
Submitted:
14 March 2023
Posted:
15 March 2023
You are already at the latest version
Abstract
Green microalgae including those from the genus Lobosphaera are exploited in biotechnology to obtain valuable fatty acids e.g., arachidonic acid (C20:4, ARA) for the production of infant formulae, food and feed additives. In nature, microalgae frequently exist in naturally immobilized state (as biofilms) with limited cell division rate and increased stress resilience. In biotechnology, immobilization of microalgae on artificial cell carriers simplifies biomass harvesting, increases culture robustness and productivity. The choice of suitable cell carrier is central to biotechnology of immobilized culture. Cell carriers based on the natural amine-containing polymer chitosan and synthetic polyethylenimine (PEI) are promising candidates for immobilization of phototrophic microorganisms. This is the first report on the effects of immobilization on PEI and chitosan on the accumulation and composition of polyunsaturated fatty acids, including ARA, in Lobosphaera sp. IPPAS C-2047. The immobilization on PEI increased ARA percentage in the total tatty acids and ARA accumulation by 72% and 81%, compared with the suspended cells cultured respectively in complete or nitrogen-deprived medium 14 days. Immobilization of Lobosphaera sp. on the chitosan-based carrier declined ARA but increased oleic and α-linoleic acid percentages. Mechanisms of the effects of immobilization on fatty acid profiles of the microalgae are discussed.
Keywords:
microalgae
; attached cultivation
; chitosan
; polyethylenimine
; fatty acids
; arachidonic acid
1. Introduction
Microalgae are used in various fields of biotechnology for the production of biofuels and valuable metabolites, bioremoval of heavy metals and surplus nutrients from wastewater [1,2,3]. Green microalgae from the genus Lobosphaera (Chlorophyta, Trebouxiophyceae) are promising objects of photobiotechnology, as they are capable of accumulating high quantities of long-chain ω6-polyunsaturated arachidonic acid (C20:4, ARA). ARA is utilized in the production of infant formulae, functional food and aquaculture feed additives as well as in pharmaceutical and cosmetology industries [4,5,6,7].
In nature, microalgae including representatives of the genus Lobosphaera frequently exist as a part of biofilms—communities of photo- and heterotrophic microorganisms formed at interphase surfaces [8,9,10]. In biofilms, the mobility and cell division rate of microorganisms embedded into the extracellular matrix are restricted [9]. Since the microorganisms living in biofilms are confined to a limited space, they can be thought as naturally immobilized cells. Theу feature increased resilience to adverse environmental conditions, so formation of biofilms is among the main strategies for the survival of microorganisms in nature [9,11,12].
Following the nature-inspire approach, microalgae immobilized either by adsorption on the surface of or entrapment within artificial cell carriers found broad application in biotechnology [13,14,15,16]. The main advantage of immobilized cells as compared to their suspended counterparts is simplified biomass harvesting, which is a key problem of microalgal biotechnology [17,18]. Other advantages include easy recycling of immobilized microalgal biomass, lack of mixing requirement, lower risk of contamination, and higher resistance to various stresses [16,17,18,19]. Notably, increased cell density, shorter lag phase, and intensified intercellular communication characteristic of immobilized cells can increase their productivity regarding valuable metabolites as compared with the corresponding suspended cultures [18,19].
The choice of biocompatible and inexpensive cell carrier characterized by high cell load capacity, reliable attachment of phototrophic cells, and free flow of nutrients as well as by transparency for adequate light energy supply to the cells is central to the biotechnology of immobilized cultures [18]. Cell carriers based on natural amine-containing polymer chitosan (CH) and synthetic polymer polyethylenimine (PEI) obtained by cryopolymerization are successfully used for the immobilization of phototrophic microorganisms [16,20]. The polycationic polymers have a high affinity for microalgal cells. It is determined by the presence of negatively charged (usually carboxyl and thiol) groups of polysaccharides, proteins, and polypeptides on the cell surface interacting with positively charged amino groups of the carrier. The highly porous polymers based on PEI and CH feature high immobilization efficiency, they are also durable, biodegradable, and non-toxic [16]. However, their effects on oleaginous microalgae such as ARA-producing Lobosphaera is underexplored.
To bridge this gap, at least partially, we studied the effects of immobilization of the microalga Lobosphaera sp. IPPAS C-2047. This is the first report on the responses of the fatty acid profile of an oleaginous microalga to immobilization on the CH- and PEI-based polymers and their modulation by availability of the nitrogen in the cultivation medium.
2. Materials and Methods
2.1. Cell carrier synthesis
The cross-linked CH-based polymeric cell carrier was synthesized in the Laboratory of Polymer Materials of the Kurchatov Institute Research Center from HQG 800 chitosan (ChitoClear, Iceland; 600 kDa molecular weight). Briefly, A 2% aqueous solution of CH was mixed with 2% acetic acid and glutaraldehyde was added (1% of the amount of chitosan, by weight). Lyophilization of the samples was carried out on a Alpha 2-4LSC (Martin Christ, Osterode am Harz, Germany) dryer (0.250 mBar vacuum) for 24 h; before extraction, the samples were kept in a vacuum of 0.001 mBar for 2 h [21].
The PEI-based polymeric cell carrier was synthesized by JSC “Advanced Medical Technologies” by cryopolymerization of highly branched PEI (MP Biomedicals, USA) with diethylene glycol diglycidyl ether (60% by weight) (“Contact”, Dzerzhinsk, Russia) at a temperature of –24 °C [22].
2.2. Strain and cultivation conditions
The unialgal culture of the green microalga Lobosphaera sp. (Chlorophyta, Trebouxiophyceae) strain IPPAS C-2047 was used in this work. The pre-culture was grown at 20 °C in an incubator shaker (Innova 44R, New Brunswick, New York, USA) at 120 rpm in 0.75 L flasks containing 350 mL of BG-11M medium [23] with an increased phosphorus content and the following composition (g L–1): NaNO3—0.74, KNO3—0.9, K2HPO4—0.181, KH2PO4—0.089, MgSO4·7H2O—0.075, CaCl2·2H2O—0.036, Citric acid—0.006, Ammonium citrate—0.006, Na2EDTA·2H2O—0.001, Na2CO3—0.02, trace element solution − 1 ml) for 10–12 days (before the onset of stationary phase).
Lobosphaera sp. cells were pelleted by centrifugation (5 min, 1000 rpm) from equal aliquots of the pre-culture. One group of cell pellets was washed with nitrogen-free BG-11 medium, BG-11M–N and resuspended in the same medium; another group of cell pellets was resuspended in complete BG-11M medium. The 1 ml of re-suspended cells were uniformly pipetted on the surface of cell carrier pieces (14 mm diameter, 10 mm thickness) for immobilization of the cells. The carriers with the cells immobilized on them were incubated in 100 mL cultivation flasks (Eppendorf, Germany). The suspended cultures of the Lobosphaera sp. cultures in the same media incubated under the same conditions (see below) were used as the controls; the experimental design and designation of the experimental treatments used in this work are shown in Figure 1. Both suspended and immobilized Lobosphaera sp. cells were cultured on the Innova-44R incubator shaker at 120 rpm, constant temperature of 20 °C, and constant illumination with white light (40 μM PAR quanta m–2 s–1 for 14 days.
