
Submitted:
02 March 2023
Posted:
07 March 2023
You are already at the latest version
Abstract
Clinical and histological similarities between sarcoidosis and tuberculosis have driven repeated investigations looking for a mycobacterial cause of sarcoidosis. Over 50 years ago “anonymous mycobacteria” were suggested to have a role in the etiology of sarcoidosis. Both tuberculosis and sarcoidosis have a predilection for lung involvement though each can be found in any area of the body. A key histopathologic feature of both sarcoidosis and tuberculosis is the granuloma – while the tuberculous caseating granuloma has an area of caseous necrosis with a cheesy consistency; the non-caseating granuloma of sarcoidosis does not have this feature. This article reviews and reiterates the complicity of the infectious agent, Mycobacterium avium ss. paratuberculosis (MAP) as a cause of sarcoidosis. MAP is involved in a parallel story as the putative cause of Crohn’s disease, another disease featuring noncaseating granulomas. MAP is a zoonotic agent infecting ruminant animals and is found in dairy products and in environmental contamination of water and air. Despite increasing evidence tying MAP to several human diseases, there is a continued resistance to embracing its pleiotropic roles. "Who Moved My Cheese" is a simple yet powerful book that explores the ways in which individuals react to change. Extending the metaphor, the “non-cheesy” granuloma of sarcoidosis actually contains the difficult-to-detect “cheese”, MAP; MAP did not move, it was there all along.
Keywords:
sarcoidosis
; Mycobacterium avium ss. paratuberculosis
; MAP
; non-caseating granuloma
; caseating granuloma
; Blau syndrome
; early onset sarcoidosis
; Crohn’s disease
; cell wall deficient (CWD)
; L-form
; Mycobacterium tuberculosis
; acid-fast bacilli
; SLC11a1 (NRAMP1)
; NOD2 (CARD15)
1. Introduction
The first clinical record of sarcoidosis dates to 1859 when Jonathan Hutchinson of the Blackfriers Hospital for Diseases of the Skin of London described a patient with raised, purple skin plaques; initially Hutchinson related them to the patient’s gout. Upon seeing another individual, a Mrs. Mortimer, with similar dermatologic findings, Hutchinson differentiated the lesions from tuberculosis and lupus and labeled the condition “Mortimer’s malady” [1]. The label sarcoidosis came in 1899 by Caesar Boeck with the description “multiple benign sarcoid of the skin” as the histology resembled sarcoma. Boeck also noted the remarkable clinical and histologic similarity to cutaneous tuberculosis and was the first to propose possible link between sarcoidosis and tuberculosis [2]. Note that pulmonary involvement was not identified in the early discussion of sarcoidosis as this era predated the clinical use of the X-ray [3]. This diagnostic uncertainty of the association of sarcoidosis and tuberculosis can be seen in the title of a small series from 1957: “Stenosing non-caseating tuberculosis (sarcoidosis) of the bronchi” [4].
Common to many inflammatory/autoimmune diseases is the concept of an “environmental trigger.” Infectious agents have been the suspected cause of sarcoidosis since the early 1900’s with mycobacteria considered the likely agent [5,6,7]. In a 1964 series, the sera of nearly 80% of 280 sarcoidosis patients revealed significant reaction to mycobacterial antigens [8]. In a 1968 case report of sarcoidosis, a non-tuberculous mycobacterium (NTM) was grown from the lymph gland of an individual with sarcoidosis – it was labeled “anonymous mycobacteria” [9]. A 1996 study demonstrated the isolation and/or identification of MAP, or a closely related M. avium complex strain, from sarcoid skin lesions and cerebrospinal fluid [10].
A hallmark of sarcoidosis is the non-caseating granuloma [11]. However, this is by no means pathognomonic; indeed, a mnemonic developed for the differential diagnosis of non-caseating granulomas used every letter of the alphabet [12]. Caseating and non-caseating granulomas are types of inflammatory lesions. Caseating granulomas are characterized by the presence of dead or necrotic tissue that has a cheesy consistency – “caseous necrosis” – where histological features are not preserved. On the other hand, non-caseating granulomas do not contain necrotic tissue and are composed of activated immune cells such as macrophages and T-lymphocytes. Both types of granulomas are found in a variety of conditions; however, caseating granulomas are more commonly associated with bacterial infections such as tuberculosis and non-tuberculous mycobacterial infections whereas non-caseating granulomas are more commonly associated with “immune-mediated disorders” such as sarcoidosis (Figure 1). Another non-caseating granulomatous disease is Crohn’s disease. Because the ever-increasing association of Mycobacterium avium ss. paratuberculosis (MAP) with Crohn’s disease, paralleling this association is an integral part of this manuscript featuring mycobacterial contribution to non-caseating granulomatous disease [13,14]. Naser made major contributions to the MAP/Crohn’s association with reports culturing MAP from the blood of Crohn’s patients [15] and by culturing MAP from the breast milk of Crohn’s patients [16]. Throughout the 1980′s, Chiodini presented case reports finding MAP in Crohn’s patients [17]. Chiodini brought his MAP experience to bear on sarcoidosis in a 1993 case series where he tested sera from sarcoid patients for reactivity against MAP antigens [18]. The 28 patients with sarcoidosis had a significantly higher IgG values compared to controls (p=0.0009) suggesting a no longer anonymous mycobacterium in sarcoidosis but identifying a “mycobacterium of interest” as MAP [18].
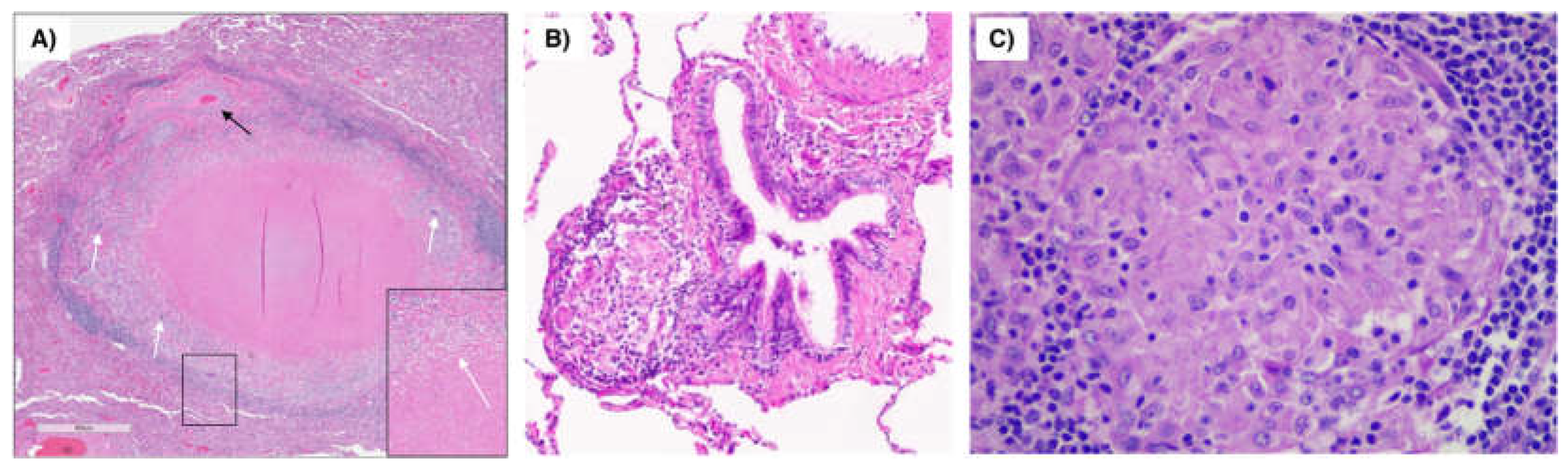
Figure 1.
