
Submitted:
01 May 2023
Posted:
02 May 2023
You are already at the latest version
Abstract
Climate change and exposure to environmental pollutants play a key role in the onset and aggravation of allergic diseases. As different climate-dependent patterns of molecular immunoglobulin E (IgE) reactivity have been regionally described, we sought to investigate the evolving allergen exposome in distinctive allergic phenotypes, and subtropical weather conditions through a Precision Allergy Molecular Diagnosis (PAMD@) model. Concurrent sensitization to several house dust mites (HDM) and storage mite molecules were broadly dominant in the investigated cohort, followed by the major cat allergen Fel d 1, and regardless of the basal allergic disease. Although a complex repertoire of allergens was recognized, a steadily increasing number of IgE binding molecules was associated with the complexity of the underlying atopic disease. Besides the highly prevalent IgE responses to HMD major allergens, Der p 21, Der p 5, and Der p 7 also showed as serodominant molecules, especially in those subjects bothered with asthma and atopic dermatitis. The accurate characterization of the external exposome at the molecular level and their putative role as clinically relevant allergens is essential to elucidate the phenotypic diversity of atopic disease in terms of personalized diagnosis and therapy.
Keywords:
Airborne allergens
; Exposome
; Climate change
; Atopy
; Allergic rhinitis
; Asthma
; Atopic dermatitis
1. Introduction
The definition of exposome was coined in 2005 to describe the totality of environmental exposures -from a variety of external and internal sources- that an individual experiences from conception throughout life [1,2]. More recently, the concept of meta-exposome refers to the bidirectional relationship of enviromental exposure on human health and the influence of human health on other living systems and genomes [3]. Along with other enviromental agents including chemical, biological, occupational and physical exposures, aeroallergens are considered part of the specific external exposome [4,5,6]. In fact, during the last decades a shap increase in the prevalence of atopic conditions and other immunomediated diseases has been assessed, turning into a relevant health and global economic burden [7,8]. In the Era of precision medicine, allergic respiratory and skin diseases can nowadays be grouped according to a combination of clinical, biological, and physiological features into specific pheno-endotypes, paving the road to targeted therapeutics [9,10].
Both epidemiological and environmental and data support that the interaction of climate change and air pollutants -including airborne allergens- play a major role in the pathobiology, evolution, clinical symptoms and long-term morbidity in human allergic disease [11,12]. Despite aeroallergens are regarded as risk factors to develop allergic disease, the relationship among allergen exposure, sentisitization and clinical symptoms is still a controversial issue [13,14].
The distribution of aeroallergens in a geographical area is highly affected by the local weather conditions, urbanization, and lifestyle, driving diverse types and degrees of specific IgE (sIgE) response dominating different areas of the world [15,16]. In fact, it has been shown that global warming favors warm and humid weather conditions, enhancing a suitable environment for the development of both indoor and outdoor allergens [17,18]. In this regard, as multiple allergens have been described capable of sensitizing and inducing clinical manifestations in genetically predisposed subjects, the accurate knowledge of the local allergen exposome turns out mandatory for the prevention, diagnosis, and treatment of allergic diseases [19,20].
Recently, the introduction of Precision Allergy Molecular Diagnosis (PAMD@) has remarkably improved the assay performance -especially in terms of sensitivity and analytic specificity- of a comprehensive assessment of the patient’s sIgE binding to a panel of individual allergens, changing our current understanding of sensitization profiles and potential cross-reactivity [21,22]. Along with these innovations in bioscience and technology, current guidelines, proposed that allergy diagnostic workup may follow a “top-down approach”, primarily from clinical symptoms to specific individual molecules [23]. In addition to former studies evaluating the local prevailing aeroallergens [24], we aimed to investigate -through a personalized PAMD@ model approach- the aeroallergen exposome and the concurrent sIgE reactivity in 3 clinical allergic phenotypes under subtropical climate conditions in Tenerife, Spain.
2. Results
2.1. Demographic Characteristics of Patients.
A total of 168 patients were screened, with 150 of them -75 females, mean age 31.8 yr. (range 8-67)- finally confirming their eligibility for the study (Figure 1) and divided into 3 different groups according to their underlying atopic disease (i.e., allergic rhinitis, allergic asthma, or atopic dermatitis).
All 150 subjects who fulfilled the correspondent ARIA, GINA, or SCORAD criteria for allergic rhinitis (AR), asthma (A), or atopic dermatitis (AD), [25,26,27] showed a positive skin prick test (SPT) to one or more of the local aeroallergens. Globally, most of the subjects (114 out of 150 patients, 76.0%) had their atopic disease onset during childhood or adolescence. All subjects were on regular daily treatment -comprising both allergen avoidance measures and conventional medical therapy- according to their allergic disease stage and severity. None of the patients were on former or current treatment with allergen immunotherapy or biologics upon inclusion in the current study. Four out 50 (8.0%) patients from the asthma cohort were on oral steroids upon inclusion in the study. Overall atopic comorbidities included 23 patients (15.33%) afflicted with food (seafood, nuts, egg and/or milk) or drug (7.33%) allergy (beta-lactam antibiotics and/or nonsteroidal anti-inflammatory drugs). Most patients (75.33%) reported a known family history of atopy.
2.2. Total IgE and blood eosinophils.
A quantitative analysis of serum total IgE was performed in order to evaluate the basal atopic status in the study population. The total IgE showed a median (range) value of 404 (41-19,993) IU/mL. Regarding basal allergic disease, a median (range) value of total IgE of 56 (23.88-1083) IU/mL was found for AR, 674 (56.5-19,993) IU/mL for A, and 898 (81.81-17,420) IU/mL for AD.
Blood eosinophils showed an overall median (range) value of 360 (20-1,880) eosinophils/μL, with a slightly higher median value (375 eosinophils/μL) in the AD group in respect to asthmatics (365 eosinophils/μL) or the AR (340 eosinophils/μL) subset (Table 1).
2.3. Prevalence, sIgE reactivity and individual molecular profile according to atopic disease.
Sensitization to aeroallergens extracts by SPT and the prevalence of 150 patients who met the inclusion criteria are summarized in Table 2.
One hundred and forty-six patients (97.33%) were independently sIgE positive (≥ 0.35 kUA/L) to one/or more of the individual molecular aeroallergens included in the multiplex array (Figure 2). Despite showing a positive SPT to one or more aeroallergen, 4 subjects (3 afflicted with A, and 1 with AD) had a sIgE ≤0.35 kUA/L to all tested allergens in the multiplex array.
2.4. Mites
Mites were identified as the most prevailing source of sensitizing airborne allergens in the present population regardless of the subjects’ atopic disease.
Sensitization to one or more out of the investigated 17 mite molecular allergens was found in 144 (96%) subjects. Six patients (1 with AR, 3 with A, and 2 AD) were not sensitized to any of the mentioned mite molecules. Although all 17 mite allergens were broadly represented throughout the investigated population, significant quantitative differences among molecules, and the involved atopic disease were confirmed.
Considering individual molecular allergens exclusively, Der f 2 was most frequently identified with sIgE ≥0.35 kUA/L in 126 out of 150 subjects (84%), Der p 2 in 125 patients (83.3%), followed Der p 23 (82%), Der p 1 (72.6%), Der f 1 (69.3%) and Der p 21 (66.6%). Regarding SM, Lep d 2 was most frequently found in 114 patients (76%), followed by Gly d 2 (67.33%), Blo t 5 (50.6%), Tyr p 2 (45.3%), and Blo t 21 (44%). Eighty-nine out of 150 (59.3%) individuals depicted one or more sIgE molecular response/s to Blo t 5, Blo t 21 and/or Blo t 10. Despite no quantitative differences (p=0.411) were depicted between the overall mean value of sIgE (kUA/L) against Der f 2 (25.67) and Der p 2 (24.12), significant differences (p<0.001) were found among both allergens and Lep d 2 (9.86), Gly d 2 (7.72), and Tyr p 2 (4.18). Significant differences (p=0.014) were also found between the overall mean values (kUA/L) of Der p 1 (14.9) and Der f 1 (9.79). A higher frequency of sIgE binding to HDM group 1 (87.03% Vs 66.66%) and 2 (88.88% Vs 79.16%), and Der p 23 (87.3% Vs 84.37%) was found in the younger subjects compared to their elder peers (≥20 yrs.).
Concerning individual mite molecules –Der f 2 (78%), Der p 2 (78%), Der p 23 (76%), Gly d 2 (74%), Lep d 2 (72%), Der p 1 (68%), Blo t 21 (54%) and Der f 1 (52%)- were considered serodominant in patients with AR. Blo t 5 was also detected in 46% patients of this group. Minor allergens -Der p 20, Der p 10, Blo t 10, and Der p 11- were identified in <15% of the investigated population.
