Submitted:
28 February 2023
Posted:
01 March 2023
You are already at the latest version
Abstract
The frequency of non-alcoholic fatty liver disease (NAFLD) has exacerbated setting diagnostic challenges, which increases the need for reliable non-invasive diagnostic tools. Due to the importance of gut-liver axis in the progression of NAFLD, studies try to reveal microbial signatures in NAFLD, evaluate them as diagnostic biomarkers and to predict the disease progression. The gut microbiome affects human physiology by processing the ingested food to bioactive metabolites. These molecules can penetrate the portal vein and the liver to promote or prevent hepatic fat accumulation. Here findings of human fecal metagenomic and metabolomic studies in relation to NAFLD are reviewed. The studies present mostly distinct and even contradictory findings on microbial metabolites and functional genes in NAFLD. The most reproducing microbial biomarkers are increased lipopolysaccharides and peptidoglycan biosynthesis, enhanced degradation of lysine, increased levels of branched chain amino acids as well as altered lipid and carbohydrate metabolism. Among other causes, the discrepancies between the studies may be related to the obesity status of the patients and severity of NAFLD. In none of the studies except one, diet was considered, though it is an important factor driving the gut microbiota metabolism. The future studies should consider diet in the analyses.
Keywords:
gut microbiota
; metabolomics
; metagenomics
; liver fat
; NAFLD
; diet
; metabolic pathways
2 Institute of Biomedicine, Medical Microbiology and Immunology, University of Turku, Turku, Finland
1. Introduction
WHO estimates that worldwide over 1.9 billion adults are overweight and consequently, the frequency of metabolic disorders, including non-alcoholic fatty liver disease (NAFLD) has exacerbated. Metabolic disorders are a cluster of physiological conditions: increased blood pressure, high blood sugar, excess body fat around the waist, and abnormal cholesterol or triglyceride levels that often occur together. Alarmingly, these diseases now also extend to children in addition to young and middle-aged population [1]. In Western countries, up to 90% of obese population are estimated to suffer from NAFLD [2]. In Nordic countries, NAFLD is the second most increasing indication for liver transplantation, and thus, a great burden to the healthcare system. Therefore, new diagnostic tools allowing early detection of the disease would be of great importance.
NAFLD is defined as excessive fat accumulation (over 5% fat in hepatocytes) in the liver without secondary causes of fat accumulation such as excessive drinking of alcohol and treatment with steatogenic drugs (e.g., methotrexate). Histopathologically, NAFLD can be categorized into simple steatosis (non-alcoholic fatty liver, NAFL), which is diagnosed as a presence of hepatic fat accumulation without any histological or biochemical injuries, and non-alcoholic steatohepatitis (NASH), which is characterized by steatosis, inflammation, and hepatocyte damage, i.e., ballooning, and can be accompanied with cirrhosis or not [3,4]. It is estimated that 3 to 5% of NAFLD patients can develop NASH [5].
The high prevalence of NAFLD sets diagnostic challenges, and there is a growing need for reliable non-invasive diagnostic tools. Due to the importance of gut-liver axis in the onset and progression of NAFLD [6], several recent studies have attempted to reveal the microbial signatures in NAFLD, evaluate their suitability as diagnostic biomarkers of NAFLD and to predict the progression of the disease. This article reviews the evidence on microbial signatures in NAFLD that have been analyzed using fecal metagenomics and metabolomics. Metagenomics refers to the analysis of gut microbiota composition and functional genes using shotgun sequencing. Metabolomics refers to quantification of the fecal metabolites using either nuclear magnetic resonance (1H-NMR) or ultra-high performance liquid chromatography (LC)/mass spectrophotometry (MS). Ultimately, pitfalls and caveats in such approaches will be summarized as well as avenues for future direction.
The literature searches for this review article were made between September and December 2022. The search words “fecal AND metabolomics AND (liver fat OR NAFLD OR NASH)” and “fecal AND metagenomics AND (liver fat OR NAFLD OR NASH)” were used both in PubMed and Ovid Medline. Studies in animals were omitted from the search results.
2. Non-Invasive, Cost-Effective, and Easy Diagnostics of NAFLD for Clinical Settings
Due to that liver biopsies are highly invasive, development of cost-effective diagnostic tools for NAFLD is important. What makes the early diagnosis difficult is that before the onset of severe fibrosis or cirrhosis, the NAFLD patients may remain asymptomatic. Therefore, the diagnosis is often made coincidentally due to abnormal findings on routine blood samples. Values above the upper limit of normal serum alanine aminotransferase (ALT, ~40 IU/l in men and ~30 IU/l in women) as well as abnormally high serum triglycerides and LDL cholesterol can be an indication of NAFLD [7]. Yet, according to some studies, the liver enzymes may be completely normal in most patients [8].
At present, there are several available non-invasive methods to diagnose NAFLD, which however, do not enable distinguishing steatosis from steatohepatitis or evaluate the severity of hepatic fibrosis. In this context, several panel markers, indexes, and scores have been developed for the diagnostics. For instance, the liver fat score includes as measured variables the presence of metabolic syndrome and type 2 diabetes (T2D), fasting serum insulin, serum aspartate aminotransferase (AST), and the AST/ALT ratio [9]. The fatty liver index (FLI) considers body mass index (BMI), waist circumference, serum triglyceride levels, and gamma-glutamyltransferase (GGT) in the general population with low prevalence of T2D [10]. To distinguish NASH from NAFL, the HAIR score, which includes determination of the presence of hypertension, elevated ALT, and insulin resistance has been employed [11]. An advanced fibrosis scoring system has also been developed and validated by McPherson and co-workers that is generated by determining age, hyperglycemia, BMI, platelet count, serum albumin, and AST/ALT ratio [12].
However, measuring variables of blood and body composition is insufficient for the ultimate diagnosis and thus, imaging techniques and other analyses are needed. While the ultrasonography is the most cost-effective and suitable in clinical practice it may fail to detect mild steatosis. One study showed that ultrasound was unable to detect steatosis present in less than 10% of hepatocytes [13]. In addition, the visual assessment of NAFLD by ultrasonography has significant substantial inter-observer variability, which makes the reproducibility of the results limited [14].
3. Physiological and Molecular Players in the Onset of NAFLD–Future for Diagnostics?
It is now well accepted that the pathogenesis of NAFLD involves multiple simultaneous ‘hits’ that are associated with environmental, host genetics and physiologic factors [15] rather than initially proposed by the “two-hit theory” [16]. The multiple hit hypothesis implies that simple hepatic steatosis may be a benign process and NASH might be a separate disease with a different pathogenesis. The pathogenic hits include: 1) inflammatory mediators derived from various tissues [17,18]; 2) increased lipid storage, lipogenesis, and (adipo)cytokines that activate endoplasmic reticulum stress [19,20,21]; 3) mitochondrial dysfunction [21] and reactive oxygen species due to lipotoxicity [22,23]; 4) nutrient sensing [24]; and 5) genetic factors [25,26,27,28]. Importantly, all these events may occur together rather than consecutively. Recent studies have also highlighted the importance of connective tissue dysfunction and insulin resistance in the onset of NAFLD [29,30,31]. However, if these above-described factors would be used for diagnostic purposes, they would all need invasive sampling and thus non-invasive alternatives are of interest.
Almost a decade ago, we [32] and Mouzaki et al. [33] were one of the pioneers in showing that the gut microbiota composition associates with hepatic fat content in humans. There are excellent, recent reviews on the topic [34], and thus this review will concentrate on the advances of fecal metabolomics and metagenomics related to NAFLD. It is increasingly accepted that the abundance and functions of many members of the gut microbiota affect the human physiology by processing the ingested food to certain bioactive metabolites [35]. These molecules can act as inter-tissue signaling messengers by penetrating the portal vein and further, the liver to promote or prevent hepatic fat accumulation (Figure 1). Collecting fecal samples for gut microbiota composition and microbial metabolite analyses would be an excellent non-invasive way for NAFLD diagnostics, which will be reviewed in the upcoming chapters. While this review concentrates on the human gut microbiota metabolism (fecal metabolomics and metagenomics) and not animal studies, one important host molecular mechanism that connect the gut microbiota and their metabolism to NAFLD is briefly introduced below as an example of animal models.
One mechanistic animal study showed that gut microbiota-dependent hepatic lipogenesis was mediated by hepatic stearoyl CoA desaturase 1 (SCD1) [36]. The authors used germ-free and conventional Toll-like receptor 5 (TLR5) deficient (T5KO) mice that are prone to develop microbiota-dependent metabolic syndrome to show first that the T5KO mice displayed elevated hepatic neutral lipid content depending on the presence of gut microbiota. TLR5 recognizes flagellin, which is the structural protein of the locomotive organelle of bacteria [37]. After the initial observations, Singh and co-workers found that colonic short-chain fatty acids (SCFA) receptors as well as hepatic lipogenic enzymes including SCD1 were upregulated in T5KO mice and that gut derived SCFA were increasingly incorporated into palmitate in liver. Dietary SCFA further aggravated hepatic steatosis and metabolic syndrome that were impeded by hepatic deletion of SCD1. All the above-mentioned effects were ablated in the germ-free mice, but when the germ-free mice were transplanted with the cecal microbiota of T5KO mice, their hepatic palmitate content doubled. The authors concluded that while several beneficial properties have been recognized for SCFA, their excess in conditions combined with innate immune deficiency and dysbiotic, over-grown microbiota due to T5KO may increase susceptibility to metabolic diseases [36].