Pulmonary granuloma due to mycobacteria vs. sarcoidosis. (A) Granuloma with central necrosis surrounded by epithelioid histiocytes, multinucleated giant cells (white arrows) and fibrosis in the resected left lower lobe of a patient with Mycobacterium avium complex lung disease. Note the adjacent pulmonary artery (thin black arrow). Inset shows a magnified multinucleated giant cell. Figure 1A is modified from Schenkel AR et al. Immune Netw 2022. (B) Sarcoid granuloma (arrow) next to a medium size ciliated bronchus. (C) Magnified view of another sarcoid granuloma. Figures 1B/C are courtesy of Carlyne Cool, MD, National Jewish Health.
Figure 1.
Pulmonary granuloma due to mycobacteria vs. sarcoidosis. (A) Granuloma with central necrosis surrounded by epithelioid histiocytes, multinucleated giant cells (white arrows) and fibrosis in the resected left lower lobe of a patient with Mycobacterium avium complex lung disease. Note the adjacent pulmonary artery (thin black arrow). Inset shows a magnified multinucleated giant cell. Figure 1A is modified from Schenkel AR et al. Immune Netw 2022. (B) Sarcoid granuloma (arrow) next to a medium size ciliated bronchus. (C) Magnified view of another sarcoid granuloma. Figures 1B/C are courtesy of Carlyne Cool, MD, National Jewish Health.
Mycobacterium avium subsp. paratuberculosis (MAP)
MAP belongs to the Mycobacterium avium complex (MAC) that comprises 24 species, some of which are pathogenic to animals and humans [19]. MAP is the cause of Johne’s disease, or paratuberculosis, a fatal enteric infectious disease studied mostly in ruminant animals [20]. Infected animals shed MAP in their milk and feces, a state commonly persisting for years prior to the animal displaying clinical Johne’s disease. Johne’s disease is common; the United States Department of Agriculture (USDA) reported that the herd-level prevalence of MAP infection in U.S. dairy herds increased from 21.6% in 1996 to 91.1% in 2007 [21].
Inefficiency of diagnostic testing for MAP along with the long latency of infection in apparent uninfected yet productive dairy animals create hesitancy by producers to test their herds; moreover, there is no mandate requiring testing. Collectively, this results in more MAP-shedding animals that propagate infection and contaminate milk and meat, promotes trade of asymptomatic animals, and delays the mainstay by which Johne’s disease is controlled, animal culling [22].
MAP is a resilient organism and shedding by infected animals is a major source of environmental MAP; once excreted, MAP can survive up to 120 weeks in soil or water [22]. MAP is found in grazing areas as well as in runoff continuing to rivers and in municipal water [23].
Both Iceland and what is now the Czech Republic were historically isolated from Johne’s disease due to limited importing/exporting of ruminant animals. For Iceland this changed in 1933 when apparently healthy, but sub-clinically MAP-infected sheep were imported [24]. Other sheep and cattle on the same importing farms acquired Johne’s disease with the same MAP strain [25]. Similarly, until 1989 Czechoslovakia was isolated; open borders and importation of livestock followed the lifting of the Iron Curtain. Crohn’s disease in both the Icelandic and Czech populations became a lagging indicator of MAP infection [26].
MAP Is Difficult to Detect
Regarding human Crohn’s samples, the basis of the hundred-year controversy is the fact that traditional culturing and staining are largely unsuccessful in identifying MAP [14,27,28]. Aside from M. leprae, the microbial cause of leprosy that has never been grown in culture, MAP is the slowest growing of the mycobacteria with some labs continuing for a year before calling a culture MAP-negative. Veterinary laboratories employing both enhanced conventional and molecular methods for identifying MAP in ruminant animals such as cattle and sheep have the greatest experience in detecting MAP [29].
In addition to being difficult to culture, MAP can exist with a modified cell wall—the component of the mycobacterium that takes up a characteristic acid-fast stain. MAP can shed its cell wall, becoming a spheroplast or L-form [30]. The bacterium is then no longer “acid-fast” and cannot be detected microscopically in the traditional manner. This morphologic change allows MAP to become spore-like. The spore morphotype capable of surviving heat and other stressors enables MAP to persist in host macrophages and in the environment [31].
Humans are exposed to MAP in contaminated food, water and air [32]. A primary reason that MAP is such a risk for human consumers is because this bacterium is particularly resistant to heat [33]. Milk and dairy products are the primary sources of MAP infection in humans [34] and pasteurization only reduces the MAP load in milk [34,35]. MAP is present in yogurt [36], muscle meat [37], and hamburgers [38]. Alarmingly, viable MAP is found in infant formula, including infant formula powder [39,40,41].
For this manuscript, MAP in cheese is featured. The U.S. Food and Drug Administration allows two options for producing safe cheese products: either use of pasteurized milk in cheese production or curing of the finished cheese for at least 60 days at temperatures of 2°C. The curing process can include low pH and various salt concentrations to lessen pathogens; like pasteurization, efforts in the curing process to curtail MAP have some, but not total success, with MAP in cheeses [42]. MAP is found in Swiss cheese [43] and persists in the manufacture and ripening of cheddar cheese [44]. Cheese from sheep and goat as well as combinations of these contain MAP [45]. Likewise, MAP is detected in the pasta-filata cheeses such as mozzarella, burrata and provolone [46].
To date, MAP has been associated with Crohn’s disease, Blau syndrome, autoimmune (Hashimoto’s) thyroiditis, autoimmune diabetes, multiple sclerosis, lupus erythematosus, rheumatoid arthritis and Parkinson’s disease [47,48]. While it is intuitive to assign a role to MAP in granulomatous disease, MAP’s contribution to inciting autoimmune disease is less obvious; it is envisaged to be due to molecular mimicry to one or more of its proteins, primarily epitope homology to its heat shock protein 65 (HSP65) [49].
Sarcoid Epidemiology
Reports on the epidemiology of sarcoidosis are quite varied. Such disparity is likely due to a number of factors including [50,51]: (i) retrospective analyses using large health administrative data with inherent inaccuracies in diagnostic coding; (ii) bias collection of case series which may not include milder cases; (iii) potential inclusion of mimics of sarcoidosis as sarcoidosis is best considered a syndrome (“sarcoidoses”); i.e., sarcoidosis is often considered to be due to an environmental exposure (along with host susceptibility) to various agents and thus different regions of the world are likely to have different inherent differences in incidence and prevalence; and (iv) greater diagnostic detection of (asymptomatic) sarcoidosis with greater use of CT scans for other reasons. Nevertheless, we have attempted to summarize below the epidemiology of sarcoidosis.
Erdal and co-workers [52] analyzed the electronic medical records of a large institution network (The Ohio State University Medical Center’s Institutional Information Warehouse) from 1995 to 2010 to determine the temporally prevalence of sarcoidosis. Using the ICD9 code for sarcoidosis (ICD9 135), they found that the prevalence increased steadily from 164 / 105 in 1995 to 330 / 105 in 2010. When they examined the prevalence of sarcoidosis in Columbus, Ohio (Franklin County) – a region with a demographic profile (race, gender, age) that is nearly identical to the U.S. as a whole – the prevalence of sarcoidosis was estimated to be 48 / 105 in 2010 (and may be as high as 200 / 105), significantly higher than the U.S. as a whole. The authors speculated that although an actual increase in the number of sarcoidosis cases could be occurring over this 15-year period, greater use of CT scans, the availability of less invasive diagnostic techniques, and perhaps increased awareness of sarcoidosis by clinicians likely played a role in the increased prevalence.
