Submitted:
03 March 2023
Posted:
07 March 2023
You are already at the latest version
Abstract
Polyploidy induction is recognized as one of the major evolutionary processes presenting remark-able morphological, physiological, and genetic variations in plants. Soybean (Glycine max L.), also known as soja bean or soya bean, is an annual leguminous crop of the pea family (Fabaceae) that shares a paleopolypoidy history, dating back to approximately 56.5 million years ago with other leguminous crops such as cowpea and other Glycine specific polyploids. This crop has been documented as one of the polyploid complex species among legumes whose gene evolution and result-ant adaptive growth characteristics, following induced polyploidization has not been fully explored. Furthermore, there are no successfully established in vivo or in vitro based polyploidy in-duction protocols that have been reported so far, particularly, with the intention to develop mutant plants showing strong resistance to abiotic salinity stress. This review, therefore, describes the role of synthetic polyploid plant production in soybean for the mitigation against high soil salt stress levels, and how this evolving approach could be used to further enhance nutritional, pharmaceutical and economic industrial value of soybeans, including addressing challenges involved during the polyploidization process.
Keywords:
Biochemical traits
; Colchicine
; Glycine max
; Morphology
; Oryzalin
; Polyploidy
; Salinity stress
; Soybean
1. Introduction
Polyploidy induction is recognized as one of the major evolutionary processes that presents a variety of remarkable morphological, physiological, and genetic variations in plants [1,2,3]. In Fabaceous crops such as soybean, also known as soja bean or soya bean, Kim et al. [1] reported a paleopolypoidy history that dates back to approximately 56.5 million years ago, which is similar to that of common bean and other Glycine specific polyploids. Such history on evolution and diversification of plant species also drove soybean domestication, and the improvement of its agronomic, as well as nutritional characteristics [4]. Both soybean’s evolutionary information and domestication made this crop to become one of the current major sources of dietary carbohydrates, fibre, minerals, oil, proteins and vitamins for human and animal consumption [5]. Generally, plant polyploidization is comprised of the multiplication of a complete set of chromosomes that co-exist within a single cell nucleus. Progenies, subsequently leading to the formation of diversified species’ mutant varieties [6], can stably inherit these newly formed genomic features. In leguminous plants, likewise in non-leguminous species like rice, wheat, tomato, cutleaf groundcherry etc., polyploidization has been widely tested using chemicals such as colchicine [7], epoxomicin [8], sodium azide [9], oryzalin [10], nitroxide [11] and ethyl methanesulfonate [12], some of which are illustrated below in Figure 1. These chemical compounds cause numerous genetic mutations that result in significant changes on the plant’s nuclear and proteome systems. Although, the induced mutations can cause genetic defects, and undesirable deformities that could be easily identified through phenotypic evaluations, changes incurred also serve as alternative means of achieving genetic variations, which contribute to the much needed species diversity. Mastuti et al. [13] reported variations in growth response characteristics of the different types of explants treated with 0.1% colchicine solution for 0, 24, 48 and 72 hours in Physalis angulate. In specifically targeting the proteasome using epoxomicin, van der Hoorn et al. [14] demonstrated unexpected changes in Arabidopsis’ protein activities during defense and stress response. Even though, epoxomicin has been widely reported mainly as an antitumoral natural product inhibiting primarily chymotrypsin-like activity [15]. Mutagenesis studies in Saccharomyces cerevisiae and other organisms, in combination with proteomics, revealed that this chemical indirectly induces irreversible mutations.
This chemical influences proteolytic systems in the cytosol and nucleus to alter cell growth and gene expression regulations in living cells [16,17,18,19]. However, nitroxide regulates plant morphogenesis and development through post-translational protein modifications, calcium pump, hormonal regulations, and reduction of reactive oxygen species (ROS) activity [18]. Meanwhile, other studies show that oryzalin induced polyploidy in plants by disrupting microtubule formations, similar to the preventative anisotropic growth effects of colchicine [20]. Interestingly, for proper plant growth, vegetative development and reproduction under high salt concentrations, mutagenic chemicals could also be used to enhance morpho-physiological and biochemical adaptive response mechanisms of plants. The ability of plants to respond and survive under salinity stress remains a priority as this abiotic condition essentially reduces cell water potential causing dehydration and ion cytotoxicity. This stress, resulting from the combinational effects of over-irrigation and poor soil drainage, disrupted soybean growth like many other plants by hindering seed germination, seedling development, flowering and fruiting [11,20,21,22,23].
This review, therefore, describes the role of synthetic polyploid plant production in soybean for the mitigation against high soil salt stress levels, and then briefly discusses how this evolving approach could be used to further enhance the nutritional, pharmaceutical and economic value of this and other leguminous crops, including addressing challenges involved during the polyploidization process under both in vivo and in vitro conditions.
2. Genetic Architecture and Response to Salinity Stress in Soybean
As a partially tetraploid, soybean forms part of the legumes belonging to the Fabales, which are comprised of more than 20,055 species, distributed within about 754 genera and nearly making up 10% of the eudicots [24,25]. Legumes experienced whole genome duplication after the Cretaceous-Paleogene (K-Pg) boundary (KPB) mass extension [25]. Especially, with allopolyploidy events giving rise to the ancestral species, Glycine soja, proposed to be a wild progenitor of Glycine max L. (soybean) [26]. Undoubtedly, species found within the Eurosid 1 angiosperm group constitute many of the most economically and agronomically important leguminous plants (Table 1 and Figure 2), in particular, species of the tribe Vicieae, Cicereae, Dalbergieae, Genisteae, Indigofereae and Phaseoleae [27,28]. Species from these selected families are used as food crops, directly or indirectly in the form of ripe-mature or unripe-immature pods, as well as mature and immature dry seeds. Figure 2 demonstrate completed inferences about a list of cultivated leguminous species used as grain and forage crops for human and animal consumption. Among them, soybean falls within the Phaseoleae tribe with prominent and widely domesticated species such as pigeon pea, dry bean, black gram, mung bean and cowpea as presented with their scientific names in Table 1. However, a species-comprehensive phylogenetic illustration (Figure 2) is important, capturing full species diversity and taxa relationships for most cultivated grain legumes belonging to all natural tribes; especially the Vicieae and Phaseoleae. Both tribes consist of species exhibiting similar phylogenetic characters needed to overcome food insecurity and limitations such as inefficient nodulation, resistance to biotic and abiotic stress constraints. Furthermore, the species within these tribes belong to a super clade called eudicot, which clearly demonstrated diversity in morphology, physiology, ecology and anatomical/ structural support mechanisms in response to stresses such as drought and salinity [29]. Like many of these plants, soybean evolutionarily contain inherent morpho-physiological and biochemical mechanisms that permit it to thrive under high salt stress environments. Taxonomically similar to other species within the tribe, which is traditionally regarded as the clade of unnatural taxon (Figure 2), with mainly edible species [30], this crop exhibit growth features required to cope with significant levels of abiotic stress.
Among the complex and elegant strategies to overcome abiotic salt stress, soybeans convert stress signals to alter gene expression in order to activate mechanisms of acclimation and tolerance. The first salt stress-sensing mechanism triggers a downstream response comprising multiple signal transduction pathways. These pathways involve activation of transcriptional regulators, ROS signaling and accumulation of secondary metabolites like hormones, among others [22]. This signaling in turn regulate metabolic and gene expression reprogramming that bring about cellular stability under salinity conditions. According to Zhu [31], stress signaling also controls protein expression, which serves as a critical mechanism for ion and water homeostasis in the cells. As a result, salinity stress-mediated transductions that emerge from these pathways in turn activate or suppress various networks within soybean that may either allow growth to continue under stress conditions or enable the plant to buffer its growth and reproduction until more favourable conditions are achieved. Therefore, the question of how soybean plants modify their morphological, physiological and molecular architecture in order to adapt to adverse environmental conditions such as salinity, drought and pathogens remains critical for agricultural productivity and sustainability.