Figure 1.
The experimental design and designation of the experimental variants.
2.3. Assay of chlorophyll and extimation of the condition of photosynthetic apparatus
Chlorophyll a + b content was determined spectrophotometrically in extracts obtained using dimethyl sulfoxide [24] or chloroform[25] (see below). The physiological condition of the culture was assessed via analysis of chlorophyll a fluorescence curves using pulse-amplitude modulation (PAM) approach [26,27]. In particular, potential maximum photochemical quantum yield of photosystem II (Qy = (Fm – Fo)/Fm = Fv/Fm and non–photochemical quenching of the electron excitation energy in the light-harvesting antenna (parameter NPQ = Fm/Fm' – 1) were monitored with a FluorCam FC 800-C (PSI, Drasov, Czech Republic) kinetic fluorescence imager. The PAM measurements of the suspended and immobilized microalgae were carried out in immunological well plates after 10-min dark adaptation. Chlorophyll fluorescence was excited at 650 nm and recorded in the red region of the spectrum (λ > 680 nm).
2.4. Analysis of cell lipid fatty acid composition
Total cell lipids were extracted from microalgae using Folch method [28]. Cells were pelleted by centrifugation, the cell pellet was homogenized in a mixture of chloroform and methanol (2:1, by volume). Distilled water was added to the homogenate in the amount of 20% of the homogenate volume, the mixture was dark-incubated overnight at a temperature of 4 °C. Chloroform was evaporated on a Heidolph Laborota 4000 rotary evaporator (Heidolph, Schwabach, Germany) at a temperature of 30 °C. As an internal standard, 50 micrograms of margaric acid (C17:0) were added to the samples. The samples were transmethylated by incubation with 2% sulfuric acid in methanol for 1.5 h at 80 °C. Fatty acid (FA) methyl esters were extracted with 2 ml of n-hexane. In the case of immobilized cells, the carriers with the cells were ground and extracted as described above.
The FA profile of total cell lipids was analyzed using an Agilent 7890A gas chromatograph (Agilent Technologies, Santa Clara, CA, USA) equipped with a 30-meter HP-5MS UI capillary column (30 m × 0.25 mm × 0.25 microns; Agilent, USA). Helium with a flow rate of 1 mL min–1 was used as a carrier gas (for more details, see [29]).
The proportion of individual FA in the total FA of the Lobosphaera sp. cell lipids was inferred from the corresponding peak area. The absolute FA contents were calculated relative to the internal standard (C17:0) peak area.
2.5. Scanning electron microscopy
The cells of the Lobosphaera sp. immobilized on the cell carriers were fixed in 2% (w/v) glutaraldehyde solution in 0.1 М sodium cacodylate buffer (pH 7.2) at room temperature for 0.5 h and then post-fixed for 4 h in 1% (w/v) ОsО4 in the same buffer, then dehydrated through graded ethanol series including anhydrous ethanol and transferred into anhydrous acetone. All samples were critical-point dried on a Dryer HCP-2 (Hitachi, Japan), coated with an Au-Pd alloy on an IB-3 Ion Coater (Eiko, Japan) and examined in a JSM-6380LA (JEOL, Tokyo, Japan) microscope at an accelerating voltage of 15 kV.
2.6. Statistical treatment
Three independent experiments were carried out, each in triplicate (n = 9). Average values ± STD are shown in the figures unless stated otherwise.
3. Results
3.1. Changes in the chlorophyll content of the culture
Effects of immobilization and N availability on the Lobosphaera sp. cultures were monitored on the chlorophyll content basis since the immobilized cells were irreversibly attached to the carriers and cannot be removed for counting. At the same time, chlorophyll is a marker sensitive enough to provide useful insights into the culture condition when counting of cell density is impossible [24,30].
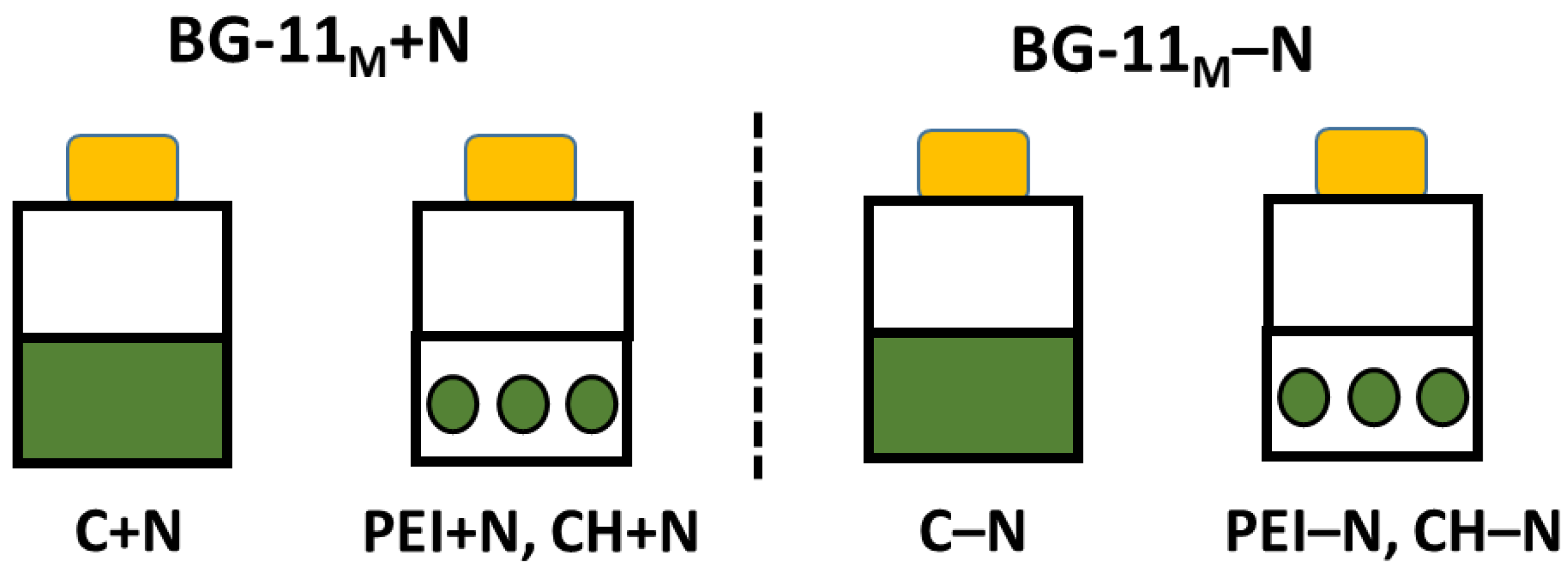
Overall, chlorophyll content in the N-deprived cultures was 35-70% less than in N-sufficient cultures (Figure 2 a, b). In turn, chlorophyll content of the PEI+N was 15-20% lower than in the N-sufficient control (C+N), whereas in the PEI–N culture it was 25-30% higher than in N-deprived suspension culture, C–N (Figure 2 a). The difference in chlorophyll content between CH-immobilized and suspended cells was insignificant regardless of N availability (Figure 2 b).
