
Submitted:
03 May 2023
Posted:
04 May 2023
You are already at the latest version
Abstract
Vaginal ecosystem is a unique environment where, in physiological conditions, lactobacilli dominate. However, also pathogenic microbial species, responsible of vaginitis and vaginosis, can harbor vaginal microbiota. To extend the data published by De Seta and Larsen in Pathogens (2021), here we analyzed both the anti-Candida and anti-inflammatory properties of the vaginal gel formulation, Respecta® Balance Gel (RBG), commercialized as an adjuvant to treat vaginitis and vaginosis. We evaluated its activity by an in vitro model where a monolayer of A-431 vaginal epithelial cells was infected by Candida albicans in the presence of RBG or the placebo formulation (pRBG). Specifically, we tested the RBG capacity to counteract C. albicans virulence factors as well as their anti-inflammatory properties. Our results show that, unlike the placebo, RBG reduces ,reduces, C. albicans adhesion, its capacity to form hyphae and C. albicans-induced vaginal cell damage. Interestingly, both RBG and pRBG reduce LPS-induced IL-8 secretion (being RBG the most effective) demonstrating that also the placebo retains anti-inflammatory properties. By our experimental approach we highlight the possible role of farnesol on such effects, but we would like to point out that lactic acid, polydextrose and glycogen too must be relevant in the actual application. Taken together, our results show that RBG impairs C. albicans virulence and is able to reduce the inflammation in the vaginal environment, ultimately allowing the establishment of a balanced vaginal ecosystem.
Keywords:
vaginal infection
; C. albicans
; dysbiosis
; topic treatments
; anti-inflammatory effect
; antifungal effect
; vaginal ecosystem
; IL-8
; vaginal epithelial cells
; dimorphic transition
; farnesol
1. Introduction
Vulvovaginal candidiasis (VVC) is a painful condition affecting up to 75% of women during their child-bearing age at least once in their lifetime [1]. Furthermore, 5 – 8% of them develop the chronic form of the disease (RVVC), namely 4 or more symptomatic episodes per year [2]. A global study published in 2018 reports the worldwide prevalence of RVVC at approximately 138 million women annually, pointing out the substantial morbidity and economic burden of the pathology [3]. Vaginal pain, burning sensation, itching and abnormal vaginal discharge are the most typical VVC symptoms and chronicity to RVVC leads to negative consequences for physical and mental health [4]. VVC onset is due to the pathological overgrowth of Candida species and in 75 – 90% of cases C. albicans is the species responsible for the disease [5]. This fungus normally dwells in the vaginal mucosa as a commensal, together with the bacterial community, where it establishes a delicate balance supervised by the immune system [6]. Candida plays a dual role in the vaginal ecosystem: it can behave as the classic opportunistic pathogen under host dysbiosis conditions, but it can also play a leading role in triggering the VVC in immunocompetent healthy women [7]. What induces C. albicans to switch from a harmless commensal to a virulent pathogen, during the VVC onset, is still unclear. It is widely demonstrated that the disease onset can be facilitated by several predisposing host factors, such as gene polymorphisms, diabetes, prolonged antibiotic therapies, oral estrogen administration, hormone replacement therapy and psychosocial stresses [8,9,10]. Being a dimorphic fungus, C. albicans can assume two different morphologies: yeast cell and hyphal form. Yeast cells are typical of the beneficial Candida colonization of the vaginal mucosa. Differently, hyphae produced by yeasts germination are common in symptomatic vaginitis [11,12]. Colonization starts when Candida cells adhere to the vaginal mucosa. Dysbiosis conditions, the presence of predisposing factors or the occurrence of other (yet unknown) events induce the yeasts-to-hyphae transition. Hyphae, which are considered the most adherent morphology of C. albicans [13], promote the invasion of the vaginal epithelium [7]. Tissue damage related to the hyphal form is also associated with the production of candidalysin, the only C. albicans toxin known to date [14], which is produced by adherent hyphae during the epithelial penetration [15]. Candidalysin exerts a lytic activity and induces neutrophils recruitment [16] due to proinflammatory cytokines released by damaged vaginal epithelium [17]. In addition, adhesion capacity, extracellular enzymes secretion (such as aspartyl proteases, and phospholipase B), biofilm production and several quorum sensing (QS) molecules secreted by C. albicans can be considered as other important virulence factors [18]. To date, only four different QS molecules have been identified in the Fungi kingdom: farnesol, tyrosol, phenylethanol and tryptophol. These molecules are all involved in the inter-microbial communication and regulate Candida morphogenesis, virulence and apoptosis [19]. Farnesol is a sesquiterpene alcohol synthesized by C. albicans starting from farnesyl pyrophosphate (FPP) [20] and it was the first identified eukaryotic quorum sensing (QS) molecule. Farnesol plays a key role in C. albicans virulence by regulating biofilm formation, yeast-to-hyphae transition [21,22] and by affecting the expression of genes involved in the protection of the fungus against oxidative stress [23]. In addition, farnesol may impact on host immune system, by affecting macrophages phagocytic and antimicrobial potential [24]. Furthermore, its capacity to downregulate the expression of several inflammatory mediators demonstrates that farnesol has also an anti-inflammatory activity [25]. The wide spectrum of farnesol biological functions, that can be beneficial both for Candida and for the host, opens to investigate if its employment in antifungal therapy may be beneficial. Despite Candida virulence factors are essential in the early phases of VVC, the chronicity of the disease is widely mediated by the host immune response [26]. Specifically, C. albicans overgrowth stimulates vaginal epithelial cells to produce alarmins and proinflammatory cytokines, which in turn lead to neutrophils recruitment and consequently to the inflammatory symptoms [27]. It is also essential to consider that resident microbiota, often dominated by Lactobacillus species [28], could play a role as well in the VVC onset. Indeed, numerous studies have demonstrated the antifungal activity of Lactobacillus species against C. albicans [29,30,31,32] and non-albicans Candida species [33,34,35,36]. Lactobacilli may inhibit Candida overgrowth by several mechanisms, including the production of lactic acid, which causes the acidification of the vaginal environment. The latter, indeed, is typically acid (pH near 4.0) during almost the entire menstrual cycle. The acidification inhibits C. albicans hyphal growth [37,38], pointing out to the importance of producing acids (such as lactic acid) by resident bacteria in counteracting fungal virulence. Nonetheless, the potential role of the healthy vaginal microbiota in the VVC onset has not been completely identified yet. Respecta® Balance Gel (RBG) is a proprietary topical gel for vaginal health produced by BioFarma S.p.A. (Udine, Italy) and commercialized as an adjuvant for the treatment of vaginitis and vaginosis. The Manufacturers claim that RBG healthful effects are due to the restoration of the physiological pH of the vaginal environment, to the increased growth of beneficial microorganisms, to the hydration and protection of the vaginal tissue and to the formation of a protective film. The active components of the gel formulation are polydextrose, lactic acid, farnesol and glycogen. Farnesol activity has been described above. Polydextrose and glycogen act both as prebiotics since they promote eubiosis [39,40]. Lactic acid maintains an acidic vaginal environment in order to counteract possible pathogens [7].
In this work, we analyzed in vitro the anti-fungal and anti-inflammatory effect of RBG.
2. Materials and Methods
2.1. Fungal strains and growth conditions
The reference strain C. albicans SC5314 (ATCC MYA-2876) and the bioluminescent strain of C. albicans gLUC59 (BLI-Ca) were employed in the experimental procedures. Both strains had been stored in frozen stocks at -80°C in Sabouraud Dextrose Broth (Condalab, Spain) supplemented with 15% glycerol. After thawing, the fungi were grown in liquid YPD medium (Yeast extract - Peptose - Dextrose, Scharlab S.L., Spain) and incubated at 37 °C under aerobic conditions for 24 hours. Fungi in the exponential growth phase were used in each experiment. C. albicans gLUC59 was employed in experimental procedures related to sections 2.5, 2.6, 2.8 and 2.10. C. albicans SC5314 was used in experiments described in sections 2.7 and 2.9.
2.2. Vaginal epithelial cells
The A-431 cell line, from vaginal epithelial squamous cell carcinoma (ATCC CLR-1555) was employed. These cells were cultured in DMEM (Dulbecco's Modified Eagle Medium, PAN Biotech) supplemented with L-glutamine (2 nM) (Euroclone SpA, Italy), penicillin (100 U/ml) (Euroclone SpA, Italy), streptomycin (100 μl/ml) (Euroclone SpA, Italy), ciprofloxacin (20 mg/ml) (Euroclone SpA, Italy) and FBS (Fetal Bovine Serum, 10% or 5%, SIGMA-Aldrich, USA). The cell line was kept viable by subculturing twice a week and incubated at 37°C with 5% CO2.