4. Fecal Metabolomics and Metagenomics Identifying the Microbial Signatures in NAFLD
The systems biological effects of gut microbial metabolism are excellently reviewed elsewhere [35], and therefore, in this review the emphasis will be in relation to NAFLD. It is likely that the severity of NAFLD affects differently the functions of the gut microbiota and vice versa. Therefore, the reviewed studies are divided below in subsections by the disease severity (steatotic without diagnosed NAFLD, NAFL, NASH, fibrosis, cirrhosis, hepatocellular carcinoma). Ultimately, also studies in children with NAFLD are reviewed. The reviewed studies are listed in Table 1, which is primarily divided by the sections below, and by the order of appearance of the references in the text.
4.1. Fecal Metabolomics and Metagenomics Identifying the Gut Microbial Signatures in Steatotic Adults without Diagnosed NAFLD
The largest metagenomic study conducted by far in fatty liver disease included a sample of 6,269 Finnish participants of which most were overweight, but according to the BMI there were also normal weight individuals included [38]. It should be noted that the functional metagenomic profiling was qualitative and not quantitative because the sequencing depth did not allow assembling contigs. In addition, instead of measuring liver fat content with imaging the authors used FLI to categorize the participants into liver fat content groups. Nevertheless, FLI is rather widely used and accepted index for NAFLD and its’ stratification [39]. Ethanol and the SCFA acetate production pathways were found to be positively associate with FLI [38]. Previously, it has been shown, for instance, that high-alcohol-producing strains of Klebsiella pneumoniae exist in humans with NAFLD [40]. Thus, it seems that endogenously produced alcohol may play a role in hepatic fat accumulation at least in some populations. Generally, SCFA are considered to exert beneficial functions in the host, such as modulating immune functions [41] and gastrointestinal permeability [42]. However, they may also negatively impact inflammatory status of the host [36].

Table 1.
The reviewed studies presenting metabolomic and/or metagenomic findings on the human gut microbiota and NAFLD.
Table 1.
The reviewed studies presenting metabolomic and/or metagenomic findings on the human gut microbiota and NAFLD.
| Reference Number in the Text |
Analysis Method | Study Patients/Adults |
|---|---|---|
| Ruuskanen et al. [38] | metagenomics | fatty liver & all normal weight, overweight or obese |
| Hoyles et al. [43] | metagenomics | fatty liver, morbidly obese |
| Driuchina et al. [47] | metabolomics | fatty liver & healthy, obese |
| Ge at al. [53] | metabolomics | NAFLD & healthy, overweight |
| Boursier et al. [57] | predicted metagenomics | NAFLD & NAFLD + fibrosis & NASH |
| Loomba et al. [58] | metagenomics | NAFLD + fibrosis |
| Lee et al. [61] | metabolomics | NAFLD + obesity + fibrosis & NAFLD + normal weight + fibrosis |
| Smirnova et al. [63] | metabolomics | NAFL, NASH, fibrosis regardless of BMI |
| Sui et al. | metabolomics | NASH & healthy, normal weight |
| Behary et al. [67] | metabolomics, metagenomics | NAFLD-HCC & healthy |
| Oh et al. [68] | metabolomics, metagenomics | NAFLD-cirrhosis & healthy |
| Reference number in the text |
Analysis method | Study patients/children |
| Michail et al. [49] | metabolomics, metagenomics | NAFLD + obese, healthy + obese & healthy + normal weight |
| Testerman et al. [71] | metagenomics | NAFLD & healthy, obese |
| Zhao et al. [72] | metagenomics | NAFLD & healthy, obese & normal weight |
| Kordy et al. [60] | metabolomics, metagenomics | NAFL, NASH & healthy, obese & normal weight |
| Yu et al. [73] | metabolomics, metagenomics | NAFLD & healthy, normal weight |
| Schwimmer et al. [74] | metagenomics | NAFL, NASH & healthy, overweight |
Fecal metagenomic signatures have been described in non-diabetic morbidly obese women with hepatic steatosis [43]. By mapping the microbial genes into Kyoto Encyclopedia of Genes and Genomes (KEGG) modules, it was found that steatosis associated positively with carbohydrate, lipid and amino acid metabolism as well as lipopolysaccharide (LPS) and peptidoglycan biosynthesis. LPS are recognized by Toll-like receptor 4 (TLR4), which expression has been shown to be increased in the liver of obese patients with NASH [44]. Peptidoglycan, in turn, is recognized by multiple pattern-recognition receptors, including nucleotide-binding oligomerization domain-containing proteins (NODs), domain-containing 3 (NLRP3) and Toll-like receptor 2 (TLR2) [45]. In addition to the above-mentioned pathways, Hoyles et al. also observed that hepatic steatosis was associated with increased number of genes related to the biosynthesis of branched-chain amino acids (BCAA) and aromatic amino acids [43]. The metagenomic findings of feces were supported further by the elevated concentrations of these particular amino acids in plasma. To show mechanistically that the microbiome contributes to the hepatic steatosis, the authors transplanted feces from steatotic human donors to mice. Indeed, the fecal transplants caused rapid hepatic fat accumulation in mice, which involved elevated circulating BCAA [43]. These observations are interesting as numerous studies have linked BCAA to obesity and NAFLD (for review [46]).
We recently compared fecal and plasma metabolomes of humans with low (<5% of fat in liver) and high (>5% of fat in liver) liver fat content without diagnosed NAFLD [47]. The study groups did not differ from each other in BMI. We found that the fecal histidine metabolism product, N-omega-acetylhistamine was markedly increased in individuals with fatty liver. In addition, another product of histidine degradation, anserine, positively associated with liver fat content [47]. Previously, plasma levels of histidine have been shown to associate with the grade of hepatic steatosis [48]. In agreement with a previous metagenome study [49], we found that the levels of lysine degradation product, saccharopine, were higher in the feces of individuals with high liver fat content [47]. As an indication of decreased steroid metabolism, the fecal levels of 6-hydroxybetatestosterone were reduced in the steatotic individuals. Contrarily, a previous study reported that low serum testosterone levels were associated with hepatic steatosis in obese male [50].
To summarize, the two metagenomic studies in individuals with hepatic steatosis show different microbial signatures, which may be due to the different obesity status of the study populations. Neither of the metagenomic studies determined dietary factors between the groups or used them as confounding factors [38,43]. In our metabolomic study, there were no major dietary differences between the high and low liver fat content groups, except that higher vitamin E and sucrose intake was observed in individuals with fatty liver [47]. Interestingly, sucrose is known to contribute to the onset of NAFLD [51], while vitamin E is considered as a possible treatment for NAFLD [52]. The main findings of the three reviewed studies in this section are presented in Figure 2.
4.2. Fecal Metabolomics and Metagenomics Identifying the Gut Microbial Signatures in Patients with NAFL, NASH and Hepatic Fibrosis
Ge and co-workers have studied fecal metabolomics in patients with NAFLD and healthy controls using UHPLC/MS/MS [53]. All participants were overweight and/or had visceral obesity. Combining the metabolite identification with KEGG pathway analysis, the authors found that the metabolism of nicotinate, nicotinamide and pyrimidine as well as signaling pathways of calcium and oxytocin and pancreatic secretion were altered in NAFLD patients compared to healthy controls. Interestingly, nicotinamide adenine dinucleotide (NAD+) is being studied in clinical trials as a potential target to treat NAFLD [54]. However, Ge et al. failed to show differences in the fecal levels of nicotinate between the healthy and NAFLD groups [53]. Altered pyrimidine metabolism was reflected in lower fecal levels of uracil in the NAFLD patients. In addition, the catabolic byproduct of purine metabolism, xanthine was also lower likely due to the lower enzymatic activity of xanthine oxidase. The authors concluded that xanthine together with the abundance of specific microbial taxa might serve in the diagnostics of NAFLD [53]. Intriguingly, it has been shown that inhibition of xanthine oxidase can ameliorate hepatic steatosis in mice [55]. This might be related to decreased production of reactive oxygen species (ROS) [56].
Based on literature, it seems that microbial functional genes can differentiate less advanced NAFLD from NASH and NAFLD with significant fibrosis [57] and the stage of fibrosis [58]. Yet, the authors of one paper did not perform shotgun metagenome sequencing but predicted the functional potential of the gut microbiota from 16S rRNA gene data using PICRUSt [57]. The name is an abbreviation for Phylogenetic Investigation of Communities by Reconstruction of Unobserved States. PICRUSt is a bioinformatics software package designed to predict metagenome functional content from marker gene (e.g., 16S rRNA gene) surveys and full genomes using a developed algorithm developed by Languille et al. [59]. However, the enriched functional categories in NASH were mostly related to carbohydrate and lipid metabolism [57], which is similar to what has been found in simple steatosis [43] and in NASH [60] by others. Interestingly, patients with fibrosis could be distinguished from the patients without fibrosis by their microbial functional genes. Fibrosis was associated with enriched functional categories related to carbohydrate and lipid metabolism [57]. A study that combined real microbial metagenomics and analysis of plasma metabolites in obese NAFLD patients [58] did not find similar microbial signatures in fibrosis as PICRUSt did [57]. Of the set of 89 metabolites that are produced both by the host and the microbiota, eleven metabolites could differentiate between mild/moderate NAFLD and advanced fibrosis. These enriched genes/metabolites were mainly related to carbon metabolism in fibrosis grade 2 as well as to nucleotide and steroid degradation in fibrosis grade 1. In addition, higher SCFA butyrate were annotated in grade 1, while the SCFA butyrate and propionate were higher in grade 2. Many findings from plasma metabolites and fecal metagenomic pathways supported each other [58]. Contrary to these findings, we have found using fecal metabolomics that testosterone metabolism in the gut was lower in obese individuals with fatty liver [47].