In a similar study from Korea of sarcoidosis between 2007 and 2016 – coded by the Korean Classification of Disease – the annual incidence rates of sarcoidosis increased from 0.85 / 105 in 2009 to 0.97 / 105 in 2015 [53]. The highest incidence rate occurred in those 50-59 years for females while biphasic peaks were seen in males (30-39 years and 60-69 years). In this population that is racially different and more homogenous than the U.S. population, the 10-year prevalence was estimated to be 11.44 / 105 in females and 7.30 / 105 in males (combined = 9.37 / 105).
In a study from Ontario, Canada, the prevalence of sarcoidosis increased from 66 / 105 in 1996 to 143 / 105 in 2015, a highly significant increase of 116% [54]. Interestingly, the incidence of sarcoidosis decreased from 7.9 / 105 in 1996 to 6.8 / 105 in 2014; when the incidence was stratified by gender, there was a 30% decrease in the incidence in females but a 5.5% increase in males. The authors speculated that the higher sarcoidosis incidence in males in the latter years examined was due to increase in occupational dust exposures as there was a significant increase in the construction industry in Ontario during this period.
Seedahmed and co-workers [55] analyzed the Veterans Health Administration electronic health record system to define the epidemiology of sarcoidosis from 2003 to 2019. Among 13 million veterans, there were 23,747 (0.20%) incident diagnosis of sarcoidosis, approximately four-fold greater than the general U.S. population. The annual incidence of sarcoidosis in the veteran population increased from 38 to 52 cases/105 person-years and the annual prevalence from 79 to 141 cases/105 persons [55]. They also found that living in the Northeast and serving in Army, Air Force, and multiple branches were more associated with a diagnosis of sarcoidosis than working in the Navy.
Genetic Risk for Sarcoidosis Overlaps with Risk for Mycobacterial Infection
Sarcoidosis is most likely the result of some form of genetic susceptibility plus an inciting environmental factor. There is a 3.7-fold increase in sarcoidosis with having at least one first degree relative with sarcoidosis [56]. While sarcoidosis may affect all races, Black race appears to have increased vulnerability. A large U.S. military veteran study found that, using multivariate analysis, Black race, female sex, and history of tobacco use were significantly associated with a diagnosis of sarcoidosis [55]. In general, Black individuals experience more severe disease, multi-organ involvement, and higher rate of hospitalization [51]. Additionally, Black females have been shown to have higher sarcoidosis mortality rates [57].
SLC11a1 (NRAMP) and Sarcoidosis
Risk of sarcoidosis is associated with polymorphisms of the SLC11A1 gene [58]. What was initially termed natural resistance-associated macrophage protein 1 (NRAMP1) is now referred to as SLC11A1 (solute carrier 11a1). The gene that encodes for this protein is recognized as having a role in the susceptibility of both humans and animals to infections, including mycobacterial infections; moreover, it is associated with several inflammatory and autoimmune diseases. In human beings, the SLC11A1 gene is located on chromosome 2q35. It encodes an integral membrane protein of 550 amino acids that is expressed exclusively in the lysosomal compartment of monocytes and macrophages [59]. The product of the SLC11A1 gene modulates the cellular environment in response to activation by intracellular pathogens by acidifying the phagosome thus killing the pathogen [60]. As such, it plays a role in host innate immunity [61]. Mutation of SLC11A1 impairs phagosome acidification yielding a permissive environment for the persistence of intracellular bacteria [62].
Susceptibility to mycobacterial diseases tuberculosis, leprosy and Buruli’s ulcer are associated with polymorphism of the SLC11A1 gene [63,64,65,66]. Similar polymorphisms are associated with Johne’s disease in cattle [67], goats [68], and sheep [69]. When researchers at the Belgium Pasteur Institute developed a murine model for MAP infection, they created an SLC11A1 defect mouse [70].
HLA Alleles and Sarcoidosis/Tuberculosis Risk
The human leukocyte antigen (HLA) system coordinates the adaptive immune response to foreign antigens, which includes both infectious and non-infectious agents. The HLA genes encode major histocompatibility complex (MHC) molecules, which are found on antigen presenting cells that process and present antigens to T-lymphocytes, resulting in the initiation of the adaptive immune response to that antigen [76].
The T-lymphocytes that proliferate in sarcoidosis often show a restricted repertoire of T-cell receptors suggesting a limited number of peptides are responsible for the expansion of these T-lymphocytes [77]. Research in sarcoidosis suggests that these peptides may have mycobacterial origins; e.g., mycobacteria can be found in the 23-30% of tissues samples from those with sarcoidosis [78].
Many HLA alleles are thought to be associated with sarcoidosis. The U.S. ACCESS study found that HLADRB1*1101, DRB1*1201, DRB1*1501, and DRB1*0402 alleles were strongly associated with sarcoidosis [79]. Additionally, HLA-DRB1*03:01 was associated with an acute form of sarcoidosis called Lofgren’s syndrome, while HLA-DRB1*1101 was associated with chronic sarcoidosis [80,81]. A large genome-wide association study of ocular sarcoidosis found a link with HLA-DRB1*04:01 and MAGI1 [82]. These findings underscore the importance of analyzing and sarcoid phenotypes separately in genetic studies.
HLA alleles are associated with different risks for tuberculosis as well. HLA-DRB1*04 and HLA-DRB1*08 genotypes seem to increase the risk for infection, while HLA-DRB1*03 and HLA-DRB1*07 genotypes seem to decrease the risk [76]. It should be noted that genotype distribution is also highly affected by ethnic differences, which makes associations difficult to generalize across populations.
Shared HLA alleles in individuals with sarcoidosis and Mycobacterium tuberculosis infections suggest a possibly common pathogenic pathway between sarcoidosis and mycobacterial infections. As mentioned before, HLA-DRB1*1101 has been associated with susceptibility to sarcoidosis. Oswald-Richter et al. demonstrated that individuals with HLA-DRB1*1101 were statistically more likely to recognize ESAT-6, a peptide derived from mycobacteria [83]. These authors also showed that antigen presenting cells expressing HLA-DRB1*1101 interact with mycobacterial peptides, ESAT-6 and katG, to elicit a TH1 polarized response from sarcoidosis T-cells [83]. Interestingly, a dysregulated TH1 response is theorized to be the immune dysfunction responsible for sarcoidosis. Another study, using a bioinformatic approach, demonstrated that individuals with sarcoidosis tended to express HLA-DR allele combinations that recognize M. tuberculosis and M. avium epitopes with greater affinity than those individuals with tuberculosis, suggesting that sarcoidosis may be a hypersensitive reaction to mycobacterial antigens [78]. These results suggest that HLA genes and mycobacterial peptides may contribute to the immune dysregulation seen in sarcoidosis and may provide a mechanistic explanation for the etiology of sarcoidosis.
HLA polymorphisms in individuals with sarcoidosis and M. tuberculosis infections may contribute to different sub-phenotypes in both diseases. In a study by Dawkin et al., it was found that certain HLA alleles were found in higher frequencies in individuals who never developed M. tuberculosis infections despite close contact with an index case and in those with acute/resolved sarcoidosis compared to individuals with M. tuberculosis infections and chronic sarcoidosis, respectively [84].