Seven molecules reached a prevalence >50% -Der f 2, Der p 2, Der p 23, Lep d 2, Der p 1 and Der f 1- in those subjects with AR. An increasing number of allergens reaching serodominance was also confirmed for the asthma group -11 individual molecules, including Der p 5 (72%), Der p 7 (58%), and Blo t 5 (56%)- and for those afflicted with AD -12 allergens, including Tyr p 2 (50%). In addition to the large number sIgE mite responses, increasing quantitative differences -from AR to AD- were observed among the basal atopic disease and the corresponding mite molecular allergen (Table 3 and Figure 3).
Patients afflicted with AD showed significantly (P< 0.05) higher serum titers to 8 mite molecules -Der f 2, Der p 2, Der p 23, Der p 1, Der f 1, Der p 21, Der p 5 and Der p 7- than subjects with AR, and to 4 allergens -Der p 23, Der p 1, Der p 21 and Der p 7- compared to the group of asthmatics. Moreover, patients with asthma outlined significantly (P< 0.05) increased serum titres of Der p 2, Der f 2, and Der f 1 than individuals with AR. No significant differences were observed among AR, A or AD in the mean levels of sIgE to Lep d 2, Gly d 2, Blo t 5, Tyr p 2, Blo t 21, Der p 20, Der p 10, Blo t 10, and Der p 11 in the present population. In addition, >50% of patients with the A and/or AD phenotype showed a more complex aggregation pattern of molecules -including concurrent sensitization to ≥10 mite allergens- in contrast to the majority of subjects (64%) with AR displaying a concomitant sIgE response to <9 molecules (Table 4).
2.5. Cat and dog epithelia
Globally, 84 out of 150 patients were sensitized to one or more out of 10 investigated epithelial molecular allergens. Considering overall sIgE epithelial response, cat allergens were most frequently identified (43.3%) compared to dog molecules (35.3%) in the studied population.
A total of 65 out of 150 subjects showed different sIgE molecular responses to any of the following cat allergens: Fel d 1 (60.39%), Fel d 7 (19.8%), Fel d 4 (14.85%) or Fel d 2 (3.96%). In addition, a total of 53 out of 150 individuals displayed diverse molecular responses to dog allergens as follows, Can f 5 (46.23%), Can f 1 (22.58%), Can f 4 (20.43%), Can f 6 (15.05%), Can f 2 (8.6%), and Can f 3 (7.52%).
Fel d 1 exhibited as the most prevalent individual epithelial allergen in 19 out of 84 (22.61%) patients, followed by Can f 5 (15.47%). One patient (1.19%) showed a monomolecular sIgE response to Fel d 7. Forty-four subjects out of 150 (29.33%) were simultaneously co-sensitized to two or more cat and dog molecular allergens. In this regard, 84 subjects out of 150 showed a total of 31 heterogenous molecular responses to one or more of the epithelial allergens as indicated in Table 5.

Table 5.
Specific IgE profiles aggregated into selected cat and dog epithelial allergens in 84 out of 150 subjects tested with microarray. Profiles are ordered by the increasing number of recognized molecules. Asterisk (*) indicates specific IgE sensitization to a single cat and/or dog molecular allergen.
Table 5.
Specific IgE profiles aggregated into selected cat and dog epithelial allergens in 84 out of 150 subjects tested with microarray. Profiles are ordered by the increasing number of recognized molecules. Asterisk (*) indicates specific IgE sensitization to a single cat and/or dog molecular allergen.
| n=84 | % | Number of molecules | Fel d 1 | Fel d 2 | Fel d 4 | Fel d 7 | Can f 1 | Can f 2 | Can f 3 | Can f 4 | Can f 5 | Can f 6 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 19 | 22.6 | 1 | * | |||||||||
| 1 | 1.1 | 1 | * | |||||||||
| 13 | 15.4 | 1 | * | |||||||||
| 2 | 2.3 | 2 | * | * | ||||||||
| 11 | 13.0 | 2 | * | * | ||||||||
| 3 | 3.5 | 2 | * | * | ||||||||
| 4 | 4.7 | 2 | * | * | ||||||||
| 1 | 1.1 | 3 | * | * | * | |||||||
| 1 | 1.1 | 3 | * | * | * | |||||||
| 1 | 1.1 | 3 | * | * | * | |||||||
| 2 | 2.3 | 3 | * | * | * | |||||||
| 1 | 1.1 | 3 | * | * | * | |||||||
| 2 | 2.3 | 3 | * | * | * | |||||||
| 1 | 1.1 | 3 | * | * | * | |||||||
| 1 | 1.1 | 3 | * | * | * | |||||||
| 3 | 3.5 | 4 | * | * | * | * | ||||||
| 1 | 1.1 | 4 | * | * | * | * | ||||||
| 1 | 1.1 | 4 | * | * | * | * | ||||||
| 2 | 2.3 | 4 | * | * | * | * | ||||||
| 1 | 1.1 | 4 | * | * | * | * | ||||||
| 1 | 1.1 | 4 | * | * | * | * | ||||||
| 1 | 1.1 | 5 | * | * | * | * | * | |||||
| 1 | 1.1 | 5 | * | * | * | * | * | |||||
| 1 | 1.1 | 6 | * | * | * | * | * | * | ||||
| 1 | 1.1 | 6 | * | * | * | * | * | * | ||||
| 1 | 1.1 | 7 | * | * | * | * | * | * | * | |||
| 1 | 1.1 | 7 | * | * | * | * | * | * | * | |||
| 2 | 2.3 | 8 | * | * | * | * | * | * | * | * | ||
| 1 | 1.5 | 8 | * | * | * | * | * | * | * | * | ||
| 1 | 1.1 | 8 | * | * | * | * | * | * | * | * | ||
| 2 | 2.3 | 9 | * | * | * | * | * | * | * | * | * |
Individuals with AD showed significantly (P< 0.05) higher serum titers to 6 epithelium molecules -Fel d 1, Fer d 7, Can f 1, Can f 2, Can f 4, and Can f 6- than subjects with AR, and to 4 allergens -Fel d 1, Can f 1, Can f 2, and Can f 4- compared to the group of patients with the asthma phenotype. No quantitative significant differences were observed among AR, A or AD in the mean levels of Fel d 2, Fel d 4, Can f 3, or Can f 5 in the studied population. Interestingly, 82 out of 84 subjects (97.61%) sensitized to epithelial molecules, showed a concurrent sIgE response to one or more of the 17 assessed mite molecular allergens.
2.6. Pollen
A total of 64 subjects out of 150 (42.6%) showed a pattern of 31 different molecular responses to one or more of the following pollen allergens: Cyn d 1 (33.7%), Par j 2 (20.22%), Lol p 1 (17.97%), Art v 1 (13.48%), Cup a 1 (12.38%), Phl p 2 (10.11%), Pla a 3 (7.86%), Phl p 1 (5.61%), Sal k 1, Ole e 1, and Bet v 6 (3.37%); Phl 5, Phl p 12, Art v 3, Bet v 2 and Pla a 1 (2.24%) and Phl p 6, Phl p 7, Pla a 2, and Bet v 1 (1.12%). No identification of Ole e 7 or Ole e 9 was spotted in the studied population (Table 5).
Table 5.
Specific IgE profiles aggregated into selected pollen allergens in 64 out of 150 subjects tested with microarray. Profiles are ordered by the increasing number of recognized molecules. Asterisk (*) indicates specific IgE sensitization to a single pollen molecular allergen.
Table 5.
