Submitted:
11 April 2023
Posted:
12 April 2023
You are already at the latest version
Abstract
Ferns and fern-allies, now known as Monylophytes, have received scant molecular attention, in relation to angiosperm group. The advent of high-throughput technologies allows to advance towards a greater knowledge of their elusive genome. In this work, samples of apogamous and sexual heart-shaped gametophytes from two ferns: the apomictic species Dryopteris affinis ssp. af-finisand its sexual relative Dryopteris oreades were extracted and identified. In total, a set of 218 proteins shared by these gametophytes were analysed by using the STRING database, and the proteome associated to metabolism, genetic information processing and responses to abitotic stress is discussed. Specifically, there are reported proteins involved in metabolism of carbohy-drates and lipids, biosynthesis of amino acids, metabolism of nucleotides, energy, and secondary compounds, oxido-reduction, transcription, translation, folding, sorting, and degradation, and response to abiotic stress. Some homologs of proteins found are MACCI-BOU (MAB1), MOSAIC DEATH 1 (MOD1), MAINTENANCE OF PHOTOSYSTEM II UNDER HIGH LIGHT 2 (MPH2), TRANSPARENT TESTA 5 (TT5), ALBINO OR GLASSY YELLOW 1 (AGY1), LEUCYL AMINOPEP-TIDASE 1 and 3 (LAP1 and LAP3), or LOW EXPRESSION OF OSMOTICALLY RESPONSIVE GENES 1 (LOS1). The interactome of the set of proteins was also studied, being the most common interac-tions database and textmining. All these data about the interactions that exist between the stud-ied proteins of the ferns D. affinis and D. oreades, together with the description of their biological function, might contribute to better understand the functioning and development of ferns as well as to fulfil gaps of knowledge in plant evolution.
Keywords:
Dryopteris affinis ssp. affinis
; Dryopteris oreades
; fern
; gametophyte
; non-seed plants
; proteome
; STRING database
1. Introduction
Ferns and fern-allies, now known as Monylophytes, represent a genetic legacy of great value, being descendants of the first plants that developed vascular tissue, about 470 million years ago. They have received scant attention, concerning the angiosperm group, which relegated them to the background, after a splendid past. The appeal of its fronds or its use to alleviate ailments in traditional medicine is all they have traditionally inspired. Only a handful of species have been used to delve into basic developmental processes, such as photomorphogenesis (Wada, 2007), spore germination (Salmi et al., 2005; Salmi et al., 2007; Suo et al., 2015), cell polarity (Salmi et al., 2010), cell wall composition (Eeckhout et al., 2014), or reproduction, focussing on the gametophyte generation, which is an autonomous-living organism, easily for in vitro culture and sample collection (Fernández and Revilla 2003; Rivera et al., 2018). Although being an individual possessing a very simple structure of a one-cell layer, the gametophyte undergoes some degree of complexity: apical-basal polarity, dorsoventral symmetry, rhizoids, meristems in the apical or lateral part, reproduction organs (antheridia and archegonia, respectively), and trichomes distributed for the entire surface.
From a metabolic point of view, ferns contain secondary metabolites: flavonoids, alkaloids, phenols, steroids, etc., and exhibit various bioactivities such as antibacterial, antidiabetic, anticancer, antioxidant, etc. (Chen et al., 2015). The therapeutic use of pteridophytes is showing a great evolution, from its use in the traditional medicine of different peoples to the current stage, in which the principles of these plants are used in the form of nanoparticles (Femi-Adepoju et al., 2019). Finally, we recently assist the use of ferns to solve interesting problems in the plant world caused by biotic and abiotic stress. Drought is one of the most severe abiotic stress factors affecting plant growth and productivity, and ferns could contribute to managing it (Wang et al., 2010). Other important adaptations of ferns to extreme environments such as salinity, heavy metal, epiphytes, or invasiveness tolerance, are summarized by Rathinasabapathi (2006). Likewise, Dhir (2018) highlights the high efficiency of many species of aquatic and terrestrial ferns, to extract various organic and inorganic pollutants from the environment.
Increasingly, researchers become more interested in these plants, and sometimes this is possible by the advent of high-throughput technologies, such as omics, allowing to advance towards a greater knowledge of their elusive genome. The variation in gene expression, induced by whatever environmental or inner conditions, can be examined in non-model organisms because these techniques have become more feasible as automation and efficiency have reduced costs. Until present, some transcriptome and proteome datasets have been published for ferns, which include the species Pteridium aquilinum (Der et al., 2011), Ceratopteris richardii (Salmi et al., 2010; Cordle et al., 2012), Blechnum spicant (Valledor et al., 2014), Lygodium japonicum (Aya et al., 2015), Dryopteris affinis ssp. affinis (Grossmann et al., 2017; Wyder et al., 2020; Fernández et al., 2021), and Dryopteris oreades (Fernández et al. 2021; Ojosnegros et al. 2022). In the last case, both transcriptomic and proteomic analyses were performed by using next-generation sequencing (NGS) and shotgun proteomics by tandem mass spectrometry.
This work expands our knowledge of the proteomic data available in non-seed plants, less explored so far than in seed plants. It is the continuation of a previous work (Ojosnegros et al., 2022) in which samples of apogamous and sexual heart-shaped gametophytes from two ferns: the apomictic species D. affinis and its sexual relative D. oreades, were extracted and identified by using a species specific transcriptome database established in a previous project (Grossmann et al., 2017). The functional annotation was inferred by blasting identified full length protein sequences. The categorization of the proteins having in common both types of gametophyte is reported. Specifically, this work reveals new proteomic information involved in the metabolism of carbohydrates and lipids, the biosynthesis of amino acids, the metabolism of nucleotides and energy, as well as of secondary compounds, such as flavonoids, terpenoids, lignans, etc., important in plant defence against stress, in addition to the oxide-reduction processes; it also reveals proteins related to transcription, translation, folding, sorting or transport, and degradation.
2. Results
A set of 218 proteins shared by the gametophytes of the apomictic fern D. affinis and its sexual relative D. oreades were analyzed by using the STRING version 11.5, and the CYTOSCAPE version 3.9.1 programs. To gain insights into their biological function, Gene Ontology (GO) and KEGG classification provided by the STRING platform were performed (Figure 1 and Figure 2). From the GO classification, most of the proteins found are involved in biological functions linked to the primary metabolism, and then, more specifically to other cellular processes, such as response to stimulus, protein metabolism, translation, etc.
In turn, the KEGG classification reveals the presence of proteins mostly associated with the biosynthesis of secondary metabolites, the ribosome, and the biosynthesis of amino acids. These processes include the building of cellular organelles such as the ribosomes (Figure 3), or the proteasomes (Figure 4). Regarding the ribosomes, there are several proteins such as nucleic acid-binding proteins, ribosomal proteins, translation elongation factors, etc. On the other hand, the proteasomes intervene in the degradation of proteins, being anotated proteins linked to the proteolytic core: the 20S particle, and also regulatory factors.
The protein domains more abundant in the studied groups are the pyruvate dehydrogenase E1 component, and the histidine and lysine active sites of the phosphoenolpyruvate carboxylase, related to carbohydrates metabolism. In the biosynthesis of amino acids, the domains more abundant are the aspartate aminotransferase and the pyridoxal phosphate-dependent transferase. In the metabolism of energy, the HAS barrel domain, and the F1 complex of the alpha and beta subunits of the ATP synthase. In the metabolism of secondary compounds, the aromatic amino acid lyase, the phenylalanine ammonia-lyase, and the N-terminal of the histidase. Finally, the domains more frequent in transcription and translation are the GTP-binding domain and domain 2 of the elongation factor Tu, and the conserved sites of the ribosomal proteins S10 and S4.
Looking at the interactome of this set of proteins, it represents a total network composed of 218 nodes and 1,792 interactions (p-value < 1 x e-16). In the group metabolism of carbohydrates, the proteins showing more interactions are CYTOSOLIC ENOLASE (ENOC) and PHOSPHOGLYCERATE KINASE 1 (PGK1), with 12 interactions each one; in the biosynthesis of amino acids is AT1G14810 with 6 interactions; in the metabolism of energy is ATPC1 with 20 interactions; in the metabolism of secondary compounds is 4-COUMARATE: COA LIGASE 3 (4CL3) with 3 interactions; in transcription and translation are SUPPRESSOR OF ACAULIS 56 (SAC56), RIBOSOMAL PROTEIN US11X (US11X), and RIBOSOMAL PROTEIN US17Y (US17Y) with 44 interactions each one; and finally, in transport is RAB GTPASE HOMOLOG 1A (RAB1A) with 4 interactions.
The strength of the interactions can be weak or strong, (Table S1) according to a scale from 0 to 1, where a weak interaction will have a score close to 0 and a strong interaction a score close to 1. Taking into account only in each group of proteins studied, the number of interactions with a total score equal to or greater than 0.99, the results show that the proteins of transcription and translation are the ones that have more strongest interactions: 554, next proteins involved on energy with 29 interactions, carbohydrates with 16 interactions, and finally biosynthesis of amino acids, and transport with 5 and 3, respectively.
In addition to the number of interactions and the strength of interactions, we considered also in this study the type of interactions that exist between the proteins, provided by STRING (Figure 5), which derived from textmining, experiments, co-expression, and databases. Specifically, we analyzed the groups of metabolism of carbohydrates (Figure 5a), metabolism of energy (Figure 5b), ribogenesis (Figure 5c), and protein degradation (Figure 5d).
Paying attention only the two main types of interactions of each group of proteins, their relations were analyzed (Figure 6). Hence, database and text mining in metabolism of carbohydrates (Figure 6a), the biosynthesis of amino acids (Figure 6b), and also in the metabolism of secondary compounds, and transport; textmining and co-expression in the metabolism of energy (Figure 6c), and experiments interactions and co-expression in transcription and translation (Figure 6d). In the metabolism of carbohydrates and transcription and translation, the two interactions are alike, meanwhile, in the biosynthesis of amino acids and the metabolism of energy, one interaction is stronger than the other, specifically the textmining interaction in the first group and the co-expression interaction in the second. On the other hand, comparing the same type of interaction among the different groups of proteins, it is observed that the neighborhood interaction is higher in the biosynthesis of amino acids and transcription and translation; gene fusion in the metabolism of carbohydrates and the biosynthesis of amino acids; co-occurrence in the biosynthesis of amino acids and the metabolism of secondary compounds; co-expression in the metabolism of energy and transcription and translation; experiments in transcription and translation, and transport; database in the metabolism of carbohydrates and transport; and finally, textmining in the metabolism of secondary compounds and transport.
3. Discussion
In the context of a proteome commitment carried out in the gametophyte generation of the species of ferns: the apomictic D. affinis and its sexual relative D. oreades, the current work provides additional information about the proteome and gives continuity to previous studies that dealt with these ferns (Grossmann et al., 2017; Wyder et al., 2020; Fernández et al., 2021; Ojosnegros et al., 2022). Specifically, the proteins discussed here are grouped into two major categories to make easier the reading: metabolism and genetic information processing (Table 1). Their biological functions and protein-protein interactions are reported next.
Metabolism comprises two main branches: primary and secondary. Primary metabolism connects to the main metabolites directly involved in plant growth (carbohydrates, lipids, amino acids, nucleotides), as well as those reactions operating in fueling them such as photosynthesis, glycolysis, tricarboxylic acid cycle, etc. In plants, there is also a secondary metabolism, which connects with other transcendental metabolic routes, governing in most cases the defence and stress responses, needed to survive, specifically when the organism is anchored to the underground.
