
Submitted:
24 April 2023
Posted:
25 April 2023
You are already at the latest version
Abstract
Neutrophils are the most abundant immune cells and make up about 70% of white blood cells in human blood and play a critical role as the first line of defense in the innate immune response. They also help regulate the inflammatory environment to promote tissue repair. However, in cancer, neutrophils can be manipulated by tumors to either promote or hinder tumor growth depending on the cytokine pool. Studies have shown that tumor-bearing mice have increased levels of neutrophils in peripheral circulation, and that neutrophil-derived exosomes can deliver various cargos, including lncRNA and miRNA, which contribute to tumor growth and degradation of extracellular matrix. Exosomes derived from immune cells generally possess anti-tumor activities and induce tumor-cell apoptosis by delivering cytotoxic proteins, ROS generation, H2O2 or activation of Fas-mediated apoptosis in target cells. Engineered exosome-like nanovesicles have been developed to deliver chemotherapeutic drugs precisely to tumor cells. However, tumor-derived exosomes can aggravate cancer-associated thrombosis through the formation of neutrophil extracellular traps. Despite the advancements in neutrophil-related research, a detailed understanding of tumor-neutrophil crosstalk is still lacking and remains a major barrier in developing neutrophil-based or targeted therapy. This review will focus on the communication pathways between tumors and neutrophils, and the role of neutrophil-derived exosomes (NDEs) in tumor growth. Additionally, potential strategies to manipulate NDEs for therapeutic purposes will be discussed.
Keywords:
Neutrophils
; Tumor associated neutrophils
; neutrophil derived exosomes
; immune suppression
; metastasis
; tumor microenvironment
; Neutrophil extracellular trap
; Trogocytosis
; ADCC and ADCP
1. Introduction
Neutrophils are the most common polymorphonuclear (PMN) leukocytes [1] which constitute around 40% to 70% of total white blood cells. Neutrophils act as an immediate innate immune responder against inflammation [2], injury and infection[3].Invading pathogens invoke an inflammatory response that recruits neutrophils to the site of inflammation following chemical signals like IL-8, Leukotriene B4, C5a or H2O2 and destroys the pathogen by phagocytosis and release of antimicrobial substances like neutrophil extracellular traps (NETs)[1]. In response to pathogens or inflammatory molecules, neutrophils get activated, marginalize themselves onto the walls of blood vessels, invade infected or inflamed tissue, frequently even in tissue environments where other molecules or cells are unable to access and begin to release proteinases, cytokines and chemokines which direct the neighboring cells function towards resolution of inflammation [4] as well as initiate an immune response to clear off the pathogen[5]. Previously, neutrophils were considered as pathogen clearing cells. However, over the last two decades, neutrophils have been recognized as a highly versatile and sophisticated population, displaying both heterogeneity and plasticity. Based on pathological niche, neutrophils can differentiate into distinct subsets like immune-suppressive neutrophils in HIV and cancer or pro-inflammatory neutrophils in SLE or psoriasis[6]. The niche dependent differentiation of neutrophils and their elevated count in cancer attracted the attention of cancer biologists in order to uncover the role of neutrophils in cancer. In most cases, immune cells can be identified by their specific surface markers. Neutrophils lack any known well-defined specific surface marker which can distinguish neutrophils from other immune cells. A number of suggested markers, including LDL receptor 1[7] and CD10[8], are under investigation. Poorly defined surface markers and short half-life are the reasons for the difficulties of neutrophil study.
Tumor microenvironment (TME) is characterized by chronic inflammation which is considered as one of the hallmarks of cancer. Approximately 25% of tumor cases are characterized by chronic inflammation and infections caused by tumors[9]. Tumor inflammation is largely promoted by infiltrating immune cells. Along with other immune cells, neutrophils are a major inflammatory cell in solid tumors[10] such as melanoma, head and neck squamous cell carcinomas (HNSCCs), bronchoalveolar carcinomas, and renal carcinomas[11]. Elevated number of neutrophils in the blood [12]and tumor [13]is associated with a poor prognosis.
Tumor associated neutrophils (TANs) seem to be an effective way to treat cancer, given the association between TAN infiltration and high neutrophil-to-lymphocyte ratio (NLR) and a poor prognosis and low recurrence-free survival. There are, however, several drugs that target neutrophils already approved by the FDA to treat various inflammatory and autoimmune diseases, such as vasculitis, gout, rheumatoid arthritis, and cancer[14,15]. Some examples of anti-neutrophil drugs are acetylsalicylic acid (aspirin), which inhibits prostaglandin synthesis by neutrophils, and CCX168, which blocks the complement 5a receptor (C5aR) on neutrophils and reduces their activation and recruitment[16]. However, one of the challenges in anti-neutrophil drug design is the difficulty in identifying and targeting different subtypes of tumor-associated neutrophils (TANs), such as the pro-tumoral N2 TANs, which have distinct phenotypes and functions from the anti-tumoral N1 TANs. Therefore, more specific and selective anti-neutrophil drugs are needed to effectively modulate neutrophil function in tumor microenvironment.
Exosomes are nano-sized vesicles that are released by neutrophils and other cells and can modulate intercellular communication by transferring proteins, lipids, and nucleic acids. In asthma, neutrophil-derived exosomes (NDEs) can enhance the proliferation of airway smooth muscle (ASM) cells, which contributes to airway remodeling and obstruction. NDEs can also affect other immune cells in the airway, such as macrophages, dendritic cells, and T cells, by altering their cytokine production, antigen presentation, and activation status demonstrating a new mechanism that links neutrophils to immune responses and tissue remodeling in asthma. In recent years, exosomes have become a hot topic among cancer biologists for modulating the TME. Exosomes, in general, contain various immunomodulatory molecules like TGFβ, IL10, Foxp3, IL6, and PGE2, which have the potential to disarm active immune cells like CD4+ T cells, NK cells, and macrophages. On the other hand, tumor cell-derived exosomes (TE) are capable of recruiting immune suppressive T-regulatory cells in the tumor microenvironment [17]. TEs can also drive neutrophil polarization towards the N2 subtype and induce autophagy via HMGB1/TLR4/NF-κB signaling pathway[18]. Tumor stem-like cells (TSLC)-derived exosomes contain various RNAs, such as microRNAs, long non-coding RNAs, and circular RNAs. These RNAs can modulate their gene expression and function. In colorectal cancer, TSLC-derived exosomal RNAs can prime neutrophils to acquire a pro-tumoral N2 phenotype[19], which is characterized by increased expression of arginase-1, vascular endothelial growth factor, and matrix metalloproteinase-9[18]. However, whether these TEs driving neutrophil polarization are amenable to target, is subject to further studies exploring tumor-neutrophil signaling network.
In addition to being subject for targeted therapy, exosomes have emerged as an efficient drug-delivery system owing to their desirable intrinsic features, such as biocompatibility, nano-scale effects, long-range targeting, and stability in circulation. Today exosomes are being engineered to deliver anti-tumor agents directly into tumor cells to reduce drug mediated toxicity. Neutrophil derived exosomes (NDEs) are known to induce anti-tumor effects via activation of the apoptotic signaling pathway [20]. NDEs can efficiently cross blood brain barrier (BBB) proving to be an effective therapeutic approach to target inflamed brain tumors[14]. Dendritic cell-derived exosomes (DCE) have been shown to promote tumor cell apoptosis through TNF superfamily ligands [21],Natural killer cell-derived exosomes (NKE) have been reported to exert cytotoxic effects through FasL and perforins [22]. Neutrophil derived exosomes have been shown to act against arthritis and sepsis setup; However, except for being used as an effective way of targeting tumors in mice[20], the potential of this method as a therapeutic tool has not been extensively investigated due to its limited lifespan and low productivity. In this review, we aim to highlight the role of neutrophils in manipulating tumor-neutrophil crosstalk, as well as possible ways to target tumor cells and associated neutrophils using exosomes.
2. Neutrophils: from generation to tumor infiltration:
Neutrophils are generated in bone marrow every day from hematopoietic stem cells [1] through lymphoid priming under the stimulation of granulocyte colony stimulating factor (G-CSF). The maturation process (Figure 1) includes stages of promyelocyte, myelocyte, metamyelocyte band cell and finally neutrophil with the nucleus transforming from a round shape to a lobed morphology[23] and changing expression markers. In neutrophil differentiation, transcription factors play a crucial role, and their expression varies with the stages of differentiation.PU.1 and C/EBPα are essential for myeloid commitment and expression of G-CSF receptor. C/EBPδ and growth factor independent-1 (Gfi-1) promote terminal differentiation[24,25]. C/EBPβ controls emergency granulopoiesis [26]. Runx1-/- and Klf6-/- neutrophils cannot migrate and recruit at an inflamed site. RELB, IRF5 and JUNB drive neutrophil effector responses. RFX2 and RELB are required for neutrophil survival during the inflammation[27]. The various stages of differentiation can be also identified by the existence of different types of granules like azurophil in myeloblast to promyelocyte or gelatinase granules during band cell stage. The mentioned granules contain myeloperoxidase (MPO) and elastase, proteolytic enzymes like cathepsin-G, proteinase-3, antimicrobial defensins and matrix metalloproteinases. Bone marrow stromal cells produce ligands specific to the receptors displayed by neutrophil progenitors for the same to be retained at place. These ligands include VCAM-1 against VLA4 and CXCL-12 against CXCR4[28]. Mature neutrophils in bone marrow are kept in place through action of the chemokine receptors CXCR2 and CXCR4[29]. G-CSF induces exit of mature neutrophil from bone marrow through disruption of CXCL12 and CXCR4 interaction, while ligands of CXCR2 are demonstrated by the endothelial cells of blood vessel. G-CSF also reduces CXCL12 expression in bone marrow stromal cells as well as CXCR4 in neutrophil. IL-17 is responsible for production of G-CSF which involves positive feedback loops along with IL-1 and IL-23. Neutrophils are thereby mobilized to the site of inflammation through leucocyte adhesion cascade. The endothelial cells of the blood vessel at or near the inflamed site express E and P-selectins which bind to the glycoprotein ligands on neutrophil making them roll on the endothelium. Eventually, ICAM-1 and ICAM-2 ligands of the endothelial cells bind to the β2 integrin receptors on the neutrophils facilitating their adherence to the wall of the blood vessel.
The mechanisms of tumor-induced neutrophilia are unclear, but tumor-derived cytokines such as G-CSF, IL-1 and IL-6 may stimulate granulopoiesis, or neutrophil development. Activated neutrophils in the peripheral tissue are expected to follow chemoattractants and show N1 phenotype in the tumor context. Chemokines such as IL-8, CXCL6, and CCL3 attract neutrophils to tumor sites [30]. Ras activates CXC chemokines, which increase neutrophil infiltration in mice [30]. CXCL5 (epithelial neutrophil-activating peptide-78) and MMP-9 also recruit neutrophils in Hepato-Cellular Carcinoma (HCC) and promote tumor progression and metastasis [30]. In breast cancer mice models, IL-17 has been shown to bring neutrophils into the TME [31]. IL-17 also has roles in antitumor responses and N2 polarization, which will be discussed later.
Human tumors contain TANs with different phenotypes: N1 (CD62Llow CD54hi) and N2 (CD45+ Lin− HLADR− CD11b+ CD33+ CD66b+ [32], which are similar to G-MDSCs. The N2 phenotype may arise from the N1 phenotype or vice versa, depending on the levels of TGFβ, G-CSF or type-I Interferons in the TME. TGFβ promotes the conversion of immature and immunosuppressive LDNs to N2 phenotype, which resembles G-MDSCs. Tumor-induced neutrophilia also correlates with increased immature myeloid cells.
3. Tumor associated neutrophils:
Tumor-associated neutrophils (TANs) can be subdivided into two subtypes like tumor-associated macrophages (TAMs)-M1 and M2; namely anti-tumorigenic N1 (Good) and pro-tumorigenic N2 (Bad) (Figure 2). Good TANs act as anti-tumorigenic when they acquire the N1 phenotype, which is characterized by killing tumor cells directly by releasing granule contents such as elastase, cathepsin G and myeloperoxidase (MPO), or indirectly by forming neutrophil extracellular traps (NETs) that trap and damage tumor cells and secretion of immunostimulatory cytokines and chemokines such as interferon-γ (IFN-γ), tumor necrosis factor-α (TNF-α) and IL-12, or by presenting tumor antigens to T cells [33]. Factors that can induce the N1 phenotype are interferon-β (IFN-β), tumor necrosis factor-α (TNF-α) and transforming growth factor-β (TGF-β) inhibitors [30,34]. These cytokines can induce the expression of pro-inflammatory molecules and cytotoxic mediators on neutrophils, such as ICAM-1, IP-10/CXCL10, TNF-α, elastase, and cathepsin G. N1 neutrophils can be generated by exposure to tumor cell lysates or apoptotic tumor cells, which can stimulate the production of interferon-γ (IFN-γ) and tumor necrosis factor-α (TNF-α) by natural killer (NK) cells and T cells[35,36]. Therefore, tumor cell lysates or apoptotic tumor cells can induce a positive feedback loop between N1 neutrophils and other immune cells to inhibit tumor progression. Some other anti-tumorigenic effects of neutrophil include; a inhibiting angiogenesis, invasion and migration of tumor cells by expressing antiangiogenic factors such as angiostatin, endostatin and thrombospondin-1 [33]; preventing metastatic colonization by blocking adhesion molecules on endothelial cells or by competing with tumor cells for attachment sites [33].
Bad neutrophils (N2 TANs) are substantially different from the N1 subtype in their activation and cytokine status. In addition, the N2 population has different morphology from the N1 subtype, with its hypersegmented nuclei compared to the circular nuclei of N1 neutrophils [37]. N2 TANs promote tumor development and progression. However, the exact mechanisms of the actions of N2 TANs are not fully understood. It is believed that N2 TANs stimulate immunosuppression, tumor growth, angiogenesis, and metastasis by DNA instability, or by cytokines and chemokines release[30,38]. N2 TANs mainly secrete immunosuppressive cytokines, matrix degrading proteases and angiogenesis-promoting molecules [39] and thus cause immune escape. Overall, the presence of N2 TANs in the tumor-microenvironment is associated with poor prognosis in various types of cancer, including lung cancer, breast cancer, and colorectal cancer. However, more research is needed to fully understand the complex interactions between N2 TANs and other immune and tumor cells, as well as to develop effective strategies to target them for cancer therapy. In addition to serving as pathogenic markers, neutrophil to lymphocyte ratio (NLR) has been already declared to be an immensely important feature to predict fatal complications [40].