The volumetric FA content of the suspended and immobilized N-deprived cultures increased 1.5–2.2 times as compared to the initial FA content (0 d). At the same time, in the N-sufficient cultures increased 3.4–4.6 times (Figure 2 c, d). Importantly, total volumetric FA content in the cells immobilized on both carrier types was 15–35% higher than that of the suspended cultures regardless of nitrogen availability after 14 d of incubation.
Considering the specific FA content expressed per unit chlorophyll, in the end of the experiment it was 22–41% lower in N-sufficient cultures than in the N-deprived ones regardless of the immobilization state due to higher chlorophyll content of the former cultures (Figure 3 d, e). In the immobilized N-sufficient cultures, the specific FA content was higher in the PEI+N cultures than in the CH+N cultures; however, the opposite trend was observed for the N-deprived cultures.
3.2. Effects of immobilization on FA profile of the cells
Analysis of the FA profile showed that palmitic (16:0), oleic (18:1), linoleic (18:2), α-linolenic (18:3), and arachidonic (20:4, ARA) acids were the major FA in the lipids of the Lobosphaera sp. cells, either suspended or immobilized on both carrier types (Fig. 3). On the 14th d of cultivation in N-replete medium, percentages of 16:0, 18:2, and 18:3 in suspended cultures was higher, although ARA and 18:1 contents were lower than in the cells cultivated in N-free medium.
The highest ARA content was found in the PEI–N (28%; Figure 3 a, c) and C–N (25-29%; Figure 3 b, d) cells after 14 d of incubation. The lowest ARA content (16–18%) was in the cells immobilized on the CH-based carrier (Figure 3 b, d). Notably, in contrast to the N-replete and N-deprived suspended cultures, the proportions of ARA in the total FA of the immobilized cells did not differ significantly regardless of nitrogen availability.
During 14 days of incubation in the N-replete medium, the contents of 18:2 and 18:3 decreased in cells immobilized on PEI, whereas ARA increased in comparison with the N-replete suspended culture (Figure 3 a, c). On the contrary, in the cells immobilized on CH-based carrier, ARA declined, but 18:2 and 18:3 increased (Figure 3 b, d).
In the N-deprived suspended or PEI-immobilized cultures, 18:1 and ARA increased, while 18:3 decreased (Figure 3 a, c). In the CH-immobilized cells, N-deprivation was accompanied by an increase in 18:2, 16:3, and 18:3 acids along with a decrease in 18:1 and ARA (Figure 3 b, d).
On the 7th d of the experiment, volumetric ARA content of the cultures immobilized on PEI was higher than that in the suspended culture by 72% and 21% for N-sufficient and N-deprived cultures, respectively. On the contrary, the ARA content for the cells immobilized on CH was 22–27% lower than that in the corresponding suspended cultures (Figure 4 a).
During the next seven days of observation, ARA content of the PEI–N cells increased significantly in comparison with that of the suspension culture. In the case of CH-immobilized cells, a slight increase in ARA (ca. 16 %) in comparison with suspended culture was observed only in N-replete medium.
Overall, the N-sufficient cells immobilized on PEI displayed a significant increase in the specific ARA (per unit chlorophyll) content (Figure 4 b). On the day 7, the specific ARA content in the CH-immobilized cells was ca. 22% lower than that of the suspended culture. Longer incubation triggered a slight increase in ARA along with an increase in total FA content.
3.3. The Condition of photosynthetic apparatus
The N-sufficient cells displayed a monotonous increase in Fv/Fm during 14 days of cultivation regardless of immobilization status and cell carrier type (Figure 5 a, b). At the same time, NPQ values remained low (0.4–0.9) indicating a low, if any stress to the culture imposed by the selected cultivation conditions (Figure 5 c, d).
In the case of N-deprived cultures, a slight decline in Fv/Fm was recorded during the first seven days of cultivation followed by its recovery by the end of observation period (Figure 5 a, b). The PEI-N cells displayed an increase in NPQ values during the first 1–3 days of cultivation and its gradual decrease to the initial values by the last day of the experiment. The suspended cultures deprived of N showed a pronounced increase in NPQ (by 4–4.5 times relative to the initial values). Judging from the condition of the photosynthetic apparatus, the N-shortage stress was, in effect, milder in the case of the immobilized cultures than in the case of suspended cultures.
3.4. Changes in the morphology of the cells as elucidated by SEM
The Lobosphaera sp. IPPAS C-2047 is a unicellular green microalga with spherical cells of 3.5-23 µm in diameter (Fig 6 a). Numerous pores of different depth and size (10–300 microns) were revealed on the surface of PEI (Figure 6 b) and CH (Figure 6 c). Lobosphaera sp. cells at various stages of its life cycle (young cells, mature cells, and sporangia) were detected in the pores and channels of the PEI (Figure 6 d, e) and CH (Figure 6 f, g). The inner part of both types of cell carriers possessed a branched system of channels of variable diameter facilitating access into the inner bulk of the polymer for the microalgal cells (Figure 6 f). The microalgal cells were strongly attached to both carrier types, judging by their tight adherence to the surface of the polymers (Figure 6 d–g). The surface of suspended cells of the Lobosphaera sp. were smooth (Figure 6 a), while the surface of the immobilized cells became corrugated (Figure 6 e–g). The cell and the polymeric carrier surfaces were connected by the strands of extracellular polymer substances which likely strengthened the attachment of cells (Figure 6 e) and manifested the onset of biofilm formation on the surface of the cell carriers.
4. Discussion
The object of this study, Lobosphaera sp. IPPAS C-2047 is an extremotolerant microalga capable of accumulating high amounts of very long-chain FA, ARA. A prominent, arguably unique feature of microalgae from this genus is the presence of ARA in the structural and reserve lipids [5,31,32]. Therefore, the biosynthesis of ARA in Lobosphaera can be up-regulated under stress, but it can be also sizeable under conditions favoring rapid cell division. Nevertheless, the most significant induction of ARA is observed under stressful irradiances and temperatures, nutrient shortage or combinations thereof [33]. The lipogenic response of Lobosphaera as well as reduction of photosynthetic apparatus serves for the protection against oxidative stress exacerbated by the diverse environmental stresses [33,34,35]. The protection is implemented by channeling the surplus photoassimilates into the biosynthesis of triacylglycerides (TAG) rich in oleate (18:1) and, in the case of Lobosphaera, in ARA [6,34]. A similar response was evidently observed in the N-deprived suspended culture under our experimental conditions (Figures 2 c-e and 3).