Proteins linked to the "metabolism of carbohydrates" turn around glycolysis, pyruvate metabolism, citrate/tricarboxylic acid cycle, pentose phosphate pathway, starch, and biosynthesis of nucleotide sugars. Glycolysis converts glucose in pyruvate, and in the gametophytes of the study, there are reported enzymatic proteins such as ATP-DEPENDENT 6-PHOSPHOFRUCTOKINASE 3(PFK3), involved in the first reaction; two enzymes participating in glycolysis and gluconeogenesis: FRUCTOSE-BISPHOSPHATE ALDOLASE 3 (FBA3), also named PIGMENT DEFECTIVE 345 (PDE345), and FRUCTOSE-BISPHOSPHATE ALDOLASE 8 (FBA8); and other catalyzing the decarboxylation of pyruvate to acetyl-CoA such as the mitochondrial component of pyruvate dehydrogenase MACCI-BOU (MAB1). Linked to pyruvate metabolism there are reported two phosphoenolpyruvate carboxylases (PPC2 and PPC3), which supply oxaloacetate for the tricarboxylic acid cycle, and the protein NAD-DEPENDENT MALIC ENZYME 1 (NAD-ME1), which is involved in the regulation of the metabolism of sugars and amino acids during the night (Tronconi et al., 2008). We mention here the protein 2,3-BIPHOSPHOGLYCERATE-INDEPENDENT PHOSPHOGLYCERATE MUTASE 1 (iPGAM1), also important in the functioning of stomatal guard cells and fertility in A. thaliana (Zhao et al., 2011).
Likewise, we identified some proteins associated with the citrate/tricarboxylic acid cycle: AT2G20420 and AT5G08300, involved in the only phosphorylation step at the substrate level of this cycle. Other protein is the cytosolic MALATE DEHYDROGENASE 1 (c-NAD-MDH1), which catalyzes a reversible NAD-dependent dehydrogenase reaction involved in central metabolism and redox homeostasis between organelle compartments (Tomaz et al., 2010). In parallel with glycolysis, it is the pentose phosphate pathway, generating NADPH and pentoses. This metabolic pathway is represented here by the proteins 6-PHOSPHOGLUCONATE DEHYDROGENASE 1 (PGD1), GLUCOSE-6-PHOSPHATE DEHYDROGENASE 6 (G6PD6), and PGK1. A mutation in the gene of the first protein might decrease cellulose synthesis and alters the structure and composition of the primary cell wall (Howles et al., 2006). G6PD6 is important for the synthesis of fatty acids and nucleic acids involved in membrane synthesis and cell division (Wakao et al., 2005). PGK1 contributes too to triggering the phosphorylation of the proteins FTSZ2-1 and FTSZ2-2, required for chloroplast division (Gargano et al., 2012). In addition, Starch is the main reserve form of carbohydrate and energy in plants, being accumulated in chloroplasts during the day, and transported and degraded to provide energy and nutritional substances for growth and metabolism. The gametophytes of our ferns count for proteins involved in its synthesis, includingSTARCH BRANCHING ENZYME 2.2 (SBE2.2), andGRANULE BOUND STARCH SYNTHASE 1 (GBSS1), required this last together with STARCH DIRECTED PROTEIN (PTST) for amylose synthesis (Seung et al., 2015).
Apart from to the proteins mentioned above, others were found associated with the biosynthesis of nucleotide sugars such as two pyrophosphorylases (ADG1 and APL1); the protein REVERSIBLY GLYCOSYLATED POLYPEPTIDE 4 (RGP4), involved in the synthesis of non-cellulosic polysaccharides of the cell wall (Rautengarten et al., 2011); and AT5G20080.
Regarding the "metabolism of lipids", three proteins are reported in this study. The first protein is MOSAIC DEATH 1 (MOD1), which catalyzes the last reduction step of the de novo fatty acid synthesis cycle and the fatty acid elongation cycle. An alteration in the gene causes a decrease in the activity of the protein, reducing the number of fatty acids, that triggers mosaic premature cell death, and finally, it changes the plant morphology, such as chlorotic and curly leaves, distorted siliques, and dwarfism (Mou et al., 2000). The second protein is ATP-CITRATE LYASE A-1 (ACLA-1), necessary for the normal growth and development of plants because synthesizes acetyl-coA, a key compound functioning in many metabolic pathways (fatty acids and glucosinolates in chloroplasts; flavonoids, sterols, and phospholipids in the cytoplasm; and ATP and amino acids carbon skeletons in mitochondria). Moreover, it is the substrate for histone acetylation and transcription factors in the nucleus and regulates their function to maintain or alter chromosome structure and transcription (Fatland et al., 2002; Fatland et al., 2005). The third protein is CITRATE SYNTHASE 2 (CSY2), which synthesizes citrate in peroxisomes for the respiration of fatty acids in seedlings, and being also needed for seed germination (Pracharoenwattana et al., 2005).
Involved in the biosynthesis of aminoacids, are reported the proteins aminotransferase ASP1; the ISOPROPYL MALATE ISOMERASE LARGE SUBUNIT 1 (IIL1), which act in the glucosinolate biosynthesis, molecules used by the plant to defend itself against insect attacks (Knill et al., 2009); the GLUTAMATE SYNTHASE 1 (GLU1), needed also for the re-assimilation of ammonium ions generated during photorespiration (Ishizaki et al., 2009); and SERINE HYDROXYMETHYLTRANSFERASE 3 (SHM3), HISTIDINOL DEHYDROGENASE (HDH), and METHIONINE OVER-ACCUMULATOR 3 (MTO3), which catalyze the formation of glycine, L-histidine, and methionine, respectively (Zhang et al., 2010; Petersen et al., 2010; Shen et al., 2002).
It is reported here some proteins associated with the"metabolism of nucleotides", specifically to the AMP synthesis, such as ADENOSINE KINASE 1 (ADK1) and ADENYLOSUCCINATE SYNTHASE (ADSS). It deserves to be also mentioned the protein DEFECTIVE 3119 (EMB3119), essential in the synthesis of numerous compounds such as purines, pyrimidines, aromatic amino acids, NAD, and NADP (Howles et al., 2006).
As it is well known, metabolism demands "energy" in the form of ATP obtained from nutrients, and it comprises a series of interconnected pathways that can function in the presence or absence of oxygen. In the gametophyte of our ferns, several mitochondrial, chloroplastic, and vacuolar ATP synthases engaged in the oxidative phosphorylation process are found. Additionally, chemical energy can be obtained through photosynthesis. The list of annotated proteins includes the protein FERREDOXIN-NADP(+)-OXIDOREDUCTASE 1 (FNR1), which regulates the flow of electrons to meet the demands of the plant for ATP and reduction power (Lintala et al., 2007), and others involved in repairing DNA damage such as DNA-DAMAGE-REPAIR/TOLERATION PROTEIN 112 (DRT112) (Kieselbach et al., 2000), PHOTOSYSTEM II SUBUNIT O-2 (PSBO2), which regulates the replacement of the protein D1 impaired by light (Lundin et al., 2007), and the protein maintenance OF PHOTOSYSTEM II UNDER HIGH LIGHT 2 (MPH2). This protein is necessary to carry out photosynthesis correctly and efficiently in two conditions: controlled photoinhibitory light and fluctuating light, since in nature plants experience rapid and extreme changes in sunlight, giving them an adaptive advantage (Liu et al., 2017). Involved in photosynthesis, there is the protein PHOTOSYNTHETIC NDH SUBCOMPLEX L 5 (Pnsl5), which modulates the conformation of the protein BRASSINAZOLE-RESISTANT 1 (BZR1) (Zhang et al., 2013). This protein binds to the promoter of the FLOWERING LOCUS D (FLD), suppresses its expression, and then FLOWERING LOCUS C (FLC) can repress flowering (Zhang et al., 2013). Finally, AT3G48420 degrades xylulose-1,5-bisphosphate, a potent inhibitor of the protein rubisco produced by itself (Bracher et al., 2015). On the other hand, photorespiration represents a waste of the energy produced by photosynthesis. The enzyme GLYCERATE KINASE (GLYK) catalyzes the final reaction of photorespiration allowing terrestrial plants to grow in an atmosphere with oxygen, reflecting the evolutionary origin of photosynthesis in an anaerobic environment towards an aerobic currently (Boldt et al., 2005). Another important protein in photorespiration is ALANINE GLYOXYLATE AMINOTRANSFERASE (AGT), which participate also in root development, both primary and lateral, in seedlings once they have germinated (Wang et al., 2019).
Likewise, there are some proteins about sulfur and nitrogen metabolism. The first one is represented by two proteins: SULFOQUINOVOSYLDIACYLGLYCEROL 1 (SQD1), which converts UDP-glucose and sulfite to the precursor of the main group of sulfolipids: UDP-sulfoquinovose, preventing it from accumulating in the cell as it is very toxic (Sanda et al., 2001); and CHLOROPLAST SULFUR E (CPSUFE), a sulfur acceptor that activates cysteine desulfurases in plastids and mitochondria, vital during embryogenesis (Ye et al., 2006). As to nitrogen metabolism, there are the proteins GLNB1 HOMOLOG (GLB1), which is a nitrogen regulatory protein and intervenes in glycosaminoglycan degradation (Ferrario-Méry et al., 2008); and NITRITE REDUCTASE 1 (NIR1), which catalyzes the reduction of nitrite to ammonium (Takahashi et al., 2001). It has been seen that if the amount of this protein in the cell increases, the tolerance and assimilation of nitrogen dioxide by the plant improve, and as nitrogen dioxide is an air pollutant caused largely by motorized vehicles, the plant could act as a sink for this substance, i.e., it could be used as a biotechnological application of bioremediation (Takahashi et al., 2001).
Concerning the “metabolism of secondary compounds”, proteins related to flavonoid biosynthesis are glossed in this work, such as TRANSPARENT TESTA 5 (TT5), responsible for the isomerization of chalcones into naringenin (Shirley et al., 1992). We found too enzymes involved in the biosynthesis of terpenoids: GERANYLGERANYL PYROPHOSPHATE SYNTHASE 1 (GGPS1); the biosynthesis of lignans: PHENYLCOUMARAN BENZYLIC ETHER REDUCTASE 1 (PCBER1); and the biosynthesis of phenylpropanoids: CINNAMYL ALCOHOL DEHYDROGENASE 9 (CAD9), very important in cell wall formation (Eudes et al., 2006). Also, the protein 4CL3, which produces coA thioesters of hydroxy- and methoxy-substituted cinnamic acids, is used to synthesize anthocyanins, flavonoids, isoflavonoids, coumarins, lignin, suberin, and phenols (Ehlting et al., 1999); and AT2G45300, which intervenes in the synthesis of chorismate, the precursor of the amino acids phenylalanine, tryptophan, and tyrosine (Klee et al., 1987). Linked to "oxido-reduction" processes, they are the proteins: GLYOXALASE 2-5 (GLX2-5), involved in cell detoxification in mitochondria (Marashinghe et al., 2005); the mitochondrial MALATE DEHYDROGENASE 1 (mMDH1), participating in redox homeostasis between organelle compartments, may limit photorespiration during the dark phase, and it is required for partitioning of carbon dioxide and energy in leaves (Lindén et al., 2016); and the transferasesGSTL2 and GSTL3, which catalyze the glutathione-dependent reduction of S-glutathionyl quercetin to quercetin (Dixon et al., 2010).
3.2. Genetic Information Processing
The processing of genetic information comprises transcription, translation, folding, sorting or transport, and degradation. In our gametophytes, two proteins are involved in "transcription", specifically the 14-3-3-like proteins: GF14 nu (GRF7) and GF14 iota (GRF12), which are associated with a DNA-binding complex that binds to the G-box, a DNA regulatory element (Rosenquist et al., 2001). Besides, and telated to "translation": RNA-BINDING GLYCINE-RICH PROTEIN A7(RBGA7), which has a role in RNA processing during stress, specifically in editing cytosine to uracil in mitochondrial RNA, controlling 6 % of mitochondrial editing sites (Shi et al., 2015); and others such as AT1G03510; RNA-BINDING PROTEIN 47B (RBP47B); and UBP1-ASSOCIATED PROTEIN 2A (UBA2A), which regulates mRNA and stabilize RNA in the nucleus (Lambermon et al., 2002). Apart from several ribosomal subunits, there are others involved in elongation, like the protein LOW EXPRESSION OF OSMOTICALLY RESPONSIVE GENES 1 (LOS1), which is also involved in the response to cold (Guo et al. 2002).
