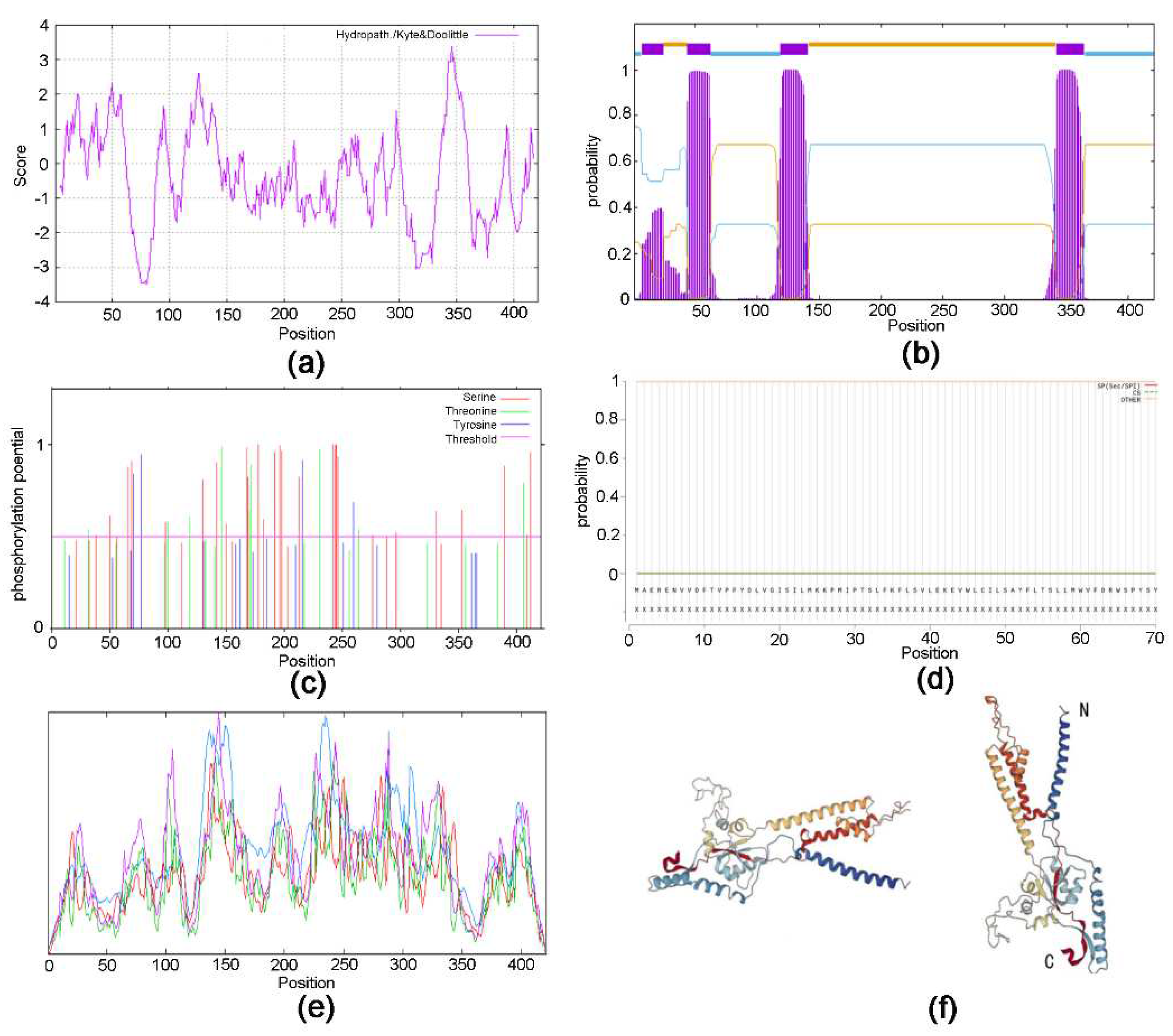
Figure 3.
