
Submitted:
23 January 2023
Posted:
27 January 2023
You are already at the latest version
Abstract
Whether the different NR-AMPs could ever be utilized as drugs not only against prokaryotic (bacteria) pathogens but eukaryotic (fungal pathogens, and parasitic protists) depends on the side effects. To get experimental experience about the option of applying EPB-produced antimicrobials to pathogens, and parasites of veterinary significance, we present here the results of an in vitro, and an accompanying in vivo study on chicken. In the in vitro study, we tested the cytotoxic potential of the cell-free conditioned culture media (CFCM) of three entomopathogenic bacterium species, - X. budapestensis, DSM16342 (EMA); X. szentirmaii DSM16338 (EMC); Photorhabdus luminescens ssp. akhurstii TT01 - on chicken tissue culture cells, namely, on the Leghorn Male Hepatoma (LMH), [92] cells, (a permanent confluent hepato-carcinoma cell line). Each CFCM proved rather cytotoxic in this test. In the in vivo study, we fed freshly hatched male broiler chickens for 42 days with XENOFOOD [39] which contained autoclaved cultures of EMA, and EMC). These bacteria were grown on standard chicken (starter and grower) [HM3] [u4] feed, and the whole culture was used as a “food supplement”. [HM5]. It had been known that these EPB species cannot grow that is, not viable) atbody temperature (above 33 C).
Keywords:
multidrug resistance
; MDR
; EPB (Xenorhabdus
; (X. budapestensis
; X. szentirmaii
; X. innexii)
; and Photorhabdus) species)
; CFCM (cell-free conditioned media)
; NR-AMP (non-ribosomal-templated antimicrobial peptides)
; anti-microbial
; (anti-bacterial
; - anti-coccidial
; -anti-protist
; - activity
; cytotoxicity
; in vitro
; in situ (local) bioavailablity
; XENOFOOD
; allometry
1. Introduction
The emergence, and re-emergence of diseases caused by multidrug-resistant (MDR) pathogens, and parasitic organisms in plants, invertebrates, vertebrate animals, and humans provide arguments for the urgent search for new antimicrobial-active drugs with novel modes of action [1,2,3,4,5,6,7,8,9,10,11,12].This study intended to contribute to the research on the drug potential of natural biosynthetic antimicrobial peptides as chemotherapeutic tools against multi-drug resistant pathogens [HM1] [u2] [13]. The term “antimicrobial peptides” (AMPs) [14] includes any polyamide (or even biopolymer with ester, thioester, or otherwise modified backbone) that can be made on a contemporary chemical peptide synthesizer. The limit in size is greater than the arbitrary cutoff of 50 amino acids set up by the US Food and Drug Administration [15] for proteins and far exceeds that of biological recognition elements [14]. Therefore, not only the gene-encoded, ribosomal templated antimicrobial peptides, (RT-AMPs) but other peptides of antimicrobial activity, including enzymatically (bio)-synthesized non-ribosomal templated antimicrobial peptide (NR-AMP) molecules should also be considered. The AMPs are of great perspectives to combat MDR prokaryotes because antibiotic-resistant bacteria perform a high frequency of collateral sensitivity to antimicrobial peptides, [16]. Furthermore, the mobility patterns of AMP-resistance genes differ from those of the antibiotics-resistance genes [17]. RT-[HM2]. AMPs are produced by all but Archea taxa, [18], and considered innate components of the innate immune systems of all known eukaryotic organisms, [19]. The RT-AMPs are usually narrow target spectral drugs, [20], but their target spectrum can be extended in vivo [21] probably due to their immune-modulatory actions,[22,23). The “secrete” of their future drug potential is their molecular versatility, allowing QSAR modeling [24], and computer-aided design of antimicrobial peptide analogs, and peptides [18],u1] ] [25], and benefiting from the well-suited and powerful tool of proteomics, for better understanding the respective study molecular responses to antimicrobial compounds, [26]. The vast majority of MPs are membrane-active [25] Others penetrate and bind to intracellular targets like proline-rich PrAMPs, [27].
Several AMPs proved efficient against pathogenic-parasitic Leishmania and Trypanosoma species.
Unlike those of the RT-AMPs, the target spectrum of most NRP-AMPs is usually large, [28] [29]). In eukaryotic targets, some induce apoptotic death of leishmanial protist parasites through calcium-dependent, caspase-independent mitochondrial toxicity mechanism [30].
Entomopathogenic bacteria (abbreviation EPBs), the obligate symbionts of entomopathogenic nematode (EPN) species synthesize and release NRP-hybrid [HM1] [u2] [u3] peptides that provide well-balanced pathobiome conditions for the respective EPN/EPB symbiotic complex in polyxenic (insect cadaver in the soil) environments [31,32]. These bacteria are considered potential sources of potent natural anti-microbial [31], and anti-protist [33] compounds. We have previously found antimicrobial active secreted peptides produce by Xenorhabdus budapestensis and X. szentirmaii [34,35] in both solid, and liquid media that were active against [HM1] [u2] Gram-positive and -negative pathogenic bacteria, [34,36]; anti-plant pathogenic bacterial [37,38], anti-coccidial [39], anti-oomycetes [37,40], as well as antifungal (Ujszegi et al., in preparation) activity.
Recently, it was discovered that the operons encoding the various enzymes of the biosynthetic pathway of each NR-AMP are globally regulated by the gene called Hfq in EPB species, providing an option to create EPB strains each of which produces only one single NR AMP molecule. (The method of constructing such a strain in the lab is called by using the „easyPACId” method) [41].