In addition to the stage of hepatic fibrosis, obesity status of the patients appears to be linked to the distinct gut microbial features in NAFLD [61]. Lee and co-workers reported that while not seen in obese individuals with NAFLD, the levels of SCFA acetate and propionate increased along with the stage of fibrosis in non-obese individuals [61]. This again indicates that the SCFA are not necessarily always beneficial for the host’s health. Lee et al. further found that the fecal levels of several conjugated and unconjugated bile acids were higher in non-obese individuals with fibrosis, while in obese individuals the levels of total conjugated bile acids inversely associated with the severity of fibrosis [61]. Supporting the findings of fecal metabolomics, the expression levels of microbial genes encoding for bile salt hydrolase and 7α-hydroxy-3-oxochol-4-en-24-oyl-CoA dehydrogenase were lower in the non-obese patients with fibrosis.
The contribution of the bile acids to the pathophysiology of NAFLD via gut-liver axis is rather well established, yet the bile acids have been mostly analyzed in plasma and not in feces in NAFLD studies [62]. However, contrary to the findings of Lee et al. [61] reviewed above, Smirnova and co-workers repor that the fecal concentrations of several secondary bile acids were lower in obese NASH patients with advanced fibrosis [63]. Yet, there is a difference between the two studies in the obesity status of the patients. In the study by Smirnova et al. especially taurine conjugated bile acids increased along the stage of fibrosis [63]. Interestingly, when the authors looked at the NAFLD activity scores (NAS), the findings were opposite. Compared to the healthy controls, the NAFLD patients had higher fecal levels of secondary bile acids and expression of microbial genes involved in biotransformation of bile acids. Further, NASH patients had higher levels of conjugated bile acids than patients with NAFL [63]. This, in turn, is contrary to another study by Sui et al. showing higher levels of primary bile acids, chenodeoxycholic acid and cholic acid, in the feces of non-diabetic individuals with NASH compared to healthy controls [64]. Nevertheless, these patients were normal weight, and therefore, the obesity status may be related to the different results between the reviewed studies. Of the other fecal metabolites, increased kynurenine and decreased L-tryptophan levels were good predictors of hepatic steatosis [64]. The latter finding is interesting as a previous study had shown that tryptophan-derived metabolite kynurenine causes hepatic steatosis in mice by activating Aryl hydrocarbon receptor signaling [65].
To summarize, while fecal bile acids seem to play an important role in NAFLD and fibrosis, the reviewed studies present different findings, which may, at least partly, depend on the obesity status of the study populations. None of the studies reported whether there were differences in dietary intakes between the study groups, though one study [64] had collected food consumption data. The main findings on the microbial metagenomes and fecal metabolomics of the studies reviewed in this section are presented in Figure 3.
4.3. Fecal Metabolomics and Metagenomics Identifying the Gut Microbial Signatures in Patients with NAFLD-Cirrhosis and NAFLD-Hepatocellular Carcinoma
With the global rise in the incidence of obesity and type 2 diabetes, NAFLD-related hepatocellular carcinoma (HCC) and NAFLD-cirrhosis are also becoming most common liver diseases [66]. By using metagenomics, Behary et al. show that the gut microbiome of the NAFLD-HCC patients was characterized by higher number of SCFA synthesizing genes [67], which is in line with what was found related to an earlier stage of the disease, namely higher FLI [38]. Compared to the NAFLD-cirrhosis patients and healthy controls, in the feces of the NAFLD-HCC patients genes related to acetate (phosphate acetyltransferase, pta), butyrate (phosphate butyryltransferase, ptb) and propionate (fumarate reductase, frd and succinate-CoA synthetase, scs) synthesis were over-expressed [67]. The gene expression findings were further confirmed using targeted LC/MS/MS and 1H-NMR quantification of the fecal metabolites. The levels of acetate and butyrate as well as oxaloacetate and acetylphosphate that are intermediates of the SCFA metabolism, were higher in the feces of the NAFLD-HCC patients, while propionate concentration did not differ between the study groups [67]. In the future, it would be interesting to investigate, which other metabolites characterize the NAFLD-HCC patients using a non-targeted metabolomic profiling.
In NAFLD-cirrhosis, the gut microbial signatures have been studied by Oh and co-workers [68]. Interestingly, similar to what has been shown in steatosis without diagnosed NAFLD [43], BCAA and aromatic amino acids were predictors of cirrhosis as shown both by metagenomic and metabolomic analyses [68]. In the NAFLD-cirrhotic patient’s fecal samples, the levels of L-tryptophan were increased due to decreased metabolism of it. This is contrary to another study showing that decreased L-tryptophan levels were good predictors of hepatic steatosis [64].
To summarize the literature reviewed above, in NAFLD-associated cirrhosis and HCC some similar metabolic signatures can be found as in hepatic steatosis without diagnosed NAFLD, namely higher fecal levels of SCFA, BCAA and aromatic amino acids. Thus, in the future it would be important to explore their role more, and to consider whether these fecal metabolites would be suitable early biomarkers of advanced liver diseases before its’ onset. It should be noted, however, as in the other studies reviewed above that none of the studies in this section considered dietary factors as confounding between the study groups.
4.4. Fecal Metagenomics and Metabolomics Identifying the Gut Microbial Signatures in Children with NAFLD
A multi-omics study by Michail et al. combined metagenomic, metabolomic and proteomic approaches to study the gut microbiome of obese children with and without NAFLD [49]. On a contrary to one targeted metabolomics study showing higher fecal acetate and propionate levels in NAFLD patients [69], the 1H-NMR quantification of the fecal metabolites revealed that acetate was lower in pediatric NAFLD patients, while propionate was unaffected [49]. As reviewed above, supporting again the role of endogenously produced ethanol in NAFLD, the fecal levels of ethanol were ~2-fold higher in obese with NAFLD than in the healthy children and obese children without NAFLD. Based on the microbial metagenomic analysis, the lysine degradation pathway was exclusively identified in children with NAFLD and not in healthy individuals [49]. This agrees with our metabolomic findings from adults showing higher degradation products of lysine in the feces of individuals with high liver fat content [47]. The metagenome shotgun sequencing of Michail et al. suggested that the microbial energy metabolism, including fatty acid and carbohydrate biosynthesis is much more enhanced in NAFLD pediatric patients compared to healthy children [49]. However, this might be solely due to the different obesity status of the patients in the studies as it has been shown that an obese microbiome more efficiently harvests energy from the diet [70].
Testerman and co-workers studied the fecal metagenomes of children with and without NAFLD [71]. They observed that multiple pathways for lysine synthesis were increased, and histidine degradation was decreased in NAFLD patients. This is contrary to our recent metabolomics finding in adults with fatty liver showing higher fecal levels of lysine and histidine degradation products [47]. However, in the NAFLD patient’s metagenomes also microbial metabolic pathways for BCAA, aromatic amino acids and peptidoglycan synthesis were enriched [71]. Interestingly, similar findings have been reported in morbidly obese adults with hepatic steatosis [43]. Thus, it could be that these metabolic pathways might help in the detection of the disease at an early age and/or stage of the disease.
Another metagenomic study compared healthy children, obese children, and obese children with NAFLD [72]. Compared to healthy children, the obese pediatric patients with and without NAFLD had lower abundance of microbial genes related to pathways of replication and reparation, folding, sorting and metabolism of amino acids. The presence of NAFLD differentiated the obese groups showing enrichment of pathways related to digestive system, immune system, and glycan biosynthesis [72]. Thus, in this case there were microbial signatures in NAFLD that were likely not dependent on the obesity status of the patients.
In agreement with the findings of Hoyles and co-workers in adults [43], Kordy et al. report that compared to the healthy BMI-matched individuals, the microbiome of the pediatric patients with NASH was characterized by increased carbohydrate, lipid and amino acid metabolism as well as LPS biosynthesis [60]. While the panels of plasma metabolites could accurately predict NASH, this was not seen with fecal metabolites analyzed from rectal swabs. The author of this current review opinions that the rectal swabs, even if stored at correct temperatures, may not be a reliable way to preserve the fecal metabolites for the subsequent analyses.
A combination of fecal metagenomics and targeted UPLC-MS/MS identified alterations in bile acid metabolism in children with NAFLD [73]. The number of genes related to the biosynthesis of secondary bile acids were more abundant in NAFLD patients, which agrees with one study in adults [63]. However, the comparison between the metagenomic and metabolomics results is a bit confusing as the fecal levels many secondary bile acids (a-hyodeoxycholic acid, 7-ketolithocholic acid, 23-nordeoxycholic acid, 7,12-diketolithocholic acid, 3-epideoxycholic acid, and dehydrocholic acid) were reduced in NAFLD, while only chenodeoxycholic acid-3-b-D-glucuronide was increased [73]. Unfortunately, the authors did not discuss these discrepant results of their publication.
One study in children with NAFL and NASH used metagenomic shotgun sequencing to reveal microbial signatures related to the disease [74]. Compared to the BMI-matched healthy controls, microbial LPS biosynthesis was significantly enriched in children with NAFL and NASH. This finding agrees with what has been found in non-diabetic obese women with hepatic steatosis [43]. Several genes related to flagellar assembly were also enriched in children with NASH and in those with moderate-to-severe fibrosis [74]. We have previously shown in an a mouse model that gut microbial flagellin causes hepatic fat accumulation, which is mediated by Vascular adhesion protein-1 [75]. Altogether these results suggest that flagellin may promote NAFLD, while other studies have suggested that a knockout of flagellin-recognizing TLR5 protects mice from NAFLD [36]. However, it should be noted that flagellin can also act via cytosolic nucleotide oligomerization domain (NOD)-like receptors to affect the inflammatory status [76]. Thus, the effects of flagellin and its’ receptors in relation to the onset of NAFLD should be explored more in the future.