4. Intra-tumoral activities of neutrophils as expected:
Tumor microenvironment (TME) is composed of a variety of cells, including tumor cells and their surrounding immune cells, tumor-associated fibroblasts, vascular endothelial cells, etc. These immune cells include innate immune cells (neutrophils, macrophages, natural killer cells, dendritic cells etc) and adaptive immune cells (T and B cells). Innate immune cells contribute to tumor suppression either through direct recognition and killing of cancer cells or by triggering a strong adaptive immune response by triggering self-activation[49].Interestingly, innate immune cells have been found to be promising tools for treating hematopoietic malignancies and solid tumors owing to their antitumor immunocompetence[50]. Neutrophils are professional phagocytes in protecting the host against microbial infection as well as in inducing inflammation. Neutrophil-mediated phagocytosis of B-cell lymphoma cells has been shown to be induced by Rituximab (CD20). Antibody-dependent cellular phagocytosis (ADCP) is in fact, one of the most important methods of clearance of tumor cells. Neutrophil-mediated ADCP has been reported for a variety of therapeutic monoclonal antibodies, including Obinutuzumab, Ofatumumab, and Trastuzumab. Although preclinical studies have confirmed the role of ADCP as a potent mechanism to eliminate tumor cells, clinical studies are yet to be conducted to ensure its safe and effective use. Antibody-based activation of the complement system as well, stimulates neutrophil-mediated anticancer immune responses. C3a and C5a are potent neutrophil chemoattractants which are formed during complement cascade, increasing tumor infiltration by neutrophils during antibody immunotherapy in mice models[51].
In principle neutrophils cannot engulf complete cancer cells (~12µM-22µM in diameter) which are typically larger than neutrophils (~8µM in diameter). To subjugate this challenge neutrophils have devised a strategy called trogocytosis. In trogocytosis, a small part of the plasma membrane of the target cell is phagocytosed. This sequential trogocytosis leads to loss of membrane integrity that finally leads to lysis of the cell. In-vitro demonstration of trogocytosis has been conducted for several clinical antibodies like trastuzumab, cetuximab, rituximab etc. Trogocytosis requires physical proximity between neutrophils and tumor cells. This interaction is dependent on both CD18/CD11b and FcγR interactions. Cell death due to trogocytosis is independent of other effector mechanisms that are employed by neutrophils. Although the clinical contribution of neutrophil-mediated trogocytosis in eliminating tumor cells is yet to be determined.
ADCC is another weapon used by neutrophils to kill cancer cells. In ADCC, neutrophils recognize the cancer cell through an antibody bound to it, and then latch onto the cancer cell and release a cocktail of chemical mediators that destroy the cancer cell. ADCC triggers the release of ROS in addition to other cytotoxic molecules such as lactoferrin, elastase, arginase, myeloperoxidase, cathepsins, defensins, and MMP9 to eliminate tumor cells. Observation has been made that the specific role played by each neutrophil Fc receptor in mediating ADCC varies depending on the type of cancer. IgG-mediated ADCC is also influenced by the local inflammatory condition. Different therapeutics are currently being used in clinics that use the ADCC pathway. Rituximab induces ADCC in B cell lymphoma [52] and cetuximab in head and neck cancer via activation of neutrophils[53]. Moreover, certain antibodies, including Trastuzumab (HER2/Neu), Zalutumumab (EGFR), and Alemtuzumab (CD52) also eliminate tumor cells by means of neutrophil dependent ADCC. Studies have ascertained that N2 neutrophils produce and release genotoxic substances like ROS and NO into the TME which increases DNA instability[54]. An interesting observation was made by Haqqani et al., which demonstrated that the number of infiltrating neutrophils into TME correlates with the number of mutations at the hypoxanthine phosphoribosyl transferase (Hprt) locus in murine subcutaneous tumors. These mutations are mainly driven by ROS and iNOS released by neutrophils[55]. Myeloperoxidase produced by neutrophils is also an important mediator of genotoxicity. Hypochlorous acid produced by myeloperoxidase is one of the major drivers of mutation in the HPRT gene.
Sun et al. demonstrated that neutrophils suppress tumor cell proliferation via Fas/FasL pathway mediated cell cycle arrest. They used Fas-knockout cells to inhibit the FasL/FasR pathway in A549 cells[56,57]. Additionally, NETs also have cytotoxic effects composed of DNA-histone protease complexes, cathepsin G, and neutrophil elastase. Defensins, another component of NET, can also cause tumor cell lysis, further epithelial cells, and blood vessels that support tumor growth may be destroyed by histones in NETs[58]. Neutrophils express cytokine receptors like conventional cytokine receptors (type I and type II), TNF-receptor superfamily members, and members of the IL-1-receptors, which may assist in anticancer immune responses. TNF-α was identified as a key regulator for the cytotoxic activity of neutrophils toward breast cancer cells by means of neutrophil transmigration and NO release that promotes cancer cell killing. IL-8 and CXCL5 secreted by renal cell carcinoma (RCC) cells have been shown to recruit neutrophils and abrogate the formation of metastases supporting the anti-tumor role of neutrophils[59].
5. Mechanism of activation and function of neutrophils:
Neutrophils perform a crucial role in initiating and regulating long-term anti-tumor immune signaling responses and participate in bidirectional interactions with other immune cells (B and T cells, NK cells, DCs)[60,61,62]. Due to their capacity to present tumor antigens[63] through RBC phagocytosis [64]and subsequent presentation through MHC and other costimulatory molecules, neutrophils play a crucial role in recruiting anti-tumor CD8+ effector T cells, DC, macrophages, and NK cells in TME. TLR-stimulated neutrophils release NETs[65], which can then activate plasmacytoid dendritic cells (pDCs) [66] and T cells. pDCs are specialized in producing type I interferons (IFN-α and IFN-β). pDCs can activate T cells, which then produce cytokines to further activate T cells[67,68]. Contrarily, GM-CSF, and INF-γ are necessary factors for the acquisition of features of antigen-presenting cells (APC) in neutrophil [64]. N1 subtype secretes CCL3, CCL9, CXCL10, TNF-α, IL12, Cathepsin G, and neutrophil elastase (NE) to recruit, activate and induce proliferation of T cells to implement anti-tumor influence and improve adaptive immune responses[69,70].
CD11b+Ly6G+ neutrophils produce IL-17 in tumor bearing mice which promotes tumor growth through an IL-6-Stat3 signaling pathway [71]. Accordingly, increase in CD8+ T cell response and reduction in tumor growth in the murine lung cancer system can be achieved by inhibition of IL17 production in CD11b+Ly6G+ neutrophils[71]. On the other hand, CD66b+ neutrophil infiltration is associated with CD8+ T cell infiltration, colocalization and improved responsiveness of CD8+ T cells to TCR stimulation[72] in colorectal cancer[73],indicating that these cells may be utilized for promoting antitumor immunity. Neutrophils can be activated by CD64, CD32a, CD16a via IgG-based antibodies. Mechanistically, like any other immune cell, Fc receptor (FcR) of neutrophil interacts with monoclonal antibodies (mAbs) by binding to their Fc domains. Interaction between neutrophil-FcR and Fc region of the antibody triggers antibody-dependent cellular cytotoxicity (ADCC), antibody-dependent cellular phagocytosis (ADCP) or trogocytosis and release of some tumoricidal mediators. Opsonization of malignant cells, which is an antibody dependent direct killing mechanism in neutrophils is facilitated by monoclonal antibodies[73]. Neutrophils express different types of FcR for IgG(FcγR) having subtypes capable of inducing FcγRI (CD64), FcγRIIa (CD32a), FcγRIIb(CD32b), FcγRIIc (CD32c), FcγRIIIa (CD16a), and FcγRIIIb (CD16b) having different holds for IgG1, IgG2, and IgG4[74] in IgG-based cancer therapies. FcγRIIa is the most abundant FcR (160,000 copies/cell) present on neutrophils and is largely responsible for anticancer activity of IgG Abs. Each Fc receptor of neutrophils has the capacity to bind to IgG-opsonizing tumor cells and to contribute to ADCC operation[75]. Neutrophils express FcγRI after activation. FcαR1 (CD89) has superior anti-cancer potential in various cancer types than CD64, which has a high affinity for the IgA. Interaction of FcαRI with IgA monoclonal antibodies (mAbs) can attract neutrophils to tumor-infiltrating effector memory T cells (TEMs). FcγRIIa and FcγRIIIb represent high and moderate affinity for IgG respectively. FcγRIIIb is abundantly present on neutrophils, where it acts as functional cross-linkers for the escalation of susceptibility of lymphoma cells to anti-CD20-mediated apoptosis by facilitating the interaction between CD20 and anti-CD20 mAbs. Neutrophils activated with G-CSF or IFN-γ strongly upregulate CD64 and inhibit the expression of CD16b[59]. This variation in Fc receptor regulatory frameworks may affect the way antibodies direct neutrophil effector functions.
6. Immune suppression by neutrophils and therapeutic possibilities:
Immunosuppression is an important hallmark of cancer. Multiple reports suggest that neutrophils can suppress both the innate and adaptive immune response during cancer initiation, progression, and metastasis. Neutrophils utilize a variety of pathways leading to immunosuppression that can be targeted in order to divert their tumor-promoting functions to the anticipated anti-tumor functions.
Neutrophils release type 1 arginase (Arg I) via degranulation for degrading arginine [76] leading to inactivation of T cells and eventual immune suppression [77]. Arginine is important for maintaining the activities of T-cells through expression of T-cell co-receptor CD3ζ. Arg I enters into the cells via transporters like CAT1 and CAT2 where it is metabolized to NO by NO synthase (NOS) [78] leading to conversion of CD4+CD25- cells into CD4+CD25+ T regulatory cells i.e. induced immune suppression [79]. Arginase inhibitors as single agents and in combination with immune checkpoint therapy are under clinical trial against solid tumors (NCT02903914). In fact, INCB001158 (an Arginase inhibitor) immunotherapy which has been reported to reverse immunosuppressive effects of neutrophils and myeloid derived suppressor cells (MDSCs) is under phase II/phase III clinical study in advanced biliary tract cancer in combination with first line chemotherapy i.e. gemcitabine/cisplatin regimen [80].
Neutrophils activated by GM-CSF have PD-L1 on their surface, which blocks T cell growth through the PD-1/PD-L1 pathway in gastric cancer and hepatocellular carcinoma. Cancer-associated fibroblasts (CAFs) also make neutrophils express PD-L1, which impairs T-cell function through IL6/Stat3 signaling. A new peptide vaccine based on Arginase-1 combined with anti-PD1 has been shown to enhance T-cell infiltration in tumor and reduce the suppression of tumor-educated myeloid cells and change the M1/M2 ratio [81].
TANs use mitochondrial fatty acid oxidation to maintain their NADPH level under low glucose and then produce ROS that impair lymphocyte functions [82]. Tumor cell-released autophagosomes (TRAPs) also influence neutrophils to produce ROS and undergo caspase-3 mediated apoptosis. These apoptotic neutrophils then suppress T cell activation and proliferation [83]. Drugs like Apocynin, DPI or TAT peptide inhibitors can target NADPH oxidase family members in cancer treatment [84].
Tumor secreted factors influence myelopoiesis and produce pathologically activated neutrophils (PMNs), also called PMN-MDSCs, from bone marrow precursors [85]. PMN-MDSCs die by ferroptosis in the tumor microenvironment and produce oxygenated lipids that suppress T cells of human and mouse origin [86]. Ferroptosis has been linked to drug resistance [87], but the effect of these drugs on PMN-MDSCs is poorly studied.
7. Good neutrophils in tumor: indication of role reversal?
Neutrophils can stimulate or suppress tumor growth [63]. During early stages of the cancer, TANs can activate cytotoxic T cell in stage I/II either by direct priming or by secreting several cytokines in human lung cancer. Elevated IFN-γ, GM-CSF and reduction of Ikaros transcription factor promote TANs to acquire a hybrid state which has neutrophil and antigen presenting capacity [88] even though neutrophils are not professional APC. Tumor draining lymph nodes as well, play crucial role in cancer progression. TANs transmigrate to lymph nodes where they modulate T cell response in a stage dependent manner. In non-metastatic stage, neutrophils in lymph nodes acquire antigen presenting phenotype (HLA-DR+CD80+CD86+ICAM1+PDL1-) and activate cytotoxic T Cells (CD27+Ki67hiPD-1-) to improve patient prognosis [89]. As tumors progress, neutrophils cease to respond in an anti-proliferative manner and display PDL1 which is driven by GM-CSF and STAT3, enriching immunosuppressive T cells (CD27+Ki67hiPD1+) [89,90]. As a result of gaining this phenotype, TANs initiate tumor promoting activities like-angiogenesis [91], vascular permeability, tumor cell dissemination to other organs and immune suppression.
In contrast to earlier mentioned immunosuppressive role of GM-CSF induced neutrophils, in lung cancer model, Granot and his co-workers demonstrated anti-metastatic activity of G-CSF activated neutrophils through ROS production in TME which were drawn by tumor-secreted CCL2. [92,93]. The extent of ROS generation in tumor indeed, appears to serve as indication of whether the response is going to be pro- or anti-tumorigenic. Since ROS generation is correlated with activation of Lyn and Akt, inhibitors of Akt pathway should be able to reduce G-CSF-induced neutrophil activities [94]. Accordingly, therapeutic regimens should be carefully designed. Interestingly, earlier mentioned DPI as well, can control ROS production thereby proving interconnecting mechanisms. IL-17 secreted by neutrophils enhances their antitumor potential through the production of MPO, ROS, Interferon (IFN), and TFN-related apoptosis-inducing ligands (TRAIL), defensins, and proteinases. The MPO found in NETs has the potential to kill melanoma cells and slow tumor growth. Azide, cyanide, or the addition of catalases inhibit MPO activity and prevent neutrophil-mediated tumor cell lysis[93] thereby indicating therapeutic approaches. Generation of ROS by the activities of earlier mentioned NADPH oxidase, also result in production of H2O2 leading to neutrophil-mediated cytotoxicity through Transient Receptor Potential Cation Channel 2 (TRPM2) activation and subsequent chemokine production by monocytes [95].