Specific IgE profiles aggregated into selected pollen allergens in 64 out of 150 subjects tested with microarray. Profiles are ordered by the increasing number of recognized molecules. Asterisk (*) indicates specific IgE sensitization to a single pollen molecular allergen.
| n=64 | % | Number of molecules | Cyn d 1 | Par j 2 | Phl p 1 | Phl p 2 | Phl p 12 | Lol p 1 | Art v 1 | Art v 3 | Cup a 1 | Sal k 1 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 | 4.6 | 1 | * | |||||||||
| 3 | 4.6 | 1 | * | |||||||||
| 2 | 3.1 | 1 | * | |||||||||
| 3 | 4.6 | 1 | * | |||||||||
| 17 | 26.5 | 1 | * | |||||||||
| 2 | 3.1 | 1 | * | |||||||||
| 3 | 4.6 | 1 | * | |||||||||
| 3 | 4.6 | 2 | * | * | ||||||||
| 1 | 1.5 | 2 | * | * | ||||||||
| 1 | 1.5 | 2 | * | * | ||||||||
| 1 | 1.5 | 2 | * | * | ||||||||
| 1 | 1.5 | 2 | * | * | ||||||||
| 2 | 3.1 | 2 | * | * | ||||||||
| 1 | 1.5 | 2 | * | * | ||||||||
| 2 | 3.1 | 2 | * | * | ||||||||
| 1 | 1.5 | 2 | * | * | ||||||||
| 1 | 1.5 | 2 | * | * | ||||||||
| 1 | 1.5 | 2 | * | * | ||||||||
| 1 | 1.5 | 2 | * | * | ||||||||
| 1 | 1.5 | 3 | * | * | * | |||||||
| 2 | 3.1 | 3 | * | * | * | |||||||
| 1 | 1.5 | 3 | * | * | * | |||||||
| 1 | 1.5 | 3 | * | * | * | |||||||
| 1 | 1.5 | 3 | * | * | * | |||||||
| 1 | 1.5 | 3 | * | * | * | |||||||
| 3 | 4.6 | 4 | * | * | * | * | ||||||
| 1 | 1.5 | 4 | * | * | * | * | ||||||
| 1 | 1.5 | 4 | * | * | * | * | ||||||
| 1 | 1.5 | 4 | * | * | * | * | ||||||
| 1 | 1.5 | 4 | * | * | * | * | ||||||
| 1 | 1.5 | 5 | * | * | * | * | * |
Twenty-eight subjects out of 64 were sensitized to a single molecular pollen allergen (43.75%). Cyn d 1 was confirmed as the most prevalent individual pollen allergen in 17 out of 64 (26.56%) patients, followed by Par j 2, Art v 1 and Cup a 1 (4.68% in all 3 cases) and Phe p 1 (3.12%). Most individuals (56.25%) with a pollen sensitization displayed a concurrent response to 2 or more molecules. No quantitative significant differences were observed among AR, A or AD in the mean levels of sIgE to 22 out 23 investigated pollen allergens. Only Cup a 1 showed significant (p=0.0307) higher sIgE values in patients with AD compared to those with AR.
No solely sensitization to pollen allergens was found in the present population. Sixty-three out of 64 subjects (98.43%) sensitized to any of the tested pollen molecules, showed a coincident sIgE response to one or more of the 17 investigated mite allergens and the remaining subject -afflicted with AD- showed a complex pollen polysensitization pattern including Cup a 1, Pla a 2, Phl p 1, Lol p 1, Cyn d 1, and Art v 1 with epithelial allergens (Fel d 1).
2.7. Mold and cockroach allergens
Regarding molds, Alt a 1 and Alt a 6 were rarely identified, in only 11 (7.3%) and 1 (0.6%) out of 150 studied subjects, respectively. No solely sensitization to Alt a 1 or Alt a 6 was found in the current study population. All 12 (100%) subjects with a sensitization to Alt a 1 and/or Alt a 6 showed a concurrent sIgE response to one or more of the 17 mite allergens.
Cockroach allergens were also scarcely depicted as Bla g 9 was only present in 10 individuals (6.6%), followed by Per a 7 in 8 individuals (5.3%), and Bla g 4 in 1 participant (0.6%). No sIgE response to Bla g 1, Bla g 2 or Bla g 5 was detected in the present population. All 18 (100%) sIgE responses to Bla g 9 and/or Per a 7 showed a concomitant polysensitization to one or more of the 17 included mite allergens.
3. Discussion
Climate change is already having an impact on the seasonality, production, and concentration, allergenicity, and geographic dissemination of airborne allergens, with subsequent consequences on human allergic disease [28,29]. Tenerife is the largest and highest oceanic island of the Canary archipelago with a climate dominated by the influence of the cool humid northeast trade winds, associated with the Azores anticyclone [30]. Despite the local levels of air pollutants are rather low, compared to continental regions of Europe, and due to the proximity of the Canary Islands to the African continent, Saharan Dust Intrusions -with PM10 concentrations higher than 50 µg/m3- have been independently related to affect human health [31].
In the present study, the assessment of aeroallergen sensitization as a core biomarker for the classification of allergic disease [32], has consistently confirmed mites as the most prevalent allergen source affecting the local population. In fact, 96% of the investigated subjects confirmed a sensitization to ≥1 individual mite molecule/s, followed by a 56% sensitized to ≥1 epithelial allergen/s, 42.66% of patients with a sIgE response to ≥1 pollen molecule/s and finally 7.3% and 6.66% sensitized to molds or cockroach, respectively.
To date, more than 30 House Dust Mites (HDM) allergens have been described, with group 1 and 2 allergens leading the sIgE response in HDM sensitized individuals, globally []. Regarding the overall prevalence of individual allergens, in the present investigation, 11 molecules were considered as serodominant (>50%) allergens in the studied population. []. Local subtropical conditions favoring an increased perennial exposure to both HDM, and Storage Mites (SM) may explain a higher prevalence of mite sIgE responses in the studied population, compared to those results reported from Australia (77%) and Singapore (63%) [33,34].
Interestingly, despite these highly prevalent allergens induce strong IgE responses, differences in the IgE antibody binding frequency have been addressed in both dissimilar and alike populations depending on the applied multiplex measurement platform [35,36]. In this regard, despite former local specific allergen profile identified identical prevalence rates for Der f 1, Der p 2 and Der f 2 -using the ADVIA Centaur ® platform- Der f 2, and Der p 2 have repeatedly exhibited a higher sIgE binding frequency compared to group 1 HDM allergens with the multiplex ALEX2 array [37,38,39]. Such variations, according to the implemented multiplex assay in the overall prevalence of these representative HDM allergens have also been addressed in former research from Spain, China and Eastern Europe [40,41,42,43].
After HDM group 2 allergens, Der p 23 was identified as the third most frequent (82%) molecule in the studied population. Despite the high prevalence of sIgE binding to Der p 23, only one individual (0.66%) afflicted with AR, showed a single sensitization to this molecule. In addition, our findings confirmed a prominent role for the so-called mid-tier HDM allergens -Der p 21, Der p 5, and Der p 7- as serodominant molecules in the studied population, especially in those subjects with A and AD. Moreover, in accordance with previously documented low cross reactivity between group 5 allergens [44,45], molecular sensitization to Der p 5 (64.0%) was significantly (p<0.05) increased compared to Blo t 5 (50.6%) across the study cohort, including both respiratory and AD subjects. Also, in line with our former observations, minor allergens such tropomyosin and Der p 11 were scarcely represented in 10.6% and 2% respectively including those patients affected with AD [46].
Although still under debate, it has been speculated that the climate change in the last decades might affect mite biology and that new allergens may become influential as sensitizing molecules [47]. Differently from B. tropicalis, that has been previously identified in nearly 24% in dust samples from local mattresses, L. destructor, G. domesticus and T. putrescetiae were all scarcely detected in less than 6% of the studied samples before 2009 [48]. Interestingly, Lep de 2 was the fourth -after Der f 2, Der p 2 and Der p 23- most frequently (76%) identified allergen in the current investigation, with more than 85% of these individuals concurrently sensitized to Gly d 2 and/or Tyr p 2 -supporting a relevant IgE-cross-reactivity among molecules- [49] and a potential role as emerging allergen sources in this area.
Remarkably, epithelial allergens were confirmed -after HDM and SM- as the second most prevalent sensitization (56.0%) in our territory, with major allergens Fel d 1 (22.61%) and Can f 5 (15.47%) as leading individual molecules. Furthermore, as previous research has elegantly shown that structurally unrelated epithelial and mite allergens can activate epithelial cells through adjuvant-like protease-independent mechanisms [50], a coincident polysensitization pattern among epithelial and mite molecules was widely identified (97.61%), contributing to depict the complexity of the indoor exposome affecting the current population. Moreover, perennial allergen sensitization has been related to an increased rate of asthma morbidity, including asthma exacerbations, hospital visits and medication needs [51,52].
Pollen showed as the third most frequently (42.6%) identified molecules with an unexpected -not previously reported in our area- 33.7% of subjects showing a sensitization to Cyn d 1, followed by Par j 2 (20.22%), Lol p 1 (17.97%), Art v 1 (13.48%), and Cup a 1 (12.38%). Concurrent sIgE responses to mite and/or epithelial allergens was found in all (100%) subjects with a pollen sensitization, giving insight into a coexisting local exposure to both perennial and seasonal molecules. Besides the exposure to pollen from the local vegetation, the island of Tenerife is also influenced by extra-regional pollen transport episodes from the Mediterranean region, the Saharan sector and the Sahel enclosing both trees and herbs pollen [53]. Despite most pollen transport occurs within the island; pollen peaks may be markedly increased during specific meteorological situations driving air masses from areas where the plants providing the depicted pollen are present. In this regard despite, it has been estimated that up to 97% of annual counts of Olea pollen in Tenerife originates from extra-regional sources [54], Ole e 1 was identified in less than 4% of the present cohort.