Once the proteins have been formed, there is a control of quality to check that they have been synthesized completely and have been folded correctly. Among the proteins playing a major role in the acceleration of folding or the degradation of misfolded proteins are GROES, AT3G10060, AT2G43560, AT2G21130, etc. The gametophyte of ferns harbor proteins linked to the sorting or transport of molecules within the cell and between inside and outside of cells. In line with it, there are ALBINO OR GLASSY YELLOW 1 (AGY1), which has a role in coupling ATP hydrolysis with protein transfer across the thylakoid membrane, participating in photosynthetic acclimation and chloroplast formation (Skalitzky et al., 2011), RAN BINDING PROTEIN 1(RANBP1), moving proteins into the nucleus (Haizel et al., 1997), IMPORTIN ALPHA ISOFORM 2 (IMPA-2), acting on nuclear localization (Bhattacharjee et al., 2008), and the proteins ADP/ATP CARRIER 2 (AAC2), mediating the import of ADP into the mitochondrial matrix (Haferkamp et al., 2002), and TRANSLOCON AT THE INNER ENVELOPE MEMBRANE OF CHLOROPLASTS 62 (Tic62), into the chloroplasts (Küchler et al., 2002). In addition, we found others associated with the transport of water and small hydrophilic molecules through the cell membrane: PLASMA MEMBRANE INTRINSIC PROTEIN 1;4 (PIP1;4) (Lee et al. 2012), and VOLTAGE DEPENDENT ANION CHANNEL 3 (VDAC3) (Berrier et al., 2015). There are also proteins bound to dilysin motifs and associated with clathrin-uncoated vesicles that are transported from the endoplasmic reticulum to the Golgi apparatus and vice versa: ALPHA1 COAT PROTEIN (alpha1-COP), GAMMA2 COAT PROTEIN (gamma2-COP), and AT5G05010. Instead, the proteins EPSIN2 (EPS2) and DYNAMIN-LIKE 3 (DL3) are related to clathrin-coated vesicles, and the last one participates also in root hair positioning during planar polarity formation in root hair-forming cells (Stanislas et al., 2015). Reviewing our proteomic profile we found proteins such as GUANOSINE NUCLEOTIDE DIPHOSPHATE DISSOCIATION INHIBITOR 1 (GDI1), which regulates the GDP/GTP exchange reaction of most RAB proteins by inhibiting GDP dissociation and subsequent GTP binding (Zarsky et al., 1997).
On the other hand, many proteins found are related to protein catabolism or degradation, being subunits of proteasomes, i.e., complexes characterized by their ability to degrade proteins or ubiquitination structures. Others such as AT1G09130; CLP PROTEASE PROTEOLYTIC SUBUNIT 2 (CLP2); CLP PROTEASE R SUBUNIT 4 (CLPR4); LON PROTEASE 1 (LON1); PRESEQUENCE PROTEASE 1 (PREP1), which degrades in mitochondria the pre-sequences of proteins that have entered after being cut, since the other option, export them to the cytoplasm, costs a lot of energy and has low efficiency (Stahl et al., 2002); and also DEGRADATION OF PERIPLASMIC PROTEINS 9 (DEG9), which degrades the A. thaliana response regulator 4 (ARR4), a regulator which participates in light and cytokinin signaling pathways by modulating the activity of phytochrome B (Ouyang et al., 2017). Plants have to deal with heat stress, and for this, the gametophytes of the study count with the aminopeptidases LEUCYL AMINOPEPTIDASE 1 and 3 (LAP1 and LAP3), probably involved in the processing and turnover of intracellular proteins, and function as molecular chaperones protecting them from heat-induced damage (Scranton et al., 2012).
3.3. Protein-Protein Interactions
Although previously the interactions that exist between the proteins of some studied selected groups, have already been commented on, it is necessary to define all the types of interactions that exist clearly for a better understanding. The interactions between proteins can be of various types: (a) Experiments interaction: referred to proteins that have been shown to have chemical, physical, or genetic interactions in laboratory experiments; (b) Databases interaction: related to proteins found in the same databases; (c) Textmining interaction: occurs between proteins that are mentioned in the same PubMed abstracts or the same articles of an internal selection of the software STRING; (d) Co-expression interaction: indicates that the gene expression of those proteins is related; (e) Neighborhood interaction: between proteins whose genes are close in the genome; (f) Gene fusion interaction: indicates that in at least one organism the orthologous genes of the genes for those proteins have been fused into a single gene; and the last, (g) Co-occurrence interaction: related to proteins that have a similar phylogenetic distribution (Crosara et al., 2018).
Analyzing with the STRING platform thoroughly one by one the interactions of the groups of proteins studied, it is observed that in the metabolism of carbohydrates, co-expression, textmining and experiments interactions are stronger between the proteins AT2G20420 and AT5G08300, and database interaction between AT2G20420 and E2-OGDH1. AT2G20420 and AT5G08300 are both mitochondrial succinate-coA ligase subunits, and E2-OGDH1 catalyzes the conversion of 2-oxoglutarate to succinyl-CoA and CO2, i.e., the three proteins intervene in the tricarboxylic acid cycle (Condori-Apfata et al., 2021). Among the proteins of biosynthesis of amino acids, co-expression interaction is stronger between the proteins AT1G14810 and DIHYDROXYACID DEHYDRATASE (DHAD); and database interaction between DHAD and 2-ISOPROPYLMALATE SYNTHASE 1 (IMS1), between IIL1 and IMS1, and between IIL1 and ISOPROPYLMALATE DEHYDROGENASE 2 (IMD2). In fact, these proteins are involved in the synthesis of numerous compounds necessary for the plant: AT1G14810 for lysine, threonine, and methionine (Zhang et al., 2018); DHAD for isoleucine and valine (Yan et al., 2018); IMS1 and IMD2 for leucine (de Kraker et al., 2007; He et al., 2011); and IIL1 for glucosinolate (Knill et al., 2009). In the metabolism of energy, co-expression interaction is stronger between the proteins ATPC1 and GLYCERALDEHYDE 3-PHOSPHATE DEHYDROGENASE A SUBUNIT 2 (GAPA-2), and between DRT112 and FED A; experiments interaction between PSAA and PSAC, because in the photosynthesis the C-terminal of PSAC interacts with PSAA and other proteins such as PSAB and PSAD for its assembly into the photosystem I complex (Varotto et al., 2000); database interaction between AGT and GLYCOLATE OXIDASE 2 (GOX2), due to both proteins are necessary for photorespiration (Wang et al., 2019); and finally: textmining, between PSBO2 and PHOTOSYSTEM II SUBUNIT P-1 (PSBP-1), as the two proteins are chloroplastic oxygen-evolving enhancers that form part of the photosystem II (Lundin et al., 2007).
Continuing with the analysis of the interactions between the proteins of the groups studied, in the fourth group: the metabolism of secondary compounds, textmining interaction is the strongest, it occurs between PHE AMMONIA LYASE 1 (PAL1) and PHENYLALANINE AMMONIA-LYASE 4 (PAL4). Both proteins participate in the synthesis from phenylalanine of numerous compounds based on the phenylpropane skeleton, necessary for the plant's metabolism (Cochrane et al., 2004). Regarding: transcription and translation, co-expression interaction is stronger between ribosomal proteins such asEL34Z and UL22Z, RPL23AB, UL11Z, EL14Z, and RPL18; among a long list of proteins forming ribosomes, essential in translation. Finally, in transport, co-expression interaction is stronger between the proteins alpha1-COP and gamma2-COP; experiments interaction between alpha1-COP and AT5G05010; and textmining between AGY1 and GET3B.
As indicated in the results, in the group metabolism of carbohydrates, the proteins with more interactions are ENOC and PGK1, because they are both involved in the process of glycolysis (Li et al., 2019). In the biosynthesis of amino acids is AT1G14810, as it intervenes in several pathways of biosynthesis: lysine, isoleucine, methionine, and threonine (Zhang et al.,2018). In the following group: the metabolism of energy, is ATPC1, surely due to it being an ATP synthase chloroplastic (Takagi et al., 2017). The protein 4CL3 is the one with more interactions in the group metabolism of secondary compounds. It is key in the synthesis of numerous secondary metabolites, such as anthocyanins, flavonoids, isoflavonoids, coumarins, lignin, suberin, and phenols (Ehlting et al., 1999). In transcription and translation are the ribosomal proteins SAC56, US11X, and US17Y, necessary for the formation of ribosomes, and thus, for the translation (Carroll et al., 2008). Finally, in transport is RAB1A, as it participates in intracellular vesicle trafficking and protein transport (Ito et al., 2011).
All these data of the different interactions that exist between the studied proteins of the ferns D. affinis and D. oreades, together with the description of the possible biological function associated, contribute to expanding the scarce information that exists to date on the development and functioning of these two species of ferns.
4. Materials and Methods
4.1. Plant Material and Growth Conditions
Spores of D. affinis were obtained from sporophytes growing in Turón valley (Asturias, Spain), 477 m.a.s.l., 43º 12’ 10 N−5º 43’ 43 W. In the case of D. oreades, spores were collected from sporophytes growing in Burgos, Neila lagoons, 1.920 m a.s.l., 42º 02’ 48N−3º 03’ 44 W. Spores were released from sporangia, soaked in water for 2 h, and then washed for 10 min with a solution of NaClO (0.5%) and Tween 20 (0.1%). Then, they were rinsed three times with sterile, distilled water. Spores were centrifuged at 1,300 g for 3 min between rinses and then cultured in 500 mL Erlenmeyer flasks containing 100 mL of liquid Murashige and Skoog (MS) medium (Murashige and Skoog, 1962). Unless otherwise noted, media were supplemented with 2% sucrose (w/v), and the pH was adjusted to 5.7 with 1 or 0.1 N NaOH. The cultures were kept at 25 ºC under cool-white fluorescent light (70 µmol m-2s-1) with a 16 h photoperiod and put on an orbital shaker (75 rpm).
Following spore germination, gametophytes develop as filamentous structures. Then, they were subcultured to 200 mL flask containing 25 mL of MS medium supplemented with 2% sucrose (w/v) and 0.7% agar. The gametophytes of D. affinis become two-dimensional, achieving the spatulate and heart shapes after 20 or 30 additional days, respectively. Gametophytes of D. oreades grow up slower and needed around six months to become cordate and reach sexual maturity (Figure 7). By using light microscopy, apogamous and sexual gametophytes were collected. In the first case, they have only female reproductive organs (i.e. archegonia); and in the case of D. affinis, cordate gametophytes have visible signs of an evolving apogamic center, composed of smaller and darker isodiametric cells. Samples of apogamous and sexual cordate gametophytes were weighed before and after being lyophilized for 48h (Telstar-Cryodos), and stored in Eppendorf tubes in a freezer at -20 ºC until required.
4.2. Protein Extraction, Separation, and In-Gel Digestion
From the cordate apogamous and cordate sexual gametophytes (three samples each), an amount of 20 mg dry weight of gametophytes was homogenized using a Silamat S5 shaker (Ivoclar Vivadent, Schaan, Liechtenstein). The protocol used for protein extract, separation, and in-gel digestion was reported by Fernández et al., (2021).
4.3. Protein Identification, Verification, and Bioinformatic Downstream Analyses
Mass spectrometry and peptide identification (Orbitrap XL) were performed accordingly (Grossmann et al., 2017). The peptide FDR and protein FDR were estimated at 2% and 1%, respectively, indicating the stringency of the analyses. To have a functional understanding of the identified proteins, we blasted the whole protein sequences of all identified proteins against S. moellendorffii and A. thaliana Uniprot sequences and we then retrieved the best matching identifier from each of them, along with the corresponding e-value, accepting blast-hits which e-value below 1E-7. These better-annotated orthologue identifiers are then used in further downstream analysis.
4.4. Protein Analysis Using the STRINGPlatform
The identifiers of the genes from the apogamous and sexual gametophyte samples were used as input for STRING platform version 11.5 analysis and a high threshold (0.700) was selected for positive interaction between a pair of genes.
5. Conclusions
The analysis of a set of 218 proteins shared by the gametophytes of the apomictic fern D. affinis and its sexual relative D. oreades reveals the presence of proteins involved in biological functions mostly associated with metabolism, and also the processing of genetic information. Some smaller groups were studied in detail: metabolism of carbohydrates, biosynthesis of amino acids, metabolism of energy and secondary compounds, transcription, translation, and transport. The interactions between the proteins were identified, the most common source of them derived from database and textmining information. The proteins involved in transcription and translation exhibit the strongest interactions. All this information about the description of the possible biological functions and the different protein-protein interactions expands the existing current knowledge about these species of ferns and plants in general.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1. Strongest STRING interactions of proteins extracted from gametophytes of D. affinis and D. oreades and classified into the following groups: metabolism of carbohydrates, biosynthesis of amino acids, metabolism of energy and secondary compounds, transcription, and translation, and transport.