To summarize, studies in children with NAFLD have reported microbial signatures of which some are similar (increased LPS biosynthesis and amino acid metabolism) and some distinct from each other. However, a few of the reviewed findings are like those seen in adults, such as higher fecal levels of ethanol in individuals with hepatic steatosis and increased LPS biosynthesis in NAFLD. None of the reviewed studies in children analyzed whether there were dietary differences between the study groups. The main microbial metagenomic and fecal metabolomic findings from children with NAFLD are summarized in Figure 4.
5. Conclusions and Future Directions
Due to the increasing incidence of NAFLD there is a growing need for non-invasive diagnostic tools. Because of the importance of gut-liver-axis in the pathophysiology of NAFLD, there is a hope that signatures of microbial metabolism could be used for diagnostic purposes. However, studies on the microbial metagenomics and fecal metabolomics in humans with NAFLD are surprisingly scarce. The existing studies reviewed here present mostly distinct findings and even contradictory findings on the microbial metabolites and functional genes in NAFLD. The most reproducing markers reviewed here are increased LPS and peptidoglycan biosynthesis, enhanced degradation of lysine, increased levels of BCAA as well as altered lipid and carbohydrate metabolism. Among many other causes, the discrepancies between the studies may be related to the obesity status of the patients, ethnicity, and severity of the disease. Hence, to reliably identify microbial metabolites as potential diagnostic biomarkers in NAFLD, studies should be done repeatedly in cohorts with same characteristics and with larger number of patients.
One of the most important factors driving the metabolism of the gut microbes is the diet. Besides the influence of long-term dietary intakes, diet can also very rapidly change the microbiome [77]. It is also well known that hypercaloric diets promote the onset of NAFLD [78]. Therefore, it is surprising that besides our study [47], none of the other studies reviewed here compared dietary intakes between the study groups. Thus, it cannot be known whether the reported metabolic differences in the gut microbiota between the study groups are solely or partly due to dietary differences. In the future, the diet should be definitely considered in studies presenting results from fecal metagenomics and metabolomics so that the microbiome could enter in the possible future diagnostics of NAFLD. Interestingly, it seems that in undernourished NAFLD patients, altered bile acid signatures are consistently reported [79]. Bauer and co-workers nicely proposed how collaborative, multi-omics approaches could improve hepatic health in undernourished population. Henceforward, similar approaches should be considered also for over nourished Western populations suffering from NAFLD.
Funding
This study was funded by the Academy of Finland funded Profiling of University of Jyväskylä and the Faculty of Sport and Health Sciences: Physical activity through life span (PACTS2) and by the Academy of Finland grant to SP (grant ID 349264).
Acknowledgments
I would like to acknowledge, and I greatly appreciate any important papers in this field that may not have been cited here because the current article is not a systematic review.
Conflicts of Interest
The author declares no conflict of interest.
References
- Pereira, K.; Salsamendi, J.; Casillas, J. The Global Nonalcoholic Fatty Liver Disease Epidemic: What a Radiologist Needs to Know. J Clin Imaging Sci 2015, 5, 32. [Google Scholar] [CrossRef]
- Araujo, A.R.; Rosso, N.; Bedogni, G.; Tiribelli, C.; Bellentani, S. Global epidemiology of non-alcoholic fatty liver disease/non-alcoholic steatohepatitis: What we need in the future. Liver Int 2018, 38 Suppl 1, 47–51. [Google Scholar] [CrossRef]
- Wong, V.W.; Wong, G.L.; Choi, P.C.; Chan, A.W.; Li, M.K.; Chan, H.Y.; Chim, A.M.; Yu, J.; Sung, J.J.; Chan, H.L. Disease progression of non-alcoholic fatty liver disease: a prospective study with paired liver biopsies at 3 years. Gut 2010, 59, 969–974. [Google Scholar] [CrossRef] [PubMed]
- McPherson, S.; Hardy, T.; Henderson, E.; Burt, A.D.; Day, C.P.; Anstee, Q.M. Evidence of NAFLD progression from steatosis to fibrosing-steatohepatitis using paired biopsies: implications for prognosis and clinical management. J Hepatol 2015, 62, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.D.; Stengel, J.; Asike, M.I.; Torres, D.M.; Shaw, J.; Contreras, M.; Landt, C.L.; Harrison, S.A. Prevalence of nonalcoholic fatty liver disease and nonalcoholic steatohepatitis among a largely middle-aged population utilizing ultrasound and liver biopsy: a prospective study. Gastroenterology 2011, 140, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Pezzino, S.; Sofia, M.; Faletra, G.; Mazzone, C.; Litrico, G.; La Greca, G.; Latteri, S. Gut-Liver Axis and Non-Alcoholic Fatty Liver Disease: A Vicious Circle of Dysfunctions Orchestrated by the Gut Microbiome. Biology (Basel) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Yki-Jarvinen, H. Diagnosis of non-alcoholic fatty liver disease (NAFLD). Diabetologia 2016, 59, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Browning, J.D.; Szczepaniak, L.S.; Dobbins, R.; Nuremberg, P.; Horton, J.D.; Cohen, J.C.; Grundy, S.M.; Hobbs, H.H. Prevalence of hepatic steatosis in an urban population in the United States: impact of ethnicity. Hepatology 2004, 40, 1387–1395. [Google Scholar] [CrossRef] [PubMed]
- Kotronen, A.; Peltonen, M.; Hakkarainen, A.; Sevastianova, K.; Bergholm, R.; Johansson, L.M.; Lundbom, N.; Rissanen, A.; Ridderstrale, M.; Groop, L.; et al. Prediction of non-alcoholic fatty liver disease and liver fat using metabolic and genetic factors. Gastroenterology 2009, 137, 865–872. [Google Scholar] [CrossRef]
- Bedogni, G.; Bellentani, S.; Miglioli, L.; Masutti, F.; Passalacqua, M.; Castiglione, A.; Tiribelli, C. The Fatty Liver Index: a simple and accurate predictor of hepatic steatosis in the general population. BMC Gastroenterol 2006, 6, 33. [Google Scholar] [CrossRef]
- Dixon, J.B.; Bhathal, P.S.; O’Brien, P.E. Nonalcoholic fatty liver disease: predictors of nonalcoholic steatohepatitis and liver fibrosis in the severely obese. Gastroenterology 2001, 121, 91–100. [Google Scholar] [CrossRef]
- McPherson, S.; Stewart, S.F.; Henderson, E.; Burt, A.D.; Day, C.P. Simple non-invasive fibrosis scoring systems can reliably exclude advanced fibrosis in patients with non-alcoholic fatty liver disease. Gut 2010, 59, 1265–1269. [Google Scholar] [CrossRef]
- Ryan, C.K.; Johnson, L.A.; Germin, B.I.; Marcos, A. One hundred consecutive hepatic biopsies in the workup of living donors for right lobe liver transplantation. Liver Transpl 2002, 8, 1114–1122. [Google Scholar] [CrossRef] [PubMed]
- Cengiz, M.; Senturk, S.; Cetin, B.; Bayrak, A.H.; Bilek, S.U. Sonographic assessment of fatty liver: intraobserver and interobserver variability. Int J Clin Exp Med 2014, 7, 5453–5460. [Google Scholar] [PubMed]
- Tilg, H.; Moschen, A.R. Evolution of inflammation in nonalcoholic fatty liver disease: the multiple parallel hits hypothesis. Hepatology 2010, 52, 1836–1846. [Google Scholar] [CrossRef] [PubMed]
- Day, C.P.; James, O.F. Steatohepatitis: a tale of two “hits”? Gastroenterology 1998, 114, 842–845. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Moschen, A.R. Inflammatory mechanisms in the regulation of insulin resistance. Mol Med 2008, 14, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ding, L.; Hassan, W.; Abdelkader, D.; Shang, J. Adipokines and hepatic insulin resistance. J Diabetes Res 2013, 2013, 170532. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, L.; Ergin, A.S.; Lu, A.; Chung, J.; Sarkar, S.; Nie, D.; Myers, M.G., Jr.; Ozcan, U. Endoplasmic reticulum stress plays a central role in development of leptin resistance. Cell Metab 2009, 9, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Sha, H.; He, Y.; Chen, H.; Wang, C.; Zenno, A.; Shi, H.; Yang, X.; Zhang, X.; Qi, L. The IRE1alpha-XBP1 pathway of the unfolded protein response is required for adipogenesis. Cell Metab 2009, 9, 556–564. [Google Scholar] [CrossRef]