Whether the different NR-AMPs could ever be utilized as drugs not only against prokaryotic (bacteria) pathogens but eukaryotic (fungal pathogens, and parasitic protists) depends on the side effects. To get experimental experience about the option of applying EPB-produced antimicrobials to pathogens, and parasites of veterinary significance, we present here the results of an in vitro, and an accompanying in vivo study on chicken. In the in vitro study, we tested the cytotoxic potential of the cell-free conditioned culture media (CFCM) of three entomopathogenic bacterium species, - X. budapestensis, DSM16342 (EMA); X. szentirmaii DSM16338 (EMC); Photorhabdus luminescens ssp. akhurstii TT01 - on chicken tissue culture cells, namely, on the Leghorn Male Hepatoma (LMH), [92] cells, (a permanent confluent hepato-carcinoma cell line). Each CFCM proved rather cytotoxic in this test. In the in vivo study, we fed freshly hatched male broiler chickens for 42 days with XENOFOOD [39] which contained autoclaved cultures of EMA, and EMC. These bacteria were grown on standard chicken (starter and grower) [HM3] [u4] feed, and the whole culture was used as a “feed supplement”. [HM5]. It had been known that these EPB species cannot grow at body temperature (above 33 °C).
2. Results.
2.1. ANticlostridial Activity Of Ema And Emc In VitrO
Based on the previous results of a repeated experiments, we choose X. budapestensis DSM16342 (EMA) and X. szentirmaii DSM 16338, (EMC) which had been identified in our laboratory, [34]. For the exact description and history of these strains, see Ref. [35]. We retested the in vitro efficiency [36,37,38] in overlay bioassays and the pepsin resistance of the active ingredient that we previously published [39], (Figure 1 1 A and B).
Both native and autoclaved CFCM samples of EMA and EMC bacterial strains were bioassayed both in liquid, and agar diffusion tests as described previously [36-39. The anticoccidial activities of native and pepsin-treated CFCM samples were compared similarly as described previously [39]. All previous data on antimicrobial activity against each of the tested substances on different clinical isolates of Gram-positive Staphylococcus aureus [35,36,37,38,39,43] was repeatedly confirmed, indicating the antimicrobial activity These data were especially important as confirming all previous findings published in [38]. The experiment also confirmed the thermostability and the pepsin durability of the CFCM of EMA, and EMC [40], since autoclaving of these CFCMs did not affect the antimicrobial activity against any of the tested organisms.
2.2. In Vitro Bioassays: Cytopathogenic Effects Of Cell-Free Condition Media (Cfcm) Of 3 Epb Species On Lmh Chicken Cell Monolayer
Two experiments were performed on this subject, (see Materials and Methods). In both experiments, permanent, confluent monolayer culture samples (developed in different Falcon flasks) were used as target organisms, and different dilutions of different CFCMs were tested.
(As described in the Materials and Methods, confluent monolayers of LMH cells were parallelly developed in 18 Falcon flasks (for the first experiment), and 36 flasks (for the second experiment). The culture media were Medium RPMI [HM1] [u2] [u3] 1640 supplemented with 10% FBS, penicillin G (200 IU/ml), and streptomycin (200 µg/ml). The incubation time was for 72 hours in both cases, in the controlled atmosphere of 5% CO2 at 37°C and around 85-90% humidity in 25 cm (2) flasks with filtered caps. After the layer formed, the liquid growth media was removed from all but 3 flasks (in the first experiment) and in 2x2 flasks (in the second experiment). Those few (3X1 and 1x2) flasks were referred to as “unchanged” control flasks [HM1] [u2]. As for other flasks, 3x1 (in the first experiment) and 1x2 (in the second experiment) were refilled e fresh culture media (referred to as M199+15% FKS control flasks) while the rest of the flasks were refilled 100 V/V% of different CFCMs (EMA, EMC, TT01 yellow and TT01 red, in the first experiment, and different dilutions of EMA and EMC CFCMs in the second experiments. As for the replicates, in the first experiment we worked with triplicates and in the second experiment with duplicates).
2.2.1. Experiment-1 aimed at comparing the cytopathogenic effects of 4 different undiluted CFCMs obtained from cultures of EMA, EMC, and TT01 yellow, and TT01 red colony color variants, respectively, (see Materials and Methods).
(As described in the Materials and Methods, the EPB cultures had been grown in the same medium and the target (LMH) cells, these cultures were made cell-free by centrifugation and following sterile filtration, and these filtrates were used as CFCMs. The incubation of the LMH cell layers with EPB CFCMs (and respective controls) was carried out exactly in the same conditions as used for developing the confluent cell layers. A prolonged incubation time was characterized by the appearance of holes in the cell monolayer, semi-quantified by using the scoring system [44] (Amin et el., 2011) presented in Supplementary Material, Table 1S).
It was found that each CFCM exerted destructive effects on the cell layer. The initial lesions on the LMH monolayer were followed by more serious ones consisting of clumps attached to the monolayer, (Score 3). The results 1 (data are given as empiric score values, [44]) of Experiment are presented in Table 1. Table 1 Results of Experiment-1. Scoring the degree of LMH monolayer destruction caused by CFCM of 3 different EPB strains representing 2 different species.
2.2.2. Experiment-2 aimed at comparing the cytopathogenic effects of serial dilutions of the CFCM filtrates of EMA, and EMC cultures, considering that both the both EMA and EMC CFCMs showed much stronger antimicrobial activity than the two TT01 CFCM. The stock solutions, (and serial dilutions of them) used in this experiment were: EMA 60%, and EMC 80%, respectively. Confluent monolayer from LMH cells was developed again in Medium RPMI 1640 supplemented with 10% FBS, penicillin G (200 IU/ml), and streptomycin (200 µg/ml) for 72 hours in a controlled atmosphere of 5% CO2 at 37°C and around 85-90% humidity in 25 cm2 Falcon flasks with filtered caps (see also in Materials, and Methods). The same score system [44] (Amin, 2012, see Table S1) was used for evaluating the results.
The results of Experiment 2 are presented in Figure 2 and Figure 3, and the data are given in Supplementary material (Table 2S & 3S).
The data may allow us to hypothesize that the in vivo application of EMA NR_AMP peptides did cause fatal or even detectable cytopathogenic in sensitive organs (like the liver) and tissues where they are present in less than 3.2 V/V % (1:25 dilution) EMA-CFCM or 2.4V/V% EMC.