2.3. Respecta® Balance Gel (RBG)
RBG was provided by BioFarma S.p.A. (Udine, Italy) and stored at +4°C. The Company provided information concerning the components of the gel, even though exact quantities of each component are proprietary information and cannot be reported. In our experiments, 2 different RBG formulations were assessed: in addition to the standard formulation (RBG), a placebo formulation (pRBG) was provided; compared to RBG, the pRBG lacks polidextrose, glycogen, farnesol and lactic acid. The composition of each RBG formulation is detailed in Table 1. Both RBG and pRBG were diluted in 50 ml of DMEM + 5% of FBS or YPD broth + 10% FBS or Roswell Park Institute da Moore (RPMI) + MOPS (35 mg/ml) + glucose (18 mg/ml) and their final pH was measured by a pH meter (Hanna Instruments, Italy). Dilutions were stored at +4°C for not more than 2 weeks.
2.4. Establishment of A-431 epithelial cells monolayer infected with C. albicans in the presence or not of RBG
Vaginal epithelial cells infections were performed using 96-well plates (Costar 3595, Corning, USA). A-431 cells (5x105 cells/ml, 200 μl per well) were seeded and then incubated overnight at 37°C with 5% CO2. Before being infected, the epithelial cell monolayer was washed with Phosphate-Buffered Saline (PBS, Dutscher, France) kept at room temperature. Then epithelial cells were infected with C. albicans, in the presence or not of RBG, by using a Multiplicity of Infection (MOI) of 1:1, namely 5x105 CFU/ml.
2.5. Assessment of vaginal epithelial cells damage
Vaginal epithelial cells damage induced by treatment with RBGs at different dilutions was evaluated after 24 hours of incubation at 37°C with 5% CO2. Cytotoxicity was quantified by analyzing lactate dehydrogenase (LDH) release in the culture medium employing a commercially available kit (Abcam, UK) and following the Manufacturer’s instructions. LDH quantification was also performed for the assessment of C. albicans-induced vaginal cells damage in the presence or not of RBG or pRBG after 24 h of infection at 37°C with 5% CO2. Percentage of damage was calculated as:
where low control is represented by uninfected cells (basal LDH release) and high control is given by uninfected cells lysed with 0.2% Triton X-100 after the incubation (maximum LDH release).
2.6. RBGs impact on C. albicans growth during vaginal epithelial cell infection
The effect of RBG on C. albicans growth capacity was assessed by using the bioluminescent strain gLUC59 (BLI-Ca). The epithelial cell monolayer was infected with 200 μl of BLI-Ca (5x105 CFU/mL) in the presence or not of RBG or pRBG. After 24 h of incubation at 37°C with 5% CO2, coelenterazine (2 μM) was added to each well and the bioluminescent signal was acquired with a Luminometer (FluoroskanTM Microplate reader, Thermo Scientific). Data were expressed as Relative Luminescence Units (RLU).
2.7. RBGs direct effect on Candida growth and metabolic activity
RBG anti-Candida direct effect and its impact on fungal metabolic activity were analyzed after 24 h of incubation. One hundred microliters of C. albicans (5x105 CFU/mL) in RPMI supplemented with MOPS and glucose were seeded in a 96-well plate in the presence or not of RBG or pRBG and incubated at 37°C for 24 h. After incubation, fungal growth was quantified by optical density (OD) measurement at 540 nm wavelength by a plate reader (Sunrise Tecan, Switzerland). Subsequently, the medium of each well was removed and 100 μl of 2,3-bis(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide (XTT) solution, supplemented with menadione (1 μM), were added. The plate was covered with tinfoil and incubated at 37°C for 3 h. Then, 80 μl were collected from each well, transferred to wells of another microtiter plate and color intensity was quantified by measuring OD at 492 nm wavelength.
2.8. Effect of RBGs on C. albicans adhesion
C. albicans adhesion was evaluated after 2 h of incubation at 37°C in the presence or not of RBGs. Two hundred microliters of C. albicans in DMEM supplemented with 5% FBS were seeded in a 96-well plate, and, after incubation, a crystal violet (CV) assay was performed. Specifically, wells were washed with PBS to remove non-adherent fungal cells then, 100 μl of 1% CV were added to each well. After 5 minutes, CV was removed and two washes with PBS were carried out before adding 100 μl of 33% acetic acid to each well. Adhesion of C. albicans was quantified by evaluating the optical density (OD) at 570 nm wavelength using a spectrophotometer (Sunrise, Tecan, Switzerland).
2.9. Evaluation of RBGs effect on C. albicans hyphal formation
RBG effect on C. albicans ability to form hyphae was assessed in YPD broth supplemented with 10% FBS. One milliliter containing 5x105 CFU of C. albicans, in the presence or not of RBG or pRBG, was added to a 24-well plate (Greiner Bio-One, Austria) and then incubated at 37°C for 4 h. After incubation, 100 μL of each sample were recovered and placed onto a glass microscope slide. Yeast cells and hyphal fragments were counted (at least 3 fields for each experimental condition) by using an optical microscope (Nikon Eclipse 80i, Nikon Corporation, Japan). Data were expressed as %
of hyphal fragments.
2.10. Quantification of IL-1β and IL-8 production after C. albicans infection and LPS stimulation of vaginal epithelial cells in the presence or not of RBGs
The production of IL-1β and IL-8 by A-431 cells monolayer was analyzed after 24 h of C. albicans infection (MOI 1:1) or LPS (10 μg/ml) and LPS+ Dexamethasone (DEX) (LPS: 10 μg/ml + DEX: 1 mM) stimulation in the presence or not of RBG or pRBG, by using specific commercially available sandwich ELISA kits (Boster Bio, USA for both IL-1β and IL-8), according to the Manufacturers’ instructions.
2.11. Statistical analysis
Shapiro-Wilk test was used to analyze data distribution within each experimental group. Data from all the experiments were normally distributed and statistical analyses were performed by One-Way ANOVA test followed by Tukey’s multiple comparisons tests. All statistical analyses were carried out using GraphPad Prism 9 software. Values of * p < 0.05; ** p < 0.01 and **** p < 0.0001 were considered statistically significant.
3. Results
3.1. RBG reduced C. albicans-induced vaginal epithelial cell damage without affecting fungal growth.
First, we set-up the experimental protocol to analyze the effect of RBG and pRBG on vaginal cells in vitro. We performed dose-dependent experiments to identify the highest RBG concentration tolerated by the vaginal cells, i.e., the one incapable to induce cell damage. RBG showed to be well tolerated by vaginal epithelial cells starting from dilution 1:300 after 24 h of contact time. Placebo RBG (pRBG) did not show any cell toxicity at any dilution tested. Therefore, the dilution 1:300 (highlighted in light green in Figure 1) was chosen for both RBG and pRBG (Figure 1), and such dilution was employed in all the subsequent experiments. Of note, the pH of RBG and pRBG at dilutions from 1:150 to 1:300 was always around 7.4, resulting superimposable to the pH of the medium alone.
Next, C. albicans-induced epithelial vaginal cells damage after 24 h of infection was assessed in the presence or absence of RBG or pRBG. Our results show that RBG, but not pRBG, was able to reduce significantly C. albicans-induced cell damage by an inhibition rate of about 23% (Figure 2A).
In parallel, fungal growth was assessed after 24 h of vaginal epithelial cells infection with BLI-C. albicans (BLI-Ca) in the presence or absence of RBG and pRBG. Results show that neither RBG nor pRBG were able to significantly impair C. albicans growth (Figure 2B) during epithelial cells infection, suggesting that the observed reduction of cell damage was not due to a direct antifungal effect.
Interestingly, by analyzing the direct antifungal activity of RBG and pRBG against C. albicans, without cells monolayer, we observed a slight but significant reduction of Candida growth with RBG , but not with pRBG (Figure 3A).
In contrast, by analyzing the metabolic activity of C. albicans under the same experimental conditions, no significant differences could be observed between C. albicans alone and RBG or pRBG-treated C. albicans (Figure 3B).
3.2. RBG reduced C. albicans adhesion and hyphae formation.
To investigate the mechanism involved in the inhibition of C. albicans-induced cell damage by RBG, we evaluated its impact on fungal adhesion. RBG was shown to reduce significantly the number of adherent C. albicans cells after 2 h of incubation at 37°C, with an inhibition rate of about 26%. The placebo formulation did not cause any inhibition (Figure 4A). As filamentation is a key virulence trait implicated in C. albicans-induced cell damage, as well as adhesion to biotic and abiotic substrates, we analyzed the capacity of RBG to modulate hyphae formation. Our results show that RBG significantly reduced C. albicans filamentation with an inhibition rate of about 31%, whereas pRBG did not cause any inhibitory effect (Figure 4B–C). Notably, by comparing pRBG with RBG, a significant difference could be observed for both adhesion and hyphal formation (Figure 4A–C).