- Begriche, K.; Massart, J.; Robin, M.A.; Bonnet, F.; Fromenty, B. Mitochondrial adaptations and dysfunctions in nonalcoholic fatty liver disease. Hepatology 2013, 58, 1497–1507. [Google Scholar] [CrossRef]
- Feldstein, A.E.; Werneburg, N.W.; Canbay, A.; Guicciardi, M.E.; Bronk, S.F.; Rydzewski, R.; Burgart, L.J.; Gores, G.J. Free fatty acids promote hepatic lipotoxicity by stimulating TNF-alpha expression via a lysosomal pathway. Hepatology 2004, 40, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Mari, M.; Caballero, F.; Colell, A.; Morales, A.; Caballeria, J.; Fernandez, A.; Enrich, C.; Fernandez-Checa, J.C.; Garcia-Ruiz, C. Mitochondrial free cholesterol loading sensitizes to TNF- and Fas-mediated steatohepatitis. Cell Metab 2006, 4, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S.; Erbay, E. Nutrient sensing and inflammation in metabolic diseases. Nat Rev Immunol 2008, 8, 923–934. [Google Scholar] [CrossRef] [PubMed]
- Romeo, S.; Kozlitina, J.; Xing, C.; Pertsemlidis, A.; Cox, D.; Pennacchio, L.A.; Boerwinkle, E.; Cohen, J.C.; Hobbs, H.H. Genetic variation in PNPLA3 confers susceptibility to nonalcoholic fatty liver disease. Nat Genet 2008, 40, 1461–1465. [Google Scholar] [CrossRef] [PubMed]
- Petersen, K.F.; Dufour, S.; Hariri, A.; Nelson-Williams, C.; Foo, J.N.; Zhang, X.M.; Dziura, J.; Lifton, R.P.; Shulman, G.I. Apolipoprotein C3 gene variants in nonalcoholic fatty liver disease. N Engl J Med 2010, 362, 1082–1089. [Google Scholar] [CrossRef] [PubMed]
- Kantartzis, K.; Peter, A.; Machicao, F.; Machann, J.; Wagner, S.; Konigsrainer, I.; Konigsrainer, A.; Schick, F.; Fritsche, A.; Haring, H.U.; et al. Dissociation between fatty liver and insulin resistance in humans carrying a variant of the patatin-like phospholipase 3 gene. Diabetes 2009, 58, 2616–2623. [Google Scholar] [CrossRef]
- Kotronen, A.; Johansson, L.E.; Johansson, L.M.; Roos, C.; Westerbacka, J.; Hamsten, A.; Bergholm, R.; Arkkila, P.; Arola, J.; Kiviluoto, T.; et al. A common variant in PNPLA3, which encodes adiponutrin, is associated with liver fat content in humans. Diabetologia 2009, 52, 1056–1060. [Google Scholar] [CrossRef]
- Bril, F.; Sninsky, J.J.; Baca, A.M.; Superko, H.R.; Portillo Sanchez, P.; Biernacki, D.; Maximos, M.; Lomonaco, R.; Orsak, B.; Suman, A.; et al. Hepatic Steatosis and Insulin Resistance, But Not Steatohepatitis, Promote Atherogenic Dyslipidemia in NAFLD. J Clin Endocrinol Metab 2016, 101, 644–652. [Google Scholar] [CrossRef]
- Bril, F.; Barb, D.; Portillo-Sanchez, P.; Biernacki, D.; Lomonaco, R.; Suman, A.; Weber, M.H.; Budd, J.T.; Lupi, M.E.; Cusi, K. Metabolic and histological implications of intrahepatic triglyceride content in nonalcoholic fatty liver disease. Hepatology 2017, 65, 1132–1144. [Google Scholar] [CrossRef]
- Lomonaco, R.; Ortiz-Lopez, C.; Orsak, B.; Webb, A.; Hardies, J.; Darland, C.; Finch, J.; Gastaldelli, A.; Harrison, S.; Tio, F.; et al. Effect of adipose tissue insulin resistance on metabolic parameters and liver histology in obese patients with nonalcoholic fatty liver disease. Hepatology 2012, 55, 1389–1397. [Google Scholar] [CrossRef]
- Munukka, E.; Pekkala, S.; Wiklund, P.; Rasool, O.; Borra, R.; Kong, L.; Ojanen, X.; Cheng, S.M.; Roos, C.; Tuomela, S.; et al. Gut-adipose tissue axis in hepatic fat accumulation in humans. J Hepatol 2014, 61, 132–138. [Google Scholar] [CrossRef]
- Mouzaki, M.; Comelli, E.M.; Arendt, B.M.; Bonengel, J.; Fung, S.K.; Fischer, S.E.; McGilvray, I.D.; Allard, J.P. Intestinal microbiota in patients with nonalcoholic fatty liver disease. Hepatology 2013, 58, 120–127. [Google Scholar] [CrossRef]
- Aron-Wisnewsky, J.; Vigliotti, C.; Witjes, J.; Le, P.; Holleboom, A.G.; Verheij, J.; Nieuwdorp, M.; Clement, K. Gut microbiota and human NAFLD: disentangling microbial signatures from metabolic disorders. Nat Rev Gastroenterol Hepatol 2020, 17, 279–297. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; Bäckhed, F. From Association to Causality: the Role of the Gut Microbiota and Its Functional Products on Host Metabolism. Molecular Cell 2020, 78, 584–596. [Google Scholar] [CrossRef]
- Singh, V.; Chassaing, B.; Zhang, L.; San Yeoh, B.; Xiao, X.; Kumar, M.; Baker, M.T.; Cai, J.; Walker, R.; Borkowski, K.; et al. Microbiota-Dependent Hepatic Lipogenesis Mediated by Stearoyl CoA Desaturase 1 (SCD1) Promotes Metabolic Syndrome in TLR5-Deficient Mice. Cell Metab 2015, 22, 983–996. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Ruuskanen, M.O.; Aberg, F.; Mannisto, V.; Havulinna, A.S.; Meric, G.; Liu, Y.; Loomba, R.; Vazquez-Baeza, Y.; Tripathi, A.; Valsta, L.M.; et al. Links between gut microbiome composition and fatty liver disease in a large population sample. Gut Microbes 2021, 13, 1–22. [Google Scholar] [CrossRef]
- Castellana, M.; Donghia, R.; Guerra, V.; Procino, F.; Lampignano, L.; Castellana, F.; Zupo, R.; Sardone, R.; De Pergola, G.; Romanelli, F.; et al. Performance of Fatty Liver Index in Identifying Non-Alcoholic Fatty Liver Disease in Population Studies. A Meta-Analysis. J Clin Med 2021, 10. [Google Scholar] [CrossRef]
- Yuan, J.; Chen, C.; Cui, J.; Lu, J.; Yan, C.; Wei, X.; Zhao, X.; Li, N.; Li, S.; Xue, G.; et al. Fatty Liver Disease Caused by High-Alcohol-Producing Klebsiella pneumoniae. Cell Metab 2019, 30, 1172. [Google Scholar] [CrossRef]
- Li, M.; van Esch, B.; Wagenaar, G.T.M.; Garssen, J.; Folkerts, G.; Henricks, P.A.J. Pro- and anti-inflammatory effects of short chain fatty acids on immune and endothelial cells. Eur J Pharmacol 2018, 831, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.X.; Lee, J.S.; Campbell, E.L.; Colgan, S.P. Microbiota-derived butyrate dynamically regulates intestinal homeostasis through regulation of actin-associated protein synaptopodin. Proceedings of the National Academy of Sciences of the United States of America 2020, 117, 11648–11657. [Google Scholar] [CrossRef] [PubMed]
- Hoyles, L.; Fernandez-Real, J.M.; Federici, M.; Serino, M.; Abbott, J.; Charpentier, J.; Heymes, C.; Luque, J.L.; Anthony, E.; Barton, R.H.; et al. Molecular phenomics and metagenomics of hepatic steatosis in non-diabetic obese women. Nat Med 2018, 24, 1070–1080. [Google Scholar] [CrossRef] [PubMed]
- Sharifnia, T.; Antoun, J.; Verriere, T.G.; Suarez, G.; Wattacheril, J.; Wilson, K.T.; Peek, R.M., Jr.; Abumrad, N.N.; Flynn, C.R. Hepatic TLR4 signaling in obese NAFLD. Am J Physiol Gastrointest Liver Physiol 2015, 309, G270–G278. [Google Scholar] [CrossRef]
- Wolf, A.J.; Underhill, D.M. Peptidoglycan recognition by the innate immune system. Nat Rev Immunol 2018, 18, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Cuomo, P.; Capparelli, R.; Iannelli, A.; Iannelli, D. Role of Branched-Chain Amino Acid Metabolism in Type 2 Diabetes, Obesity, Cardiovascular Disease and Non-Alcoholic Fatty Liver Disease. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Driuchina, A.; Hintikka, J.; Lehtonen, M.; Keski-Rahkonen, P.; O’Connell, T.; Juvonen, R.; Kuula, J.; Hakkarainen, A.; Laukkanen, J.A.; Makinen, E.; et al. Identification of Gut Microbial Lysine and Histidine Degradation and CYP-Dependent Metabolites as Biomarkers of Fatty Liver Disease. mBio 2023, e0266322. [Google Scholar] [CrossRef]
- Grenier-Larouche, T.; Coulter Kwee, L.; Deleye, Y.; Leon-Mimila, P.; Walejko, J.M.; McGarrah, R.W.; Marceau, S.; Trahan, S.; Racine, C.; Carpentier, A.C.; et al. Altered branched-chain alpha-keto acid metabolism is a feature of NAFLD in individuals with severe obesity. JCI Insight 2022, 7. [Google Scholar] [CrossRef]
- Michail, S.; Lin, M.; Frey, M.R.; Fanter, R.; Paliy, O.; Hilbush, B.; Reo, N.V. Altered gut microbial energy and metabolism in children with non-alcoholic fatty liver disease. FEMS Microbiol Ecol 2015, 91, 1–9. [Google Scholar] [CrossRef]
- Van de Velde, F.; Bekaert, M.; Hoorens, A.; Geerts, A.; T’Sjoen, G.; Fiers, T.; Kaufman, J.M.; Van Nieuwenhove, Y.; Lapauw, B. Histologically proven hepatic steatosis associates with lower testosterone levels in men with obesity. Asian J Androl 2020, 22, 252–257. [Google Scholar] [CrossRef]
- Jensen, T.; Abdelmalek, M.F.; Sullivan, S.; Nadeau, K.J.; Green, M.; Roncal, C.; Nakagawa, T.; Kuwabara, M.; Sato, Y.; Kang, D.H.; et al. Fructose and sugar: A major mediator of non-alcoholic fatty liver disease. J Hepatol 2018, 68, 1063–1075. [Google Scholar] [CrossRef] [PubMed]