2.3. Results Of The In Vivo Xenofood-Feeding Experiment
2.3.1 Gastrointestinal activity of XENOFOOD
To examine whether the gastrointestinal potential of the XENOFOOD was considerable we counted Clostridium CFUs in the lower ileum (see Figure 4) of XENOFOOD-fed and untreated animals
Using ANOVA statistical analysis, we compared the average of the lower ileal Clostridium CFUs (see Figure 2) of the XENOFOOD-fed animals (48.1) with those of the controls (149.9). We found that despite the 3X difference, only a slightly significant could be demonstrated, probably due to the large standard deviation in the control values (Table 4).
The data show that the average weights of the spleens were smaller, while the average measurements of the sizes of the bursas were larger in the XENOFOOD-fed group, and the differences were significant. The same can be stated about the Clostridium colony-forming units, (CFUs): there were fewer in the XENOFOOD-fed group (on average). The XENOFOOD exerted unambiguously detectable gastrointestinal antimicrobial activity. But the original infestation of the population from which the experimental animals were selected rate was very low. These results allow concluding that XENOFOOD feeding was harmless. We suppose that the bursa-dependent humoral immune system must have neutralized the cytotoxic peptide-like molecules (including the GI-active NR-APMPs) of the XENOFOOD in the blood, protecting the sensitive tissues and organs in such a way.
2.3.2. Growth rate, feed consumption and the post-mortal data
During the 42-day-long experiment, no animal was lost. We did not detect any significant differences either in the body weight, (Table 5), in the feed conversion rates between the control and XENOFOOD-fed groups, or the growth rates of the two groups (Table 6).
As for the organ weight, they followed the allometric rules, according to which the regular and systematic patterns of growth such as the mass or size of any organ or part of a body can be expressed about the total mass or size of the entire organism according to the allometric equation: Y = bxα, where Y = mass of the organ, x = mass of the organism, α = growth coefficient of the organ, and b = a constant (only bursa- and spleen data are given). This fact itself excludes any organ-development abnormality of the treated (XENOFOOD-fed) animals. But the larger bursa weight and the smaller spleen weights, as well as larger bursa/spleen ratios in the XENOFOOD-fed groups may indicate indirectly an intensified humoral immune activity, leading to the neutralization of the cytotoxic AMP-components in the blood (Table 6).
3. Discussion.
AMPs comprise a potentially promising toolkit combatting MDR because antibiotic-resistant bacteria perform a high frequency of collateral sensitivity to antimicrobial peptides, [13,16,17]. X. budapestensis (EMA) and X. szentirmaii (EMC) are abundant sources of NR-AMP molecules of large target spectra and strong anti-pathogen potential, [35]. Each NR-AMP molecule in the Xenorhabdus CFCM is the end-product of a biosynthetic pathway. Each biosynthetic pathway is encoded by genes clustered in the respective operon (biosynthetic gene complex, BCG). In EMC 71 BCGs have been identified, [70]. Their large spectral NR-AMP products of overlapping activities and cooperating potential are coexisting as ingredients of the CFCM. Theoretically, there are two alternative ways of research philosophy for the geneticist of agricultural or veterinary commitments. One option is to focus on searching for individual molecules as new potential drugs, benefitting the global (co)regulation of the biosynthesis of different NR_AMPs, searching for or constructing EPB strains synthesizing only single NR-AMP molecules. As for this alternative, the recent discovery, according to which the different NRPS-encoding operons are under the global control of the post-transcriptional regulator, Hfq, provides an option for constructing Xenorhabdus strains producing single NR-AMP only, [41]. It is a great challenge! Another strategy is to try to benefit from the co-existence of naturally cooperating NR_AMP molecules in the CFCMs by producing harmless but effective preventive biopreparations like food supplements either as food supplements for veterinary, or soil additives for plant protection goals against pathogens and parasites. This latter option may be discussed here in light of the experiment's results where chickens were fed by XENOFOOD, a food supplement, in which in vitro active ut heat-sterlized NR-AMPs were present as ingredients. We know about these compounds that they were of peptides, [28], in vitro anti-coccidial active, [39] which did not lose their antimicrobial potential either when heat-sterilized [36].or when subjected to pepsin digestion, [39]. But in the in vitro experiment we presented here we found that each EPB CCM we tested exerted strong cytopathogenic effects on LMH permanent confluent chicken cells. We do not know whether one single (most probably the fabclavine) [48,49,50,51] molecule, or more than one molecule can be taken as responsible for the cyto-pathogenecity. There are several candidates (see Table 7).
It is not known whether the mechanism of the antimicrobial action of a given NR-AMP molecule is the same as those causing cytopathogenicity in the organisms. But the XENOFOOD food supplement proved in this experiment harmless and GI active against a bacterium pathogen, Clostridium perfringens. But in our in vivo feeding experiment the lower number of Clostridia does not automatically indicate a “protection” exerted by the XENOFOOD, since the natural infestation rate of the experimental animals was very low, but we do not see any other plausible explanation. The moderate but detectable anti-Clostridial effect without any detectable harmful side effects may be explained in more ways than one. An explanation may come from the previous experiments on Eimeria (unpublished) that the effective cytotoxic concentration is much higher than the effective anti-clostridial dose. Another possible explanation is that the actual local NP-AMR concentration (bioavailability) was much lower in the blood and the other organs and tissues than in the gut. We suppose that the second option is more probable because the immune system of the birds is probably capable of neutralizing these peptide-like compounds. The higher average weights of the bursas in the T-groups suggest that humoral immunity is mainly responsible for that.
As for the perspetives, see Conclusons.
4. Material, and Method
- Anti-clostridial potential of EPB CFCM: The respective methods have been published in [39]. Briefly, Clostridium perfringens NCAIM 1417 strain was obtained from the National Collection of Agricultural and Industrial Microorganisms – WIPO (of Hungary, Faculty of Food Sciences, Szent István University Somlói út 14-16 1118 Budapest, Hungary). Clostridium perfringens LH1-LH8; LH11-LH16; LH19, and LH20 are of chicken origin, and LH24 came from a pig; each has been deposited in the (frozen) stock collection of the Department of Microbiology and Infectious Diseases, University of Veterinary Medicine Budapest, Hungary. Xenorhabdus strains, X. budapestensis DSM 16342(T) (Lengyel) (EMA), and X. szentirmaii DSM 16338 (T) (Lengyel)(EMC) [34,35]. of EMA and EMC CFCM bioassays were tested on different Gram-positive strains including Clostridium perfringens strains were carried out as described before, [35,36,37,38,39].
