
Submitted:
01 February 2023
Posted:
03 February 2023
You are already at the latest version
Abstract
Calcium-activated potassium channels (KCa) are the important participants in calcium signaling pathways due to their ability to be activated by increase of the intracellular free calcium concentration. KCa channels are involved in the regulation of various processes in the cells under normal, as well as pathophysiological conditions, including oncotransformation. Previously, with the use of patch-clamp method, we registered the KCa activity in the plasma membrane of human myeloid leukemia K562 cell line. Here, we performed the molecular and functional identification of KCa channels and have uncovered their role in proliferation, migration and invasion of K562 cells. Using a combined approach, we identified the functional activity of SK2, SK3 and IK channels in the plasma membrane of the cells. Selective SK and IK channel inhibitors, apamin and TRAM-34, reduced the proliferative, migratory and invasive capabilities of human myeloid leukemia cells. At the same time, the viability of K562 cells was not affected by KCa channel inhibitors. Our data imply that SK/IK channel inhibitors could be used to slow down the proliferation and spreading of leukemia cells that express functionally active KCa channels in the plasma membrane.
Keywords:
Calcium-activated potassium channels
; SK channels
; IK channels
; human leukemia cells
; cell migration
; cancer cell invasion
1. Introduction
Ion channels are transmembrane protein complexes involved in the regulation of many physiological functions of living cells in health and disease. Different types of cells and tissues are characterized by various repertoires of specific ion channels that could define cellular reactions and signaling events. It is assumed that the migration and proliferation of cancer cells (including blood malignancies) could be critically dependent on the membrane potential, mechanical properties of the plasma membrane and cellular volume changes [1,2]. The control of the above functions of malignant cells is closely related to the activity of various types of ion channels.
Calcium-activated potassium channels (KCa channels) are a unique class among other K+ channels that are stimulated by the increase in intracellular Ca2+ concentration ([Ca2+]i). They are divided into three sub-groups based on their single-channel properties: big (BK), intermediate (IK) and small (SK) conductance. Among them, BK channels could be activated by the synergy of membrane potential (are voltage-dependent) and Ca2+, whereas SK and IK channels are voltage-independent and are gated solely by Ca2+ that bind to channel-associated calmodulin. Small- and intermediate-conductance KCa channels (KCa2/SK and KCa3.1/IK channels), which are gated by sub micromolar Ca2+ concentrations, are involved in the regulation of cell volume and mediated Ca2+-induced membrane hyperpolarization [3,4,5]. These channels are encoded by four KCNN genes: three subtypes for SK channels, KCNN1–3 for KCa2.1–2.3 (SK1–SK3) and KCNN4 for KCa3.1 (IK or Gardos channels), respectively [4]. Importantly, the SK and IK channel activity could be reliably distinguished by highly selective pharmacological channel inhibitors. Moreover, the activity of the members of the SK family could also be separated from each other due to their different sensitivity to the component of bee venom - a selective SK channel blocker apamin [4]. Particularly, SK2 channels are the most sensitive to apamin with a half maximal blocking concentration (IC50: 27-140 pM, SK1 are less sensitive (IC50: 0.7-12 nM), and SK3 show an intermediate sensitivity to apamin (IC50: 0.6-4 nM). Importantly, apamin has no inhibitory effects on other types of K+ channels as well as on ion channels from other families as it was confirmed recently on a broad repertoire of channel proteins [6]. For functional identification of IK activity, TRAM-34, the analog of clotrimazole, a potent and selective blocker of IK (KCa3.1), is widely used [7,8].
The involvement of SK and IK channels in the regulation of various processes in blood cells was investigated. Moreover, activation of KCa current was demonstrated, for the first time, in red blood cells [9]. In T lymphocytes SK channels have been shown to be involved in the maintenance of Ca2+ signaling [10,11]. In human leukemic cell lines CEM and MOLT-3 and healthy T cells the expression of only IK channels was found while apamin-sensitive SK2 currents predominated in Jurkat leukemic cells [12]. IK (Gardos) channels regulate erythrocytes volume by the transfer of Ca2+ uptake into K+ and loss of water: Ca2+- dependent activation of IK and subsequent K+ efflux cause erythrocyte dehydration (cell shrinkage) [13]. Impairment of K+ transport and fluid flow associated with mutations in IK channels may lead to erythrocyte disorders such as hereditary xerocytosis or stomatocytosis [14].
Dyrda et al. using patch-clamp technique found that local deformation of red blood cells (RBC) membrane (induced by the application of a negative pressure pulse during patch formation) may activate Ca2+-dependent K+ currents due to mechano-dependent Ca2+ entry [15]. These experiments suggest that mechanosensitive mechanisms may play an important role in controlling the shape and volume of RBC. Accordingly, mechanosensitive Ca2+-permeable Piezo1 channels have been shown to be implicated in RBC volume homeostasis, inducing Ca2+ entry, which, in turn, activates IK channels and triggers erythrocyte dehydration [16]. Importantly, in our previous studies on human myeloid leukemia K562 cell line, which are of the erythroleukemia type, the activity of mechanosensitive Ca2+-permeable (MSCa) ion channels with biophysical properties close to Piezo family was discovered [17,18,19]. Moreover, coupled activation of MSCas and KCa channels was found in K562 cells [20] indicating the presence of functional complexes of Ca2+-permeable and KCa channels in the plasma membrane. It was shown that such complexes could be formed by the Ca2+ channels and KCa channels of various subtypes and could contribute to cancer-associated functions of the cells [21]. The purpose of this work is to reveal the molecular identity of KCa channels in human leukemia K562 cells and to identify their possible contribution to K+ currents and regulation of pathophysiological functions of the cells.
2. Results
2.1. Selective SK and IK channel blockers inhibit whole-cell currents in the plasma membrane of human myeloid leukemia cells
The first series of experiments were carried out to determine the level of functional expression of KCa channels in the plasma membrane of human leukemia K562 cells. Based on previously reported single-channel conductance (about 10 pS in near-physiological Na-based bath solution [20]), the KCa currents could be mediated by SK or IK channels. Thus, we designed the whole-cell patch-clamp experiments in which we were able to distinguish between SK and IK channels using highly specific inhibitory analysis. As selective blockers of SK and IK channels, 300 nM apamin and 1 μM TRAM-34 were used, respectively. Both pore blockers act from the extracellular side of the membrane [22,23,24], thus we added them to the extracellular media in the course of sequential changes of bath solution during the recording. It is known that SK and IK channels are activated by cytoplasmic Ca2+ (from about 0.1 μM or more) via calmodulin-binding mechanism [25]. Thus, we used the pipette cytosol-like solution containing high concentration of intracellular free Ca2+ (10-6 M=1 μM or pCa6, see Materials and Methods) for reliable activation of both SK and IK currents. The K+ outward currents in the plasma membrane of K562 cells were recorded at positive membrane potentials in the range of +20 to +80 mV.
After establishing the whole-cell configuration, firstly we have recorded outward K+ currents under control conditions (145 mM NaCl in bath solution; KAsp-based pipette solution, pCa6, Figure 1). After that, we changed the bath solution to an apamin-containing one, and then to the solution with both apamin and TRAM-34 to avoid the possibility of re-activation of SK channels by wash-out of apamin. In whole-cell experiments we have observed a stepward decrease of ion currents in response to the subsequent addition of the inhibitors (Figure 1A,B) indicating the presence of both SK- and IK-mediated K+ channel activity in the plasma membrane. Particularly, the application of 300 nM apamin (that is sufficient to block all SK subtypes) resulted in significant reduction of K+ currents allowing to demonstrate SK-mediated components. The addition of 1 μM TRAM-34 (together with 300 nM apamin) resulted in further decrease in outward currents (Figure 1B) that demonstrate the contribution of IK channels. Thus, our whole-cell patch-clamp experiments using selective inhibitors unequivocally demonstrated the functional activity of both SK and IK channels in the plasma membrane of K562 cells.
2.2. Functional expression of SK2, SK3 and IK channels in K562 cells
As a next step, using RT-PCR analysis, we identified what subtypes of SK channels (SK1, SK2 or SK3) are expressed in K562 cells and showed the presence of mRNA encoding IK channels. Particularly, we detected mRNA for SK2 (KCa2.1, encoded by the KCNN2 gene), SK3 (KCa2.3, encoded by the KCNN3 gene) and IK (KCa3.1, encoded by the KCNN4 gene), whereas SK1 was not detected by the reaction (Figure 1C). The immunofluorescent staining of SK2, SK3 and IK with specific antibodies confirmed the presence of the channel proteins in K562 cells (Figure 1D).
