
Submitted:
07 March 2023
Posted:
08 March 2023
You are already at the latest version
Abstract
During the progression from ductal carcinoma in situ (DCIS) to invasive breast cancer (IBC), cells have to overcome the physically restraining basement membrane (BM) which compartmentalizes the epithelium from the stroma. Since the extracellular matrix (ECM) of the epithelial and stromal compartment is biochemically and physically distinct from one another, the progression demands a certain degree of cellular plasticity being essentially required for a primary tumor to become invasive. The Epithelial-to-Mesenchymal Transition depicts such a cell program equipping cancer cells with features allowing for dissemination from the epithelial entity and stromal invasion on the single-cell level. Here, we investigated the reciprocal interference between an altering tumor microenvironment and the EMT-phenotype in vitro. BM-typical collagen IV and stroma-typical collagen I coatings were applied as provisional 2-D matrices. Pro-inflammatory growth factors were introduced to improve tissue mimicry. Whereas the growth on coated surfaces did only slightly affect the EMT-phenotype, the combinatorial action of collagen with growth factor TGF-β1 induced prominent phenotypic changes. However, the EMT-induction was independent of the collagen type and cellular accessibility for EMT-like changes was strongly cell line dependent. Summarizing the entire body of data, we computed an EMT-phenotyping model that was used to decide on cellular EMT-status and estimate EMT-like changes. We confirmed that miR200c-mediated reversion of mesenchymal MDA-MB-231 cells is reflected by our EMT-phenotype model emphasizing its potential to predict the therapeutic efficacy of EMT-targeting drugs in the future.
Keywords:
breast cancer
; EMT
; tumor microenvironment
; collagen coatings
; EMT-phenotyping
; shape factors
1. Introduction
Epithelial-to-Mesenchymal Transition (EMT) is known as a crucial part of embryogenesis for nearly half a century, but its critical role in cancer metastasis was revealed only recently in the early 2000’s [1,2]. Since then, a continuous increase in the interest on understanding the role of EMT in cancer metastasis has been reflected in about 6000 publications in 2019 [3]. While it is controversially discussed how to understand EMT and its relevance during and for metastasis [4], three key features are commonly attributed to EMT or EMT-like changes.
First, the loss of proteins characterizing an epithelial phenotype and the acquisition of mesenchymal proteins is considered the basis of EMT (EMT-hallmarks) [5]. Driven by signals received from the tumor microenvironment (TME) [6], EMT-relevant transcription factors (e.g. SNAIL, SLUG, TWIST) downregulate epithelial and/or upregulate mesenchymal genes that cause the re-organization of the cell cytoskeleton [1]. The resultant phenotype is giving up its cobblestone-like epithelial morphology as the cell-cell junctions are abrogated and cells adopt a more spindle-like, elongated shape with a front-back polarity (Morphology). Third, as a consequence of epithelial depletions and mesenchymal fortifications, cellular motility is highly increased (Motility). An additional feature is the enhanced secretion of extracellular matrix (ECM)-degrading enzymes, promoting motility and helping cells to better cope with migration and invasion that accompanies metastasis [7].
Importantly, EMT has to be understood as a concept of cellular plasticity (Epithelial-to-Mesenchymal Plasticity (EMP)) and adaptability. It is a reversible and non-binary transition that is not necessarily completed but rather partially fulfilled (partial EMT) [3,8]. Intermediate hybrid states (E/M-states) possess both, epithelial and mesenchymal features and the degree of transition is governed by the contextuality of signaling within the tumor microenvironment, the developmental lineage of the distinct cancer types and (epi-) genetic alterations and regulations [3,9,10,11]. This trans-differentiation between epithelial and mesenchymal phenotypes has been described for all kinds of carcinoma [12]. Aside from lung cancer, colorectal cancer, hepatocellular carcinoma and prostate cancer it is breast cancer that has been tremendously studied and proven clinically relevant in the context of EMT [12,13,14,15,16]. Based on gene expression profiling and clinical outcome, breast cancer can be classified into four intrinsic subtypes with specific molecular marker expression and increasing malignancies [17]. Luminal A and Luminal B subtypes express the estrogen receptor (ER) and the progesterone receptor (PR) (Luminal A > Luminal B) and both subtypes are fairly sensitive to endocrine therapies. HER2 subtype lacks the latter sensitivity (as being ER/PR negative) but displays an overexpression of human epidermal growth factor receptor 2 (HER2) [18,19]. Absence of ER and HER2 is descriptive for the basal-like subtype, which is considered a phenotype with high mutation load and poor prognosis [20,21]. Triple negative breast cancer (TNBC), a subgroup, representing 70 - 80% of basal-like breast cancers, is characterized negative for ER, PR and HER2 [19,22]. It is the most aggressive form of breast cancer due to a synergism of poor treatment options and a high metastatic potential, which presumably relies on EMT-like changes enabling DCIS-to-IBC progression [23,24,25].
The transmembrane protein E-cadherin can be considered as “the guardian” of an epithelial phenotype. The extracellular domains of E-cadherin of each cell entangle with the extracellular domains on neighboring cells, leading to the establishment of adherens junctions [26]. Once downregulated, it is not only the physical/mechanical rupture that dissolves the epithelial phenotype. Intracellularly bound β-catenin (within the cytoplasmic cell adhesion complex) can translocate into the nucleus once E-cadherin is internalized, and act as a transcription factor towards EMT [27,28].
It is well accepted, that a switch in cellular intermediate filament (IF) usage from cytokeratin to vimentin occurs during EMT [3,29]. Vimentin is a network-forming type III intermediate filament and may be considered as the counterpart to E-cadherin, also because its expression is mainly restricted to mesenchymal cells [30,31]. By maintaining cytoskeletal integrity and mechanical strength, vimentin cushions traction stress during single-cell migration [32]. Apart from this load-bearing function, vimentin promotes microtubule polarity which is a prerequisite for directed cell migration [30,33]. Vimentin IF (VIF) maturation depends on microtubular transport. Whilst providing the infrastructure for VIF network assembly, a long-lasting template of microtubule’s architecture is simultaneously formed. Considering the fast turnover of microtubules (10 times faster than VIF), it appears, that vimentin’s “memory” function eventually guides and enables persistent microtubules-mediated cell polarization and consequently directional migration [34].
The EMT machinery can be induced by extracellular stimuli in multiple ways, mainly by soluble factors like TGF-β, EGF or HGF [5,35]. However, it was shown that solid components of the ECM trigger EMT in lung cancer. Cells, cultured on a type I collagen gel activated autocrine TGF-β3 signaling, that in turn induced EMT, which is based on collagen I fiber recognition via integrins [35]. Likewise, Shawn P. Carey et al. created a 3-D collagen I matrix with defined mechanical properties, mimicking the stroma of the mammary gland. Incorporation of non-malignant breast epithelial cells into this scaffold upregulated mesenchymal genes, which was attributed to both, biochemical and mechanical stimuli of the matrix. Interestingly, insertion of the same cell line into a basement membrane - mimicking Matrigel (containing mainly collagen IV and laminin) could not provoke EMT-like changes. The authors concluded, that the distinct ECM composition of epithelial (Matrigel) and stromal tissue (collagen I) differentiated between EMT or not [36]. Furthermore, using a xenograft breast cancer mouse model, M. Vidal et al. reported that only cancer cells at the interface of the tumor and its stroma, i.e. cells that are directly exposed to the ECM of the stromal compartment express vimentin whereas cells in the core region of the tumor maintain cytokeratin expression [37]. Taken together, it appears that signals cancer cells receive from the distinct ECM comprised within the epithelial (DCIS) and stromal (IBC) compartment ultimately dictate the present phenotype and phenotypic changes. It remains elusive to what extent the combinatorial action of solid and soluble factors in each compartment participate during initial as well as sustained EMT induction.
In this study, we used two common collagen coatings (Globular type IV collagen and fibrillar type I collagen) as provisional matrices to depict the two opposing ECM components in the mammary gland (Figure 1). Globular type IV collagen is used as a part of the basement membrane (BM), the physiological substrate of the (myo-) epithelial layer. As described above, type I collagen is the main extracellular compound within the stromal fraction. In a healthy tissue, it is well-separated from the epithelium via the BM (Figure 1), but leakage during tumor progression causes cell exposure to it.
To stress and assess the concept of “contextuality of signaling” within the TME and its importance for EMT [4,10], breast cancer cells of distinct intrinsic subtypes were subjected to a combinatorial treatment with the different collagens and the prominent EMT-inducers, TGF-β1 or EGF. Single treatment with either soluble or solid ECM components served as reference. Initially, we defined the EMT-status of the four breast cancer cell lines based on morphological aspects, EMT-marker (E-cadherin, CDH1 and Vimentin, VIM) expression and migratory behavior. MCF7 (Luminal A) and HCC1954 (HER2) cells were found to hold strong epithelial characteristics, whereas the MDA-MB-231 cell line (TNBC) was clearly attributed a mesenchymal phenotype. The second TNBC cell line MDA-MB-468 featured both, epithelial and mesenchymal characteristics (epithelial > mesenchymal), and therefore was considered an E/M-hybrid. We then monitored the three mentioned EMT features under exposure to a simplified model, taking MDA-MB-231 attributes as a positive control/reference. Cellular shape factor image analysis served as a powerful tool to predict EMT-like changes. Together with the input from protein and migration analysis, we were able to build up a computational EMT-model that hopefully helps to further understand the complexity of EMT. Besides using the model to predict EMT-phenotype and phenotypic changes that arise during cellular stimulations we exemplified its possible relevance in a miR200c inducible cell line for the facile evaluation of therapeutic efficacy of new drug candidates targeting EMT. Furthermore, our findings highlight that cellular responses are affected by combinatorial action of growth factors and ECM components requiring future 2D EMT-research to consider this context-dependency by applying similar experimental set-ups.
2. Results
2.1. EMT-status
2.1.1. EMT-Marker
EMT is characterized by phenotypic changes in protein expression relying on extrinsic (GFs, hypoxia, ECM) and intrinsic (epigenetic) regulation [40,41]. To define the EMT-status of our four breast cancer (BC) cell lines, assessing the expression of phenotypic protein markers (EMT-markers), is of fundamental importance. As described above, high levels of CDH1 are representative for an epithelial phenotype, whereas expression of VIM is a mesenchymal attribute. To this end, we performed Western blotting and qPCR analysis of the four breast cancer cell lines MCF7, HCC1954, MDA-MB-468 and MDA-MB-231 (Figure 2b,c). Both, the Luminal A and the HER2-positive cell line strongly expressed E-Cadherin, whereas they lacked vimentin expression. Considerably lower level of E-Cadherin was detected for MDA-MB-468 (2-fold lower than for MCF7), but they co-expressed VIM. Furthermore, MDA-MB-231 was the only cell line to exhibit high levels of VIM showing absence of CDH1. We confirm that the intrinsic subtype nomenclature for breast cancer matches with EMT-marker expression and malignancy correlates with the less epithelial but rather mesenchymal phenotypic marker expression. To improve this still binary system, we decided to take additional aspects into account to define the EMT-status.
2.1.2. Migration
Cellular migration is strongly dependent on cell-cell and cell-ECM adhesion [42]. Considering transcriptional repression of CDH1 and other cell-cell adhesion molecules as part of EMT, it appears logical that the resultant phenotype exhibit increased motility. VIM and cytoskeletal-dependent cell polarization further enhance the migratory ability of cancer cells during the transition. Thus, to estimate the cellular EMT-status we also evaluated the migratory behavior of the different cell lines using a wound healing assay. Migration potential was quantified by means of percentage gap-closure over time starting from 0 % at t0. Figure 2d shows that MCF7 and HCC1954 cells were able to close the gap at a similar rate. On the other hand, the other two cell lines MDA-MB-468 and MDA-MB-231 were 2 and 3 times faster, respectively. In a spheroid-based migration assay we confirmed that MDA-MB-231 cells, which fail to express CDH1 spread significantly faster (5-10 times) as the CDH1-positive cell line MCF7 (Supplementary data, Figure S1).
2.1.3. Morphology
Another feature of EMT is the re-structuring of the cytoskeleton based on spatiotemporal organization of actomyosin, microtubules, IF and other functional proteins [27,33,43,44]. Thereby, the intrinsic mechanical properties of cells are altered, which lead to changes in cellular shape [45,46]. Actin filament polymerization is a driving force for the switch from a basolateral to a front-back polarity during EMT [47,48]. Fluorescently labeling the actin cytoskeleton allows for monitoring changes in cellular shape via confocal microscopy. To describe morphological changes in cancer cells the aspect ratio AR, the cellular minor axis divided by its major axis (AR = dmin/dmax), has been used [38,49,50,51]. In the context of EMT, a value close to 1 is attributed to an epithelial, cobblestone-like morphology, whereas values close to 0 describe a spindle-like appearance. We further evaluated nuclear pleomorphisms via nuclear circularity (CN). Nuclear pleomorphisms are established to be clinically relevant in diseased tissues [52,53], but unfortunately, standard 2-D in vitro cell culture results are not often able to link circularity to phenotypic changes [52,54]. Since nuclear envelop proteins are directly physically entangled with cytoskeletal proteins and since nuclear dynamics are connected to cytoskeleton-mediated migration [55,56], we expected changes in nuclear circularity during EMT progression.