- El Hadi, H.; Vettor, R.; Rossato, M. Vitamin E as a Treatment for Nonalcoholic Fatty Liver Disease: Reality or Myth? Antioxidants (Basel) 2018, 7. [Google Scholar] [CrossRef]
- Ge, H.; Wei, W.; Tang, L.; Tian, Y.; Zhu, Y.; Luo, Y.; Liu, S. CONSORT-Characteristics and metabolic phenotype of gut microbiota in NAFLD patients. Medicine (Baltimore) 2022, 101, e29347. [Google Scholar] [CrossRef] [PubMed]
- Dall, M.; Hassing, A.S.; Treebak, J.T. NAD(+) and NAFLD - caution, causality and careful optimism. J Physiol 2022, 600, 1135–1154. [Google Scholar] [CrossRef]
- Nishikawa, T.; Nagata, N.; Shimakami, T.; Shirakura, T.; Matsui, C.; Ni, Y.; Zhuge, F.; Xu, L.; Chen, G.; Nagashimada, M.; et al. Xanthine oxidase inhibition attenuates insulin resistance and diet-induced steatohepatitis in mice. Sci Rep 2020, 10, 815. [Google Scholar] [CrossRef] [PubMed]
- Ucar, F.; Sezer, S.; Erdogan, S.; Akyol, S.; Armutcu, F.; Akyol, O. The relationship between oxidative stress and nonalcoholic fatty liver disease: Its effects on the development of nonalcoholic steatohepatitis. Redox Rep 2013, 18, 127–133. [Google Scholar] [CrossRef]
- Boursier, J.; Mueller, O.; Barret, M.; Machado, M.; Fizanne, L.; Araujo-Perez, F.; Guy, C.D.; Seed, P.C.; Rawls, J.F.; David, L.A.; et al. The severity of nonalcoholic fatty liver disease is associated with gut dysbiosis and shift in the metabolic function of the gut microbiota. Hepatology 2016, 63, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Loomba, R.; Seguritan, V.; Li, W.; Long, T.; Klitgord, N.; Bhatt, A.; Dulai, P.S.; Caussy, C.; Bettencourt, R.; Highlander, S.K.; et al. Gut Microbiome-Based Metagenomic Signature for Non-invasive Detection of Advanced Fibrosis in Human Nonalcoholic Fatty Liver Disease. Cell Metab 2017, 25, 1054–1062 e1055. [Google Scholar] [CrossRef]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat Biotechnol 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Kordy, K.; Li, F.; Lee, D.J.; Kinchen, J.M.; Jew, M.H.; La Rocque, M.E.; Zabih, S.; Saavedra, M.; Woodward, C.; Cunningham, N.J.; et al. Metabolomic Predictors of Non-alcoholic Steatohepatitis and Advanced Fibrosis in Children. Front Microbiol 2021, 12, 713234. [Google Scholar] [CrossRef]
- Lee, G.; You, H.J.; Bajaj, J.S.; Joo, S.K.; Yu, J.; Park, S.; Kang, H.; Park, J.H.; Kim, J.H.; Lee, D.H.; et al. Distinct signatures of gut microbiome and metabolites associated with significant fibrosis in non-obese NAFLD. Nat Commun 2020, 11, 4982. [Google Scholar] [CrossRef] [PubMed]
- Albillos, A.; de Gottardi, A.; Rescigno, M. The gut-liver axis in liver disease: Pathophysiological basis for therapy. J Hepatol 2020, 72, 558–577. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, E.; Muthiah, M.D.; Narayan, N.; Siddiqui, M.S.; Puri, P.; Luketic, V.A.; Contos, M.J.; Idowu, M.; Chuang, J.C.; Billin, A.N.; et al. Metabolic reprogramming of the intestinal microbiome with functional bile acid changes underlie the development of NAFLD. Hepatology 2022, 76, 1811–1824. [Google Scholar] [CrossRef] [PubMed]
- Sui, G.; Jia, L.; Quan, D.; Zhao, N.; Yang, G. Activation of the gut microbiota-kynurenine-liver axis contributes to the development of nonalcoholic hepatic steatosis in nondiabetic adults. Aging (Albany NY) 2021, 13, 21309–21324. [Google Scholar] [CrossRef]
- Rojas, I.Y.; Moyer, B.J.; Ringelberg, C.S.; Wilkins, O.M.; Pooler, D.B.; Ness, D.B.; Coker, S.; Tosteson, T.D.; Lewis, L.D.; Chamberlin, M.D.; et al. Kynurenine-Induced Aryl Hydrocarbon Receptor Signaling in Mice Causes Body Mass Gain, Liver Steatosis, and Hyperglycemia. Obesity (Silver Spring) 2021, 29, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nat Rev Dis Primers 2021, 7, 6. [Google Scholar] [CrossRef]
- Behary, J.; Amorim, N.; Jiang, X.T.; Raposo, A.; Gong, L.; McGovern, E.; Ibrahim, R.; Chu, F.; Stephens, C.; Jebeili, H.; et al. Gut microbiota impact on the peripheral immune response in non-alcoholic fatty liver disease related hepatocellular carcinoma. Nat Commun 2021, 12, 187. [Google Scholar] [CrossRef]
- Oh, T.G.; Kim, S.M.; Caussy, C.; Fu, T.; Guo, J.; Bassirian, S.; Singh, S.; Madamba, E.V.; Bettencourt, R.; Richards, L.; et al. A Universal Gut-Microbiome-Derived Signature Predicts Cirrhosis. Cell Metab 2020, 32, 878–888. [Google Scholar] [CrossRef]
- Da Silva, H.E.; Teterina, A.; Comelli, E.M.; Taibi, A.; Arendt, B.M.; Fischer, S.E.; Lou, W.; Allard, J.P. Nonalcoholic fatty liver disease is associated with dysbiosis independent of body mass index and insulin resistance. Sci Rep 2018, 8, 1466. [Google Scholar] [CrossRef]
- Backhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc Natl Acad Sci USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef]
- Testerman, T.; Li, Z.; Galuppo, B.; Graf, J.; Santoro, N. Insights from shotgun metagenomics into bacterial species and metabolic pathways associated with NAFLD in obese youth. Hepatol Commun 2022, 6, 1962–1974. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhou, J.; Liu, J.; Wang, Z.; Chen, M.; Zhou, S. Metagenome of Gut Microbiota of Children With Nonalcoholic Fatty Liver Disease. Front Pediatr 2019, 7, 518. [Google Scholar] [CrossRef]
- Yu, J.; Zhang, H.; Chen, L.; Ruan, Y.; Chen, Y.; Liu, Q. Disease-Associated Gut Microbiota Reduces the Profile of Secondary Bile Acids in Pediatric Nonalcoholic Fatty Liver Disease. Front Cell Infect Microbiol 2021, 11, 698852. [Google Scholar] [CrossRef]
- Schwimmer, J.B.; Johnson, J.S.; Angeles, J.E.; Behling, C.; Belt, P.H.; Borecki, I.; Bross, C.; Durelle, J.; Goyal, N.P.; Hamilton, G.; et al. Microbiome Signatures Associated With Steatohepatitis and Moderate to Severe Fibrosis in Children With Nonalcoholic Fatty Liver Disease. Gastroenterology 2019, 157, 1109–1122. [Google Scholar] [CrossRef]
- Toivonen, R.; Vanhatalo, S.; Hollmén, M.; Munukka, E.; Keskitalo, A.; Pietilä, S.; Elo, L.; Huovinen, P.; Jalkanen, S.; Pekkala, S. Vascular Adhesion Protein 1 Mediates Gut Microbial Flagellin-Induced Inflammation, Leukocyte Infiltration, and Hepatic Steatosis. Sci 2021, 3, 13. [Google Scholar] [CrossRef]
- Tourlomousis, P.; Wright, J.A.; Bittante, A.S.; Hopkins, L.J.; Webster, S.J.; Bryant, O.J.; Mastroeni, P.; Maskell, D.J.; Bryant, C.E. Modifying bacterial flagellin to evade Nod-like Receptor CARD 4 recognition enhances protective immunity against Salmonella. Nat Microbiol 2020, 5, 1588–1597. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Hydes, T.; Alam, U.; Cuthbertson, D.J. The Impact of Macronutrient Intake on Non-alcoholic Fatty Liver Disease (NAFLD): Too Much Fat, Too Much Carbohydrate, or Just Too Many Calories? Front Nutr 2021, 8, 640557. [Google Scholar] [CrossRef] [PubMed]
- Bauer, K.C.; Littlejohn, P.T.; Ayala, V.; Creus-Cuadros, A.; Finlay, B.B. Nonalcoholic Fatty Liver Disease and the Gut-Liver Axis: Exploring an Undernutrition Perspective. Gastroenterology 2022, 162, 1858–1875. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The abundance of many members of the gut microbiota affects the human physiology by processing the ingested food into certain bioactive metabolites. These molecules can act as inter-tissue signaling messengers by penetrating the portal vein and further, the liver to promote or prevent hepatic fat accumulation. For instance, dietary fibre is metabolized by the gut microbiota into short chain fatty acids (SCFA, mainly butyrate, acetate, and propionate), and ethanol is produced from dietary carbohydrates. Gut microbiota produce trimethyl amine (TMA) from dietary choline that is mainly derived from meat, yolk, and dairy products. TMA is converted further into trimethyl amineoxide (TMAO) in liver. Primary bile acids (BA) are synthesized in liver, stored in the gallbladder, and then released into the gut, where they are converted into secondary BA by the gut microbiota. In liver, ethanol can increase fat accumulation by reducing beta-oxidation. TMAO has been shown to induce insulin resistance (IR), and further fat accumulation in the liver. Controversial findings on the role of BA and SCFA in hepatic fat accumulation exist. These will be reviewed in this article. The figure was created using BioRender.