Based on the results of RT-PCR, which allowed us to detect the presence of mRNA of SK2 and SK3 channels, we decided to show the impact of each of the channel to whole-cell K+ currents. This could be done due to the different levels of SK channel sensitivity to apamin: the SK2 channels are sensitive to picomolar (pM) concentrations (IC50=27-140 pM) whereas SK3 channels are sensitive in nanomolar (nM) range (IC50=0.6-4 nM, [4]). In whole-cell experiments we have sequentially applied two apamin concentrations: 300 pM to reliably block SK2, and 8 nM for SK3 inhibition, respectively (Figure 2). Consistently, we observed a decrease of whole-cell currents in response to both concentrations of apamin that confirms the functional activity of SK2 and SK3 in the plasma membrane of K562 cells (Figure 2). Taken together, our electrophysiological assay evidenced the presence of functionally active SK2 and SK3 in human myeloid leukemia cells.
2.3. The effects of selective SK and IK channel inhibitors on cell proliferation and viability
It is known that Ca2+ is a main second messenger that plays a crucial role in tumorigenesis, such as proliferation, migration, invasion, etc. of malignant cells [27,28]. KCa channels, a components of Ca2+ signaling pathways that are activated by the rise of [Ca2+]i, were found in various cancer cell types where they could control several pathophysiological reactions [21,27]. The increase of SK3 channel expression is an unfavorable prognostic marker in renal cancer (The Human Protein Atlas, https://www.proteinatlas.org/ENSG00000143603-KCNN3/pathology), whereas IK channel expression is a poor prognosis marker for urothelial, renal, glioma and pancreatic cancer (https://www.proteinatlas.org/ENSG00000104783-KCNN4/pathology). We hypothesized that identified SK and IK channels could be possibly involved in the proliferation, cell viability, migration and invasion of human leukemia cells. Firstly, we assessed the proliferation of K562 cells under control conditions (full media) and in the presence of KCa blockers (apamin or TRAM-34) in culture media. As an additional control, the culture media containing tetraethylammonium bromide (TEA, 10 mM) that is widely used as a non-selective blocker of K+ channels from various families [29], was used to test the impact of K+ currents to the proliferative activity of K562 cells. We observed a significant decrease in proliferation in the presence of 300 nM apamin and 10 mM TEA after 24 h incubation of K562 cells with channel inhibitors, whereas TRAM-34 also had a tendency to decrease of proliferation but the effect was not statistically significant (p=0.084) at this time point. Further incubation of the cells with specific blockers (for 48 h) allowed to detect a significant reduction of cell number under all experimental conditions as compared to control (Figure 3A). The block of K+ currents by TEA had a most prominent effect on cell proliferation that was almost completely inhibited as compared to control (see Figure 3). Next, we addressed the question if the observed effects of SK/IK blockers or TEA on cell proliferation could be due to the decrease of cell viability. Our experiments demonstrated that there were no statistically significant decreases in a number of viable cells in 300 nM apamin- or 5 mM TRAM-34-treated experimental groups even after 48 h of incubation, whereas 10 mM TEA slightly but significantly different (as compared to control) influenced K562 cell viability after 48 h (Figure 3B).
2.4. Role of SK and IK channels in migration and invasion of leukemia cells
In our last part of experiments, we aimed to elucidate if SK and IK channels may also be implicated in the migration and invasion of human myeloid leukemia К562 cells. We performed the cell migration and invasion assays through microporous transwell membranes (Transwell assay, with 8 μM membrane pore sizes) using serum gradient as a stimulator of cell migration/invasion (see Materials and Methods). To determine K562 invasion, the upper chambers of transwell inserts were coated with Matrigel Matrix (Corning, NY, USA) before the addition of the cells. The inhibitors of SK or IK channel activity (apamin or TRAM-34, respectively) were added to upper and lower wells of the transwell system installed in 4-well cell culture plates. We counted the number of migrated/invaded cells in the lower chamber after 4 h from the start of the experiment. Our data showed that apamin and TRAM-34 inhibited both the migration and invasive abilities of K562 cells. Importantly, the inhibition of cell migration/invasion was observed in the presence of 300 pM of apamin (concentration that affects only SK2 channels). This could indicate the dominant role of SK2 channels in studied processes as no significant differences in the effects of higher apamin concentrations (8 nM and 300 nM) were observed comparing to 300 pM. 5 mM of TRAM-34 had more prominent effect of K562 migration comparing to 1 mM, whereas both concentrations inhibited the invasion to a similar extent (Figure 4).
3. Discussion
In the current study using electrophysiological experiments with selective pharmacological channel inhibitors, we evidenced the presence of functionally active SK2, SK3 and IK channels in human myeloid leukemia K562 cells. It is of specific interest why K562 cells need the functional expression of three different subtypes of KCa channels in the plasma membrane. A possible explanation is that the SK and IK channels are known to be activated by slightly different [Ca2+]i levels: from about 100 nM for IK channels (that is a near-basal [Ca2+]i in most cell types), whereas 300-700 nM of Ca2+ was evidenced to cause 50% activity of SK channels [30,31]. Thus, the presence of both SK and IK channels in the plasma membrane of the same cell allows it to react to a broader range of Ca2+ concentrations and to fine-tune cellular responses to the degree of Ca2+ increase.
We separated the activity of SK channel subtypes and showed the presence of SK2- and SK3-mediated K+ currents utilizing their different sensitivity to apamin. We found the sequential decrease of whole-cell currents in response to the addition of apamin in picomolar and then in nanomolar concentrations. It should be specifically noted that SK channels could co-assemble as a combination of the subtypes (i.e., form hybrid heteromeric SK2/SK3 channel complex, [22,32,33]). Such hybrid channels were reported to have intermediate sensitivity to apamin compared to SK2 and SK3 homomers, and we could not exclude this situation in K562 cells. Nevertheless, the observed inhibition of SK currents both by picomolar and nanomolar ranges of apamin concentration indicate that homomeric SK2 and SK3 channels are definitely present in the plasma membrane.
The presence of mRNA of SK2, SK3 and IK was confirmed by RT-PCR, whereas SK1 mRNA was not detected. Evidently, there is specific importance for the use of combinative approach of molecular biology supplemented with electrophysiological analysis to identify the presence of functionally active ion channels in the plasma membrane. As an illustrative example, in glioma cells, the PCR analysis detected the presence of mRNA of all KCa subtypes (SK1-3, IK and BK channels) [34]. However, only pharmacological inhibitors of BK channels blocked K+ currents and, at the same time, SK/IK inhibitors had no effect that directly indicated the absence of functionally active SK/IK channels in the plasma membrane of glioma cells [34]. Thus, the detection of ion channel mRNA in cell lysates does not guarantee the expression of the channel proteins, and a combinatory approach is necessary to confirm the functional activity of ion channels in the plasma membrane.
In the current work, the immunofluorescent staining using specific antibodies allowed to visualize SK2, SK3 and IK channel proteins in K562 cells, whereas whole-cell patch-clamp experiments clearly evidenced the activity of functional channels in the plasma membrane. It should be noted, that the fluorescent staining was clearly observed in the cytosol of the cells indicating the localization of the channels in intracellular compartments (Figure 1D). Indeed, the presence of SK and IK channels was reported in endoplasmic reticulum, cell nuclei and mitochondria [35] as well as in vesicular transporting organelles [36]. Thus, the observed cytosolic staining could be due to interaction of specific antibodies with intracellular SK2, SK3 and IK channel proteins.
Our results demonstrate that blocking of SK or IK channels in K562 cells reduced their proliferative capabilities without any significant effect on cell viability. Consistently, the inhibition of cell proliferation after blocking of IK currents was previously observed in pancreatic, prostate, colorectal cancer and lung adenocarcinoma cells [37,38,39,40], and was recently reviewed by Mohr et al. [41]. In the paper by Millership et al. [42] it was documented that TRAM-34 reduced cell proliferation of human embryonic kidney (HEK293) cells transfected with recombinant human IK channels. At the same time, the overexpression of IK mutants (that are unable to conduct K+ or with impaired membrane trafficking) increased cell proliferation, thus indicating putative non-channel functions of IK proteins. Importantly, TRAM-sensitive K+ currents were not observed in HEK293 cells with the expression of both mutants indicating that K+-selective transporting pathway formed by IK channels is not crucial for the control of cell proliferation. In further experiments the authors showed the direct interaction of IK proteins with ERK1/2 and JNK pathways [42]. The non-channel function of IK protein as regulator of cell proliferation via ERK1/2/JNK signaling pathways is a very intriguing discovery, and this type of regulation could also work in the K562 leukemia cell line. Nevertheless, our data demonstrate (1) specific channel activity of IK channels in K562 cells that could be selectively inhibited by TRAM-34 in whole-cell patch-clamp assay and (2) the significant decrease of cell proliferation in response to 5 mM TRAM-34. The identification of alternative signaling pathways that could mediate the regulation of cell proliferation by IK channels (and are not linked with their channel function) in K562 cells could be specifically addressed in future studies.