Confocal imaging revealed pronounced clustering of cells for MCF7, HCC1954 and MDA-MB-468 cells, even though the latter cells appeared to bundle less tight (Figure 3). MDA-MB-231 did not cluster at all, but showed an elongated shape which is also confirmed by the image analysis. The aspect ratio of MDA-MB-231 (0.240 ± 0.157) was significantly lower in comparison to the other 3 cell lines, MCF7 (0.654 ± 0.136), HCC1954 (0.707 ± 0.128) and MDA-MB-468 (0.683 ± 0.170) which showed no significant differences amongst each other. A similar tendency was observed for the morphological assessment of the nucleus. Here, a circularity of 1 matches a perfect circle and decreasing values describe progression to ellipsoid shapes. Nuclei of MDA-MB-231 showed to have the most ellipsoid shapes (0.735 ± 0.101) that significantly differed from the other cell lines. Trending mean values of 0.808 ± 0.053 for HCC1954 and 0.877 ± 0.046 for MDA-MB-468 implied a stronger heterogeneity of nuclear circularity amongst the other 3 cell lines as compared to the AR value.
2.2. EMT-induction
To understand which factors potentially play a role for EMT induction in the mammary gland, it is crucial to become acquainted with the tissue’s architecture. As shown in Figure 1, the epithelium of the mammary duct comprises a layer of milk-producing luminal cells, which is surrounded by myoepithelial cells. Apart from their contractile competence during lactation, myoepithelial cells produce the substrate of the epithelium, the basement membrane. This thin, but dense layer consists of collagen IV, laminin and proteoglycans and is bordering epithelial from stromal compartment [6]. The ducts are encircled by the microenvironment comprising ECM, predominantly collagen I, and stromal cells (e.g. fibroblasts, endothelial cells, leukocytes). In healthy tissues and DCIS, the luminal epithelial cells will not experience the environment of the stromal compartment. It is not before the breakage of the BM of an invading tumor until epithelial cancer cells get in contact to collagen I and the multitude of factors (growth factors, cytokines and enzymes) secreted by cancer-associated cells during tumor progression [57,58,59,60]. It is strikingly evident that the resultant inflamed and desmoplastic stroma (Figure 1) bears a high potential to induce EMT.
2.2.1. EMT-marker
In the first place, to estimate the impact of the acellular stromal fraction of the mammary gland on our EMT-model, we confronted the three cell lines with collagen I (as major ECM component of the stroma) and the growth factors EGF and TGF-β1, respectively. In addition, we tried to further provoke EMT-like changes by combinatorial stimulation with all factors (Figure 4).
MCF7 only showed minor changes in protein marker expression in comparison to untreated cells after 72h incubation (Figure 4a). Normalization with the housekeeping protein revealed that CDH1 protein levels of cells grown on collagen I were reduced to a similar extend as for TGF-β1 treatment (0.41 vs. 0.54). EGF appeared to have no impact on the EMT-markers. It rather abolished the effect of collagen I and TGF-β1 on CDH1 as part of a co-treatment. Combinatorial stimulation with collagen I and TGF-β1 did not further decrease E-cadherin expression (0.52). On the contrary, none of the treatments affected VIM levels.
Protein levels of the HCC1954 cell line demonstrated the importance of the concept of contextuality of signaling for EMT induction (Figure 4b). Single treatments and treatment of collagen I with EGF were inefficient to mediate phenotypic marker changes. Interestingly, the coaction of TGF-β1 with collagen I did imposingly enhance VIM protein expression (75-fold). This was also true when EGF was additionally added to the latter two. However, CDH1 protein levels were not strongly altered. Taken together, it appears that the growth on collagen I allowed for TGF-β1 to unfold its EMT-inducing capacities even if it only caused an incomplete pEMT.
In accordance to what has been reported elsewhere [61], EGF stimulation was able to drive EMT in MDA-MB-468 cells as demonstrated in Figure 4c. Both, CDH1 and VIM protein levels underwent EMT-typical changes. VIM was highly upregulated (9.4-fold) and CDH1 levels faded considerably (0.37). Combinatorial treatments with EGF did vaguely foster EMT induction in MDA-MB-468. Minor increase of VIM protein levels was observed under treatment with collagen I or TGF-β1 or with both.
As described above, invading tumors have to face the influence of collagen I once they have degraded the BM and enter the stromal compartment. Anyway, growing in situ carcinomas arising from luminal epithelial cells eventually interfere with collagen IV within the BM as they displace myoepithelial cells or the latter are depleted as a result of tumor progression [57,58]. Consequently, examining the impact of collagen IV exerted on cancer cells which are under exposure to growth factors and comparing the outcome to growth on collagen I is of interest here.
Therefore, we included collagen IV in our EMT-marker study. Trilateral treatments (TGF-β1 + EGF + collagen) were excluded as they seemed to have no additive effect on EMT induction (data not shown). For MCF7 and MDA-MB-468 we confirmed phenotypic changes of EMT-protein levels of earlier findings (Supplementary data, Figure S2). In MCF7, after normalization, E-cadherin levels decreased for cell growth on collagen I under TGF-β1 stimulation to 60%. This decrease was even more pronouncedly on collagen IV (40%). On the contrary, VIM expression was unaffected by the tested treatments. Both collagens in combination with EGF did induce EMT-like changes in MDA-MB-468, but EGF was definitely the main cause to drive EMT (Supplementary data, Figure S2) as its single treatment exhibited the strongest effect on CDH1 and VIM levels.
Surprisingly, EMT induction experiments in HCC1954 cell line revealed that vimentin protein increment was independent of the collagen type when co-treated with TGF-β1 (Figure 4e). Thus, the data underline the necessity of collagen coatings for TGF-β1 to drive EMT. Again, it was mostly vimentin expression that changed whereas E-cadherin levels did hardly differ from untreated samples. EGF stimulation did not lead to phenotypic changes independently of the collagen type cells were seeded on (Figure 4d).
2.2.2. Morphology
The re-organization of the cytoskeleton as part of the EMT program converts cells into a phenotype with an increased motility and a spindle-like shape. As shown above, morphology of mesenchymal MDA-MB-231 cells significantly differed from the other three cell lines based on aspect ratio AR and nuclear circularity CN. EMT induction is expected to entail AR and CN approaching values comparable to those of MDA-MB-231 cells. The morphological analysis of confocal images is depicted in Figure 5.
Neither the AR nor the CN were significantly altered in the induction study for the MCF7 cell line (Figure 5a). The AR of the sample co-treated with collagen IV and TGF-β1 (0.520 ± 0.195) showed the strongest deviation from the untreated cells (0.659 ± 0.122) whereas all CN values remained comparable to control cells (0.801 ± 0.079).
Morphological assessment of confocal images of HCC1954 cells (Figure 5b) displayed significant decrease in nuclear circularity for TGF-β1 stimulated samples seeded on collagen I (P < 0.05) and collagen IV (P < 0.001). Remarkably, the AR of the latter samples also strongly deviated from the untreated cells (0.801 ± 0.077) with 0.709 ± 0.062 and 0.689 ± 0.099, respectively, even if not statistically significantly. As can be seen in Figure 5d, cells treated solely with TGF-β1 did neither change shape nor cytoskeletal architecture in comparison to untreated cells. Once grown on either collagen I or collagen IV, actin bundles (stress fibers) were formed, and the cellular outgrowth and shape differed from the control.
Furthermore, analysis of MDA-MB-468 cells revealed that CN was significantly reduced for all samples treated with EGF (P < 0.001) in comparison to the untreated cells (Figure 5c). Similarly to what has been observed in HCC1954, decrease in CN was accompanied by a significant reduction of the aspect ratio, most prominently for EGF stimulation alone (Control: 0.695 ± 0.161 vs. EGF: 0.358 ± 0.129). Cell-cell contacts diminished, and cellular shape became elongated when exposed to EGF (Figure 5e). AR and CN values of the other treatments varied marginally from those of untreated cells. In another experiment (Supplementary data, Figure S3) we showed, that the degree of morphological change depended on the concentration of EGF. Applying concentrations from 10 ng/ml to up to 50 ng/ml during a 72 h incubation period resulted in continuous reduction of AR and CN values implying a more pronounced EMT induction for higher concentrations of the growth factor. However, the highest concentration of EGF (100 ng/ml) reverted the effects on AR and CN and cells exhibited rounded shapes similar to untreated cells. Consequently, there might be an optimal range of EGF concentration to induce EMT-like changes.
2.2.3. Migration
Based on our previous findings, we conducted migration assays with a focus on treatments that have shown considerable effects on EMT-marker expression and morphology during EMT induction.
Apart from moderate changes in CDH1 levels after exposure to TGF-β1 or collagen I/IV, MCF7 cells mainly retained their phenotype. According to the migration analysis depicted in Figure 6a, these changes were insufficient to alter the migratory behavior of MCF7.
Additionally, we included combinatorial treatments for HCC1954 cells (Figure 6b). Interestingly, coaction of TGF-β1 and collagen I enhanced cellular motility. Time for completing gap closure was reduced to 56 h (vs. 96 h for untreated cells). No other treatment provoked similar changes. Even the combination of TGF-β1 with collagen IV had no impact on migration, in contrast to the morphological changes discussed above.
A striking effect on migration was observed for EGF treatment in MDA-MB-468 cells. Its presence accelerated cellular migration almost twofold. The time necessary to close the gap was reduced by 46% (28 h vs. 52 h). Of note, the migration under exposure to collagen IV was strongly decelerated. Cells required 72 h for a 100% gap closure. TGF-β1 and collagen I had no influence on the cellular motility in MDA-MB-468 (Figure 6c) which is in line with the lack of morphological effects discussed above.
2.3. EMT-phenotyping model and its application
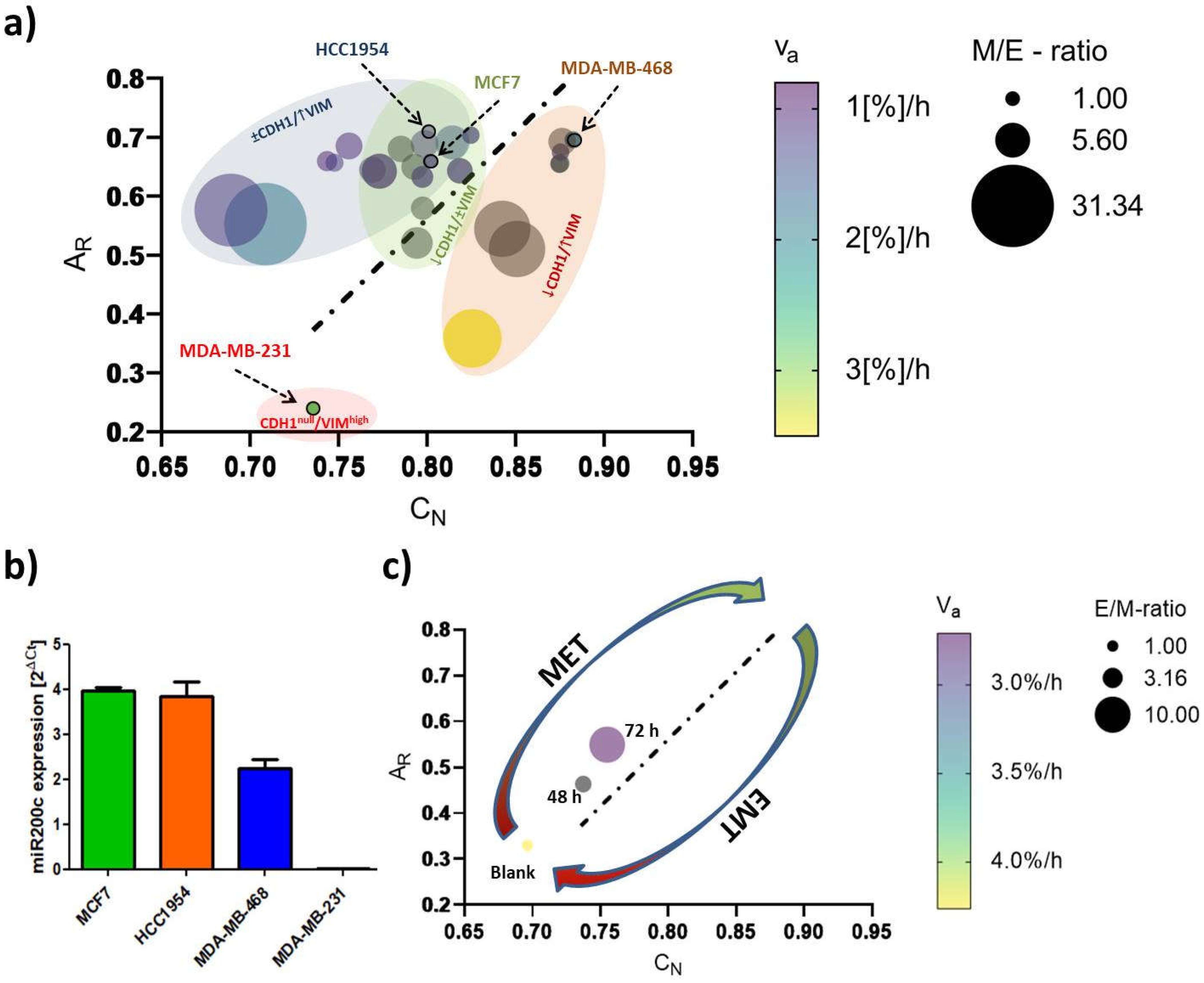
As indicated in Figure 7, we propose a phenotyping model to monitor EMT-status and EMT-like changes, which is essentially based on nuclear circularity (CN) and cellular aspect ratio (AR). We expanded it by queuing data from the aforementioned cellular EMT-protein marker settings and migratory behavior. Providing the M/E-ratio (normalized mesenchymal vimentin protein levels (M) divided by normalized E-cadherin protein levels (E)) for each treatment enabled us to correlate the magnitude of phenotypic transition on the protein level with the image-based shape analysis. Cellular motility, expressed as the apparent velocity νa ([%] gap-closure per hour) further contributed to this model. Accordingly, rounded, slowly migrating epithelial-like cells are found in the upper right corner and highly motile mesenchymal-like cells in the lower left corner of the plot. Alongside the linear regression diagonal (AR/CN) of the control cells (dotted line), cellular phenotype is transiting from a low EMT- to a high EMT-status. A decrease in E-cadherin level is accompanied by a lowering of the AR whereas the deformation of the nucleus (CN values) showed a better correlation with vimentin upregulation.