IL-17 production by γδ T cells has been also shown to be responsible for G-CSF-dependent expansion of neutrophils and N2 polarization in mammary tumor [96] indicating differential role of IL-17 in neutrophils and γδ T cells. However, this tumor-promoting role was observed in early stages in contrast to late stages when depletion of neutrophil did not affect metastasis again suggesting either role reversal of the same after significant progression of the disease or loss of signal transduction pathway molecules specifically involved in tumor-cell-neutrophil interactions. Noteworthy, in a path breaking discovery, neutrophils have been shown to accumulate in premetastatic lung and inhibit metastatic seeding in mammary tumor model of mice [93] supporting the above-mentioned role-reversal hypothesis. The evidence mentioned, also suggests possibilities of successful detection of metastatic sites through neutrophil tracking and accordingly designing targeted delivery of drugs.
The phenotype and function of TANs are determined by the tumor microenvironment, which can induce different polarization states of neutrophils, ranging from N1 (anti-tumoral) to N2 (pro-tumoral). The balance between the pro-tumor and anti-tumor effects of neutrophils is influenced by several factors, including the tumor type, the stage of disease, and the patient's immune status. In some cases, neutrophils may promote tumor growth, while in other cases they may have anti-tumor effects.
8. Neutrophil-derived exosomes: emerging players in cancer metastasis
Exosomes were first reported in 1987 during in-vitro culture of sheep reticulocytes and these small vesicles were able to perform different membrane receptor activities including acetylcholinesterase, cytochalasin B binding (glucose transporter) nucleoside binding (i.e. nucleoside transporter), Na+-independent amino acid transport and the transferrin receptor[97]. Exosomes are formed by a double invagination of the plasma membrane (PM) and formation of multivesicular bodies (MVB) which contain intraluminal vesicles (ILV). The ILVs sizing about 40-160nm are then released in surrounding tissue environment or in circulation through fusion of the MVBs with the PM and eventual exocytosis [98] which are termed exosomes. Exosomes are usually referred as cargo vesicles which can carry proteins, lipid, genetic material, RNAs or membrane receptors [99]. The composition of exosomes is dependent on the cell type and tissue from which it originates. However, proteome profile showed that 72% proteins are common to most exosomes and these proteins contribute to their biogenesis [100]. Exosomes execute their mode of action by two ways: either fusion with to the recipient cell, since as they have a membrane surrounding the material they carry or by communicating with the receptors present on neighboring or distant cells, whereby they can reprogram the recipient cell.
Exosomes may arise de novo inside cells or may be consumed by an endosome. During invagination of the PM, proteins, lipids, DNA or RNA are incorporated inside the vesicular formation. These vesicles can be considered as mini parent cells which provide a novel way to communicate with neighboring or distant cells. Almost all types of cells can generate exosomes, such as epithelial, mesenchymal, stromal, lymphocytes, monocytes, macrophages, NK, dendritic, endothelial and tumor cells [101,102,103,104,105,106,107]. The preferred naming for exosome is based on the cell origin. Recent studies have shown that neutrophils also release exosomes [107] and can play a role independently outside neutrophils and orchestrate adaptive immune responses by influencing some cell types[14]. Exosomes possess functional resemblance to their parental cells as they do carry partial cellular component of parental cells. However, they do not reconstitute parental cell[108].
9. Unravelling the intricate role of neutrophil-derived exosomes in cancer progression:
Neutrophil derived exosomes (NDEs) play important roles in a variety of physiological and pathological processes, including inflammation, wound healing, and cancer[109,110]. In inflammation, NDEs can promote the recruitment of other immune cells to the site of inflammation where they release pro-inflammatory cytokines[111]. In wound healing, NDE promotes the proliferation and migration of fibroblasts. These migrated fibroblasts release growth factors that stimulate growth of new tissue[112]. In the context of cancer, NDEs have been shown to play a role in promoting tumor growth and metastasis, as well as suppressing the immune response to cancer cells. However, NDEs have been rarely studied for tumor treatment [20]. Recently Genschmer et al., showed that NDEs contain NE on its surface. These NE-bound NDEs bind and degrade ECM via the integrin Mac-1. NE has been found to be associated with metastasis of non-small lung cancer into the aorta[113]. Similarly, in prostate and breast cancer NE activates MAPK to induce ERK phosphorylation enhancing tumorigenesis. The differential polarization of neutrophil also leads to skewed production of exosomes. N1 neutrophils produce N1 NDEs, whose molecular contents are biased towards inflammatory biomolecules, whereas N2 neutrophils produce N2 NDEs, whose molecular contents are biased towards regulatory biomolecules. N1 NDEs promote macrophage activation and T cell proliferation, whereas N2 NDEs promote M2 polarization of macrophage and also reactivate tumor cells[114]. Proteases derived from NDEs play an important role in chronic inflammatory diseases of the lung, such as chronic obstructive pulmonary disease (COPD) and acute respiratory distress syndrome (ARDS), by contributing to tissue damage, mucus hypersecretion, and impaired host defense [115,116]. N2 NDEs also contain IL6 which promotes neutrophil polarization to N2 via the IL-6-STAT3-ERK1/2 signaling[117].Conversely, N1 NDEs transport the Defensin 1 that recruits lymphocytes. N1 NDEs transport miR-223 which plays a critical role in the development of the myeloid lineage cells, promote immune cell recruitment and carry factors such as RAP1A which is known to prevent metastasis. N1 NDEs can also carry pro-inflammatory cytokines, such as IL-1β, IL-2 and IL-4. Recently an interesting observation was made by Kolonics et al. which suggested that neutrophils produce EVs based on the surrounding environment which might function as an indicator of tumor progression. When N1 neutrophils enter the TME they release N1 NDEs with anti-tumor signals, but as the tumor grows and escapes immune surveillance, the same reprograms the immune cells around itself. At this point N1 neutrophils are switched to N2 phenotype and release N2 NDEs which promote tumor growth [114].
Although NDEs have been used previously to treat arthritis and sepsis, their role in cancer treatment remains unexplored mainly due to the low yield of NDEs and the short half-life of neutrophils themselves.
10. NDEs: Driving tumor invasion and metastasis:
Cancer metastasis consists of sequential steps of digestion of extracellular matrix (ECM) and dissemination of tumor cells from primary tumor, intravasation, transportation to a suitable niche, extravasation and adaptation to the secondary site and subsequent proliferation. The mentioned steps require secretion of matrix metalloproteinases for dissolving the ECM, manipulation of adhesion molecules and transportation. NDEs are extracellular vesicles that can influence cancer progression and metastasis by carrying various bioactive molecules. For example, Zhu et al. (2022) showed that these exosomes carried miR-23a-3p that targeted PTEN and enhanced liver cancer cell invasion [20]. Wang et al. (2019) demonstrated that NDEs carried S100A8/A9 that facilitated lung metastasis by creating pre-metastatic niches [118]. Park et al. (2019) revealed that NDEs carried CD177 that induced regulatory T cell differentiation and suppressed anti-tumor immunity [119]. Tian et al. (2018) reported that NDEs carried MMP9 that stimulated angiogenesis and increased gastric cancer cell invasion and metastasis[120].
Importantly, exosomes derived from the ascites of patients with ovarian cancer patients, activated CD8+ T cells [121] and Treg cells [122] secrete MMP9. Exosomes-derived from tumor cells, mesenchymal stem cells, Treg cells[123], mast cells are known to secrete VEGF [124]. Cathepsin G is secreted by neutrophils and other immune cells. It has been reported that cathepsin G can be released by exosomes. Cathepsin G may promote angiogenesis, the formation of new blood vessels by cleaving and activating VEGF[125]. Cathepsin G also induces the expression of MMPs[126]. Cathepsin G has been shown to stimulate TGF-β signaling for upregulating VEGF and MCP-1 [127]. Therefore, analysis of neutrophil-derived exosomes for presence of these factors may serve in understanding disease prognosis and open new avenues in disease treatment. These studies highlight the diverse roles of NDEs in promoting cancer metastasis through different mechanisms.
11. NDEs as therapeutic tools in cancer:
Understanding the role of NDEs in tumor progression has the potential to lead to new treatments that can target these processes and improve outcomes for cancer patients. One promising approach is to develop therapies that can inhibit the production or function of N2 NDEs, preventing them from promoting immune system suppression and tumor growth. Another approach is to use NDEs as a delivery system for cancer treatments. NDEs can transport molecules directly to cancer cells, allowing for targeted and efficient delivery of cancer therapies. Additionally, NDEs can be engineered to target specific cancer cells, further increasing their effectiveness as a delivery system for cancer treatments. In fact, NDEs have been decorated with superparamagnetic iron oxide nanoparticles (SPIONs) to achieve higher tumor-targeting therapeutic effect [20]. Some potential approaches (Figure 3) that are currently being investigated to treat solid cancers are being discussed below:
Inhibition of exosome release: Exosome biogenesis and release inhibitors are a new class of cancer therapies that aim to block the production or release of exosomes from cancer cells or other tumor supporting cells like N2 neutrophil. N2 TANs derived exosomes can carry tumor-promoting factors, such as growth factors and cytokines, to other cells and stimulate angiogenesis, invasion, and metastasis[128]. Additionally, NDEs can suppress the immune system by inducing apoptosis of T cells, inhibiting NK cell activity, and promoting the expansion of regulatory T cells and myeloid-derived suppressor cells [129]. Therefore, blocking N2-TANs-derived exosome and tumor cell derived exosome is a rational strategy to inhibit tumor progression. Exosome biogenesis and release inhibitors can interfere with the endosomal sorting complex required for transport (ESCRT) machinery or other proteins involved in exosome biogenesis and secretion, such as TSG101, CD9, Alix, SMase2, Rab11, ARF6, and Rab27[130,131] which can help to slow down or stop tumor growth. Rab27a and b are involved in the docking and fusion of neutrophil granules with the plasma membrane, which is an essential step in the release of neutrophil degranulation enzymes into the extracellular environment. A recent study has demonstrated that the knocking down of Rab27a and b can inhibit exosome secretion[132]. In the case of Rab27b more research is needed to investigate the potential role of Rab27b overexpression in NDEs in the context of tumor growth and treatment. In melanoma, Rab27a overexpression is correlated with decreased patient survival [133]. Blockage of the ARF6-based pathway causes mitochondrial aggregation near the microtubule-organizing center, and subsequently induces detrimental reactive oxygen species (ROS) production[134]. Inhibition of exosome release by tumor cells can inhibit NET formation and release of mature N2 neutrophil at the tumor site. Some of the most promising exosome biogenesis and release inhibitors include Manumycin A, Tipifarnib, Macitentan, Nexinhib20, ARF6 inhibitors etc. Manumycin A suppresses Ras/Raf/ERK1/2 signaling and hnRNP H1, a splicing factor that regulates exosome cargo[135]. Tipifarnib reduces ESCRT-0 proteins HRS and ALIX, as well as Rab27a [130]. Macitentan block the activation of endothelin receptors and downstream signaling pathways, such as ERK1/2 and Akt[131]. Nexinhib20 reduces ceramide levels and prevents MVB formation and ARF6 inhibitors impair the fusion of MVBs with the plasma membrane.
Modulation of exosome content: Modulation of the content of NDEs can be a potential strategy to target cancer. NDEs contain various bioactive molecules, such as microRNAs (miRNAs), proteins, and lipids, which can promote or inhibit cancer growth and metastasis. By modulating the content of NDEs, it may be possible to alter their effect on cancer cells and enhance their therapeutic potential. Load NDEs with tumor-suppressive molecules such as miRNAs[136], which can inhibit the expression of genes involved in cancer progression. For example, loading NDEs with miRNAs that target oncogenes or promote tumor suppressor pathways could help to suppress tumor growth and metastasis. Block NDEs to release of pro-tumorigenic molecules such as TGF-β[137] in later stage of cancer, which is a growth factor that promotes cancer cell proliferation, migration, and invasion. Inhibition of TGF-β release from NDEs could help to reduce the pro-tumorigenic effects of NDEs on cancer cells[138]. Furthermore, engineering NDEs to express specific proteins or peptides that target cancer cells, or their microenvironment could be another strategy to enhance their therapeutic potential. For instance, NDEs could be decorated with specific ligands that bind to receptors on cancer cells, which can increase their uptake by cancer cells and improve their efficacy as a delivery system for cancer treatments.
Neutrophil-derived exosomes: Targeting NDEs is a promising approach for cancer therapy. It can be achieved by using the antibodies that recognize and bind to specific surface proteins on NDEs, such as CD63, CD9 and CD81[139,140,141,142]. LFA-1 (lymphocyte function-associated antigen-1) is another receptor present on the surface of neutrophils and plays a role in neutrophil adhesion and migration[143]. These antibodies can be conjugated to drugs or other therapeutic agents, allowing them to be delivered specifically to cancer cells via N2 NDEs. We can also use nanoparticles, such as liposomes or gold nanoparticles, to deliver drugs or other therapeutics to NDEs. These nanoparticles can be engineered to target NDEs specifically, coat them with molecules that bind to surface proteins on NDEs. Once taken up by NDEs, the nanoparticles can deliver their cargo specifically to cancer cells, resulting in targeted and efficient delivery of cancer therapies.