There is some contradictory data on the role of SK channels in proliferation of cancer cells. In mammary cancer cells, no inhibition of cell proliferation was observed in the presence of apamin [43], whereas cell migration was significantly inhibited. At the same time, the inhibition of SK channels by a glycerophospholipid Edelfosine in the urothelial carcinoma cell line resulted in an antiproliferative effect on the cells [44]. Also, apamin was demonstrated to slow down the growth of human melanoma IGR1 cells under hypoxic conditions [45]. In our experiments we have evidenced the apamin-dependent decrease of K562 cell proliferation that could indicate the role of SK channels in the division of leukemia cells. We did not observe any significant changes in cell viability after 24h or 48 h incubation with apamin or TRAM-34. At the same time, treatment of the cells with 10 mM TEA slightly lowered the viability of K562 cells after 48 h. The more significant effect of 10 mM TEA on cancer cell viability was reported earlier [46]: about 50% of cervical cancer HeLa cells were dead after treatment with 10 mM TEA for 48 h. In our assay, 10 mM TEA almost completely blocked cell proliferation after 24 h and 48 h of incubation, whereas negative effect on cell viability was observed only after 48 h. The inhibition of proliferation by TEA was previously shown on C6, 9L glioma, cervical carcinoma SiHA and endometrial adenocarcinoma HEC1-A cell lines [47]. In glioma cells, treatment of the cells with TEA significantly increased the number of reactive oxygen species (ROS) and alteration of B-cell lymphoma protein 2/B-cell lymphoma protein-associated X (Bcl-2/Bax) balance, which further resulted in cell apoptosis [48]. In cervical carcinoma, cell cycle arrest in Go/G1 phase due to inhibition of voltage-gated potassium (Kv) currents by 10 mM TEA was reported [49]. Since TEA could inhibit a broad spectrum of potassium channels from different families, the specific channel targets of TEA as well as signaling pathways affected by TEA treatment in human leukemia cells could hardly be reliably determined. Nevertheless, the role of K+ channels in the proliferation of K562 cells was demonstrated in our experiments.
We observed that apamin or TRAM-34 inhibited both the migratory and invasive abilities of K562 cells. The effect of apamin was prominent at low concentration (300 pM, block of SK2 channels) that could indicate the dominant role of SK2 channels in studied processes. Our data is in principal agreement with several studies, where SK channels were shown to be involved in cell migration of tumor breast tissues [43] and invasion of urothelial carcinoma cells [44]. Also, the inhibition of cancer cell invasion and metastasis by selective IK channel inhibitors, including TRAM-34, was observed for several cancer cell lines [41]. Interestingly, the anomalous effects of TRAM-34 on pancreatic cancer cells were reported [38]. Particularly, 10 μM TRAM-34 was shown to inhibit IK currents, and in the same time, stimulated migration and invasion whereas the proliferation of pancreatic cancer cells decreased. In our experiments, the inhibition of migration and invasion of K562 cells was observed in the presence of 1 μM and 5 μM of TRAM-34, and 5 μM of TRAM-34 had a more prominent effect on cell migration. Thus, IK channels could play a complex role in cancer cell migration, and the potentially unfavorable effects of IK channel inhibition should be taken into account. Most likely, the effects of KCa inhibitors could be cell-type specific and are dependent on the specific signaling pathways in which the channels are integrated.
KCa channels can be considered as an attractive target in pharmacology for the treatment of various diseases and are among FDA-approved drug targets (see KCNN1-4, https://www.proteinatlas.org/search/protein_class%3AFDA+approved+drug+targets/10). For instance, IK channels appear to play a more significant role in the development of different pathological conditions compared the normal physiology [50]. KCa3.1 blocker clotrimazole has been clinically tested for the treatment of rheumatoid arthritis [51] and RBC dehydration in sickle cell disease [52]. Although clotrimazole was effective in treatment, it had many side effects that limited its use. The discovery of clotrimazole analogs TRAM-34 and senicapoc [53], that are more selective for the KCa3.1 channel, may have clinical significance. The intravenous administration of TRAM-34 (0.5 mg/kg; 29 μM) had no toxic effect on CF-1BR mice at a concentration 500-100 times higher than the dose that is sufficient to block IK channels [54]. Moreover, this concentration of TRAM-34 has been shown to have several protective effects on different cells and tissues: immunosuppressive effect, lymphocyte mitogenesis [54] and reverse renal damage on mice [55] and infarction and neurological deficit on ischemia/reperfusion stroke rat model [56]. Therefore, the administration of high doses of TRAM-34 to mice and rats during the week had no toxic effect and minor side effects. Apamin was shown as a potential pharmacological agent to treat apoptosis, fibrosis and the diseases of central nervous system (recently reviewed by Gu et al., [57]). To summarize, the IK and SK channel blockers TRAM-34 and apamin were tested and showed several positive modulatory effects on various diseases. It can be assumed that KCa channels can be used clinically also to treat cancer patients. In our experiments, we observed a decrease of K562 migration and invasion in the presence of selective blockers of SK or IK channels. Our data imply that SK/IK channel inhibitors could be used to slow down the proliferation and spreading of leukemia cells that express functionally active channels in the plasma membrane.
4. Materials and Methods
4.1. Cell Culture and Reagents
Human myeloid leukemia K562 cells were obtained from the shared research facility “Vertebrate cell culture collection” (supported by the Ministry of Science and Higher Education of Russian Federation, Agreement №075-15-2021-683) of the Institute of Cytology (St. Petersburg, Russia). Cells were grown in full RPMI-1640 medium containing 10% fetal bovine serum (FBS, Biolot, St. Petersburg, Russia) and 80 mg/mL antibiotic gentamicin in a humidified incubator at 37 °C and 5% CO2. Before the electrophysiological and immunocytochemical experiments, the cells were plated on poly-DL-lysine-coated (Sigma-Aldrich, St. Louis, MO, USA, Cat. no. P4158) glass coverslips of 0.4 x 0.4 cm or 1.2 x 1.2 cm, respectively. Selective SK blocker (apamin, Santa Cruz Biotechnology, California, CA, USA), and IK blocker (TRAM-34, Tocris, Bristol, UK) were used for the inhibitory analysis in all experiments. Tetraethylammonium bromide (TEA-Br) was purchased from Santa Cruz Biotechnology (Dallas, TX, USA). Stock solutions of apamin and TEA were prepared by the dilution in distilled water, stock solution of TRAM-34 was prepared using DMSO (Sigma-Aldrich, USA). Working solutions were prepared prior to the experiments.
4.2. Electrophysiology
The patch-clamp set-up was based on the Axopatch 200B operational amplifier and Axon Digidata 1550A (Molecular Devices, San Jose, CA, USA) analog–digital converter controlled by a Windows-based personal computer with Axon PClamp 10.7 Software Suite (Molecular Devices LLC, San Jose, CA, USA) for data acquisition, filtration, processing and analysis. Pipettes were pulled from borosilicate glass capillaries with filament (BF-150-86-10, Sutter Instruments, Novato, CA, USA) to a resistance of ~5-10 MΩ when filled with a cytosol-like solution (see Solutions). All experiments were performed at room temperature (22-23 °C). Whole-cell currents were recorded using a whole-cell configuration of the patch-clamp technique (Hamill et al., 1981) according to the following protocol: current traces were obtained from voltage steps from +20 to +80 mV with 10 mV increments (holding potential was 0 mV), Sampling interval was 1.75 s.
4.3. Solutions
For whole-cell experiments, micropipettes were filled with a cytosol-like solution containing (in mM): 140 KAsp, 5 NaCl, 2 EGTA/KOH,1 MgCl2, 20 HEPES/TrisOH with necessary amount of CaCl2 to establish free ionized calcium concentration [Ca2+]i at 1 μM (pCa6). The experimental chamber solution (extracellular) contained (in mM): 145 NaCl, 2 CaCl2, 1 MgCl2 and 10 HEPES/TrisOH. The pH of all solutions was maintained at 7.3.
4.4. Reverse transcription polymerase chain reaction (RT-PCR)
Total RNA was isolated using the RNeasy Mini Kit (Qiagen, Germantown, MD, USA) according to the manufacturer’s instructions. Synthesis of the first-strand cDNA was performed using a commercial kit (OT-1, Syntol, Moscow, Russia). The reverse transcription reaction was carried out by mixing 1 µg of RNA with 0.5 µg random hexaprimers, 100 units reverse transcriptase (revertase) MMLV and 2.5x of the reaction buffer in a total volume of 25 µl followed by incubation at 37°C for 1 hour. For the negative control (RT-), a PCR without MMLV revertase was used. The concentrated cDNA was diluted to a working concentration of 100 ng/ml. The PCR primers were designed using the GeneRunner v5.0.59 software. The primer sequences for amplification of the gene of interest and expected reaction product sizes (bp) are presented in Table 1. PCR was performed in a volume of 15 µl using 1.5 µl diluted cDNA, 0.3 µM each primer, 200 µM dNTP, 2 mM MgCl2, 1 unit Hot-Taq polymerase and 1×Hot-Taq polymerase buffer (Syntol, Moscow, Russia). PCR was performed according to the following scheme: 9 min 30 s at 94°C; 35 cycles of 40 s at 94°C, 30 s at 60 °C and 30 s at 72 °C; 5 min at 72°C. To determine the presence of a gene product, 10 μl of the PCR was subjected to electrophoresis in 6% polyacrylamide gel. The gel was stained with DNA-binding dye GelRed (1:10000, Biotium, Fremont, CA, USA). The results of the reaction were visualized in UV light using the E-Gel Imager System (Thermo Fisher Scientific, Waltham, MA, USA).