Several clinical consequences may be outlined form this investigation. Firstly, the proposed panel accurately identified (≥98.0%) the molecular profile in patients from the same region but affected with distinct atopic diseases. Secondly, and in line with previous reports [55,56], we confirmed an increased number of sIgE binding molecules associated with the complexity of the underlying atopic phenotype. Moreover, significant (p<0.05) quantitative higher titers were gradually found to major and mid-tier allergens in subjects with AD against those with A or AR, and in the group of asthmatics in respect to those with only AR. Transcutaneous sensitization to these allergens has been previously related to the pathogenesis of AD, which manifests with a genetically predetermined skin barrier defect, leading to an overexpression of pro-inflammatory cytokines, and the subsequent activation of innate and adaptative immune responses [57] Also, as previously reported, a higher frequency of sIgE binding -especially to mite molecules- was found in the younger participants of the study, regardless of their underlying allergic condition [58,59,60].
A few limitations should be mentioned in the current study as 4 out of 150 individuals (2.66%) -3 asthmatics and 1 with AD- with a positive SPT to local aeroallergens could not be identified by the multiplex array, and a restricted number of subjects were finally included in the cross-sectional analysis.
The identification of these highly prevalent molecules remains mandatory not only to describe part of the local external exposome, but also to investigate their role as clinically relevant allergen sources and risk assessment in different allergic phenotypes [61]. In fact, the clinical relevance of only a few molecules presents in the current assay -Der p 1, Der p 2, Der p 5, Blo t 5, Per a 10, Alt a 1, Fel d 1, Can f 1, Can f 2, Can f 3, Can f 4 and Can f 5- has been intrinsically proved, moving away from the traditional assumption that only allergens with high (>50%) sIgE binding frequencies are relevant [62].
4. Materials and Methods
4.1. Subjects
We consecutively recruited children and adult patients with an Allergist-confirmed diagnosis of atopic disease -i.e. allergic rhinitis (AR), asthma (A), and/or atopic dermatitis (AD)- from the Outpatient Allergy Clinic and Severe Asthma Unit at Hospital Universitario de Canarias (Tenerife, Spain), from December 2021 to January 2023. The present investigation was previously evaluated and authorized by the domestic Ethical Committee and the corresponding informed consent documents were properly signed by all participants -and parents/guardians for those participants <18 y.o.- upon inclusion in the study. Onset of referred clinical symptoms only after a minimum of 3 years of local residency and was also required to meet the inclusion criteria in the present investigation. Severity and staging of allergic diseases were also clinically evaluated according to specific Guidelines [25,26,27].
The following clinical data were collected from the patients’ medical records: Sociodemographic data, clinical profile including past medical conditions and current allergy diagnosis, and the characteristics of the associated medication, forced expiratory volume in the first second (FEV1), a validated Asthma Control Test (ACT), the SCORAD index, and SPT results. Following routine clinical practise, only subjects with a positive SPT and/or a specific sIgE to the corresponding aeroallergen extract -mites, pollens, molds, and/or fur animals’ epithelium- were included in the study. Patients under treatment with past or current allergen immunotherapy or monoclonal antibodies -biologics- were excluded. Pregnant and breast-feeding women were also excluded from the study.
4.2. Skin Prick Test.
Percutaneous testing was carried out according to European standards [63], enclosing a diagnostic panel (Inmunotek, Madrid, Spain) with standardized raw extracts (Dermatophagoides pteronyssinus (D. pteronyssinus), Blomia tropicalis (B. tropicalis), Lepidoglyphus destructor (L. destructor), Tyrophagus putrescentiae (T. putrescentiae), cat and dog dander, grass mix (Poa pratensis, Dactilis glomerata, Lolium perenne, Phleum pratense and Festuca pratensis), olive, Parietaria judaica, Artemisa vulgaris, Alternaria alternata, Aspergillus fumigatus, Cladosporium herbarum and Blatella. Histamine (10 mg/ml) and saline were used as positive and negative controls as usually. Antihistamines were withdrawn a week before the SPT, and wheal diameters were immediately measured after 20 minutes with those diameters greater than 3 mm regarded as positive.
4.3. Mite allergenic extracts.
Proteins from mite bodies of D. pteronyssinus, B. tropicalis, and L. destructor were manufactured using standardized internal protocols (Inmunotek S.L., Madrid, Spain). The extracts were prepared extracting this material in 0.01 M phosphate-buffered saline buffer (PBS; 1/5 wt:vol) , pH 7.4, for 4-6 hours at 5±3°C under magnetic starring. Then, the extracts were centrifugated at 16,000 x g for 30 min at 4°C. Afterwards, the supernatants were recollected. The pellet was reconstituted and extracted in 0.01 M PBS (1/5 wt:vol) overnight at 5±3°C. Then, the extract was centrifugated again and the supernatant separated from pellet. Both supernatants were mixed and subsequent dialysis by tangential ultrafiltration against highly purified water, using Omega polyethersulfone membranes (TFF Cassette T series, Pall Life Sciences) with a pore size of 100 kDa. Finally, native extracts were frozen, and lyophilized. The protein content was measured by Bradford method.
4.4. SDS PAGE and IgE Western Blot.
Proteins from D. pteronyssinus B. tropicalis, and L. destructor extracts were separated by 12% polyacrylamide gel with sodium dodecylsulfate (SDS-PAGE) under reducing conditions according to Laemmli´s method [64]. Proteins were visualized when gel was stained with GelCode Blue stain reagent (Life Technologies, Carlsbad, CA, USA For the Western Blot, proteins from the gel electrophoresis were electrotransferred to nitrocellulose membranes 0,45µm (Bio-Rad) and the binding of IgE antibody to allergens was analyzed using individual patients´ sera and anti-human IgE peroxidase conjugate (Southern Biotech, Birmingham, USA). Chemiluminescence detection reagents (Western Lightning® Plus-ECL. Perkin Elmer. Waltham, MA, USA) were added following the manufacturer’s instructions and image was analyzed in Image Lab Touch software 3.0.1.14. IgE binding bands were identified using the BioRad Diversity database program.
4.5. Serological analysis.
Blood samples were obtained from all participating individuals, identified with a code label, stored at -40° C and thawed immediately prior to the in vitro assay. Total IgE levels, and sIgE were measured (ALEX MacroArray Diagnostics, Vienna, Austria) according to the manufacturer´s instructions in all included subjects. In brief, ALEX is a multiplex array containing 282 reagents (157 whole allergens and 125 molecular components). The different allergens and components are coupled onto polystyrene nano-beads, and then the allergen beads are deposited on a nitrocellulose membrane, as formerly published [65]. Total IgE levels were expressed in international units per unit volume (IU/mL), sIgE levels were expressed in kUA/L. Values ≥0.35 kUA/L were considered positive. A total of 17 mite molecular allergens were included: Der p 1, Der p 2, Der p 5, Der p 7, Der p 10, Der p 11, Der p 20, Der p 21, Der p23, Der f 1, Der f 2, Blo t 5, Blot 10, Blo t 21, Lep d 2, Gly d 2, and Tyr p 2. A total of 10 cat and dog epithelial molecular allergens were included: Fel d 1, Fel d 2, Fel d 4, fel d 7, Can f 1, Can f 2, Can f 3, Can f 4, Can f 5, and Can f 6. A total of 23 pollen allergens included: Bet v 1, Bet v 2, Bet v 6, Cup a 1, Pla a 1, Pla a 2, Pla a 3, Ole e 1, Ole e 7, Ole e 9, Phl p 1, Phl p 2, Phl p 5, Phl p 6, Phl p 7, Phl p 12, Lol p 1, Cyn d 1, Sal k 1, Pla l 1, Par j 2, Art v 1, and Art v 3. Alt a 1 and Alt a 6 were investigated for mould sensitization while Bla g 1, Bla g 2, Bla g 4, Bla g 5, Bla g 9 and Per a 7 were designed to evaluate cockroach sensitization.
4.6. Statistical analysis.
Demographic features were summarized by medians and standard deviations for continuous variables and percentages for categorical variables. To compare differences analysis of variance, Kruskal-Wallis, Mann-Whitney U and Chi-square tests are required for parametric continuous, nonparametric continuous, and categorical variables respectively. A P value of less than 0.05 was considered statistically significant. All statistical data were analysed using GraphPad Prism version 8.0.0 for Windows, GraphPad Software, La Jolla California USA.
5. Conclusion
The present study is, to the best of our knowledge, the first to investigate a real-life global molecular response to a comprehensive panel of aeroallergens in our local territory, in subjects afflicted with different atopic conditions. The outlined PAMD@ panel performed as a sensitive and highly specific tool to a precise molecular diagnosis in allergic patients strongly influenced by a complex external exposome.