Amino acid sequence alignment of EonuIR25a with other Hemiptera IRs. note:HvitIR25a, HvitIR25a-like, Homalodisca vitripennis (KAG8282323.1,XP_046658708.1); SyanIR25a, Subpsaltria yangi (AXY87920.1); HhalIR25a, Halyomorpha halys (XP_024219750.1); AlucIR25a, Apolygus lucorum (QFU27937.1); AlinIR25a, Adelphocoris lineolatus (APZ81419.1); ApisIR25a, Acyrthosiphon pisum (XP_008183092.2). Black areas indicate 100% similarity, pink areas indicate more than 75% similarity, blue areas indicate more than 50% similarity.
Figure 3.
Amino acid sequence alignment of EonuIR25a with other Hemiptera IRs. note:HvitIR25a, HvitIR25a-like, Homalodisca vitripennis (KAG8282323.1,XP_046658708.1); SyanIR25a, Subpsaltria yangi (AXY87920.1); HhalIR25a, Halyomorpha halys (XP_024219750.1); AlucIR25a, Apolygus lucorum (QFU27937.1); AlinIR25a, Adelphocoris lineolatus (APZ81419.1); ApisIR25a, Acyrthosiphon pisum (XP_008183092.2). Black areas indicate 100% similarity, pink areas indicate more than 75% similarity, blue areas indicate more than 50% similarity.
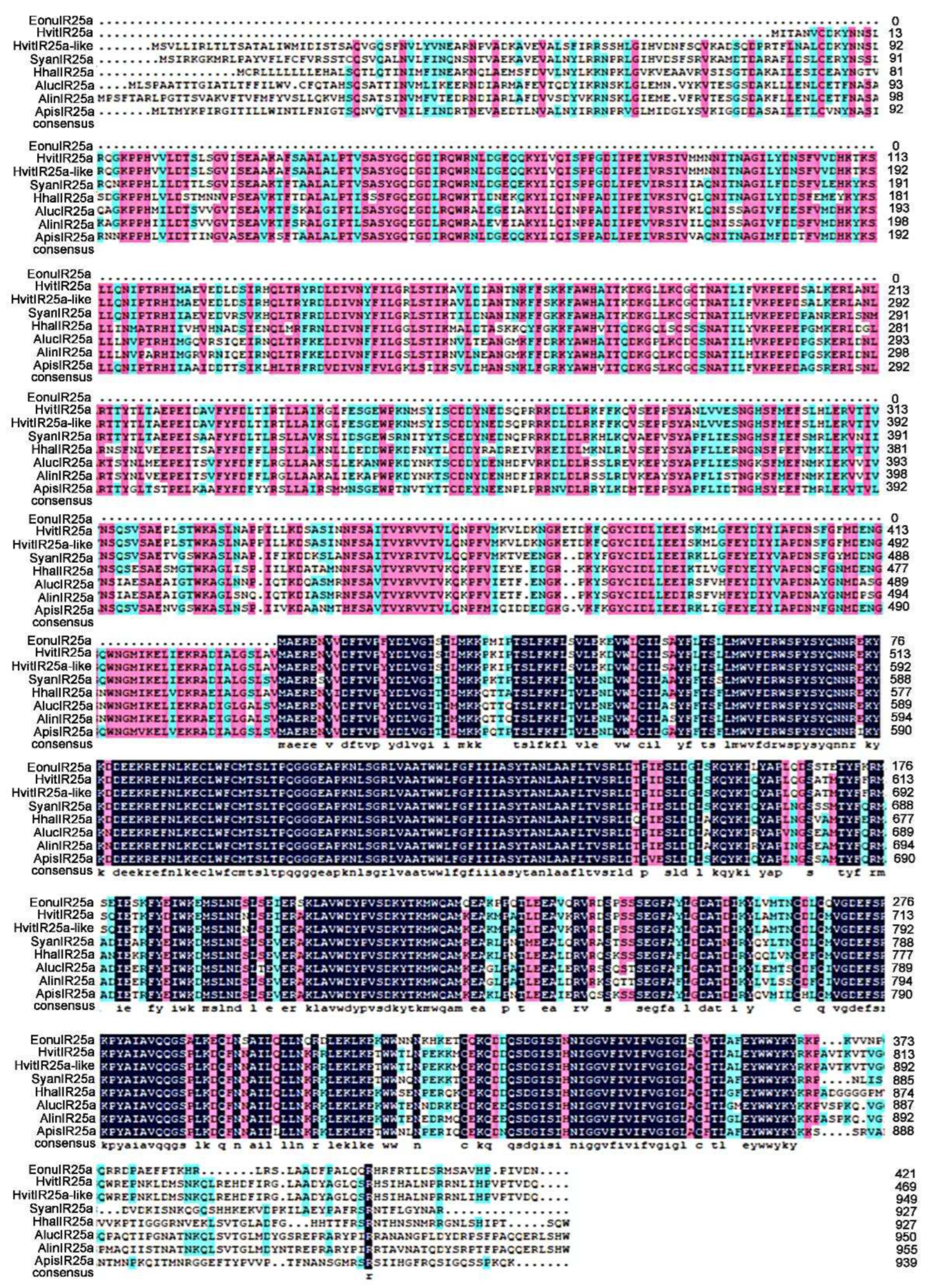
Figure 4.
Phylogenetic tree of EonuIR25a with other Hemiptera IRs based on amino acid sequences. note: different species in the evolutionary tree are indicated by different colors. Black, Empoasca onukii; Red, Halyomorpha halys; Purple, Subpsaltria yangi; Yellow, Homalodisca vitripennis; Brown, Apolygus lucorum; Aquamarine, Adelphocoris lineolatus; Blue, Bemisia tabaci; Blue black, Nilaparvata lugens; Navy blue, Aphis gossypii; Green, Acyrthosiphon pisum. The target gene is indicated with a pentagram☆.
Figure 4.