Author Contributions
Conceptualization, H.F., and U.G.; methodology, H.F., J.G., V.G., J.M.A., and S.O.; formal analysis, J.G., and H.F.; writing—original draft preparation, H.F., and S.O.; writing—review and editing, U.G., J.G., V.G., L.G.Q. and J.M.A.; funding acquisition, H.F., and U.G.; resources, L.G.Q. and U.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the University of Zurich, University of Oviedo: Grant CESSTT1819 for International Mobility of Research Staff, and European Union’s 7th Framework Program: PRIME-XS-0002520.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The concatenated dDB is available online at http://fgcz-r-021.uzh.ch/fasta/p1222_combo_NGS_n_Viridi_20160205.fasta (accessed on 9 November 2022).
Acknowledgments
We thank the University of Oviedo for a grant from International Mobility of Research Staff, according to the collaboration agreement CESSTT1819, and the Functional Genomics Center Zurich for access to its excellent infrastructure. We also thank Hanspeter Schöb for his logistics upport during visits to the Grossniklaus laboratory
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aya, K.; Kobayashi, M.; Tanaka, J.; Ohyanagi, H.; Suzuki, T.; Yano, K.; Takano, T.; Yano, K.; Matsuoka, M. De novo transcriptome assembly of a fern, Lygodium japonicum, and a web resource database Ljtrans DB. Plant Cell Physiol. 2015, 56, e5. [CrossRef]
- Berrier, C.; Peyronnet, R.; Betton, J.M.; Ephritikhine, G.; Barbier-Brygoo, H.; Frachisse, J.M.; Ghazi, A. Channel characteristics of VDAC-3 from Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2015, 459, 24–28. [CrossRef]
- Bhattacharjee, S.; Lee, L.Y.; Oltmanns, H.; Cao, H.; Veena; Cuperus, J.; Gelvin, S.B. IMPa-4, an Arabidopsis importin α isoform, is preferentially involved in agrobacterium-mediated plant transformation. Plant Cell. 2008, 20, 2661–2680. [CrossRef]
- Boldt, R.; Edner, C.; Kolukisaoglu, Ü.; Hagemann, M.; Weckwerth, W.; Wienkoop, S.; Morgenthal, K.; Bauwe, H. D-glycerate 3-kinase, the last unknown enzyme in the photorespiratory cycle in Arabidopsis, belongs to a novel kinase family. Plant Cell. 2005, 17, 2413–2420. [CrossRef]
- Bracher, A.; Sharma, A.; Starling-Windhof, A.; Hartl, F.U.; Hayer-Hartl, M. Degradation of potent rubisco inhibitor by selective sugar phosphatase. Nat. Plants. 2014, 112015, 1–7. [CrossRef]
- Carroll, A. J.; Heazlewood, J. L.; Ito, J.; Millar, A. H. Analysis of the Arabidopsis cytosolic ribosome proteome provides detailed insights into its components and their post-translational modification. Mol.Cell Proteomics. 2008, 7, 347–369. [CrossRef]
- Chen, C.-Y.; Chiu, F.-Y.; Lin, Y.; Huang, W.-J.; Hsieh, P.-S.; Hsu, F.-L. Chemical constituents analysis and antidiabetic activity validation of four fern species from Taiwan. Int. J. Mol. Sci. 2015, 16, 2497–2516. [CrossRef]
- Cochrane, F. C.; Davin, L. B.; Lewis, N. G. The Arabidopsis phenylalanine ammonia lyasegene family: kinetic characterization of the four PAL isoforms. Phytochemistry. 2004, 65, 1557–1564. [CrossRef]
- Condori-Apfata, J. A.; Batista-Silva, W.; Medeiros, D. B.; Vargas, J. R.; Valente, L. M. L.; Pérez-Díaz, J. L.; Fernie, A. Condori-Apfata, J. A.; Batista-Silva, W.; Medeiros, D. B.; Vargas, J. R.; Valente, L. M. L.; Pérez-Díaz, J. L.; Fernie, A. R,; Araújo, W. L.; Nunes-Nesi, A. Downregulation of the E2 subunit of 2-oxoglutarate dehydrogenase modulates plant growth by impacting carbon-nitrogen metabolism in Arabidopsis thaliana. Plant Cell Physiol 2021, 62, 79. [Google Scholar]
- Cordle, A.; Irish, E.; Cheng, C.L. Gene expression associated with apogamy commitment in Ceratopteris richardii. Sex. Plant Reprod. 2012, 25, 293–304. [CrossRef]
- Crosara, K.T.B.; Moffa, E.B.; Xiao, Y.; Siqueira, W.L. Merging in-silico and in vitrosalivary protein complex partners using the STRING database: a tutorial. J. Proteomics. 2018, 171, 87–94. [CrossRef]
- Der, J.P.; Barker, M.S.; Wickett, N.J.; dePamphilis, C.W.; Wolf, P.G. De novo characterization of the gametophyte transcriptome in bracken fern, Pteridium aquilinum. BMC Genomics. 2011, 12, 99. [CrossRef]
- Dhir, B. Role of ferns in environmental cleanup. In: Current Advances in Fern Research; Fernández, H., Ed.; Springer International Publishing: Cham, 2018; pp. 517–531.
- Dixon, D.P.; Edwards, R. Roles for stress-inducible lambda glutathione transferases in flavonoid metabolism in plants as identified by ligand fishing. J. Biol. Chem. 2010, 285, 36322–36329. [CrossRef]
- Eeckhout, S.; Leroux, O.; Willats, W. G.; Popper, Z. A.; Viane, R. L. Comparative glycan profiling of Ceratopteris richardii 'C-fern' gametophytes and sporophytes links cell-wall composition to functional specialization. Ann Bot. 2014, 114, 1295–1307. [CrossRef]
- Ehlting, J.; Büttner, D.; Wang, Q.; Douglas, C.J.; Somssich, I.E.; Kombrink, E. Three 4-coumarate:coenzyme A ligases in Arabidopsis thaliana represent two evolutionarily divergent classes in angiosperms. Plant J. 1999, 19, 9–20. [CrossRef]
- Eudes, A.; Pollet, B.; Sibout, R.; Do, C.T.; Séguin, A.; Lapierre, C.; Jouanin, L. Evidence for a role of AtCAD 1 in lignification of elongating stems of Arabidopsis thaliana. Planta. 2006,225, 23–39. [CrossRef]
- Fatland, B.L.; Ke; Anderson, J.; Mentzen, M.D.; Cui, W.I.; Wei; Allred, L.; Christy; Johnston, C.; Nikolau, J.L.; Wurtele, B.J.; Syrkin; LWC, E.; Biology; Physiol, M. Fatland, B.L.; Ke, J.; Anderson, M.D.; Mentzen, W.I.; Wei Cui, L.; Christy Allred, C.; Johnston, J.L.; Nikolau, B.J.; Syrkin Wurtele, E.; Biology LWC, M. Molecular characterization of a heteromeric ATP-citrate lyase that generates cytosolic acetyl-coenzyme A in Arabidopsis. Plant Physiol. 2002. [Google Scholar] [CrossRef]
- Fatland, B.L.; Nikolau, B.J.; Wurtele, E. S. Reverse genetic characterization of cytosolic acetyl-CoA generation by ATP-citrate lyase in Arabidopsis. PlantCell. 2005, 17, 182–203. [CrossRef]
- Femi-Adepoju, A.G.; Dada, A.O.; Otun, K.O.; Adepoju, A.O.; Fatoba, O. P. Green synthesis of silver nanoparticles using terrestrial fern (Gleichenia pectinata (Willd.) C. Presl.): characterization and antimicrobial studies. Heliyon. 2019, 5, e01543. [CrossRef]
- Fernández, H.; Grossmann, J.; Gagliardini, V.; Feito, I.; Rivera, A.; Rodríguez, L.; Quintanilla, L.G.; Quesada, V.; Cañal, M.J.; Grossniklaus, U. Sexual and apogamous species of woodferns show different protein and phytohormone profiles. Front. PlantSci. 2021, 12. [CrossRef]
- Fernández, H.; Revilla, M.A. Fernández, H.; Revilla, M.A. In vitroculture of ornamental ferns. Plant Cell. Tissue Organ Cult. 2003, 73, 1–13.
- Ferrario-Méry, S.; Meyer, C.; Hodges, M. Chloroplast nitrite uptake is enhanced in Arabidopsis PII mutants. FEBS Lett. 2008, 582, 1061–1066. [CrossRef]
- Gargano, D.; Maple-Grødem, J.; Møller, S.G. In vivophosphorylation of FtsZ2 in Arabidopsis thaliana. Biochem. J. 2012, 446, 517–521. [CrossRef]
- Gaudet, P.; Livstone, M.S.; Lewis, S.E.; Thomas, P.D. Phylogenetic-based propagation of functional annotations within the gene ontology consortium. Brief. Bioinform. 2011, 12, 449–462. [CrossRef]
- Grossmann, J.; Fernández, H.; Chaubey, P. M.; Valdés, A. E.; Gagliardini, V.; Cañal, M. J.; Russo, G.; Grossniklaus, U. Proteogenomic analysis greatly expands the identification of proteins related to reproduction in the apogamous fern Dryopterisaffinis ssp. affinis. Front. Plant Sci. 2017, 8, 336. [CrossRef]
- Guo, Y.; Xiong, L.; Ishitani, M.; Zhu, J.K. An Arabidopsis mutation in translation elongation factor 2 causes superinduction of CBF/DREB1 transcription factor genes but blocks the induction of their downstream targets under low temperatures. Proc. Natl. Acad. Sci. USA. 2002, 99, 7786–7791. [CrossRef]
- Haferkamp, I.; Hackstein, J.H.P.; Voncken, F.G.J.; Schmit, G.; Tjaden, J. Functional integration of mitochondrial and hydrogenosomal ADP/ATP carriers in the Escherichia coli membrane reveals different biochemical characteristics for plants, mammals and anaerobic chytrids. Eur. J. Biochem. 2002, 269, 3172–3181. [CrossRef]
- Haizel, T.; Merkle, T.; Pay, A.; Fejes, E.; Nagy, F. Characterization of proteins that interact with the GTP-bound form of the regulatory GTPase Ran in Arabidopsis. Plant J. 1997, 11, 93–103. [CrossRef]