Use targeted antibodies: Antibodies are a promising new therapeutic agent for cancer, and they are being investigated in a number of clinical trials[144]. An antibody that targets a cancer cell surface protein can be used to activate T cells, which can then attack and kill the cancer cells. There are just a few of the studies that have been done on the use of antibodies in NDEs to fight cancer and inflammation. Here are some of the specific antibodies that are being used in NDEs. such as Anti-EGFR, PD1 and CTLA-4. EGFR is a protein that is often overexpressed on cancer cells. Anti-EGFR antibodies can be used to target NDEs to cancer cells, or they can be used to deliver drugs or other therapeutics to cancer cells[145]. PD-1 (Programmed cell death protein 1) is expressed by T Cells and helps to regulate the immune response. When PD-1 binds to its ligands, PD-L1 or PD-L2, which are often overexpressed on cancer cells[146], it can inhibit T cell activation and promote immune evasion by cancer cells. pembrolizumab and nivolumab, have been developed to block the interaction between PD-1 and its ligands, thereby releasing the brakes on the immune system and allowing T cells to attack cancer cells[147]. These antibodies have been shown to be effective in the treatment of several types of cancer, including melanoma, non-small cell lung cancer, and bladder cancer. Another receptor which is expressed by T cells which act as a negative regulator for T cell activation and proliferation by binding to its ligand B7-1 and B7-2 present on cancer cell. Ipilimumab, an anti-CTLA-4 antibody which can block the interaction between CTLA-4 and its ligands, thereby enhancing T cell activation and promoting immune response[148].
Engineer NDEs to express anti-cancer protein: NDEs can be engineered to express genes that encode for anti-cancer proteins such as TRAIL protein which can be used to kill cancer cells. TRAIL binds to cell surface death receptors, TRAIL-R1 (DR4) and TRAIL-R2 (DR5) and facilitates formation of death-inducing signaling complex (DISC), eventually activating the p53-independent apoptotic cascade[149]. This unique mechanism makes TRAIL a potential anticancer therapeutic especially for p53-mutated tumors. Further research is needed to determine the feasibility and efficacy of engineering NDEs to express anti-cancer proteins such as TRAIL. While this approach holds promise, there are still significant challenges to be addressed, including optimizing the delivery and targeting of these therapeutic agents. In an exceptional study carfilzomib, a second-generation proteasome inhibitor, was loaded into nitrogen cavitation (NC)-derived extracellular vehicles (EVs) which efficiently depleted circulating Tumor cells (CTCs) and inhibited metastasis in murine models[150]. NC instantly disrupts neutrophils to form nanosized membrane vesicles. NC-derived EVs are similar to naturally secreted EVs by neutrophils but unlike natural EVs they include less nuclear acids and organelles. However, NC enhanced EV production by 16 times, opening up an avenue for achieving higher production of EVs for eventual encapsulation. Eventually, NC-derived EVs loaded with anti-inflammatory drug piceatannol inhibited lung inflammation and sepsis induced by lipopolysaccharide (ref). The above-mentioned approach may be also used to deliver siRNA-based cancer vaccines given the ability of NDEs to accumulate at Tumor site in contrast to nanoparticles which have the problem of accumulating in the liver. Further research is needed to investigate the feasibility and safety of these approaches and to identify the most effective cargo for NDEs in cancer therapy.
Immune modulation: Neutrophil-derived exosomes can modulate the immune response, and this property can be utilized to target solid cancers. NDEs have been shown to have immunomodulatory properties that can be harnessed for cancer therapy. One approach involves loading NDEs with immune-stimulatory molecules, such as toll-like receptor (TLR) ligands[151], to activate the immune response against cancer cells. TLR ligands can activate immune cells by binding to TLRs, which are expressed on the surface of various immune cells, including dendritic cells and T cells. By loading NDEs with TLR ligands, it is possible to stimulate the immune response and enhance anti-tumor immunity. NDEs loaded with a TLR9 ligand could activate dendritic cells and induce anti-tumor immunity in a mouse model of melanoma[152]. Another example, CD11b, which is surface receptor integrin on neutrophils. It has been shown that CD11b activation can promote anti-tumor immunity by stimulating pro-inflammatory macrophage polarization[153] and recruitment to the tumor microenvironment and their interaction with cancer cells[30].
Combination therapy: Combining different strategies has been shown to enhance the efficacy of neutrophil-derived exosome-based therapies. neutrophil-derived exosomes were loaded with the chemotherapy drug doxorubicin and a small interfering RNA (siRNA) targeting the anti-apoptotic protein Bcl-2, as well as immune-stimulatory CpG oligonucleotides. In a mouse model of melanoma, these exosomes induced apoptosis and enhanced anti-tumor immunity, resulting in a significant reduction in tumor growth[154]. The combination of doxorubicin and Bcl-2-targeting siRNA enhances the cytotoxic effects of chemotherapy and promotes apoptosis in tumor cells[155]. Additionally, the inclusion of CpG oligonucleotides in the exosome cargo can stimulate the immune system, leading to an increased anti-tumor response [156]. Neutrophil-derived exosomes are an attractive therapeutic option due to their ability to target cancer cells selectively and efficiently[20].
Overall, the role of neutrophil-derived exosomes in therapy is still in the early stages of investigation. However, their potential therapeutic applications are promising and warrant further research to fully understand their mechanisms and potential clinical applications.
12. Current advancements and future perspectives:
Numerous evidence supports the pro-tumor role of N2 neutrophils but targeting them remains a challenge due to the lack of clear molecular discrimination between TANs, especially PMN-MDSCs, from the remaining neutrophil populations. Technological advances, especially in single cell profiling techniques, such as single cell proteomics and single cell RNA-seq, may greatly enhance the abilities to classify heterogeneous neutrophil populations, into “bad neutrophils” or N2 neutrophils which can be targeted for therapy, and “good neutrophils” or N1 neutrophils which needs to be spared to prevent neutropenia which is a major side-effect of neutrophil-targeted therapy.
The mentioned roles that neutrophils take on when dealing with cancer emphasize their adaptability and capacity to react to a wide range of targets including modified self-cells through pattern recognition molecules (PRMs) like collectins, picolins and pentraxins[157,158] both inside and outside the TME. At an early stage before metastasis, neutrophils have been already shown to transmigrate to lymph nodes for anti-tumor T cell priming, which is converted into immunosuppressive functions after stimulation by GM-CSF/STAT3[159]. Therapies aimed to target N2 neutrophils are being assessed in clinical trials. CD33+ in combination with HLA-DR− CD15+ is used to identify N2 neutrophils. A novel tetravalent bispecific antibody AMV564 targeting CD33/CD3 has been developed and is currently being evaluated in early phase clinical trials (NCT03144245) for relapsed or refractory acute myeloid leukemia (AML). The efficacy of LXRβ agonists, such as GW3965 and RGX-104, for depletion of pro-tumor PMN-MDSCs are also being tested. Clinical trials determining the efficacy of blocking PMN-MDSC recruitment in enhancing cancer immunotherapy (NCT03161431) are underway.
In addition to research focusing towards selectively targeting the N2 neutrophils, the population as a whole are being considered to carry anti-tumor drugs owing to their mobility and responsiveness. A study by Wang et al., demonstrated that activated neutrophils could uptake drug-loaded albumin nanoparticles (NP) via Fcγ receptors. Chu et al., showed that NP can be efficiently delivered across the blood to the tumor tissue via neutrophils after induction of inflammation. Neutrophils have been shown to efficiently deliver anti-CD11b antibody-linked gold nanorods or pyropheophorbide-a loaded NPs in tumor cells inducing anti-tumor effects [150]. Noteworthy, acute stimulation of inflammation is required for neutrophil recruitment, either via monoclonal antibody or photosensitization. Neutrophils can also efficiently deliver drugs across the blood brain barrier (BBB) to inflamed brain tumors. For example, neutrophils have been reported to deliver paclitaxel (PTX)containing liposomes to the brain and prevent recurrence of glioma in murine models after surgical resection of tumors and thereby improving survival. Xue et al., showed that PTX liposomes exert negligible toxicity on neutrophils[150] High concentration of inflammatory signals triggered infiltration of neutrophils and the release of liposomal PTX. However, the delivery of drugs via neutrophils remains challenging owing to the short half-life. and anti-cancer drugs like PTX can significantly affect and impair neutrophil functions, viability and its migratory potential.
Evidence demonstrated that under the influence of the external magnetic field, superparamagnetic iron oxide nanoparticles (SPIONs) modified by NDE loaded with the chemotherapy drug doxorubicin (DOX) can be highly targeted and enriched at tumor sites, inhibiting tumor growth to a significant degree [11]. Intravenous injection of DOX-loaded NDEs suppressed tumor growth and increased survival time in murine models[20]. In another interesting study, Gao et al., demonstrated a unique strategy to generate extracellular vehicles (EVs) using nitrogen cavitation (NC)which instantly disrupts neutrophils to form nanosized membrane vesicles.
Overall, despite significant advancement in understanding the characteristics of TANs, their importance in both Tumor promotion and anti-tumor functions and therapeutic approaches targeting N2 neutrophils, implementation of the findings still needs clinical adaptability. In this context, identification of N1 and N2 NDE contents, relevance of the NDE contents with progression of the disease and further exploration regarding planned delivery systems of NDE-based nanotherapeutics seem extremely relevant and necessary for future research.
Author Contributions
AD: drafted the manuscript; SB and SP: assisted in drafting the manuscript; DS: designing, and critical analysis and drafting the manuscript. DKB: designing, critical analysis and drafting the final version and figures.
Funding
This work was supported by NIH NIDDK R01 DK120989 Grant.
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
we would like to acknowledge the Gastroenterology department of university of Pennsylvania for proving the facility to complete this review manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rosales, C. Neutrophil: A Cell with Many Roles in Inflammation or Several Cell Types? Front Physiol. 2018, 9, 113. [Google Scholar] [CrossRef] [PubMed]
- Liew, P.X.; Kubes, P. The Neutrophil’s Role During Health and Disease. Physiol. Rev. 2019, 99, 1223–1248. [Google Scholar] [CrossRef] [PubMed]
- Kubes, P. The enigmatic neutrophil: What we do not know. Cell Tissue Res. 2018, 371, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Greenlee-Wacker, M.C. Clearance of apoptotic neutrophils and resolution of inflammation. Immunol. Rev. 2016, 273, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Witter, A.R.; Okunnu, B.M.; Berg, R.E. The Essential Role of Neutrophils during Infection with the Intracellular Bacterial Pathogen Listeria monocytogenes. J. Immunol. 2016, 197, 1557–1565. [Google Scholar] [CrossRef]