The different phenotypes of sarcoidosis may be the result of how different HLA alleles respond to mycobacterial peptides. Grosser et al found that using HLA-DRB1 alleles in combination with M. tuberculosis status enabled the authors to predict the sarcoidosis phenotype: sarcoidosis cases with either HLA-DRB1*03 or -DRB1*04 allele and the absence of M. tuberculosis DNA were associated with acute/resolved disease, while cases with HLA-DRB1*11, -DRB1*15 and the presence of M. tuberculosis disease were associated with chronic sarcoidosis [85]. The authors hypothesized that M. tuberculosis could be an antigen that produces a more severe clinical course in sarcoidosis with certain HLA alleles [85]. The study by Saltini et al supports this hypothesis with a study that showed individuals with acute sarcoidosis had HLA-DR genomes with an exaggerated ability to recognized M. tuberculosis and M. avium epitopes, while those with chronic versions of the disease had HLA alleles with lower affinity to the same epitopes [78]. The authors posited this could represent an efficient immune response to mycobacterial antigens in acute sarcoidosis with subsequent clearance and resolution of disease. The HLA alleles associated with chronic sarcoidosis may produce a less efficient response to mycobacterial antigens – due in part to a lower affinity for these epitopes – resulting in antigen persistence, continuous granuloma formation, and chronic disease [78].
CARD15 (NOD2)–Early Onset Sarcoidosis (EOS) and Blau Syndrome
The CARD15 gene is part of the ancestral innate immune system that senses and eliminates bacteria [86]; originally referred to as the NOD2, it is now known as the CARD15 gene [87]. Blau syndrome is a rare disease with the clinical triad of dermatitis, arthritis, and uveitis. It is the only inflammatory disease that is autosomal dominantly inherited and is monogenic. Blau syndrome has generated interest in medical literature because of the discovery that places its genetic defect with the same Crohn’s susceptibility CARD15 gene [88,89]. The Blau syndrome susceptibility component of CARD15 is found within the nucleotide binding site domain [90] while the Crohn’s susceptibility is in the N-terminal leucine- rich repeat domain [91]. In a small series looking for MAP in six individual Blau tissues, including skin, synovium and granulomas of the liver and kidney, all six tissues had the DNA of MAP [92]. Early onset sarcoidosis (EOS) is the identical clinical presentation as Blau syndrome but without the family history [93,94].
An Illustrative Case of Cardiac Sarcoidosis
Cardiac involvement is an increasingly recognized and serious manifestation of sarcoidosis ranging from silent myocardial granulomas leading to sudden death [95,96,97] to symptomatic arrhythmias and heart failure [98]. In isolation, cardiac sarcoidosis can be very difficult to detect [99]. In a patient with sarcoidosis who died of a heart attack, MAP was detected in the noncaseating granuloma that infiltrated the cardiac tissue [100].
A particularly instructional sarcoidosis case was reported in 2018: Case Study: Cardiac sarcoidosis resolved with Mycobacterium avium paratuberculosis antibiotics (MAP) [101]. The patient, a physician, had inactive sarcoidosis later followed by acute cardiac sarcoidosis manifest as complete heart block, cardiomyopathy, and heart failure. His own literature search led him to the prospect of MAP as a cause for sarcoidosis and the analogous use of MAP-targeting antibiotics for Crohn’s disease [102] as a possible treatment. He sent blood to the only commercial lab culturing human samples for MAP, Otakaro Pathways in New Zealand (https://otakaropathways.co.nz). His blood was positive for MAP. The MAP-targeting anti-mycobacterial treatment resulted in complete resolution of his cardiac sarcoidosis and no residual pulmonary nodules nor mediastinal or distant lymphadenopathy [101]. This parallels similar MAP-targeting antibiotic therapy for Crohn’s disease [103,104,105].
2. Discussion
An entrenched tenant is that sarcoidosis is a disease of undetermined etiology. While the parallel non-granulomatous story of MAP and Crohn’s disease has garnered an increasingly tighter association, a MAP/sarcoidosis association remains to be made known; this article proposes a causal role for MAP in sarcoidosis. As presented in various sections of this paper, MAP has been found in juvenile sarcoidosis tissues and anti-MAP antibiotic therapy has successfully treated cardiac sarcoidosis. Revealed in the epidemiology section of this article, sarcoidosis is increasing in its prevalence; also, the genetics section of this article indicates that the risks for sarcoidosis are shared risks for mycobacterial infection. Parsimony favors a causal role for mycobacteria in sarcoidosis. A mycobacterium to which humans are regularly exposed is MAP, a mycobacterium that is difficult to detect. A consensus article calling for MAP zoonotic designation was the product of a 2017 international assembly of MAP researchers – such precautionary measures have yet to be taken [106].
The book “Who Moved My Cheese?” by Spencer Johnson is about four characters who live in a maze and rely on finding cheese to survive [107]. The cheese in the story represents what people seek and the way that different individuals respond to change and uncertainty in their lives. The book encourages readers to be adaptable and flexible in the face of change, and to embrace new opportunities even when they may be uncomfortable or unfamiliar.
This article, like the book, asks the reader to consider something unfamiliar: the inertia of MAP’s causal involvement in an increasing list of inflammatory and autoimmune diseases as it applies to sarcoidosis [48]. Understanding the limitations of conventional laboratory investigations as they relate to finding MAP, molecular investigation employed by dedicated veterinary laboratories are the key to exposing MAP as the previous anonymous mycobacterium associated with sarcoidosis.
In the book, characters searching the maze for cheese leave messages on the wall – they do this in hopes that those who follow will “see the writing on the wall.” Extending the metaphor, the writing on the wall of this article is that although there is no apparent cheesy necrosis in the noncaseous granulomas of sarcoidosis, the sought-after cheese (“MAP”) was there all along.
Author Contributions
CTD conceptualized the content of the paper; CTD, NWL, and EDC all participated in writing the manuscript.
Abbreviations
HLA = human leukocyte antigen; MAP = Mycobacterium avium subsp Paratuberculosis; NTM = non-tuberculous mycobacteria
References
- Spagnolo, P. Sarcoidosis: a Critical Review of History and Milestones. Clin Rev Allergy Immunol. 2015,49,1-5. [CrossRef]
- Gupta, D. Tuberculosis and sarcoidosis: The continuing enigma. Lung India. 2009,26,1-2. [CrossRef]
- Howell, J.D. Early Clinical Use Of The X-Ray. Trans Am Clin Climatol Assoc. 2016,127,341-49.
- Citron, K.M.;Scadding, J.G. Stenosing non-caseating tuberculosis (sarcoidosis) of the bronchi. Thorax. 1957,12,10-17. [CrossRef]
- Rook, G.A.;Stanford, J.L. Slow bacterial infections or autoimmunity? Immunol Today. 1992,13,160-64.