3. Improvement of Salt Stress Tolerance via Polyploidy Induction in Plants
As the yield of soybean is significantly decreased by salinity stress, especially by influencing negatively on the vegetative growth stages of this crop as represented in Figure 3. Salinity was predicted to affect at least 50% of cultivated land worldwide by the year 2050 [32]. This stress causes primary negative effects by reducing cell water potential, causing dehydration and ion cytotoxicity, and secondary effects leading to ROS production, membrane destabilization, protein degradation and inhibition of photosynthesis (Figure 2). As previously emphasized, salinity inhibits growth and development of the whole plant by causing ROS accumulation, water and ionic imbalances in plant cells [21,32,33]. The Foyer-Halliwell-Asada pathway also known as the glutathione-ascorbate cycle that detoxifies ROS is essentially a one major mechanism used by plants to tolerate and survive salinity stress [34]. Ascorbate-glutathione or glutathione-ascorbate cycle eliminates ROS through the activity of ascorbate peroxidase, monodehydroascorbate reductase, dehydroascorbate reductase and glutathione reductase. These enzymes play a vital role in detoxifying ROS effects as indicated in Figure 3, especially during abiotic stress where their harmful oxygen free radical production is increasing, even surpassing the antioxidant defense capacity of plants [34,35].
Apart from ROS detoxification, plants also use compartmentalization or exportation of ions to different internal/external structures to achieve proper osmotic adjustments. In this way, plants are generally able to regulate Na+/ Cl- ion uptake, carryout long distance transport and intracellular compartmentalization in the vacuole and other specialized tissues to avoid excessive salt accumulation and cell damage by Na+/Cl- influx [36]. For example, halophytes such as Aeluropus, Astriplex, Cakila, Mesembryanthemum, Imperata, Salicornia, Suaeda and Trellungiella have greater abilities to coordinate the distribution of salts through unique structures such as bladders, salt glands and succulence, to excrete salts from their internal organs [36,37].
3.1. Vegetative Growth Characteristics for Salinity Tolerance
Induced polyploidisation as a tool for improving crop traits was discovered in 1907, and was thought to be responsible for heritability in genomic characteristics [2]. It was then later demonstrated that different ploidy levels caused different effects on the morphology and physiology of plants. As reported earlier by Alam et al. [38], tetraploid and triploid Camellia sinensis mutant plants showed more vigour and leaf hardness due to increased sizes of cortical and mesophyll cells. However, evidence of implications of synthetic polyploidy application in legumes, particularly, in soybean is very limited. A few detailed and insightful scholarly works that are available about polyploidy at all levels clearly show that this phenomenon induces changes in plant phenotypes via altered genome, also influencing interactions with abiotic and biotic environmental stress factors [39]. Forrester and Ashman [39] reported that polyploidy directly increased quantity and quality of rhizobia symbionts found in legumes such as Glycine wightii, Medicago sativa, Stylosanthes hamata and several Trifolium spp. resulting in enhanced nitrogen (N) fixation due to larger quantities of nodules, improved nodule sizes and higher root density. In cowpea, attempts have been made using this tool to improve primitive and long-existing characteristics such as small seed size, hairiness, and exined pollen grain surfaces [40].
Vicia cracca, another Fabaceae, was also used by Munzbergova [41] as a model taxon to reveal larger seed sizes and stomata, which were influenced by colchicine application during polyploidisation that was performed as described by Pavlikova et al. [42]. Even though, a larger number of morphological effects have been detected in non-leguminous but mostly ornamental plant species, such horticultural and ornamental plants with commercial value that include Rhododendron fortunei [43], Trachyspermum ammi L. [44], Taraxacum kok-saghyz [45], Lilium regale [46], Chrysanthemum carinatum L. [47], as well as other species that were not cited in this paper. In some of these species, like the Chinese privet (Ligustrum sinense Lour.), higher frequencies of pure tetraploids (30.8%) at lower seedling mortality after treatment with 0.1 or 0.2%, resulted in reduced morphological changes (compact plants, shorter, with fewer branches), but with larger leaves and higher fresh as well as dry weights. However, in view of above-mentioned reports, compared to soybean, artificially multiplying chromosome complement to influence morphological changes and responses to abiotic salinity stress remains a daunting task. This is due to the formation of aneuploidy as earlier reported by Palmer [48]. In most cases, aneuploidy leads to the lack of morphological variations among the expected polyploidy progenies, loss of pollen fertility, reduced number of seeds and plant growth retardation.
3.2. Enhanced Biochemical and Physiological Responses
Generally, polyploidisation, leading to changes in the number of chromosomes serves as a critical source of genetic variations required to improve desirable traits in many crop species. Changes in chromosome number affect nuclear genome, which in turn influences enzyme production and activity, then finally affecting the traits. The chenges in the chromosome number also leads to quantifiable effects on the physiological and biochemical chacteristics of the plant. Apart from the nuclear DNA content changes induced during polyploidisation, Luo et al. [45] reported significant fluctuations in the concentrations of inulin, sugar and resin in Taraxacum kok-saghz seedlings developed by treating seeds with 0 to 0.5% colchicine for 12 to 96 hours. Noori et al. [44] also reported two-and-a half times more oil yield in colchicine induced tetraploid than diploid control plants using 0.025 to 0.5% for 6 to 48 hours in ajowan (Trachyspermum ammi L.). In R. fortunii, polyploidised tetraploids and octoploids were found to contain significantly higher amounts of total chlorophyll with 76.37 and 117.80 mg/g, compared to their diploid counterparts with 41.19 mg/g [43]. The application of colchicine as the most common proceedure used for chromosome doubling, enhancing both biochemical and physiologicl parameters has proved especially successful in monocot species, such as barlley, maize and wheat. Allowing an increased gene scope and expression using this technique as reported by Rauf et al. [49]. The approach also reduced deleterious effects induced by genomic and epigenomic instability. Crops such as maize, barley and wheat showed increased leaf protein content, total soluble solids, sucrose and water soluble carbohydrates following induced polyploidy germplasm [49,50,51]. Although, polyploidy induction has been used to improve a number of biochemical parameters in many species, over 75% efficiency of genome duplication has been induced in over 120000 species of grasses and legumes mainly used as forage crops [50,51]. Such improvements in the performance of plants can be attributed to the biochemical and phsyiological changes induced anti-tubulin effects of acetamide, oryzalin (Figure 1), colchicine (Figure 1), amiprophos-methyl and trifluralin [49,52].
4. Ploidy Stability and Molecular Profile for Salinity Stress Resistance in Soybean
As already defined, polyploidy is a natural phenomenon responsible for reduplication of sets of chromosomes in plants species through autopolyploidy or allopolyploidy. However, as a result of genetic instability and sterility, many of these polyploid species require propagation through vegetative cloning. For instance, in banana, maximum vigour and quality of bananas are maintained generations after generations through vegetative propagation with the triploid state. As reported by Lestari et al. [53] similarities in the number of banana tillers, brix percentage, fruit peel/ mesocarp thickness and fruit length served as indicators of genetic stability, wherein genetic similarity coefficient ranged between 0.9 and 1.0. Nevertheless, under stressful conditions such as salinity stress, polyploidised plants are expected to demonstrate high adaptive value or fitness (Figure 3) than their diploid counterparts. Furthermore, in the case of triploid bananas being sterile, the resultant fruit becomes seedless and more commercially acceptable. Other polyploid species include commercial cultivars of potato and sugarcane that are vegetatively propagated to maintain their genetic stability and integrity. The pollen and ovule development of these cultivars can be ignored, even if they are normal and viable, particularly in order to keep a good genetic stock. Therefore, in soybean, like many other legumes of Fabales (Figure 2), polyploidy could be explored for the development of unique bean germplasm that would morphologically and physiologically contribute to improved yield under biotic and abiotic stress conditions. Additionally, this could also be based on the need to increase genetic diversity and improve the genetic base that may come with enhanced architectural and yield traits. Nevertheless, polyploidy is well tolerated in many crop species, with the majority of plants being descendants from polyploidized ancestors. Other long-term or short-term effects such as genomic stability, gene expression and genetic redundancy still need to be fully evaluated.