3.3. RBG effects on IL-1β and IL-8 production by vaginal epithelial cells
We next analyzed the capacity of RBG to modulate epithelial immune response to C. albicans infection and to LPS stimulation. Specifically, we assessed IL-1β and IL-8 production after 24 h infection of vaginal epithelial cells with C. albicans in the presence or absence of RBG and pRBG. Our results show that C. albicans could induce the release of a significant (albeit modest) amount of IL-1β by vaginal epithelial cells and that such amount was not modulated by either RBG or pRBG. In addition, the 24 h stimulation of vaginal epithelial cells with LPS induced the secretion of a small amount of IL-1β, not significantly different from the amount secreted by cells alone. No modulation of IL-1β secretion was observed after epithelial stimulation with LPS+DEX or LPS+RBGs; moreover, no modulation of IL-1β secretion was detected in the presence of RBG or pRBG alone (Figure 5A).
Differently, C. albicans did not induce the secretion of IL-8 by vaginal epithelial cells after 24h of infection under our experimental conditions. Accordingly, no effect could be observed by infecting vaginal epithelial cells with C. albicans + RBG or pRBG. However, LPS strongly stimulated IL-8 production by vaginal epithelial cells after 24 h. By adding the RBG and pRBG in combination with LPS, we observed a significant reduction of IL-8 with respect to the stimulation by LPS alone. As expected, the addition of DEX reduced the IL-8 levels in a highly significant manner. Interestingly, such reduction was comparable to the reduction induced by RBG (Figure 5B).
4. Discussion
In this work, we have tested the anti-fungal and anti-inflammatory properties of the vaginal gel RBG, a commercially available formulation used as an adjuvant in the treatment of vaginitis and vaginosis. The Manufacturer claims that this gel helps to restore the physiological pH of the vaginal environment and to promote the growth of a balanced microbiota [41]. De Seta and Larsen have recently shown in an in vitro study that RBG gel has antimicrobial properties against several microorganisms relevant to vaginal infections, such as Candida spp. and Gardnerella vaginalis. They conclude that the RBG formulation contains different components, including some excipients (such as EDTA), that can play a role in counteracting microbial growth. Although they ascribe the antimicrobial effect mainly to the lactic acid, they hypothesize an additional mechanistic and possible synergistic interaction between active components (polydextrose, lactic acid, farnesol) that may occur when RBG encounters vaginal fluids [41]. We exclude an effect of lactic acid in our experimental conditions, since the pH of both RBG and the placebo formulation is around 7.4 when diluted from 1:150 to 1:300, and therefore superimposable to the pH of the medium alone. Moreover, RBG contains prebiotic compounds (such as glycogen and polydextrose) [39,40] that can promote the growth of a balanced microbiota, thus contributing to counteract potentially pathogenic microorganisms in the vaginal environment. Therefore, it is very likely that the main role of glycogen, polydextrose, and lactic acid must be exerted in the actual application.
In this work, we provide new data on RBG, studying its activity against C. albicans by an in vitro model that could mimic, as closely as possible, a physiological situation. Specifically, a monolayer of epithelial vaginal cells was infected with C. albicans in the presence or absence of the commercially available RBG (RBG) and a placebo formulation (pRBG). Our results clearly show that only RBG, but not pRBG, is able to reduce Candida-induced cell damage. It should be noted that RBG has been employed at a dilution of 1:300, i.e., the highest non-toxic dose for epithelial cells. This dilution is far below the doses tested by De Seta and Larsen to study the direct antimicrobial activity in vitro [41].
In parallel, during epithelial cells infection, Candida growth has not been affected by the addition of any of the 2 RBG formulations. Therefore, we hypothesize that the observed reduction of cell damage, in our experimental model, is not due to a direct antifungal effect exerted by RBG, but rather to a reduction in the ability of Candida to express its virulence potential (for example, its capacity to adhere and/or to produce hyphae). There are two possible reasons that may explain this apparent discrepancy in our results, with respect to De Seta and Larsen’s. First, as mentioned above, the dilution used in our protocol (1:300) is far below the doses tested by De Seta and Larsen. Second, the effect of the gel formulation on Candida, when it infects an epithelium monolayer, mirrors a situation more similar to the one occurring in vivo, when the fungus dwells within a biological environment where it can react more efficiently. It should be pointed out that, in line with the data obtained by De Seta and Larsen, by analyzing the direct antifungal activity of RBG against C. albicans, i.e., assessed without the vaginal cells monolayer, a slight, albeit significant, reduction of overall Candida growth has been detected after incubation with RBG, but not with pRBG. However, the results of XTT test, used to evaluate the metabolic activity of the fungus, reveal that no significant differences could be observed between live C. albicans cells in the presence or absence of both RBG and pRBG.
The morphological transition from yeasts to hyphae represents a key step of C. albicans virulence, because it increases deep tissues invasiveness and resistance to multiple environmental and chemical stresses [42,43]. Therefore, an effective strategy against C. albicans should include the reduction of its capacity to adhere and to form hyphae, rather than the mere growth reduction. Indeed, C. albicans could be part of the vaginal microbiota of healthy women, and in its yeast form it is considered as a harmless commensal [7].
In line with this idea, our results show that RBG significantly reduces the capacity of C. albicans to adhere; in addition, RBG impairs C. albicans capacity to form hyphae. We tend to ascribe these effects essentially to farnesol since, as mentioned above, the role played by the other 3 active components (lactic acid, polydextrose and glycogen) would become evident during the treatment. Farnesol is a quorum-sensing (QS) molecule that, together with tyrosol, is involved in biofilm formation through the modulation of several virulence factors of the fungus, including the dimorphic transition. In particular, while tyrosol stimulates hyphae production, therefore exerting a pivotal role in biofilm formation, farnesol counteracts yeast-to-hyphae transition (one of the effects of RBG) and inhibits biofilm formation [44]. In addition, glycogen and polydextrose are two active components of RBG, with prebiotic activity. Both may favor the proliferation of “beneficial” bacteria, which are part of the healthy vaginal microbiota. Indeed, a glycogen-rich vaginal milieu has been demonstrated to favor the proliferation of lactobacilli, which in turn increase the production of lactic acid and decrease the pH [45]. Therefore, the role of glycogen in the RBG formulation must be an indirect one, by exerting a positive effect on the vaginal microbiota. It is well-known that a lactobacilli-dominated microbiota provides an effective first line of defense against invading pathogens, including Candida [46], through several mechanisms: competitions for adhesion sites on the host cells, competition for nutrients, and release of antimicrobial compounds such as bacteriocins, hydrogen peroxide and organic acids [47]. Similarly, polydextrose too should be considered a prebiotic component, capable to play a role in vivo stimulating the growth of the “good” bacteria.
The role of inflammatory response on vaginal infections onset is well documented [7,48]. Concerning vulvovaginal candidiasis (VVC), although fungal virulence factors are important for triggering the disease, its spread is largely mediated by the host immune system. In particular, C. albicans overgrowth, yeast-to-hyphae transition and candidalysin production all stimulate the epithelial cells to secrete antimicrobial peptides and proinflammatory cytokines and chemokines, such as IL-1β and IL-8. This leads to a massive recruitment of non-protective neutrophils that in turn generate a hyper-inflammatory environment mainly responsible of the VVC symptoms [7]. Similarly, numerous studies have demonstrated the occurrence of high levels of cytokines and chemokines, such as IL-1β, TNF-α, IL-6 and IL-8, in vaginal fluids of women with bacterial vaginosis (BV) [48].
By our in vitro model, we have analyzed the capacity of RBG to modulate cytokines and chemokines production by epithelial cells during C. albicans infection or after treatment with LPS. Our results show that both RBG and pRBG display some degree of activity in reducing LPS-induced IL-8 production. Notably, although both RBG and its placebo significantly reduce IL-8 secretion by LPS-stimulated epithelial cells, the reduction observed after RBG treatment is comparable to that observed after DEX treatment. EDTA (an excipient contained in both RBG formulations) has been reported to enhance the activity of host defense factors, including lysozyme and lactoferrin [49], both of which are also components of vaginal secretions [50,51]. Therefore, our hypothesis is that by potentiating the epithelial barrier, both RBG and pRBG are able to reduce LPS-induced IL-8 secretion.
5. Conclusions
These results expand the data by De Seta and Larsen [41], because RBG has the capacity to impair Candida virulence by counteracting C. albicans-induced vaginal epithelial cell damage. Such effect seems to be achieved through an impairment of specific virulence factors of the fungus, i.e., the reduction of its capacity to adhere and to undergo the yeasts-to-hyphae morphological transition, which are two key factors in fungal pathogenesis. Interestingly, the gel formulations seem not to impair neither fungal growth, nor its metabolic activity. Consequently, the reduced epithelial cell-damage exerted by RBG, but not by pRBG, is not achieved through a reduction of the fungal burden. It is our hypothesis that, among the active compounds, the main role may be played by farnesol, which is a well-known QS Candida molecule whose role in promoting yeast morphology has been described [44]. Accordingly, literature data report that farnesol does not impair Candida growth [22]. The effects of RBG in our experimental system are summarized in Figure 6. This mechanism of action highlights the impact of farnesol without including the role of lactic acid, polydextrose and glycogen. As mentioned above, the effects of such 3 active components are relevant mainly in vivo.