- He, Y.; Chen, L.; Zhou, Y.; Mawhinney, T. P.; Chen, B.; Kang, B. H.; Hauser, B. A.; Chen, S. Functionalcharacterization of Arabidopsis thaliana isopropylmalate dehydrogenases reveals their important roles in gametophyte development. New Phytol. 2011, 189, 160–175. [CrossRef]
- Howles, P.A.; Birch, R.J.; Collings, D.A.; Gebbie, L.K.; Hurley, U.A.; Hocart, C.H.; Arioli, T.; Williamson, R. E. A mutation in an Arabidopsis ribose 5-phosphate isomerase reduces cellulose synthesis and is rescued by exogenous uridine. Plant J. 2006, 48, 606–618. [CrossRef]
- Ishizaki, T.; Ohsumi, C.; Totsuka, K.; Igarashi, D. Analysis of glutamate homeostasis by overexpression of Fd-GOGAT gene in Arabidopsis thaliana. Amin. Acids. 2009,38, 943–950. [CrossRef]
- Ito, J.; Batth, T. S.; Petzold, C. J.; Redding-Johanson, A. M.; Mukhopadhyay, A.; Verboom, R.; Meyer, E. H.; Millar, A. H.; Heazlewood, J. L. Analysis of the Arabidopsis cytosolic proteome highlights subcellular partitioning of central plant metabolism. J. Proteome Res. 2011, 10, 1571–1582. [CrossRef]
- Kieselbach, T.; Bystedt, M.; Hynds, P.; Robinson, C.; Schröder, W.P. A peroxidase homologue and novel plastocyanin located by proteomics to the Arabidopsis chloroplast thylakoid lumen. FEBS Lett. 2000, 480, 271–276. [CrossRef]
- Klee, H.J.; Muskopf, Y.M.; Gasser, C.S. Cloning of an Arabidopsis thalianagene encoding 5-enolpyruvylshikimate-3-phosphate synthase: sequence analysis and manipulation to obtain glyphosate-tolerant plants. Mol. Gen. Genet. 1987,210, 437–442. [CrossRef]
- Knill, T.; Reichelt, M.; Paetz, C.; Gershenzon, J.; Binder, S. Arabidopsis thaliana encodes a bacterial-type heterodimeric isopropylmalate isomerase involved in both Leu biosynthesis and the Met chain elongation pathway of glucosinolate formation. Plant Mol. Biol. 2009, 71, 227–239. [CrossRef]
- Kraker, J. W. de; Luck, K.; Textor, S.; Tokuhisa, J. G.; Gershenzon, J. Two Arabidopsis genes (IPMS1 andIPMS2) encode isopropylmalate synthase, the branchpoint step in the biosynthesis of leucine. Plant Physiol. 2007, 143, 970–986. [CrossRef]
- Küchler, M.; Decker, S.; Hörmann, F.; Soll, J.; Heins, L. Protein import into chloroplasts involves redox-regulated proteins. EMBOJ. 2002, 21, 6136–6145. [CrossRef]
- Lambermon, M.H.L.; Fu, Y.; Kirk, D.A.W.; Dupasquier, M.; Filipowicz, W.; Lorković, Z.J. UBA1 and UBA2, two proteins that interact with UBP1, a multifunctional effector of pre-mRNA maturation in plants. Mol. Cell. Biol. 2002, 22, 4346–4357. [CrossRef]
- Lee, S.H.; Chung, G.C.; Jang, J.Y.; Ahn, S.J.; Zwiazek, J.J. Overexpression of PIP2;5 aquaporin alleviates effects of low root temperature on cell hydraulic conductivity and growth in Arabidopsis. Plant Physiol. 2012, 159, 479–488. [CrossRef]
- Li, R.; Qiu, Z.; Wang, X.; Gong, P.; Xu, Q.; Yu, Q. B.; Guan, Y. Pooled CRISPR/Cas9 reveals redundant roles of plastidial phosphoglycerate kinases in carbon fixation and metabolism. Plant J. 2019,98, 1078-1089. [CrossRef]
- Lindén, P.; Keech, O.; Stenlund, H.; Gardeström, P.; Moritz, T. Reduced mitochondrial malate dehydrogenase activity has a strong effect on photorespiratory metabolism as revealed by 13C labelling. J. Exp. Bot. 2016, 67, 3123–3135. [CrossRef]
- Lintala, M.; Allahverdiyeva, Y.; Kidron, H.; Piippo, M.; Battchikova, N.; Suorsa, M.; Rintamäki, E.; Salminen, T.A.; Aro, E. M.; Mulo, P. Structural and functional characterization of ferredoxin-NADP+-oxidoreductase using knock-out mutants of Arabidopsis. Plant J. 2007, 49, 1041–1052. [CrossRef]
- Liu, J.; Last, R.L. A chloroplast thylakoid lumen protein is required for proper photosynthetic acclimation of plants under fluctuating light environments. Proc. Natl. Acad. Sci. 2017, 114. [CrossRef]
- Lundin, B.; Hansson, M.; Schoefs, B.; Vener, A. V.; Spetea, C. The Arabidopsis PsbO2 protein regulates dephosphorylation and turnover of the photosystem II reaction centre D1 protein. Plant J. 2007, 49, 528–539. [CrossRef]
- Marasinghe, G.P.K.; Sander, I.M.; Bennett, B.; Periyannan, G.; Yang, K. W.; Makaroff, C.A.; Crowder, M. W. Structural studies on a mitochondrial glyoxalase II. J. Biol. Chem. 2005, 280, 40668–40675. [CrossRef]
- Mou, Z.; He, Y.; Dai, Y.; Liu, X.; Li, J. Deficiency in fatty acid synthase leads to premature cell death and dramatic alterations in plant morphology. Plant Cell. 2000, 12, 405–417. [CrossRef]
- Murashige, T.; Skoog, F. Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Plant Physiol. 1962, 473–497.
- Ojosnegros, S.; Alvarez, J. M.; Grossmann, J.; Gagliardini, V.; Quintanilla, L. G.; Grossniklaus, U.; Fernández, H. The shared proteome of the apomictic fern Dryopteris affinisssp. affinisand its sexual relativeDryopteris oreades. Int. J. Mol. Sci. 2022, 23, 14027. [CrossRef]
- Ouyang, M.; Li, X.; Zhao, S.; Pu, H.; Shen, J.; Adam, Z.; Clausen, T.; Zhang, L. The crystal structure of Deg9 reveals a novel octameric-type HtrA protease. Nat. Plants. 2017, 3, 973–982. [CrossRef]
- Petersen, L.N.; Marineo, S.; Mandalà, S.; Davids, F.; Sewell, B.T.; Ingle, R.A. The missing link in plant histidine biosynthesis: Arabidopsis myoinositol monophosphatase-like2 encodes a functional histidinol-phosphate phosphatase. Plant Physiol. 2010, 152, 1186–1196. [CrossRef]
- Pracharoenwattana, I.; Cornah, J.E.; Smith, S.M. Arabidopsis peroxisomal citrate synthase is required for fatty acid respiration and seed germination. Plant Cell. 2005, 17, 2037–2048. [CrossRef]
- Rathinasabapathi, B. Ferns represent an untapped biodiversity for improving crops for environmental stress tolerance. New Phytol. 2006, 172, 385–390. [CrossRef]
- Rautengarten, C.; Ebert, B.; Herter, T.; Petzold, C.J.; Ishii, T.; Mukhopadhyay, A.; Usadel, B.; Scheller, H.V. The interconversion of UDP-arabinopyranose and UDP-arabinofuranose is indispensable for plant development in Arabidopsis. Plant Cell. 2011, 23, 1373–1390. [CrossRef]
- Rivera, A.; Cañal, M. J.; Grossniklaus, U.; Fernández, H. The gametophyte of fern: born to reproduce. In: Current Advances in Fern Research. Fernández, H., Ed.; Springer International Publishing: New York, 2018, pp. 3–19.
- Rosenquist, M.; Alsterfjord, M.; Larsson, C.; Sommarin, M. Data mining the Arabidopsis genome reveals fifteen 14-3-3 genes. expression is demonstrated for two out of five novel genes. Plant Physiol. 2001, 127, 142–149. [CrossRef]
- Salmi, M. L.; Bushart, T. J. Cellular, molecular, and genetic changes during the development of Ceratopteris richardii gametophytes. In: Working with ferns. Issues and applications; Fernández, H., Kumar, A., Eds.; Springer International Publishing: New York, 2010; pp. 11–24.
- Salmi, M.L.; Bushart, T.; Stout, S.; Roux, S. Profile and analysis of gene expression changes during early development in germinating spores of Ceratopteris richardii. Plant Physiol. 2005, 138, 1734–1745. [CrossRef]
- Salmi, M.L.; Morris, K.E.; Roux, S.J.; Porterfield, D.M. Nitric oxide and CGMP signaling in calcium-dependent development of cell polarity in Ceratopteris richardii. Plant Physiol. 2007, 144, 94–104. [CrossRef]
- Sanda, S.; Leustek, T.; Theisen, M.J.; Garavito, R.M.; Benning, C. Recombinant Arabidopsis SQD1 converts UDP-glucose and sulfite to the sulfolipid head group precursor UDP-sulfoquinovose in vitro. J. Biol. Chem. 2001, 276, 3941–3946. [CrossRef]
- Scranton, M.A.; Yee, A.; Park, S.Y.; Walling, L. L. Plant leucine aminopeptidases moonlight as molecular chaperones to alleviate stress-induced damage. J. Biol. Chem. 2012, 287, 18408–18417. [CrossRef]
- Seung, D.; Soyk, S.; Coiro, M.; Maier, B.A.; Eicke, S.; Zeeman, S. C. PROTEIN TARGETING TO STARCH is required for localising GRANULE-BOUND STARCH SYNTHASE to starch granules and for normal amylose synthesis in Arabidopsis. PLOS Biol. 2015, 13, e1002080. [CrossRef]
- Shen, B.; Li, C.; Tarczynski, M.C. Shen, B.; Li, C.; Tarczynski, M.C. High free-methionine and decreased lignin content result from a mutation in the Arabidopsis S-adenosyl-L-methionine synthetase 3 gene. Plant J. 2002, 29, 371–380.
- Shi, X.; Hanson, M.R.; Bentolila, S. Two RNA recognition motif-containing proteins are plant mitochondrial editing factors. Nucleic Acids Res. 2015, 43, 3814–3825. [CrossRef]
- Shirley, B.W.; Hanley, S.; Goodman, H. M. Effects of ionizing radiation on a plant genome: analysis of two Arabidopsis transparent testa mutations. Plant Cell. 1992, 4, 333–347. [CrossRef]