- Gupta, D.; Agarwal, R.; Aggarwal, A.N.;Jindal, S.K. Molecular evidence for the role of mycobacteria in sarcoidosis: a meta-analysis. Eur Respir J. 2007,30,508-16. [CrossRef]
- Brownell, I.; Ramírez-Valle, F.; Sanchez, M.;Prystowsky, S. Evidence for mycobacteria in sarcoidosis. Am J Respir Cell Mol Biol. 2011,45,899-905. [CrossRef]
- Chapman, J.S.;Speight, M. Further studies of mycobacterial antibodies in the sera of sarcoidosis patients. Acta Med Scand. 1964,Suppl 425,61-67. [CrossRef]
- Berger, H.W.; Zaldivar, C.;Chusid, E.L. Anonymous mycobacteria in the etiology of sarcoidosis. Ann Intern Med. 1968,68,872-74. [CrossRef]
- el-Zaatari, F.A.; Naser, S.A.; Markesich, D.C.; Kalter, D.C.; Engstand, L.;Graham, D.Y. Identification of Mycobacterium avium complex in sarcoidosis. J Clin Microbiol. 1996,34,2240-45. [CrossRef] [PubMed]
- Llanos, O.;Hamzeh, N. Sarcoidosis. Med Clin North Am. 2019,103,527-34.
- Mehta, A.C.;Ali, S.R. Mnemonic for the differential diagnosis of non-caseating granulomas. Sarcoidosis Vasc Diffuse Lung Dis. 2017,34,200-07. [CrossRef]
- Jeyanathan, M.; Alexander, D.C.; Turenne, C.Y.; Girard, C.;Behr, M.A. Evaluation of in situ methods used to detect Mycobacterium avium subsp. paratuberculosis in samples from patients with Crohn’s disease. J Clin Microbiol. 2006,44,2942-50. [CrossRef]
- Sechi, L.A.;Dow, C.T. Mycobacterium avium ss. paratuberculosis Zoonosis - The Hundred Year War - Beyond Crohn’s Disease. Front Immunol. 2015,6,96. [CrossRef]
- Naser, S.A.; Ghobrial, G.; Romero, C.;Valentine, J.F. Culture of Mycobacterium avium subspecies paratuberculosis from the blood of patients with Crohn’s disease. . Lancet. 2004,364,1039-44. [CrossRef] [PubMed]
- Naser, S.A.; Schwartz, D.;Shafran, I. Isolation of Mycobacterium avium subsp paratuberculosis from breast milk of Crohn’s disease patients. Am J Gastroenterol. 2000,95,1094-95. [CrossRef]
- Chiodini, R.J. Crohn’s disease and the mycobacterioses: a review and comparison of two disease entities. . Clin Microbiol Rev. 1989,2,90-117. [CrossRef]
- Reid, J.D.;Chiodini, R.J. Serologic reactivity against Mycobacterium paratuberculosis antigens in patients with sarcoidosis. Sarcoidosis. 1993,10,32-35.
- Biet, F.; Boschiroli, M.L.; Thorel, M.F.;Guilloteau, L.A. Zoonotic aspects of Mycobacterium bovis and Mycobacterium avium-intracellulare complex (MAC). Vet Res. 2005,36,411-36. [CrossRef]
- Whittington, R.J.;Sergeant, E.S. Progress towards understanding the spread, detection and control of Mycobacterium avium subsp paratuberculosis in animal populations. Aust Vet J. 2001,79,267-78. [CrossRef]
- Lombard, J.E.; Gardner, I.A.; Jafarzadeh, S.R.; Fossler, C.P.; Harris, B.; Capsel, R.T.; Wagner, B.A.;Johnson, W.O. Herd-level prevalence of Mycobacterium avium subsp. paratuberculosis infection in United States dairy herds in 2007. Prev Vet Med. 2013,108,234-38. [CrossRef]
- Garvey, M. Mycobacterium avium paratuberculosis: A Disease Burden on the Dairy Industry. . Animals (Basel). 2020,10,1773. [CrossRef] [PubMed]
- Beumer, A.; King, D.; Donohue, M.; Mistry, J.; Covert, T.;Pfaller, S. Detection of Mycobacterium avium subsp. paratuberculosis in drinking water and biofilms by quantitative PCR. Appl Environ Microbiol. 2010,76,7367-70. [CrossRef]
- Fridriksdottir, V.; Gunnarsson, E.; Sigurdarson, S.;Gudmundsdottir, K.B. Paratuberculosis in Iceland: epidemiology and control measures, past and present. Vet Microbiol. 2000,77,263-67. [CrossRef] [PubMed]
- Whittington, R.J.; Taragel, C.A.; Ottaway, S.; Marsh, I.; Seaman, J.;Fridriksdottir, V. Molecular epidemiological confirmation and circumstances of occurrence of sheep (S) strains of Mycobacterium avium subsp. paratuberculosis in cases of paratuberculosis in cattle in Australia and sheep and cattle in Iceland. Vet Microbiol. 2001,79,311-22. [CrossRef]
- Dow, C.T.;Alvarez, B.L. Mycobacterium paratuberculosis zoonosis is a One Health emergency. . Ecohealth. 2022,19,164-74. [CrossRef] [PubMed]
- Chamberlin, W.; Borody, T.;Naser, S. MAP-associated Crohn’s disease MAP, Koch’s postulates, causality and Crohn’s disease. Dig Liver Dis. 2007,39,792-94. [CrossRef]
- Hansen, R.; Thomson, J.M.; El-Omar, E.M.;Hold, G.L. The role of infection in the aetiology of inflammatory bowel disease. J Gastroenterol. 2010,45,266-76. [CrossRef] [PubMed]
- Harris, N.B.;Barletta, R.G. Mycobacterium avium subsp. paratuberculosis in Veterinary Medicine. . Clin Microbiol Rev. 2001,14,489-512. [CrossRef]
- Hines, M.E., 2nd.;Styer, E.L. Preliminary characterization of chemically generated Mycobacterium avium subsp. paratuberculosis cell wall deficient forms (spheroplasts). Vet Microbiol. 2003,95,247-58. [CrossRef]
- Lamont, E.A.; Bannantine, J.P.; Armién, A.; Ariyakumar, D.S.;Sreevatsan, S. Identification and characterization of a spore-like morphotype in chronically starved Mycobacterium avium subsp. paratuberculosis cultures. PLoS One. 2012,7,e30648. [CrossRef]
- Waddell, L.; Rajić, A.; Stärk, K.;McEwen, S.A. Mycobacterium avium ssp. paratuberculosis detection in animals, food, water and other sources or vehicles of human exposure: A scoping review of the existing evidence. . Prev Vet Med. 2016,132,32-48. [CrossRef]
- Mullan, W. Are we closer to understanding why viable cells of Mycobacterium avium subsp. paratuberculosis are still being reported in pasteurised milk? Int J Dairy Technol. 2019,72,332–44. [CrossRef]
- Gill, C.O.; Saucier, L.;Meadus, W.J. Mycobacterium avium subsp. paratuberculosis in dairy products, meat, and drinking water. J Food Prot. 2011,74,480-99. [CrossRef]
- Eltholth, M.M.; Marsh, V.R.; Van Winden, S.;Guitian, F.J. Contamination of food products with Mycobacterium avium paratuberculosis: a systematic review. J Appl Microbiol. 2009,107,1061-71. [CrossRef] [PubMed]
- Van Brandt, L.; Coudijzer, K.; Herman, L.; Michiels, C.; Hendrickx, M.;Vlaemynck, G. Survival of Mycobacterium avium ssp. paratuberculosis in yoghurt and in commercial fermented milk products containing probiotic cultures. . J Appl Microbiol. 2011,110,1252-61. [CrossRef]
- Alonso-Hearn, M.; Molina, E.; Geijo, M.; Vazquez, P.; Sevilla, I.; Garrido, J.M.;Juste, R.A. Isolation of Mycobacterium avium subsp. paratuberculosis from muscle tissue of naturally infected cattle. Foodborne Pathog Dis. 2009,6,513-18. [CrossRef]
- Hammer, P.; Walte, H.G.; Matzen, S.; Hensel, J.;Kiesner, C. Inactivation of Mycobacterium avium subsp. paratuberculosis during cooking of hamburger patties. J Food Prot. 2013,76,1194-201. [CrossRef] [PubMed]
- Hruska, K.; Bartos, M.; Kralik, P.;Pavlik, I. Mycobacterium avium subsp. paratuberculosis in powdered infant milk: Paratuberculosis in cattle—The public health problem to be solved. Vet Med. 2005,50,327–35.