In order to validate our model in the therapeutic context, we first quantified miR200c (microRNA200c) expression in the four cell lines. miR200c is known to fulfill a regulatory function regarding the epithelial-mesenchymal state of a cell as it directly inhibits the activity of pro-EMT transcription factors ZEB1 and ZEB2 [38]. Therefore, miR200c expression is thought to correlate with E-cadherin expression and consequently with an epithelial phenotype. Indeed, miRNA expression analysis (Figure 7b) demonstrated high levels of miR200c to occur in epithelial MCF7 and HCC1954 cell lines. Moreover, miR200c expression was about halved in E/M-hybrid cell line MDA-MB-468 and absent in mesenchymal MDA-MB-231 cells. Hence, we clearly confirmed the correlation of miR200c expression levels and the underlying E/M phenotype.
Recently, it was show that the re-expression of miR-200c in vitro partially reverses the mesenchymal phenotype of MDA-MB-231 cancer cells, i.e. leading to a Mesenchymal-to-Epithelial transition (MET) [38]. Using a miR200c-inducible MDA-MB-231 cell line, we sought to exemplify a therapeutic intervention of targeting EMT i.e., by inducing MET (Figure 7c). Strikingly, applying the miR200c inducible cell line for 48 h and 72 h to our model, we monitored morphological features to follow the linear regression diagonal (y = 2.935x – 1.787) calculated above. Setting the CN-values in the equation predicted AR-values of 0.245, 0.387 and 0.438 for 0 h, 48 h and 72 h time points, respectively, which were in relatively good agreement to the actual mean AR-values (0.319, 0.421 and 0.546). Concomitantly, increased E/M-ratio and decreased apparent velocity were indicative for a transition towards a more epithelial phenotype, i.e. for the success of the modelled therapeutic intervention.
3. Discussion
Here, we established a simplified, EMT-relevant breast cancer model comprising 4 breast cancer cell lines. The triad of EMT-marker expression, morphology and migration was found to allow for a reasonable approximation of the present cellular EMT-status merging their input into a computational model phenotyping EMT in breast cancer, which we believe can serve as new platform to support EMT-related research. Moreover, biomimetic collagen coatings were shown to partially (cell line dependent) but not necessarily influence cellular EMT-phenotype upon growth factor stimulation. Surprisingly, other than reported elsewhere [36], the distinct collagen types chosen to represent the compartmentalization of the mammary gland (DCIS vs. IBC) did not differentially affect the phenotype and phenotypic transitions. Presumably, including in vivo-like cues such as 3-dimensionality and stiffness of the ECM would help to draw a final conclusion.
Underlining the current understanding of EMT in cancer research [4,5,7,8,14,27,62,63], (high) vimentin expression and absence of E-cadherin determined a fairly migratory phenotype with an elongated shape as was shown for the MDA-MB-231 cell line. Expression of the epithelial protein marker E-cadherin appeared to have a strong impact on morphological features as the three E-cadherin+ cell lines studied here exhibited rounded shapes with AR and CN values close to a value of 1. Further, its expression was correlating with an immotile phenotype as long as vimentin was not co-expressed. Indeed, even low levels of vimentin protein could be correlated with increased cellular motility over vimentin- cells as was demonstrated for MDA-MB-468 cell line. In parallel to the classification of the intrinsic subtype nomenclature, epithelial characteristics vanished towards more malignant phenotypes whereas mesenchymal features concentrated in both TNBC cell lines (Figure 2 and Figure 3).
In a recent study it was reported that Slug-mediated downregulation of E-cadherin, together with upregulation of vimentin impaired cellular morphology and increased cellular motility of the non-malignant breast epithelial cell line MCF10A [64]. Cellular circularity, which is interchangeable with the aspect ratio, was strongly decreased. Subsequent RNA interference with vimentin-targeting siRNA not only restored the circular cellular shape but also decelerated cellular motility. The authors attributed vimentin a crucial role in influencing cellular morphology and motility, also because it may directly inhibit E-cadherin expression. In another study on breast cancer E-cadherin expression was proposed to be obligatory for a round polygon shape [38]. However, the findings of our EMT induction study suggest that an upregulation of vimentin and a concomitant downregulation of E-cadherin protein levels drive important morphological changes and strengthen migratory behavior, whereas merely downregulation of E-cadherin, concluded from the MCF7 cell line, was insufficient to significantly alter either of the latter two. As seen for MDA-MB-468, EGF stimulation significantly elongated cellular and nuclear morphology as part of a pronounced EMT-induction (↓ E-cadherin, ↑↑ vimentin). The AR and CN values of MDA-MB-231blank (AR = 0.240 ± 0.157; CN = 0.735 ± 0.101) and MDA-MB-468+EGF (AR = 0.358 ± 0.129; CN = 0.825 ± 0.102) essentially converged in a concentration-dependent manner (Figure S2) implying a shift from an E/M-hybrid with mostly epithelial characteristics towards a phenotype endowed with dominant mesenchymal functionalities (Figure 5c and Figure 7a). Interestingly, upon EGF stimulation in MDA-MB-468, cellular migration was superior or at least the same to what has been observed for untreated MDA-MB-231. It took 24-28 h to close the gap in comparison to 30-35h for the already mesenchymal cell line. Moreover, we successfully transformed HCC1954 cells into a hybrid E/M phenotype entailing alterations of both shape factors and an acceleration of cellular motility. The wound healing migration assay revealed similar migratory properties under combinatorial treatment of TGF-β1 with collagen I, as was detected for MDA-MB-468blank cells (56h vs. 50h). It appears that the strongly elevated vimentin protein level is the main driver for increased cellular motility, as E-cadherin level remained unaffected. Contradictory to these findings, combinatorial treatment of TGF-β1 with type IV collagen also shown to upregulate vimentin expression did not result in increased motility.
We can only speculate about the mechanistic causality explaining how a combinatorial treatment can provoke EMT-like changes while the single components fail to do so. In accordance to our data, Stephen T. Buckley et al. examined TGF-β1-induced EMT in human alveolar epithelial cells (AEC) and found that following TGF-β1 stimulation EMT induction was enhanced when cells were grown on a collagen I matrix as compared to growth on a glass surface [43]. They based their findings on changes in cellular shape factor and cellular stiffness to be superior to the simple growth factor treatment and emphasized the role of integrins (“ECM-receptors”) for EMT induction as was reported elsewhere [65]. In this work on EMT in fibrosis, the authors elegantly ruled out the possibility that growth of AECs on the ECM would lead to increased secretion of TGF-β1 which in turn drives EMT. Instead, they provided strong evidence that the αvβ6 integrin, a receptor that binds fibronectin, activates latent TGF-β1 signaling that causes cells to undergo EMT. Still, this cannot be the explanation in our case. Firstly, αvβ6 integrin is not known for binding any collagen type and secondly, their observation was independent of TGF-β1 supply. Nevertheless, literature offers two other explanations for how integrin-growth factor receptor (GFR) interplay may modulate cellular phenotypes [66]. GFR-ligand interactions can lead to cytosolic inside-out integrin receptor activation or changes in expression pattern of integrin subunits [67]. Referring to our data, TGF-β1 signaling may have caused enhanced integrin activation/expression that would have resulted in intensified integrin signaling (through collagen-integrin interaction), finally leading to EMT-like changes. On the other hand, integrins may co-opt in GFR signaling cascades. Signals emitting from integrin activation participate in downstream processes of GFR stimulation. Consequently, those kinds of interactions may not be the driving force for EMT, but rather scale up its dimension. Hence, we have to consider, that the 48 h to 72 h time scope of our EMT induction study was insufficiently long to detect TGF-β1-mediated EMT in HCC1954. However, our findings, together with the cited literature highlight the evident role of both contextuality of signaling and the tumor microenvironment for EMT induction in vitro. Such combinatorial actions might induce EMT in DCISs helping to overcome the physiological barrier, i.e. the basement membrane, whilst sustaining a (more) mesenchymal phenotype during stromal invasion.
Confocal imaging combined with image data analysis has been proven to be a powerful tool for addressing many kinds of biological questions. In the recent years, imaging has become increasingly relevant for studies on “phenomics”, the quantification of the plurality of phenotypes that fully characterizes an organism [68]. Collective cellular properties such as cellular and subcellular morphologies are the result of genotypic expression pattern, whose quantification is readily susceptible via image data analysis [69]. In the context of EMT, a thoroughly planed study by Weikang Wang et al. stunningly demonstrated how live-cell imaging with subsequent deep image analysis disclosed heterogeneous transition dynamics upon TGF-β stimulation within one cell line [70]. They described a cell state in a 309-dimensional composite feature space of cell morphology and vimentin texture features and further revealed that spatiotemporal vimentin distribution allows for recording phenotypic alteration. The EMT-phenotyping model presented in our work is rather a snapshot approach as comparison of shape factors (CN, AR) was conducted at a specific time-point. As indicated in Figure 9, the morphological features coincided with EMT-marker expression and cellular motility. It is noteworthy that this computational model not only ranges EMT-phenotype and phenotypic changes, but presumably permits to differentiate between cell line specific transition-dynamics as indicated by the elliptic, colored areas in Figure 7a. To exemplify how this model can be used, we applied it to a miR200c inducible MDA-MB-231 cell line (Figure 7c, Figure S4). miR200c expression, which is absent in untreated MDA-MB-231 cells, correlated with an epithelial phenotype (Figure 7b). The height of miR200c expression levels further discriminated between fully epithelial and partial epithelial characteristics. Induction of miR200c in the MDA-MB-231 cell line resulted in MET, resembling a potential therapeutic intervention targeting EMT. The success of miRNA induction after 48 h and 72 h resulting in increased nuclear circularity, and cellular aspect ratios closer to 1.0 can be appreciated as the shift from the left to the right alongside the regression diagonal. Simultaneously, the increasing E/M-ratio (= reciprocal M/E-ratio) and the decrease in νa are testifying to MET. Therefore, it may be sufficient to analyze one of the aforementioned shape factors in order to predict the cellular E/M character as well as cellular motility. Thus, our model can estimate the impact of therapeutic approaches that target EMT-relevant factors on the EMT phenotype and may serve as an indicator to decide over new drug candidates during screening processes.