Wendel et al. [54] reported that polyploidization has both immediate as well as long-term genomic and transcriptomic consequences, which lead to different fitness, biased expression of homologs (gene duplicated by polyploidy) and loss of certain sequences from the duplicated genome. As reported by Kolar et al. [55], polyploidy lead to a unique form of genomic and phenotypic variation that influences crop species interactions with their environment. This process may result into high crop species fitness, postulated to confer resistance to abiotic constraints such as salinity stress. In Malus domestica (apple), cultivar Hanfu and Gala, induced autotetraploid plants were found to respond better to salt stress (200 mmolL-1 NaCl) than that of diploid plants [56]. Similarly, tetraploids of Carrizo citrange seedlings were reported to be more tolerant to salt stress (40 mM NaCl for 20 days) [57] and Citrus limonia Osb seedlings more adapted to nutrient deficiency for over 7 months than their diploid genotypes [58]. Although, numerous studies already provide insights into polyploidy-induced mechanism of salinity tolerance in citrus crops, better understanding and evidence of induced changes in the physiological functions, as well as gene expressions are required in leguminous crops such as soybean. Less clear and still needing investigation is the magnitude of stress tolerance conferred by polyploidized species and how potential barriers such as genetic instability or redundancy can be overcome. Although, polyploidy can be achieved in most crop species of angiosperm dicots, their potential to improve tolerance to environmental stresses still prompt more investigations, and perhaps tolerance to heat, drought and salt stress will be found in legumes, including important horticultural species such as Lycium ruthenicum [59], Asparagus officinalis L. [60], Ziziphus jujube Mill. Var. spinosa [61] and Malus prunifolia [62].
5. Potential Undesirable Ploidy Effects
Although, polyploidy remains well tolerated in many eukaryotic organisms, as well as the majority of angiosperms descended from polyploid ancestors as previously indicated [63]. The changes in genome structure may lead to immediate unintended undesirable effects on the genotype, phenotype and fitness of individual plant species. Such changes may allow beneficial evolutionary transitions that were not previously possible to occur, but with emerging heavy consequences. Polyploidization can stimulate further structural changes in the genomes, leading to disrupted normal cell growth, metabolism and regulation of the mitotic or meiotic cell cycle. This has been particularly, observed in animal cells than other living organisms, including plants [55,63]. At an ecosystem level, Weiss-Schneeweiss et al. [64] highlighted several fitness disadvantages such as stigma clogging, dispersal limitations and higher water-use efficiency of tetraploid plants. Moreover, this involves genome rearrangement that include repetitive DNA and locus silencing (nuclear dominance), interlocus recombination and complete or near-complete repeat replacement or redistribution [64] that may lead to considerable losses of chromosomes. In soybean, aneuploidy commonly occurs, which is primarily a trisomy comprising an extra complete chromosome (2n = 2x +1 = 41) observed from asynaptic and desynaptic mutant progeny [65]. This cause of meiotic mutation often lead to low fitness and poor reproductive capacity of soybean mutant polyploids.
For instance, in animals, aneuploidy or any ploidy changes are typically fatal with polyploids dying early during their fetal development [66]. However, the available data indicate that polyploidization contributes to evolutionary successes by increasing embryo survival, seed longevity and species richness of polyploids compared to non-polyploidized individuals within many genera in plants. Furthermore, polyploidy shapes the geographical range and distribution of species [67]. Taxonomically, phylogenetic data also revealed that the frequency of polyploids increased with increasing latitude or any forms of environmental extremeness, such as high temperature, drought and salinity [68]. Nevertheless, the better colonizing capabilities of polyploids especially under abovementioned stressful environmental conditions affect polyploid and non-polyploid plants differently. These effects involve polyploidy response to stress, which also include salinity stress. Moreover, polyploidy, particularly, the synthetic production of mutant plants through the use of chemicals such as colchicine still have the potential to play a significant role in the physiology and development of plants through cellular, metabolic and genetic effects [69].
6. Future Prospects and Final Considerations
Van de Peer et al. [68] reported genome duplication in plants as a phenomenon that has reached a dead-end, particularly under stable conditions. The study postulated that, under natural conditions, changes in whole genome duplication predominantly coincided with the periods of major global climatic fluctuations. For instance, these variations overlapped with the Cretaceous-Paleogene boundary. This implied that the frequency of polyploidy formation in living organisms was driven primarily by external environmental stimuli such as stress, especially at a level of cataclysmic events [70]. The fact that ploidy number show remarkable changes in response to environmental stress among plant species suggest that this can be further explored from an adaptation and mitigation point of view. Such predictable consequences of polyploidization resulting from the fusion or fission of chromosomes have been recorded over time as shown by the readily available taxonomic data across groups of ferns and angiosperms. Approximations of widespread natural polyploidization occurrences in plants is illustrated in Figure 4, together with the estimates of the proportions of polyploids found among some of the major families of angiosperms. This include the Fabaceae family comprising soybeans and other leguminous crop species. As incidences of polyploidization are mapped according to taxonomic groups, phylogenetic position, habitat and phytogeographical region. The proportion of herbaceous leguminous species of Fabales, particularly, Phaseolus (tribe of Cajan, Glycine, Phaseolus and Vigna) (Figure 3) exhibited higher basic chromosome numbers with considerable polyploidy of about 40%. This proportional percentage includes species of the subfamily Caesalpinioideae, Mimosoideae and Papilionoideae [66,71].
With genomic studies having revealed that soybean developed from a tetraploid ancestor, wherein cytogenetics and molecular analysis showed that allopolyploidization played a key role in the speciation of this crop. Furthermore, more wild perennial species of the subgenus Glycine forming tetraploid and aneutetraploid ploidy complexes were also discovered [72]. Polyploids are a prevalent source and rich reservoir of desirable agronomic genes. Therefore, both naturally occurring and synthetic polyploids could be explored to unlock these valuable genes, which has so far not been successful. As such, this aneupolyploidy, including other genetic barriers, have caused the improvement of soybean to lag behind other economically important crops such as barley, faba bean, maize, tomato, rice and wheat. Wide crosses and dual polyploidization have been explored for yield improvement in rice. Cai et al. [73] earlier reported selective breeding of two polyploid lines PMeS-1 and PMeS-2 using polyploid meiosis stability (PMeS) genes to breed japonica rice with higher rate of seed set. Compared to monocot species such as barley, maize and rice, seldomly, leguminous dicot polyploid lines of faba bean (Vicia faba L.) showed a strong interaction between resistance genes and avirulence pathogen genes conferring resistance to Uromyces viciae-fabae [74]. This study demonstrated the significant that polyploid lines may play in crops threatened by abiotic stresses and diseases. Such mutations can also be beneficially utilised to genetically improve varieties of soybeans, especially for enhancing adaptive properties of the crop plant against stresses such as drought and salinity.
5. Conclusions
The genetic base of soybean, particularly cultivars that are currently grown for both commercial and subsistence farming are extremely narrow. This makes soybean a very poor reservoir for genes conferring tolerance to abiotic and biotic stress. While the effects of these stresses are widely documented, especially the influence of salinity on soybean growth, development and yield [11,21,22,24,33]. The potential for induced polyploidy in soybean against salinity stress remains less recognized and largely unexplored. This paper demonstrate without doubt that polyploidy can be used to confer resistance to different kinds of environmental stresses, and can act as a reliable source of mutations to improve genetic variability in soybean. However, evidence is still required that may provide further insights on the mechanisms underlying the responses of polyploids to growth and stress related phenomena, which might not differ greatly between polyploidised plants and their diploid counterparts. Under natural conditions, evolutionary changes in recalcitrant legumes like soybean gradually takes place due to aneuploidy or aneutetraploidy that has restricted genetic variability and wider geographical distribution for most of the Glycine species [68]. In contrast, mutagenic chemical such as those shown in Figure 1 can be used to successfully induce the desired genomic changes under controlled environmental conditions, also, in a short space of time.