- Scapini, P.; Cassatella, M.A. Social networking of human neutrophils within the immune system. Blood 2014, 124, 710–719. [Google Scholar] [CrossRef]
- Condamine, T.; Dominguez, G.A.; Youn, J.-I.; Kossenkov, A.V.; Mony, S.; Alicea-Torres, K.; Tcyganov, E.; Hashimoto, A.; Nefedova, Y.; Lin, C.; et al. Lectin-type oxidized LDL receptor-1 distinguishes population of human polymorphonuclear myeloid-derived suppressor cells in cancer patients. Sci. Immunol. 2016, 1, aaf8943–aaf8943. [Google Scholar] [CrossRef]
- Marini, O.; Costa, S.; Bevilacqua, D.; Calzetti, F.; Tamassia, N.; Spina, C.; De Sabata, D.; Tinazzi, E.; Lunardi, C.; Scupoli, M.T.; et al. Mature CD10+ and immature CD10− neutrophils present in G-CSF–treated donors display opposite effects on T cells. Blood 2017, 129, 1343–1356. [Google Scholar] [CrossRef]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Jaillon, S.; Ponzetta, A.; Di Mitri, D.; Santoni, A.; Bonecchi, R.; Mantovani, A. Neutrophil diversity and plasticity in tumour progression and therapy. Nat. Rev. Cancer 2020, 20, 485–503. [Google Scholar] [CrossRef]
- Shen, M.; Hu, P.; Donskov, F.; Wang, G.; Liu, Q.; Du, J. Tumor-Associated Neutrophils as a New Prognostic Factor in Cancer: A Systematic Review and Meta-Analysis. PLoS ONE 2014, 9, e98259. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.; Bastholt, L.; Geertsen, P.; Christensen, I.J.; Larsen, S.; Gehl, J.; von der Maase, H. Elevated neutrophil and monocyte counts in peripheral blood are associated with poor survival in patients with metastatic melanoma: A prognostic model. Br. J. Cancer 2005, 93, 273–278. [Google Scholar] [CrossRef]
- Sagiv, J.Y.; Michaeli, J.; Assi, S.; Mishalian, I.; Kisos, H.; Levy, L.; Damti, P.; Lumbroso, D.; Polyansky, L.; Sionov, R.V.; et al. Phenotypic diversity and plasticity in circulating neutrophil subpopulations in cancer. Cell Rep. 2015, 10, 562–573. [Google Scholar] [CrossRef]
- Wang, H.; Zang, J.; Zhao, Z.; Zhang, Q.; Chen, S. The Advances of Neutrophil-Derived Effective Drug Delivery Systems: A Key Review of Managing Tumors and Inflammation. Int. J. Nanomed. 2021, ume 16, 7663–7681. [Google Scholar] [CrossRef]
- Mutua, V.; Gershwin, L.J. A Review of Neutrophil Extracellular Traps (NETs) in Disease: Potential Anti-NETs Therapeutics. Clin. Rev. Allergy Immunol. 2020, 61, 194–211. [Google Scholar] [CrossRef] [PubMed]
- Kimoto, Y.; Horiuchi, T. The Complement System and ANCA Associated Vasculitis in the Era of Anti-Complement Drugs. Front. Immunol. 2022, 13, 926044. [Google Scholar] [CrossRef]
- Hou, P.-P.; Chen, H.-Z. Extracellular vesicles in the tumor immune microenvironment. Cancer Lett. 2021, 516, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, H.; Yuan, X.; Jiang, P.; Qian, H.; Xu, W. Tumor-derived exosomes induce N2 polarization of neutrophils to promote gastric cancer cell migration. Mol. Cancer 2018, 17, 146. [Google Scholar] [CrossRef]
- Hwang, W.-L.; Lan, H.-Y.; Cheng, W.-C.; Huang, S.-C.; Yang, M.-H. Tumor stem-like cell-derived exosomal RNAs prime neutrophils for facilitating tumorigenesis of colon cancer. J. Hematol. Oncol. 2019, 12, 1–17. [Google Scholar] [CrossRef]
- Zhang, J.; Ji, C.; Zhang, H.; Shi, H.; Mao, F.; Qian, H.; Xu, W.; Wang, D.; Pan, J.; Fang, X.; et al. Engineered neutrophil-derived exosome-like vesicles for targeted cancer therapy. Sci. Adv. 2022, 8, eabj8207. [Google Scholar] [CrossRef]
- Munich, S.; Sobo-Vujanovic, A.; Buchser, W.J.; Beer-Stolz, D.; Vujanovic, N.L. Dendritic cell exosomes directly kill tumor cells and activate natural killer cells via TNF superfamily ligands. OncoImmunology 2012, 1, 1074–1083. [Google Scholar] [CrossRef] [PubMed]
- Lugini, L.; Cecchetti, S.; Huber, V.; Luciani, F.; Macchia, G.; Spadaro, F.; Paris, L.; Abalsamo, L.; Colone, M.; Molinari, A.; et al. Immune surveillance properties of human NK cell-derived exosomes. J. Immunol. 2012, 189, 2833–2842. [Google Scholar] [CrossRef] [PubMed]
- Borregaard, N. Neutrophils, from Marrow to Microbes. Immunity 2010, 33, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Evrard, M.; Kwok, I.W.H.; Chong, S.Z.; Teng, K.W.W.; Becht, E.; Chen, J.; Sieow, J.L.; Penny, H.L.; Ching, G.C.; Devi, S.; et al. Developmental Analysis of Bone Marrow Neutrophils Reveals Populations Specialized in Expansion, Trafficking, and Effector Functions. Immunity 2018, 48, 364–379. [Google Scholar] [CrossRef] [PubMed]
- Hock, H.; Hamblen, M.J.; Rooke, H.M.; Traver, D.; Bronson, R.T.; Cameron, S.; Orkin, S.H. Intrinsic Requirement for Zinc Finger Transcription Factor Gfi-1 in Neutrophil Differentiation. Immunity 2003, 18, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, R.; Barlow, C.; Lekstrom-Himes, J.; Castilla, LH.; Liu, PP.; Eckhaus, M; et al. Impaired granulopoiesis, myelodysplasia, and early lethality in CCAAT/enhancer binding protein epsilon-deficient mice. Proc Natl Acad Sci U S A. 1997, 94, 13187–13192. [Google Scholar] [CrossRef] [PubMed]
- Khoyratty, T.E.; Ai, Z.; Ballesteros, I.; Eames, H.L.; Mathie, S.; Martín-Salamanca, S.; Wang, L.; Hemmings, A.; Willemsen, N.; von Werz, V.; et al. Distinct transcription factor networks control neutrophil-driven inflammation. Nat. Immunol. 2021, 22, 1093–1106. [Google Scholar] [CrossRef]
- Petty JM, Lenox CC, Weiss DJ, Poynter ME, Suratt BT. Crosstalk between CXCR4/stromal derived factor-1 and VLA-4/VCAM-1 pathways regulates neutrophil retention in the bone marrow. J Immunol. 2009, 182, 604–612. [CrossRef]
- Furze, R.C.; Rankin, S.M. Neutrophil mobilization and clearance in the bone marrow. Immunology 2008, 125, 281–288. [Google Scholar] [CrossRef]
- Masucci MT, Minopoli M, Carriero MV. Tumor Associated Neutrophils. Their Role in Tumorigenesis, Metastasis, Prognosis and Therapy. Front Oncol. 2019, 9, 1146. [Google Scholar] [CrossRef]
- SenGupta, S.; Hein, L.E.; Parent, C.A. The Recruitment of Neutrophils to the Tumor Microenvironment Is Regulated by Multiple Mediators. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, Z.; Wu, L.; Zhang, M.; Li, W.; Ding, J.; Zhu, J.; Wei, H.; Zhao, K. Circulating and Tumor-Infiltrating Myeloid-Derived Suppressor Cells in Patients with Colorectal Carcinoma. PLoS ONE 2013, 8, e57114. [Google Scholar] [CrossRef] [PubMed]
- Shaul, M.E.; Fridlender, Z.G. Tumour-associated neutrophils in patients with cancer. Nat. Rev. Clin. Oncol. 2019, 16, 601–620. [Google Scholar] [CrossRef] [PubMed]
- Subhan MA, Torchilin VP. Neutrophils as an emerging therapeutic target and tool for cancer therapy. Life Sci. 2021, 285, 119952. [Google Scholar] [CrossRef] [PubMed]
- Schimek, V.; Strasser, K.; Beer, A.; Göber, S.; Walterskirchen, N.; Brostjan, C.; Müller, C.; Bachleitner-Hofmann, T.; Bergmann, M.; Dolznig, H.; et al. Tumour cell apoptosis modulates the colorectal cancer immune microenvironment via interleukin-8-dependent neutrophil recruitment. Cell Death Dis. 2022, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Shaul ME, Levy L, Sun J, Mishalian I, Singhal S, Kapoor V, et al. Tumor-associated neutrophils display a distinct N1 profile following TGFbeta modulation: A transcriptomics analysis of pro- vs. antitumor TANs. Oncoimmunology. 2016, 5, e1232221. [CrossRef] [PubMed]
- Ohms, M.; Möller, S.; Laskay, T. An Attempt to Polarize Human Neutrophils Toward N1 and N2 Phenotypes in vitro. Front. Immunol. 2020, 11, 532. [Google Scholar] [CrossRef]
- Lau D, Lechermann LM, Gallagher FA. Clinical Translation of Neutrophil Imaging and Its Role in Cancer. Mol Imaging Biol. 2022, 24, 221–234. [Google Scholar] [CrossRef]
- Fridlender ZG, Sun J, Kim S, Kapoor V, Cheng G, Ling L, et al. Polarization of tumor-associated neutrophil phenotype by TGF-beta: "N1" versus "N2" TAN. Cancer Cell. 2009, 16, 183–194. [CrossRef]
- Chiang, C.-C.; Korinek, M.; Cheng, W.-J.; Hwang, T.-L. Targeting Neutrophils to Treat Acute Respiratory Distress Syndrome in Coronavirus Disease. Front. Pharmacol. 2020, 11, 572009. [Google Scholar] [CrossRef]
- Ellis TN, Beaman BL. Interferon-gamma activation of polymorphonuclear neutrophil function. Immunology. 2004, 112, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Gomes, N.E.; Brunialti, M.K.; Mendes, M.E.; Freudenberg, M.; Galanos, C.; Salomão, R. Lipopolysaccharide-induced expression of cell surface receptors and cell activation of neutrophils and monocytes in whole human blood. Braz. J. Med. Biol. Res. 2010, 43, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Andzinski L, Kasnitz N, Stahnke S, Wu CF, Gereke M, von Kockritz-Blickwede M, et al. Type I IFNs induce anti-tumor polarization of tumor associated neutrophils in mice and human. Int J Cancer. 2016, 138, 1982–1993. [CrossRef]
- Gerlini, G.; Tun-Kyi, A.; Dudli, C.; Burg, G.; Pimpinelli, N.; Nestle, F.O. Metastatic Melanoma Secreted IL-10 Down-Regulates CD1 Molecules on Dendritic Cells in Metastatic Tumor Lesions. Am. J. Pathol. 2004, 165, 1853–1863. [Google Scholar] [CrossRef] [PubMed]
- Greenhough A, Smartt HJ, Moore AE, Roberts HR, Williams AC, Paraskeva C, et al. The COX-2/PGE2 pathway: Key roles in the hallmarks of cancer and adaptation to the tumour microenvironment. Carcinogenesis. 2009, 30, 377–386. [CrossRef] [PubMed]
- Joshita, S.; Nakazawa, K.; Sugiyama, Y.; Kamijo, A.; Matsubayashi, K.; Miyabayashi, H.; Furuta, K.; Kitano, K.; Kawa, S. Granulocyte-Colony Stimulating Factor-Producing Pancreatic Adenosquamous Carcinoma Showing Aggressive Clinical Course. Intern. Med. 2009, 48, 687–691. [Google Scholar] [CrossRef]
- Romero-Garcia, S.; Moreno-Altamirano, M.M.B.; Prado-Garcia, H.; Sánchez-García, F.J. Lactate Contribution to the Tumor Microenvironment: Mechanisms, Effects on Immune Cells and Therapeutic Relevance. Front. Immunol. 2016, 7, 52. [Google Scholar] [CrossRef]
- Leone, R.D.; Emens, L.A. Targeting adenosine for cancer immunotherapy. J. Immunother. Cancer 2018, 6, 57. [Google Scholar] [CrossRef]
- Woo SR, Corrales L, Gajewski TF. Innate immune recognition of cancer. Annu Rev Immunol. 2015, 33, 445–474. [Google Scholar]
- Bonneville M, Scotet E. Human Vgamma9Vdelta2 T cells: Promising new leads for immunotherapy of infections and tumors. Curr Opin Immunol. 2006, 18, 539–546. [Google Scholar] [CrossRef]
- Hong, F.; Yan, J.; Baran, J.T.; Allendorf, D.J.; Hansen, R.D.; Ostroff, G.R.; Xing, P.X.; Cheung, N.-K.V.; Ross, G.D. Mechanism by Which Orally Administered β-1,3-Glucans Enhance the Tumoricidal Activity of Antitumor Monoclonal Antibodies in Murine Tumor Models. J. Immunol. 2004, 173, 797–806. [Google Scholar] [CrossRef]
- Hernandez-Ilizaliturri FJ, Jupudy V, Ostberg J, Oflazoglu E, Huberman A, Repasky E, et al. Neutrophils contribute to the biological antitumor activity of rituximab in a non-Hodgkin's lymphoma severe combined immunodeficiency mouse model. Clin Cancer Res. 2003, 9, 5866–5873.
- Vermorken, J.; Specenier, P. Cetuximab: Its unique place in head and neck cancer treatment. Biol. Targets Ther. 2013, ume 7, 77–90. [Google Scholar] [CrossRef]
- Xiong, S.; Dong, L.; Cheng, L. Neutrophils in cancer carcinogenesis and metastasis. J. Hematol. Oncol. 2021, 14, 173. [Google Scholar] [CrossRef]
- Sandhu, J.K.; Privora, H.F.; Wenckebach, G.; Birnboim, H.C. Neutrophils, Nitric Oxide Synthase, and Mutations in the Mutatect Murine Tumor Model. Am. J. Pathol. 2000, 156, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Tazawa, H.; Okada, F.; Kobayashi, T.; Tada, M.; Mori, Y.; Une, Y.; Sendo, F.; Kobayashi, M.; Hosokawa, M. Infiltration of Neutrophils Is Required for Acquisition of Metastatic Phenotype of Benign Murine Fibrosarcoma Cells: Implication of Inflammation-Associated Carcinogenesis and Tumor Progression. Am. J. Pathol. 2003, 163, 2221–2232. [Google Scholar] [CrossRef]
- Sun, B.; Qin, W.; Song, M.; Liu, L.; Yu, Y.; Qi, X.; Sun, H. Neutrophil Suppresses Tumor Cell Proliferation via Fas /Fas Ligand Pathway Mediated Cell Cycle Arrested. Int. J. Biol. Sci. 2018, 14, 2103–2113. [Google Scholar] [CrossRef] [PubMed]
- Garley M, Jablonska E, Dabrowska D. NETs in cancer. Tumour Biol. 2016, 37, 14355–14361.