Under optimal conditions, the biosynthesis of structural lipids, including glycolipids of chloroplast membranes enriched with 18:3 and 18:2, prevails. Stresses shift the lipid metabolism towards the biosynthesis of reserve TAG [34,36,37,38]. Stress e.g., N deprivation promotes degradation of chlorophyll and dismantling of chloroplast membranes manifesting itself as a decline in 18:2 and 18:3 [39,40], as was the case in our study (Figure 3). The build-up of ARA during stress-induced decline in 18:3 might reflect the role of the polyunsaturated FA in protecting photosystem I against oxidative stress [41]. Similarity of the FA profile changes observed in the PEI+N cultures (increase in 18:1 and decline in 18:2 and 18:3) and in the C–N cultures support the similarity of stress responses of the Lobosphaera sp. under those conditions. As was shown previously [42,43], immobilization of cells on PEI-based carriers is stressful per se: thus, it increased accumulation of atsaxanthin in Haematococcus pluvialis in comparison with its suspended culture. Likewise, immobilization of the Lobosphaera sp. cells on PEI might augment ARA accumulation, which can be even more significant under nitrogen deprivation. This is in line with an increase in total FA and FA/chlorophyll ratio observed in the cultures immobilized on PEI (Figure 2 c, e).
The signs of stress such as decrease in chlorophyll showed by the cells immobilized on PEI when nitrogen was ample in the medium (Figure 2 a) might stem from the restriction imposed on cell division by spatial constraints and strong binding of the microalgal cells to the carrier. Effectively, the slowdown of cell division induced by the immobilization can shrink the metabolic sink for the photoassimilates similarly to other stresses. In turn, this would trigger general stress responses including an increase in the biosynthesis of reserve lipids enriched in ARA.
An important outcome of this study is pinpointing the profound difference of the effect of immobilization on PEI and CH on lipid metabolism of Lobosphaera sp. Unlike PEI, immobilization on CH declined ARA but increased 18:2, 18:3, and 16:3 content. Possible explanations of this effect include inhibitory effect of CH on the ARA biosynthesis, modulation of lipid composition of the cell membranes. However, in contrast to the suspension culture, the nitrogen deprivation of the cells immobilized on both carriers did not augment the ARA percentage of total FA. It is therefore likely that the nitrogen shortage in the immobilized cells was partially relieved by s supply of nitrogen-containing products of the carrier biodegradation which is rich in nitrogen itself [16].
Interestingly, the FA content of the cells immobilized on CH was higher than that of the suspended culture (Figure 2 d, f). This increase was not due to increase in ARA, but to enhancement of 18:2, 18:3, and 16:3, the valuable FA harbored mostly by chloroplast membrane lipids. This might indicate that the cells immobilized on CH either divided more rapidly or possessed larger thylakoid membranes to capture more light under condition of self-shading of the cells crowded on the surface or within the volume of the carrier. The retention of the photosynthetic apparatus functionality under our experimental conditions was supported by the lack of profound decline in photosystem II efficiency (high Fv/Fm) in the immobilized cells (Figure 5 a, b). A slight decrease in Fv/Fm was noted in the suspended and immobilized cells on the first day of cultivation in N-free medium but even there it recovered rapidly. As expected, nitrogen shortage promoted an increase in thermal dissipation of the absorbed light energy manifested by an increase in NPQ [27]. However, only a moderate an increase in NPQ was observed suggesting that the stress imposed on the cultures was rather mild, and the microalgal cells successfully acclimated to it (Figure 5 c, d). On the other hand, the functional intactness of the photosynthetic apparatus of the immobilized cells (Figure 5) confirmed the suitability of the studied CH and PEI carriers for immobilization of microalgal cultures [16,20]. Indeed, these carriers turned to be sufficiently transparent and biocompatible, and their porosity determined by ice microcrystals formed during cryopolymerization facilitating the entrapment of the cells by the carrier [21,22].
Judging by the presence of numerous sporangia and young cells released from them (Figure 6 e-g), the immobilized Lobosphaera sp. cells retained the ability to grow and divide despite their strong attachment to the surface of both carrier types. Still the surface of mature cells and sporangia immobilized on the carriers was frequently more corrugated and deformed in comparison with that of the suspended cells (Figure 6 e-g). These changes might result either from shear forces resulting from strong binding to the carrier or from enhanced excretion of the extracellular polymeric substances (EPS), or from both processes [16]. The enhancement of EPS excretion by microalgal cells upon their immobilization on hard surfaces is well known; it was also documented in our experimental system (Figure 6 e). One can think that the EPS makes the attachment of cells to the carrier stronger or even irreversible facilitating the formation of biofilm [16]. This hypothesis was supported by the presence of cellular aggregates consisting of microalgae and the associated heterotrophic bacteria attached to the surface of chitosan (Figure 6 e). The EPS components (exopolysaccharides, proteins, amino acids) can be mobilized by the bacteria as additional nutrient sources [10,11].
5. Conclusions
Our results confirmed the suitability of the studied CH- and PEI-based polymeric cell carriers for immobilization of microalgal cultures. Moreover, immobilization on the PEI-based carrier augmented the accumulation of ARA and total FA accumulation by 72% (cultivation in the N-replete medium) or 81% (cultivation in the N-free medium). Surprisingly, immobilization of the Lobosphaera sp. cells on a CH-based sorbent declined ARA percentage (likely due to supply of nitrogen from the biodegradation of the carrier itself). At the same time, the CH-immobilized cells possessed higher amounts of other valuable FA such as oleic (18:2), α-linoleic (18:3), and hexadecatrienoic (16:3) than cells from the suspended cultures. In view of the advantages of attached cultivation, immobilization of Lobosphaera sp. IPPAS C-2047 on the CH- and PEI-based polymeric cell carriers is a promising avenue for the development of viable technology of the “green” ARA production from microalgae.
Author Contributions
Conceptualization, S.V. and E.L.; methodology, K.A. and S.V.; validation, A.S.; formal analysis, O.C.; investigation, K.S. and O.C.; resources, S.V. and A.M.; data curation, A.S.; writing—original draft preparation, S.V.; writing—review and editing, A.S.; visualization, S.V.; supervision, E.L.; funding acquisition, A.S. All authors have read and agreed to the published version of the manuscript.