Funding
The APC was funded by the Department of Research Administration and Development of the University of Limpopo, Turfloop Campus, South Africa.
Informed Consent Statement
Not applicable.
Acknowledgments
The author would like to thank colleagues and all students in the Department of Biodiversity for their support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kim, K.D.; El-Baidouri, M.; Abernathy, B.; Iwato-Otsubo, A.; Chavarro, C.; Gonzales, M.; Libaut, M.; Grimwood, J.; Jackson, S.A. A comparative epigeonomic analysis of polyploidy-derived genes in soybean and common bean. Plant Physiol 2015, 168(4), 1433–1447. [CrossRef]
- Manzoor, A.; Ahmad, T.; Bashir, A.; Hafiz, I.A.; Silvestri, C. Studies on colchicine induced chromosome doubling for enhancement of quality traits in ornamental plants. Plants (Basel) 2019, 8(7), 194. [CrossRef]
- Juliao, S.A.; Ribeiro, C.V.; Lopes, J.M.L.; de Matos, E.M.; Reis, A.C.; Peixoto, P.H.P.; Machado, M.A.; Azevedo, A.L.S.; Grazul, R.M.; Campos, J.M.S.; Viccini, L.F. Induction of synthetic polyploids and assessment of genomic stability in Lippia alba. Front Plant Sci 2020, 11, 292. [CrossRef]
- Sedivy, E.J.; Wu, F.; Hanzawa, Y. Soybean domestication: The origin, genetic architecture and molecular bases. New Physiol 2017, 214(2), 539–553. [CrossRef]
- Oyegbami, A.; Fadairo, A.O.; Oyedokun, M.O. Women’s knowledge of the nutritional benefits and perceived constraints in soybean utilization in Oyo State, Nigeria. S. Afr J Agric Ext 2020, 48(2), 166–175.
- Kang, Z.; Xiaowu, W.; Feng, C. Plant polyplidy: Origin, evolution and its influence on crop domestication. Horti Plant J 2019, 5(6), 231–239. [CrossRef]
- Al Nebaihi, H.; Le, T.S.; Davies, N.M.; Brocks, D.R. Liquid chromatography tandem mass spectrometric analytical method for study of colchicine in rats given low doses. Processes 2021, 9(11), 2007. [CrossRef]
- Katukojvala, S.; Barlett, K.N.; Lotesta, S.D.; Williams, L.J.; Spirodiepoxides in total synthesis: Epoxomicin. J Am Chem Soc 2004, 126(47), 15348–15349. [CrossRef]
- Kantin, G.P.; Krasavin, M.Y. Microwave promoted reaction of N-(alk-1-enyl)chloroacetamides with sodium azide unexpectedly yields 1H-imidazol-5(4H)-ones. Mendocomm 2017, 27(1), 95–96. [CrossRef]
- Honma, M. An assessment of mutagenicity of chemical substances by (quantitative) structure-activity relationship. Gene Environ 2020, 42, 23. [CrossRef]
- Simontacchi, M.; Galatro, A.; Ramos-Artuso, F.; Santa-Maria, GE. Plant survival in changing environment: The role of nitric oxide in plant response to abiotic stress. Front Plant Sci 2015, 6, 977. [CrossRef]
- Espina, M.J.; Ahmed, C.M.S.; Bernardini, A.; Adeleke, E.; Yadegari, Z.; Arielli, P.; Pantalone, V.; Taheri, A. Development and phenotypic screening of ethyl methane sulfonate mutant population in soybean. Front Plant Sci 2018, 9, 394. DOI: 3389/fpls.2018.00394.
- Mastuti, R.; Munawarti, A.; Afiyanti, M. The effect of colchicine on in vitro growth of ciplukan (Physalis angulate). IOC Conf Ser: Earth Environ Sci 2022, 1097, 1–7. [CrossRef]
- Van der Hoorn, R.A.L.; Colby, T.; Nickel, S.; Richau, K.H.; Schmidt, J.; Kaiser, M. Mining the active proteome of Arabidopsis thaliana. Front Plant Sci 2021, 2, 89. [CrossRef]
- Schwarz, K.; Giul, R.; Schmidtke, G.; Kostka, S.; van der Broek, M.; Kim, K.B.; Crews, C.M.; Kraft, R.; Groettrup, M. The selective proteasome inhibitors lactacystatin and epoxomicin can be used to either up or down regulate antigen presentation at nontoxic does. J Immunol 2000, 164(12), 6147–6157. [CrossRef]
- Kisselev, A.F.; Goldberg, A.L. Proteosome inhibitor: From research tools to drug candidates. Chem Biol 2001, 8(8), 739–758.
- Levie, J.; Belvalet, H.D.; Sonon, S.; Ion, A.M.; Dumon, E.; Melser, S.; Lacombe, D.; Dupuy, J-W.; Lalou, C.; Benard, G. Ubiquitin-dependent degradation of mitochondrial protein regulates energy metabolism. Cell Rep 2018, 23, 2852–2862. [CrossRef]
- Meng, L.; Mohan, R.; Kwok, B.H.B.; Crews, C.M. Epoxomicin, a potent and selective proteasome inhibitor, exhibit in vivo anti-inflammatory activity. PNAS 1999, 96(18), 10403–10408. [CrossRef]
- Bouyer, D.; Geier, F.; Kragler, F.; Schnittger, A.; Pesch, M.; Wester, K.; Balkunde, R.; Timmer, J.; Fleck, C.; Hulskamp, M. Two-dimensional patterning by a trapping/ depletion mechanism: The role of TTGI and GL3 in Arabidopsis trichome formation. PLoS Biol 2008, 6(6), e141. [CrossRef]
- Silalahi, C.B.; Sinuraya, M.; Hanafiah, D.S.; Sipayung, R. The influence of oryzalin concentration on the plant growth of two tomato (Solanum lycopersicum L.) varieties. IOP Con Ser: Earth Environ Sci 2019, 454, 1–6. [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of plant response to salt stress. Int J Mol Sci 2021, 22, 4609. [CrossRef]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant salinity stress: Many unanswered questions remain. Front Plant Sci 2019, 10, 80. [CrossRef]
- Zahra, N.; Raza, Z.A.; Mahmood, S. Effect of salinity stress on various growth and physiological attributes of two contrasting maize genotypes. Braz Arch Biol Technol 2020, 63, e20200072.