- .In vitro experiments: Cell-free conditioned culture media (CFCM), of antibiotic-producing bacteria, were tested, and LHM (tissue culture) cells were in two different experiments.
- Experiment 1: Testing cytotoxicity of different EPB CFCM on confluent LMH (leghorn male hepatoma, LMH) cell line [42]: Developing Confluent layers of LMH (leghorn male hepatoma, LMH) cell line [42]: (LMH; ATCC Number: CRL-2117™) were developed culture Falcon flasks in Medium RPMI 1640 (Invitrogen/GIBCO), supplemented with 10% heat-inactivated fetal bovine serum FBS (Invitrogen/GIBCO), penicillin G (200 IU/ml) and streptomycin (200 µg/ml), respectively.
- In detail, cells were inoculated into 25 cm (2) flasks with filtered caps (Sarstedt) containing an end volume of 7 ml culture and incubated in a controlled atmosphere of 5% CO2 at 37 °C and around 85-90% humidity. After 72 hours of incubation, a confluent monolayer of LMH cells was obtained per flask.
- Altogether 18 Falcon tubes, - each containing 9 ml Medium 199 with Earle’s Salts, L-glutamine, 25 mM HEPES and L-amino acids (Invitrogen/GIBCO) and supplemented with 15% heat-inactivated fetal bovine serum FBS (Invitrogen/GIBCO) and 0.22% rice starch were used, 2x3 controls and 4X3 experimental tubes.
- Preparation of CFCMs: Tubes with thw same media (except fo for streptomicine, were inoculated with 4 different bacterial strains, EMA, EMC, TT01 yellow or TT01 red, representing 2 Xenorhabdus, (X. budapestensis nov. DSM16342(T), Lengyel) (EMA), [34,35], and X. szentirmaii nov. DSM16338 (T) (Lengyel) (EMC, [34,35], and 1 Photorhabdus (P. luminescens ssp. akhurstii TT01, [71] species. The latter is obtained from the Boemare laboratory (Montpellier, France). TT01 yellow and TT01 red names were used for two colony-color variants segregating spontaneously in McConkey agar plates, (P. Ganas, unpublished). (Xenorabdus, and Photorhabdus are penicillin resistant species).
- These antibiotic-producing bacteria were freshly taken from frozen cultures and grown on the bacterial species grown on MacConkey agar plates before they were transferred to the liquid medium as described before, [35,36,37,38,39,40]. The bacterial species were grown on MacConkey agar plates before they were transferred to the liquid medium, and unexpectedly two different types of colonies for TT01 were observed on the agar plates: red-brown colored colonies which adsorbed the neutral red from the MacConkey agar and yellow-colored colonies which did not so. Both types of colonies were tested for the effect of cell-free filtrates on LMH monolayers. In this particular experiment, the antibiotic-producing bacteria were cultured at 28 oC. The bacteria were incubated for 65 hours at 30 °C in a shaker (225 rpm. Bacterial cultures were then centrifuged at 3,300xg for 5 min and then the supernatants from the cultures were filtered through 0.22 µm cellulose acetate filters (Millipore).
- Experimental design: From all but 3 of the Falcon flasks (with the 72-hrs old LMH-layers), the culture medium of the LMH monolayers was removed from the flasks and replaced by the 4 CFCMs. Each of the CFCMs was tested in triplicates. There were two sets of controls. There were also 3 flasks with fresh medium (fresh Medium 199 supplemented with 15% FBS) without CFCM, and another 3 with the original, ("unchanged", that is 72h-old) culture media. All these 4X3 experimental and 2X3 control flasks were incubated for another 72hrs. The cultures were incubated in a controlled atmosphere of 5% CO2 at 37 °C and around 85-90% humidity.
- In vitro Experiment 2: permanent chicken liver cells (LMH; ATCC Number: CRL-2117™) [42] were grown in Medium RPMI 1640 (Invitrogen/GIBCO) supplemented with 10% heat-inactivated fetal bovine serum FBS (Invitrogen/GIBCO), penicillin G (200 IU/ml) and streptomycin (200 µg/ml).
- Cells were inoculated into 25 cm2 flasks with filtered caps (Sarstedt) containing an end volume of 7 ml culture and incubated in a controlled atmosphere of 5% CO2 at 37 °C and around 85-90% humidity.
- After 72 hours of incubation, a confluent monolayer of LMH cells was obtained per flask.
- All but the so-called “Unchanged culture flasks” LMH monolayers, the culture medium of the LMH monolayers was removed from the flasks and replaced by something.
- The control flasks were refilled with fresh media (Medium RPMI 1640 (Invitrogen/GIBCO) supplemented with 10% heat-inactivated fetal bovine serum FBS (Invitrogen/GIBCO), penicillin G (200 IU/ml) and streptomycin (200 µg/ml).
- The experimental flasks were refilled with cell-free (centrifuged and filtered serially diluted stock solutions (EMA: stock solution: 60V/V; EMC stock solution 80V/V%). The dilutions were 1:2.5; 1:5; 1:10 1:.25; 1:.50; 1:75, and 1:100, respectively.
- The EPB cells had been also cultured in Medium RPMI 1640 supplemented with 10% FBS, penicillin G, without streptomycin, (Xenorhabdus are pecicillin resistant).
- Each of the different filtrate analyses and the controls was performed in duplicate.
- The cultures were incubated in a controlled atmosphere of 5% CO2 at 37 °C and around 85-90% humidity.
- Each monolayer was investigated visually by an inverted light microscope to detect the effect of the cell-free filtrates on LMH monolayers. According to the degree of monolayer destruction, the following scoring system was established and also published, [44], (See Supplementary Material, Table S1).
- XENOFOOD preparation: XENOFOOD contained 5% soy-meal, which had been suspended with an equal amount (w/w) of EMA and another 5% suspended in an equal amount (w/w) of EMC cells obtained from 5 days-old shaken (2000 rpm) liquid cultures by high-speed (Sorwall; for 30 minutes) centrifugation.