Funding
The synthesis of polymers, the estimation of fatty acids and pigments in suspended and immobilized microalgae were funded by the Russian Science Foundation, grant number 23-44-00006. The analysis of photosynthetic apparatus condition and electron microscopy studies were funded by the Russian Science Foundation, grant number 23-24-00122. The electron microscopy studies were carried out at the User Facilities Center of M.V. Lomonosov Moscow State University.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data are available from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Barbosa, M.J.; Janssen, M.; Sudfeld, C.; D'Adamo, S.; Wijffels, R.H. Hypes, hopes, and the way forward for microalgal biotechnology. Trends Biotechnol 2023. [CrossRef]
- Shahi Khalaf Ansar, B.; Kavusi, E.; Dehghanian, Z.; Pandey, J.; Asgari Lajayer, B.; Price, G.W.; Astatkie, T. Removal of organic and inorganic contaminants from the air, soil, and water by algae. Environmental science and pollution research international 2022. [CrossRef]
- Brodie, J.; Chan, C.X.; De Clerck, O.; Cock, J.M.; Coelho, S.M.; Gachon, C.; Grossman, A.R.; Mock, T.; Raven, J.A.; Smith, A.G., et al. The Algal Revolution. Trends Plant Sci 2017. [CrossRef]
- Cohen, Z.; Khozin-Goldberg, I. Searching for PUFA-rich microalgae. In Single Cell Oils, 2 ed.; Cohen, Z., Ratledge, C., Eds. American Oil Chemists’ Society: Champaign IL, 2010; pp. 201-224.
- Bigogno, C.; Khozin-Goldberg, I.; Boussiba, S.; Vonshak, A.; Cohen, Z. Lipid and fatty acid composition of the green oleaginous alga Parietochloris incisa, the richest plant source of arachidonic acid. Phytochemistry 2002, 60, 497-503. [CrossRef]
- Khozin-Goldberg, I.; Iskandarov, U.; Cohen, Z. LC-PUFA from photosynthetic microalgae: occurrence, biosynthesis, and prospects in biotechnology. Applied microbiology and biotechnology 2011, 10.1007/s00253-011-3441-x, 1-11. [CrossRef]
- Dumancas, G.G. Arachidonic Acid: Dietary Sources and General Functions; nova science publishers, incorporated: 2012.
- Dragos, A.; Kiesewalter, H.; Martin, M.; Hsu, C.Y.; Hartmann, R.; Wechsler, T.; Eriksen, C.; Brix, S.; Drescher, K.; Stanley-Wall, N., et al. Division of Labor during Biofilm Matrix Production. Curr Biol 2018, 28, 1903-1913 e1905. [CrossRef]
- Branda, S.S.; Vik, Å.; Friedman, L.; Kolter, R. Biofilms: the matrix revisited. Trends in microbiology 2005, 13, 20-26. [CrossRef]
- Nozhevnikova, A.; Botchkova, E.; Plakunov, V. Multi-species biofilms in ecology, medicine, and biotechnology. Microbiology 2015, 84, 731-750. [CrossRef]
- Roeselers, G.; van Loosdrecht, M.C.; Muyzer, G. Heterotrophic pioneers facilitate phototrophic biofilm development. Microbial Ecology 2007, 54, 578-585. [CrossRef]
- Costerton, J.W. Overview of microbial biofilms. Journal of Industrial Microbiology and Biotechnology 1995, 15, 137-140. [CrossRef]
- Danaee, S.; Heydarian, S.M.; Ofoghi, H.; Varzaghani, N.B. Optimization, upscaling and kinetic study of famine technique in a microalgal biofilm-based photobioreactor for nutrient removal. Environmental Technology & Innovation 2021. [CrossRef]
- Orfanos, A.G.; Manariotis, I.D. Algal biofilm ponds for polishing secondary effluent and resource recovery. Journal of Applied Phycology 2019. [CrossRef]
- Kesaano, M.; Sims, R.C. Algal biofilm based technology for wastewater treatment. Algal Research 2014, 5, 231-240. [CrossRef]
- Vasilieva, S.; Lobakova, E.; Solovchenko, A. Biotechnological Applications of Immobilized Microalgae. Environmental Biotechnology Vol. 3 2021, 193-220. [CrossRef]
- de-Bashan, L.E.; Bashan, Y. Immobilized microalgae for removing pollutants: review of practical aspects. Bioresource Technology 2010, 101, 1611-1627. [CrossRef]
- Eroglu, E.; Smith, S.M.; Raston, C.L. Application of Various Immobilization Techniques for Algal Bioprocesses. In Biomass and Biofuels from Microalgae, Springer: 2015; pp. 19-44.
- Moreno-Garrido, I. Microalgae immobilization: current techniques and uses. Bioresource technology 2008, 99, 3949-3964. [CrossRef]
- Vasilieva, S.; Shibzukhova, K.; Morozov, A.; Solovchenko, A.; Bessonov, I.; Kopitsyna, M.; Lukyanov, A.; Chekanov, K.; Lobakova, E. Immobilization of microalgae on the surface of new cross-linked polyethylenimine-based sorbents. Journal of biotechnology 2018, 281, 31-38. [CrossRef]
- Romanova, O.; Grigor’ev, T.; Goncharov, M.; Rudyak, S.; Solov’yova, E.; Krasheninnikov, S.; Saprykin, V.; Sytina, E.; Chvalun, S.; Pal’tsev, M. Chitosan as a modifying component of artificial scaffold for human skin tissue engineering. Bulletin of experimental biology and medicine 2015, 159, 557-566. [CrossRef]
- Nuzhdina, A.V.; Morozov, A.S.; Kopitsyna, M.N.; Strukova, E.N.; Shlykova, D.S.; Bessonov, I.V.; Lobakova, E.S. Simple and versatile method for creation of non-leaching antimicrobial surfaces based on cross-linked alkylated polyethyleneimine derivatives. Materials Science and Engineering: C 2017, 70, 788-795. [CrossRef]
- Stanier, R.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Microbiology and Molecular Biology Reviews 1971, 35, 171-205. [CrossRef]
- Solovchenko, A.; Merzlyak, M.; Khozin-Goldberg, I.; Cohen, Z.; Boussiba, S. Coordinated carotenoid and lipid syntheses induced in Parietochloris incisa (Chlorophyta, Trebouxiophyceae) mutant deficient in Δ5 desaturase by nitrogen starvation and high light. Journal of Phycology 2010, 46, 763-772. [CrossRef]
- Wellburn, A. The spectral determination of chlorophyll a and chlorophyll b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. Journal of Plant Physiology 1994, 144, 307-313. [CrossRef]
- Strasser, R.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a fluorescence: a signature of photosynthesis, Papageorgiou, G., Govindjee, Eds. Springer: 2004; pp. 321–362.