- Avtenyuk, N.U.; Visser, N.; Bremer, E.; Wiersma, V.R. The Neutrophil: The Underdog That Packs a Punch in the Fight against Cancer. Int. J. Mol. Sci. 2020, 21, 7820. [Google Scholar] [CrossRef] [PubMed]
- Shaul ME, Zlotnik A, Tidhar E, Schwartz A, Arpinati L, Kaisar-Iluz N, et al. Tumor-Associated Neutrophils Drive B-cell Recruitment and Their Differentiation to Plasma Cells. Cancer Immunol Res. 2021, 9, 811–824. [CrossRef]
- Oberg, H.-H.; Wesch, D.; Kalyan, S.; Kabelitz, D. Regulatory Interactions Between Neutrophils, Tumor Cells and T Cells. Front. Immunol. 2019, 10, 1690. [Google Scholar] [CrossRef]
- Furumaya, C.; Martinez-Sanz, P.; Bouti, P.; Kuijpers, T.W.; Matlung, H.L. Plasticity in Pro- and Anti-tumor Activity of Neutrophils: Shifting the Balance. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Granot, Z. Neutrophils as a Therapeutic Target in Cancer. Front. Immunol. 2019, 10, 1710. [Google Scholar] [CrossRef]
- Meinderts SM, Baker G, van Wijk S, Beuger BM, Geissler J, Jansen MH, et al. Neutrophils acquire antigen-presenting cell features after phagocytosis of IgG-opsonized erythrocytes. Blood Adv. 2019, 3, 1761–1773. [CrossRef] [PubMed]
- Hayashi, F.; Means, T.K.; Luster, A.D. Toll-like receptors stimulate human neutrophil function. Blood 2003, 102, 2660–2669. [Google Scholar] [CrossRef] [PubMed]
- Lande, R.; Ganguly, D.; Facchinetti, V.; Frasca, L.; Conrad, C.; Gregorio, J.; Meller, S.; Chamilos, G.; Sebasigari, R.; Riccieri, V.; et al. Neutrophils Activate Plasmacytoid Dendritic Cells by Releasing Self-DNA–Peptide Complexes in Systemic Lupus Erythematosus. Sci. Transl. Med. 2011, 3, 73ra19–73ra19. [Google Scholar] [CrossRef] [PubMed]
- Psarras, A.; Antanaviciute, A.; Alase, A.; Carr, I.; Wittmann, M.; Emery, P.; Tsokos, G.C.; Vital, E.M. TNF-α Regulates Human Plasmacytoid Dendritic Cells by Suppressing IFN-α Production and Enhancing T Cell Activation. J. Immunol. 2021, 206, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Riise, R.E.; Bernson, E.; Aurelius, J.; Martner, A.; Pesce, S.; Della Chiesa, M.; Marcenaro, E.; Bylund, J.; Hellstrand, K.; Moretta, L.; et al. TLR-Stimulated Neutrophils Instruct NK Cells To Trigger Dendritic Cell Maturation and Promote Adaptive T Cell Responses. J. Immunol. 2015, 195, 1121–1128. [Google Scholar] [CrossRef]
- Uribe-Querol, E.; Rosales, C. Neutrophils in Cancer: Two Sides of the Same Coin. J. Immunol. Res. 2015, 2015, 1–21. [Google Scholar] [CrossRef]
- Que, H.; Fu, Q.; Lan, T.; Tian, X.; Wei, X. Tumor-associated neutrophils and neutrophil-targeted cancer therapies. Biochim. et Biophys. Acta (BBA) - Rev. Cancer 2022, 1877, 188762. [Google Scholar] [CrossRef]
- Liu, Y.; O'Leary, C.E.; Wang, L.-C.S.; Bhatti, T.R.; Dai, N.; Kapoor, V.; Liu, P.; Mei, J.; Guo, L.; Oliver, P.M.; et al. CD11b+Ly6G+ cells inhibit tumor growth by suppressing IL-17 production at early stages of tumorigenesis. OncoImmunology 2015, 5, e1061175. [Google Scholar] [CrossRef]
- Bert, S.; Nadkarni, S.; Perretti, M. Neutrophil-T cell crosstalk and the control of the host inflammatory response. Immunol. Rev. 2023, 314, 36–49. [Google Scholar] [CrossRef]
- Heemskerk N, van Egmond M. Monoclonal antibody-mediated killing of tumour cells by neutrophils. Eur J Clin Invest. 2018, 48 Suppl 2, e12962. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn F, Ravetch JV. Fcgamma receptors as regulators of immune responses. Nat Rev Immunol. 2008, 8, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Wang Y, Jonsson F. Expression, Role, and Regulation of Neutrophil Fcgamma Receptors. Front Immunol. 2019, 10, 1958. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez PC, Zea AH, Culotta KS, Zabaleta J, Ochoa JB, Ochoa AC. Regulation of T cell receptor CD3zeta chain expression by L-arginine. J Biol Chem. 2002, 277, 21123–21129. [CrossRef] [PubMed]
- Marti ILAA, Reith W. Arginine-dependent immune responses. Cell Mol Life Sci. 2021, 78, 5303–5324. [Google Scholar] [CrossRef]
- Luiking, Y.C.; Ten Have, G.A.; Wolfe, R.R.; Deutz, N.E. Arginine de novo and nitric oxide production in disease states. Am J Physiol Endocrinol Metab. 2012, 303, E1177–E1189. [Google Scholar] [CrossRef]
- Niedbala, W.; Cai, B.; Liew, F.Y. Role of nitric oxide in the regulation of T cell functions. Ann. Rheum. Dis. 2006, 65 (Suppl 3), iii37–iii40. [Google Scholar] [CrossRef]
- Javle MM, Bridgewater JA, Gbolahan OB, Jungels C, Cho MT, Papadopoulos KP, et al. A phase I/II study of safety and efficacy of the arginase inhibitor INCB001158 plus chemotherapy in patients (Pts) with advanced biliary tract cancers. J Clin Oncol. 2021, 39. [CrossRef]
- Aaboe Jorgensen M, Ugel S, Linder Hubbe M, Carretta M, Perez-Penco M, Weis-Banke SE, et al. Arginase 1-Based Immune Modulatory Vaccines Induce Anticancer Immunity and Synergize with Anti-PD-1 Checkpoint Blockade. Cancer Immunol Res. 2021, 9, 1316–1326. [CrossRef]
- Rice, C.M.; Davies, L.C.; Subleski, J.J.; Maio, N.; Gonzalez-Cotto, M.; Andrews, C.; Patel, N.L.; Palmieri, E.M.; Weiss, J.M.; Lee, J.-M.; et al. Tumour-elicited neutrophils engage mitochondrial metabolism to circumvent nutrient limitations and maintain immune suppression. Nat. Commun. 2018, 9, 5099. [Google Scholar] [CrossRef]
- Wu, M.; Ma, M.; Tan, Z.; Zheng, H.; Liu, X. Neutrophil: A New Player in Metastatic Cancers. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Weyemi, U.; Redon, C.E.; Parekh, P.R.; Dupuy, C.; Bonner, W.M. NADPH Oxidases NOXs and DUOXs as putative targets for cancer therapy. Anticancer Agents Med Chem. 2013, 13, 502–514. [Google Scholar]
- Raskov, H.; Orhan, A.; Gaggar, S.; Gögenur, I. Neutrophils and polymorphonuclear myeloid-derived suppressor cells: An emerging battleground in cancer therapy. Oncogenesis 2022, 11, 22. [Google Scholar] [CrossRef]
- Kim, R.; Hashimoto, A.; Markosyan, N.; Tyurin, V.A.; Tyurina, Y.Y.; Kar, G.; Fu, S.; Sehgal, M.; Garcia-Gerique, L.; Kossenkov, A.; et al. Ferroptosis of tumour neutrophils causes immune suppression in cancer. Nature 2022, 612, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, X.; Jin, S.; Chen, Y.; Guo, R. Ferroptosis in cancer therapy: A novel approach to reversing drug resistance. Mol. Cancer 2022, 21, 1–12. [Google Scholar] [CrossRef]
- Singhal, S.; Bhojnagarwala, P.S.; O’Brien, S.; Moon, E.K.; Garfall, A.L.; Rao, A.S.; Quatromoni, J.G.; Stephen, T.L.; Litzky, L.; Deshpande, C.; et al. Origin and role of a subset of tumor-associated neutrophils with antigen-presenting cell features in early-stage human lung cancer. Cancer Cell 2016, 30, 120–135. [Google Scholar] [CrossRef] [PubMed]
- Pylaeva, E.; Korschunow, G.; Spyra, I.; Bordbari, S.; Siakaeva, E.; Ozel, I.; Domnich, M.; Squire, A.; Hasenberg, A.; Thangavelu, K.; et al. During early stages of cancer, neutrophils initiate anti-tumor immune responses in tumor-draining lymph nodes. Cell Rep. 2022, 40, 111171. [Google Scholar] [CrossRef]
- Wang, T.-T.; Zhao, Y.-L.; Peng, L.-S.; Chen, N.; Chen, W.; Lv, Y.-P.; Mao, F.-Y.; Zhang, J.-Y.; Cheng, P.; Teng, Y.-S.; et al. Tumour-activated neutrophils in gastric cancer foster immune suppression and disease progression through GM-CSF-PD-L1 pathway. Gut 2017, 66, 1900–1911. [Google Scholar] [CrossRef]
- Jablonska J, Leschner S, Westphal K, Lienenklaus S, Weiss S. Neutrophils responsive to endogenous IFN-beta regulate tumor angiogenesis and growth in a mouse tumor model. J Clin Invest. 2010, 120, 1151–1164. [CrossRef] [PubMed]
- Andrés, C.M.C.; de la Lastra, J.M.P.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. Myeloid-Derived Suppressor Cells in Cancer and COVID-19 as Associated with Oxidative Stress. Vaccines 2023, 11, 218. [Google Scholar] [CrossRef] [PubMed]
- Granot, Z.; Henke, E.; Comen, E.A.; King, T.A.; Norton, L.; Benezra, R. Tumor entrained neutrophils inhibit seeding in the premetastatic lung. Cancer Cell 2011, 20, 300–314. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.-S.; Xia, L.; Mills, G.B.; Lowell, C.A.; Touw, I.P.; Corey, S.J. G-CSF induced reactive oxygen species involves Lyn-PI3-kinase-Akt and contributes to myeloid cell growth. Blood 2006, 107, 1847–1856. [Google Scholar] [CrossRef] [PubMed]
- Hack CT, Buck T, Bagnjuk K, Eubler K, Kunz L, Mayr D, et al. A Role for H(2)O(2) and TRPM2 in the Induction of Cell Death: Studies in KGN Cells. Antioxidants (Basel) 2019, 8.
- Coffelt SB, Kersten K, Doornebal CW, Weiden J, Vrijland K, Hau CS, et al. IL-17-producing gammadelta T cells and neutrophils conspire to promote breast cancer metastasis. Nature. 2015, 522, 345–348. [CrossRef] [PubMed]
- Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem. 1987, 262, 9412–9420. [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 19. [Google Scholar] [CrossRef]
- Mathivanan, S.; Fahner, C.J.; Reid, G.E.; Simpson, R.J. ExoCarta 2012: Database of exosomal proteins, RNA and lipids. Nucleic Acids Res. 2012, 40, D1241–D1244. [Google Scholar] [CrossRef]
- Cheng, L.; Zhang, K.; Wu, S.; Cui, M.; Xu, T. Focus on Mesenchymal Stem Cell-Derived Exosomes: Opportunities and Challenges in Cell-Free Therapy. Stem Cells Int. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Hua, W.; Liu, J.; Fan, L.; Wang, H.; Sun, G. Exosomes derived from endoplasmic reticulum-stressed liver cancer cells enhance the expression of cytokines in macrophages via the STAT3 signaling pathway. Oncol. Lett. 2020, 20, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, Y.; Jin, X.; Hu, D.; Xia, C.; Xu, H.; Hu, J. NK cell-derived exosomes carry miR-207 and alleviate depression-like symptoms in mice. J. Neuroinflammation 2020, 17, 1–19. [Google Scholar] [CrossRef]
- Zhao D, Yu Z, Li Y, Wang Y, Li Q, Han D. GelMA combined with sustained release of HUVECs derived exosomes for promoting cutaneous wound healing and facilitating skin regeneration. J Mol Histol. 2020, 51, 251–263. [CrossRef]
- Li, H.; Feng, Y.; Zheng, X.; Jia, M.; Mei, Z.; Wang, Y.; Zhang, Z.; Zhou, M.; Li, C. M2-type exosomes nanoparticles for rheumatoid arthritis therapy via macrophage re-polarization. J. Control. Release 2021, 341, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Ramos, T.; Parekh, M.; Kaye, S.B.; Ahmad, S. Epithelial Cell-Derived Extracellular Vesicles Trigger the Differentiation of Two Epithelial Cell Lines. Int. J. Mol. Sci. 2022, 23, 1718. [Google Scholar] [CrossRef] [PubMed]
- Vargas, A.; Roux-Dalvai, F.; Droit, A.; Lavoie, J.-P. Neutrophil-Derived Exosomes: A New Mechanism Contributing to Airway Smooth Muscle Remodeling. Am. J. Respir. Cell Mol. Biol. 2016, 55, 450–461. [Google Scholar] [CrossRef]
- Ren, K. Exosomes in perspective: A potential surrogate for stem cell therapy. Odontology 2019, 107, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Cheng, C.; Wei, Y.; Yang, F.; Li, G. The Role of Exosomes in Inflammatory Diseases and Tumor-Related Inflammation. Cells 2022, 11, 1005. [Google Scholar] [CrossRef]
- Al-Nedawi, K.; Meehan, B.; Micallef, J.; Lhotak, V.; May, L.; Guha, A.; Rak, J. Intercellular transfer of the oncogenic receptor EGFRvIII by microvesicles derived from tumour cells. Nat. Cell Biol. 2008, 10, 619–624. [Google Scholar] [CrossRef]
- Noonin, C.; Thongboonkerd, V. Exosome-inflammasome crosstalk and their roles in inflammatory responses. Theranostics 2021, 11, 4436–4451. [Google Scholar] [CrossRef]
- Arishe, O.O.; Priviero, F.; Wilczynski, S.A.; Webb, R.C. Exosomes as Intercellular Messengers in Hypertension. Int. J. Mol. Sci. 2021, 22, 11685. [Google Scholar] [CrossRef]
- Yamashita, J.-I.; Ogawa, M.; Abe, M.; Hayashi, N.; Kurusu, Y.; Kawahara, K.; Shirakusa, T. Tumor Neutrophil Elastase Is Closely Associated With the Direct Extension of Non-small Cell Lung Cancer Into the Aorta. Chest 1997, 111, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Rubenich, D.S.; Omizzollo, N.; Szczepański, M.J.; Reichert, T.E.; Whiteside, T.L.; Ludwig, N.; Braganhol, E. Small extracellular vesicle-mediated bidirectional crosstalk between neutrophils and tumor cells. Cytokine Growth Factor Rev. 2021, 61, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Rivot, A.; Jugniot, N.; Jacoutot, S.; Vanthuyne, N.; Massot, P.; Mellet, P.; Marque, S.R.; Audran, G.; Voisin, P.; Delles, M.; et al. Magnetic Resonance Imaging of Protease-Mediated Lung Tissue Inflammation and Injury. ACS Omega 2021, 6, 15012–15016. [Google Scholar] [CrossRef] [PubMed]
- Jasper, A.E.; McIver, W.J.; Sapey, E.; Walton, G.M. Understanding the role of neutrophils in chronic inflammatory airway disease. F1000Research 2019, 8, F1000. [Google Scholar] [CrossRef]
- Contesini, F.J.; de Melo, R.R.; Sato, H.H. An overview of Bacillus proteases: From production to application. Crit. Rev. Biotechnol. 2018, 38, 321–334. [Google Scholar] [CrossRef]
- Xue, R.; Zhang, Q.; Cao, Q.; Kong, R.; Xiang, X.; Liu, H.; Feng, M.; Wang, F.; Cheng, J.; Li, Z.; et al. Liver tumour immune microenvironment subtypes and neutrophil heterogeneity. Nature 2022, 612, 141–147. [Google Scholar] [CrossRef]
- Kim, H.Y.; Min, H.-K.; Song, H.-W.; Yoo, A.; Lee, S.; Kim, K.-P.; Park, J.-O.; Choi, Y.H.; Choi, E. Delivery of human natural killer cell-derived exosomes for liver cancer therapy: An in vivo study in subcutaneous and orthotopic animal models. Drug Deliv. 2022, 29, 2897–2911. [Google Scholar] [CrossRef]
- Tian, Y.; Li, S.; Song, J.; Ji, T.; Zhu, M.; Anderson, G.J.; Wei, J.; Nie, G. A doxorubicin delivery platform using engineered natural membrane vesicle exosomes for targeted tumor therapy. Biomaterials 2014, 35, 2383–2390. [Google Scholar] [CrossRef]
- Hazrati, A.; Soudi, S.; Malekpour, K.; Mahmoudi, M.; Rahimi, A.; Hashemi, S.M.; Varma, R.S. Immune cells-derived exosomes function as a double-edged sword: Role in disease progression and their therapeutic applications. Biomark. Res. 2022, 10, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Asemani, Y.; Najafi, S.; Ezzatifar, F.; Zolbanin, N.M.; Jafari, R. Recent highlights in the immunomodulatory aspects of Treg cell-derived extracellular vesicles: Special emphasis on autoimmune diseases and transplantation. Cell Biosci. 2022, 12, 67. [Google Scholar] [CrossRef] [PubMed]
- Lee JK, Park SR, Jung BK, Jeon YK, Lee YS, Kim MK, et al. Exosomes derived from mesenchymal stem cells suppress angiogenesis by down-regulating VEGF expression in breast cancer cells. PLoS ONE. 2013, 8, e84256.