In addition, some of the excipients contained in both RBG and pRBG (such as EDTA) may help in reducing the IL-8 secretion by LPS-stimulated epithelial cells, and such effect is superimposable to the same inhibition obtained after treatment with DEX. The resulting IL-8 secretion inhibition may be responsible (in combination with other mechanisms) for the recovery of a low inflammatory vaginal environment, which in turn is the starting point for the establishment of vaginal eubiosis. In addition, such effect may be strengthened by glycogen, polydextrose and lactic acid contained in RBG that, by helping the proliferation of lactobacilli, provide another mean for the vaginal environment to prevent invasion of potential pathogens like Candida.
Author Contributions
L.S., A.A., F.D.S. and E.P. have contributed significantly to this work including conceptualization, original draft, review, and editing. E.B. contributed to final review and editing. L.S., G.B.S.R., C.A.S.R. and N.P. were responsible for methodology, laboratory investigation and research. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding, however, BioFarma S.p.A. (Udine, Italy), provided the Respecta® Balance Gel for laboratory use as described in the Methods section. E.P. was supported by FAR instrumentation 2022.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Cinzia Rosselli from the University of Modena and Reggio Emilia, “Centro Linguistico di Ateneo”, for proof-reading the English text.
Conflicts of Interest
Financial support was not received for the conduct of this research; apart from BioFarma S.p.A. (Udine, Italy) providing Respecta® Balance Gel, all materials and equipment used in this research came from supplies purchased by E.P. and E.B. F.D.S. provided medical advice to BioFarma S.p.A. (Udine, Italy) limited to clinical research issues. BioFarma S.p.A. had no role in the development or funding of this project and was not allowed to review this manuscript before publication.
References
- Blostein F, Levin-Sparenberg E, Wagner J, Foxman B. 2017. Recurrent vulvovaginal candidiasis. Ann Epidemiol 27:575-582.e3.
- Cassone, A. 2015. Vulvovaginal Candida albicans infections: pathogenesis, immunity and vaccine prospects. BJOG: Int J Obstet Gy 122:785–794.
- Denning DW, Kneale M, Sobel JD, Rautemaa-Richardson R. 2018. Global burden of recurrent vulvovaginal candidiasis: a systematic review. Lancet Infect Dis 18:e339–e347.
- 2020. Vaginitis in Nonpregnant Patients: ACOG Practice Bulletin, Number 215. Obstetrics & Gynecology 135:e1–e17.
- Rosati D, Bruno M, Jaeger M, ten Oever J, Netea MG. 2020. Recurrent Vulvovaginal Candidiasis: An Immunological Perspective. Microorganisms 8:144.
- Miró MS, Caeiro JP, Rodriguez E, Vargas L, Vigezzi C, Icely PA, Castillo GDV, Azcurra AI, Abiega CD, Riera FO, Sotomayor CE. 2021. Candida albicans Modulates Murine and Human Beta Defensin-1 during Vaginitis. JoF 8:20.
- Ardizzoni A, Wheeler RT, Pericolini E. 2021. It Takes Two to Tango: How a Dysregulation of the Innate Immunity, Coupled With Candida Virulence, Triggers VVC Onset. Front Microbiol 12:692491.
- Farr A, Effendy I, Frey Tirri B, Hof H, Mayser P, Petricevic L, Ruhnke M, Schaller M, Schaefer APA, Sustr V, Willinger B, Mendling W. 2021. Guideline: Vulvovaginal candidosis (AWMF 015/072, level S2k). Mycoses 64:583–602.
- McClelland RS, Richardson BA, Hassan WM, Graham SM, Kiarie J, Baeten JM, Mandaliya K, Jaoko W, Ndinya-Achola JO, Holmes KK. 2009. Prospective study of vaginal bacterial flora and other risk factors for vulvovaginal candidiasis. J Infect Dis 199:1883–1890.
- Liu M-B, Xu S-R, He Y, Deng G-H, Sheng H-F, Huang X-M, Ouyang C-Y, Zhou H-W. 2013. Diverse vaginal microbiomes in reproductive-age women with vulvovaginal candidiasis. PLoS One 8:e79812.
- Roselletti E, Perito S, Sabbatini S, Monari C, Vecchiarelli A. 2019. Vaginal Epithelial Cells Discriminate Between Yeast and Hyphae of Candida albicans in Women Who Are Colonized or Have Vaginal Candidiasis. J Infect Dis 220:1645–1654.
- Pericolini E, Perito S, Castagnoli A, Gabrielli E, Mencacci A, Blasi E, Vecchiarelli A, Wheeler RT. 2018. Epitope unmasking in vulvovaginal candidiasis is associated with hyphal growth and neutrophilic infiltration. PLoS One 13:e0201436.
- Naglik JR, Moyes DL, Wächtler B, Hube B. 2011. Candida albicans interactions with epithelial cells and mucosal immunity. Microbes Infect 13:963–976.
- Moyes DL, Wilson D, Richardson JP, Mogavero S, Tang SX, Wernecke J, Höfs S, Gratacap RL, Robbins J, Runglall M, Murciano C, Blagojevic M, Thavaraj S, Förster TM, Hebecker B, Kasper L, Vizcay G, Iancu SI, Kichik N, Häder A, Kurzai O, Luo T, Krüger T, Kniemeyer O, Cota E, Bader O, Wheeler RT, Gutsmann T, Hube B, Naglik JR. 2016. Candidalysin is a fungal peptide toxin critical for mucosal infection. Nature 532:64–68.
- Ho J, Camilli G, Griffiths JS, Richardson JP, Kichik N, Naglik JR. 2021. Candida albicans and candidalysin in inflammatory disorders and cancer. Immunology 162:11–16.
- Ho J, Yang X, Nikou S-A, Kichik N, Donkin A, Ponde NO, Richardson JP, Gratacap RL, Archambault LS, Zwirner CP, Murciano C, Henley-Smith R, Thavaraj S, Tynan CJ, Gaffen SL, Hube B, Wheeler RT, Moyes DL, Naglik JR. 2019. Candidalysin activates innate epithelial immune responses via epidermal growth factor receptor. Nat Commun 10:2297.
- Richardson JP, Willems HME, Moyes DL, Shoaie S, Barker KS, Tan SL, Palmer GE, Hube B, Naglik JR, Peters BM. 2018. Candidalysin Drives Epithelial Signaling, Neutrophil Recruitment, and Immunopathology at the Vaginal Mucosa. Infect Immun 86.
- Calderone RA, Fonzi WA. 2001. Virulence factors of Candida albicans. Trends in Microbiology 9:327–335.
- Ardizzoni A, Boaretto G, Pericolini E, Pinetti D, Capezzone de Joannon A, Durando L, Ragni L, Blasi E. 2022. Effects of benzydamine and mouthwashes containing benzydamine on Candida albicans adhesion, biofilm formation, regrowth, and persistence. Clin Oral Invest 26:3613–3625.
- Hornby JM, Kebaara BW, Nickerson KW. 2003. Farnesol Biosynthesis in Candida albicans : Cellular Response to Sterol Inhibition by Zaragozic Acid B. Antimicrob Agents Chemother 47:2366–2369.
- Polke M, Leonhardt I, Kurzai O, Jacobsen ID. 2018. Farnesol signalling in Candida albicans – more than just communication. Critical Reviews in Microbiology 44:230–243.
- Hornby JM, Jensen EC, Lisec AD, Tasto JJ, Jahnke B, Shoemaker R, Dussault P, Nickerson KW. 2001. Quorum sensing in the dimorphic fungus Candida albicans is mediated by farnesol. Appl Environ Microbiol 67:2982–2992.
- Deveau A, Piispanen AE, Jackson AA, Hogan DA. 2010. Farnesol Induces Hydrogen Peroxide Resistance in Candida albicans Yeast by Inhibiting the Ras-Cyclic AMP Signaling Pathway. Eukaryot Cell 9:569–577.
- Abe S, Tsunashima R, Iijima R, Yamada T, Maruyama N, Hisajima T, Abe Y, Oshima H, Yamazaki M. 2009. Suppression of anti- Candida activity of macrophages by a quorum-sensing molecule, farnesol, through induction of oxidative stress. Microbiology and Immunology 53:323–330.
- Jung Y, Hwang S, Sethi G, Fan L, Arfuso F, Ahn K. 2018. Potential Anti-Inflammatory and Anti-Cancer Properties of Farnesol. Molecules 23:2827.
- Casadevall A, Pirofski LA. 1999. Host-pathogen interactions: redefining the basic concepts of virulence and pathogenicity. Infect Immun 67:3703–3713.
- Yano J, Lilly E, Barousse M, Fidel PL. 2010. Epithelial cell-derived S100 calcium-binding proteins as key mediators in the hallmark acute neutrophil response during Candida vaginitis. Infect Immun 78:5126–5137.
- Ravel J, Gajer P, Abdo Z, Schneider GM, Koenig SSK, McCulle SL, Karlebach S, Gorle R, Russell J, Tacket CO, Brotman RM, Davis CC, Ault K, Peralta L, Forney LJ. 2011. Vaginal microbiome of reproductive-age women. Proc Natl Acad Sci U S A 108 Suppl 1:4680–4687.