Despite our results confirmed an increasingly pleomorphic profile of sIgE binding molecules in connection with the complexity of the allergic phenotype, no single molecular allergen could be identified as a surrogate marker for any of the studied atopic conditions. In relation to the aggregation of molecules and evolving profiles, concurrent polysensitization to unrelated indoor allergenic sources -especially mites and epithelia- were broadly predominant, making it challenging to propose an individualized treatment -i.e., allergen immunotherapy- for most patients.
Finally, as climate change impacts on airborne allergens in terms of allergenicity, seasonality, production, and atmospheric concentration -leading to subsequent outcomes in human health- the identification of the local external exposome stands out to a personalized diagnosis and therapy in clinically distinctive allergic phenotypes.
Author Contributions
Conceptualization, RG-P, PP-G and FP; methodology, RG-P, PP-G and FP; software, FP, TG, EA-F and MJ-M; validation and formal analysis, IS-M, FP; investigation, IS-M, EM-L, RG-P, PP-G, TG, EA-F, MJ-M and FP; resources, IS-M, PF, FP, MC; data curation, RG-P, PP-G, TG, EA-F, EM-L, MJ-M and FP; writing—original draft preparation, RG-P and PP-G; writing—review and editing, RG-P, PP-G and FP; project administration RG-P, PP-G and IS-M; funding acquisition RG-P, PP-G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fundación Canaria Instituto de Investigación Sanitaria de Canarias (FIISC), Servicio Canario de Salud, grant number OA17/042.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Institutional Ethics Committee of CEIC Hospital Universitario de Canarias, Tenerife, Spain with the reference number P.I.-2017/72 on October 30, 2017.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data that support the findings of this study are available from Servicio Canario de Salud but restrictions apply to the availability of these data, which were used under license for the current study, and so are not publicly available. Data are however available from the authors upon reasonable request and with permission of Servicio Canario de Salud.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Wild, C.P. Complementing the genome with an "exposome": the outstanding challenge of environmental exposure measurement in molecular epidemiology. Cancer Epidemiol Biomarkers Prev. 2005, 14, 1847–1850. [Google Scholar] [CrossRef] [PubMed]
- Wild, C.P. The exposome: from concept to utility. Int J Epidemiol. 2012, 41, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Renz, H.; Holt, P.G.; Inouye, M.; Logan, A.C.; Prescott, S.L.; Sly, P.D. An exposome perspective: early-life events and immune development in a changing world. J Allergy Clin Immunol. 2017, 140, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Celebi Sozener, Z.; Ozdel Ozturk, B.; Cerci, P.; Turk, M.; Gorgulu Akin, B.; Akdis, M.; et al. Epithelial barrier hypothesis: Effect of the external exposome on the microbiome and epithelial barriers in allergic disease. Allergy 2022, 77, 1418–1449. [Google Scholar] [CrossRef] [PubMed]
- Custovic, A. To what extent is allergen exposure a risk factor for the development of allergic disease? Clin Exp Allergy 2015, 45, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Putcha, N.; Woo, H.; McCormack, M.C.; Fawzy, A.; Romero, K.; Davis, M.F.; et al. Home Dust Allergen Exposure Is Associated with Outcomes among Sensitized Individuals with Chronic Obstructive Pulmonary Disease. Am J Respir Crit Care Med. 2022, 205, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Ellwood, P.; Asher, M.I.; Billo, N.E.; Bissell, K.; Chiang, C.Y.; Ellwood, E.M.; et al. The Global Asthma Network rationale and methods for phase I global surveillance: prevalence, severity, management and risk factors. Eur Respir J. 2017, 49, 1601605. [Google Scholar] [CrossRef] [PubMed]
- Pawankar, R. Allergic diseases and asthma: a global public health concern and a call to action. World Allergy Organ J. 2014, 7, 12. [Google Scholar] [CrossRef]
- Kuruvilla, M.E.; Lee, F.E.; Lee, G.B. Understanding Asthma Phenotypes, Endotypes, and Mechanisms of Disease. Clin Rev Allergy Immunol. 2019, 56, 219–233. [Google Scholar] [CrossRef]
- Kim, J.; Ahn, K. Atopic dermatitis endotypes: knowledge for personalized medicine. Curr Opin Allergy Clin Immunol. 2022, 22, 153–159. [Google Scholar] [CrossRef]
- Perera, F.; Nadeau, K. Climate Change, Fossil-Fuel Pollution, and Children's Health. N Engl J Med. 2022, 386, 2303–2314. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Khatri, S.B. The Exposome and Asthma. Clin Chest Med. 2019, 40, 107–23. [Google Scholar]
- Huang, C.F.; Chie, W.C.; Wang, I.J. Effect of environmental exposures on allergen sensitization and the development of childhood allergic diseases: A large-scale population-based study. World Allergy Organ J. 2021, 14, 100495. [Google Scholar] [CrossRef] [PubMed]
- Schoos, A.M.; Chawes, B.L.; Jelding-Dannemand, E.; Elfman, L.B.; Bisgaard, H. Early indoor aeroallergen exposure is not associated with development of sensitization or allergic rhinitis in high-risk children. Allergy 2016, 71, 684–691. [Google Scholar] [CrossRef] [PubMed]
- Weghofer, M.; Thomas, W.R.; Kronqvist, M.; Mari, A.; Purohit, A.; Pauli, G.; et al. Variability of IgE reactivity profiles among European mite allergic patients. Eur J Clin Invest. 2008, 38, 959–965. [Google Scholar] [CrossRef] [PubMed]
- Stemeseder, T.; Schweidler, B.; Doppler, P.; Klinglmayr, E.; Moser, S.; Lueftenegger, L.; et al. Exposure to Indoor Allergens in Different Residential Settings and Its Influence on IgE Sensitization in a Geographically Confined Austrian Cohort. PLoS One 2017, 12, e0168686. [Google Scholar] [CrossRef]
- D'Amato, G.; Chong-Neto, H.J.; Monge Ortega, O.P.; Vitale, C.; Ansotegui, I.; Rosario, N.; et al. The effects of climate change on respiratory allergy and asthma induced by pollen and mold allergens. Allergy 2020, 75, 2219–2228. [Google Scholar] [CrossRef]
- Paudel, B.; Chu, T.; Chen, M.; Sampath, V.; Prunicki, M.; Nadeau, K.C. Increased duration of pollen and mold exposure are linked to climate change. Sci Rep. 2021, 11, 12816. [Google Scholar] [CrossRef]
- Miller, J.D. The Role of Dust Mites in Allergy. Clin Rev Allergy Immunol. 2019, 57, 312–329. [Google Scholar] [CrossRef]
- Andiappan, AK; Puan, KJ; Lee, B; Nardin, A; Poidinger, M; Connolly, J; et al. Allergic airway diseases in a tropical urban environment are driven by dominant mono-specific sensitization against house dust mites. Allergy 2014, 69, 501–509. [Google Scholar]
- Anto, J.M.; Bousquet, J.; Akdis, M.; Auffray, C.; Keil, T.; Momas, I.; et al. Mechanisms of the Development of Allergy (MeDALL): Introducing novel concepts in allergy phenotypes. J Allergy Clin Immunol. 2017, 139, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, J.; Melén, E.; Haahtela, T.; Koppelman, G.H.; Togias, A.; Valenta, R.; et al. Rhinitis associated with asthma is distinct from rhinitis alone: The ARIA-MeDALL hypothesis. Allergy 2023. [CrossRef] [PubMed]
- Dramburg, S.; Hilger, C.; Santos, A.; et al. EAACI Molecular Allergology User's Guide 2.0. Pediatr Allergy Immunol. 2023, 34 (Suppl. 28), e13854. [Google Scholar] [CrossRef] [PubMed]
- https://www.aemet.es/documentos/es/conocermas/recursos_en_linea/publicaciones_y_estudios/publicaciones/Aerobiologia/Aerobiologia.pdf Last accessed, April 14, 2023.
- Bousquet, J.; Schünemann, H.J.; Togias, A.; Bachert, C.; Erhola, M.; Hellings, P.W.; et al. Allergic Rhinitis and Its Impact on Asthma Working Group. Next-generation Allergic Rhinitis and Its Impact on Asthma (ARIA) guidelines for allergic rhinitis based on Grading of Recommendations Assessment, Development and Evaluation (GRADE) and real-world evidence. J Allergy Clin Immunol. 2020, 145, 70–80. [Google Scholar] [PubMed]
- https://ginasthma.org/gina-reports/ Last accessed March 15, 2023.