Phylogenetic tree of EonuIR25a with other Hemiptera IRs based on amino acid sequences. note: different species in the evolutionary tree are indicated by different colors. Black, Empoasca onukii; Red, Halyomorpha halys; Purple, Subpsaltria yangi; Yellow, Homalodisca vitripennis; Brown, Apolygus lucorum; Aquamarine, Adelphocoris lineolatus; Blue, Bemisia tabaci; Blue black, Nilaparvata lugens; Navy blue, Aphis gossypii; Green, Acyrthosiphon pisum. The target gene is indicated with a pentagram☆.
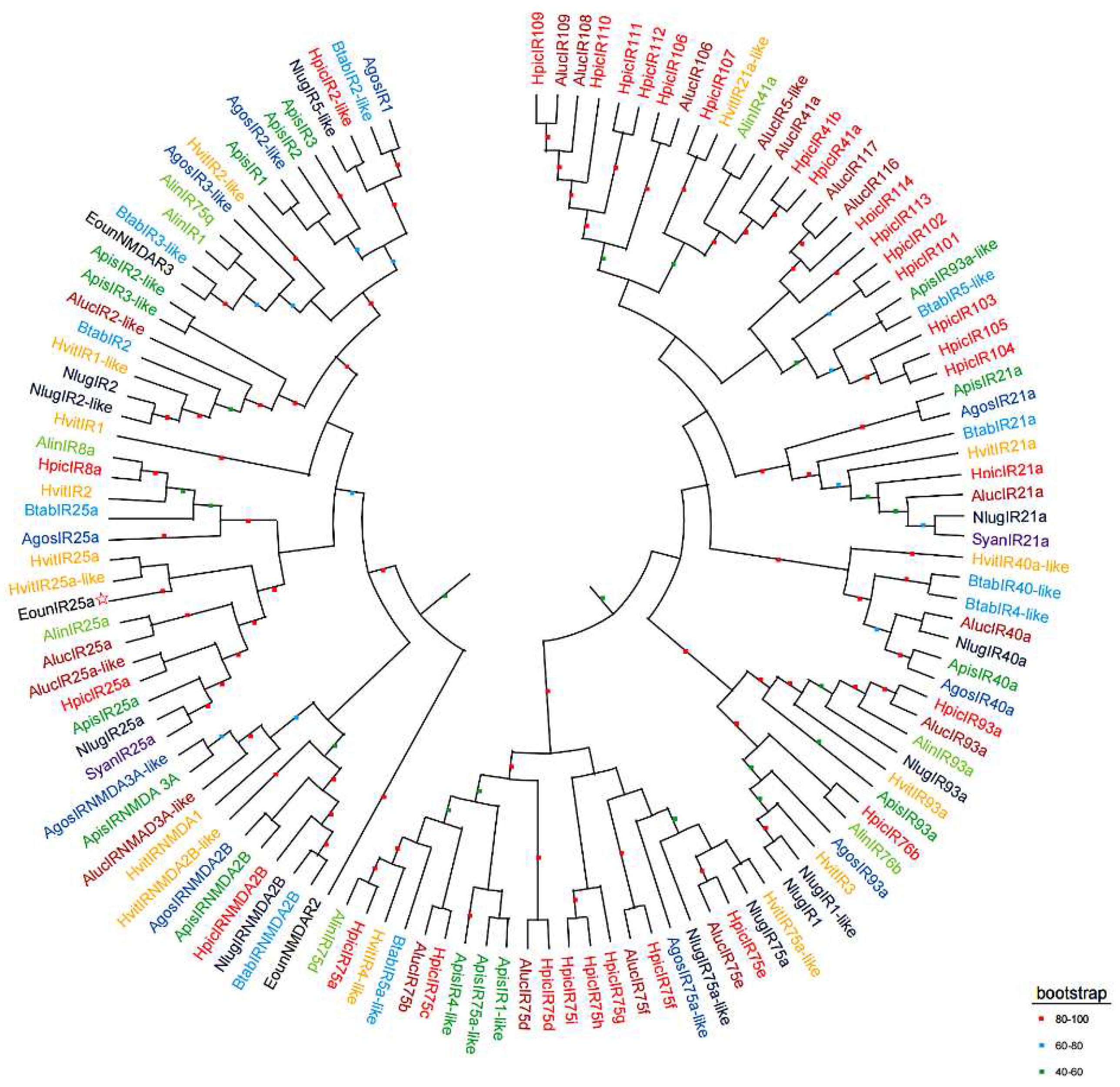
Figure 5.
Relative expression levels of EonuIR25a in different tissues of Empoasca onukii (Matsuda).
Figure 5.
Relative expression levels of EonuIR25a in different tissues of Empoasca onukii (Matsuda).
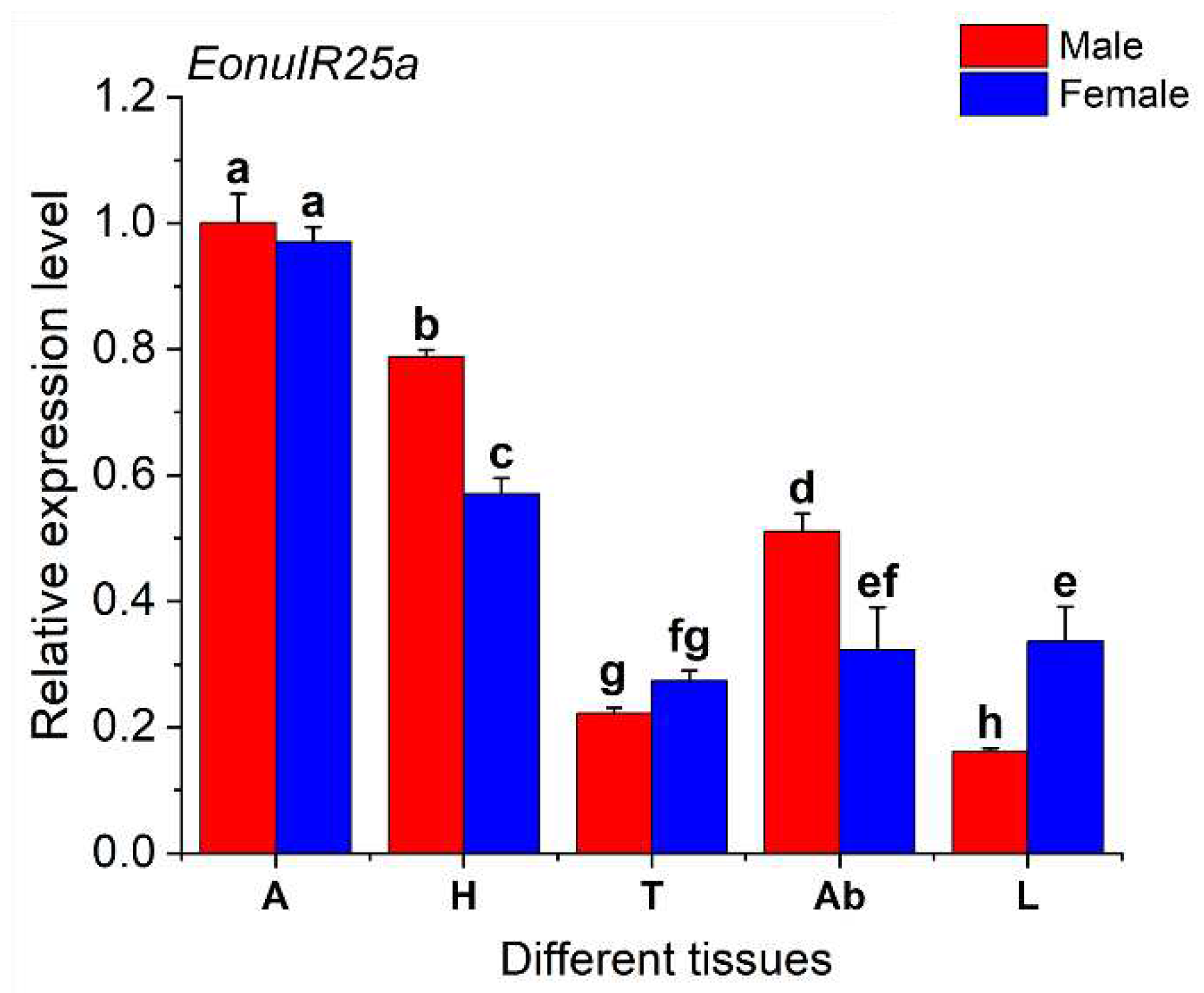
Figure 6.
Responses of E. onukii to different concentrations of compounds in field trials and olfactometer bioassays. (a): Response of E. onukii to compounds in the field. (b): Response of E. onukii to different concentrations of compounds with paraffin in a Y-tube olfactometer. note: asterisks indicate significant differences within a choice test (*P<0.05; **P<0.01); N. S indicates no significant difference.
Figure 6.
Responses of E. onukii to different concentrations of compounds in field trials and olfactometer bioassays. (a): Response of E. onukii to compounds in the field. (b): Response of E. onukii to different concentrations of compounds with paraffin in a Y-tube olfactometer. note: asterisks indicate significant differences within a choice test (*P<0.05; **P<0.01); N. S indicates no significant difference.