4.5. Immunofluorescence
For immunofluorescent staining, specific primary antibodies against the intracellular loop of SK2 (Alomone Labs, Jerusalem, Israel, cat. no. #APC-028), SK3 (Alomone Labs, Jerusalem, Israel, cat. no. #APC-025) channels and extracellular loop of IK channels (conjugated to ATTO Fluor 488 dye, Alomone Labs, Jerusalem, Israel, cat. no. #ALM-051-AG) were used. Cell fixation was performed in 3.7% paraformaldehyde for 10 min at room temperature (RT), then the cells were permeabilized with 0.25% Tween-20 (only before the antibodies against intracellular epitope (SK2 and SK3), for 10 min at RT). Nonspecific binding of the antibodies was blocked by incubating the samples in phosphate-buffered saline (PBS) containing 10% goat serum for 1 hour at RT. Then, cells were incubated with SK2, SK3 or IK-targeting primary antibodies overnight at +4 °C. Staining with fluorescently labeled secondary antibodies (Goat anti-rabbit conjugated to Cy3, Santa Cruz, USA) was performed for detection of SK2 and SK3 channels for 40 min at RT in the dark. Cell nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI, Sigma-Aldrich, St. Louis, MO, USA, 0.05 μg/ml, 30 min at RT). The specificity of antibodies used was determined by eliminating the primary antibodies followed by incubation cells only with secondary antibodies (fluorescent signal was not observed under these conditions). Visualization of the fluorescently labeled preparations were performed on Olympus FV3000 (Olympus, Shinjuku, Tokyo, Japan) confocal microscope. Images were analyzed in ImageJ software (NIH, Bethesda, MD, USA).
4.6. Cell proliferation and viability assays
Cells were seeded in 4-well plates at the concentration of 5×105 cells per well and were incubated with the reagents (apamin, TRAM-34 or TEA) for 48 h. Cell counting was performed at 24 and 48 h time points using Countess II Automated Cell Counter (ThermoFisher, Waltham, Massachusetts, USA). To assess cell viability a Trypan Blue solution (0.4%, Thermo Fisher Scientific, USA) was added, and a number of trypan-positive cells was calculated for each experimental condition.
4.7. Transwell migration and invasion assays
Cell migration and invasion assays were assessed using Transwell inserts (Corning, NY, USA) with semi-permeable microporous membrane (10 µm thickness, 8 µm pore diameter). Before the experiments the inserts were placed into 4-well plates (Jet Biofil, Guangzhou, China). For invasion experiments, transwells were pre-coated with 100 µl of Matrigel (1:40 dilution, 30 min at 37 °C, Corning, NY, USA). Serum gradient (10% FBS in lower well) was used to induce K562 cell migration or invasion. Cells were resuspended in RPMI culture media without FBS. A total volume of 400 µL containing 5×105 cells was placed to the upper well of insert and 750 µL of full RPMI medium was added to the lower well. Cell culture plates were transferred to the incubator (5% CO2, 37 °C) for 4 hours. Selective channel inhibitors were added in the upper and lower well of the transwell system. The number of migrated or invaded cells were counted on Countess II Automated Cell Counter. The number of cells for each experimental condition was normalized to the mean number of cells in control.
4.8. Statistics
The statistical analysis was performed using GraphPad Prism 8.0 software (GraphPad Software). One-way ANOVA with multiple comparisons (to control values) was used to compare the number of cells in the presence of channel inhibitors with control conditions in proliferation/viability/migration and invasion assays. The effects of different concentrations of the reagent were compared using one-way ANOVA with multiple comparisons of the mean values between each concentration tested and to control conditions, p<0.05 was considered as significant.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Vladislav Chubinskiy-Nadezhdin; Data curation, Valeria Vasileva and Vladislav Chubinskiy-Nadezhdin; Formal analysis, Valeria Vasileva, Anastasia Sudarikova and Vladislav Chubinskiy-Nadezhdin; Funding acquisition, Vladislav Chubinskiy-Nadezhdin; Investigation, Valeria Vasileva, Zuleikha Khairullina and Vladislav Chubinskiy-Nadezhdin; Methodology, Valeria Vasileva and Vladislav Chubinskiy-Nadezhdin; Project administration, Vladislav Chubinskiy-Nadezhdin; Resources, Anastasia Sudarikova and Vladislav Chubinskiy-Nadezhdin; Supervision, Valeria Vasileva and Vladislav Chubinskiy-Nadezhdin; Validation, Valeria Vasileva, Anastasia Sudarikova and Vladislav Chubinskiy-Nadezhdin; Visualization, Valeria Vasileva, Zuleikha Khairullina, Anastasia Sudarikova and Vladislav Chubinskiy-Nadezhdin; Writing – original draft, Valeria Vasileva, Zuleikha Khairullina, Anastasia Sudarikova and Vladislav Chubinskiy-Nadezhdin; Writing – review & editing, Valeria Vasileva, Anastasia Sudarikova and Vladislav Chubinskiy-Nadezhdin.
Funding
This research was funded by the Russian Science Foundation (RSF grant № 22-74-10037, https://rscf.ru/en/project/22-74-10037/).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available within the article and upon request.
Conflicts of Interest
The authors declare no conflict of interest. The funder had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Arcangeli, A.; Becchetti, A. Novel perspectives in cancer therapy: Targeting ion channels. Drug Resist Updat 2015, 21-22, 11-9. [CrossRef]
- Rafieemehr, H.; Samimi, A.; Maleki Behzad, M.; Ghanavat, M.; Shahrabi, S. Altered expression and functional role of ion channels in leukemia: bench to bedside. Clin Transl Oncol 2020, 22(3), 283-293. [Google Scholar] [CrossRef]
- Köhler, M.; Hirschberg, B.; Bond, C.T.; Kinzie, J.M.; Marrion, N.V.; Maylie, J.; Adelman, J.P. Small-conductance, calcium-activated potassium channels from mammalian brain. Science 1996, 273(5282), 1709-14. [Google Scholar] [CrossRef]
- Pedarzani, P.; Stocker., M. Molecular and cellular basis of small- and intermediate-conductance, calcium-activated potassium channel function in the brain. Cell Mol Life Sci 2008, 65, 3196-3217. [CrossRef]
- Brown, B.M.; Shim, H.; Christophersen, P.; Wulff, H. Pharmacology of Small- and Intermediate-Conductance Calcium-Activated Potassium Channels. Annu Rev Pharmacol Toxicol 2020, 60, 219-240. [CrossRef]
- Kuzmenkov, A.I.; Peigneur, S.; Nasburg, J.A.; Mineev, K.S.; Nikolaev, M.V.; Pinheiro-Junior, E.L.; Arseniev, A.S.; Wulff, H.; Tytgat, J.; Vassilevski, A.A. Apamin structure and pharmacology revisited. Front Pharmacol 2022, 13, 977440. [CrossRef]
- Agarwal, J.J.; Zhu, Y.; Zhang, Q.Y.; Mongin, A.A.; Hough, L.B. TRAM-34, a putatively selective blocker of intermediate-conductance, calcium-activated potassium channels, inhibits cytochrome P450 activity. PLoS One 2013, 8(5), e63028. [CrossRef]
- Wulff, H.; Kolski-Andreaco, A.; Sankaranarayanan, A.; Sabatier, J.M.; Shakkottai, V. Modulators of small- and intermediate-conductance calcium-activated potassium channels and their therapeutic indications. Curr Med Chem 2007, 14(13), 1437-57. [CrossRef]
- Gardos, G. The function of calcium in the potassium permeability of human erythrocytes. Biochim Biophys Acta 1958, 30(3), 653-4. [CrossRef]
- Fanger, C.M.; Rauer, H.; Neben, A.L.; Miller, M.J.; Rauer, H.; Wulff, H.; Rosa, J.C.; Ganellin, C.R.; Chandy, K.G.; Cahalan, M.D. Calcium-activated potassium channels sustain calcium signaling in T lymphocytes. Selective blockers and manipulated channel expression levels. J Biol Chem 2001, 276(15), 12249-56. [Google Scholar] [CrossRef]
- Feske, S.; Skolnik, E.Y.; Prakriya, M. Ion channels and transporters in lymphocyte function and immunity. Nat Rev Immunol 2012, 12(7), 532-47. [Google Scholar] [CrossRef]
- Valle-Reyes, S.; Valencia-Cruz, G.; Liñan-Rico, L.; Pottosin, I.; Dobrovinskaya, O. Differential Activity of Voltage- and Ca2+-Dependent Potassium Channels in Leukemic T Cell Lines: Jurkat Cells Represent an Exceptional Case. Front Physiol 2018, 9, 499. [CrossRef]
- Kaestner, L.; Bogdanova, A.; Egee, S. Calcium Channels and Calcium-Regulated Channels in Human Red Blood Cells. Adv Exp Med Biol 2020, 1131, 625-648. [CrossRef]