4. Materials and Methods
Materials and cell culture: Formaldehyde solution (≥ 36%), 4′,6–diamidino–2-phenylindole dihydrochloride (DAPI), FluorSave reagent, DNase I (recombinant, RNase-free), cOmplete™, EDTA-free Protease Inhibitor Cocktail, Phosphatase Inhibitor Cocktail 2, RIPA buffer, Tris buffered saline powder, Ponceau S Stain, Tween 20, Amersham™ Protran® Western-Blotting-Membrane (nitrocellulose) and for cell culture Eagle’s Minimum Essential Medium (EMEM), RPMI-1640 Medium, Dulbecco’s modified eagle’s medium (DMEM), fetal bovine serum (FBS), Penicillin-Streptomycin (Pen/Strep) solution, Dulbecco’s Phosphate Buffered Saline (PBS), trypsin-EDTA solution 0.05 and 0.25%, 200 mM of L-glutamine solution and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich (Taufkirchen, Germany). GAPDH Monoclonal Antibody (ZG003), Pierce™ BCA Protein Assay Kit, Novex™ 10% Tris-Glycine Mini Gels (WedgeWell™ format, 15-well), Novex™ Value™ 4-20% Tris-Glycine Mini Gels (1.0 mm, 10-well), Page Ruler™ Plus Prestained Protein Ladder 10 to 250 kDa, Tris Glycin transfer buffer, SuperSignal™ West Pico PLUS Chemiluminescent Substrate, Rhodamine Phalloidin, High capacity cDNA synthesis kit, Power SYBR™ Green PCR Master Mix, PureLink™ RNA Mini Kit and for cell culture Leibovitz's L-15 Medium and MEM Non-Essential Amino Acids Solution (100X) were purchased from Thermo Fisher Scientific (Waltham, MA, USA). Hs_CDH1_Primer Assay (QT00080143), Hs_VIM_Primer Assay (QT00095795) and Hs_GAPDH_1_SG QuantiTect Primer Assay (QT00079247) were purchased from Qiagen (Hilden, Germany). Collagen I (sc-136154), Collagen IV (sc-29010), m-IgGκ BP-HRP (sc-516102), E-cadherin Antibody (G-10) and Vimentin Antibody (V9) were ordered from Santa Cruz Biotechnology (Dallas, Texas, USA). Rotiphorese 10x SDS Page, Rotilabo®-Blotting Papers and Methanol (blotting grade) were purchased from Carl Roth (Karlsruhe, Germany). rh-TGF-β 1 (Transforming Growth Factor beta 1) and rh-EGF (Epidermal Growth Factor) were purchased from ImmunoTools (Friesoythe, Germany). HyClone trypan blue solution 0.4% in phosphate-buffered saline was obtained from FisherScientific (Hampton, NH, USA). Culture-Insert 2 Well in µ-Dish 35 mm was purchased from Ibidi (Gräfelfing, Germany). Laemmli loading buffer (4x) was purchased from VWR (Allison Park, PA, USA).
MCF7 (and MCF7 miR200c_KO) cells, a Luminal A breast cancer cell line, were cultured in EMEM supplemented with 10% FBS, 1x Pen/Strep, 1x MEM Non-Essential Amino Acids Solution and 2 mM glutamine. The HER2-positive breast cancer cell line HCC1954 was grown in RPMI-1640 Medium supplemented with 10% FBS and 1x Pen/Strep. MDA-MB-231 (and MDA-MB-231 i-miR200c) cells, a triple negative breast cancer (TNBC) cell line, were cultured in high glucose (4500 mg/L) DMEM. 10% FBS, 1x Pen/Strep and 2 mM glutamine were added to the medium. For miRNA induction medium is equipped with 5 µg/ml doxycycline hydrochloride as was described elsewhere.[38] The latter three cell lines were cultured in a humidified atmosphere with 5% CO2 at 37°C. The second TNBC cell line MDA-MB-468 was grown in L-15 Medium supplemented with 20% FBS and 1x Pen/Strep. Those cells were held in a humidified incubator with 0% CO2 at 37°C.
EMT marker – gene expression analysis: To compare EMT-marker RNA expression among the four cell lines, RT-qPCR was performed. Of each cell line 200.000 cells were seeded in a 6-well plate and cultured for 24 h. After the incubation, cells were harvested, and total RNA was isolated using the PureLink RNA mini kit according to the manufacturer’s protocol with additional DNAse I digestion. Subsequently, 1000 ng of RNA was used to synthesize cDNA using the High capacity cDNA synthesis kit. In the following, E-cadherin- and vimentin-specific primers were used to amplify and quantify RNA using Power SYBR™ Green PCR Master Mix and the qTOWER real-time PCR thermal cycler (Analytik Jena, Jena, Germany). Ct values were normalized to GAPDH RNA expression ,and delta Ct values were calculated for the comparison.
In order to quantify the miR200c expression levels, RNA was isolated using the peqGOLD Micro RNA kit (Peqlab Biotechnology GmbH, Erlangen, Germany), according to the manufacturer protocol. cDNA was synthesized with the qScript microRNA cDNA synthesis kit (Quantabio, Beverly, MA, USA). Since microRNAs are not polyadenylated, the polyA tailing reaction was performed by mixing 1 µg of RNA, 2 µL of Poly(A) Tailing Buffer, 1 µL Poly(A) polymerase, nuclease-free water up to 10 µL and incubated 60 minutes at 37°C followed by 5 minutes at 70°C. Subsequently, 9 µL of microRNA cDNA reaction mix was mixed with 1 µL reverse transcriptase and incubated 20 minutes at 42°C, plus 5 minutes at 85°C. RT-qPCR was performed in triplicates. The microRNA-191 was used as a housekeeper and each sample was analyzed in triplicates.
EMT marker – protein level analysis: To define the EMT status, protein levels of CDH1 and VIM of the 4 cell lines were analyzed via Western blotting. Of each cell line 300.000 cells were seeded in a 6-well plate and cultured for 24h. Total protein extract was isolated after incubation. Briefly, cells were washed 3 times with PBS prior to cell lyses. To each well, 70 µl of proteinase- and phosphatase-inhibitor containing RIPA buffer was added, and cells were kept on ice for 30 minutes. Hereinafter, wells were thoroughly scraped, and the extracts were transferred into 1.5 ml Eppendorf tubes. After a 10 min centrifugation step at 4°C, total protein concentration was assessed according to the manufacturer’s protocol (Pierce™ BCA Protein Assay Kit). Gels were loaded with 30 µg protein per sample and electrophoresis was run for 90 min at 120 mV. Subsequent to 1 h of protein transfer at 100 mV, blots were washed, blocked and incubated overnight using E-cadherin-, vimentin- and GAPDH-specific antibodies. HRP-bound secondary antibody was added for 1 h under exclusion of light before blots were developed.
As part of the EMT induction study, we first optimized the timepoints when to extract protein data. It should be noted that cell lines do not facultatively perform EMT and may exhibit different “transition dynamics” (fast vs. slow) upon GF treatment. Therefore we chose an optimized time point (72 h) and GF concentrations that allowed to detect EMT-like changes on the protein level in the cell lines used.
To do so, 300.000 cells were seeded to attach for 4 h. Samples included untreated cells, cells stimulated with growth factors and cells grown in collagen-coated wells (+/- growth factors). Afterwards control cells and cells growing only on collagen were washed with PBS, and pre-warmed medium was refilled. At this step, growth factors were included. Samples were either supplied with 10 ng/ml of TGF-β1 or 25 ng/ml EGF or both. Collagen I and IV coatings (2 µg collagen/cm2) of the respective wells were prepared following the manufacturer’s protocol in advance to the seeding. After 72 h incubation, samples were subjected to the aforementioned Western blotting protocol, and E-cadherin and vimentin protein levels were normalized to GAPDH-housekeeping protein level using the Image Lab™ software (Bio-Rad Laboratories, Hercules, CA, USA).
Confocal scanning microscopy – Morphological analysis: Confocal image analysis was used to assess and quantify morphological differences between cell lines and treatments. Experimental set-up was performed as described above. Briefly, sterile glass coverslips were distributed in a 24-well plate. Collagen coatings were conducted for the respective wells. Thereafter, 40.000 cells were seeded and attached for 4 h. After the growth factor treatment, cells were incubated for 72 h. Hereinafter, wells were washed 3 times with PBS before cells were fixed for 15 min with a 4% formaldehyde solution. To stain the actin cytoskeleton, cells were incubated with 8.25 µM rhodamine phalloidin solution for 40 min. Hereinafter, cells were washed another 3 times with PBS. Nuclei staining was achieved by 10 min incubation with a 0.5 µg/ml DAPI solution. Finally, after an additional washing step, samples were mounted on glass slides using FluorSave and stored at 4°C until the next day. Fluorescence images were acquired using a laser scanning microscope (Leica SP8 inverted, Software: LAS X, Leica microsystems GmbH, Wetzlar, Germany) equipped with a HC PL APO CS2 40x/1.30 and 63x/1.40 oil immersion objective. Diode lasers (405 nm) and a semiconductor laser OPSL (552 nm) were chosen for excitation, emission was detected in blue (PMT1: 410–520 nm) and yellow (PMT2: 560nm – 760nm), respectively. Images were further processed with Fiji image analysis software [39]. Nuclear circularity was calculated as , and the cellular aspect ratio as . The axis (dmin and dmax) were drawn manually.
Ibidi® migration assay: Migratory properties were analyzed as follows: To define the cellular EMT-status, 25.000 cells of each cell line were seeded in both wells of the Ibidi culture-insert. After 24h the inserts were carefully removed and the time until gap closure in between the two wells was monitored for up to 120 h using the Keyence BZ81000 Fluorescence microscope (Keyence, Osaka, Japan). Three to four pictures of different parts of the gaps were taken for each time point. The cell free area was analyzed based on a custom-made macro within the Fiji imaging software: 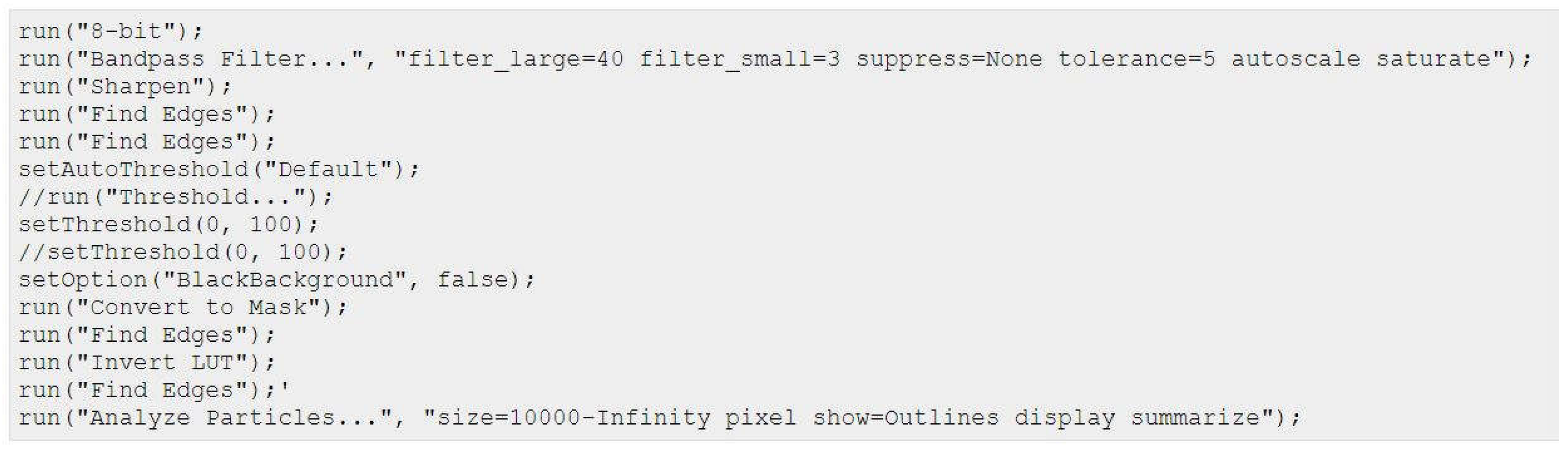
Percentage gap closure was calculated according to the following equation:
As part of the EMT induction study, cells were seeded at a density of 15.000 cells per well. Collagen coatings were prepared in advance. Similar to what has been described for the protein analysis growth factors were supplemented after cell-attachment. Samples were incubated for 48 h before the culture-inserts were removed and migration was analyzed.
5. Conclusions
The Epithelial-to-Mesenchymal transition is an ambivalent issue in the field of cancer research particularly when it comes to its impact for breast cancer metastasis and the transition from DCIS to IBC. Studies, which show the independence of metastasis from EMT-phenotypic alterations, are scrutinizing the importance of EMT.[41,71,72] Nevertheless, the concept of pEMT has broadened up its interpretation and the increasing amount of literature assessing EMT from other perspectives (e.g. epigenetics, phenomics, biomechanics) is revitalizing the field of EMT. Even conventional cell culture set-up enable to study more complex interrelations such as the here presented contextual EMT induction which was partially dependent on the combinatorial action of collagen and soluble growth factors. Establishing a more biosimilar context and illuminating EMT from different angles provided us with data allowing for our own interpretation of phenotypic changes during EMT. We hope this model will contribute to a better understanding of EMT in breast cancer and that it potentially bears relevance for therapeutic applications.