- Morales, M.; Oakley, L.; Sartori, A.L.B.; Mogni, U=V.Y.; Atahuachi, M.; Vanni, R.O.; FRortunato, R.H.; Prado, D.E. Diversity and conservation of legumes in the Gran Chaco and biogeographical influences. PLoS One 2019, 14(8), e0220151. [CrossRef]
- Koenen, E.J.M.; Ojeda, D.I.; Bakker, F.T.; Weringa, J.J.; Kidner, C.; Hardy, O.J.; Pennington, R.T.; Herendeen, P.S.; Bruneau, A.; Hughes, C.E. The origin of the legumes is a complex paleopolyploid phylogenomic tangle closely associated with the Cretaceous-paleogene (K-Pg) mass extinction event. Systematic Biol 2020, 70(3), 508–526. [CrossRef]
- Gill, N.; Findley, S.; Walling, J.G.; Hans, C.; Ma, J.; Doyle, J.; Stacey, G.; Jackson, S.A. Molecular and chromosomal evidence for allopolyploidy in soybean. Plant Physiol 151(3), 1167–1174. [CrossRef]
- de la Estrella, M.; Forest, F.; Klitgard, B.; Lewis, G.P.; Mackinder, B.A.; de Queiroz, L.P.; Wieringa, J.J.; Bruneau, A. A new phylogeny-based tribal classification of subfamily Detarioideae, an early branching clade of florally diverse tropical arborescent legumes. Sci Rep 2018, 8, 6884. [CrossRef]
- Trytsman, M.; van Wyk, A.E.; Masemola, L. Systematics, diversity and forage value of indigenous legumes of South Africa, Lesotho and Swaziland. Afr J Biotechnol 2011, 10(63), 13773–13779. [CrossRef]
- Idowu, G.A.; Fletcher, A.J. The manufacture and characterization of Rosid angiosperms-derived biochars applied to water treatment. BioEnergy Res 2020, 13, 387–396. [CrossRef]
- Viviani, T.; Conte, L.; Cristofolini, G.; Speranza, M. Sero-systematic studies on the Phaseoleae (Leguminosae) and related tribes. Bot J Linnean Soc 2008, 105(2), 113–136. [CrossRef]
- Zhu, J-K. Abiotic stress signaling and responses in plants. Cell 2016, 167(2), 313–324. [CrossRef]
- Pi, E.; Qu, L.; Hu, J.; Huang, Y.; Qiu, L.; Lu, H.; Jiang, B.; Liu, C.; Peng, T.; Zhao, Y.; Wang, H.; Tsai, S-H.; Ngai, S.; Du, L. Mechanism of soybean roots’ tolerance to salinity revealed by proteomic and phosphoproteomic comparison between two cultivars. Mol Cell Proteomics 2016, 15(1), 266–288. [CrossRef]
- Cai, X.; Jia, B.; Sun, X. Insights into the regulation of wild soybean tolerance to salt-alkaline stress. Front Plant Sci 2022, 13, 1002302. [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidant 2019, 8(9), 384. [CrossRef]
- Lopez-Huertas, E.; Palma, J.M. Changes in glutathione, ascorbate, and antioxidant enzymes during olive fruit ripening. J Agric Food Chem 2020, 68(44), 12221–12228. [CrossRef]
- Peng, Z.; He, S.; Sun, J.; Pan, Z.; Gong, W.; Lu, Y.; Du, X. Na+ compartmentalization related to salinity stress tolerance in upland cotton (Gossypium hirsutum) seedlings. Sci Rep 2016, 6, 34548. [CrossRef]
- Mishra, A.; Tanna, B. Halophytes: Potential resources for salt stress tolerance genes and promoters. Front Plant Sci 2017, 8, 829. [CrossRef]
- Alam, H.; Razaq, M.; Salahuddin. Induced polyploidy as a tool for increasing tea (Camellia sinensis L.) production. J Northest Agric Univ 2015, 22(3), 43–47. [CrossRef]
- Forrester, N.J.; Ashman, T-L. The direct effects of plant polyploidy on the legume-rhizobia mutualism. Ann Bot 2028, 121(2), 209–220. [CrossRef]
- Boukar, O.; Abberton, M.; Oyatomi, O.; Togola, A.; Tripathi, L.; Fatokum, C. Introgression breeding in cowpea [Vigna unguiculata (L.) Walp.]. Front Plant Sci 2020, 11, 567425. [CrossRef]
- Munzbergova Z. Colchicine application significantly affect plant performance in the second generation of synthetic polyploids and its effects vary between populations. Ann Bot 2017, 120, 329–339. [CrossRef]
- Pavlikova, Z.; Pastova, L.; Munzbergova, Z. Synthetic polyploids in Vicia cracca: Methodology, effects on plant performance and aneuploidy. Plant Syst Evol 2017, 303, 827–839. [CrossRef]
- Mo, L.; Chen, J.; Lou, X.; Xu, Q.; Dong, R.; Tong, Z.; Huang, H.; Lin, E. Colchicine-induced polyploidization in Rhododendron fortunei Lindl. Plant 2020, 9, 424. [CrossRef]
- Noori, S.A.S.; Norouzi, M.; Karimzadeh, G.; Shirkool, K.; Niazian, M. Effect of colchicine-induced polyploidy on morphological characteristics and essential oil composition of ajowan (Trachyspermum ammi L.). Plant Cell Tiss Org Cult 2017, 130, 534–551. [CrossRef]
- Luo, Z.; Iaffaldano, B.J.; Cornish, K. Colchicine-induced polyploidy has the potential to improve rubber yield in Taraxacum kok-saghyz. Ind Crops Prod 2018, 112, 75–81. [CrossRef]
- Jeloudar, N.I.; Chamani, E.; Shokouhian, A-a.; Zakaria, R.A. Induction and identification of polyploidy by colchicine treatment in Lilium regale. Cytologia 2019, 84(3), 271–276. [CrossRef]
- Kushwah, K.S.; Verma, R.C.; Patel, S.; Jain, N.K. Colchicine induced polyploidy in Chrysanthemum carinatum L. Phylogenetics Evol Biol 2018, 6, 1. [CrossRef]
- Palmar, R.G. Aneuploids in the soybean, Glycine max. Genome 2011, 16(2), 441–447. [CrossRef]
- Rauf, S.; Ortiz, R.; Malinowski, D.P.; Clarindo, W.R.; Kainat, W.; Schehzad, M.; Waheed, U.; Hassan, S.W. Induced polyploidy: A tool for forage species improvement. Agric 2021, 11(3), 210. [CrossRef]
- Debkeviciene, G.; Kemesyte, V.; Statkeviciute, G.; Lemeziene, N.; Brazauskas, G. Authopolyploids in fodder grass breeding: Induction and field performance. Span J Agric Res 2017, 15(4), e0706. [CrossRef]
- Innes, L.A.; Denton, M.D.; Dundas, I.S.; Pech, D.M.; Humphries, A.W. The effect of ploidy number on vigor, productivity, and potential adaptation to climate change in annual Medicago species. Crop Sci 2021, 61(1), 89–103. [CrossRef]
- Venial, L.R.; Mendonca, M.A.C.; Amaral-Silva, P.M.; Canal, G.B.; Passos, A.B.R.J.; Ferreira, A.; Soures, T.C.B.; Clarindo, W.R. Autotetraploid Coffea canephora and auto-alloctapleid Coffea arabica from in vitro chromosome set doubling: New germplasm for Coffea. Front Plant Sci 2020, 11, 154. [CrossRef]