- Wollenberg, A.; Barbarot, S.; Bieber, T.; Christen-Zaech, S.; Deleuran, M.; Fink-Wagner, A.; et al. Consensus-based European guidelines for treatment of atopic eczema (atopic dermatitis) in adults children: part, I. J Eur Acad Dermatol Venereol. 2019, 32, 657–682. [Google Scholar] [CrossRef]
- Goodess, C.M.; Giorgi, F.; Hamaoui-Laguel, L.; Semenov, M.A.; Solmon, F.; Storkey, J.; Vautard, R.; Epstein, M.M. Climate Change and Future Pollen Allergy in Europe. Environ Health Perspect. 2017, 125, 385–391. [Google Scholar]
- Beggs, P.J.; Clot, B.; Sofiev, M.; Johnston, F.H. Climate change, airborne allergens, and three translational mitigation approaches. EBioMedicine. 2023, 104478. [Google Scholar] [CrossRef]
- Martín, J.L.; Bethencourt, J.; Cuevas-Agulló, E. Assessment of global warming on the island of Tenerife, Canary Islands (Spain). Trends in minimum, maximum and mean temperatures since 1944. Climatic Change 2012, 114, 343–355. [Google Scholar] [CrossRef]
- Dominguez-Rodriguez, A.; Baez-Ferrer, N.; Rodríguez, S.; Avanzas, P.; Abreu-Gonzalez, P.; Terradellas, E.; Cuevas, E.; Basart, S.; Werner, E. Saharan Dust Events in the Dust Belt -Canary Islands- and the Observed Association with in-Hospital Mortality of Patients with Heart Failure. J. Clin. Med. 2020, 9, 376. [Google Scholar] [CrossRef]
- Szefler, S.J.; Wenzel, S.; Brown, R.; Erzurum, S.C.; Fahy, J.V.; Hamilton, R.G.; et al. Asthma outcomes: biomarkers. J Allergy Clin Immunol. 2012, 129 (3 Suppl), S9–23. [Google Scholar] [CrossRef]
- Kidon, M.I.; Chiang, W.C.; Chin, C.W.; Liew, W.K.; Kang, L.W.; Ong, T.C.; et al. Mite componentspecific IgE repertoire and phenotypes of allergic disease in childhood: the tropical perspective. Pediatr Allergy Immunol. 2011, 22, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Oksel, C.; Haider, S.; Fontanella, S.; Frainay, C.; Custovic, A. Classification of pediatric asthma: from phenotype discovery to clinical practice. Front Pediatr. 2018, 6, 258. [Google Scholar] [CrossRef] [PubMed]
- Scala, E.; Caprini, E.; Abeni, D.; Meneguzzi, G.; Buzzulini, F.; Cecchi, L.; Villalta, D.; Asero, R. A qualitative and quantitative comparison of IgE antibody profiles with two multiplex platforms for component-resolved diagnostics in allergic patients. Clin Exp Allergy 2021, 51, 1603–1612. [Google Scholar] [CrossRef] [PubMed]
- Platteel, A.C.M.; van der Pol, P.; Murk, J.L.; Verbrugge-Bakker, I.; Hack-Steemers, M.; Roovers, T.H.W.M.; Heron, M. A comprehensive comparison between ISAC and ALEX2 multiplex test systems. Clin Chem Lab Med. 2022, 60, 1046–1052. [Google Scholar] [CrossRef] [PubMed]
- Barber, D.; Arias, J.; Boquete, M.; Cardona, V.; Carrillo, T.; Gala, G.; et al. Analysis of mite allergic patients in a diverse territory by improved diagnostic tools. Clin Exp Allergy 2012, 42, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- González-Pérez, R.; Pineda, F.; Poza-Guedes, P.; Castillo, M.; Matheu, V.; Sánchez-Machín, I. Molecular Allergen Profiling of Dual Mite Sensitization in Severe Allergic Rhinitis. J Investig Allergol Clin Immunol. 2020, 30, 421–429. [Google Scholar] [CrossRef] [PubMed]
- González-Pérez, R.; Poza-Guedes, P.; Pineda, F.; Forstenlechner, P.; Castillo, M.; Mederos-Luís, E.; Aumayr, M.; Matheu, V.; Alava-Cruz, C.; Sánchez-Machín, I. Mite Molecular Profile in the Th2-Polarized Moderate-to-Severe Persistent Asthma Endotype Subjected to High Allergen Exposure. Int Arch Allergy Immunol. 2021, 182, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Vidal, C.; Lojo, S.; Juangorena, M.; Gonzalez-Quintela, A. Association Between Asthma and Sensitization to Allergens of Dermatophagoides pteronyssinus. J Investig Allergol Clin Immunol. 2016, 26, 304–309. [Google Scholar] [CrossRef]
- López-Rodríguez, R.; Rial, M.J.; Esteban-Gorgojo, I.; Veleiro Pérez, B.; López-Araújo, G.A.; Pérez-Quintero, O.; Carballas, C.; Parra, A.; Pineda, F. Serodominance Profile in a Dust Mite Complex Region. Int Arch Allergy Immunol. 2022, 183, 843–851. [Google Scholar] [CrossRef]
- Zeng, G.; Luo, W.; Zheng, P.; Wei, N.; Huang, H.; Sun, B.; Zhao, X. Component-Resolved Diagnostic Study of Dermatophagoides Pteronyssinus Major Allergen Molecules in a Southern Chinese Cohort. J Investig Allergol Clin Immunol. 2015, 25, 343–351. [Google Scholar]
- Rodinkova, V.V.; Yuriev, S.D.; Kryvopustova, M.V.; Mokin, V.B.; Kryzhanovskyi, Y.M.; Kurchenko, A.I. Molecular Profile Sensitization to House Dust Mites as an Important Aspect for Predicting the Efficiency of Allergen Immunotherapy. Front Immunol. 2022, 13, 848616. [Google Scholar] [CrossRef] [PubMed]
- Manolio, T.A.; Barnes, K.C.; Naidu, R.P.; Levett, P.N.; Beaty, T.H.; Wilson, A.F. Correlates of sensitization to Blomia tropicalis and Dermatophagoides pteronyssinus in asthma in Barbados. Int Arch Allergy Immunol 2003, 131, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.H.; Zhou, Q.; Prabhu, N.; Furuhashi, K.; Chua, Y.L.; Grotenbreg, G.M.; et al. Blomia tropicalis allergen 5 (Blo t 5) T-cell epitopes and their ability to suppress the allergic immune response. Immunology. 2017, 152, 344–355. [Google Scholar] [CrossRef] [PubMed]
- González-Pérez, R.; Poza-Guedes, P.; Pineda, F.; Castillo, M.; Sánchez-Machín, I. House Dust Mite Precision Allergy Molecular Diagnosis (PAMD@) in the Th2-prone Atopic Dermatitis Endotype. Life (Basel). 2021, 11, 1418. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, N.; Zakzuk, J.; Caraballo, L. House Dust Mite Allergy Under Changing Environments. Allergy Asthma Immunol Res. 2019, 11, 450–469. [Google Scholar] [CrossRef] [PubMed]
- Iraola Calvo, V.; Fernández-Caldas, E. Mapa acarológico de España; Laboratorios Leti: Barcelona, 2009; ISBN 978-84-69173336. [Google Scholar]
- Gafvelin, G.; Johansson, E.; Lundin, A.; Smith, A.M.; Chapman, M.D.; Benjamin, D.C.; et al. Cross-reactivity studies of a new group 2 allergen from the dust mite Glycyphagus domesticus, Gly d 2, and group 2 allergens from Dermatophagoides pteronyssinus, Lepidoglyphus destructor, and Tyrophagus putrescentiae with recombinant allergens. J Allergy Clin Immunol. 2001, 107, 511–518. [Google Scholar] [CrossRef]
- Osterlund, C.; Grönlund, H.; Gafvelin, G.; Bucht, A. Non-proteolytic aeroallergens from mites, cat and dog exert adjuvant-like activation of bronchial epithelial cells. Int Arch Allergy Immunol. 2011, 155, 111–118. [Google Scholar] [CrossRef]
- Gruchalla, R.S.; Pongracic, J.; Plaut, M.; et al. Inner City Asthma Study: relationships among sensitivity, allergen exposure, and asthma morbidity. J Allergy Clin Immunol 2005, 115, 478–485. [Google Scholar] [CrossRef]
- Subramanian, A.; Khatri, S.B. The Exposome and Asthma. Clin Chest Med. 2019, 40, 107–123. [Google Scholar] [CrossRef]
- Available online: https://izana.aemet.es/wp-content/docs/Izana_Report_2017_2018.pdf Last accessed March 3, 2023.