- The liquid cultures were in 2XLB; (DIFCO); supplemented with meat extract equivalent amount to the yeast extract. The 5 days had been proven optimal for antimicrobial substance production at 25 oC in these conditions).
- It had previously been discovered that both EMA and EMC grow and produce antimicrobial substances in autoclaved soy meal containing some water and yeast extract or autoclaved yeast, (in 0.5 w/w %).
- Therefore, the original chicken food served as a semi-solid culture media of Xenorhabdus cells. Both EMA and EMC culturing semi-solid chicken food, which was prepared daily, and have been incubated in sterile conditions for another five days; then the EMA and EMC culturing media were united; autoclaved (20 min, 121 oC), and then dried by heat overnight. The Xenorhabdus cells were killed in such a way, while the heat stabile [36] antimicrobial compounds remained active
- XENOFOOD in vivo feeding experiment:Experimental animals: One-day-old male broiler chickens (N=2x34 = 68) were equally distributed into two groups: Control (C) and Treated (T) groups. The latter was fed with XENOFOOD. The C group was kept on a normal starter (1-10-d) and grower (11-42d) diet according to the standard international protocol.
- Food, feeding, evaluations: The T Group T was kept on “starter (1-10-d) XENOFOOD”, and “grower XENOFOOD” (11-42d). Body weights were measured daily between 1-42 days. Growth and FCR were monitored for 24-d. In the in vivo feeding experiment, we fed 39 birds with XENOFOOD, and there were 39 control birds.
- Body weights were measured daily between 1-42 days. Growth, - and food-conversion rates (GR and FCR respectively, were monitored for every 24-d.
- Dissection, Post mortam data: Not all but a sample (N=2x10=20) of 42-day-old birds were dissected on the 42nd day are presented here. We dissected a sample of 10 birds from the XENOFOOD-fed and a sample of 10 birds from the Control groups. [HM1] [u2] That is, not all but samples (N=2x10) of the 2x34) from 42-day-old birds were dissected to get post-mortam data about a few body organs as well as about the number of Clostridium germs in their ilea. The body weights of these selected animals did not differ from the average of their respective (C, or T) experimental groups. After dissection, the weights of the different organs were measured. The absolute and relative weights of the spleens and the bursae of Fabricii are presented in the Results section.
- CFU determination:The content of the lower ileum was washed taken out, diluted, and equilibrated and the colony-forming units were determined on BAM Media M75: Lactose-Gelatin Medium.
- Statistical AnalysisANOVA procedure was carried out by using the respective propositions of the SAS 9.4, see Acknowledgment section Software mostly due to an unbalanced data set. The significant differences (α = 0.05) between treatment means were assessed using the Least Significant Difference (LSD).
5. Conclusions
The work presented here can be considered as a contribution to the field of application of biosynthetic non-ribosomal encoded antimicrobial peptides (NR-AMPs) controlling multi-drug resistance (MDR). Regarding the future potential application of natural compounds, several aspects have to be taken into consideration: (A) the antimicrobial potential; (B) the durability, thermotolerance, and bioavailability; (C) cytotoxicity, and another, unwanted side effects, especially if one group of target organisms are of eukaryotic (protist) pathogens. We have not found any publication concerning the comparisons of the in vitro and the in vivo effects of antimicrobial active NR-AMP molecules of EPB origin on the same eukaryote (host) organism to be protected from pathogens or parasites.
Herein we demonstrated that both the heat-inactivated and the native cell-free conditioned culture media (CFCM) of Xenorhabdus budapestensis DSM16342 (EMA), and that of Xenorhabdus szentirmaii DSM16338 (EMC) which has broad-spectrum antimicrobial activity against several prokaryotic and eukaryotic microbial pathogens, also exerted a robust cytopathogenic effect on confluent (cultured) chicken cell layer, LHM).
The results of the in vivo experiment presented in Table 4, Table 5 and Table 6 may allow concluding that XENOFOOD feeding was harmless. It means that the NR-AMPs, present in the CFCM of X. budapestensis and X. szentirmaii - at least when applied as autoclaved food ingredients - did not cause any detectable side effects in vivo in broilers.
Their metabolic incorporation of undecomposed NR-AMPs into proteins or other tissue components is biochemical nonsense. However, their deposition into the egg yolk cannot theoretically be ruled out, therefore, we may recommend XENOFOOD as protecting food supplement only for cockerels but not for pullets.
The XENOFOOD exerted gastrointestinal antimicrobial activity in the GI has been reflected in the reduced average value of the Clostridium CFUs in the XENOFOOD-fed group. We suppose that the bursa-dependent humoral immune system neutralized the cytotoxic side components (of peptide nature) of the XENOFOOD, (that is, cytotoxic NR-AMPs of CFCMs in the blood (but, naturally, not in the gut).
The presence of the active antimicrobial compounds in the GI are proved by the significantly reduced number of colony-forming germs in the lower jejunum of the XENOFOOD-fed (T-group of) animals compared to that of the controls, (C-group). Based on those facts, we may recommend XENOFOOD as a useful food supplement to protect broiler cockerels against coccidiosis.
However, before drawing that conclusion, we recommend another experiment on experimentally Clostridium-pre-infested chickens, (also on cockerels, not pullet) to reproducibly determine the in vivo anti-Clostridial effects of the XENOFOOD diet quantitatively. (As mentioned above, we do not recommend the use of pullets, because the deposition of the XENOFOOD ingredients is unpredictable). We have been looking for a cooperating partner for carrying on this final experiment, and this is one of our strongest arguments for publishing these data in Pathogens
6. Patents
This work has not resulted in any patented result, we offer XENOFOOD recipe for the Scientific Community for free. Conflict of Interest Statement: The authors declare that the research was conducted without any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material: Scores of HML Monolayer Destruction caused by by EPB CFCMs.