- Wang S, Wang Q, Yang E, Yan L, Li T, Zhuang H. 2017. Antimicrobial Compounds Produced by Vaginal Lactobacillus crispatus Are Able to Strongly Inhibit Candida albicans Growth, Hyphal Formation and Regulate Virulence-related Gene Expressions. Front Microbiol 08.
- Matsuda Y, Cho O, Sugita T, Ogishima D, Takeda S. 2018. Culture Supernatants of Lactobacillus gasseri and L. crispatus Inhibit Candida albicans Biofilm Formation and Adhesion to HeLa Cells. Mycopathologia 183:691–700.
- Jang SJ, Lee K, Kwon B, You HJ, Ko G. 2019. Vaginal lactobacilli inhibit growth and hyphae formation of Candida albicans. Sci Rep 9:8121.
- De Seta F, Parazzini F, De Leo R, Banco R, Maso GP, De Santo D, Sartore A, Stabile G, Inglese S, Tonon M, Restaino S. 2014. Lactobacillus plantarum P17630 for preventing Candida vaginitis recurrence: a retrospective comparative study. European Journal of Obstetrics & Gynecology and Reproductive Biology 182:136–139.
- Spaggiari L, Sala A, Ardizzoni A, De Seta F, Singh DK, Gacser A, Blasi E, Pericolini E. 2022. Lactobacillus acidophilus, L. plantarum, L. rhamnosus, and L. reuteri Cell-Free Supernatants Inhibit Candida parapsilosis Pathogenic Potential upon Infection of Vaginal Epithelial Cells Monolayer and in a Transwell Coculture System In Vitro. Microbiol Spectr 10:e02696-21.
- Parolin C, Croatti V, Laghi L, Giordani B, Tondi MR, De Gregorio PR, Foschi C, Vitali B. 2021. Lactobacillus Biofilms Influence Anti-Candida Activity. Front Microbiol 12:750368.
- Chew SY, Cheah YK, Seow HF, Sandai D, Than LTL. 2015. Probiotic L actobacillus rhamnosus GR -1 and L actobacillus reuteri RC -14 exhibit strong antifungal effects against vulvovaginal candidiasis-causing C andida glabrata isolates. J Appl Microbiol 118:1180–1190.
- Salari S, Ghasemi Nejad Almani P. 2020. Antifungal effects of Lactobacillus acidophilus and Lactobacillus plantarum against different oral Candida species isolated from HIV/ AIDS patients: an in vitro study. Journal of Oral Microbiology 12:1769386.
- Noverr MC, Huffnagle GB. 2004. Regulation of Candida albicans morphogenesis by fatty acid metabolites. Infect Immun 72:6206–6210.
- Liang W, Guan G, Dai Y, Cao C, Tao L, Du H, Nobile CJ, Zhong J, Huang G. 2016. Lactic acid bacteria differentially regulate filamentation in two heritable cell types of the human fungal pathogen Candida albicans. Mol Microbiol 102:506–519.
- Goh YJ, Klaenhammer TR. 2014. Insights into glycogen metabolism in Lactobacillus acidophilus: impact on carbohydrate metabolism, stress tolerance and gut retention. Microb Cell Fact 13:94.
- do Carmo MMR, Walker JCL, Novello D, Caselato VM, Sgarbieri VC, Ouwehand AC, Andreollo NA, Hiane PA, Dos Santos EF. 2016. Polydextrose: Physiological Function, and Effects on Health. Nutrients 8:553.
- De Seta F, Larsen B. 2021. Antimicrobial Activity of a Vaginal Gel Formulation: Considerations Related to Vaginal Infection and Dysbiosis. Pathogens 10:1576.
- Tsui C, Kong EF, Jabra-Rizk MA. 2016. Pathogenesis of Candida albicans biofilm. Pathogens and Disease 74:ftw018.
- Desai, J. 2018. Candida albicans Hyphae: From Growth Initiation to Invasion. JoF 4:10.
- Kovács R, Majoros L. 2020. Fungal Quorum-Sensing Molecules: A Review of Their Antifungal Effect against Candida Biofilms. J Fungi (Basel) 6.
- Amabebe E, Anumba DOC. 2018. The Vaginal Microenvironment: The Physiologic Role of Lactobacilli. Front Med 5:181.
- Petrova MI, Lievens E, Malik S, Imholz N, Lebeer S. 2015. Lactobacillus species as biomarkers and agents that can promote various aspects of vaginal health. Front Physiol 6.
- Sun Z, Ge X, Qiu B, Xiang Z, Jiang C, Wu J, Li Y. 2023. Vulvovaginal candidiasis and vaginal microflora interaction: Microflora changes and probiotic therapy. Front Cell Infect Microbiol 13:1123026.
- Hedges SR, Barrientes F, Desmond RA, Schwebke JR. 2006. Local and Systemic Cytokine Levels in Relation to Changes in Vaginal Flora. J INFECT DIS 193:556–562.
- Li X, Wang X, Gao X. 2015. [Synergistic effects of lysozyme with EDTA-2Na on antibacterial activity]. Beijing Da Xue Xue Bao Yi Xue Ban 47:52–56.
- Schumacher GFB, Kim MH, Hosseinian AH, Dupon C. 1977. Immunoglobulins, proteinase inhibitors, albumin, and lysozyme in human cervical mucus. American Journal of Obstetrics and Gynecology 129:629–636.
- Mitsukawa K, Otsuki K, Yanaihara A, Sawada M, Iwasaki S, Okai T. 2006. Concentration of lactoferrin and interleukin-6 in cervical mucus from patients being treated for infertility: Lactoferrin and IL-6 in cervical mucus. Reproductive Medicine and Biology 5:105–109.
Figure 1.
Effect of RBG on vaginal epithelial cells. Cell damage was evaluated by the quantification of lactate dehydrogenase (LDH) release in the culture medium after 24 h of treatment with RBG and pRBG. The graph reports the mean percentage of damage SD from triplicate samples of at least 3 different experiments (range dilutions 1:250-1:350) and from triplicate samples of at least 2 different experiments (dilution 1:150; 1:200 and from 1:400).
Figure 1.
Effect of RBG on vaginal epithelial cells. Cell damage was evaluated by the quantification of lactate dehydrogenase (LDH) release in the culture medium after 24 h of treatment with RBG and pRBG. The graph reports the mean percentage of damage SD from triplicate samples of at least 3 different experiments (range dilutions 1:250-1:350) and from triplicate samples of at least 2 different experiments (dilution 1:150; 1:200 and from 1:400).
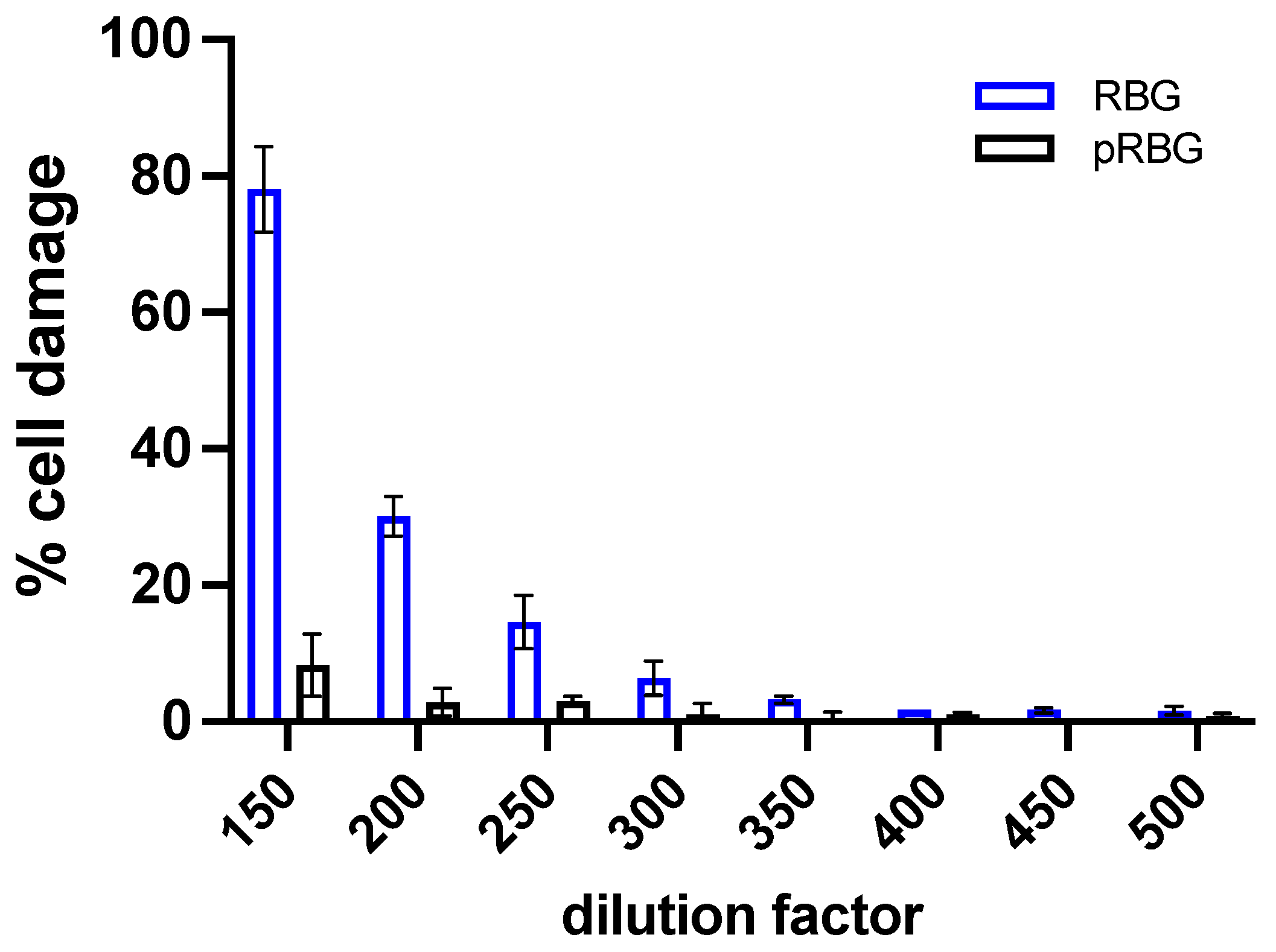
Figure 2.