- Izquierdo, R.; Belmonte, J.; Avila, A.; Alarcón, M.; Cuevas, E.; Alonso-Pérez, S. Source areas and long-range transport of pollen from continental land to Tenerife (Canary Islands). Int J Biometeorol. 2011, 55, 67–85. [Google Scholar]
- Posa, D.; Perna, S.; Resch, Y.; Lupinek, C.; Panetta, V.; Hofmaier, S.; et al. Evolution and predictive value of IgE responses toward a comprehensive panel of house dust mite allergens during the first 2 decades of life. J Allergy Clin Immunol. 2017, 139, 541–549.e8. [Google Scholar] [CrossRef] [PubMed]
- Custovic, A.; Murray, C.S. The effect of allergen exposure in early childhood on the development of atopy. Curr Allergy Asthma Rep. 2002, 2, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Stefanovic, N.; Flohr, C.; Irvine, A.D. The exposome in atopic dermatitis. Allergy 2020, 75, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Bronnert, M.; Mancini, J.; Birnbaum, J.; Agabriel, C.; Liabeuf, V.; Porri, F.; et al. Component-resolved diagnosis with commercially available D. pteronyssinus Der p 1, Der p 2 and Der p 10: relevant markers for house dust mite allergy. Clin Exp Allergy 2012, 42, 1406–1415. [Google Scholar]
- Celi, G.; Brusca, I.; Scala, E.; Villalta, D.; Pastorello, E.; Farioli, L.; et al. House dust mite allergy in Italy-Diagnostic and clinical relevance of Der p 23 (and of minor allergens): A real-life, multicenter study. Allergy 2019, 74, 1787–1789. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, A.; Utsumi, D.; Lund, K.; Okano, M.; Ohashi-Doi, K.; Okubo, K. Correlation between sensitization to house dust mite major allergens, age, and symptoms in Japanese house dust mite allergic subjects. Int Immunopharmacol. 2022, 107, 108640. [Google Scholar] [CrossRef] [PubMed]
- Walsemann, T.; Böttger, M.; Traidl, S.; Schwager, C.; Gülsen, A.; Freimooser, S.; Roesner, L.M.; Werfel, T.; Jappe, U. Specific IgE against the house dust mite allergens Der p 5, 20 and 21 influences the phenotype and severity of atopic diseases. Allergy 2023, 78, 731–742. [Google Scholar] [CrossRef]
- Caraballo, L.; Valenta, R.; Puerta, L.; Pomés, A.; Zakzuk, J.; Fernandez-Caldas, E.; et al. The allergenic activity and clinical impact of individual IgE-antibody binding molecules from indoor allergen sources. World Allergy Organ J. 2020, 13, 100118. [Google Scholar] [CrossRef]
- Heinzerling, L.; Mari, A.; Bergmann, K.C.; Bresciani, M.; Burbach, G.; Darsow, U.; et al. The skin prick test - European standards. Clin Transl Allergy 2013, 3, 3. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 227, 680–685. [CrossRef] [PubMed]
- Bojcukova, J.; Vlas, T.; Forstenlechner, P.; Panzner, P. Comparison of two multiplex arrays in the diagnostics of allergy. Clin Transl Allergy 2019, 9, 31. [Google Scholar] [CrossRef]
Figure 1.
Flow diagram of study selection.
Figure 2.
Specific IgE serodominance in a comprehensive panel of molecular aeroallergens.
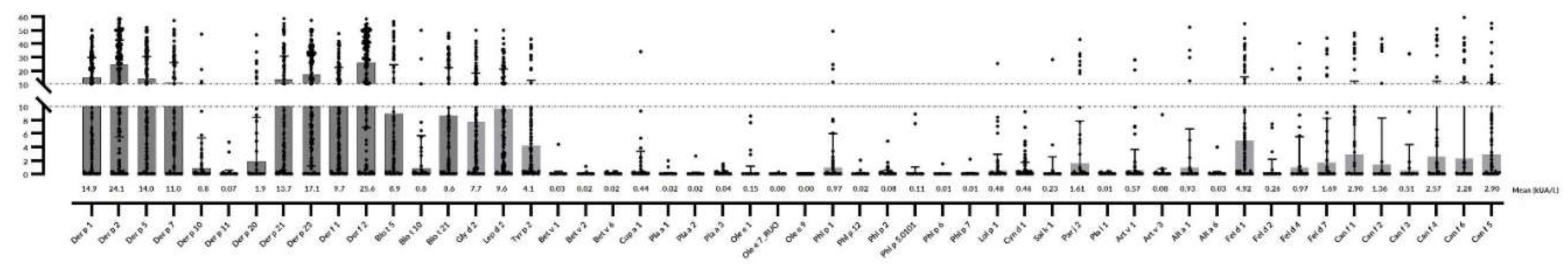
Figure 3.
Sensitization profile to specific IgE (sIgE) (A) heatmap (B) scatter plots with bars to a comprehensive panel of molecular allergens of Dermatophagoides pteronyssinus, Blomia tropicalis, Lepidoglyphus destructor, and Tyrophagus putrescentiae in 50 rhinitis, 50 rhinitis and asthma, 50 rhinitis and asthma and atopic dermatitis patients. (C) IgE- western blot of the different groups included against Dermatophagoides pteronyssinus (DPT), Blomia tropicalis (BTR) and Lepidoglyphus destructor (LDE). Broadly different patterns of sIgE binding were identified for each group of patients.
Figure 3.
Sensitization profile to specific IgE (sIgE) (A) heatmap (B) scatter plots with bars to a comprehensive panel of molecular allergens of Dermatophagoides pteronyssinus, Blomia tropicalis, Lepidoglyphus destructor, and Tyrophagus putrescentiae in 50 rhinitis, 50 rhinitis and asthma, 50 rhinitis and asthma and atopic dermatitis patients. (C) IgE- western blot of the different groups included against Dermatophagoides pteronyssinus (DPT), Blomia tropicalis (BTR) and Lepidoglyphus destructor (LDE). Broadly different patterns of sIgE binding were identified for each group of patients.
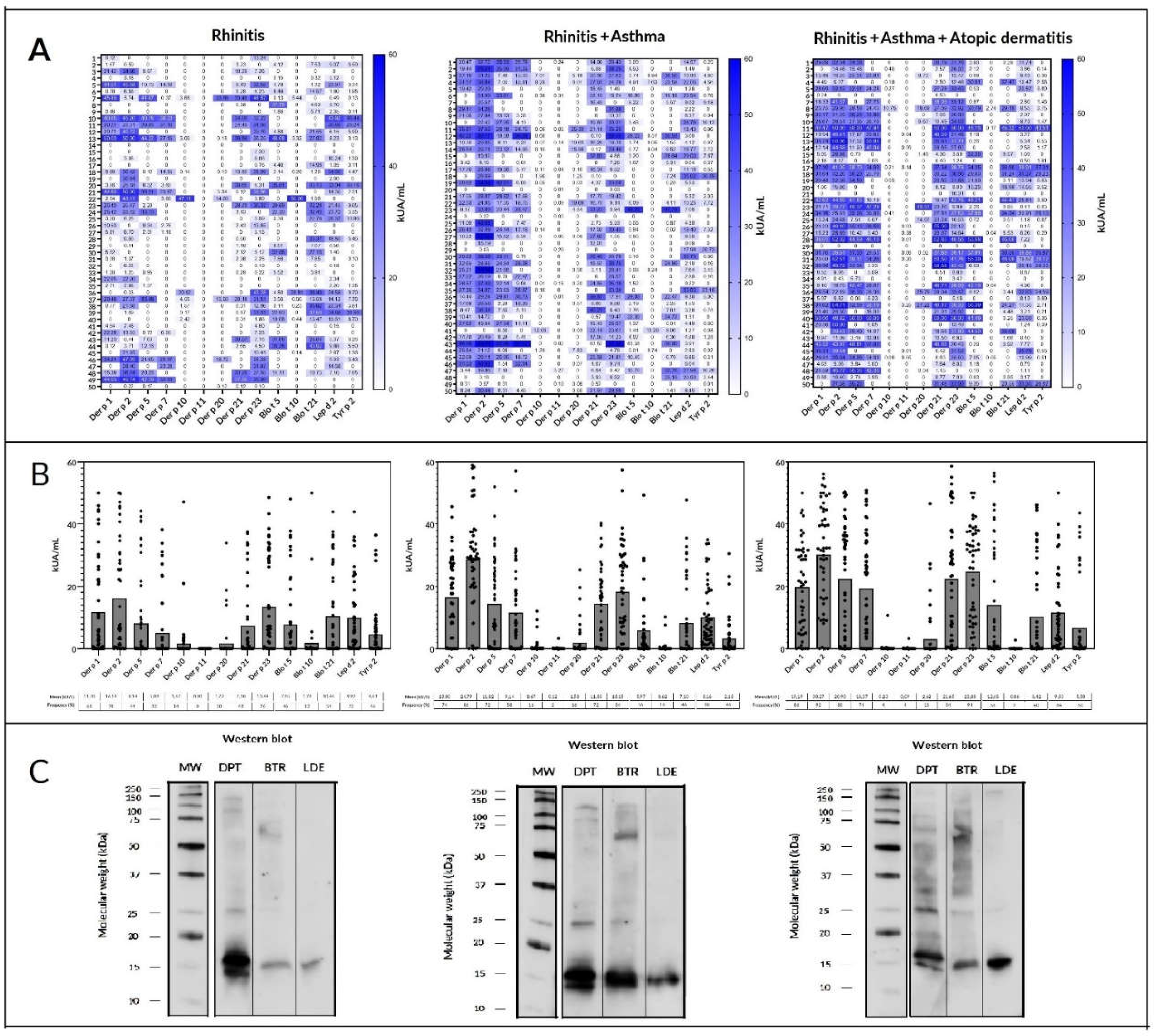
Table 1.