Author Contributions
Conceptualization, ANDRÁS FODOR, László Makrai, Károly Dublecz and Michael Hess; Data curation, Sándor Józsa; Formal analysis, Claudia Hess, László Pál and Michael Klein; Funding acquisition, Tibor Vellai; Investigation, Claudia Hess, Petra Ganes and Andor Molnár; Methodology, Petra Ganes, László Makrai, László Pál, Andor Molnár, Sándor Józsa and Csaba Pintér; Project administration, László Makrai; Resources, Tibor Vellai and Károly Dublecz; Validation, Claudia Hess, László Fodor and Sándor Józsa; Writing – original draft, ANDRÁS FODOR; Writing – review & editing, Tibor Vellai, Claudia Hess, Michael Klein, Eustachio Tarascoi and Michael Hess. All authors will be informed about each step of manuscript processing including submission, revision, revision reminder, etc. via emails from our system or assigned Assistant Editor. The final order of the authors is as follows: ANDRÁS FODOR*; Tibor Vellai; Claudia Hess; Petra Ganes; László MAKRAI*; Károly DUBLECZ*; László Pál; Andor Molnár; László Fodor; Sándor Józsa; Csaba Pintér; Michael Gardner KLEIN*; Eustachio Tarasco; Michael Hess. Author Credit Statement. The authors confirm their proportional but e important, equally essential contribution to the paper. The authors demonstrate that they were sharing work contributed to the report as follows: Conceptualization: AF, LM, MH, KD; Data curation: SJ; Formal analysis: MGK, LF; Funding acquisition: collective, TV, LM Methodology: LM, (Clostridium in vitro, colony counting), CH, PG, (in vitro bioassays in LMH), LP (feeding experiments, food conversion), LM (post mortem examinations); AF (CFCM preparations); CSP (photo), SJ (statistics). The first draft was written by AF, corrected, and improved by TV, MGK, CH, and MH; Supplementary Material: PG
Funding
This work was carried out in collaboration with all authors. Resources: BSM, TÄMOP (see Funding). Fulbright, CEPO. Acknowledgements: In Budapest, at the Department of Microbiology and Infectious Diseases, University of Veterinary Medicine, Budapest, Hungary, (where coauthors Prof. László Makrai and László Fodor's work and in vitro Clostridium experiments were carried out), we would like to express thanks and appreciation for their professional technical help to the lab technicians for taking care of the bacterium strains for long years, and their invaluable technical help; especially to Miss Teréz Halasi, Éva Kolozsvárim Miss Alexandra Jutasi. In Vienna, at the Clinic for Poultry and Fish Medicine, Department for Farm Animals and Veterinary Public Health, University of Veterinary Medicine (Vetmeduni), Vienna), Austria; we would like to express thanks and appreciation not only for their professional technical but the intellectual help to the whole research team of the Hess laboratory, especially to Aziza Amin, and Irina Prokofieva for running the in vitro cytopathogenicity experiments, on the LMH cells. Also in Austria, we have to thank Dr. Klaus Teichmann (Biomin, Tulln, Austria), for his testing the same CFCM samples on Eimeria cells, repeating our cytotoxicity tests on chicken cells confirming our findings, and sharing his conclusions with us unselfishly, declaring that the effective anti-Eimeria was an order of magnitude lower than the cytotoxic dose. In Keszthely, at the Institute of Physiology and Nutrition, Georgikon Campus of Hungarian University of Agriculture and Life Sciences,(MATE), Hungary, where the in vivo feeding experiments were going on, we would like to express thanks and appreciation not only for their professional technical but the intellectual help to the whole research team of the Dublecz team, especially for the technical help of Mrs. Judit Varga, and Mrs. Cseh, for her professional and administrative help Mrs. Angéla Vörös – Mezei. Finally, as the first author, A. Fodor would like to thank each of the coauthors as well his wife Mrs. Andrea Máthé-Fodor, for their professional, and intellectual, but for emotional- psychological support. Funding. This research received no external funding” except for some parts parts of this study (The in vitro Clostridium bioassays preceding XENOFOOD experiments) were supported by the TÁMOP-4.2.2/B-10/1-2010-0025 project based on the financial support the State of Hungary and the EU. The chicken feeding experiments included two (control (34, chicken)) and a XENOFOOD-fed (34 chicken) groups of another feeding experiment of the respective Department of the University of used-to-be Pannonia, Keszthely, Hungary, (recently a joint unit of the Hungarian University of Agriculture and Life Sciences (MATE).
Institutional Review Board Statement
The in vivo feeding experiments were actually carried out according to the protocol that was asked for at the University of Veterinary Medicine, Budapest, Hungary; (Attached). We are aware that the Editorial Office might ask you for further information and are ready to provide it. We are aware that the Editorial Office might ask you for further information and are ready to provide it. A copy of the official permission is available.