- Lestari, P.; Roostika, I.; Nugroho, K.; Edison, H.S.; Rijzaani, H.; Mastar, M. Genetic stability of banana plant regenerated from floral axis organogenesis assessed by newly developed SSR markers. AGRIVTA J Agric Sci 2019, 41(2): 302–315. [CrossRef]
- Wendel, J.F.; Lisch, D.; Hu, G.; Manson, A.S. The long and short of doubling down: Polyploidy, epigenetics and the temporal dynamics of genome fractionation. Curr Opin Gnetic Dev 2018, 48: 1–7. [CrossRef]
- Kolar, F.; Certner, M.; Suda, J.; Schonswetter, P.; Husband, B.C. Mixed ploidy species: Progress and opportunities in polyploid research. Trends Plant Sci 2017; 22(12): 1041–1055. [CrossRef]
- Xue, H.; Zhang, F.; Zhang, Z-H.; Fu, J-F.; Wang, F.; Zhang, B.; Ma, Y. Differences in salt tolerance between diploid and autotetraploid apple seedlings exposed to salt stress. Sci Hort 2015, 190: 24–30. [CrossRef]
- Ruiz, M.; Quimones, A.; Martinez-Cuenca, M.R.; Aleza, P.; Morillon, R.; Navarro, L.; Primo-Millo, E.; Martinez-Alcantara, B. Tetraploidy enhances the ability to exclude chloride from leaves in Carrizo citrange seedlings. J. Plant Physiol 2016, 205: 1–10. [CrossRef]
- Oustric, J.; Quilichini, Y.; Morillon, R.; Herbette, S.; Luro, F.; Giannettini, J.; Berti, L.; Santini, J. Tetraploid Citrus seedlings subjected to long-term nutrient deficiency are less affected at the ultrastructural, physiological and biochemical levels than diploid ones. Plant Physiol Biochem 2019, 135: 375–384. [CrossRef]
- Rao, S.; Tian, Y.; Xia, X.; Li, Y.; Chen, J. Chromosome doubling mediates superior drought tolerance in Lycium ruthenicum via abscisic acid signaling. Hort Res 2020, 7: 40. [CrossRef]
- Chen, H.; Lu, Z.; Wang, J.; Chen, T.; Gao, J.; Zheng, J.; Zhang, J.; Zhang, S.; Xi, J.; Huang, X.; Guo, A.; Yi, K. Induction of new tetraploid genotypes and heat tolerance assessment in Asparagus officinalis L. Sci Hort 2020, 264: 109168. [CrossRef]
- Li, M.; Zhang, C.; Hou, L.; Yang, W.; Liu, S.; Pang, X.; Li, Y. Multiple responses contribute to the enhanced drought tolerance of the autotetraploid Ziziphus jujube Mill. Var. spinosa. Cell Biosci 2021, 11: 119. [CrossRef]
- Jin, Y.; Zhao, Y.; Ai, S.; Chen, X.; Liu, X.; Wang, H.; Han, Y.; Ma, F.; Li, C. Induction of polyploid Malus prunifolia and analysis of its salt tolerance. Tree Physiol 2022, 42(10): 2100–2115. [CrossRef]
- Otto, S.P. The evolutionary consequences of polyploidy. Cell 2007, 131: 452–462. [CrossRef]
- Weiss-Schneeweiss, H.; Emadzade, K.; Jang, T.S.; Schneeweiss, G.M. Evolutionary consequences, constraints and potential of polyplody in plants. Cytogenet Genome Res 2013, 140: 137–150. [CrossRef]
- Palmer, R.G.; Sandhu, D.; Curran, K.; Bhattacharyya, M.K. Molecular mapping of 36 soybean male-sterile, female-sterile mutants. Theor Appl Genet 2008, 117: 711–719. [CrossRef]
- Fox, D.T.; Soltis, D.E.; Solitis, P.S.; Ashman, T-L.; Van de Peer, Y. Polyploidy: A biological force from cells to ecosystems. Trend Cell Biol 2020, 30(9): 688–694. [CrossRef]
- Spoelhof, J.P.; Soltis, D.E.; Soltis, P.S. Habitat shape affects polyploid establishment in a spatial, stochastic model. Front Plant Sci 2020, 11: 592356. [CrossRef]
- Van de Peer, Y. Ashman, T-L.; Soltis, P.S.; Soltis, D.E. Polyploidy: An evolutionary and ecological force in stressful times. Plant Cell 2021, 33: 11–26. [CrossRef]
- Scholes, D.R.; Paige, K.N. Plasticity in ploidy: A generalized response to stress. Trend Plant Sci 2015; 20(3): 165–175. [CrossRef]
- Tossi, V.; Tosar, L.J.M.; Laino, L.E.; Lannicelli, J.; Regalado, J.J.; Escandon, A.S.; Baroli, I.; Causin, H.F.; Pitta-Alvarez, S.I. Impact of polyploidy on plant tolerance to abiotic and biotic stress. Front Plant Sci 2022, 13: 869423. [CrossRef]
- Khatoon, S.; Ali, S.I. Chromosome numbers and polyploidy in the legumes of Pakistan. Pak J Bot 2006, 38(4): 935–945.
- Singh, R.J.; Chung, G.H. Cytogenetics of soybean: Progress and perspectives. Nucleus 2007, 50(3): 403–425.
- Cai, D.T.; Chen, J.G.; Chen, D.L.; Dai, B.C.; Zhang, W.; Song, Z.J.; Yan, Z.F.; Du, C.D.; Tang, Z.Q.; He, Y.C.; Zhang, D.S.; He, G.C.; Zhu, Y.G. The breeding of two polyploid rice lines with the characteristics of polyploid meiosis stability. Sci China C Life Sci 2007, 50(3): 356–366. [CrossRef]
- Ijaz, U.; Adhikari, K.N.; Stoddard, F.L.; Trethwan, R.M. Rust resistance in faba bean (Vicia faba L.): Status and strategies form improvement. Aust Plant Pathol 2018, 47(1): 71–81. [CrossRef]
Figure 1.
Chemical structure depictions of the most commonly used compounds for mutagenic crop improvement against biotic and abiotic stresses. Oryzalin with molecular weight (MW) of 346.36 g/mol, colchicine with MW of 399.437 g/mol, sodium azide with MW of 65.0099 g/mol, hydrogen fluoride with MW of 20.0064 g/mol, ethyl methanesulfonate (Ethyl-MS) with MW of 124.16 g/mol, methyl methanesulfonate (methyl-MS) with MW of 110.13 g/mol, N-Methyl-N-Nitrosourea with MW of 103.08 g/mol and hydroxylamine with MW of 33.03 mg/mol. .
Figure 1.
Chemical structure depictions of the most commonly used compounds for mutagenic crop improvement against biotic and abiotic stresses. Oryzalin with molecular weight (MW) of 346.36 g/mol, colchicine with MW of 399.437 g/mol, sodium azide with MW of 65.0099 g/mol, hydrogen fluoride with MW of 20.0064 g/mol, ethyl methanesulfonate (Ethyl-MS) with MW of 124.16 g/mol, methyl methanesulfonate (methyl-MS) with MW of 110.13 g/mol, N-Methyl-N-Nitrosourea with MW of 103.08 g/mol and hydroxylamine with MW of 33.03 mg/mol. .