- Maxwell, K.; Johnson, G. Chlorophyll fluorescence-a practical guide. Journal of Experimental Botany 2000, 51, 659-668. [CrossRef]
- Folch, J.; Lees, M.; Sloane-Stanley, G. A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem 1957, 226, 497–509. [CrossRef]
- Solovchenko, A.; Gorelova, O.; Selyakh, I.; Pogosyan, S.; Baulina, O.; Semenova, L.; Chivkunova, O.; Voronova, E.; Konyukhov, I.; Scherbakov, P. A novel CO2-tolerant symbiotic Desmodesmus (Chlorophyceae, Desmodesmaceae): Acclimation to and performance at a high carbon dioxide level. Algal Research 2015, 399–410. [CrossRef]
- Solovchenko, A.; Khozin-Goldberg, I.; Cohen, Z.; Merzlyak, M. Carotenoid-to-chlorophyll ratio as a proxy for assay of total fatty acids and arachidonic acid content in the green microalga Parietochloris incisa. Journal of Applied Phycology 2009, 21, 361-366. [CrossRef]
- Khozin-Goldberg, I.; Bigogno, C.; Shrestha, P.; Cohen, Z. Nitrogen starvation induces the accumulation of arachidonic acid in the freshwater green alga Parietochloris incisa (Trebuxiophyceae). Journal of Phycology 2002, 38, 991-994. [CrossRef]
- Bigogno, C.; Khozin-Goldberg, I.; Cohen, Z. Accumulation of arachidonic acid-rich triacylglycerols in the microalga Parietochloris incisa (Trebuxiophyceae, Chlorophyta). Phytochemistry 2002, 60, 135-143. [CrossRef]
- Solovchenko, A.; Khozin-Goldberg, I.; Didi-Cohen, S.; Cohen, Z.; Merzlyak, M. Effects of light intensity and nitrogen starvation on growth, total fatty acids and arachidonic acid in the green microalga Parietochloris incisa. Journal of Applied Phycology 2008, 20, 245-251. [CrossRef]
- Solovchenko, A. Physiological role of neutral lipid accumulation in eukaryotic microalgae under stresses. Russian Journal of Plant Physiology 2012, 59, 167-176. [CrossRef]
- Solovchenko, A.E.; Merzlyak, M.N.; Chivkunova, O.B.; Reshetnikova, I.V.; Khozin-Goldberg, I.; Didi-Cohen, S.; Cohen, Z. Effects of light irradiance and nitrogen starvation on theaccumulation of arachidonic acid by the microalga Parietochloris incisa. Vestnik Moskovskogo Universiteta Seriya 16 Biologiya 2008, 49-49.
- Li-Beisson, Y.; Thelen, J.J.; Fedosejevs, E.; Harwood, J.L. The lipid biochemistry of eukaryotic algae. Prog Lipid Res 2019, 74, 31-68. [CrossRef]
- Wacker, A.; Piepho, M.; Harwood, J.L.; Guschina, I.A.; Arts, M.T. Light-induced changes in fatty acid profiles of specific lipid classes in several freshwater phytoplankton species. Frontiers in Plant Science 2016, 7. [CrossRef]
- Guschina, I.A.; Harwood, J.L. Algal Lipids and Their Metabolism. In Algae for Biofuels and Energy, Borowitzka, M.A., Moheimani, N.R., Eds. Springer: Dordrecht, Heidelberg, New York, London, 2013; pp. 17-36.
- Kokabi, K.; Gorelova, O.; Zorin, B.; Didi-Cohen, S.; Itkin, M.; Malitsky, S.; Solovchenko, A.; Boussiba, S.; Khozin-Goldberg, I. Lipidome Remodeling and Autophagic Respose in the Arachidonic-Acid-Rich Microalga Lobosphaera incisa Under Nitrogen and Phosphorous Deprivation. Frontiers in plant science 2020, 11. [CrossRef]
- Kugler, A.; Zorin, B.; Didi-Cohen, S.; Sibiryak, M.; Gorelova, O.; Ismagulova, T.; Kokabi, K.; Kumari, P.; Lukyanov, A.; Boussiba, S. Long-chain polyunsaturated fatty acids in the green microalga Lobosphaera incisa contribute to tolerance to abiotic stresses. Plant and Cell Physiology 2019, 60, 1205-1223. [CrossRef]
- Klyachko-Gurvich, G.; Tsoglin, L.; Doucha, J.; Kopetskii, J.; Shebalina, I.; Semenenko, V. Desaturation of fatty acids as an adaptive response to shifts in light intensity 1. Plant Physiol 1999, 107, 240-249. [CrossRef]
- Lee, J.B.; Park, Y.H.; Jeon, M.S.; Kim, S.; Choi, Y.-E. Polyethylenimine linked with chitosan improves astaxanthin production in Haematococcus pluvialis. Applied Microbiology and Biotechnology 2022, 1-12. [CrossRef]
- Yoshitomi, T.; Shimada, N.; Iijima, K.; Hashizume, M.; Yoshimoto, K. Polyethyleneimine-induced astaxanthin accumulation in the green alga Haematococcus pluvialis by increased oxidative stress. Journal of bioscience and bioengineering 2019, 128, 751-754. [CrossRef]
Figure 2.
Total chlorophyll (a, b) and (c, d) fatty acid content as well as (d, e) their ratios in the suspended (C+N, C-N) and immobilized (PEI+N, PEI-N, CH+N, CH-N) either on PEI (a, c, e) or CH (b, d, f) Lobosphaera sp. IPPAS C–2047 cells grown either in N-replete (C+N, PEI+N, CH+N) or N-free (C-N, PEI-N, CH-N). Data are presented as mean ± STD (n = 9). Lowercase letters indicate significant differences between experimental variants at p < 0.05.
Figure 2.
Total chlorophyll (a, b) and (c, d) fatty acid content as well as (d, e) their ratios in the suspended (C+N, C-N) and immobilized (PEI+N, PEI-N, CH+N, CH-N) either on PEI (a, c, e) or CH (b, d, f) Lobosphaera sp. IPPAS C–2047 cells grown either in N-replete (C+N, PEI+N, CH+N) or N-free (C-N, PEI-N, CH-N). Data are presented as mean ± STD (n = 9). Lowercase letters indicate significant differences between experimental variants at p < 0.05.