References
- Huang W, Reyes-Caldas P, Mann M, Seifbarghi S, Kahn A, Almeida RPP, Béven L, Heck M, Hogenhout SA, Coaker G. Bacterial Vector-Borne Plant Diseases: Unanswered Questions and Future Directions. Mol Plant. 2020, 13, 1379–1393. [CrossRef] [PubMed]
- Białas A, Zess EK, De la Concepcion JC, Franceschetti M, Pennington HG, Yoshida K, Upson JL, Chanclud E, Wu CH, Langner T, Maqbool A, Varden FA, Derevnina L, Belhaj K, Fujisaki K, Saitoh H, Terauchi R, Banfield MJ, Kamoun S. Lessons in Effector and NLR Biology of Plant-Microbe Systems. Mol Plant Microbe 3.Interact. 2018, 31, 34–45. [CrossRef] [PubMed]
- Castro Monzon F, Rödel MO, Jeschke JM. Tracking Batrachochytrium dendrobatidis Infection Across the Globe. Ecohealth. 2020, 17, 270–279. [CrossRef] [PubMed]
- Kolářová I, Valigurová A. Hide-and-Seek: A Game Played between Parasitic Protists and Their Hosts. Microorganisms. 2021, 9, 2434. [CrossRef] [PubMed]
- Itoïz S, Metz S, Derelle E, Reñé A, Garcés E, Bass D, Soudant P, Chambouvet A. Emerging Parasitic Protists: The Case of Perkinsea. Front Microbiol. 2022, 12, 735815. [CrossRef] [PubMed] [PubMed Central]
- Preston S, Jabbar A, Gasser RB. A perspective on genomic-guided anthelmintic discovery and repurposing using Haemonchus contortus. Infect Genet Evol. 2016, 40, 368–373. [CrossRef] [PubMed]
- Zuo Y, Shi Y, Zhang F, Guan F, Zhang J, Feyereisen R, Fabrick JA, Yang Y, Wu Y. Genome mapping coupled with CRISPR gene editing reveals a P450 gene confers avermectin resistance in the beet armyworm. PLoS Genet. 2021, 17, e1009680. [CrossRef] [PubMed] [PubMed Central]
- Sokól R, Galecki R. The resistance of Eimeria spp. to toltrazuril in black grouse (Lyrurus tetrix) kept in an aviary. Poult Sci. 2018, 97, 4193–4199. [CrossRef] [PubMed]
- Akhter MJ, Aziz FB, Hasan MM, Islam R, Parvez MMM, Sarkar S, Meher MM. Comparative effect of papaya (Carica papaya) leaves' extract and Toltrazuril on growth performance, hematological parameter, and protozoal load in Sonali chickens infected by mixed Eimeria spp. J Adv Vet Anim Res. 2021, 8, 91–100. [CrossRef] [PubMed] [PubMed Central]
- Hawkins NJ, Fraaije BA. Contrasting levels of genetic predictability in the evolution of resistance to major classes of fungicides. Mol Ecol. 2021, 30, 5318–5327. [CrossRef] [PubMed]
- Knutie SA, Wilkinson CL, Kohl KD, Rohr JR. Early-life disruption of amphibian microbiota decreases later-life resistance to parasites. Nat Commun. 2017, 8, 86. [CrossRef] [PubMed] [PubMed Central]
- Uddin, T.M.; Chakraborty, A.J.; Khusro, A.; Zidan, B.R.M.; Mitra, S.; Emran, T.B.; Dhama, K.; Ripon, M.K.H.; Gajdács, M.; Sahibzada, M.U.K.; et al. Antibiotic resistance in microbes: History, mechanisms, therapeutic strategies and future prospects. J. Infect. Public Health 2021, 14, 1750–1766. [Google Scholar] [CrossRef] [PubMed]
- Fodor A, Abate BA, Deák P, Fodor L, Gyenge E, Klein MG, Koncz Z, Muvevi J, Ötvös L, Székely G, Vozik D, Makrai L. Multidrug Resistance (MDR) and Collateral Sensitivity in Bacteria, with Special Attention to Genetic and Evolutionary Aspects and to the Perspectives of Antimicrobial Peptides-A Review. Pathogens. 2020, 9, 522. [CrossRef] [PubMed] [PubMed Central]
- Ötvös L Jr, Wade JD. Current challenges in peptide-based drug discovery . Front Chem. 2014, 2, 62. [CrossRef] [PubMed] [PubMed Central]
- Carton J. M., Strohl W. R. (2013). Protein therapeutics (Introduction to biopharmaceuticals), in Biological and Small Molecule Drug Research and Development, eds Ganelin R., Jefferts R., Roberts S. (Waltham, MA: Academic Press; ), 127–159.
- Lázár V, Martins A, Spohn R, Daruka L, Grézal G, Fekete G, Számel M, Jangir PK, Kintses B, Csörgő B, Nyerges Á, Györkei Á, Kincses A, Dér A, Walter FR, Deli MA, Urbán E, Hegedűs Z, Olajos G, Méhi O, Bálint B, Nagy I, Martinek TA, Papp B, Pál C. Antibiotic-resistant bacteria show widespread collateral sensitivity to antimicrobial peptides. Nat Microbiol. 2018, 3, 718–731. [CrossRef] [PubMed]
- Kintses B, Jangir PK, Fekete G, Számel M, Méhi O, Spohn R, Daruka L, Martins A, Hosseinnia A, Gagarinova A, Kim S, Phanse S, Csörgő B, Györkei Á, Ari E, Lázár V, Nagy I, Babu M, Pál C, Papp B. Chemical-genetic profiling reveals limited cross-resistance between antimicrobial peptides with different modes of action. Nat Commun. 2019, 10, 5731. [CrossRef] [PubMed] [PubMed Central]
- Mojsoska B, Jenssen H Peptides and Peptidomimetics for Antimicrobial Drug Design . Pharmaceuticals (Basel). 2015, 8, 366–415. [CrossRef] [PubMed] [PubMed Central]
- Jenssen H, Hamill P, Hancock RE. Peptide antimicrobial agents. Clin Microbiol Rev. 2006, 19, 491–511. [CrossRef] [PubMed] [PubMed Central]
- Ostorházi E, Hoffmann R, Herth N, Wade JD, Kraus CN, Otvos L Jr. Advantage of a Narrow Spectrum Host Defense (Antimicrobial) Peptide Over a Broad Spectrum Analog in Preclinical Drug Development. Front Chem. 2018, 6, 359. [CrossRef] [PubMed] [PubMed Central]
- Ostorházi E, Holub MC, Rozgonyi F, Harmos F, Cassone M, Wade JD, Otvos L Jr. Broad-spectrum antimicrobial efficacy of peptide A3-APO in mouse models of multidrug-resistant wound and lung infections cannot be explained by in vitro activity against the pathogens involved. Int J Antimicrob Agents. 2011, 37, 480–4. [CrossRef] [PubMed]
- Ötvös L., Jr. Immunomodulatory effects of anti-microbial peptides. Acta Microbiol Immunol Hung. 2016, 63, 257–277. [Google Scholar] [CrossRef] [PubMed]
- Petkovic M, Mouritzen MV, Mojsoska B, Jenssen H. Immunomodulatory Properties of Host Defence Peptides in Skin Wound Healing. Biomolecules. 2021, 11, 952. [CrossRef] [PubMed] [PubMed Central]
- Jenssen, H, Fjell, C. D.; Cherkasov, A.; Hancock, R.E. QSAR modeling and computer-aided design of antimicrobial peptides. J. Pept. Sci. 2008, 14, 110–114.