- Hruska, K.; Slana, I.; Kralik, P.;Pavlik, I. Mycobacterium avium subsp. paratuberculosis in powdered infant milk: F57 competitive real time PCR. Vet Med. 2011,56,226–30. [CrossRef]
- Botsaris, G.; Swift, B.M.; Slana, I.; Liapi, M.; Christodoulou, M.; Hatzitofi, M.; Christodoulou, V.;Rees, C.E. Detection of viable Mycobacterium avium subspecies paratuberculosis in powdered infant formula by phage-PCR and confirmed by culture. Int J Food Microbiol. 2016,216,91-94. [CrossRef] [PubMed]
- Sung, N.;Collins, M.T. Effect of three factors in cheese production (pH, salt, and heat) on Mycobacterium avium subsp. paratuberculosis viability. Appl Environ Microbiol. 2000,66,1334-39. [CrossRef] [PubMed]
- Spahr, U.;Schafroth, K. Fate of Mycobacterium avium subsp. paratuberculosis in Swiss hard and semihard cheese manufactured from raw milk. . Appl Environ Microbiol. 2001,67,4199-205. [CrossRef]
- Donaghy, J.A.; Totton, N.L.;Rowe, M.T. Persistence of Mycobacterium paratuberculosis during manufacture and ripening of cheddar cheese. Appl Environ Microbiol. 2004,70,4899-905. [CrossRef]
- Galiero, A.; Fratini, F.; Mataragka, A.; Turchi, B.; Nuvoloni, R.; Ikonomopoulos, J.;Cerri, D. Detection of Mycobacterium avium subsp. paratuberculosis in cheeses from small ruminants in Tuscany. Int J Food Microbiol. 2016,217,195-99. [CrossRef]
- Barsi, F.; Dalzini, E.; Russo, S.; Cosciani-Cunico, E.; Monastero, P.; Arrigoni, N.; Garbarino, C.A.; Cortimiglia, C.; Losio, M.N.;Ricchi, M. Isothermal inactivation of Mycobacterium avium subsp. paratuberculosis in curd simulating the stretching phase in pasta-filata cheese process. Front Microbiol. 2022,13,1052222. [CrossRef]
- Dow, C.T.;Sechi, L.A. Cows Get Crohn’s Disease and They’re Giving Us Diabetes. Microorganisms. 2019,7,466. [CrossRef]
- Ekundayo, T.C.;Okoh, A.I. Systematic Assessment of Mycobacterium avium Subspecies Paratuberculosis Infections from 1911-2019: A Growth Analysis of Association with Human Autoimmune Diseases. Microorganisms. 2020,8,1212. [CrossRef]
- Dow, C.T. M. paratuberculosis Heat Shock Protein 65 and Human Diseases: Bridging Infection and Autoimmunity. Autoimmune Dis. 2012,2012,150824. [CrossRef] [PubMed]
- Arkema, E.V.;Cozier, Y.C. Sarcoidosis epidemiology: recent estimates of incidence, prevalence and risk factors. Curr Opin Pulm Med. 2020,26,527-34. [CrossRef]
- Hena, K.M. Sarcoidosis Epidemiology: Race Matters. Front Immunol. 2020,11,537382. [CrossRef]
- Erdal, B.S.; Clymer, B.D.; Yildiz, V.O.; Julian, M.W.;Crouser, E.D. Unexpectedly high prevalence of sarcoidosis in a representative U.S. Metropolitan population. Respir Med. 2012,106,893-99. [CrossRef] [PubMed]
- Yoon, H.Y.; Kim, H.M.; Kim, Y.J.;Song, J.W. Prevalence and incidence of sarcoidosis in Korea: a nationwide population-based study. Respir Res. 2018,19,158. [CrossRef] [PubMed]
- Fidler, L.M.; Balter, M.; Fisher, J.H.; To, T.; Stanbrook, M.B.;Gershon, A. Epidemiology and health outcomes of sarcoidosis in a universal healthcare population: a cohort study. Eur Respir J. 2019,54,1900444. [CrossRef]
- Seedahmed, M.I.; Baugh, A.D.; Albirair, M.T.; Luo, Y.; Chen, J.; McCulloch, C.E.; Whooley, M.A.; Koth, L.L.;Arjomandi, M. Epidemiology of Sarcoidosis in U.S. Veterans from 2003 to 2019. Ann Am Thorac Soc. 2023,E-pub. [CrossRef]
- Rossides, M.; Grunewald, J.; Eklund, A.; Kullberg, S.; Di Giuseppe, D.; Askling, J.;Arkema, E.V. Familial aggregation and heritability of sarcoidosis: a Swedish nested case-control study. Eur Respir J.52,1800385. [CrossRef]
- Swigris, J.J.; Olson, A.L.; Huie, T.J.; Fernandez-Perez, E.R.; Solomon, J.; Sprunger, D.;Brown, K.K. Sarcoidosis-related mortality in the United States from 1988 to 2007. Am J Respir Crit Care Med. 2011,183,1524-30. [CrossRef]
- Dubaniewicz, A.; Jamieson, S.E.; Dubaniewicz-Wybieralska, M.; Fakiola, M.; Miller, E.N.;Blackwell, J.M. Association between SLC11A1 (formerly NRAMP1) and the risk of sarcoidosis in Poland. Eur J Hum Genet. 2005,13,829-34. [CrossRef]
- Canonne-Hergaux, F.; Gruenheid, S.; Govoni, G.;Gros, P. The Nramp1 protein and its role in resistance to infection and macrophage function. Proc Assoc Am Physicians. 1999,111,283-89. [CrossRef]
- Lapham, A.S.; Phillips, E.S.;Barton, C.H. Transcriptional control of Nramp1: a paradigm for the repressive action of c-Myc. Biochem Soc Trans. 2004,32(Pt 6),1084-86. [CrossRef] [PubMed]
- Wyllie, S.; Seu, P.;Goss, J.A. The natural resistance-associated macrophage protein 1 Slc11a1 (formerly Nramp1) and iron metabolism in macrophages. Microbes Infect. 2002,4,351-59. [CrossRef] [PubMed]
- Hackam, D.J.; Rotstein, O.D.; Zhang, W.; Gruenheid, S.; Gros, P.;Grinstein, S. Host resistance to intracellular infection: mutation of natural resistance-associated macrophage protein 1 (Nramp1) impairs phagosomal acidification. J Exp Med. 1998,188,351-64. [CrossRef] [PubMed]
- Stienstra, Y.; van der Werf, T.S.; Oosterom, E.; Nolte, I.M.; van der Graaf, W.T.; Etuaful, S.; Raghunathan, P.L.; Whitney, E.A.; Ampadu, E.O.; Asamoa, K.; et al. Susceptibility to Buruli ulcer is associated with the SLC11A1 (NRAMP1) D543N polymorphism. Genes Immun. 2006,7,185-89. [CrossRef]
- Brochado, M.J.; Gatti, M.F.; Zago, M.A.;Roselino, A.M. Association of the solute carrier family 11 member 1 gene polymorphisms with susceptibility to leprosy in a Brazilian sample. Mem Inst Oswaldo Cruz. 2016,111,101-05. [CrossRef] [PubMed]
- Hu, B.; Wang, Y.; Wang, Z.; He, X.; Wang, L.; Yuan, D.; He, Y.; Jin, T.;He, S. Association of SLC11A1 Polymorphisms With Tuberculosis Susceptibility in the Chinese Han Population. Front Genet. 2022,13,899124. [CrossRef]