- Fermo, E.; Bogdanova, A.; Petkova-Kirova, P.; Zaninoni, A.; Marcello, A.P.; Makhro, A.; Hänggi, P.; Hertz, L.; Danielczok, J.; Vercellati, C.; Mirra, N.; Zanella, A.; Cortelezzi, A.; Barcellini, W.; Kaestner, L.; Bianchi, P. 'Gardos Channelopathy': a variant of hereditary Stomatocytosis with complex molecular regulation. Sci Rep 2017, 7(1), 1744. [CrossRef]
- Dyrda, A.; Cytlak, U.; Ciuraszkiewicz, A.; Lipinska, A.; Cueff, A.; Bouyer, G.; Egée, S.; Bennekou, P.; Lew, V.L.; Thomas, S.L. Local membrane deformations activate Ca2+-dependent K+ and anionic currents in intact human red blood cells. PLoS One 2010, 5(2), e9447. [CrossRef]
- Cahalan, S.M.; Lukacs, V.; Ranade, S.S.; Chien, S.; Bandell, M.; Patapoutian, A. Piezo1 links mechanical forces to red blood cell volume. Elife 2015, 4, e07370. [CrossRef]
- Staruschenko, A.V.; Vedernikova, E.A. Mechanosensitive cation channels in human leukaemia cells: calcium permeation and blocking effect. J Physiol 2002, 541(Pt 1), 81-90. [CrossRef]
- Staruschenko, A.; Negulyaev, Y.A.; Morachevskaya, E.A. Actin cytoskeleton disassembly affects conductive properties of stretch-activated cation channels in leukaemia cells. Biochim Biophys Acta 2005, 1669(1), 53-60. [CrossRef]
- Chubinskiy-Nadezhdin, V.I.; Negulyaev, Y.A.; Morachevskaya, E.A. Cholesterol depletion-induced inhibition of stretch-activated channels is mediated via actin rearrangement. Biochem Biophys Res Commun 2011, 412(1), 80-5. [CrossRef]
- Chubinskiy-Nadezhdin, V.I.; Negulyaev, Y.A.; Morachevskaya, E.A. Functional Coupling of Ion Channels in the Process of Mechano-Dependent Activation in the Membrane of K562 Cells. Cell Tiss. Biol 2019, 13, 470–477. [CrossRef]
- Guéguinou, M.; Chantôme, A.; Fromont, G.; Bougnoux, P.; Vandier, C.; Potier-Cartereau, M. KCa and Ca(2+) channels: the complex thought. Biochim Biophys Acta 2014, 1843(10), 2322-33. [CrossRef]
- Weatherall, K.L.; Seutin, V.; Liégeois, J.F.; Marrion, N.V. Crucial role of a shared extracellular loop in apamin sensitivity and maintenance of pore shape of small-conductance calcium-activated potassium (SK) channels. Proc Natl Acad Sci U S A 2011, 108(45), 18494-9. [CrossRef]
- Lamy, C.; Goodchild, S.J.; Weatherall, K.L.; Jane, D.E.; Liégeois, J.F.; Seutin, V.; Marrion, N.V. Allosteric block of KCa2 channels by apamin. J Biol Chem 2010, 285(35), 27067-27077. [CrossRef]
- Wulff, H.; Gutman, G.A.; Cahalan, M.D.; Chandy, K.G. Delineation of the clotrimazole/TRAM-34 binding site on the intermediate conductance calcium-activated potassium channel, IKCa1. J Biol Chem 2001, 276(34), 32040-5. [CrossRef]
- Maylie, J.; Bond, C.T.; Herson, P.S.; Lee, W.S.; Adelman, J.P. Small conductance Ca2+-activated K+ channels and calmodulin. J Physiol 2004, 554(Pt 2), 255-61. [CrossRef]
- Qi, M.M.; Qian, L.L.; Wang, R.X. Modulation of SK Channels: Insight Into Therapeutics of Atrial Fibrillation. Heart Lung Circ 2021, 30(8),1130-1139. [CrossRef]
- Bong, A.H.L.; Monteith, G.R. Calcium signaling and the therapeutic targeting of cancer cells. Biochim Biophys Acta Mol Cell Res 2018, 1865(11 Pt B), 1786-1794. [CrossRef]
- Wu, L.; Lian, W.; Zhao, L. Calcium signaling in cancer progression and therapy. FEBS J 2021, 288(21), 6187-6205. [CrossRef]
- Khodakhah, K.; Melishchuk, A.; Armstrong, C.M. Killing K channels with TEA+. Proc Natl Acad Sci U S A 1997, 94(24), 13335-8. [CrossRef]
- Xia, X.M.; Fakler, B.; Rivard, A.; Wayman, G.; Johnson-Pais, T.; Keen, J.E.; Ishii, T.; Hirschberg, B.; Bond, C.T.; Lutsenko, S.; Maylie, J.; Adelman, J.P. Mechanism of calcium gating in small-conductance calcium-activated potassium channels. Nature 1998, 395(6701), 503-7. [CrossRef]
- Joiner, W.J.; Wang, L.Y.; Tang, M.D.; Kaczmarek, L.K. hSK4, a member of a novel subfamily of calcium-activated potassium channels. Proc Natl Acad Sci U S A 1997, 94(20), 11013-8. [CrossRef]
- Strassmaier, T.; Bond, C.T.; Sailer, C.A.; Knaus, H.G.; Maylie, J.; Adelman, J.P. A novel isoform of SK2 assembles with other SK subunits in mouse brain. J Biol Chem 2005, 280(22), 21231-6. [CrossRef]
- Girault, A.; Haelters, J.P.; Potier-Cartereau, M.; Chantôme, A.; Jaffrés, P.A.; Bougnoux, P.; Joulin, V.; Vandier, C. Targeting SKCa channels in cancer: potential new therapeutic approaches. Curr Med Chem 2012, 19(5),697-713. [CrossRef]
- Weaver, A.K.; Bomben, V.C.; Sontheimer, H. Expression and function of calcium-activated potassium channels in human glioma cells. Glia 2006, 54(3), 223-33. [CrossRef]
- Checchetto, V.; Teardo, E.; Carraretto, L.; Leanza, L.; Szabo, I. Physiology of intracellular potassium channels: A unifying role as mediators of counterion fluxes? Biochimica et biophysica acta 2016, 1857(8), 1258–1266. [CrossRef]
- Capera, J.; Serrano-Novillo, C.; Navarro-Pérez, M.; Cassinelli, S.; Felipe, A. The Potassium Channel Odyssey: Mechanisms of Traffic and Membrane Arrangement. International journal of molecular sciences 2019, 20(3), 734. [CrossRef]
- Lallet-Daher, H.; Roudbaraki, M.; Bavencoffe, A.; Mariot, P.; Gackière, F.; Bidaux, G.; Urbain, R.; Gosset, P.; Delcourt, P.; Fleurisse, L.; Slomianny, C.; Dewailly, E.; Mauroy, B.; Bonnal, J.L.; Skryma, R.; Prevarskaya, N. Intermediate-conductance Ca2+-activated K+ channels (IKCa1) regulate human prostate cancer cell proliferation through a close control of calcium entry. Oncogene 2009, 28(15), 1792-806. [CrossRef]
- Bonito, B.; Sauter, D.R.; Schwab, A.; Djamgoz, M.B.; Novak, I. KCa3.1 (IK) modulates pancreatic cancer cell migration, invasion and proliferation: anomalous effects on TRAM-34. Pflugers Arch 2016, 468(11-12), 1865-1875. [CrossRef]
- Lai, W.; Chen, S.; Wu, H.; Guan, Y.; Liu, L.; Zeng, Y.; Zhao, H.; Jiang, J.; Chu, Z. PRL-3 promotes the proliferation of LoVo cells via the upregulation of KCNN4 channels. Oncol Rep 2011, 26(4), 909-17. [CrossRef]
- Bulk, E.; Ay, A.S.; Hammadi, M.; Ouadid-Ahidouch, H.; Schelhaas, S.; Hascher, A.; Rohde, C.; Thoennissen, N.H.; Wiewrodt, R.; Schmidt, E.; Marra, A.; Hillejan, L.; Jacobs, A.H.; Klein, H.U.; Dugas, M.; Berdel, W.E.; Müller-Tidow, C.; Schwab, A. Epigenetic dysregulation of KCa 3.1 channels induces poor prognosis in lung cancer. Int J Cancer 2015, 137(6), 1306-17. [CrossRef]
- Mohr, C.J.; Steudel, F.A.; Gross, D.; Ruth, P.; Lo, W.Y.; Hoppe, R.; Schroth, W.; Brauch, H.; Huber, S.M.; Lukowski, R. Cancer-Associated Intermediate Conductance Ca2+-Activated K⁺ Channel KCa3.1. Cancers (Basel) 2019, 11(1), 109. [CrossRef]
- Millership, J.E.; Devor, D.C.; Hamilton, K.L.; Balut, C.M.; Bruce, J.I.; Fearon, I.M. Calcium-activated K+ channels increase cell proliferation independent of K+ conductance. Am J Physiol Cell Physiol 2011, 300(4), C792-802. [CrossRef]
- Potier, M.; Joulin, V.; Roger, S.; Besson, P.; Jourdan, M.L.; Leguennec, J.Y.; Bougnoux, P.; Vandier, C. Identification of SK3 channel as a new mediator of breast cancer cell migration. Mol Cancer Ther 2006, 5(11), 2946-53. [CrossRef]
- Steinestel, K.; Eder, S.; Ehinger, K.; Schneider, J.; Genze, F.; Winkler, E.; Wardelmann, E.; Schrader, A.J.; Steinestel, J. The small conductance calcium-activated potassium channel 3 (SK3) is a molecular target for Edelfosine to reduce the invasive potential of urothelial carcinoma cells. Tumour Biol 2016, 37(5), 6275-83. Erratum in: Tumour Biol 2016, 37(2), 2773. [CrossRef]
- Tajima, N.; Schönherr, K.; Niedling, S.; Kaatz, M.; Kanno, H.; Schönherr, R.; Heinemann, S.H. Ca2+-activated K+ channels in human melanoma cells are up-regulated by hypoxia involving hypoxia-inducible factor-1alpha and the von Hippel-Lindau protein. J Physiol 2006, 571(Pt 2), 349-59. [CrossRef]
- Huang, L.; Huang, Q.Y.; Huang, H.Q. The evidence of HeLa cell apoptosis induced with tetraethylammonium using proteomics and various analytical methods. J Biol Chem 2014, 289(4), 2217-29. [CrossRef]
- Erdem Kış, E.; Tiftik, R.N.; Al Hennawi, K.; Ün, İ. The role of potassium channels in the proliferation and migration of endometrial adenocarcinoma HEC1-A cells. Mol Biol Rep 2022, 49(8), 7447-7454. [CrossRef]
- Yang, K.B.; Zhao, S.G.; Liu, Y.H.; Hu, E.X.; Liu, B.X. Tetraethylammonium inhibits glioma cells via increasing production of intracellular reactive oxygen species. Chemotherapy 2009, 55(5), 372-80. [CrossRef]
- Han, X.B.; Wang, F.; Yao, W.X.; Liu, Y.Q.; Wang, G.; Ma, D. Inhibitory effects of tetraethylammonium on proliferation and voltage-gated potassium channels in human cervical carcinoma cell line SiHa. Ai Zheng 2006, 25(4), 451-5. Chinese.