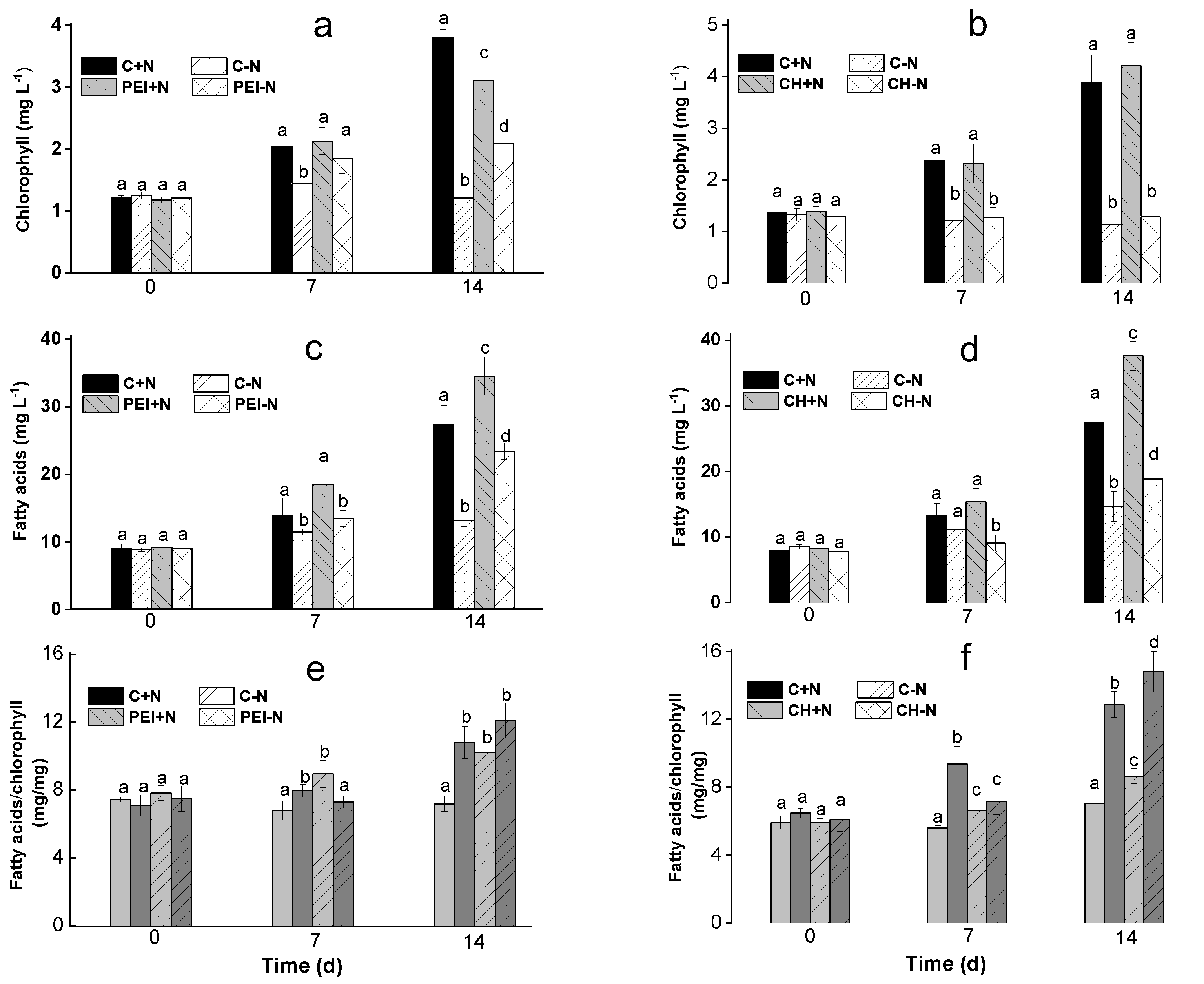
Figure 3.
The profile of the FA most abundant in total lipids of suspended (C+N, C-N) and immobilized (PEI+N, PEI-N, CH+N, CH-N , 4) either on PEI (a, c) or CH (b, d) cells of Lobosphaera sp. IPPAS C-2047 incubated in complete (C+N, PEI+N, CH+N) or N-free (C-N, PEI-N, CH-N) BG11M medium for 7 (a, b) or 14 days (c, d) of incubation. Data are presented as mean ± STD (n = 9). Lowercase letters indicate significant differences between experimental variants at p < 0.05.
Figure 3.
The profile of the FA most abundant in total lipids of suspended (C+N, C-N) and immobilized (PEI+N, PEI-N, CH+N, CH-N , 4) either on PEI (a, c) or CH (b, d) cells of Lobosphaera sp. IPPAS C-2047 incubated in complete (C+N, PEI+N, CH+N) or N-free (C-N, PEI-N, CH-N) BG11M medium for 7 (a, b) or 14 days (c, d) of incubation. Data are presented as mean ± STD (n = 9). Lowercase letters indicate significant differences between experimental variants at p < 0.05.
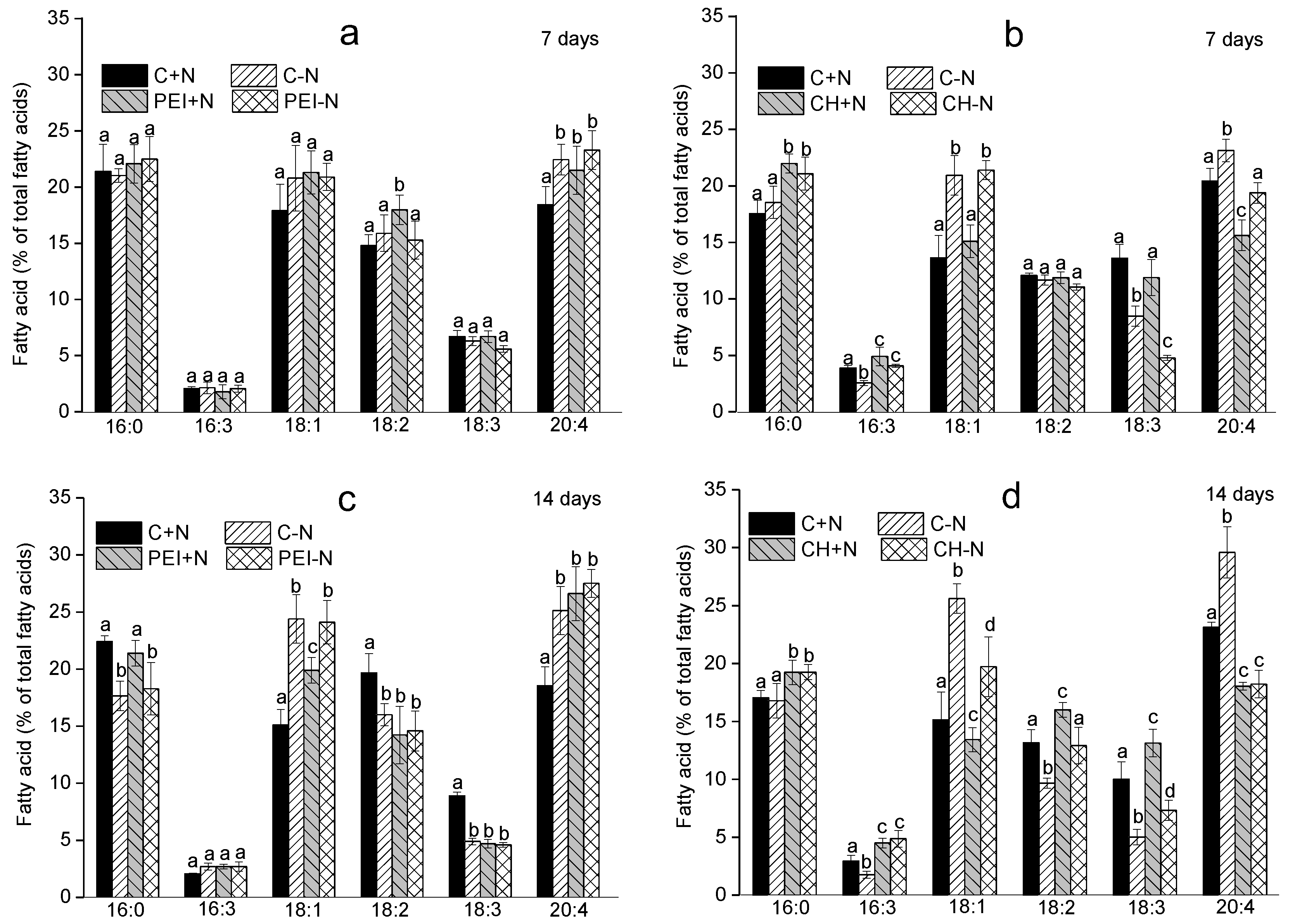
Figure 4.