- Skalitzky, C.A.; Martin, J.R.; Harwood, J.H.; Beirne, J.J.; Adamczyk, B.J.; Heck, G.R.; Cline, K.; Fernandez, D. E. Plastids contain a second Sec translocase system with essential functions. Plant Physiol. 2011, 155, 354–369. [CrossRef]
- Ståhl, A.; Moberg, P.; Ytterberg, J.; Panfilov, O.; Von Löwenhielm, H. B.; Nilsson, F.; Glaser, E. Isolation and identification of a novel mitochondrial metalloprotease (PreP) that degrades targeting presequences in plants. J. Biol. Chem. 2002, 277, 41931–41939. [CrossRef]
- Stanislas, T.; Hüser, A.; Barbosa, I.C.R.; Kiefer, C.S.; Brackmann, K.; Pietra, S.; Gustavsson, A.; Zourelidou, M.; Schwechheimer, C.; Grebe, M. Arabidopsis D6PK is a lipid domain-dependent mediator of root epidermal planar polarity. Nat. Plants. 2015,1, 1–9. [CrossRef]
- Suo, J.; Zhao, Q.; Zhang, Z.; Chen, S.; Cao, J.; Liu, G.; Wei, X.; Wang, T.; Yang, C.; Dai, S. Cytological and proteomic analyses of Osmunda cinnamomea germinating spores reveal characteristics of fern spore germination and rhizoid tip growth. Mol. Cell. Proteomics. 2015, 14, 2510–2534. [CrossRef]
- Takagi, D.; Amako, K.; Hashiguchi, M.; Fukaki, H.; Ishizaki, K.; Goh, T.; Fukao, Y.; Sano, R.; Kurata, T.; Demura, T.; Sawa, S.; Miyake, C. Chloroplastic ATP synthase builds up a proton motive force preventing production of reactive oxygen species in photosystem I. Plant J. 2017, 91, 306–324. [CrossRef]
- Takahashi, M.; Sasaki, Y.; Ida, S.; Morikawa, H. Nitrite reductase gene enrichment improves assimilation of NO(2) in Arabidopsis. Plant Physiol. 2001, 126, 731–741. [CrossRef]
- Tomaz, T.; Bagard, M.; Pracharoenwattana, I.; Lindén, P.; Lee, C.P.; Carroll, A.J.; Ströher, E.; Smith, S.M.; Gardeström, P.; Millar, A.H. Mitochondrial malate dehydrogenase lowers leaf respiration and alters photorespiration and plant growth in Arabidopsis. Plant Physiol. 2010, 154, 1143–1157. [CrossRef]
- Tronconi, M.A.; Fahnenstich, H.; Gerrard Weehler, M.C.; Andreo, C. S.; Flügge, U.I.; Drincovich, M.F.; Maurino, V.G. Arabidopsis NAD-malic enzyme functions as a homodimer and heterodimer and has a major impact on nocturnal metabolism. Plant Physiol. 2008, 146, 1540. [CrossRef]
- Valledor, L.; Menéndez, V.; Canal, M.J.; Revilla, A.; Fernández, H. Proteomic approaches to sexual development mediated by antheridiogen in the fern Blechnum spicant L. Proteomics 2014, 14. [CrossRef]
- Varotto, C.; Pesaresi, P.; Meurer, J.; Oelmüller, R.; Steiner-Lange, S.; Salamini, F.; Leister, D. Disruptionof the Arabidopsis photosystem I gene psaE1 affects photosynthesis and impairs growth. Plant J. 2000, 22, 115–124. [CrossRef]
- Wada, M. The fern as a model system to study photomorphogenesis. J. Plant Res. 2007, 120, 3–16. [CrossRef]
- Wakao, S.; Benning, C. Genome-wide analysis of glucose-6-phosphate dehydrogenases in Arabidopsis. Plant J. 2005, 41, 243–256. [CrossRef]
- Wang, R.; Yang, L.; Han, X.; Zhao, Y.; Zhao, L.; Xiang, B.; Zhu, Y.; Bai, Y.; Wang, Y. Overexpression of AtAGT1 promoted root growth and development during seedling establishment. Plant Cell Rep. 2019, 38, 1165–1180. [CrossRef]
- Wang, X.; Chen, S.; Zhang, H.; Shi, L.; Cao, F.; Guo, L.; Xie, Y.; Wang, T.; Yan, X.; Dai, S. Desiccation tolerance mechanism in resurrection fern-ally Selaginella tamariscina revealed by physiological and proteomic analysis. J. Proteome Res. 2010, 9, 6561–6577. [CrossRef]
- Wyder, S.; Rivera, A.; Valdés, A.E.; Cañal, M. J.; Gagliardini, V.; Fernández, H.; Grossniklaus, U. Plant physiology and biochemistry differential gene expression profiling of one- and two-dimensional apogamous gametophytes of the fern Dryopteris affinis ssp. affinis. Plant Physiol. Biochem. 2020, 148, 302–311. [CrossRef]
- Yan, Y.; Liu, Q.; Zang, X.; Yuan, S.; Bat-Erdene, U.; Nguyen, C.; Gan, J.; Zhou, J.; Jacobsen, S. E.; Tang, Y.Resistance-gene-directed discovery of a natural-product herbicide with a new mode of action. Nature. 2018, 559, 415–418. [CrossRef]
- Ye, H.; Abdel-Ghany, S. E.; Anderson, T. D.; Pilon-Smits, E. A.; Pilon, M. CpSufE activates the cysteine desulfurase CpNifS for chloroplastic Fe-S cluster formation. J Biol Chem. 2006, 281, 8958–8969. [CrossRef]
- Žárský, V.; Cvrčková, F.; Bischoff, F.; Palme, K. At-GDI1 from Arabidopsis thalianaencodes a Rab-specific GDP dissociation inhibitor that complements the Sec19 mutation of Saccharomyces cerevisiae. FEBS Lett. 1997, 403, 303–308. [CrossRef]
- Zhang, Y.; Li, B.; Xu, Y.; Li, H.; Li, S.; Zhang, D.; Mao, Z.; Guo, S.; Yang, C.; Weng, Y.; Chong, K. The cyclophilin CYP20-2 modulates the conformation of BRASSINAZOLE-RESISTANT1, which binds the promoter of FLOWERING LOCUS D to regulate flowering in Arabidopsis. Plant Cell. 2013, 25, 2504–2521. [CrossRef]
- Zhang, Y.; Sun, K.; Sandoval, F. J.; Santiago, K.; Roje, S. One-carbon metabolism in plants: characterization of a plastid serine hydroxymethyltransferase. Biochem. J. 2010, 430, 97–105. [CrossRef]
- Zhang, Y.; Swart, C.; Alseekh, S.; Scossa, F.; Jiang, L.; Obata, T.; Graf, A.; Fernie, A. R. The extra-pathway interactome of the TCA cycle: expected and unexpected metabolic interactions. Plant Physiol. 2018, 177, 966–979. [CrossRef]
- Zhao, Z.; Assmann, S.M. The glycolytic enzyme, phosphoglycerate mutase, has critical roles in stomatal movement, vegetative growth, and pollen production in Arabidopsis thaliana. J. Exp. Bot. 2011, 62, 5179. [CrossRef]
Figure 1.
GO enrichment terms of the shared proteomes obtained from gametophytes of Dryopteris affinis and D. oreades according to the category biological function, analyzed with the STRING and CYTOSCAPE programs. Yellow color refers to the metabolism of energy, pink to transcription and translation, green to protein degradation, and blue to the metabolism of carbohydrates.
Figure 1.
GO enrichment terms of the shared proteomes obtained from gametophytes of Dryopteris affinis and D. oreades according to the category biological function, analyzed with the STRING and CYTOSCAPE programs. Yellow color refers to the metabolism of energy, pink to transcription and translation, green to protein degradation, and blue to the metabolism of carbohydrates.
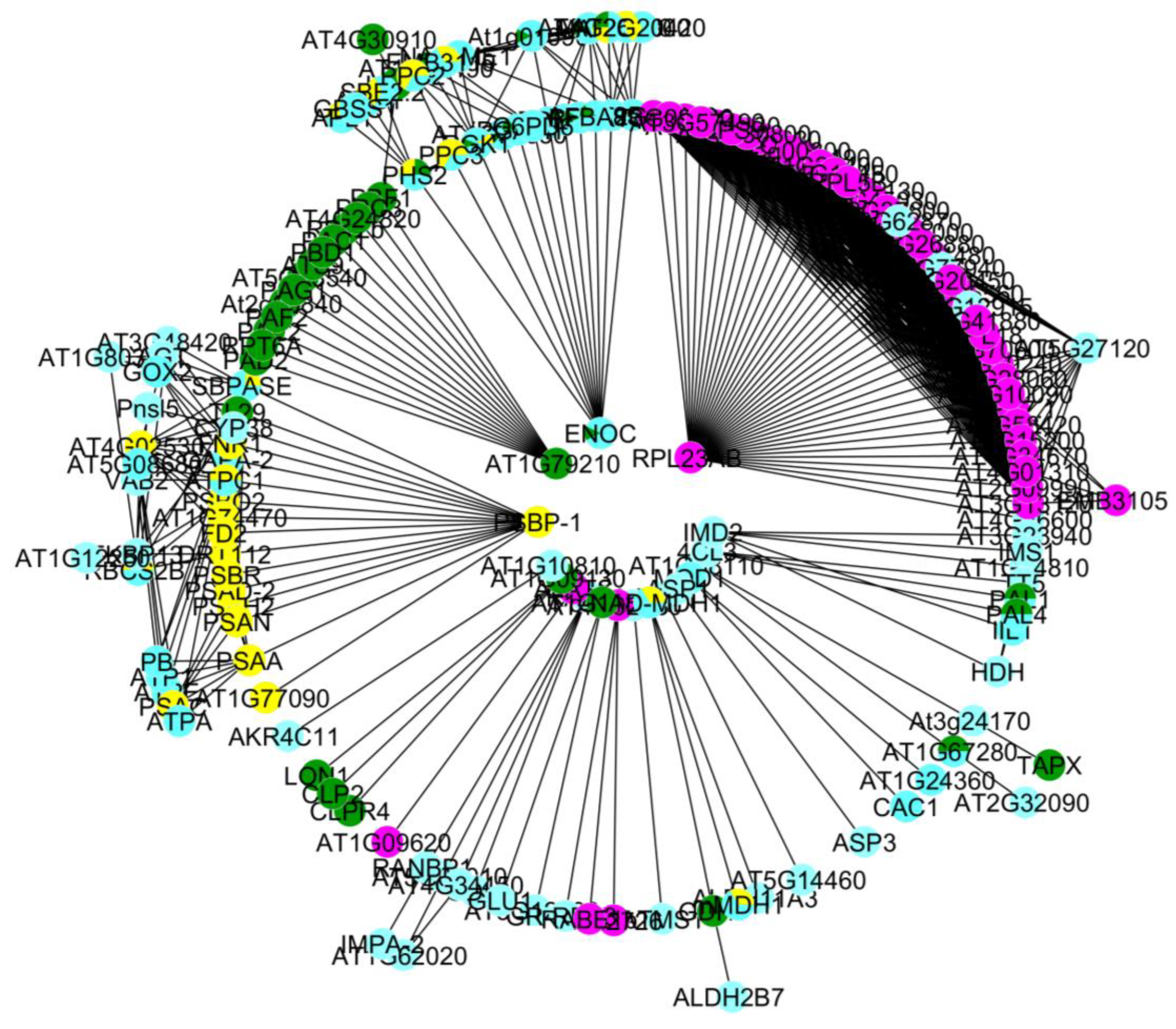
Figure 2.
KEGG enrichment terms of the shared proteomes obtained from the gametophytes of Dryopteris affinisand D. oreades, analyzed with the STRING platform.
Figure 2.
KEGG enrichment terms of the shared proteomes obtained from the gametophytes of Dryopteris affinisand D. oreades, analyzed with the STRING platform.
Figure 3.
Proteins involved in ribogenesis found in the gametophyte of the ferns Dryopteris affinis and D. oreades. Imaged provided by STRING platform according to KEGG dataset. “Light-green” highlighted boxes are the identified proteins.
Figure 3.
Proteins involved in ribogenesis found in the gametophyte of the ferns Dryopteris affinis and D. oreades. Imaged provided by STRING platform according to KEGG dataset. “Light-green” highlighted boxes are the identified proteins.

Figure 4.
Proteins involved in proteasome found in the gametophyte of the ferns Dryopteris affinis and D. oreades. Imaged provided by STRING platform according to KEGG dataset. “Light-green” highlighted boxes are the identified proteins.
Figure 4.
Proteins involved in proteasome found in the gametophyte of the ferns Dryopteris affinis and D. oreades. Imaged provided by STRING platform according to KEGG dataset. “Light-green” highlighted boxes are the identified proteins.
Figure 5.
Circular representations obtained through STRING and CYTOSCAPE programs of proteins shared by the gametophytes Dryopteris affinis and D. oreades and classified in the following groups: (a) metabolism of carbohydrates, (b) metabolism of energy, (c) ribogenesis, and (d) protein degradation. The pinklines refer to experiments interactions, the green lines to textmining interactions, the black lines to co-expression interactions, and the blue lines to databases interactions.
Figure 5.
Circular representations obtained through STRING and CYTOSCAPE programs of proteins shared by the gametophytes Dryopteris affinis and D. oreades and classified in the following groups: (a) metabolism of carbohydrates, (b) metabolism of energy, (c) ribogenesis, and (d) protein degradation. The pinklines refer to experiments interactions, the green lines to textmining interactions, the black lines to co-expression interactions, and the blue lines to databases interactions.