- Roach, K. M.; Bradding, P. Ca2+ signalling in fibroblasts and the therapeutic potential of KCa3.1 channel blockers in fibrotic diseases. British journal of pharmacology 2020,177(5), 1003–1024. [CrossRef]
- Wojtulewski, J. A.; Gow, P. J.; Walter, J.; Grahame, R.; Gibson, T.; Panayi, G. S.; Mason, J. Clotrimazole in rheumatoid arthritis. Annals of the rheumatic diseases 1980, 39(5), 469–472. [CrossRef]
- Brugnara, C.; Gee, B.; Armsby, C. C.; Kurth, S.; Sakamoto, M.; Rifai, N.; Alper, S. L.; Platt, O. S. Therapy with oral clotrimazole induces inhibition of the Gardos channel and reduction of erythrocyte dehydration in patients with sickle cell disease. The Journal of clinical investigation 1996, 97(5), 1227–1234. [CrossRef]
- Ataga, K. I.; Smith, W. R.; De Castro, L. M.; Swerdlow, P.; Saunthararajah, Y.; Castro, O.; Vichinsky, E.; Kutlar, A.; Orringer, E. P.; Rigdon, G. C.; Stocker, J. W. ICA-17043-05 Investigators. Efficacy and safety of the Gardos channel blocker, senicapoc (ICA-17043), in patients with sickle cell anemia. Blood 2008,111(8), 3991–3997. [CrossRef]
- Wulff, H.; Miller, M. J.; Hansel, W.; Grissmer, S.; Cahalan, M. D.; Chandy, K. G. Design of a potent and selective inhibitor of the intermediate-conductance Ca2+-activated K+ channel, IKCa1: a potential immunosuppressant. Proceedings of the National Academy of Sciences of the United States of America 2000, 97(14), 8151–8156. [CrossRef]
- Huang, C.; Zhang, L.; Shi, Y.; Yi, H.; Zhao, Y.; Chen, J.; Pollock, C. A.; Chen, X. M. The KCa3.1 blocker TRAM34 reverses renal damage in a mouse model of established diabetic nephropathy. PloS one 2018, 13(2), e0192800. [CrossRef]
- Chen, Y. J.; Raman, G.; Bodendiek, S.; O'Donnell, M. E.; Wulff, H. The KCa3.1 blocker TRAM-34 reduces infarction and neurological deficit in a rat model of ischemia/reperfusion stroke. Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism 2011, 31(12), 2363–2374. [CrossRef]
- Gu, H.; Han, S. M.; Park, K. K. Therapeutic Effects of Apamin as a Bee Venom Component for Non-Neoplastic Disease. Toxins 2020, 12(3), 195. [CrossRef]
Figure 1.
Ca2+-dependent potassium (KCa) channels in the plasma membrane of K562 cells. (A) Representative whole-cell currents in the control conditions (pCa6) and after subsequent addition of apamin (300 nM, selective inhibitor of SK channels) and TRAM-34 (1 μM, inhibitor of IK channels) recorded in the range of holding membrane potentials from +20 mV to +80 mV, with 10 mV increments. (B) The mean (± SEM) whole-cell currents decreased after the subsequent addition of apamin and TRAM-34. (C) RT-PCR analysis of KCa channel mRNA expression: M – size marker, KCNN2 (SK2) – 423 bp, KCNN3 (SK3) – 134 bp, KCNN4 (IK) – 325 bp; KCNN1 (SK1, 282 bp) was not detected. Shown are cropped gels with enhanced contrast. Original gels are available as Supplementary Figure S1. (D) Immunofluorescent staining confirmed the presence of SK2, SK3 and IK proteins in K562 cells. Blue channel – cell nuclei (DAPI), red channel – SK2 and SK3, green channel – IK. Scale bar is 30 µm.
Figure 1.
Ca2+-dependent potassium (KCa) channels in the plasma membrane of K562 cells. (A) Representative whole-cell currents in the control conditions (pCa6) and after subsequent addition of apamin (300 nM, selective inhibitor of SK channels) and TRAM-34 (1 μM, inhibitor of IK channels) recorded in the range of holding membrane potentials from +20 mV to +80 mV, with 10 mV increments. (B) The mean (± SEM) whole-cell currents decreased after the subsequent addition of apamin and TRAM-34. (C) RT-PCR analysis of KCa channel mRNA expression: M – size marker, KCNN2 (SK2) – 423 bp, KCNN3 (SK3) – 134 bp, KCNN4 (IK) – 325 bp; KCNN1 (SK1, 282 bp) was not detected. Shown are cropped gels with enhanced contrast. Original gels are available as Supplementary Figure S1. (D) Immunofluorescent staining confirmed the presence of SK2, SK3 and IK proteins in K562 cells. Blue channel – cell nuclei (DAPI), red channel – SK2 and SK3, green channel – IK. Scale bar is 30 µm.