- Shahzad, F.; Bashir, N.; Ali, A.; Nadeem, A.; Ammar, A.; Kashif, M.; Javaid, K.; Jahan, S.; Tahir, R.; Rizwan, M.; et al. SLC11A1 genetic variation and low expression may cause immune response impairment in TB patients. Genes Immun. 2022,23,85-92. [CrossRef] [PubMed]
- Ruiz-Larrañaga, O.; Garrido, J.M.; Manzano, C.; Iriondo, M.; Molina, E.; Gil, A.; Koets, A.P.; Rutten, V.P.; Juste, R.A.;Estonba, A. Identification of single nucleotide polymorphisms in the bovine solute carrier family 11 member 1 (SLC11A1) gene and their association with infection by Mycobacterium avium subspecies paratuberculosis. J Dairy Sci. 2010,93,1713-21. [CrossRef]
- Korou, L.M.; Liandris, E.; Gazouli, M.;Ikonomopoulos, J. Investigation of the association of the SLC11A1 gene with resistance/sensitivity of goats (Capra hircus) to paratuberculosis. . Vet Microbiol. 2010,144,353-58. [CrossRef]
- Purdie, A.C.; Plain, K.M.; Begg, D.J.; de Silva, K.;Whittington, R.J. Candidate gene and genome-wide association studies of Mycobacterium avium subsp. paratuberculosis infection in cattle and sheep: a review. Comp Immunol Microbiol Infect Dis. 2011,34,197-208. [CrossRef]
- Roupie, V.; Rosseels, V.; Piersoel, V.; Zinniel, D.K.; Barletta, R.G.;Huygen, K. Genetic resistance of mice to Mycobacterium paratuberculosis is influenced by Slc11a1 at the early but not at the late stage of infection. Infect Immun. 2008,76,2099-105. [CrossRef]
- Ates, O.; Dalyan, L.; Müsellim, B.; Hatemi, G.; Türker, H.; Ongen, G.; Hamuryudan, V.;Topal-Sarikaya, A. NRAMP1 (SLC11A1) gene polymorphisms that correlate with autoimmune versus infectious disease susceptibility in tuberculosis and rheumatoid arthritis. Int J Immunogenet. 2009,36,15-19. [CrossRef] [PubMed]
- Sechi, L.A.; Gazouli, M.; Sieswerda, L.E.; Molicotti, P.; Ahmed, N.; Ikonomopoulos, J.; Scanu, A.M.; Paccagnini, D.;Zanetti, S. Relationship between Crohn’s disease, infection with Mycobacterium avium subspecies paratuberculosis and SLC11A1 gene polymorphisms in Sardinian patients. World J Gastroenterol. 2006,12,7161-64. [CrossRef]
- Kotlowski, R.; Bernstein, C.N.; Silverberg, M.S.;Krause, D.O. Population-based case-control study of alpha 1-antitrypsin and SLC11A1 in Crohn’s disease and ulcerative colitis. Inflamm Bowel Dis. 2008,14,1112-17. [CrossRef]
- Kotze, M.J.; de Villiers, J.N.; Rooney, R.N.; Grobbelaar, J.J.; Mansvelt, E.P.; Bouwens, C.S.; Carr, J.; Stander, I.;du Plessis, L. Analysis of the NRAMP1 gene implicated in iron transport: association with multiple sclerosis and age effects. Blood Cells Mol Dis. 2001,27,44-53. [CrossRef]
- Takahashi, K.; Satoh, J.; Kojima, Y.; Negoro, K.; Hirai, M.; Hinokio, Y.; Kinouchi, Y.; Suzuki, S.; Matsuura, N.; Shimosegawa, T.; et al. Promoter polymorphism of SLC11A1 (formerly NRAMP1) confers susceptibility to autoimmune type 1 diabetes mellitus in Japanese. Tissue Antigens. 2004,63,231-36. [CrossRef]
- Malkova, A.; Starshinova, A.; Zinchenko, Y.; Basantsova, N.; Mayevskaya, V.; Yablonskiy, P.;Shoenfeld, Y. The opposite effect of human leukocyte antigen genotypes in sarcoidosis and tuberculosis: a narrative review of the literature. ERJ Open Res. 2020,6,00155-2020. [CrossRef]
- Grunewald, J.; Spagnolo, P.; Wahlström, J.;Eklund, A. Immunogenetics of Disease-Causing Inflammation in Sarcoidosis. Clin Rev Allergy Immunol. 2015,49,19-35. [CrossRef]
- Saltini, C.; Pallante, M.; Puxeddu, E.; Contini, S.; Voorter, C.E.; Drent, M.;Amicosante, M. M. avium binding to HLA-DR expressed alleles in silico: a model of phenotypic susceptibility to sarcoidosis. . Sarcoidosis Vasc Diffuse Lung Dis. 2008,25,100-16.
- Rossman, M.D.; Thompson, B.; Frederick, M.; Iannuzzi, M.C.; Rybicki, B.A.; Pander, J.P.; Newman, L.S.; Rose, C.; Magira, E.;Monos, D. ACCESS Group. HLA and environmental interactions in sarcoidosis. Sarcoidosis Vasc Diffuse Lung Dis. 2008,25,125-32.
- Rossman, M.D.; Thompson, B.; Frederick, M.; Maliarik, M.; Iannuzzi, M.C.; Rybicki, B.A.; Pandey, J.P.; Newman, L.S.; Magira, E.; Beznik-Cizman, B.; et al. ACCESS Group. HLA-DRB1*1101: a significant risk factor for sarcoidosis in blacks and whites. Am J Hum Genet. 2003,73,720-35. [CrossRef]
- Grunewald, J.;Eklund, A. Löfgren’s syndrome: human leukocyte antigen strongly influences the disease course. . Am J Respir Crit Care Med. 2009,179,307-12. [CrossRef]
- Garman, L.; Pezant, N.; Pastori, A.; Savoy, K.A.; Li, C.; Levin, A.M.; Iannuzzi, M.C.; Rybicki, B.A.; Adrianto, I.;Montgomery, C.G. Genome-Wide Association Study of Ocular Sarcoidosis Confirms HLA Associations and Implicates Barrier Function and Autoimmunity in African Americans. Ocul Immunol Inflamm. 2021,29,244-49. [CrossRef]
- Oswald-Richter, K.; Sato, H.; Hajizadeh, R.; Shepherd, B.E.; Sidney, J.; Sette, A.; Newman, L.S.;Drake, W.P. Mycobacterial ESAT-6 and katG are recognized by sarcoidosis CD4+ T cells when presented by the American sarcoidosis susceptibility allele, DRB1*1101. J Clin Immunol. 2010,30,157-66. [CrossRef]
- Dawkins, B.A.; Garman, L.; Cejda, N.; Pezant, N.; Rasmussen, A.; Rybicki, B.A.; Levin, A.M.; Benchek, P.; Seshadri, C.; Mayanja-Kizza, H.; et al. Novel HLA associations with outcomes of Mycobacterium tuberculosis exposure and sarcoidosis in individuals of African ancestry using nearest-neighbor feature selection. Genet Epidemiol. 2022,46,463-74. [CrossRef]
- Grosser, M.; Luther, T.; Fuessel, M.; Bickhardt, J.; Magdolen, V.;Baretton, G. Clinical course of sarcoidosis in dependence on HLA-DRB1 allele frequencies, inflammatory markers, and the presence of M. tuberculosis DNA fragments. Sarcoidosis Vasc Diffuse Lung Dis. 2005,22,66-74.