- Detoraki, A.; Staiano, R.I.; Granata, F.; Giannattasio, G.; Prevete, N.; de Paulis, A.; Ribatti, D.; Genovese, A.; Triggiani, M.; Marone, G. Vascular endothelial growth factors synthesized by human lung mast cells exert angiogenic effects. J. Allergy Clin. Immunol. 2009, 123, 1142–1149. [Google Scholar] [CrossRef] [PubMed]
- Olejarz, W.; Kubiak-Tomaszewska, G.; Chrzanowska, A.; Lorenc, T. Exosomes in Angiogenesis and Anti-angiogenic Therapy in Cancers. Int. J. Mol. Sci. 2020, 21, 5840. [Google Scholar] [CrossRef]
- Bausch, D.; Pausch, T.; Krauss, T.; Hopt, U.T.; Fernandez-Del-Castillo, C.; Warshaw, A.L.; Thayer, S.P.; Keck, T. Neutrophil granulocyte derived MMP-9 is a VEGF independent functional component of the angiogenic switch in pancreatic ductal adenocarcinoma. Angiogenesis 2011, 14, 235–243. [Google Scholar] [CrossRef]
- Wilson TJ, Nannuru KC, Futakuchi M. Cathepsin G-mediated enhanced TGF-beta signaling promotes angiogenesis via upregulation of VEGF and MCP-1. Cancer Lett. 2010, 288, 162–169. [Google Scholar] [CrossRef]
- Labani-Motlagh, A.; Ashja-Mahdavi, M.; Loskog, A. The Tumor Microenvironment: A Milieu Hindering and Obstructing Antitumor Immune Responses. Front. Immunol. 2020, 11, 940. [Google Scholar] [CrossRef]
- Dai, J.; Su, Y.; Zhong, S.; Cong, L.; Liu, B.; Yang, J.; Tao, Y.; He, Z.; Chen, C.; Jiang, Y. Exosomes: Key players in cancer and potential therapeutic strategy. Signal Transduct. Target. Ther. 2020, 5, 145. [Google Scholar] [CrossRef]
- Datta, A.; Kim, H.; McGee, L.; Johnson, A.E.; Talwar, S.; Marugan, J.; Southall, N.; Hu, X.; Lal, M.; Mondal, D.; et al. High-throughput screening identified selective inhibitors of exosome biogenesis and secretion: A drug repurposing strategy for advanced cancer. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Andreu, Z.; Masiá, E.; Charbonnier, D.; Vicent, M.J. A Rapid, Convergent Approach to the Identification of Exosome Inhibitors in Breast Cancer Models. Nanotheranostics 2023, 7, 1–21. [Google Scholar] [CrossRef]
- Zhang, H.; Lu, J.; Liu, J.; Zhang, G.; Lu, A. Advances in the discovery of exosome inhibitors in cancer. J. Enzym. Inhib. Med. Chem. 2020, 35, 1322–1330. [Google Scholar] [CrossRef]
- Guo, D.; Lui, G.Y.; Lai, S.L.; Wilmott, J.S.; Tikoo, S.; Jackett, L.A.; Quek, C.; Brown, D.L.; Sharp, D.M.; Kwan, R.Y.; et al. RAB27A promotes melanoma cell invasion and metastasis via regulation of pro-invasive exosomes. Int. J. Cancer 2019, 144, 3070–3085. [Google Scholar] [CrossRef] [PubMed]
- Onodera, Y.; Nam, J.-M.; Horikawa, M.; Shirato, H.; Sabe, H. Arf6-driven cell invasion is intrinsically linked to TRAK1-mediated mitochondrial anterograde trafficking to avoid oxidative catastrophe. Nat. Commun. 2018, 9, 2682. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, C.-H.; Baek, M.-C. Dissecting exosome inhibitors: Therapeutic insights into small-molecule chemicals against cancer. Exp. Mol. Med. 2022, 54, 1833–1843. [Google Scholar] [CrossRef] [PubMed]
- Graner, M.W.; Schnell, S.; Olin, M.R. Tumor-derived exosomes, microRNAs, and cancer immune suppression. Semin. Immunopathol. 2018, 40, 505–515. [Google Scholar] [CrossRef]
- Syed, V. TGF-beta Signaling in Cancer. J Cell Biochem. 2016, 117, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chengalvala, V.; Hu, H.; Sun, D. Tumor-derived exosomes: Nanovesicles made by cancer cells to promote cancer metastasis. Acta Pharm. Sin. B 2021, 11, 2136–2149. [Google Scholar] [CrossRef]
- Pesce E, Manfrini N, Cordiglieri C, Santi S, Bandera A, Gobbini A, et al. Exosomes Recovered From the Plasma of COVID-19 Patients Expose SARS-CoV-2 Spike-Derived Fragments and Contribute to the Adaptive Immune Response. Front Immunol. 2021, 12, 785941. [CrossRef]
- He, G.; Peng, X.; Wei, S.; Yang, S.; Li, X.; Huang, M.; Tang, S.; Jin, H.; Liu, J.; Zhang, S.; et al. Exosomes in the hypoxic TME: From release, uptake and biofunctions to clinical applications. Mol. Cancer 2022, 21, 19. [Google Scholar] [CrossRef]
- Im, Y.; Yoo, H.; Ko, R.-E.; Lee, J.Y.; Park, J.; Jeon, K. Exosomal CD63 in critically ill patients with sepsis. Sci. Rep. 2021, 11, 20300. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Salam, B.K.; Ebaid, H. Expression of CD11b and CD18 on polymorphonuclear neutrophils stimulated with interleukin-2. Cent Eur J Immunol 2014, 39, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Awla, D.; Abdulla, A.; Zhang, S.; Roller, J.; Menger, M.D.; Regnér, S.; Thorlacius, H. Lymphocyte function antigen-1 regulates neutrophil recruitment and tissue damage in acute pancreatitis. Br. J. Pharmacol. 2011, 163, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Sun, Y.; Liang, X.; Gu, X.; Ning, J.; Xu, Y.; Chen, S.; Pan, L. Emerging new therapeutic antibody derivatives for cancer treatment. Signal Transduct. Target. Ther. 2022, 7, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Abdollahzadeh, F.; Nejatollahi, F. Anti-Proliferative Effect of Specific Anti-EGFR Single Chain Antibody on Triple Negative Breast Cancer Cells. Rep. Biochem. Mol. Biol. 2020, 9, 180–187. [Google Scholar] [CrossRef]
- Diskin, B.; Adam, S.; Cassini, M.F.; Sanchez, G.; Liria, M.; Aykut, B.; Buttar, C.; Li, E.; Sundberg, B.; Salas, R.D.; et al. PD-L1 engagement on T cells promotes self-tolerance and suppression of neighboring macrophages and effector T cells in cancer. Nat. Immunol. 2020, 21, 442–454. [Google Scholar] [CrossRef]
- Robert, C.; Ribas, A.; Wolchok, J.D.; Hodi, F.S.; Hamid, O.; Kefford, R.; Weber, J.S.; Joshua, A.M.; Hwu, W.-J.; Gangadhar, T.C.; et al. Anti-programmed-death-receptor-1 treatment with pembrolizumab in ipilimumab-refractory advanced melanoma: A randomised dose-comparison cohort of a phase 1 trial. Lancet 2014, 384, 1109–1117. [Google Scholar] [CrossRef]
- Hodi, F.S.; O’Day, S.J.; McDermott, D.F.; Weber, R.W.; Sosman, J.A.; Haanen, J.B.; Gonzalez, R.; Robert, C.; Schadendorf, D.; Hassel, J.C.; et al. Improved Survival with Ipilimumab in Patients with Metastatic Melanoma. N. Engl. J. Med. 2010, 363, 711–723. [Google Scholar] [CrossRef]
- Thapa, B.; Kc, R.; Uludağ, H. TRAIL therapy and prospective developments for cancer treatment. J. Control. Release 2020, 326, 335–349. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, G.; Sun, M.; Lu, X. Targeting and exploitation of tumor-associated neutrophils to enhance immunotherapy and drug delivery for cancer treatment. Cancer Biol. Med. 2020, 17, 32–43. [Google Scholar] [CrossRef]
- Liu, Y.; Gu, Y.; Han, Y.; Zhang, Q.; Jiang, Z.; Zhang, X.; Huang, B.; Xu, X.; Zheng, J.; Cao, X. Tumor Exosomal RNAs Promote Lung Pre-metastatic Niche Formation by Activating Alveolar Epithelial TLR3 to Recruit Neutrophils. Cancer Cell 2016, 30, 243–256. [Google Scholar] [CrossRef]
- Reilley, M.J.; Morrow, B.; Ager, C.R.; Liu, A.; Hong, D.S.; Curran, M.A. TLR9 activation cooperates with T cell checkpoint blockade to regress poorly immunogenic melanoma. J. Immunother. Cancer 2019, 7, 1–9. [Google Scholar] [CrossRef]
- Schmid, M.C.; Khan, S.Q.; Kaneda, M.M.; Pathria, P.; Shepard, R.; Louis, T.L.; Anand, S.; Woo, G.; Leem, C.; Faridi, M.H.; et al. Integrin CD11b activation drives anti-tumor innate immunity. Nat. Commun. 2018, 9, 5379. [Google Scholar] [CrossRef]
- Lu H-H, Liu HW, Dinh TK, Huang C-H, Huang H-C, Tseng Y-C, et al. pH-Responsive, two-in-one doxorubicin and Bcl-2 siRNA-loaded micelleplexes for triple-negative breast cancer therapy. Polymer Chemistry. 2022, 13, 5568–5578. [CrossRef]
- Takahashi, M.; Saito, H.; Atsukawa, K.; Ebinuma, H.; Okuyama, T.; Ishii, H. Bcl-2 prevents doxorubicin-induced apoptosis of human liver cancer cells. Hepatol. Res. 2003, 25, 192–201. [Google Scholar] [CrossRef] [PubMed]
- 156. Li, L.; Zuo, X.; Xiao, Y.; Liu, D.; Luo, H.; Zhu, H. Neutrophil-derived exosome from systemic sclerosis inhibits the proliferation and migration of endothelial cells. Biochem. Biophys. Res. Commun. 2020, 526, 334–340. [Google Scholar] [CrossRef]
- Foo SS, Reading PC, Jaillon S, Mantovani A, Mahalingam S. Pentraxins and Collectins: Friend or Foe during Pathogen Invasion? Trends Microbiol. 2015, 23, 799–811. [PubMed]
- Jaillon, S.; Ponzetta, A.; Magrini, E.; Barajon, I.; Barbagallo, M.; Garlanda, C.; Mantovani, A. Fluid phase recognition molecules in neutrophil-dependent immune responses. Semin. Immunol. 2016, 28, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Zahid, K.R.; Raza, U.; Tumbath, S.; Jiang, L.; Xu, W.; Huang, X. Neutrophils: Musketeers against immunotherapy. Front. Oncol. 2022, 12, 975981. [Google Scholar] [CrossRef]
Figure 1.
Schematic showing development stages of neutrophils. Neutrophil development in bone marrow occurs in three phases: the stem cell pool, the mitotic pool, and the post-mitotic pool. The stem cell pool includes the hematopoietic stem cell (HSC) to the granulocyte and macrophage precursor (GMP). The mitotic pool holds the myeloblast, promyelocyte, and myelocyte stages. The third phase, the post-mitotic pool, covers the metamyelocyte to mature neutrophil stages. Different transcription factors come into play at different stages of neutrophil development. PU.1 promotes MPP conversion to LMP, while Runx1 and c-myb stimulate myelocyte proliferation and azurophilic granule synthesis. The downregulation of Runx1 and c-Myb promotes myeloblast to promyeloblast conversion. Only C/EBPα remains constant throughout the subsequent differentiation stages of neutrophils. c/EBP-β, c/EBP-γ, and c/EBP-δ levels increase as terminal differentiation proceeds. Stromal cells secrete ligands like VCAM-1 and CXCL-12 specific to the receptor to be displayed by mature neutrophils. Further developmental stages of neutrophils include the myeloblast, promyelocyte, myelocyte, metamyelocyte, and band cell stages. Finally, band cells are converted into mature neutrophils, which are released into circulation.
Figure 1.
Schematic showing development stages of neutrophils. Neutrophil development in bone marrow occurs in three phases: the stem cell pool, the mitotic pool, and the post-mitotic pool. The stem cell pool includes the hematopoietic stem cell (HSC) to the granulocyte and macrophage precursor (GMP). The mitotic pool holds the myeloblast, promyelocyte, and myelocyte stages. The third phase, the post-mitotic pool, covers the metamyelocyte to mature neutrophil stages. Different transcription factors come into play at different stages of neutrophil development. PU.1 promotes MPP conversion to LMP, while Runx1 and c-myb stimulate myelocyte proliferation and azurophilic granule synthesis. The downregulation of Runx1 and c-Myb promotes myeloblast to promyeloblast conversion. Only C/EBPα remains constant throughout the subsequent differentiation stages of neutrophils. c/EBP-β, c/EBP-γ, and c/EBP-δ levels increase as terminal differentiation proceeds. Stromal cells secrete ligands like VCAM-1 and CXCL-12 specific to the receptor to be displayed by mature neutrophils. Further developmental stages of neutrophils include the myeloblast, promyelocyte, myelocyte, metamyelocyte, and band cell stages. Finally, band cells are converted into mature neutrophils, which are released into circulation.