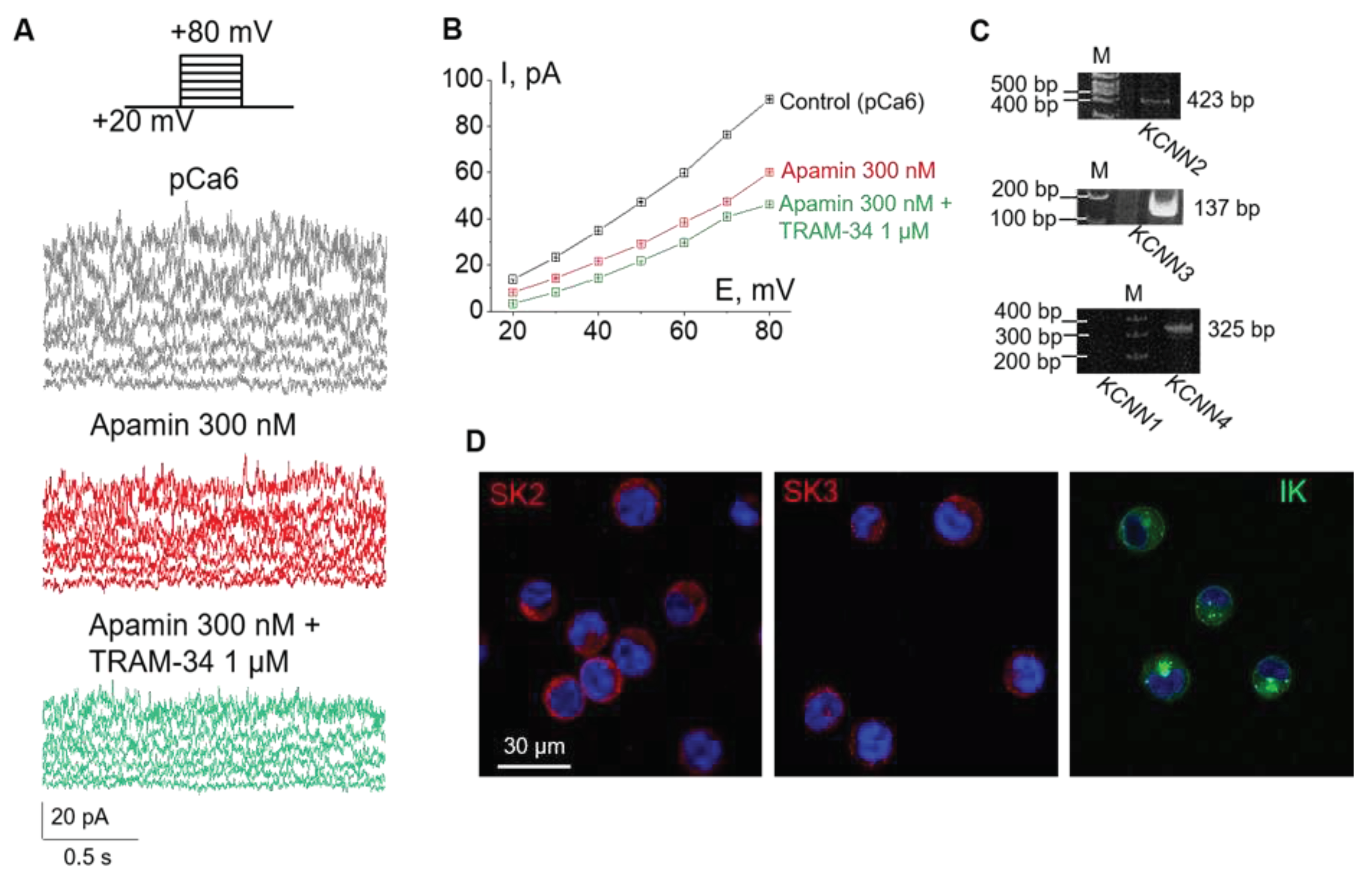
Figure 2.
Pharmacological separation of SK2 and SK3 activity in the plasma membrane of K562 cells due to different sensitivity of SK channel subtypes to apamin. (A) Representative whole-cell currents recorded in control conditions (pCa 6) and in the presence of 300 pM (blocks SK2 currents) or 8 nM (blocks SK3 currents) of apamin. (B) The mean (± SEM) whole-cell currents in control and after the addition of apamin (300 pM and 8 nM).
Figure 2.
Pharmacological separation of SK2 and SK3 activity in the plasma membrane of K562 cells due to different sensitivity of SK channel subtypes to apamin. (A) Representative whole-cell currents recorded in control conditions (pCa 6) and in the presence of 300 pM (blocks SK2 currents) or 8 nM (blocks SK3 currents) of apamin. (B) The mean (± SEM) whole-cell currents in control and after the addition of apamin (300 pM and 8 nM).
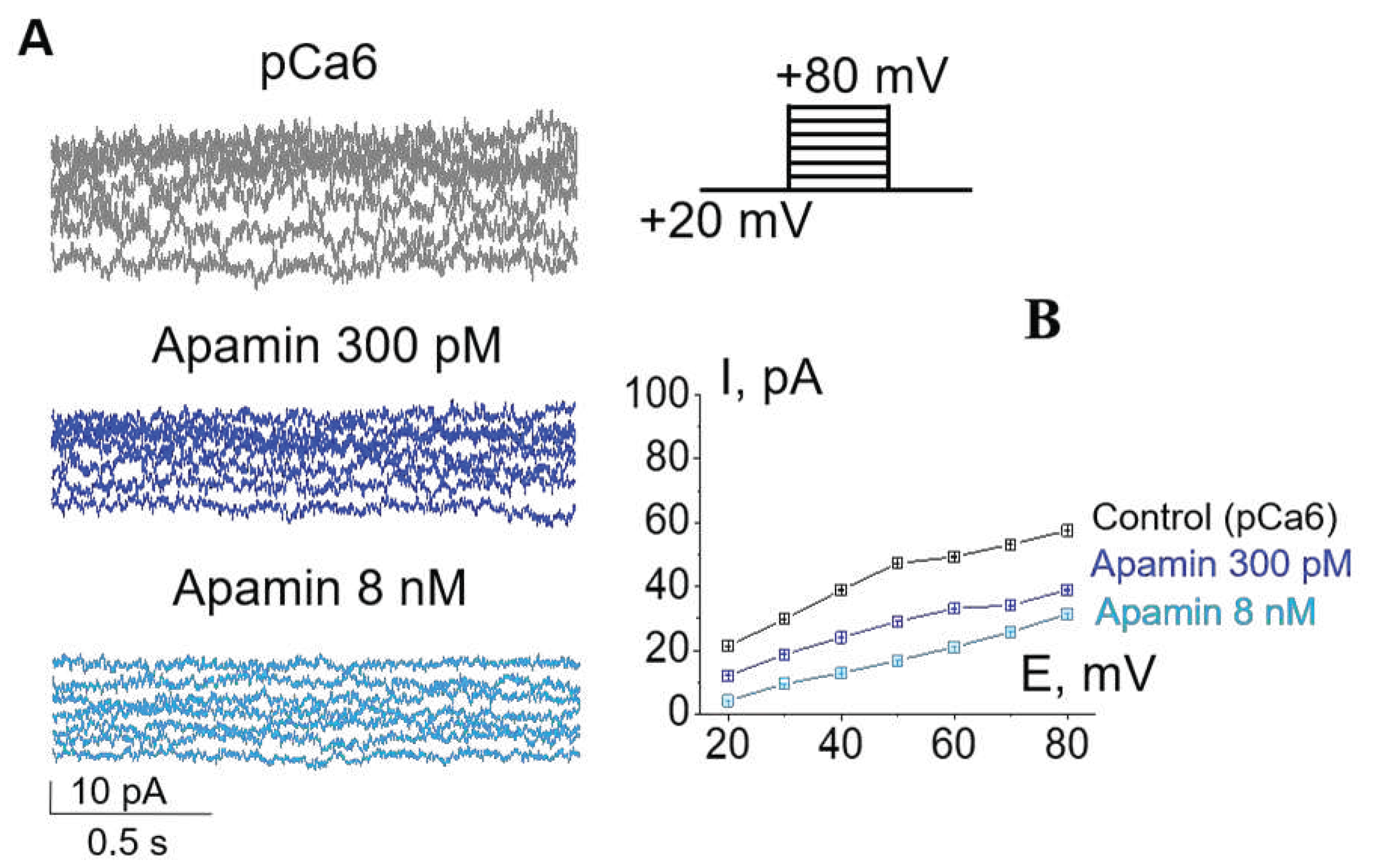
Figure 3.
Effects of KCa inhibitors on the proliferation and viability of leukemia K562 cells. (A) The bar graph represents the changes in the number of cells after 24 h and 48 h from the addition of selective KCa inhibitors. 500 000 cells were seeded at the starting point (0 h). Tetraethylammonium (TEA) was used as a non-selective blocker of K+ channels. Shown is the mean number of cells ± SD (n = 8-14) in each experimental condition. *(p<0.05), ** (p<0.01), *** (p<0.001), **** (p<0.0001) significantly different compared to control, Student’s t-test. (B) No significant changes (ns) in the viability of cells were observed in the presence of selective KCa inhibitors. Note the slight decrease (88.9% viable with TEA vs 96.3% in control) of K562 viability by TEA after 48 h. Shown is the mean percent ± SD (n=4). **** (p<0.0001) significantly different compared to control, One-way ANOVA.
Figure 3.
Effects of KCa inhibitors on the proliferation and viability of leukemia K562 cells. (A) The bar graph represents the changes in the number of cells after 24 h and 48 h from the addition of selective KCa inhibitors. 500 000 cells were seeded at the starting point (0 h). Tetraethylammonium (TEA) was used as a non-selective blocker of K+ channels. Shown is the mean number of cells ± SD (n = 8-14) in each experimental condition. *(p<0.05), ** (p<0.01), *** (p<0.001), **** (p<0.0001) significantly different compared to control, Student’s t-test. (B) No significant changes (ns) in the viability of cells were observed in the presence of selective KCa inhibitors. Note the slight decrease (88.9% viable with TEA vs 96.3% in control) of K562 viability by TEA after 48 h. Shown is the mean percent ± SD (n=4). **** (p<0.0001) significantly different compared to control, One-way ANOVA.