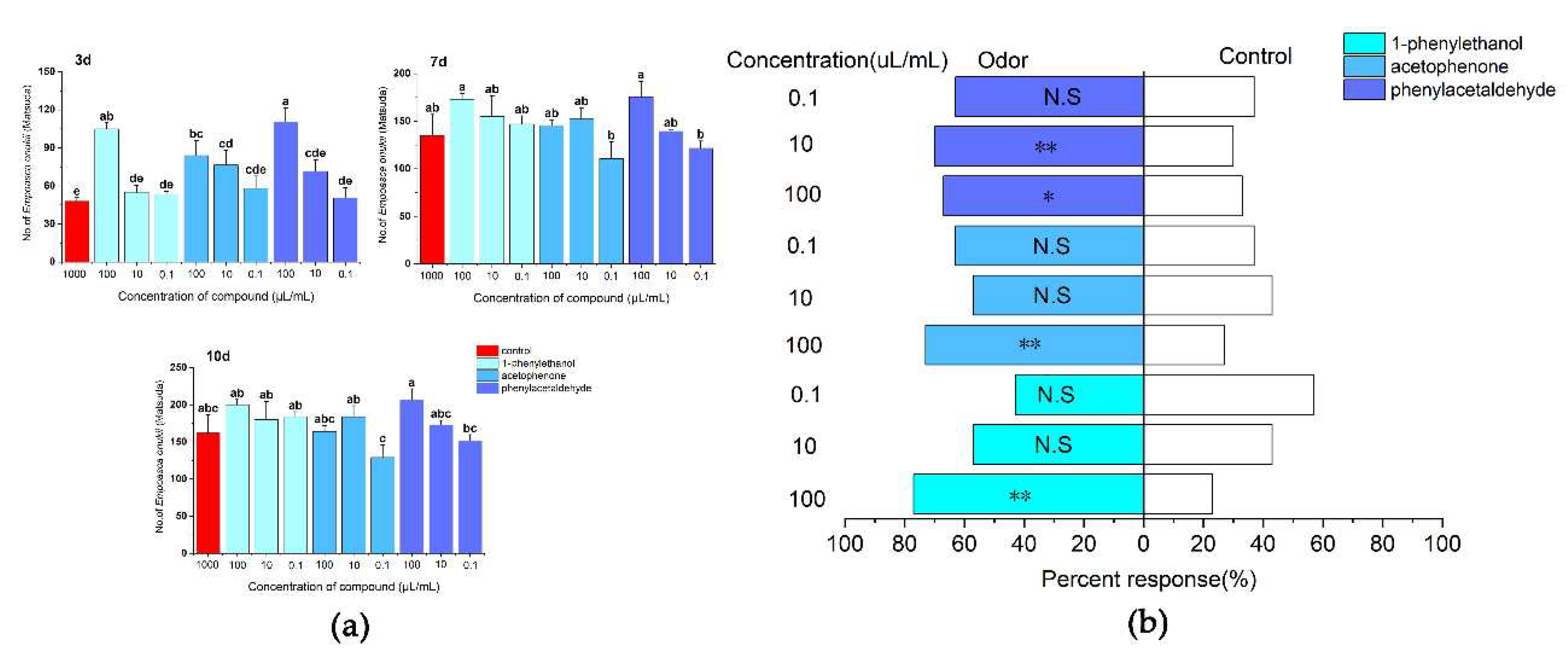
Figure 7.
EonuIR25a was interferenced by RNAi technology. (a): Identification of PCR products of dsEonuIR25a. (b): Relative expression of EonuIR25a was detected by root soak and foliar spray treatment for different time periods, respectively. note: data in the graph are mean ± SE. Different lowercase letters on the color bar indicate that the relative expression levels of EonuIR25a were significantly different at different time periods by Duncan's test (P<0.05).
Figure 7.
EonuIR25a was interferenced by RNAi technology. (a): Identification of PCR products of dsEonuIR25a. (b): Relative expression of EonuIR25a was detected by root soak and foliar spray treatment for different time periods, respectively. note: data in the graph are mean ± SE. Different lowercase letters on the color bar indicate that the relative expression levels of EonuIR25a were significantly different at different time periods by Duncan's test (P<0.05).