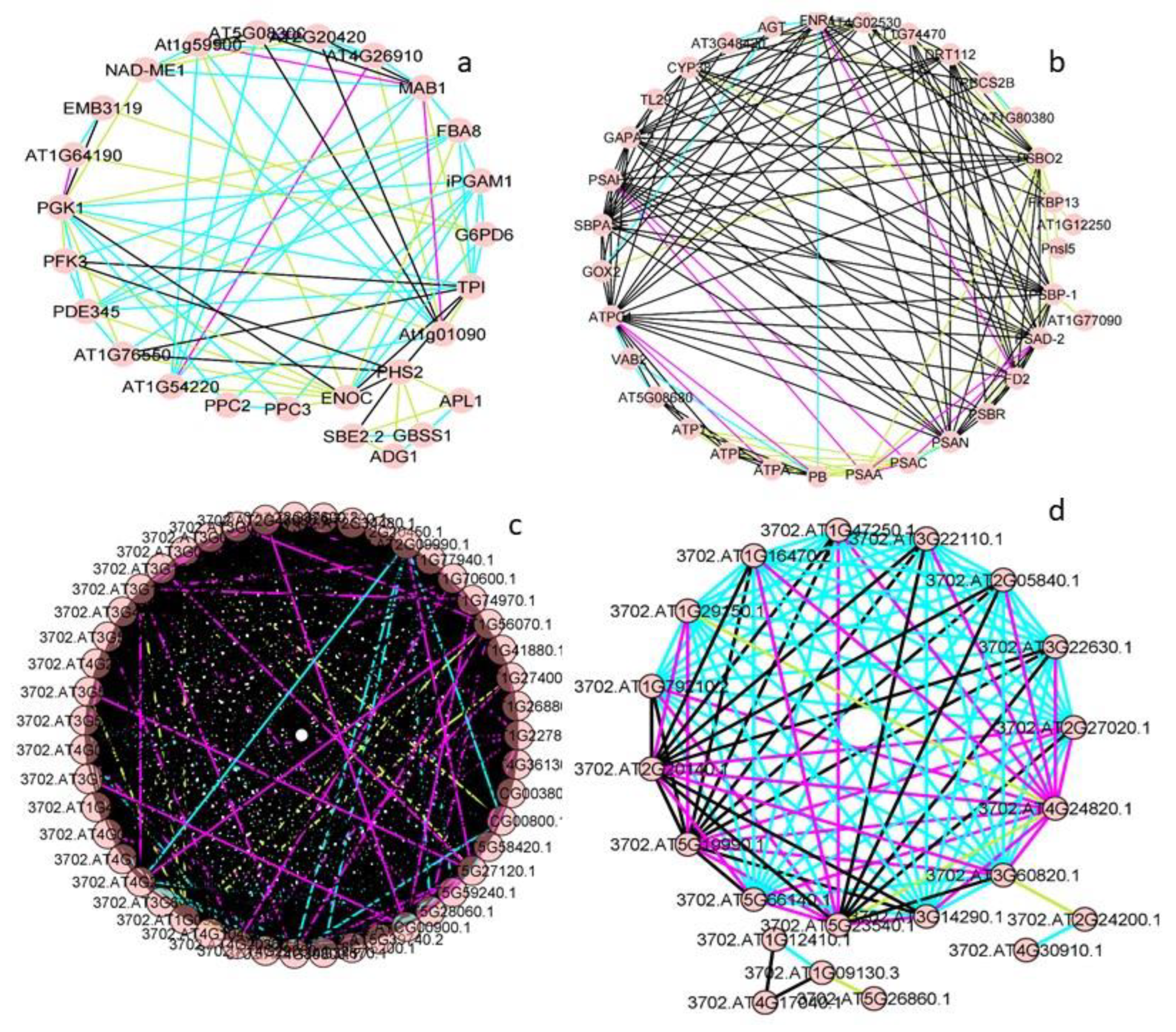
Figure 6.
Figure 6. Plots of the two main types of interactions in the groups of proteins shared by the gametophytes of Dryopteris affinis and D. oreades: (a) metabolism of carbohydrates, (b) biosynthesis of amino acids, (c) metabolism of energy, and (d) transcription and translation. Each spot represents the intersection ofthe interactions between two proteins.
Figure 6.
Figure 6. Plots of the two main types of interactions in the groups of proteins shared by the gametophytes of Dryopteris affinis and D. oreades: (a) metabolism of carbohydrates, (b) biosynthesis of amino acids, (c) metabolism of energy, and (d) transcription and translation. Each spot represents the intersection ofthe interactions between two proteins.
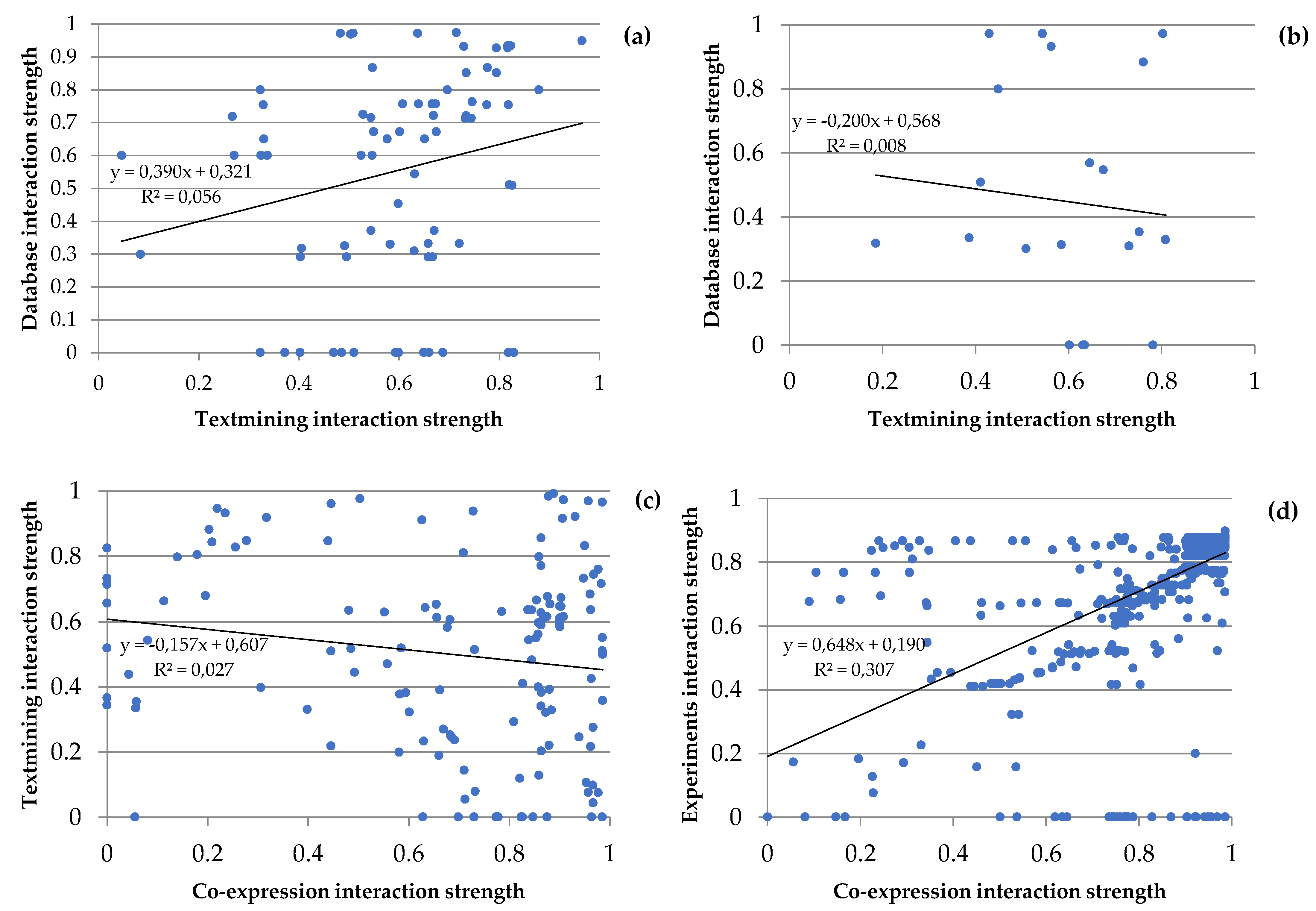
Figure 7.
Figure 7. Morphological features in the apogamous fern Dryopteris affinis and its sexual relative D. oreades. From left to right, and from top to below (a) typical kidney sori on the leaf underside; (b) confocal image of spores; (c) gametophytes growing up in a Petri dish; (d) and (e) images under light microscope of one- and two-dimensional gametophytes of D. affinis; and (f) female sexual organs or archegonia in the gametophyte of D. oreades.
Figure 7.
Figure 7. Morphological features in the apogamous fern Dryopteris affinis and its sexual relative D. oreades. From left to right, and from top to below (a) typical kidney sori on the leaf underside; (b) confocal image of spores; (c) gametophytes growing up in a Petri dish; (d) and (e) images under light microscope of one- and two-dimensional gametophytes of D. affinis; and (f) female sexual organs or archegonia in the gametophyte of D. oreades.
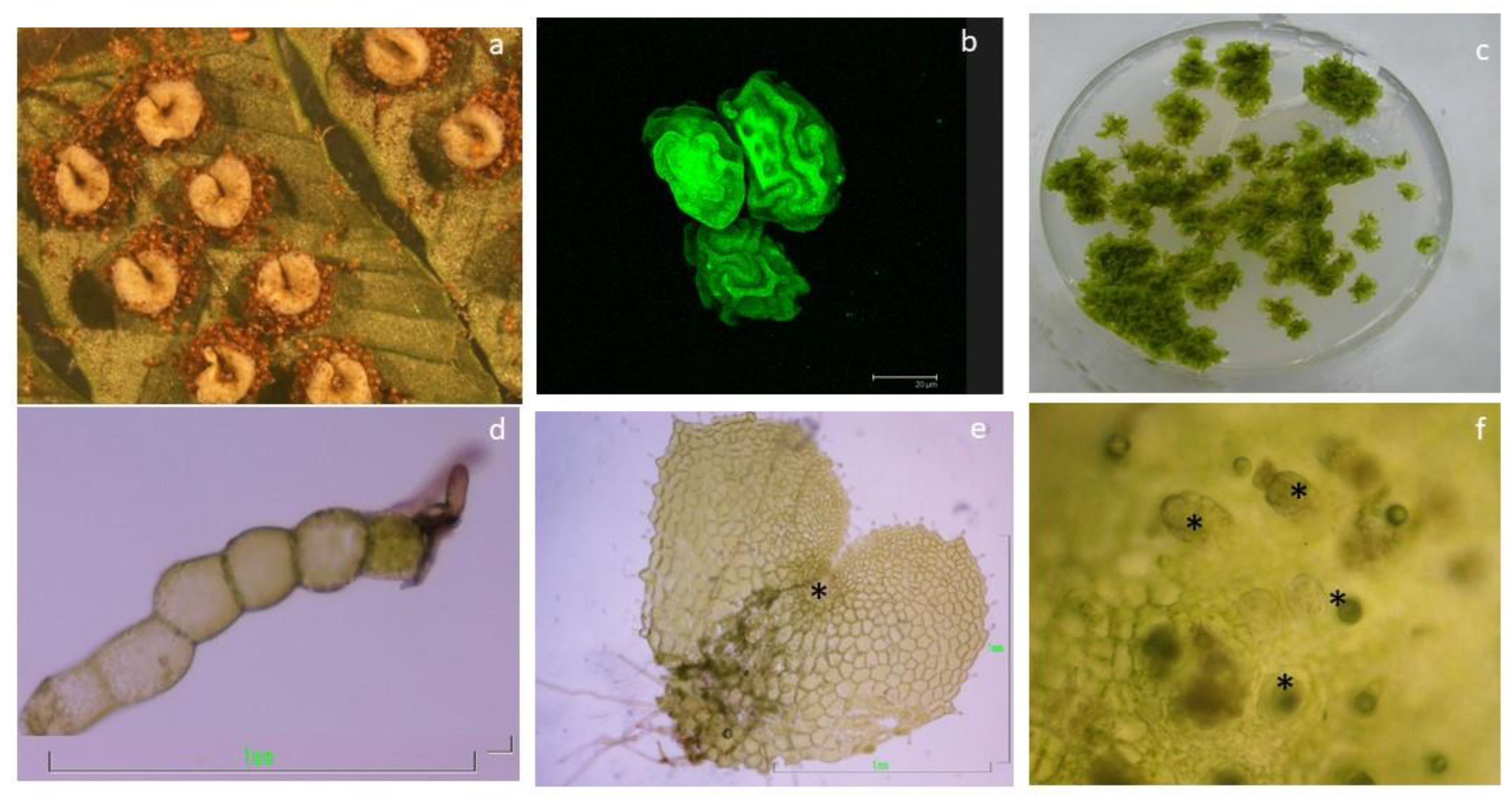

Table 1.