Figure 1.
The abundance of many members of the gut microbiota affects the human physiology by processing the ingested food into certain bioactive metabolites. These molecules can act as inter-tissue signaling messengers by penetrating the portal vein and further, the liver to promote or prevent hepatic fat accumulation. For instance, dietary fibre is metabolized by the gut microbiota into short chain fatty acids (SCFA, mainly butyrate, acetate, and propionate), and ethanol is produced from dietary carbohydrates. Gut microbiota produce trimethyl amine (TMA) from dietary choline that is mainly derived from meat, yolk, and dairy products. TMA is converted further into trimethyl amineoxide (TMAO) in liver. Primary bile acids (BA) are synthesized in liver, stored in the gallbladder, and then released into the gut, where they are converted into secondary BA by the gut microbiota. In liver, ethanol can increase fat accumulation by reducing beta-oxidation. TMAO has been shown to induce insulin resistance (IR), and further fat accumulation in the liver. Controversial findings on the role of BA and SCFA in hepatic fat accumulation exist. These will be reviewed in this article. The figure was created using BioRender.
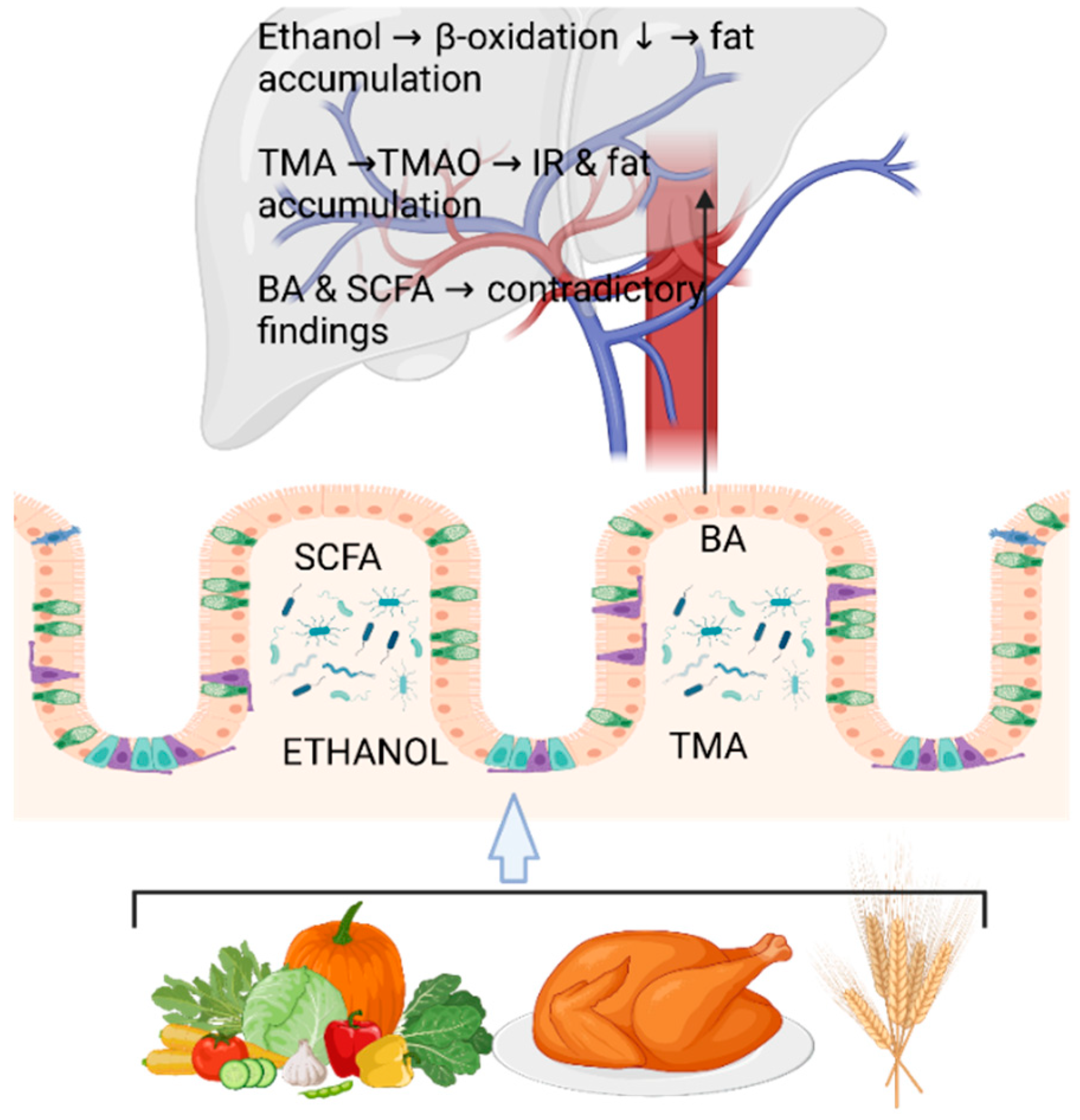
Figure 2.
Main findings from two fecal metagenomic studies and one metabolomics study in individuals with hepatic steatosis and without diagnosed NAFLD. CH, carbohydrates; AA, amino acids; BCAA, branched chain amino acids; AAA, aromatic amino acids; LPS, lipopolysaccharides. The figure was created using BioRender.
Figure 2.
Main findings from two fecal metagenomic studies and one metabolomics study in individuals with hepatic steatosis and without diagnosed NAFLD. CH, carbohydrates; AA, amino acids; BCAA, branched chain amino acids; AAA, aromatic amino acids; LPS, lipopolysaccharides. The figure was created using BioRender.
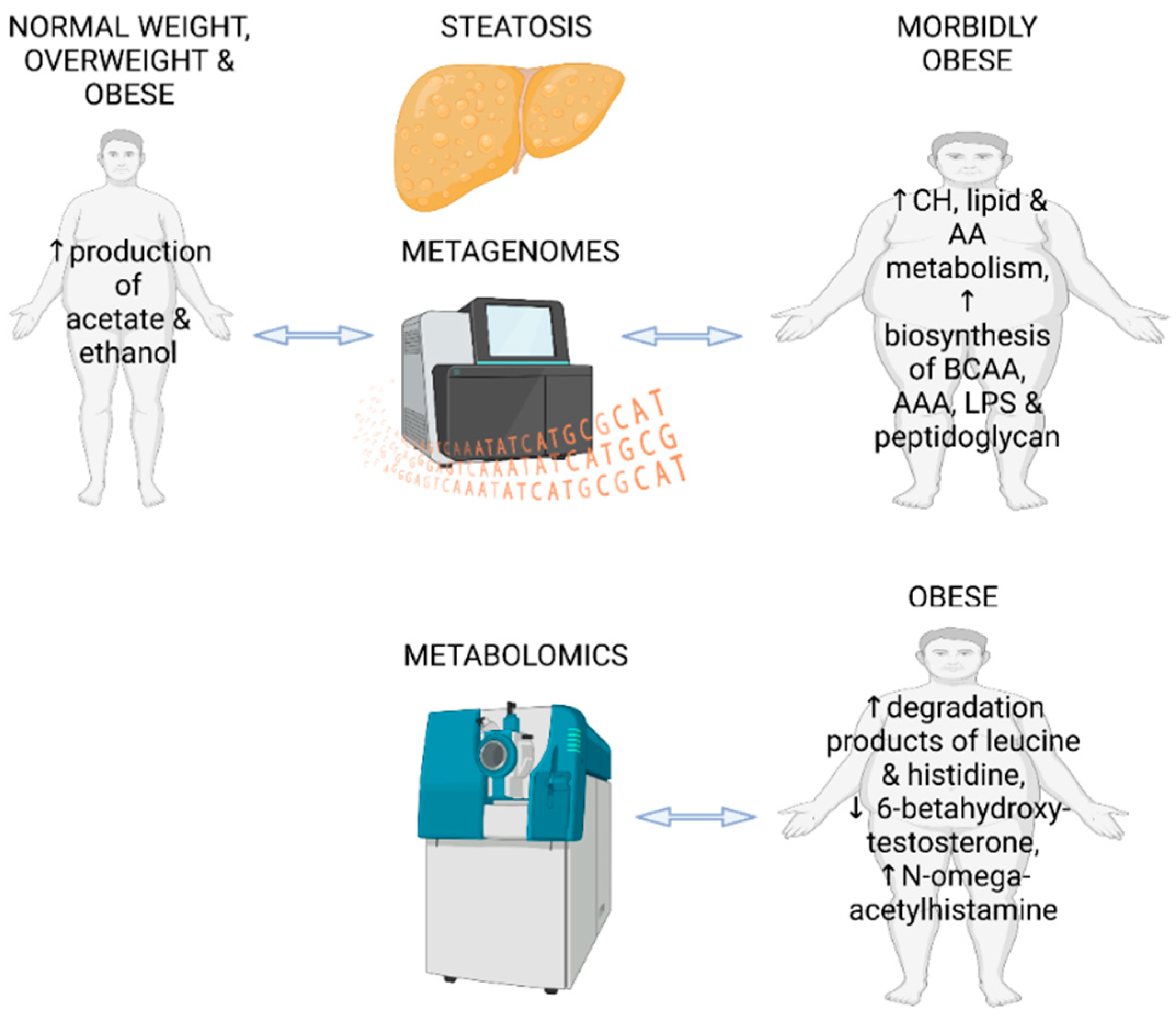
Figure 3.