Effect of RBG on C. albicans-induced cell damage and on fungal growth during vaginal cells infection. (A) Epithelial cells damage was evaluated by the quantification of lactate dehydrogenase (LDH) release in the culture supernatants after 24 h of C. albicans infection, in the presence or absence of RBG and pRBG. Data reported in the violin-plot are from triplicate samples of at least 3 different experiments. (B) C. albicans growth was analyzed by the determination of Relative Luminescence Units (RLU) emitted from live fungal cells after 24 h of infection of vaginal epithelial cells in the presence or absence of RBG and pRBG. The graph reports the mean RLU SD from triplicate samples of 3 different experiments.
Figure 2.
Effect of RBG on C. albicans-induced cell damage and on fungal growth during vaginal cells infection. (A) Epithelial cells damage was evaluated by the quantification of lactate dehydrogenase (LDH) release in the culture supernatants after 24 h of C. albicans infection, in the presence or absence of RBG and pRBG. Data reported in the violin-plot are from triplicate samples of at least 3 different experiments. (B) C. albicans growth was analyzed by the determination of Relative Luminescence Units (RLU) emitted from live fungal cells after 24 h of infection of vaginal epithelial cells in the presence or absence of RBG and pRBG. The graph reports the mean RLU SD from triplicate samples of 3 different experiments.
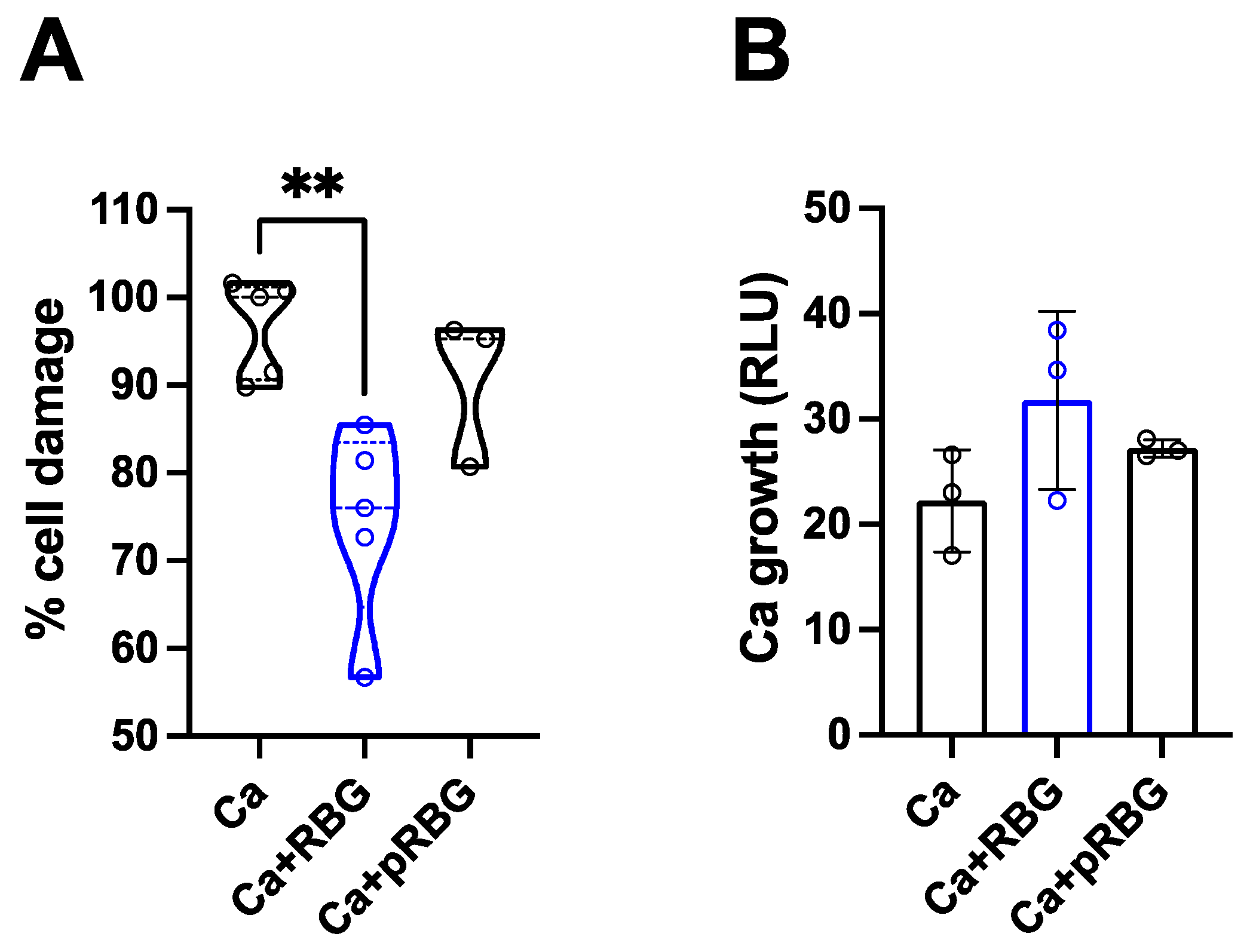
Figure 3.
RBG effects on C. albicans growth and metabolic activity. (A) The anti-Candida effect of RBG was analyzed by culturing C. albicans yeast cells in the presence or absence of RBG and pRBG for 24 h. After incubation, absorbance (OD540) was spectrophotometrically quantified. Data in the box-plots are from triplicate samples of 4 different experiments. (B) The metabolic activity of C. albicans after 24 h of incubation with RBG and pRBG was analyzed by XTT assay. Data in the box-plots are from triplicate samples of 4 different experiments.
Figure 3.
RBG effects on C. albicans growth and metabolic activity. (A) The anti-Candida effect of RBG was analyzed by culturing C. albicans yeast cells in the presence or absence of RBG and pRBG for 24 h. After incubation, absorbance (OD540) was spectrophotometrically quantified. Data in the box-plots are from triplicate samples of 4 different experiments. (B) The metabolic activity of C. albicans after 24 h of incubation with RBG and pRBG was analyzed by XTT assay. Data in the box-plots are from triplicate samples of 4 different experiments.