Supplementary Materials
Supplementary Information including four figures.
Author Contributions
Conceptualization, L.I., O.M. and A.R.; methodology, L.I., A.M., A.O. and G.L.; software, L.I.; validation, LI., A.M. and O.M.; formal analysis, L.I..; investigation, L.I., A.M., G.L. and A.O.; resources, O.M. and A.R..; data curation, L.I, G.L. and A.O.; writing—original draft preparation, L.I., A.M. and O.M.; writing—review and editing, O.M. and A.R.; visualization, L.I..; supervision, A.M. and O.M.; project administration, O.M.; funding acquisition, O.M.; All authors have read and agreed to the published version of the manuscript.
Funding
Please add: This research was funded by the NanoSystems Initiative Munich (NIM) and Center for NanoScience (CeNS) at Ludwig-Maximilians-Universität and partially by the European Research Council, grant number ERC-2014-StG – 637830.
Data Availability Statement
Data are available from the authors upon request.
Acknowledgments
Expert technical support by Bianca Köhler (Pharmaceutical Biotechnology, LMU) is gratefully acknowledged.
Conflicts of Interest
Olivia Merkel is a Scientific Board Member for Coriolis Pharma GmbH, AMW GmbH and Carver Biosciences and an Advisor for PARI Pharma GmbH, Boehringer-Ingelheim International GmbH, and AbbVie Deutschland GmbH on unrelated projects.
References
- Cano, M.A. Pérez-Moreno, I. Rodrigo, A. Locascio, M.J. Blanco, M.G. del Barrio, F. Portillo, M.A. Nieto, Nat. Cell Biol. 2000, 2(2): p. 76-83. [CrossRef]
- J.P. Thiery, Nat. Rev. Cancer 2002, 2(6): p. 442-54. [CrossRef]
- J. Yang, P. Antin, G. Berx, C. Blanpain, T. Brabletz, M. Bronner, K. Campbell, A. Cano, J. Casanova, G. Christofori, S. Dedhar, R. Derynck, H.L. Ford, J. Fuxe, A. García de Herreros, G.J. Goodall, A.-K. Hadjantonakis, R.Y.J. Huang, C. Kalcheim, R. Kalluri, Y. Kang, Y. Khew-Goodall, H. Levine, J. Liu, G.D. Longmore, S.A. Mani, J. Massagué, R. Mayor, D. McClay, K.E. Mostov, D.F. Newgreen, M.A. Nieto, A. Puisieux, R. Runyan, P. Savagner, B. Stanger, M.P. Stemmler, Y. Takahashi, M. Takeichi, E. Theveneau, J.P. Thiery, E.W. Thompson, R.A. Weinberg, E.D. Williams, J. Xing, B.P. Zhou, G. Sheng, E.M.T.I.A. On behalf of the, Nat. Rev. Mol. Cell Biol. 2020, 21(6): p. 341-352. [CrossRef]
- T. Brabletz, R. Kalluri, M.A. Nieto, R.A. Weinberg, Nat. Rev. Cancer 2018, 18(2): p. 128-134. [CrossRef]
- J. Banyard, D.R. Bielenberg, Connect. Tissue Res. 2015, 56(5): p. 403-413. [CrossRef]
- M.W. Conklin, P.J. Keely, Cell Adhes. Migr. 2012, 6(3): p. 249-260. [CrossRef]
- H. Son, A. Moon, Toxicological research 2010, 26(4): p. 245-252. [CrossRef]
- R. Kalluri, R.A. Weinberg, J. Clin. Invest. 2009, 119(6): p. 1420-8. [CrossRef]
- W.L. Tam, R.A. Weinberg, Nat. Med. 2013, 19: p. 1438. [CrossRef]
- R.A. Weinberg, The Biology of Cancer, 2014.
- T.Z. Tan, Q.H. Miow, Y. Miki, T. Noda, S. Mori, R.Y. Huang, J.P. Thiery, EMBO Mol. Med. 2014, 6(10): p. 1279-93. [CrossRef]
- S.V. Vasaikar, A.P. Deshmukh, P. den Hollander, S. Addanki, N.A. Kuburich, S. Kudaravalli, R. Joseph, J.T. Chang, R. Soundararajan, S.A. Mani, Br. J. Cancer 2021, 124(1): p. 259-269. [CrossRef]
- J.H. Taube, J.I. Herschkowitz, K. Komurov, A.Y. Zhou, S. Gupta, J. Yang, K. Hartwell, T.T. Onder, P.B. Gupta, K.W. Evans, B.G. Hollier, P.T. Ram, E.S. Lander, J.M. Rosen, R.A. Weinberg, S.A. Mani, Proc. Natl. Acad. Sci. U. S. A. 2010, 107(35): p. 15449-54. [CrossRef]
- S.K. Bhowmik, E. Ramirez-Peña, J.M. Arnold, V. Putluri, N. Sphyris, G. Michailidis, N. Putluri, S. Ambs, A. Sreekumar, S.A. Mani, Oncotarget 2015, 6(40): p. 42651-60. [CrossRef]
- A. Prat, C.M. Perou, Mol. Oncol. 2011, 5(1): p. 5-23. [CrossRef]
- M. Scimeca, C. Antonacci, D. Colombo, R. Bonfiglio, O.C. Buonomo, E. Bonanno, Tumour Biol. 2016, 37(4): p. 5427-35. [CrossRef]
- T. Bao, N.E. Davidson, Adv. Surg. 2008, 42: p. 249-260. [CrossRef]
- G. Arpino, D. Generali, A. Sapino, L. Del Matro, A. Frassoldati, M. de Laurentis, P. Pronzato, G. Mustacchi, M. Cazzaniga, S. De Placido, P. Conte, M. Cappelletti, V. Zanoni, A. Antonelli, M. Martinotti, F. Puglisi, A. Berruti, A. Bottini, L. Dogliotti, The Breast 2013, 22(2): p. 109-120. [CrossRef]
- L. Yin, J.-J. Duan, X.-W. Bian, S.-c. Yu, Breast Cancer Res. 2020, 22(1): p. 61. [CrossRef]
- T. Sorlie, R. Tibshirani, J. Parker, T. Hastie, J.S. Marron, A. Nobel, S. Deng, H. Johnsen, R. Pesich, S. Geisler, J. Demeter, C.M. Perou, P.E. Lønning, P.O. Brown, A.L. Børresen-Dale, D. Botstein, Proc. Natl. Acad. Sci. U. S. A. 2003, 100(14): p. 8418-23. [CrossRef]
- T.A. O'Meara, S.M. Tolaney, Oncotarget 2021, 12(5): p. 394-400. [CrossRef]
- P. Alluri, L.A. Newman, Surg. Oncol. Clin. N. Am. 2014, 23(3): p. 567-577. [CrossRef]
- D. Sarrió, S.M. Rodriguez-Pinilla, D. Hardisson, A. Cano, G. Moreno-Bueno, J. Palacios, Cancer Res. 2008, 68(4): p. 989-97. [CrossRef]
- M. Koleckova, J. Ehrmann, J. Bouchal, M. Janikova, A. Brisudova, J. Srovnal, K. Staffova, M. Svoboda, O. Slaby, L. Radova, K. Vomackova, B. Melichar, L. Veverkova, Z. Kolar, Sci. Rep. 2021, 11(1): p. 5145. [CrossRef]
- Q. Zeng, W. Li, D. Lu, Z. Wu, H. Duan, Y. Luo, J. Feng, D. Yang, L. Fu, X. Yan, Proc. Natl. Acad. Sci. U. S. A. 2012, 109(4): p. 1127-1132. [CrossRef]
- J.J. Christiansen, A.K. Rajasekaran, Cancer Res. 2006, 66(17): p. 8319-26. [CrossRef]
- M. Yilmaz, G. Christofori, Cancer Metastasis Rev. 2009, 28(1): p. 15-33. [CrossRef]
- J.M. Lee, S. Dedhar, R. Kalluri, E.W. Thompson, J. Cell Biol. 2006, 172(7): p. 973-981. [CrossRef]
- M.I. Kokkinos, R. Wafai, M.K. Wong, D.F. Newgreen, E.W. Thompson, M. Waltham, Cells Tissues Organs 2007, 185(1-3): p. 191-203. [CrossRef]
- R.A. Battaglia, S. Delic, H. Herrmann, N.T. Snider, 2018, 7. [CrossRef]
- H. Herrmann, H. Bär, L. Kreplak, S.V. Strelkov, U. Aebi, Nat. Rev. Mol. Cell Biol. 2007, 8(7): p. 562-573. [CrossRef]
- N. Costigliola, L. Ding, C.J. Burckhardt, S.J. Han, Proc. Natl. Acad. Sci. U. S. A. 2017, 114(20): p. 5195-5200. [CrossRef]
- C.-Y. Liu, H.-H. Lin, M.-J. Tang, Y.-K. Wang, Oncotarget 2015, 6(18): p. 15966-15983.
- Z. Gan, L. Ding, Christoph J. Burckhardt, J. Lowery, A. Zaritsky, K. Sitterley, A. Mota, N. Costigliola, Colby G. Starker, Daniel F. Voytas, J. Tytell, Robert D. Goldman, G. Danuser, Cell Syst. 2016, 3(3): p. 252-263.e8. [CrossRef]
- Y. Shintani, M. Maeda, N. Chaika, K.R. Johnson, M.J. Wheelock, Am. J. Respir. Cell Mol. Biol. 2008, 38(1): p. 95-104. [CrossRef]
- S.P. Carey, K.E. Martin, C.A. Reinhart-King, Sci. Rep. 2017, 7: p. 42088. [CrossRef]
- M. Vidal, L. Salavaggione, L. Ylagan, M. Wilkins, M. Watson, K. Weilbaecher, R. Cagan, Am. J. Pathol. 2010, 176(6): p. 3007-3014. [CrossRef]
- Ljepoja, C. Schreiber, F.A. Gegenfurtner, J. García-Roman, B. Köhler, S. Zahler, J.O. Rädler, E. Wagner, A. Roidl, PLoS One 2019, 14(11): p. e0224314. [CrossRef]
- J. Schindelin, I. Arganda-Carreras, E. Frise, V. Kaynig, M. Longair, T. Pietzsch, S. Preibisch, C. Rueden, S. Saalfeld, B. Schmid, J.-Y. Tinevez, D.J. White, V. Hartenstein, K. Eliceiri, P. Tomancak, A. Cardona, Nat. Methods 2012, 9(7): p. 676-682. [CrossRef]
- S. Bhatia, P. Wang, A. Toh, E.W. Thompson, Front. Mol. Biosci. 2020, 7. [CrossRef]
- M.K. Jolly, T. Celià-Terrassa, J. Clin. Med. 2019, 8(10): p. 1542.
- Schaeffer, J.A. Somarelli, G. Hanna, G.M. Palmer, M.A. Garcia-Blanco, Mol. Cell. Biol. 2014, 34(18): p. 3486-3499. [CrossRef]
- S.T. Buckley, C. Medina, A.M. Davies, C. Ehrhardt, Nanomed. Nanotechnol. Biol. Med. 2012, 8(3): p. 355-364. [CrossRef]
- M.F. Coughlin, D.R. Bielenberg, G. Lenormand, M. Marinkovic, C.G. Waghorne, B.R. Zetter, J.J. Fredberg, Clin. Exp. Metastasis 2013, 30(3): p. 237-50. [CrossRef]
- Paluch, C.-P. Heisenberg, Curr. Biol. 2009, 19(17): p. R790-R799. [CrossRef]
- T. Lecuit, P.-F. Lenne, Nat. Rev. Mol. Cell Biol. 2007, 8(8): p. 633-644. [CrossRef]
- S.E. Leggett, A.M. Hruska, M. Guo, I.Y. Wong, Cell Communication and Signaling 2021, 19(1): p. 32. [CrossRef]
- J. Shankar, I.R. Nabi, PLoS One 2015, 10(3): p. e0119954-e0119954. [CrossRef]
- Z.-X. Ren, H.-B. Yu, J.-S. Li, J.-L. Shen, W.-S. Du, Biosci. Rep. 2015, 35(3): p. e00202. [CrossRef]
- A. Sarkar, A. Barui, S. Sengupta, J. Chatterjee, S. Ghorai, A. Mukherjee. in 2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC). 2015. [CrossRef]
- J. Kim, Y. Cao, C. Eddy, Y. Deng, H. Levine, W.-J. Rappel, B. Sun, Proc. Natl. Acad. Sci. U. S. A. 2021, 118(10): p. e2024780118. [CrossRef]
- M. Schöchlin, S.E. Weissinger, A.R. Brandes, M. Herrmann, P. Möller, J.K. Lennerz, J. Pathol. Inform. 2014, 5(1): p. 40. [CrossRef]
- G. Bussolati, F. Maletta, S. Asioli, L. Annaratone, A. Sapino, C. Marchiò, E.C. Schirmer and J.I. de las Heras, Editors. 2014, Springer New York: New York, NY. p. 101-121. [CrossRef]
- C. Rena, P. Hara, T.P. A., Biochem. Cell Biol. 2014, 92(4): p. 287-295.