Descriptive statistics regarding basal comorbid conditions and associated clinical features of the studied population (n=150).
Table 1.
Descriptive statistics regarding basal comorbid conditions and associated clinical features of the studied population (n=150).
| Allergic Rhinitis | Allergic Asthma | Atopic Dermatitis | |
|---|---|---|---|
| n=150 | 50 | 50 | 50 |
| Age (y.o.) median (range) | 27.5 (9-70) | 32 (8-67) | 26 (9-62) |
| <20 y.o. (n= 54) | 22 (40.7%) | 16 (29.6%) | 16 (29.6%) |
| ≥20 y.o. (n=96) | 28 (29,1%) | 34 (35.4%) | 34 (35.4%) |
| Sex (F/M) | 29/21 | 32/18 | 22/28 |
| Food Allergy (n=23) | 8 (34.7%) | 6 (20.0%) | 9 (39.1%) |
| Drug Allergy (n=11) | 2 (18.1%) | 6 (54.5%) | 3 (27.27%) |
| SPT+ to any aeroallergen | 50 (100%) | 50 (100%) | 50 (100%) |
| Total IgE (IU/ml) median (range) | 56 (23.88-1083) | 674 (56.5-19,993) | 898 (81.81-17,420) |
| Blood Eosinophils/mm3 median (range) | 340 (140-530) | 365 (20-1,290) | 375 (25-1,880) |
| Family History of Atopy | 36 (72%) | 38 (76%) | 39 (78%) |
SPT: Skin Prick Test. Median values (ranges) are shown.
Table 2.
Prevalence of sensitization to grouped local aeroallergens by Skin Prick Test (n= 150).
| Positive SPT | N (%) |
|---|---|
| HDM and/or SM | 146 (97.33) |
| Cat and/or dog dander | 46 (30.66) |
| Pollen | 14 (9.33) |
| Cockroach | 9 (6) |
| Molds | 8 (5.33) |
SPT: Skin Prick Test. HDM: House Dust Mites. SM: Storage Mites.
Table 3.
Serological analysis of specific IgE (sIgE) responses (kU/L) to mite molecular allergens in patients with allergic rhinitis (AR, n=50), asthma (A, n=50), or atopic dermatitis (AD, n=50). The number (%) of subjects (n=150) sensitized to the corresponding mite molecular allergen is shown. Bold figures indicate quantitative significant differences (P< 0.05) in mean sIgE to mite molecular allergen among atopic conditions.
Table 3.
Serological analysis of specific IgE (sIgE) responses (kU/L) to mite molecular allergens in patients with allergic rhinitis (AR, n=50), asthma (A, n=50), or atopic dermatitis (AD, n=50). The number (%) of subjects (n=150) sensitized to the corresponding mite molecular allergen is shown. Bold figures indicate quantitative significant differences (P< 0.05) in mean sIgE to mite molecular allergen among atopic conditions.
| Mite allergen |
Mean sIgE (M±SD) | No. of sensitized patients (All) | Mean sIgE in AR (M±SD) |
No. of sensitized patients (AR) |
Mean sIgE in asthma (M±SD) | No. of sensitized patients (A) | Mean sIgE in atopic dermatitis (M±SD) | No. of sensitized patients (AD) |
|---|---|---|---|---|---|---|---|---|
| Der f 2 | 25.6±14.2 | 126 (84) | 17.17±0.2 | 39 (78) | 26.2±11.41 | 43 (86) | 32.7±0.31 | 46 (92) |
| Der p 2 | 24.1±15.3 | 125 (83) | 16.19±0.1 | 39 (78) | 24.79±12.3 | 43 (86) | 30.27±2.96 | 46 (92) |
| Der p 23 | 17.15±0.3 | 123 (82) | 13.44±1.2 | 38 (76) | 13.15±6.25 | 42 (84) | 23.88±12.7 | 47 (94) |
| Lep d 2 | 9.68± 1.5 | 114 (76) | 9.92±0.2 | 36 (72) | 8.16±5.23 | 40 (80) | 9.53±7.47 | 42 (84) |
| Der p 1 | 14.9±3.34 | 109 (72.6) | 11.7±4.3 | 34 (68) | 13±9.3 | 37 (74) | 19.19±14.5 | 43 (86) |
| Der f 1 | 9.79±2.51 | 104 (69.3) | 3.63±0.89 | 26 (52) | 10.36±8.32 | 38 (76) | 15.2±4.1 | 43 (86) |
| Gly d 2 | 7.72±1.53 | 101 (67.3) | 7.34±0.34 | 37 (74) | 5.03±3.9 | 32 (64) | 9.5±3.81 | 36 (72) |
| Der p 21 | 13.71±3.6 | 100 (66.6) | 7.38±0.36 | 24 (48) | 11.85±4.17 | 36 (72) | 21.65±0.22 | 42 (84) |
| Der p 5 | 14.07±1.0 | 96 (64) | 8.14±0.72 | 22 (44) | 11.82±1.66 | 36 (72) | 20.9±2.3 | 40 (80) |
| Der p 7 | 11.01±1.9 | 82 (54.6) | 5.08±0.39 | 16 (32) | 9.14±4.21 | 29 (58) | 18.37±1.9 | 37 (74) |
| Blo t 5 | 8.96±2.84 | 76 (50.6) | 7.85±0.53 | 23 (46) | 5.97±0.2 | 28 (56) | 12.65±2.75 | 28 (56) |
| Tyr p 2 | 4.18±0.2 | 68 (45.3) | 4.61±1,02 | 23 (46) | 2.15±2 | 23 (46) | 5.5±0.22 | 25 (50) |
| Blo t 21 | 8.64±0.31 | 66 (44) | 10.46±0.8 | 27 (54) | 7.1±5.3 | 23 (46) | 8.42±0.1 | 20 (40) |
| Der p 20 | 1.94±0.5 | 22 (14.6) | 1.72±0.08 | 5 (10) | 1.53±0.09 | 8 (16) | 2.62±0.5 | 9 (18) |
| Der p 10 | 0.86±0.23 | 16 (10.6) | 1.67±0.06 | 7 (14) | 0.67±2.3 | 8 (16) | <0.35 | 2 (4) |
| Blo t 10 | 0.83±0.41 | 14 (9.3) | 1.78±0.09 | 6 (12) | 0.62±0.2 | 8 (16) | <0.35 | 1 (2) |
| Der p 11 | 0.75±0.3 | 3 (2.0) | <0.35 | 0 (0) | <0.35 | 1 (2) | <0.35 | 2 (4) |
Table 4.
Number of identified mite molecular allergens and corresponding basal atopic disease (Allergic rhinitis, asthma and atopic dermatitis) in 150 patients studied with microarray. Most subjects (64%) with allergic rhinitis displayed a specific IgE response <9 mite molecules, in contrast to patients with asthma (58%), or atopic dermatitis (68%) showing a polysensitization profile to ≥10 individual mite allergens. .
Table 4.
Number of identified mite molecular allergens and corresponding basal atopic disease (Allergic rhinitis, asthma and atopic dermatitis) in 150 patients studied with microarray. Most subjects (64%) with allergic rhinitis displayed a specific IgE response <9 mite molecules, in contrast to patients with asthma (58%), or atopic dermatitis (68%) showing a polysensitization profile to ≥10 individual mite allergens. .
| Number of identified mite allergens | Allergic Rhinitis | Asthma | Atopic Dermatitis |
|---|---|---|---|
| 0 | 0 | 3 | 1 |
| 1 | 1 | 1 | 2 |
| 2 | 0 | 1 | 0 |
| 3 | 2 | 1 | 1 |
| 4 | 5 | 1 | 0 |
| 5 | 3 | 0 | 0 |
| 6 | 5 | 4 | 1 |
| 7 | 6 | 1 | 0 |
| 8 | 3 | 4 | 5 |
| 9 | 7 | 5 | 5 |
| 10 | 4 | 3 | 5 |
| 11 | 6 | 11 | 11 |
| 12 | 5 | 5 | 10 |
| 13 | 1 | 7 | 7 |
| 14 | 1 | 2 | 0 |
| 15 | 1 | 0 | 1 |
| 16 | 0 | 1 | 1 |
| 17 | 0 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