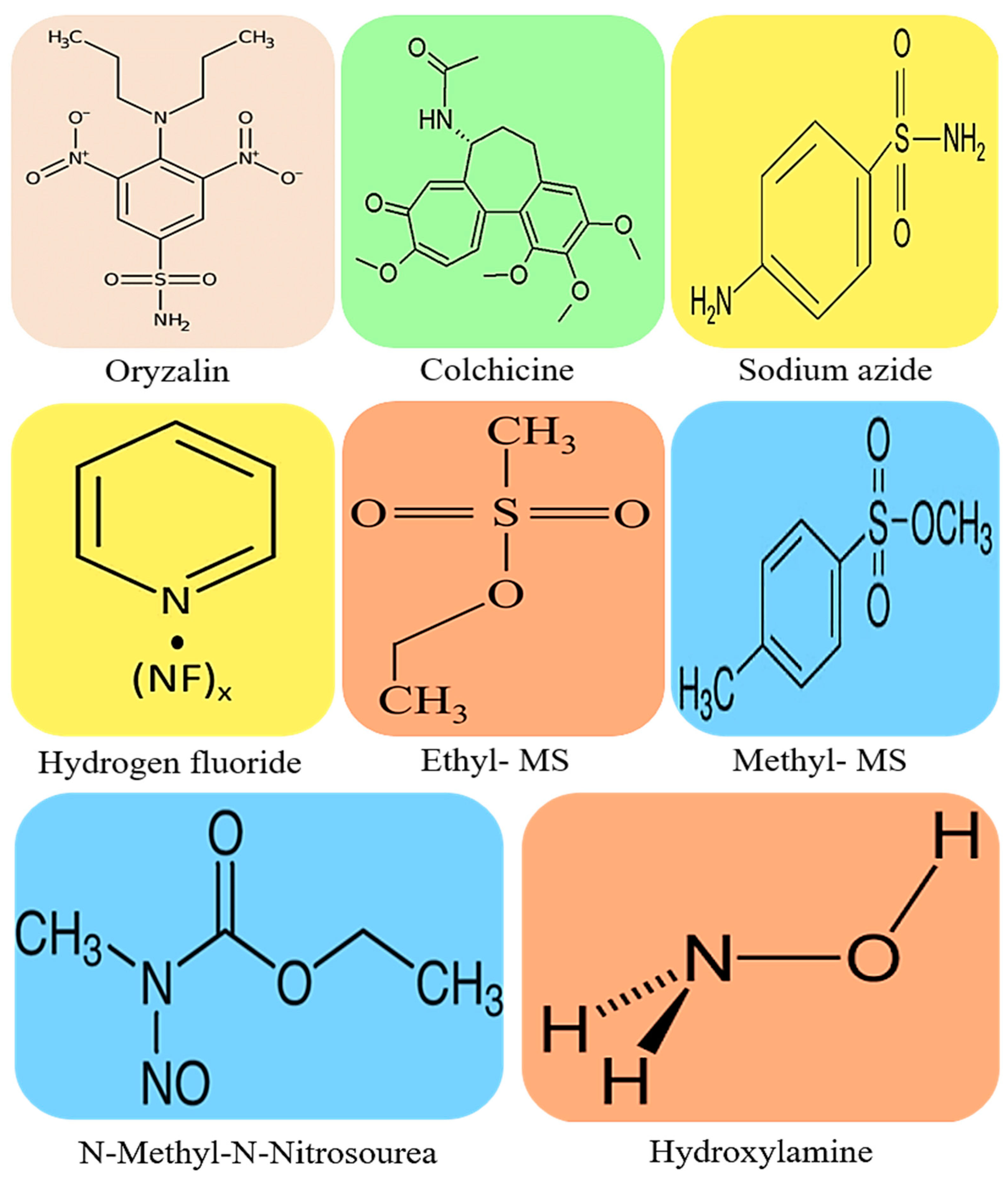
Figure 2.
A summary of a taxonomic classification scheme highlighting the position of soybean (Glycine max L.) and other selected grain leguminous crops, as well as their botanical names (genus and species).
Figure 2.
A summary of a taxonomic classification scheme highlighting the position of soybean (Glycine max L.) and other selected grain leguminous crops, as well as their botanical names (genus and species).
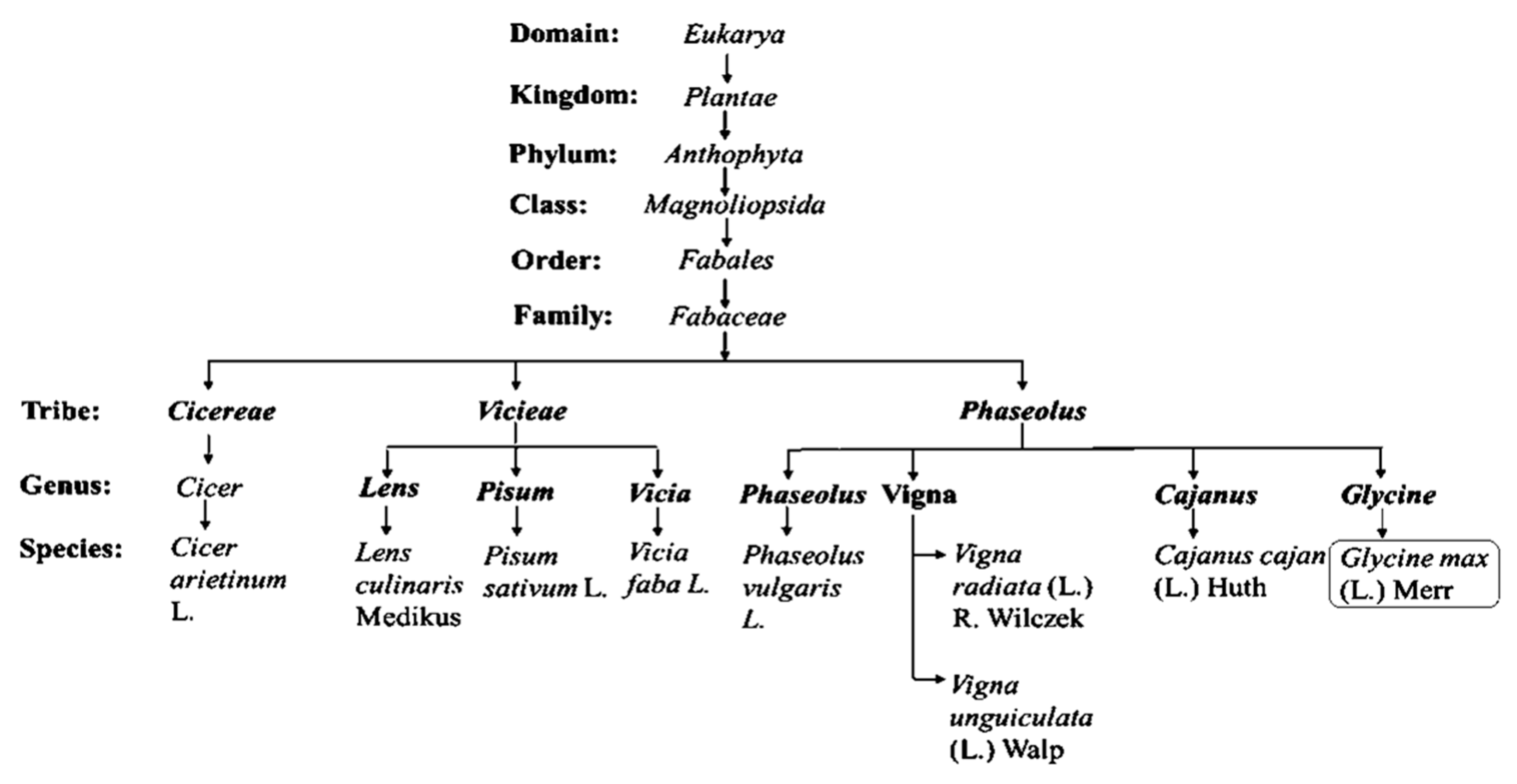
Figure 3.
Potential effects of salinity on the growth and development of polyploidized and diploid soybean plants.
Figure 3.
Potential effects of salinity on the growth and development of polyploidized and diploid soybean plants.