- P. Friedl, K. Wolf, J. Lammerding, Curr. Opin. Cell Biol. 2011, 23(1): p. 55-64. [CrossRef]
- T. Fischer, A. Hayn, C.T. Mierke, Front. Cell Dev. Biol. 2020, 8(393). [CrossRef]
- J.B. Kim, R. Stein, M.J. O'Hare, Tumour Biol. 2005, 26(4): p. 173-85. [CrossRef]
- K. Polyak, R. Kalluri, Cold Spring Harb. Perspect. Biol. 2010, 2(11): p. a003244. [CrossRef]
- Y. Mao, E.T. Keller, D.H. Garfield, K. Shen, J. Wang, Cancer metastasis reviews 2013, 32(1-2): p. 303-315. [CrossRef]
- B.S. Wiseman, Z. Werb, Science (New York, N.Y.) 2002, 296(5570): p. 1046-1049. [CrossRef]
- J.J. Campbell, A. Husmann, R.D. Hume, C.J. Watson, R.E. Cameron, Biomaterials 2017, 114: p. 34-43. [CrossRef]
- M.A. Nieto, R.Y. Huang, R.A. Jackson, J.P. Thiery, Cell 2016, 166(1): p. 21-45. [CrossRef]
- B.D. Craene, G. Berx, Nat. Rev. Cancer 2013, 13: p. 97. [CrossRef]
- K. Vuoriluoto, H. Haugen, S. Kiviluoto, J.P. Mpindi, J. Nevo, C. Gjerdrum, C. Tiron, J.B. Lorens, J. Ivaska, Oncogene 2011, 30(12): p. 1436-1448. [CrossRef]
- K.K. Kim, M.C. Kugler, P.J. Wolters, L. Robillard, M.G. Galvez, A.N. Brumwell, D. Sheppard, H.A. Chapman, Proc. Natl. Acad. Sci. U. S. A. 2006, 103(35): p. 13180-13185. [CrossRef]
- J.S. Munger, D. Sheppard, Cold Spring Harb. Perspect. Biol. 2011, 3(11): p. a005017-a005017. [CrossRef]
- G. Zambruno, P.C. Marchisio, A. Marconi, C. Vaschieri, A. Melchiori, A. Giannetti, M. De Luca, J. Cell Biol. 1995, 129(3): p. 853-65. [CrossRef]
- Houle, D.R. Govindaraju, S. Omholt, Nat. Rev. Genet. 2010, 11(12): p. 855-866. [CrossRef]
- K. Smith, F. Piccinini, T. Balassa, K. Koos, T. Danka, H. Azizpour, P. Horvath, Cell Syst. 2018, 6(6): p. 636-653. [CrossRef]
- W. Wang, D. Douglas, J. Zhang, S. Kumari, M.S. Enuameh, Y. Dai, C.T. Wallace, S.C. Watkins, W. Shu, J. Xing, Sci. Adv. 2020, 6(36): p. eaba9319. [CrossRef]
- H. Tian, R. Lian, Y. Li, C. Liu, S. Liang, W. Li, T. Tao, X. Wu, Y. Ye, X. Yang, J. Han, X. Chen, J. Li, Y. He, M. Li, J. Wu, J. Cai, Nat. Commun. 2020, 11(1): p. 5127. [CrossRef]
- M.K. Jolly, K.E. Ware, S. Gilja, J.A. Somarelli, H. Levine, Mol. Oncol. 2017, 11(7): p. 755-769. [CrossRef]
Figure 1.
Schematic cross-section of a mammary duct during cancer progression; The left panel shows a healthy tissue in the mammary gland. The mammary duct is formed by an epithelial layer surrounded by myoepithelial cells that are framed by the basement membrane. The stromal compartment is well separated from the epithelium which is also the case for DCIS. The right panel demonstrates tissue alteration as part of tumor progression as occur for IBCs. Defects in basement membrane and/or myoepithelial layer arise. Stromal and epithelial compartments are exposed to each other. The lower table lists the most prominent changes of the depicted tissue during tumor progression.
Figure 1.
Schematic cross-section of a mammary duct during cancer progression; The left panel shows a healthy tissue in the mammary gland. The mammary duct is formed by an epithelial layer surrounded by myoepithelial cells that are framed by the basement membrane. The stromal compartment is well separated from the epithelium which is also the case for DCIS. The right panel demonstrates tissue alteration as part of tumor progression as occur for IBCs. Defects in basement membrane and/or myoepithelial layer arise. Stromal and epithelial compartments are exposed to each other. The lower table lists the most prominent changes of the depicted tissue during tumor progression.
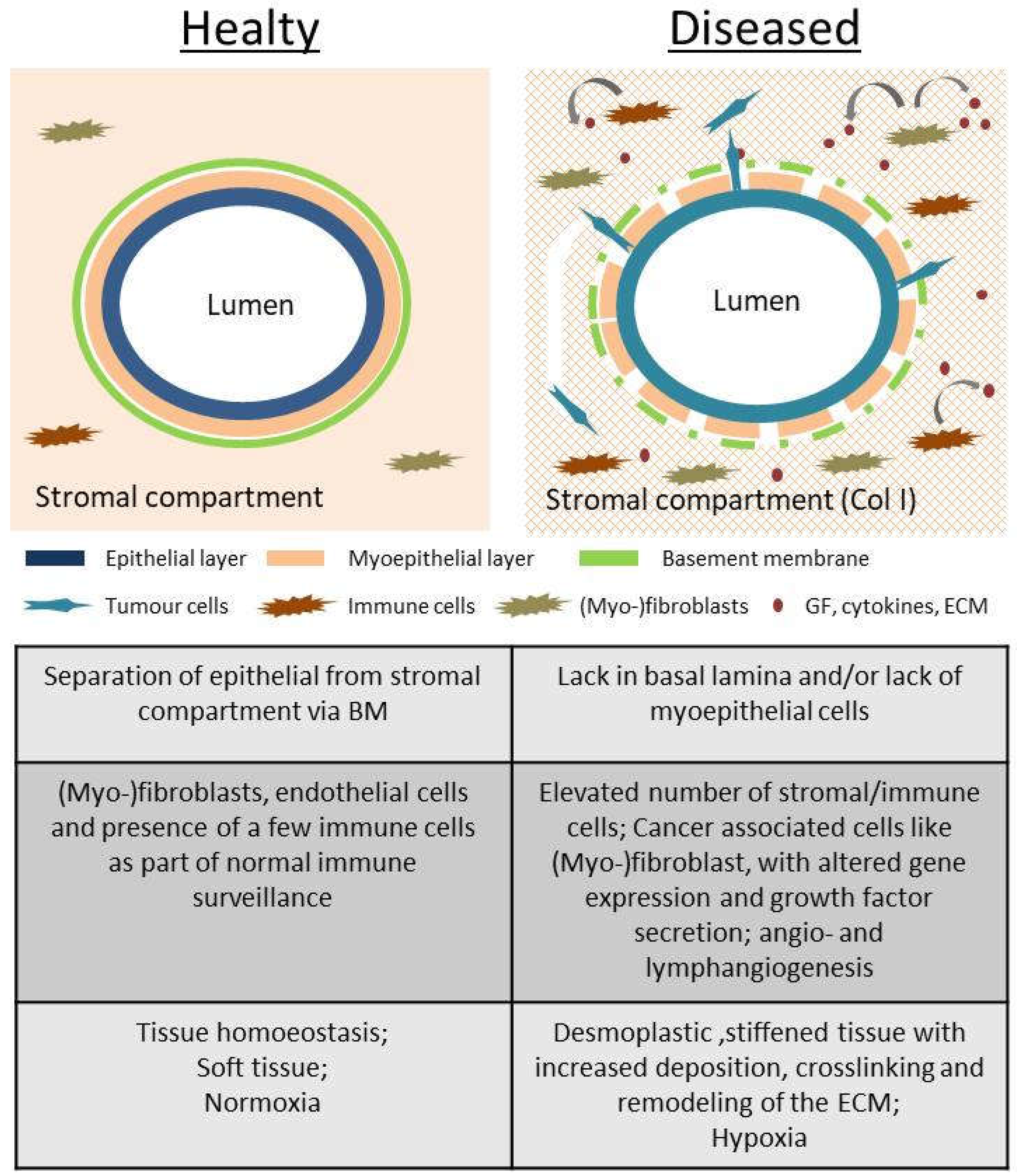
Figure 2.
Defining EMT-status of 4 breast cancer cell line. (a) Summarising and defining the EMT-status according to the available data consisting of morphological, EMT-marker and motility assessment. MCF7 and HC1954 cell lines were shown to have epithelial (E) characteristics throughout the experiments. MDA-MB-468 cells were considered an E/M-hybrid cell line. MDA-MB-231 cell line consistently demonstrated mesenchymal (M) characteristics. They are considered an EMT-positive cell line. (b) Protein levels of CDH1, VIM and GAPDH (housekeeping) of the 4 cell lines. (c) mRNA expression of CDH1 and VIM gene are shown as 2-∆Ct normalized to GAPDH mRNA expression. (d) Ibidi® migration assay performed on 4 BC cell lines. Migration is analysed by comparing the percentage of gap closure [%] over time (h). Area/timepoint is calculated based on three marked spots within each gap (n = 2). Error bars represent SD.
Figure 2.
Defining EMT-status of 4 breast cancer cell line. (a) Summarising and defining the EMT-status according to the available data consisting of morphological, EMT-marker and motility assessment. MCF7 and HC1954 cell lines were shown to have epithelial (E) characteristics throughout the experiments. MDA-MB-468 cells were considered an E/M-hybrid cell line. MDA-MB-231 cell line consistently demonstrated mesenchymal (M) characteristics. They are considered an EMT-positive cell line. (b) Protein levels of CDH1, VIM and GAPDH (housekeeping) of the 4 cell lines. (c) mRNA expression of CDH1 and VIM gene are shown as 2-∆Ct normalized to GAPDH mRNA expression. (d) Ibidi® migration assay performed on 4 BC cell lines. Migration is analysed by comparing the percentage of gap closure [%] over time (h). Area/timepoint is calculated based on three marked spots within each gap (n = 2). Error bars represent SD.
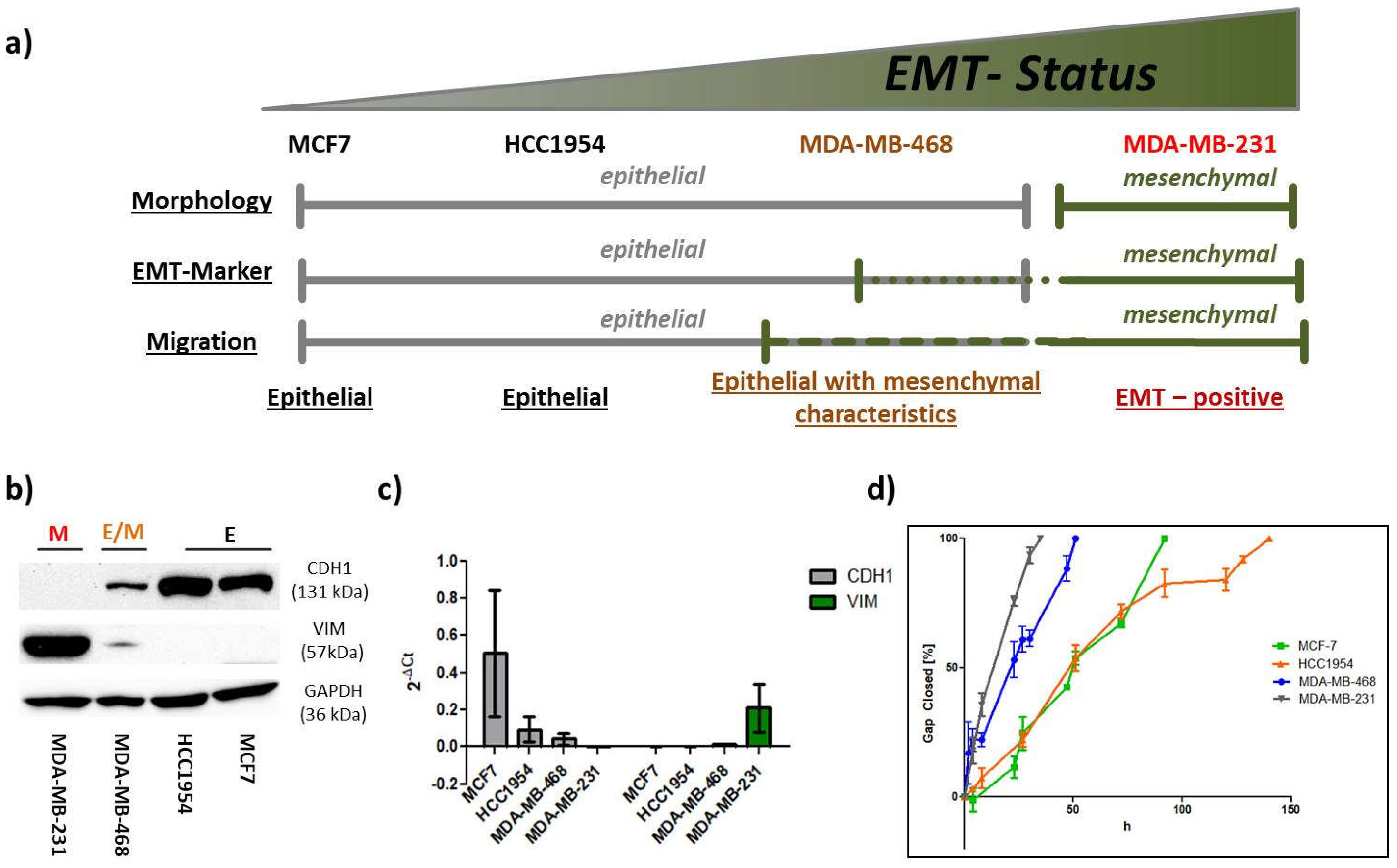
Figure 3.