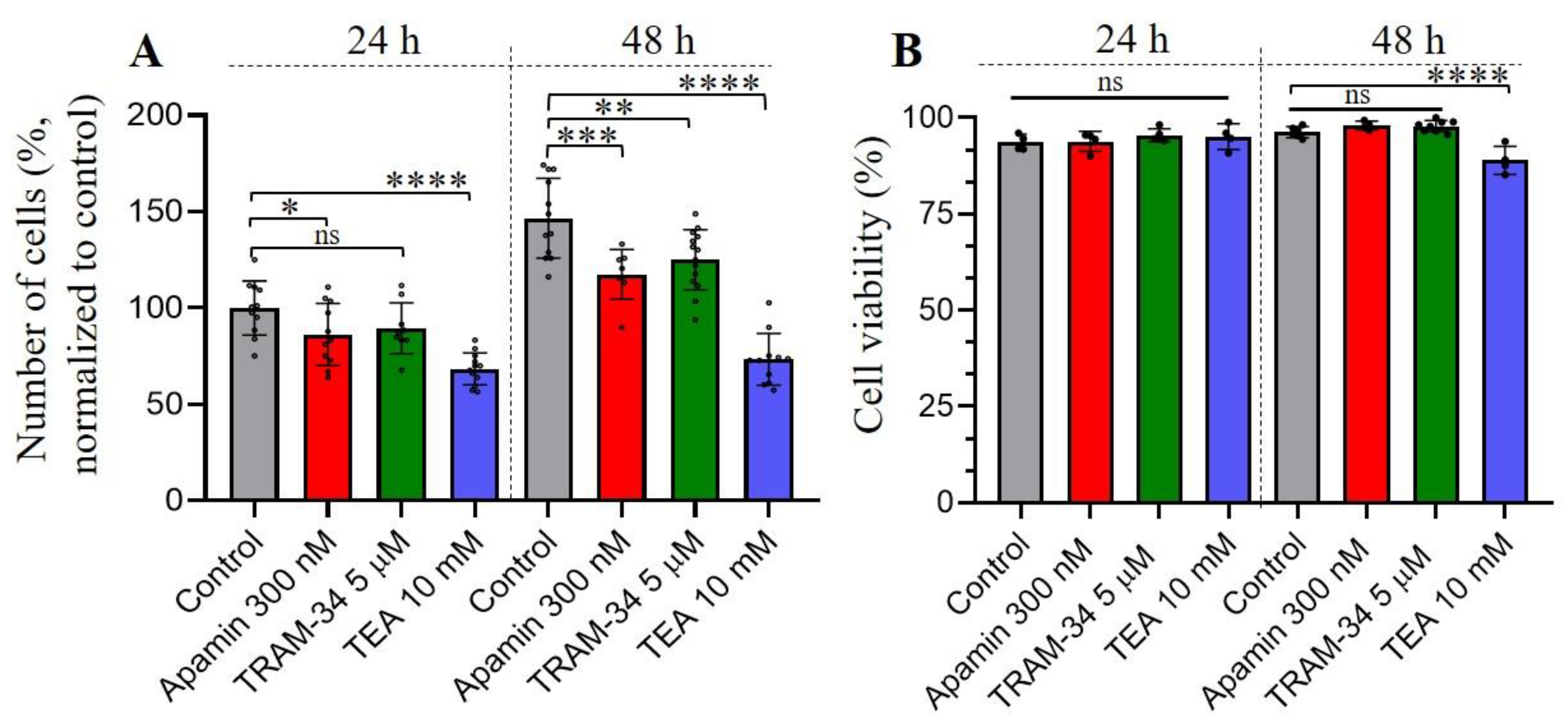
Figure 4.
Effects of KCa inhibitors on the migration (A) and invasion (B) of leukemia K562 cells. The selective inhibitors were added to the culture media in upper and lower chambers. The graphs summarize the results of independent assays, each of the values within each assay were normalized to respective control values. The percent of migrated (A) and invaded (B) cells is presented as means ± SD, all datapoints are shown on the graph. ns - not significant, **** (p<0.0001) significantly different, One-way ANOVA test.
Figure 4.
Effects of KCa inhibitors on the migration (A) and invasion (B) of leukemia K562 cells. The selective inhibitors were added to the culture media in upper and lower chambers. The graphs summarize the results of independent assays, each of the values within each assay were normalized to respective control values. The percent of migrated (A) and invaded (B) cells is presented as means ± SD, all datapoints are shown on the graph. ns - not significant, **** (p<0.0001) significantly different, One-way ANOVA test.
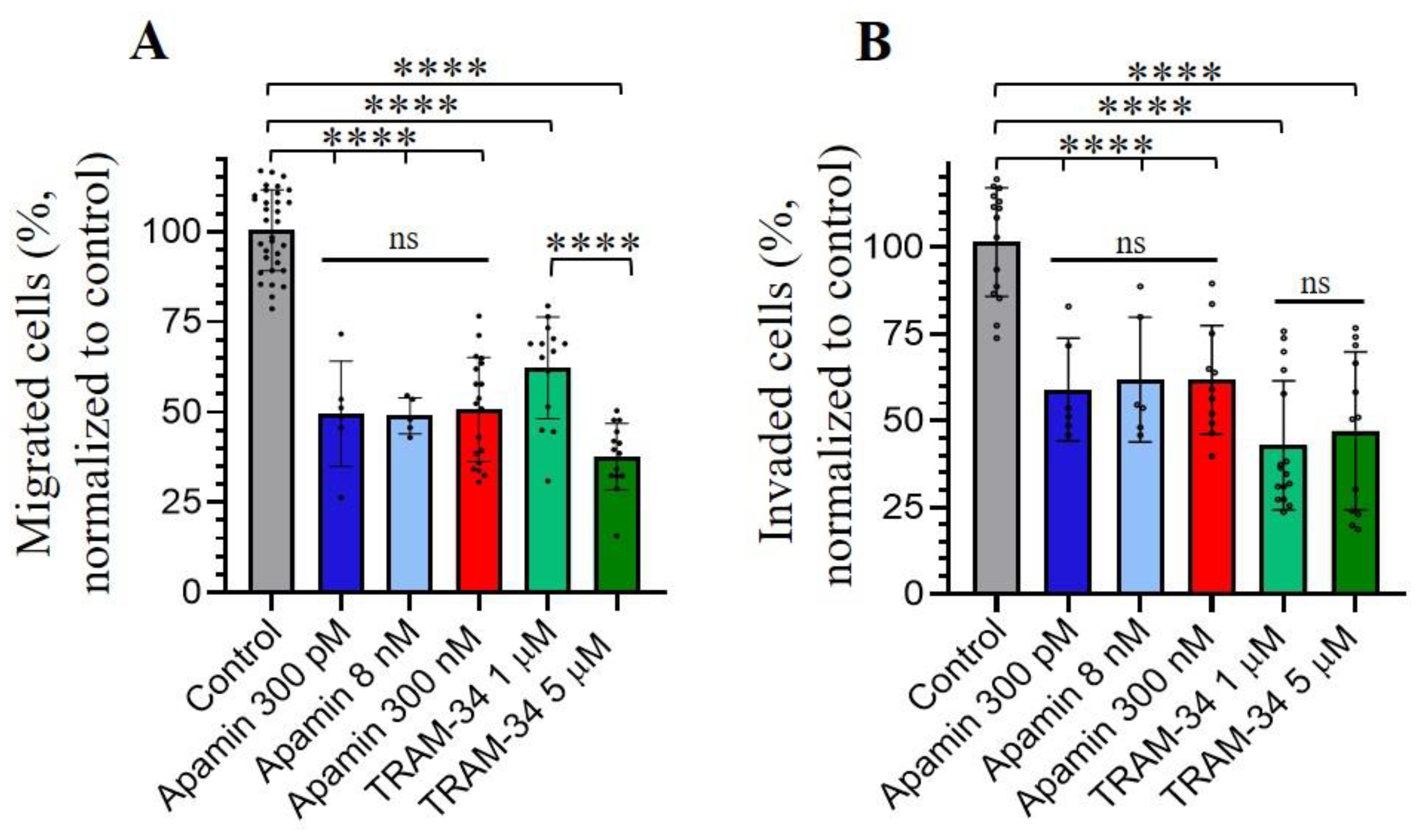

Table 1.
Primer sequences for KCNN1-4 genes used in this study.
| Gene (KCa type) |
Forward primer Reverse primer |
Predicted product size, bp |
|---|---|---|
|
hKCNN1 (KCa2.1, SK1) |
3’-AGA ACA GCA AGA CAT ATC CG-5’ 5’-ATT GTA GCT GTG GCT GTT CA-3’ |
282 |
|
hKCNN2 (KCa2.2, SK2) |
3’-CTT ATC AGT CTC TCC ACG ATC-5’ 5’-TAC AGT TCC TGG GCA TAT AG-3’ |
423 |
|
hKCNN3 (KCa2.3, SK3) |
3’-CGA CTG AGT GAC TAT GCT C-5’ 5’-GTG GAC AGA CTG ATA AGG C-3’ |
137 |
|
hKCNN4 (KCa3.1, IK) |
5’-ATC ATG AAG TTG TGC ACG TG-3’ 5’-ATC ATG AAG TTG TGC ACG TG-3’ |
325 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