- Inohara, N.; Ogura, Y.; Fontalba, A.; Gutierrez, O.; Pons, F.; Crespo, J.; Fukase, K.; Inamura, S.; Kusumoto, S.; Hashimoto, M.; et al. Host recognition of bacterial muramyl dipeptide mediated through NOD2. Implications for Crohn’s disease. J Biol Chem. 2003,278,5509-12. [CrossRef]
- Hugot, J.P.; Chamaillard, M.; Zouali, H.; Lesage, S.; Cézard, J.P.; Belaiche, J.; Almer, S.; Tysk, C.; O’Morain, C.A.; Gassull, M.; et al. Association of NOD2 leucine-rich repeat variants with susceptibility to Crohn’s disease. . Nature. 2001,411,599-603. [CrossRef]
- Miceli-Richard, C.; Lesage, S.; Rybojad, M.; Prieur, A.M.; Manouvrier-Hanu, S.; Häfner, R.; Chamaillard, M.; Zouali, H.; Thomas, G.;Hugot, J.P. CARD15 mutations in Blau syndrome. Nat Genet. 2001,29,19-20. [CrossRef]
- Hampe, J.; Grebe, J.; Nikolaus, S.; Solberg, C.; Croucher, P.J.; Mascheretti, S.; Jahnsen, J.; Moum, B.; Klump, B.; Krawczak, M.; et al. Association of NOD2 (CARD 15) genotype with clinical course of Crohn’s disease: a cohort study. Lancet. 2002,359,1661-65. [CrossRef]
- Wang, X.; Kuivaniemi, H.; Bonavita, G.; Mutkus, L.; Mau, U.; Blau, E.; Inohara, N.; Nunez, G.; Tromp, G.;Williams, C.J. CARD15 mutations in familial granulomatosis syndromes: a study of the original Blau syndrome kindred and other families with large-vessel arteritis and cranial neuropathy. Arthritis Rheum. 2002,46,3041-45. [CrossRef]
- Lesage, S.; Zouali, H.; Cézard, J.P.; Colombel, J.F.; Belaiche, J.; Almer, S.; Tysk, C.; O’Morain, C.; Gassull, M.; Binder, V.; et al. EPWG-IBD Group; EPIMAD Group; GETAID Group. CARD15/NOD2 mutational analysis and genotype-phenotype correlation in 612 patients with inflammatory bowel disease. Am J Hum Genet. 2002,70,845-57. [CrossRef]
- Dow, C.T.;Ellingson, J.L. Detection of Mycobacterium avium ss. Paratuberculosis in Blau Syndrome Tissues. . Autoimmune Dis. 2010,2011,127692.
- Caso, F.; Galozzi, P.; Costa, L.; Sfriso, P.; Cantarini, L.;Punzi, L. Autoinflammatory granulomatous diseases: from Blau syndrome and early-onset sarcoidosis to NOD2-mediated disease and Crohn’s disease. RMD Open. 2015,1,e000097. [CrossRef]
- Takeuchi, Y.; Shigemura, T.; Kobayashi, N.; Kaneko, N.; Iwasaki, T.; Minami, K.; Kobayashi, K.; Masumoto, J.;Agematsu, K. Early diagnosis of early-onset sarcoidosis: a case report with functional analysis and review of the literature. Clin Rheumatol. 2017,36,1189-96. [CrossRef]
- Matsui, Y.; Iwai, K.; Tachibana, T.; Fruie, T.; Shigematsu, N.; Izumi, T.; Homma, A.H.; Mikami, R.; Hongo, O.; Hiraga, Y.; et al. Clinicopathological study of fatal myocardial sarcoidosis. Ann N Y Acad Sci. 1976,278,455-69. [CrossRef]
- Roberts, W.C.; McAllister, H.A., Jr.;Ferrans, V.J. Sarcoidosis of the heart. A clinicopathologic study of 35 necropsy patients (group 1) and review of 78 previously described necropsy patients (group 11). Am J Med. 1977,63,86-108. [CrossRef]
- Silverman, K.J.; Hutchins, G.M.;Bulkley, B.H. Cardiac sarcoid: a clinicopathologic study of 84 unselected patients with systemic sarcoidosis. Circulation. 1978,58,1204-11. [CrossRef]
- Dubrey, S.W.;Falk, R.H. Diagnosis and management of cardiac sarcoidosis. Prog Cardiovasc Dis. 2010,52,336-46. [CrossRef]
- Kandolin, R.; Lehtonen, J.; Graner, M.; Schildt, J.; Salmenkivi, K.; Kivistö, S.M.;Kupari, M. Diagnosing isolated cardiac sarcoidosis. . J Intern Med. 2011,270,461-68. [CrossRef]
- Dow, C.T.;Collins, M.T. Detection of M. paratuberculosis in Sarcoisosis Cardiac Tissue. 3rd International WASOG Conference on Diffuse Lung Diseases. Catania, ItalyJune 21, 2006.
- Celler, B.G. Case Study: Cardiac sarcoidosis resolved with Mycobacterium avium paratuberculosis antibiotics (MAP). Sarcoidosis Vasc Diffuse Lung Dis. 2018,35,171-77. [CrossRef]
- Borody, T.J.; Leis, S.; Warren, E.F.; Surace, R. Treatment of severe Crohn’s disease using antimycobacterial triple therapy--approaching a cure? Dig Liver Dis. 2002,34,29-38. [CrossRef]
- Borody, T.J.; Bilkey, S.; Wettstein, A.R.; Leis, S.; Pang, G.; Tye, S. Anti-mycobacterial therapy in Crohn’s disease heals mucosa with longitudinal scars. Dig Liver Dis. 2007,39,438-44. [CrossRef]
- Alcedo, K.P.; Thanigachalam, S.; Naser, S.A. RHB-104 triple antibiotics combination in culture is bactericidal and should be effective for treatment of Crohn’s disease associated with Mycobacterium paratuberculosis. Gut Pathog. 2016, 8, 32. [Google Scholar] [CrossRef]
- Agrawal, G.; Clancy, A.; Huynh, R.; Borody, T. Profound remission in Crohn’s disease requiring no further treatment for 3-23 years: a case series. Gut Pathog. 2020, 12, 16. [Google Scholar] [CrossRef]
- Kuenstner, J.T.; Naser, S.; Chamberlin, W.; Borody, T.; Graham, D.Y.; McNees, A.; Hermon-Taylor, J.; Hermon-Taylor, A.; Dow, C.T.; Thayer, W.; et al. The Consensus from the Mycobacterium avium ssp. paratuberculosis (MAP) Conference 2017. Front Public Health, 2017,5,208. [CrossRef]
- Johnson, S. Who Moved My Cheese? Putnam Adult; 1998.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