Main findings from studies reporting fecal metagenomics and/or metabolomics in NAFLD and advanced fibrosis. NAFL, non-alcoholic fatty liver; NASH, non-alcoholic steatohepatosis; CH, carbohydrates; BA, bile acids. The figure was created using BioRender.
Figure 3.
Main findings from studies reporting fecal metagenomics and/or metabolomics in NAFLD and advanced fibrosis. NAFL, non-alcoholic fatty liver; NASH, non-alcoholic steatohepatosis; CH, carbohydrates; BA, bile acids. The figure was created using BioRender.
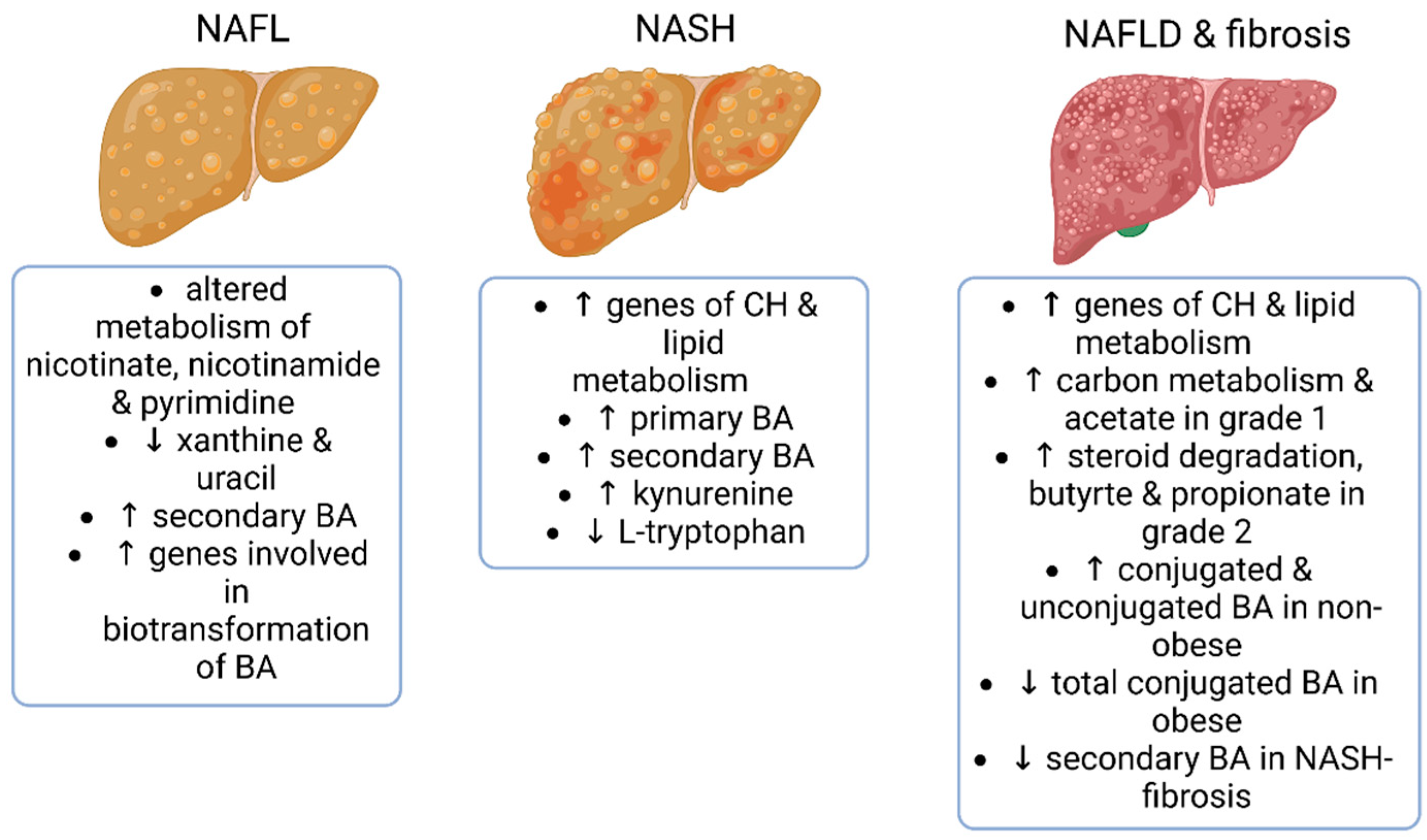
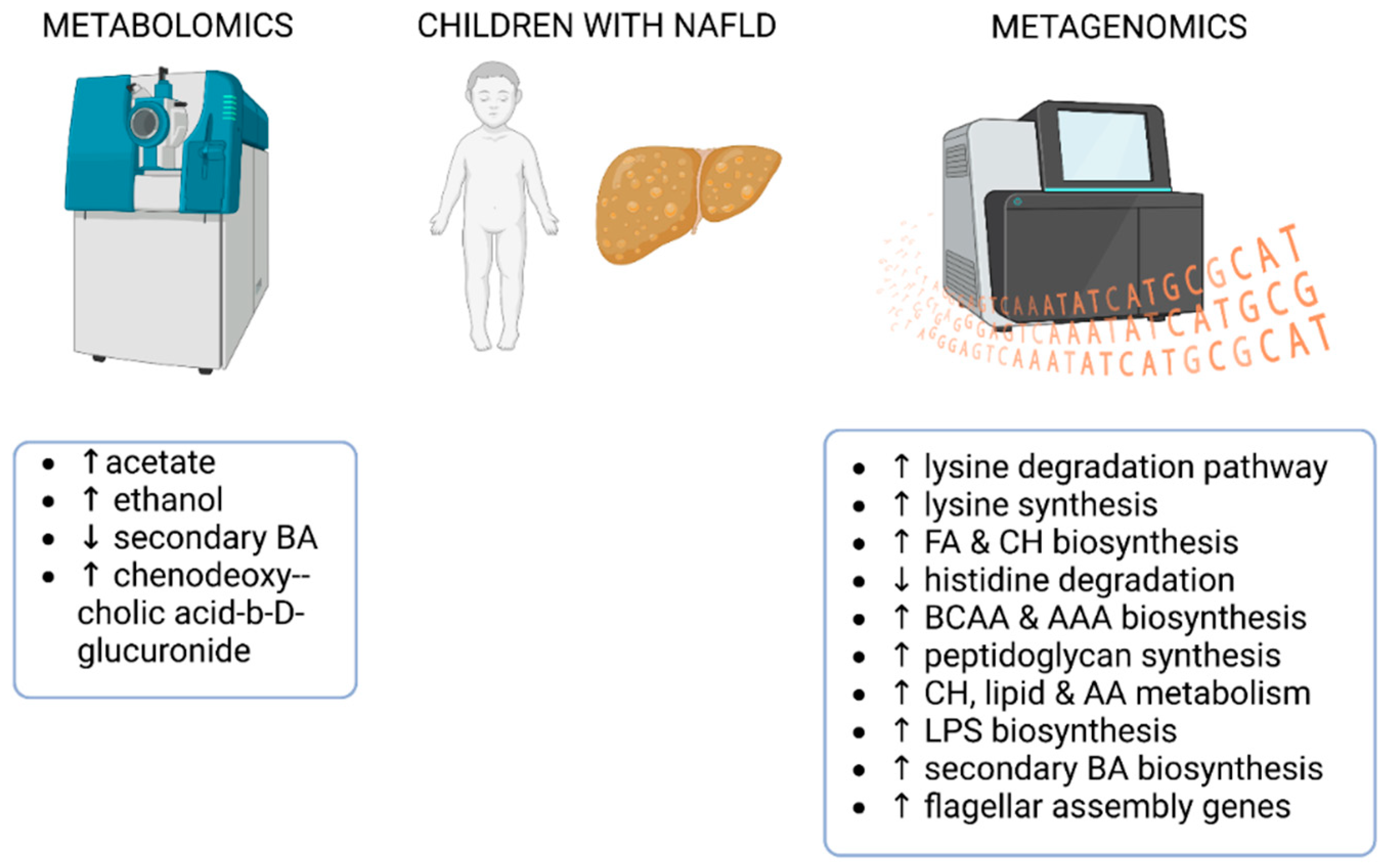
Figure 4.
Main findings from studies reporting fecal metagenomics and/or metabolomics in children with NAFLD. FA, fatty acids; CH, carbohydrates; BA, bile acids; AA, amino acids, BCAA, branched chain amino acids; AAA, aromatic amino acids; LPS, lipopolysaccharides. The figure was created using BioRender.
Figure 4.
Main findings from studies reporting fecal metagenomics and/or metabolomics in children with NAFLD. FA, fatty acids; CH, carbohydrates; BA, bile acids; AA, amino acids, BCAA, branched chain amino acids; AAA, aromatic amino acids; LPS, lipopolysaccharides. The figure was created using BioRender.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