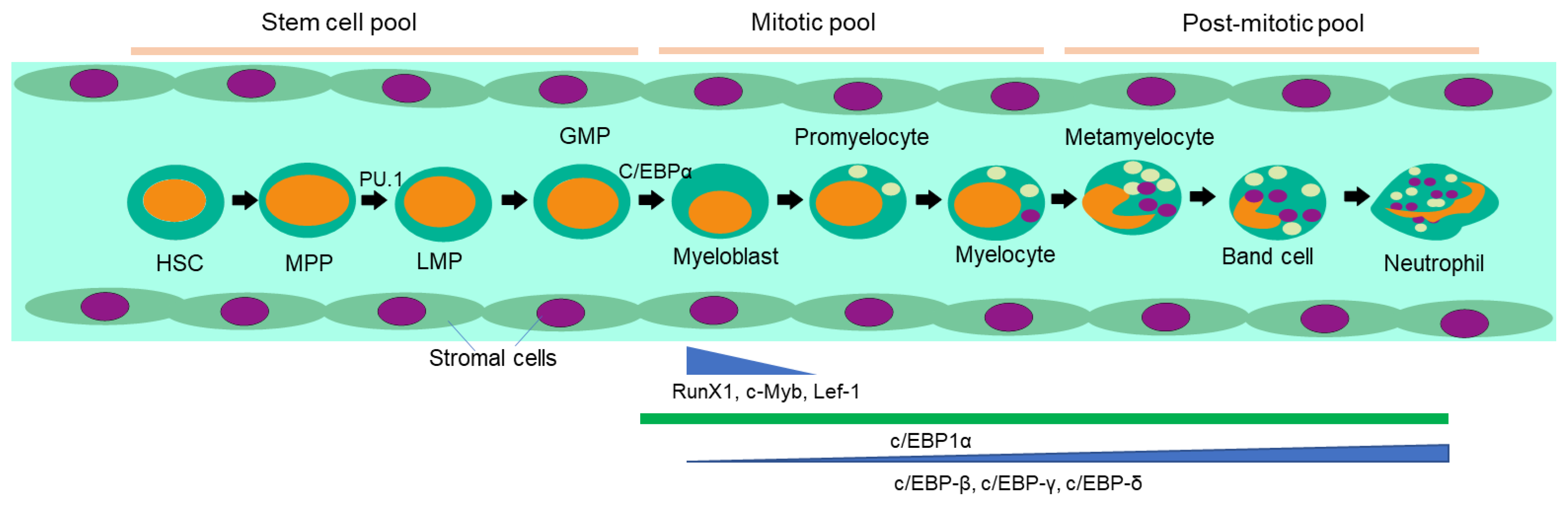
Figure 2.
Anti-tumor and pro-tumor activities of neutrophils. The N1 subtype helps to generate an anti-tumorigenic niche. These cells can kill tumor cells directly by secreting elastase, cathepsin G, and myeloperoxidase. They can also act as non-professional APCs to activate other immune cells. TLR-stimulated neutrophils generate neutrophil extracellular traps (NETs). FcR I and FcR II can bind to the Fc region of mAbs and promote antibody-dependent cellular cytotoxicity, phagocytosis, and trogocytosis of tumor cells. On the other hand, N2-TANs influence several mechanisms that promote tumor spread. These cells release MMP-9, which frees VEGF from the ECM and stimulates angiogenesis. N2 TANs also secrete cytokines (IL-1β, TNF-α, IL-6, and IL-12) that cause chronic inflammation and arginase 1, which blocks CD8+ T cells, prevents NK cells and APCs from reaching the tumor site or presenting antigens to cytotoxic T cells, creating an immunosuppressive environment. Moreover, they produce ROS that can cause DNA damage and genotoxic effects on tumor cells. Tumor cells also secrete G-CSF, IL-1, TNF-α, and IL-6, which promote the differentiation of N2 subtype neutrophils. The LTB4 and MMP9 secreted by neutrophils support metastasis-initiating cell survival and bring the tumor cells into circulation.
Figure 2.
Anti-tumor and pro-tumor activities of neutrophils. The N1 subtype helps to generate an anti-tumorigenic niche. These cells can kill tumor cells directly by secreting elastase, cathepsin G, and myeloperoxidase. They can also act as non-professional APCs to activate other immune cells. TLR-stimulated neutrophils generate neutrophil extracellular traps (NETs). FcR I and FcR II can bind to the Fc region of mAbs and promote antibody-dependent cellular cytotoxicity, phagocytosis, and trogocytosis of tumor cells. On the other hand, N2-TANs influence several mechanisms that promote tumor spread. These cells release MMP-9, which frees VEGF from the ECM and stimulates angiogenesis. N2 TANs also secrete cytokines (IL-1β, TNF-α, IL-6, and IL-12) that cause chronic inflammation and arginase 1, which blocks CD8+ T cells, prevents NK cells and APCs from reaching the tumor site or presenting antigens to cytotoxic T cells, creating an immunosuppressive environment. Moreover, they produce ROS that can cause DNA damage and genotoxic effects on tumor cells. Tumor cells also secrete G-CSF, IL-1, TNF-α, and IL-6, which promote the differentiation of N2 subtype neutrophils. The LTB4 and MMP9 secreted by neutrophils support metastasis-initiating cell survival and bring the tumor cells into circulation.
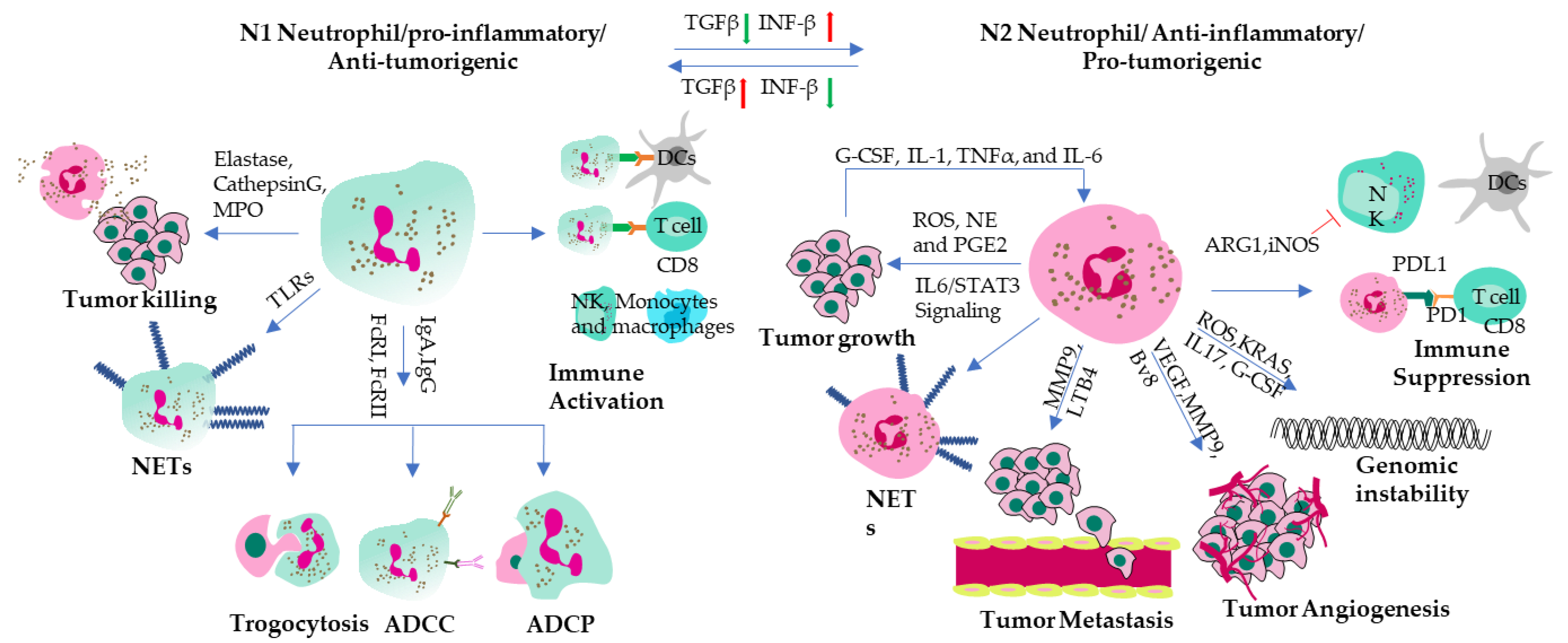
Figure 3.
Strategies to use NDEs as therapeutic tools: Tumor cells, N2 TANs, and TANs-derived NDEs can be targeted in several ways. Exosome release can be inhibited using different drugs that are involved in their formation and secretion, or by modulating their uptake by tumor cells, or by inducing uptake by phagocytic cells. NDEs can be used to deliver miRNAs to target oncogenic proteins like TGFβ in advanced stages of cancer. N2 TANs NDEs can also be directly targeted using drug-loaded antibodies or nanoparticles against CD63, CD9, CD81, and LFA1. Using specific antibodies like anti-PD-1 and CTLA4 can help in releasing the brakes on immune suppression. Tumors with mutated p53 can be targeted using Trail receptor loaded NDEs.
Figure 3.
Strategies to use NDEs as therapeutic tools: Tumor cells, N2 TANs, and TANs-derived NDEs can be targeted in several ways. Exosome release can be inhibited using different drugs that are involved in their formation and secretion, or by modulating their uptake by tumor cells, or by inducing uptake by phagocytic cells. NDEs can be used to deliver miRNAs to target oncogenic proteins like TGFβ in advanced stages of cancer. N2 TANs NDEs can also be directly targeted using drug-loaded antibodies or nanoparticles against CD63, CD9, CD81, and LFA1. Using specific antibodies like anti-PD-1 and CTLA4 can help in releasing the brakes on immune suppression. Tumors with mutated p53 can be targeted using Trail receptor loaded NDEs.
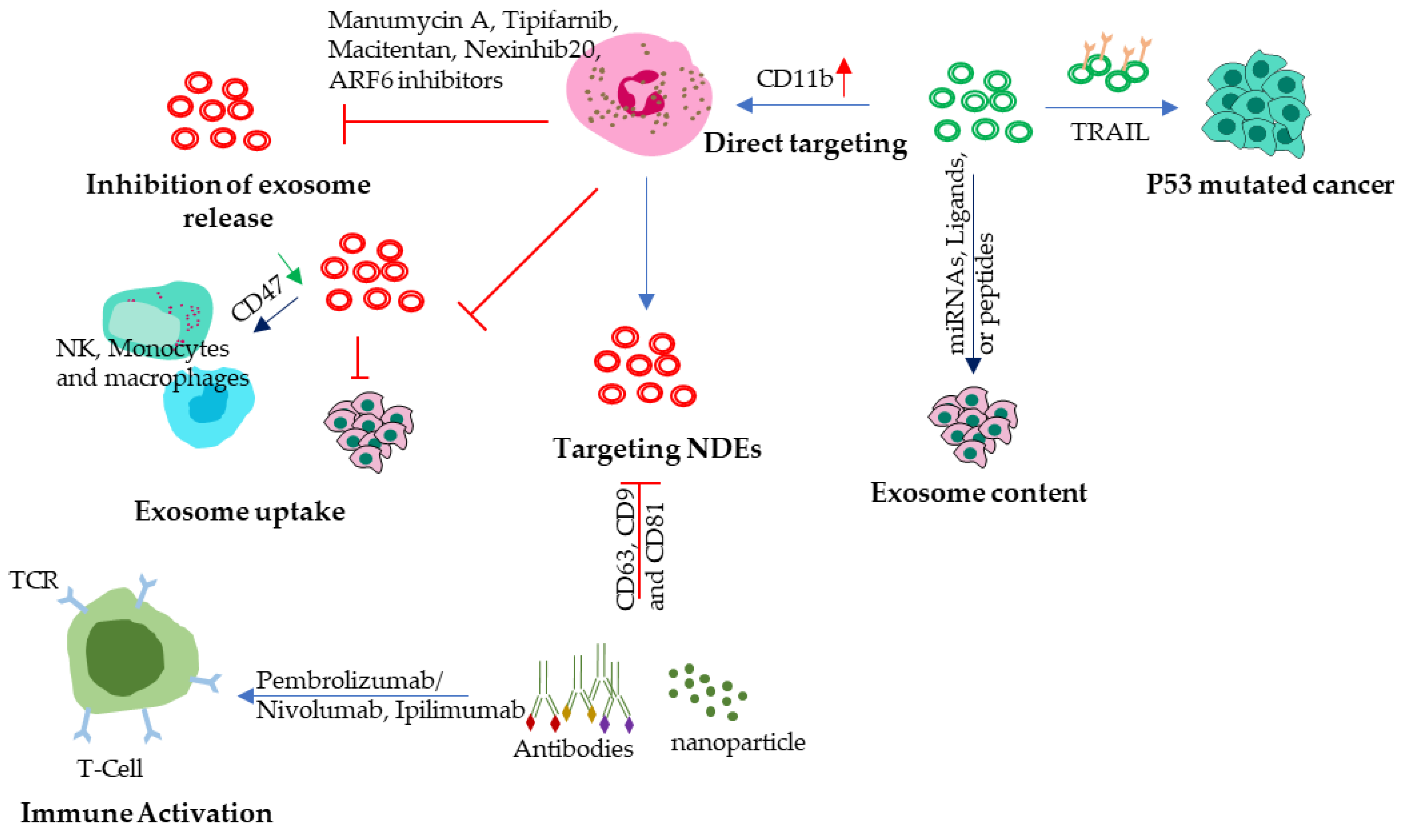
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