Morphological analysis of the 4 BC cell lines. (a) Confocal images of fixed cells recorded with a 63x objective. Nuclei are shown in blue (DAPI) and the actin cytoskeleton in red (TRITC). Cellular clustering and cuboidal shapes are typical epithelial features as shown for MCF7, HCC1954 and MDA-MB-468. F-Actin was predominantly organized in cortical bundles tightly associated with cell–cell adhesions. Mesenchymal MDA-MB-231 cells exhibited spindle-shaped morphologies and failed to form cellular islets. (b) Image analysis via Fiji software is presented as Whiskers plot with 5-95 percentiles. Upper panel shows nuclear circularity CN of the 4 cell lines (nnuclei = 33 - 74). Nuclear circularities between 0.8 - 0.9 were assigned to epithelial cells (dotted lines). The lower panel shows cellular aspect ratio AR of the 4 cell lines (ncells = 25). AR-values from 0.6 to 0.8 were attributed to epithelial cell shapes as indicated by the dotted lines. One-way ANOVA with Bonferroni multiple comparison test was performed in GraphPad Prism software (Graph Pad Software, La Jolla, CA, USA) to calculate P-values at 95% confidence interval.
Figure 3.
Morphological analysis of the 4 BC cell lines. (a) Confocal images of fixed cells recorded with a 63x objective. Nuclei are shown in blue (DAPI) and the actin cytoskeleton in red (TRITC). Cellular clustering and cuboidal shapes are typical epithelial features as shown for MCF7, HCC1954 and MDA-MB-468. F-Actin was predominantly organized in cortical bundles tightly associated with cell–cell adhesions. Mesenchymal MDA-MB-231 cells exhibited spindle-shaped morphologies and failed to form cellular islets. (b) Image analysis via Fiji software is presented as Whiskers plot with 5-95 percentiles. Upper panel shows nuclear circularity CN of the 4 cell lines (nnuclei = 33 - 74). Nuclear circularities between 0.8 - 0.9 were assigned to epithelial cells (dotted lines). The lower panel shows cellular aspect ratio AR of the 4 cell lines (ncells = 25). AR-values from 0.6 to 0.8 were attributed to epithelial cell shapes as indicated by the dotted lines. One-way ANOVA with Bonferroni multiple comparison test was performed in GraphPad Prism software (Graph Pad Software, La Jolla, CA, USA) to calculate P-values at 95% confidence interval.
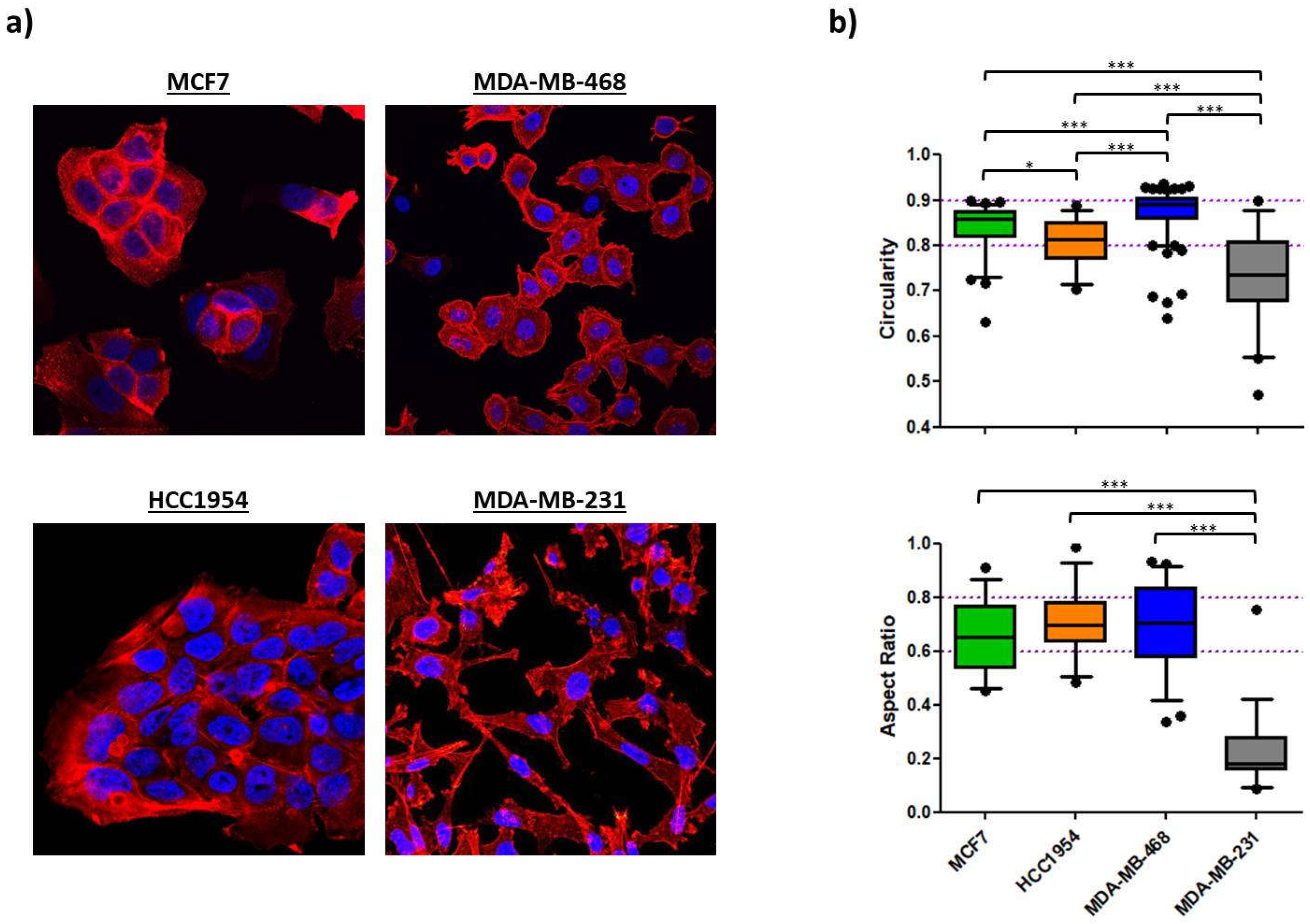
Figure 4.
Protein levels of CDH1, VIM and GAPDH (housekeeping) were assessed via Western blotting from 30 µg of the total protein extracts of MCF7, HCC1954 and MDA-MB-468 cell line as part of the EMT-induction study. Cells were subjected to multiple treatments as indicated. Numbers in the blots indicate fold-changes in protein levels normalized to untreated control cells (Blank) after 72 h incubation; (a) EMT induction study in MCF7; (b) EMT induction study in HCC1954; (c) EMT induction study in MDA-MB-468. (d) EGF stimulation in HCC1954 dependent on collagen type; (e) TGF-β1 stimulation in HCC1954 dependent on collagen type.
Figure 4.
Protein levels of CDH1, VIM and GAPDH (housekeeping) were assessed via Western blotting from 30 µg of the total protein extracts of MCF7, HCC1954 and MDA-MB-468 cell line as part of the EMT-induction study. Cells were subjected to multiple treatments as indicated. Numbers in the blots indicate fold-changes in protein levels normalized to untreated control cells (Blank) after 72 h incubation; (a) EMT induction study in MCF7; (b) EMT induction study in HCC1954; (c) EMT induction study in MDA-MB-468. (d) EGF stimulation in HCC1954 dependent on collagen type; (e) TGF-β1 stimulation in HCC1954 dependent on collagen type.
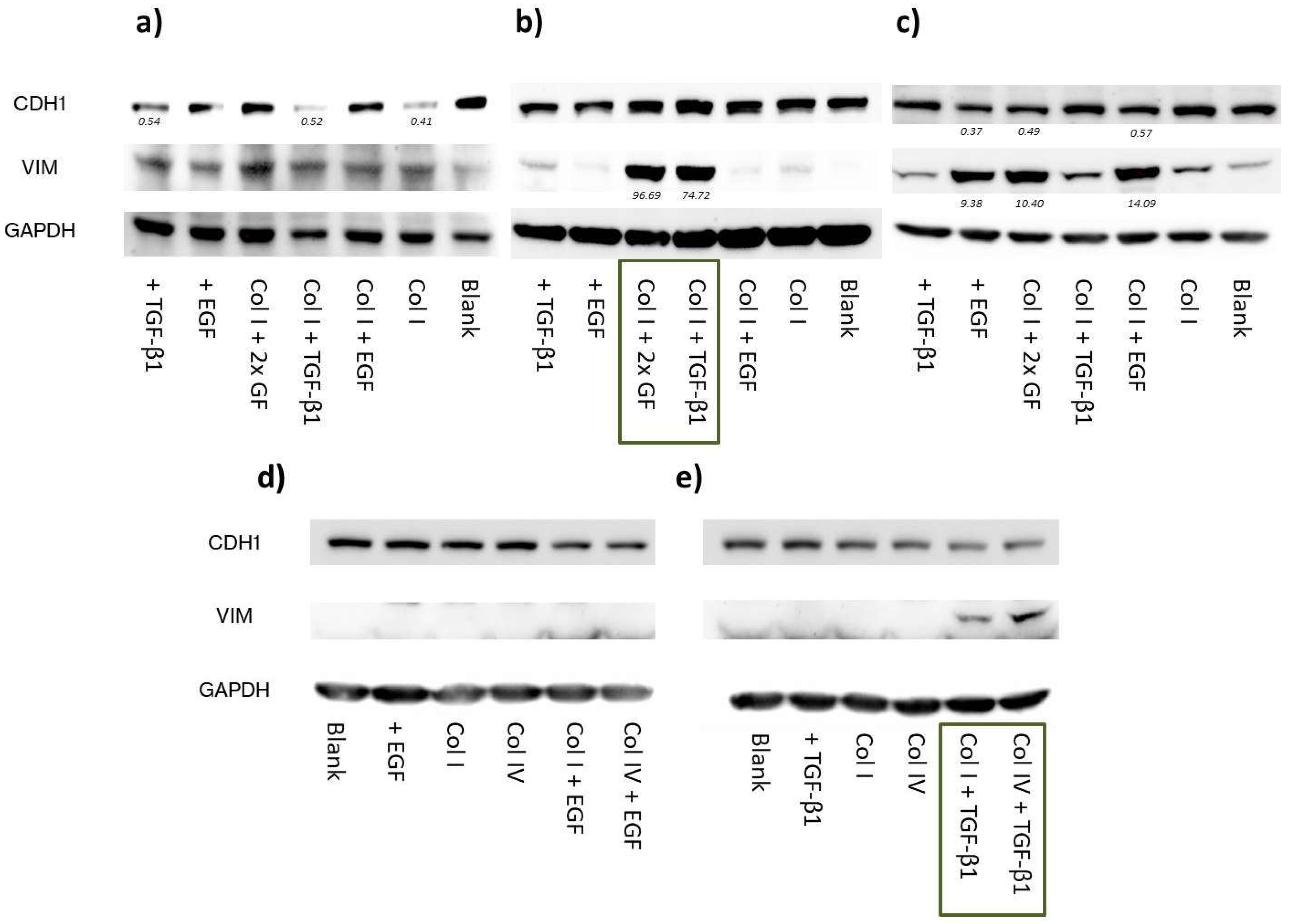
Figure 5.