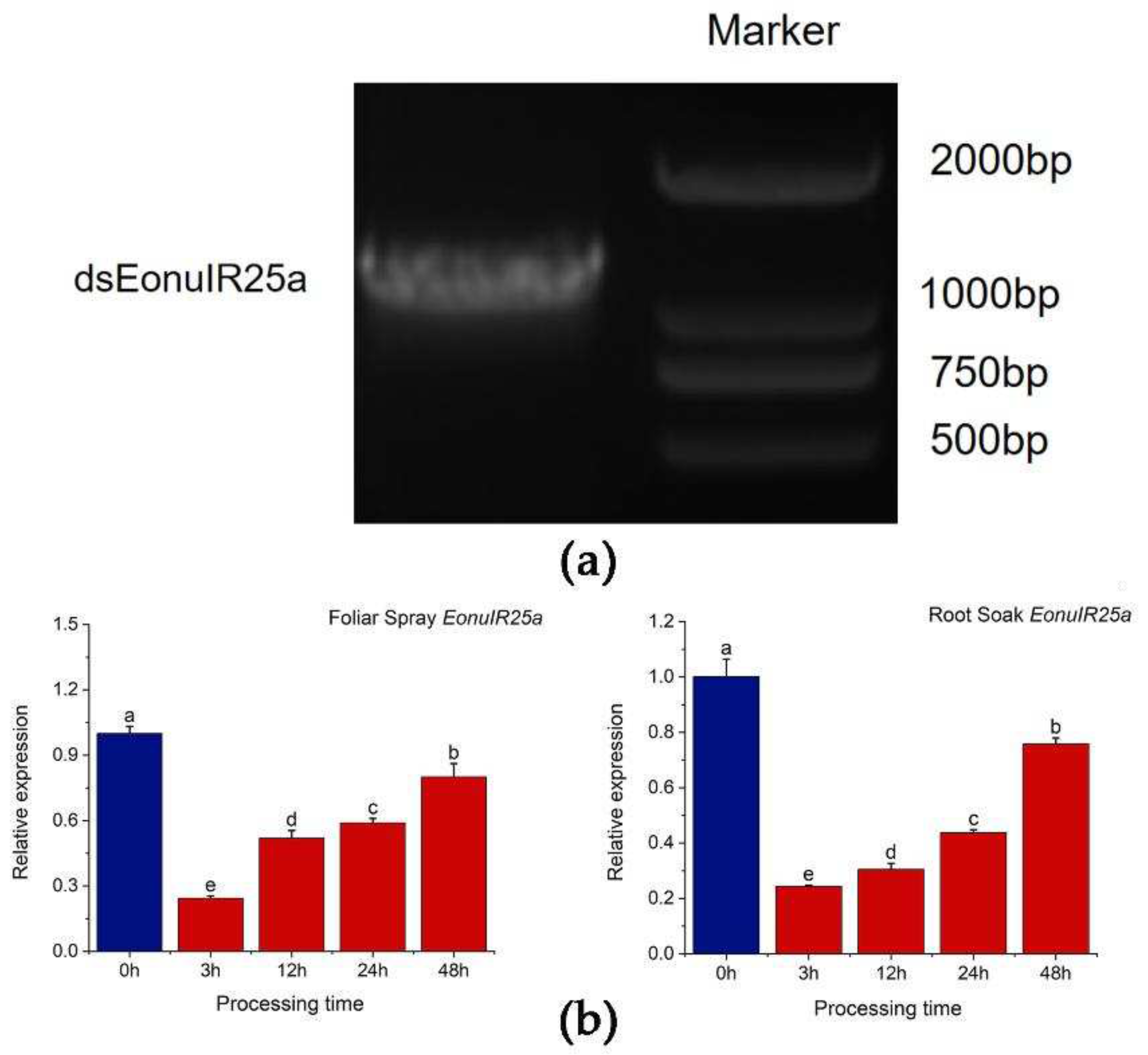
Figure 8.
Responses of E. onukii to compounds versus paraffin in a Y-tube olfactometer. (a): Response rate of E. onukii to compounds after 3 h of interference with EonuIR25a. (b): Response rate of E. onukii to compounds after 12 h of interference with EonuIR25a. note: asterisks indicate significant differences within a choice test (*P<0.05; **P<0.01); N. indicates no significant difference.
Figure 8.
Responses of E. onukii to compounds versus paraffin in a Y-tube olfactometer. (a): Response rate of E. onukii to compounds after 3 h of interference with EonuIR25a. (b): Response rate of E. onukii to compounds after 12 h of interference with EonuIR25a. note: asterisks indicate significant differences within a choice test (*P<0.05; **P<0.01); N. indicates no significant difference.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