The difference of arachidonic acid proportion of total fatty acids in the immobilized Lobosphaera sp. IPPAS C-2047 cells expressed (a) per unit culture volume or (b) per unit chlorophyll and calculated as percentages of the corresponding values recorded for the suspended cultures incubated under the same conditions for 7 d or 14 d
Figure 4.
The difference of arachidonic acid proportion of total fatty acids in the immobilized Lobosphaera sp. IPPAS C-2047 cells expressed (a) per unit culture volume or (b) per unit chlorophyll and calculated as percentages of the corresponding values recorded for the suspended cultures incubated under the same conditions for 7 d or 14 d
Figure 5.
Changes in (a, b) Fv/Fm and (c, d) NPQ of Lobosphaera sp. IPPAS C-2047 cells either suspended (C+N, C-N) or immobilized (PEI+N, PEI-N, CH+N, CH-N) on PEI (a, c) or CH (b, d) cells incubated in complete (C+N, PEI+N, CH+N) or N-free (C-N, PEI-N, CH-N) BG11M medium.
Figure 5.
Changes in (a, b) Fv/Fm and (c, d) NPQ of Lobosphaera sp. IPPAS C-2047 cells either suspended (C+N, C-N) or immobilized (PEI+N, PEI-N, CH+N, CH-N) on PEI (a, c) or CH (b, d) cells incubated in complete (C+N, PEI+N, CH+N) or N-free (C-N, PEI-N, CH-N) BG11M medium.
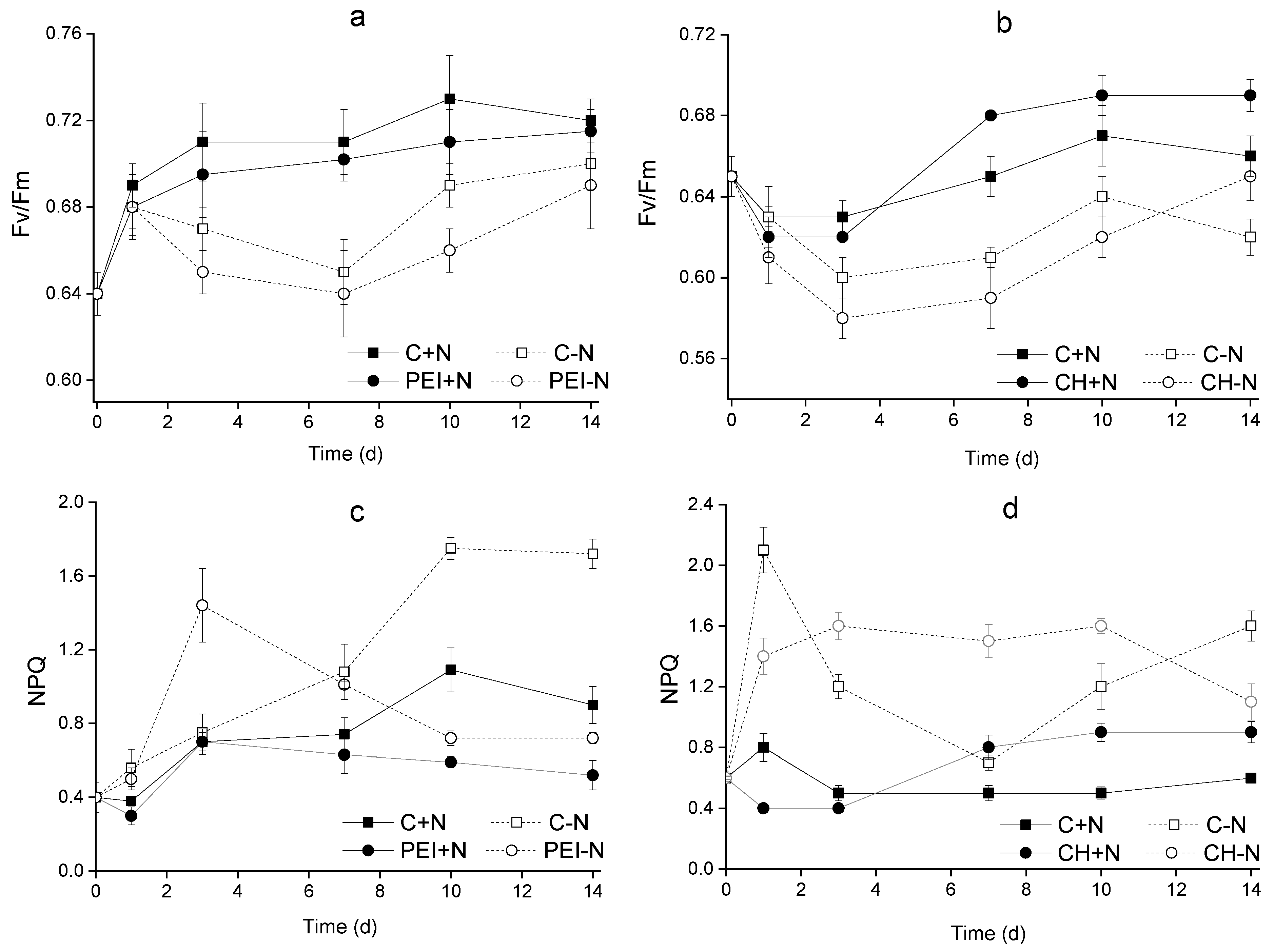
Figure 6.
Scanning electron micrographs of suspended Lobosphaera sp. IPPAS C-2047 cells (a), the surface of PEI (b) and CH (c) carriers and Lobosphaera sp. cells immobilized on PEI (d, e) and CH (f, g). VC vegetative cells, S sporangia, EPS extracellular polymeric substances. Asterisks indicate the pores on the carrier surface.
Figure 6.
Scanning electron micrographs of suspended Lobosphaera sp. IPPAS C-2047 cells (a), the surface of PEI (b) and CH (c) carriers and Lobosphaera sp. cells immobilized on PEI (d, e) and CH (f, g). VC vegetative cells, S sporangia, EPS extracellular polymeric substances. Asterisks indicate the pores on the carrier surface.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