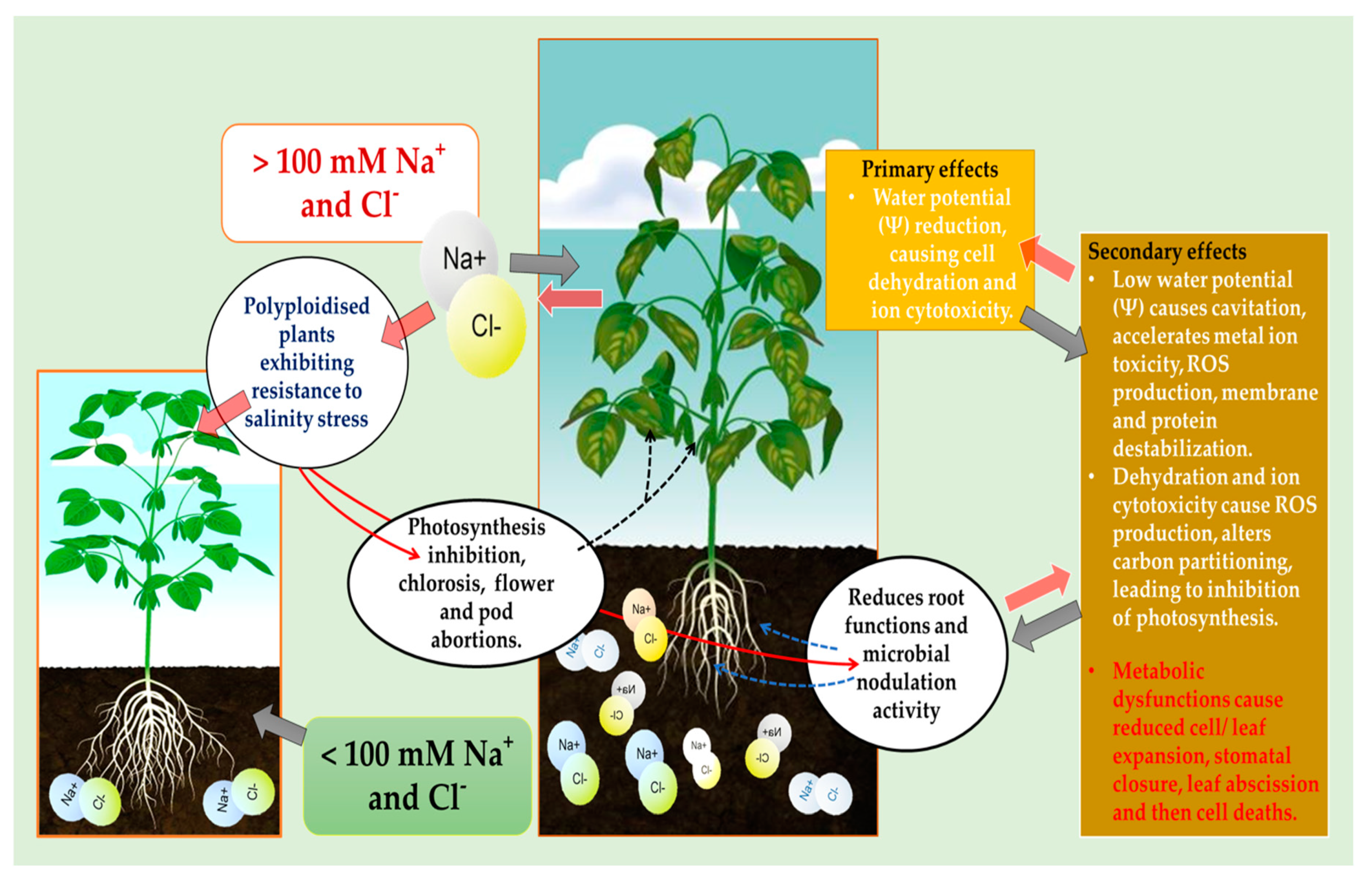
Figure 4.
Approximation of the frequency (percentage) of natural polyploidization occurrences in vascular seed and seedless plants (A) [66] and among the major families of angiosperms comprising species used as food and feed crops (B) [71].
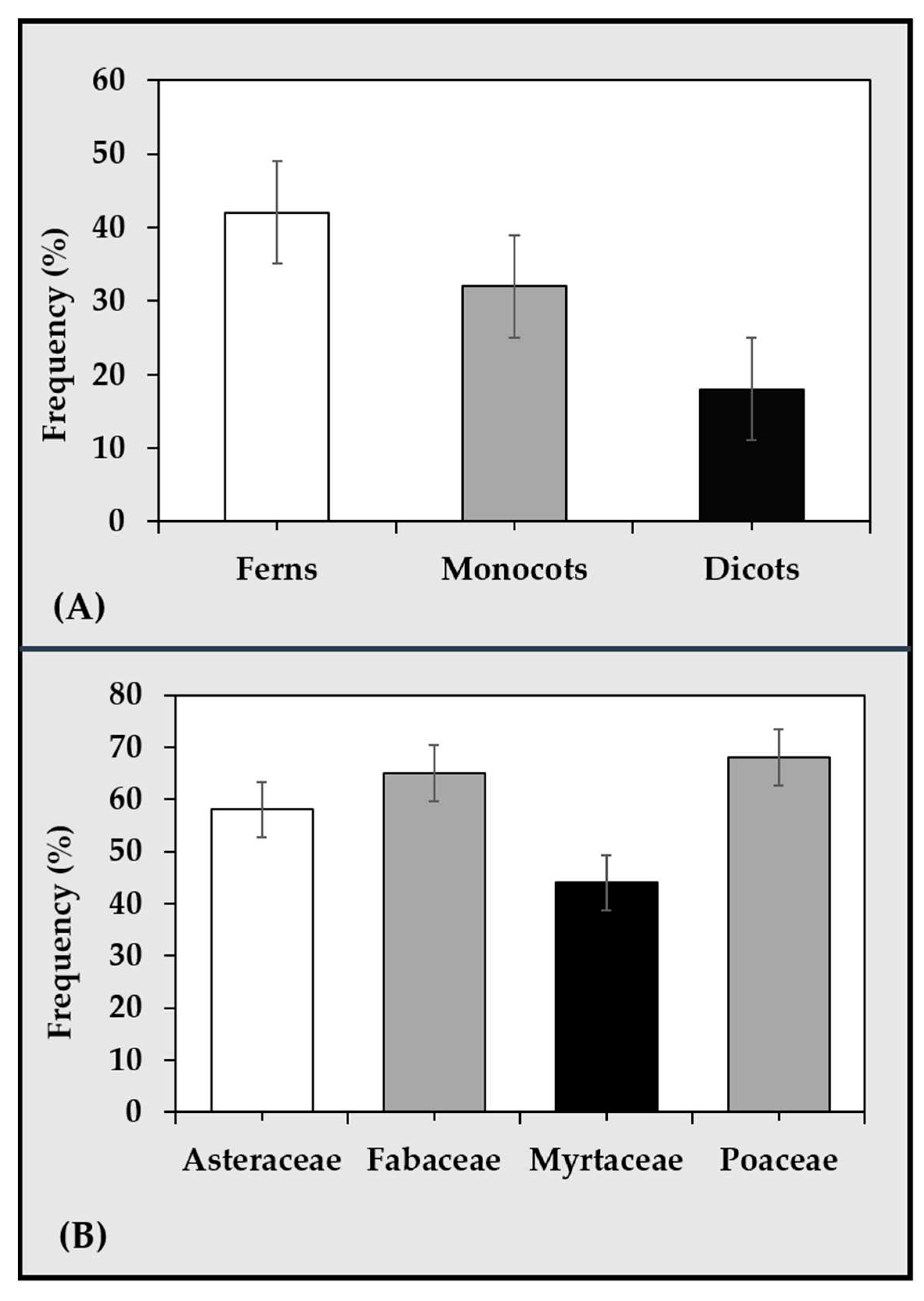

Table 1.
Species of legumes found within the Eurosid I angiosperm group (Fabidae) constituting some of the most economically important Fabale crops, globally [28].
Table 1.
Species of legumes found within the Eurosid I angiosperm group (Fabidae) constituting some of the most economically important Fabale crops, globally [28].
| Tribe | Species name | Common name |
|---|---|---|
| Phaseoleae | Cajanus cajan | Pigeon pea |
| Canavalia ensiformis | Horse bean | |
| Glycine max | Soybean | |
| Mucuna pruriens | Velvet bean | |
| Phaseolus lanatus | Butter bean | |
| Phaseolus acutifolius | Tepary bean | |
| Phaseolus vulgaris | Dry bean | |
| Vigna radiata | Mung bean | |
| Vigna mungo | Black gram | |
| Vigna umbellate | Rice bean | |
| Vigna unguiculata | Cowpea | |
| Vicieae | Lens culinaris | Lentil |
| Pisum sativum | Pea | |
| Vicia faba | Broad bean | |
| Cicereae | Cicer arietinum | Chickpea |
| Dalbergieae | Arachis hypogeae | Peanut |
| Genisteae | Lupinus luteus | Yellow lupin |
| Indigofereae | Cyamopsis tetragonoloba | Guar |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