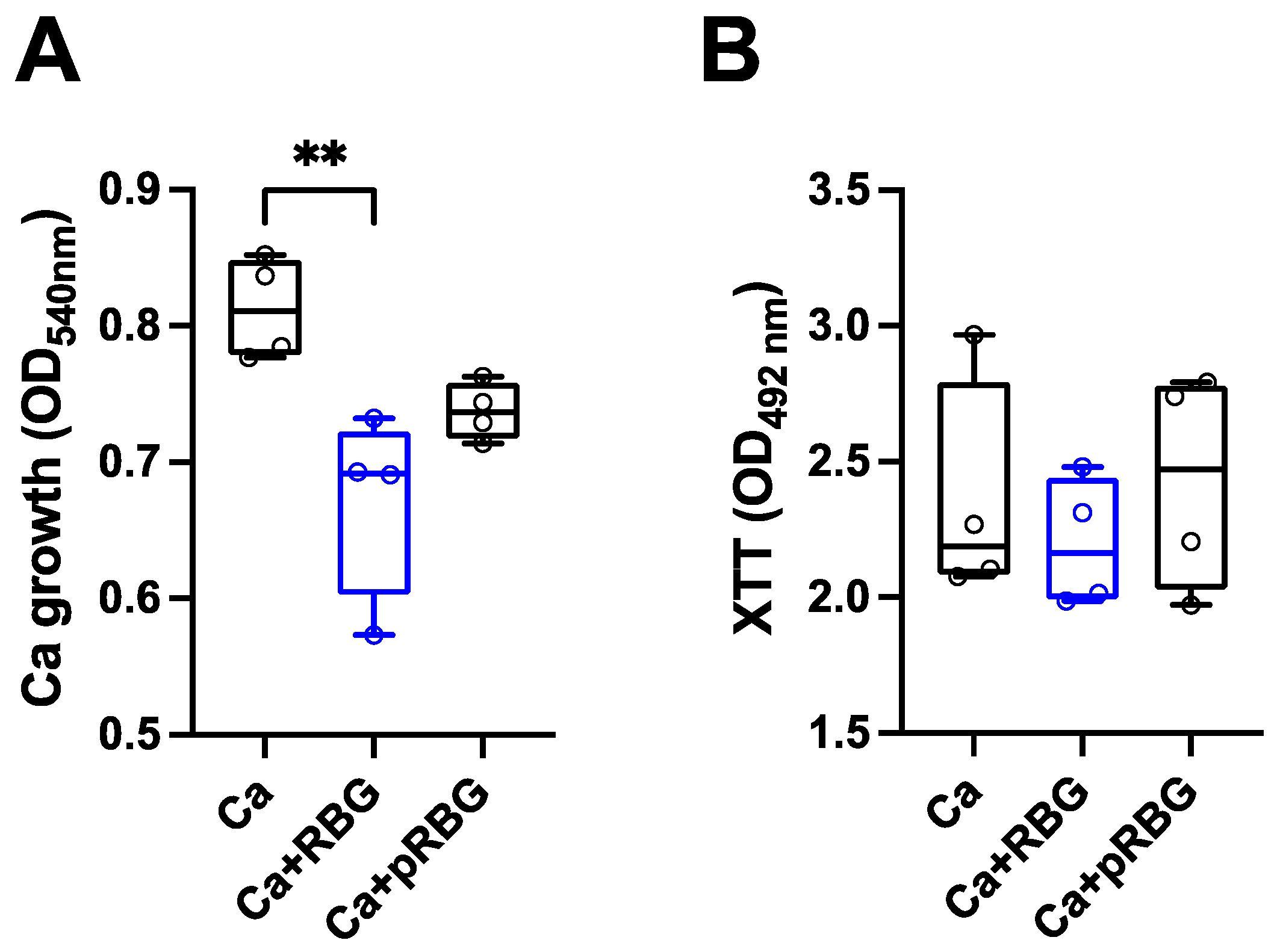
Figure 4.
RBG effect on C. albicans adhesion and hyphae formation. (A) Effect of RBG on C. albicans adhesion to abiotic surface after 2 h of incubation at 37°C in DMEM supplemented with 5% FBS. Data in the box-plots are from triplicate samples from 4 different experiments. (B) RBG effect on C. albicans hyphal formation after 4 h of incubation at 37°C in YPD liquid medium + 10% FBS. Data are expressed as the mean percentage of hyphal fragments SD from at least 3 analyzed fields for each condition from 3 different experiments. (C) Representative fields of RBG or pRBG effect on C. albicans hyphal formation as detailed above. Magnification 20 X. Scale bar.
Figure 4.
RBG effect on C. albicans adhesion and hyphae formation. (A) Effect of RBG on C. albicans adhesion to abiotic surface after 2 h of incubation at 37°C in DMEM supplemented with 5% FBS. Data in the box-plots are from triplicate samples from 4 different experiments. (B) RBG effect on C. albicans hyphal formation after 4 h of incubation at 37°C in YPD liquid medium + 10% FBS. Data are expressed as the mean percentage of hyphal fragments SD from at least 3 analyzed fields for each condition from 3 different experiments. (C) Representative fields of RBG or pRBG effect on C. albicans hyphal formation as detailed above. Magnification 20 X. Scale bar.
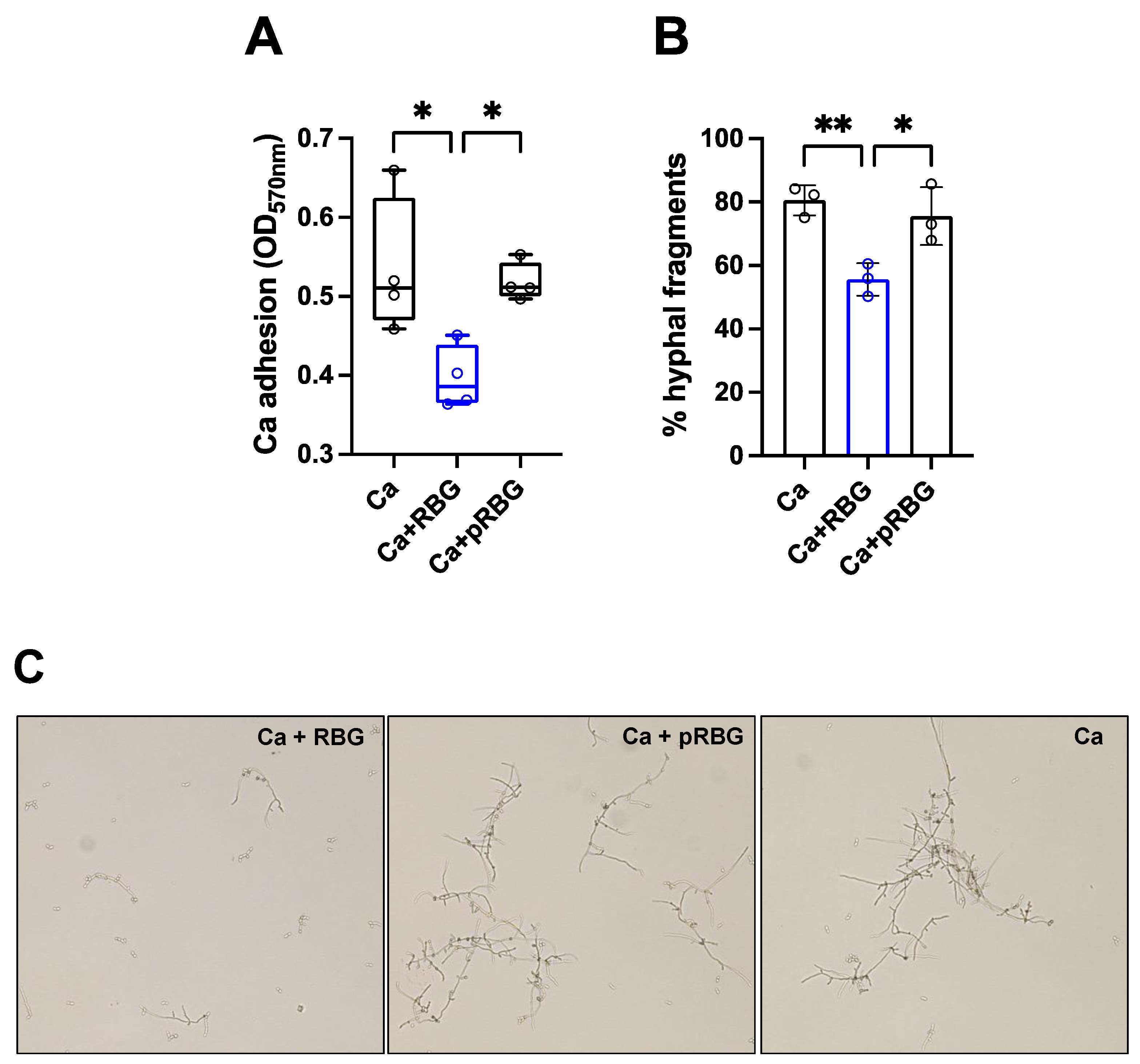
Figure 5.
Interleukins production. IL-1β (A) and IL-8 (B) production by vaginal epithelial cells after 24 h of C. albicans (Ca) infection or LPS treatment at 37°C plus 5% CO2 in the presence or absence of RBG, pRBG and dexamethasone (DEX). Violin-plot show the interleukins amounts released in the culture medium. Data are expressed as pg/ml of duplicate samples from at least 2 different experiments.
Figure 5.
Interleukins production. IL-1β (A) and IL-8 (B) production by vaginal epithelial cells after 24 h of C. albicans (Ca) infection or LPS treatment at 37°C plus 5% CO2 in the presence or absence of RBG, pRBG and dexamethasone (DEX). Violin-plot show the interleukins amounts released in the culture medium. Data are expressed as pg/ml of duplicate samples from at least 2 different experiments.