Selected proteins equally regulated in gametophytes of Dryopteris affinisand D. oreades.
| Category | Accession Number | UniProtKB/ Swiss-Prot |
Gene Name | Protein Name | MW (kDa) | Amino Acids |
|---|---|---|---|---|---|---|
| Carbohydrates | 58787-330_2_ORF2 | Q94AA4 | PFK3 | Phosphofructokinase 3 | 53 | 489 |
| Carbohydrates | 135690-210_1_ORF2 | Q9ZU52 | PDE345 | Pigment defective 345 | 42 | 391 |
| Carbohydrates | tr|A9NMQ0|A9NMQ0_PICSI | Q9LF98 | FBA8 | Fructose-bisphosphate aldolase 8 | 38 | 358 |
| Carbohydrates | 38153-411_5_ORF2 | Q38799 | MAB1 | Macci-bou | 39 | 363 |
| Carbohydrates | 83096-276_3_ORF2 | Q5GM68 | PPC2 | Phosphoenolpyruvate carboxylase 2 | 109 | 963 |
| Carbohydrates | 54280-344_1_ORF1 | Q84VW9 | PPC3 | Phosphoenolpyruvate carboxylase 3 | 110 | 968 |
| Carbohydrates | 113756-233_2_ORF1 | Q9SIU0 | NAD-ME1 | NAD-dependent malic enzyme 1 | 69 | 623 |
| Carbohydrates | 102811-246_6_ORF2 | O04499 | iPGAM1 | 2,3-biphosphoglycerate-independent phosphoglycerate mutase 1 | 60 | 557 |
| Carbohydrates | 70011-302_2_ORF1 | O82662 | AT2G20420 | - | 45 | 421 |
| Carbohydrates | 8279-816_3_ORF2 | P68209 | AT5G08300 | - | 36 | 347 |
| Carbohydrates | 222487-119_2_ORF2 | P93819 | c-NAD-MDH1 | Cytosolic-NAD-dependent malate dehydrogenase 1 | 35 | 332 |
| Carbohydrates | 156827-185_4_ORF1 | Q9SH69 | PGD1 | 6-phosphogluconate dehydrogenase 1 | 53 | 487 |
| Carbohydrates | 12493-682_6_ORF2 | Q9FJI5 | G6PD6 | Glucose-6-phosphate dehydrogenase 6 | 59 | 515 |
| Carbohydrates | 20760-547_4_ORF1 | Q9LD57 | PGK1 | Phosphoglycerate kinase 1 | 50 | 481 |
| Carbohydrates | 69882-302_6_ORF2 | Q9LZS3 | SBE2.2 | Starch branching enzyme 2.2 | 92 | 805 |
| Carbohydrates | tr|Q5PYJ7|Q5PYJ7_9MONI | Q9MAQ0 | GBSS1 | Granule bound starch synthase 1 | 66 | 610 |
| Carbohydrates | tr|A9SGH8|A9SGH8_PHYPA | P55228 | ADG1 | ADP glucose pyrophosphorylase 1 | 56 | 520 |
| Carbohydrates | 181563-155_3_ORF2 | P55229 | APL1 | ADP glucose pyrophosphorylase large subunit 1 | 57 | 522 |
| Carbohydrates | tr|D7MQA6|D7MQA6_ARALL | Q9LUE6 | RGP4 | Reversibly glycosylated polypeptide 4 | 41 | 364 |
| Carbohydrates | 162660-176_6_ORF1 | P83291 | AT5G20080 | - | 35 | 328 |
| Lipids | 20213-554_2_ORF1 | Q9SLA8 | MOD1 | Mosaic death 1 | 41 | 390 |
| Lipids | 387953-27_4_ORF1 | Q9SGY2 | ACLA-1 | ATP-citrate lyase A-1 | 49 | 443 |
| Category | Accession Number |
UniProtKB/ Swiss-Prot |
Gene Name | Protein Name | MW (kDa) |
Amino Acids |
| Lipids | 211149-128_1_ORF1 | Q9LXS6 | CSY2 | Citrate synthase 2 | 56 | 514 |
| Amino acids | 47558-369_4_ORF2 | P46643 | ASP1 | Aspartate aminotransferase 1 | 47 | 430 |
| Amino acids | 72506-296_4_ORF1 | Q94AR8 | IIL1 | Isopropyl malate isomerase large subunit 1 | 55 | 509 |
| Amino acids | 125905-219_3_ORF2 | Q9ZNZ7 | GLU1 | Glutamate synthase 1 | 179 | 1,648 |
| Amino acids | 393073-25_4_ORF2 | Q94JQ3 | SHM3 | Serine hydroxymethyltransferase 3 | 57 | 529 |
| Amino acids | tr|D8RLH8|D8RLH8_SELML | Q9C5U8 | HDH | Histidinol dehydrogenase | 50 | 466 |
| Amino acids | 294436-71_4_ORF2 | Q9LUT2 | MTO3 | Methionine over-accumulator 3 | 42 | 393 |
| Nucleotides | 2121-1366_3_ORF2 | Q9SF85 | ADK1 | Adenosine kinase 1 | 37 | 344 |
| Nucleotides | 59309-329_5_ORF1 | Q96529 | ADSS | Adenylosuccinate synthase | 52 | 490 |
| Nucleotides | 152024-193_3_ORF2 | Q9S726 | EMB3119 | Embryo defective 3119 | 29 | 276 |
| Energy | 164104-175_1_ORF1 | Q9FKW6 | FNR1 | Ferredoxin-NADP(+)-oxidoreductase 1 | 40 | 360 |
| Energy | sp|Q7SIB8|PLAS_DRYCA | P42699 | DRT112 | DNA-damage-repair/toleration protein 112 | 16 | 167 |
| Energy | 154679-189_1_ORF2 | Q9S841 | PSBO2 | Photosystem II subunit O-2 | 35 | 331 |
| Energy | 218625-122_1_ORF2 | O22773 | MPH2 | Maintenance of photosystem II under high light 2 | 23 | 216 |
| Energy | 6036-926_2_ORF1 | Q9ASS6 | Pnsl5 | Photosynthetic NDH subcomplex l 5 | 28 | 259 |
| Energy | 250817-99_2_ORF2 | Q94K71 | AT3G48420 | - | 34 | 319 |
| Energy | tr|A9RDI1|A9RDI1_PHYPA | Q944I4 | GLYK | Glycerate kinase | 51 | 456 |
| Energy | 297118-70_2_ORF2 | Q56YA5 | AGT | Alanine:glyoxylate aminotransferase | 44 | 401 |
| Energy | 33137-439_6_ORF2 | O48917 | SQD1 | Sulfoquinovosyldiacylglycerol 1 | 53 | 477 |
| Energy | 227095-115_1_ORF2 | Q84W65 | CPSUFE | Chloroplast sulfur E | 40 | 371 |
| Energy | 311596-62_2_ORF2 | Q9ZST4 | GLB1 | GLNB1 homolog | 21 | 196 |
| Energy | 318906-58_1_ORF1 | Q39161 | NIR1 | Nitrite reductase 1 | 65 | 586 |
| Secondary compounds | 156331-186_3_ORF2 | P41088 | TT5 | Transparent testa 5 | 26 | 246 |
| Secondary compounds | 230420-113_2_ORF2 | P34802 | GGPS1 | Geranylgeranyl pyrophosphate synthase 1 | 40 | 371 |
| Secondary compounds | 85783-271_1_ORF2 | Q9T030 | PCBER1 | Phenylcoumaran benzylic ether reductase 1 | 34 | 308 |
| Category | Accession Number |
UniProtKB/ Swiss-Prot |
Gene Name | Protein Name | MW (kDa) |
Amino Acids |
| Secondary compounds | 153413-190_1_ORF2 | P42734 | CAD9 | Cinnamyl alcohol dehydrogenase 9 | 38 | 360 |
| Secondary compounds | 156554-185_2_ORF1 | Q9S777 | 4CL3 | 4-coumarate:coA ligase 3 | 61 | 561 |
| Secondary compounds | 223603-118_1_ORF1 | P05466 | AT2G45300 | - | 55 | 520 |
| Oxido -reduction |
133847-212_2_ORF2 | Q9SID3 | GLX2-5 | Glyoxalase 2-5 | 35 | 324 |
| Oxido -reduction |
tr|E1ZRS4|E1ZRS4_CHLVA | Q9ZP06 | mMDH1 | Mitochondrial malate dehydrogenase 1 | 35 | 341 |
| Oxido -reduction |
34437-432_2_ORF1 | Q9M2W2 | GSTL2 | Glutathione transferase lambda 2 | 33 | 292 |
| Oxido -reduction |
115571-230_4_ORF1 | Q9LZ06 | GSTL3 | Glutathione transferase L3 | 27 | 235 |
| Transcription | tr|A2X6N1|A2X6N1_ORYSI | Q96300 | GRF7 | General regulatory factor 7 | 29 | 265 |
| Transcription | 287872-75_1_ORF1 | Q9C5W6 | GRF12 | General regulatory factor 12 | 30 | 268 |
| Translation | 209284-130_2_ORF2 | Q9FNR1 | RBGA7 | MA-binding glycine-rich protein A7 | 29 | 309 |
| Translation | 293356-72_1_ORF1 | Q9LR72 | AT1G03510 | - | 47 | 429 |
| Translation | 26795-487_6_ORF2 | Q0WW84 | RBP47B | MA-binding protein 47B | 48 | 435 |
| Translation | 20230-554_5_ORF2 | Q9LES2 | UBA2A | UBP1-associated protein 2A | 51 | 478 |
| Translation | 174433-162_1_ORF1 | Q9ASR1 | LOS1 | Low expression of osmotically responsive genes 1 | 93 | 843 |
| Folding | 26640-489_1_ORF2 | Q9M1C2 | GROES | - | 15 | 138 |
| Folding | 189606-147_1_ORF2 | Q9SR70 | AT3G10060 | - | 24 | 230 |
| Folding | 149253-199_6_ORF2 | O22870 | AT2G43560 | - | 23 | 223 |
| Folding | 2524-1285_6_ORF2 | Q9SKQ0 | AT2G21130 | - | 18 | 174 |
| Transport | 19573-562_5_ORF2 | Q9SYI0 | AGY1 | Albino or glassy yellow 1 | 117 | 1,042 |
| Transport | 248569-101_3_ORF1 | P92985 | RANBP1 | RAN binding protein 1 | 24 | 219 |
| Transport | 146969-201_2_ORF1 | F4JL11 | IMPA-2 | Importin alpha isoform 2 | 58 | 535 |
| Transport | 151836-193_1_ORF2 | P40941 | AAC2 | ADP/ATP carrier 2 | 41 | 385 |
| Category | Accession Number |
UniProtKB/ Swiss-Prot |
Gene Name | Protein Name | MW (kDa) |
Amino Acids |
| Transport | 161087-178_2_ORF2 | Q8H0U5 | Tic62 | Translocon at the inner envelope membrane of chloroplasts 62 | 68 | 641 |
| Transport | 82340-277_1_ORF2 | Q39196 | PIP1;4 | Plasma membrane intrinsic protein 1;4 | 30 | 287 |
| Transport | 154825-188_3_ORF2 | Q9SMX3 | VDAC3 | Voltage dependent anion channel 3 | 29 | 274 |
| Transport | 272341-85_2_ORF2 | Q94A40 | alpha1-COP | Alpha1 coat protein | 136 | 1,216 |
| Transport | 29489-466_3_ORF1 | Q0WW26 | gamma2-COP | Gamma2 coat protein | 98 | 886 |
| Transport | 38639-409_2_ORF3 | Q93Y22 | AT5G05010 | - | 57 | 527 |
| Transport | 43675-385_1_ORF2 | Q67YI9 | EPS2 | Epsin2 | 95 | 895 |
| Transport | 68824-304_5_ORF2 | Q9LQ55 | DL3 | Dynamin-like 3 | 100 | 920 |
| Transport | - | F4J3Q8 | GET3B | Guided entry of tail-anchored proteins 3B | 47 | 433 |
| Transport | 3434-1154_1_ORF2 | Q96254 | GDI1 | Guanosine nucleotide diphosphate dissociation inhibitor 1 | 49 | 445 |
| Degradation | 141778-205_4_ORF2 | Q8L770 | AT1G09130 | - | 40 | 370 |
| Degradation | 172993-163_5_ORF1 | Q9XJ36 | CLP2 | CLP protease proteolytic subunit 2 | 31 | 279 |
| Degradation | 72587-296_2_ORF2 | Q8LB10 | CLPR4 | CLP protease R subunit 4 | 33 | 305 |
| Degradation | 17420-593_1_ORF2 | P93655 | LON1 | LON protease 1 | 109 | 985 |
| Degradation | tr|A9SF86|A9SF86_PHYPA | Q9LJL3 | PREP1 | Presequence protease 1 | 121 | 1,080 |
| Degradation | 186732-150_2_ORF2 | Q9FL12 | DEG9 | Degradation of periplasmic proteins 9 | 65 | 592 |
| Degradation | 170504-166_2_ORF2 | P30184 | LAP1 | Leucyl aminopeptidase 1 | 54 | 520 |
| Degradation | 170504-166_2_ORF2 | Q944P7 | LAP3 | Leucyl aminopeptidase 3 | 61 | 581 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