Morphological analysis as part of the EMT-induction study. Confocal image analysis of fixed samples treated for 72 h. Cells were subjected to multiple treatments as indicated. Nuclear circularity CN (upper panel) and cellular aspect ratio AR (lower panel) were calculated using the Fiji software. Data is presented as Whiskers plot with 10-90 percentiles. One-way ANOVA with Dunnett's Multiple Comparison Test was performed in GraphPad Prism software to calculate P-values at 95% confidence interval; (a) Shape factor analysis of MCF7 cell treated with growth factors and/or grown on collagen demonstrated no significant changes in cellular morphologies; (b) Shape factor analysis of HCC1954 cell line showed significant alterations as part of a combinatorial treatment of TGF-β1 with collagen coatings (highlighted in blue); (c) Shape factor analysis of MDA-MB-468 cells revealed significant decrease of CN and AR for EGF stimulation (highlighted in orange); (d) Confocal images of fixed cells were recorded with a 63xobjective. Nuclei are shown in blue (DAPI) and the actin cytoskeleton in red (Phalloidin-TRITC). Cell growth of HCC1954 cells subjected to TGF-β1 treatment either grown on conventional glass coverslips or on collagen coated dishes. Combinatorial treatment resulted in restructuring of the cytoskeleton and stress fiber formation as part of the EMT program; (e) Confocal images of fixed cells were recorded with a 40x objective. MDA-MB-468 cell growth comparison between untreated cells (Blank) and EGF-treated cells. EGF stimulation resulted in loosened cell-cell junctions. Cells disseminated from epithelial clusters and exhibited more elongated shapes as compared to untreated cells.
Figure 5.
Morphological analysis as part of the EMT-induction study. Confocal image analysis of fixed samples treated for 72 h. Cells were subjected to multiple treatments as indicated. Nuclear circularity CN (upper panel) and cellular aspect ratio AR (lower panel) were calculated using the Fiji software. Data is presented as Whiskers plot with 10-90 percentiles. One-way ANOVA with Dunnett's Multiple Comparison Test was performed in GraphPad Prism software to calculate P-values at 95% confidence interval; (a) Shape factor analysis of MCF7 cell treated with growth factors and/or grown on collagen demonstrated no significant changes in cellular morphologies; (b) Shape factor analysis of HCC1954 cell line showed significant alterations as part of a combinatorial treatment of TGF-β1 with collagen coatings (highlighted in blue); (c) Shape factor analysis of MDA-MB-468 cells revealed significant decrease of CN and AR for EGF stimulation (highlighted in orange); (d) Confocal images of fixed cells were recorded with a 63xobjective. Nuclei are shown in blue (DAPI) and the actin cytoskeleton in red (Phalloidin-TRITC). Cell growth of HCC1954 cells subjected to TGF-β1 treatment either grown on conventional glass coverslips or on collagen coated dishes. Combinatorial treatment resulted in restructuring of the cytoskeleton and stress fiber formation as part of the EMT program; (e) Confocal images of fixed cells were recorded with a 40x objective. MDA-MB-468 cell growth comparison between untreated cells (Blank) and EGF-treated cells. EGF stimulation resulted in loosened cell-cell junctions. Cells disseminated from epithelial clusters and exhibited more elongated shapes as compared to untreated cells.
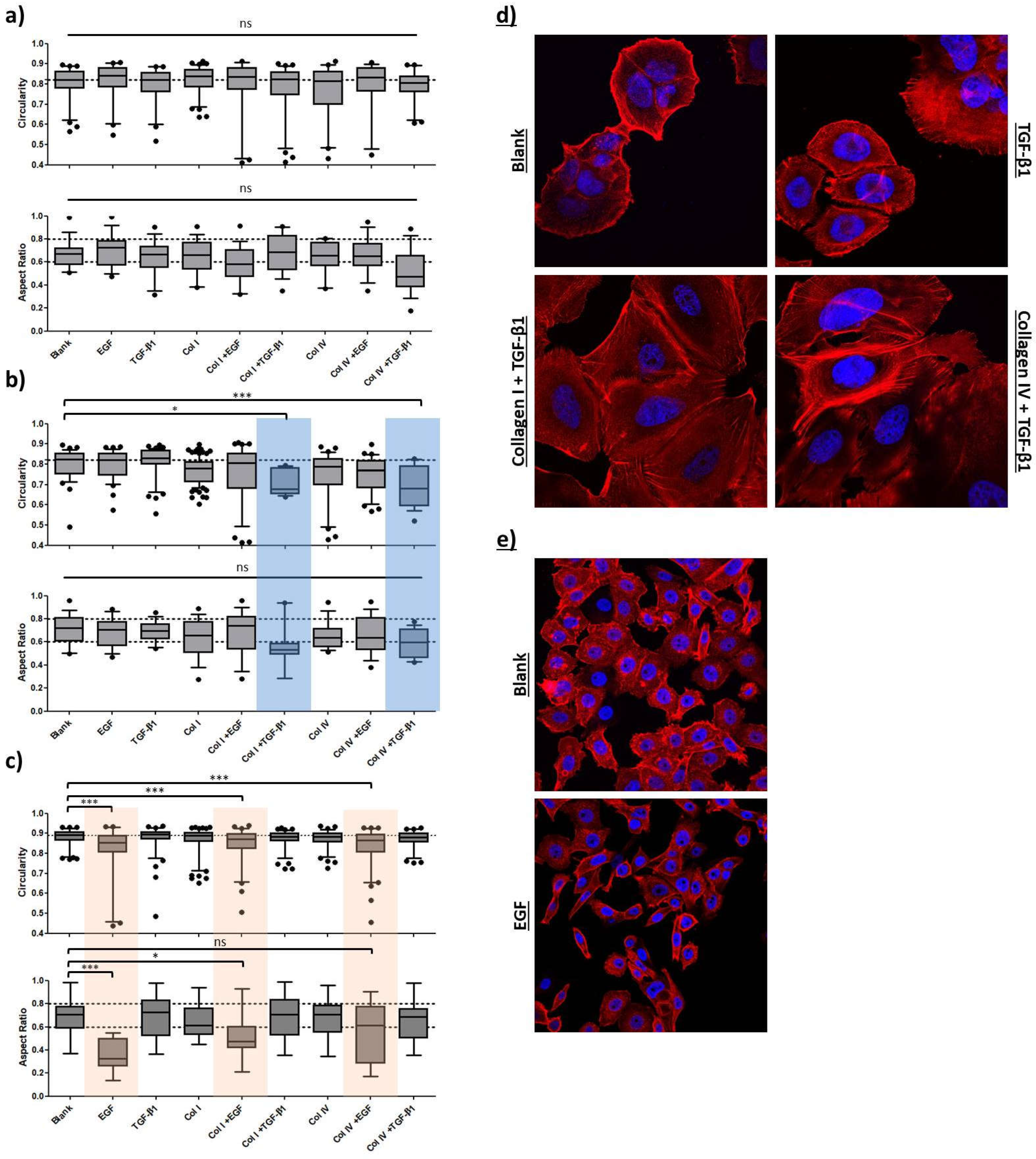
Figure 6.
Influence of EMT induction on migratory properties of BC cell lines. Ibidi® migration assay of MCF7 (a), HCC1954 (b) and MDA-MB-468 (c) are shown after 48 h incubation. Cells were subjected to multiple treatments as indicated. Migration is analyzed by comparing the percentage of gap closure [%] over time [h]. Dotted lines depict time point of 100% gap closure of faster moving cells highlighting treatments that influence cellular migration.
Figure 6.
Influence of EMT induction on migratory properties of BC cell lines. Ibidi® migration assay of MCF7 (a), HCC1954 (b) and MDA-MB-468 (c) are shown after 48 h incubation. Cells were subjected to multiple treatments as indicated. Migration is analyzed by comparing the percentage of gap closure [%] over time [h]. Dotted lines depict time point of 100% gap closure of faster moving cells highlighting treatments that influence cellular migration.
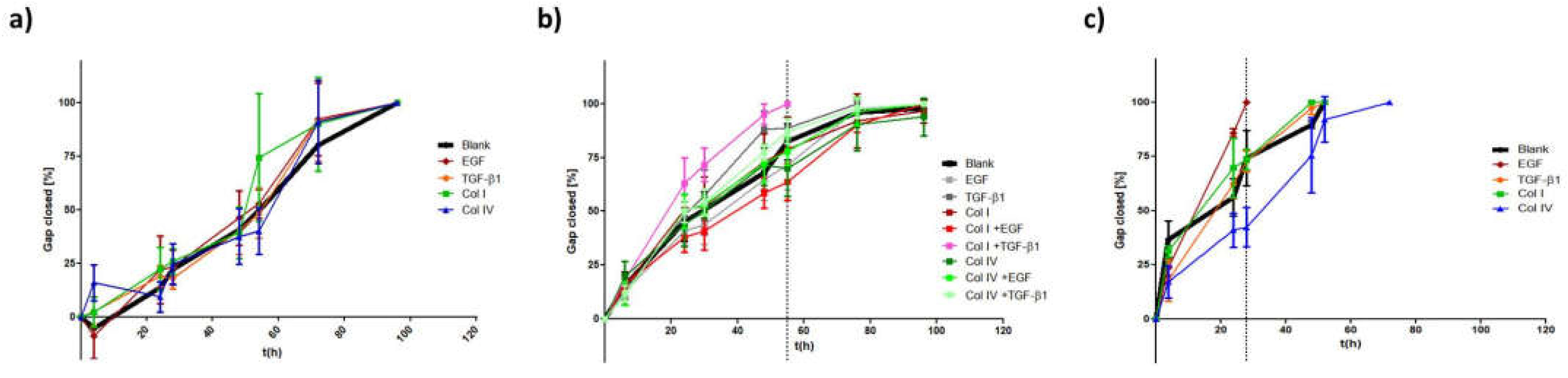
Figure 7.
(a) EMT-phenotyping model for breast cancer. Summary of the data obtained from the cell lines merging EMT-marker protein levels, morphological and migration analysis; (b) miR200c expression levels of the four cell lines; (c) Applying miR-200c induction in MDA-MB-231 cells to test the EMT-model. Data was extracted after 48 h and 72 h of DOX-dependent miR200c induction and compared to untreated (Blank) cells; Mean values of nuclear circularity CN, cellular aspect ratio AR, apparent velocity νa ([%] gap-closure per hour) and E/M- or M/E-ratio (= mesenchymal/epithelial-ratio on the protein level (VIM/CDH1)) were plotted in a multiple variable bubble plot. Results of the EMT induction study were included in (a). M/E-ratio (E/M) of 1 indicates untreated control cells of each cell line and the respective controls are further indicated by an arrow. With increasing size of the bubbles, cells are approaching a protein set-up phenotypic for mesenchymal (a) or epithelial (c) cells. Colors of the bubbles indicate the migratory behavior with yellow representing the fastest moving cells. Grey bubbles were not attributed an apparent velocity. Ellipsoid, colored accentuations depict cell line-specific phenotypic changes with green representing MCF7 cells, purple HCC1954 cells, orange MDA-MB-468 cell line and red untreated sample of MDA-MB-231 cell line. The black dotted line shows the linear regression diagonal (AR/CN) of the control cells (y = 2.935x – 1.787; R2 = 0.62)
Figure 7.
(a) EMT-phenotyping model for breast cancer. Summary of the data obtained from the cell lines merging EMT-marker protein levels, morphological and migration analysis; (b) miR200c expression levels of the four cell lines; (c) Applying miR-200c induction in MDA-MB-231 cells to test the EMT-model. Data was extracted after 48 h and 72 h of DOX-dependent miR200c induction and compared to untreated (Blank) cells; Mean values of nuclear circularity CN, cellular aspect ratio AR, apparent velocity νa ([%] gap-closure per hour) and E/M- or M/E-ratio (= mesenchymal/epithelial-ratio on the protein level (VIM/CDH1)) were plotted in a multiple variable bubble plot. Results of the EMT induction study were included in (a). M/E-ratio (E/M) of 1 indicates untreated control cells of each cell line and the respective controls are further indicated by an arrow. With increasing size of the bubbles, cells are approaching a protein set-up phenotypic for mesenchymal (a) or epithelial (c) cells. Colors of the bubbles indicate the migratory behavior with yellow representing the fastest moving cells. Grey bubbles were not attributed an apparent velocity. Ellipsoid, colored accentuations depict cell line-specific phenotypic changes with green representing MCF7 cells, purple HCC1954 cells, orange MDA-MB-468 cell line and red untreated sample of MDA-MB-231 cell line. The black dotted line shows the linear regression diagonal (AR/CN) of the control cells (y = 2.935x – 1.787; R2 = 0.62)
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