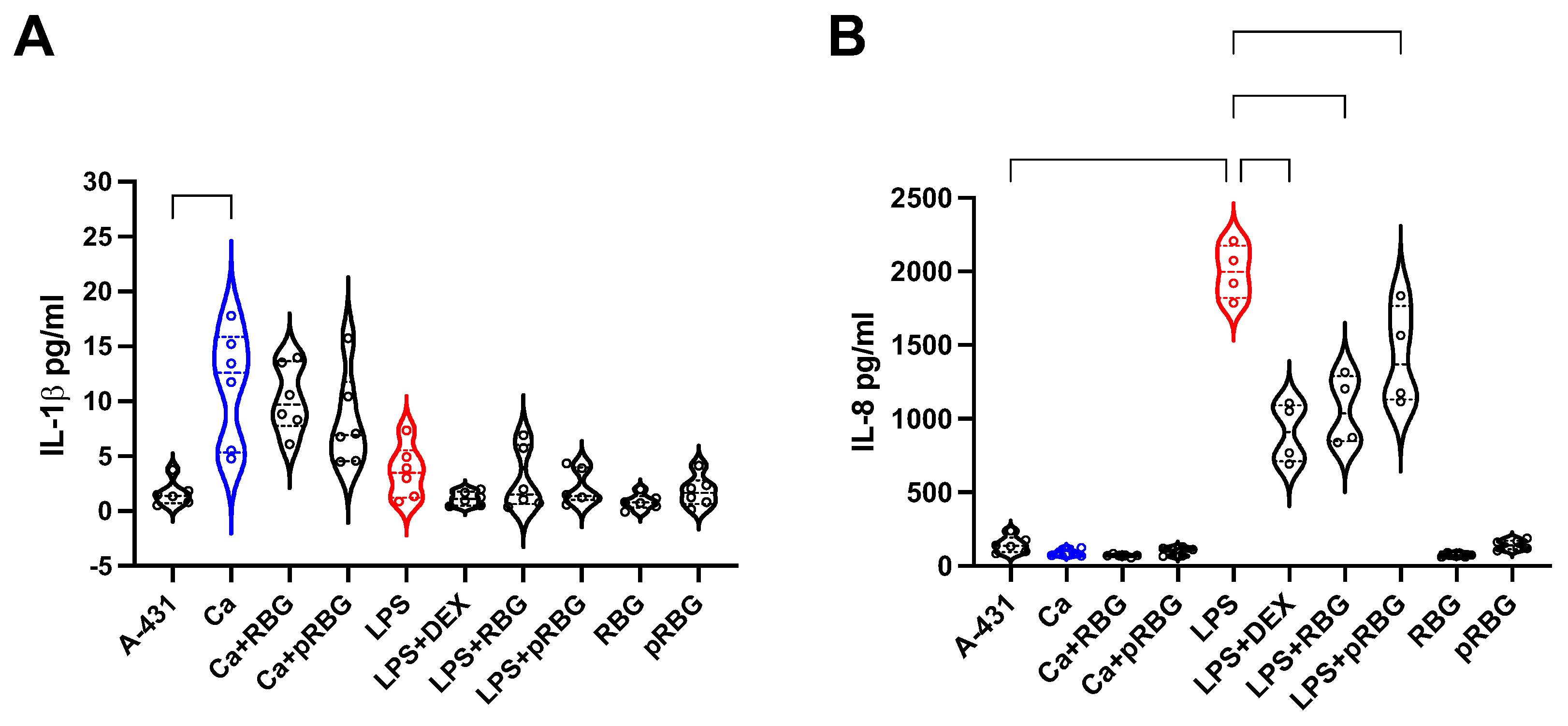
Figure 6.
Proposed model of RBG effects on C. albicans. Farnesol included in RBG counteracts yeast-to-hyphae transition and C. albicans adhesion without affecting Candida growth. This leads to a reduced capacity of C. albicans to damage vaginal epithelial cells. This scheme highlights the impact of farnesol without including the role of lactic acid, polydextrose and glycogen. However, the effects of such 3 components must be relevant in the actual application (left panel). pRBG is not able to modulate C. albicans morphological transition, adhesion, and growth. Hence, C. albicans can induce cell damage unchallenged (right panel).
Figure 6.
Proposed model of RBG effects on C. albicans. Farnesol included in RBG counteracts yeast-to-hyphae transition and C. albicans adhesion without affecting Candida growth. This leads to a reduced capacity of C. albicans to damage vaginal epithelial cells. This scheme highlights the impact of farnesol without including the role of lactic acid, polydextrose and glycogen. However, the effects of such 3 components must be relevant in the actual application (left panel). pRBG is not able to modulate C. albicans morphological transition, adhesion, and growth. Hence, C. albicans can induce cell damage unchallenged (right panel).
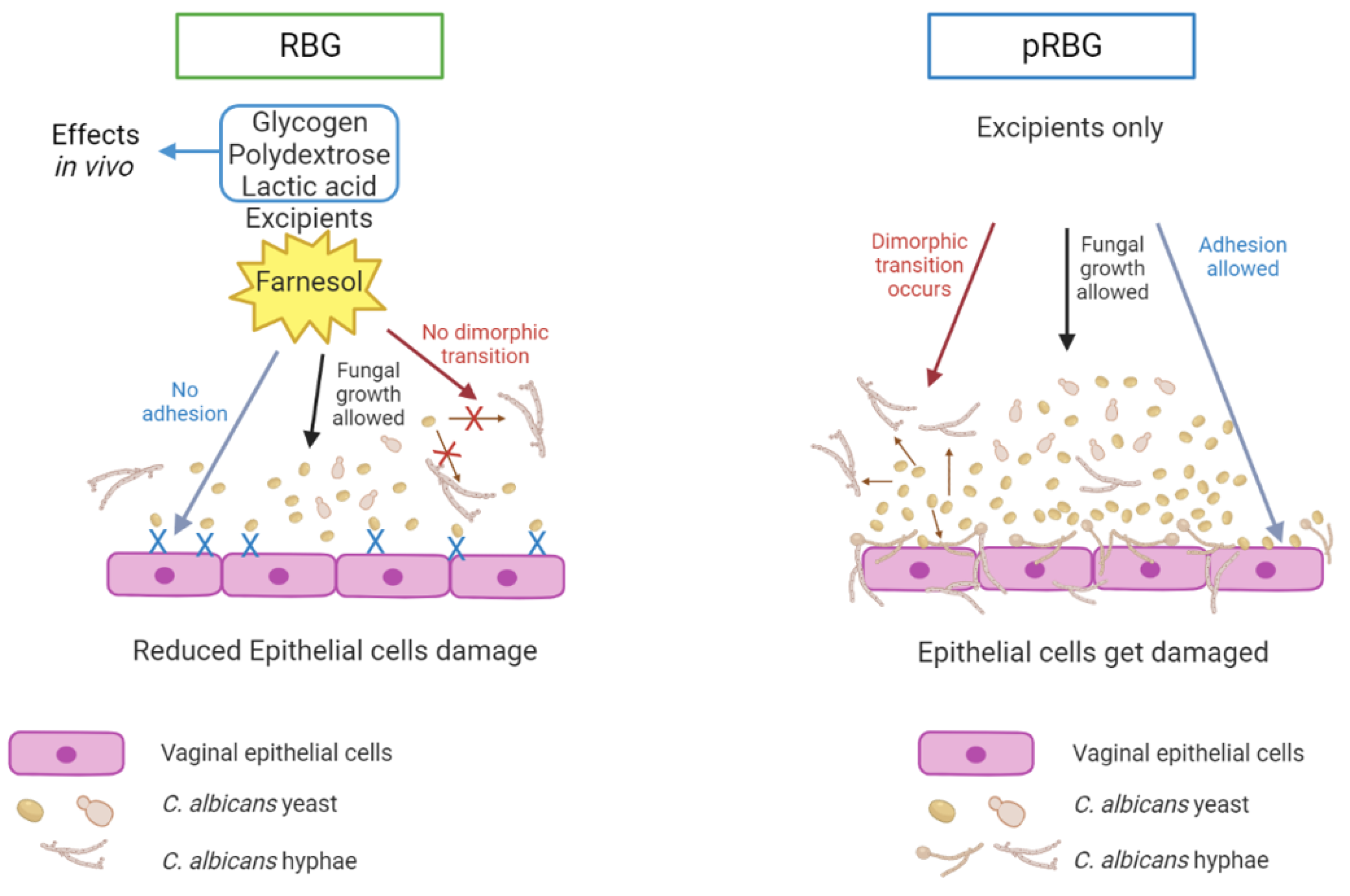

Table 1.
Composition of RBG and pRBG. Components identified as active compounds are written in bold blue.
Table 1.
Composition of RBG and pRBG. Components identified as active compounds are written in bold blue.
| RBG | pRBG |
| Water | Water |
| Disodium EDTA 0.2% Xanthan gum Sodium hyaluronate Propylene glycol Decylene glycol Hydroxyacetophenone Hydrogenated castor oil Tocopherol acetate PEG-40 Polydextrose Lactic acid Farnesol Glycogen |
Disodium EDTA 0.2% Xanthan gum Sodium hyaluronate Propylene glycol Decylene glycol Hydroxyacetophenone Hydrogenated castor oil Tocopherol acetate PEG-40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