- Mojsoska B, Zuckermann RN, Jenssen H. Structure-activity relationship study of novel peptoids that mimic the structure of antimicrobial peptides. Antimicrob Agents Chemother. 2015, 59, 4112–20. [CrossRef] [PubMed]
- Tsakou F, Jersie-Christensen R, Jenssen H, Mojsoska B. The Role of Proteomics in Bacterial Response to Antibiotics. Pharmaceuticals (Basel). 2020, 13, 214. [CrossRef] [PubMed] [PubMed Central]
- Ötvös, L., Jr. The short proline-rich antibacterial peptide family. Cell Mol. Life Sci. 2002, 59, 1138–1150. [Google Scholar] [CrossRef] [PubMed]
- Chang KP, Reed SG, McGwire BS, Soong L. Leishmania model for microbial virulence: the relevance of parasite multiplication and pathoantigenicity. Acta Trop. 2003, 85, 375–90. [CrossRef] [PubMed]
- McGwire BS, Kulkarni MM Interactions of antimicrobial peptides with Leishmania and trypanosomes and their functional role in host parasitism . Exp Parasitol. 2010, 126, 397–405. [CrossRef] [PubMed]
- Kulkarni MM, McMaster WR, Kamysz W, McGwire BS. Antimicrobial peptide-induced apoptotic death of leishmania results from calcium-de pendent, caspase-independent mitochondrial toxicity. J Biol Chem. 2009, 284, 15496–504. [CrossRef] [PubMed]
- Akhurst, R.J. Antibiotic activity of Xenorhabdus spp. , bac 190 teria symbiotically associated with insect pathogenic nematodes of the families Heterorhabditidae and Steinernematidae. J. Gen.Microbiol. 1982, 128, 3061. [Google Scholar]
- Ogier, J.C. , Pagès, S. ; Frayssinet, M.; Gaudriault, S. Entomopathogenic nematode-associated microbiota: From monoxenic paradigm to pathobiome. Microbiome 2020, 8, 25. [Google Scholar] [CrossRef] [PubMed]
- McGwire BS, Olson CL, Tack BF, Engman DM. Killing of African trypanosomes byantimicrobial peptides. J Infect Dis. 2003, 188, 146–52. [CrossRef] [PubMed]
- Lengyel, K. , Lang, E. , Fodor, A.; Szállás, E, Schumann, P, Stackebrandt, E. Description of four novel species of Xenorhabdus, family Enterobacteriaceae: Xenorhabdus budapestensis sp. nov., Xenorhabdus ehlersii sp. nov., Xenorhabdus innexi sp. nov., and Xenorhabdus szentirmaii sp. nov. Syst. Appl. Microbiol. 2005, 28, 115–122. [Google Scholar] [PubMed]
- Fodor A, Gualtieri M, Zeller M, Tarasco E, Klein MG, Fodor AM, Haynes L, Lengyel K, Forst SA, Furgani GM, Karaffa L, Vellai T. Type Strains of Entomopathogenic Nematode-Symbiotic Bacterium Species, Xenorhabdus szentirmaii (EMC) and X. budapestensis (EMA), Are Exceptional Sources of Non-Ribosomal Templated, Large-Target-Spectral, Thermotolerant-Antimicrobial Peptides (by Both), and Iodinin (by EMC). Pathogens. 2022, 11, 342. [CrossRef] [PubMed] [PubMed Central]
- Furgani, G, Böszörményi, E. , Fodor, A, Máthé-Fodor, A. Forst, S, Hogan, J.S., Katona, Z, Klein, M.G.; Stackebrandt, E.; Szentirmai, A. et al. Xenorhabdus antibiotics: A comparative analysis and potential utility for controlling mastitis caused by bacteria. J. Appl. Microbiol. 2008, 104, 745–758. [CrossRef] [PubMed]
- Böszörményi E, Érsek T, Fodor A, Fodor AM, Földes LS, Hevesi M, Hogan JS, Katona Z, Klein MG, Kormány A, Pekár S, Szentirmai A, Sztaricskai F, Taylor RA. Isolation and activity of Xenorhabdus antimicrobial compounds against the plant pathogens Erwinia amylovora and Phytophthora nicotianae. J Appl Microbiol. 2009, 107, 746–59. [CrossRef] [PubMed]
- Vozik, D, Bélafi-Bakó, K, Hevesi, M, Böszöményi, E. , Fodor, A. Effectiveness of a peptide-rich fraction from Xenorhabdus budapestensis culture against fire blight disease on apple blossoms. Not. Bot. Horti Agrobo 2015, 43, 547–553. [CrossRef]
- Fodor, A, Makrai L, Fodor L, Venekei, I, Pál, L, Husvéth, F, Molnár, A, Dublecz, K, Pintér, Cs, Józsa, S, Klein, MG. Anti-Coccidiosis Potential of Autoclaveable Antimicrobial Peptides from Xenorhabdus budapestensis Resistant to Proteolytic (Pepsin, Trypsin) Digestion Based on In vitro Studies. Microbiology Research Journal International, Page 1-17. [CrossRef]
- Fodor, A; Varga, I. ; Hevesi, M; Máthé-Fodor, A, Racsko, J, Eds.; Hogan, J.A. Novel anti-microbial peptides of Xenorhabdus origin against multidrug resistant plant pathogens. In: V. Bobbarala (Ed.) A Search for Antibacterial Agents. London, United Kingdom, IntechOpen, 2012 [Online]. pp. 3–32. Available from: https://www.intechopen.com/books/2129. [Google Scholar] [CrossRef]
- Bode, E, Heinrich, AK, Hirschmann, M, Abebew, D, Shi, YN Vo, T. D.; Wesche, F.; Shi, Y.M.; Grün, P.; Simonyi, S.; et al. Promoter activation in Δhfq mutants as an efficient tool for specialized metabolite production enabling direct bioactivity testing. Angew. Chem. Int. Ed. Engl. 2019, 58, 18957–18963. [CrossRef]
- Kawaguchi T, Nomura K, Hirayama Y, Kitagawa T. Establishment and characterization of a chicken hepatocellular carcinoma cell line, LMH. Cancer Res. 1987, 47, 4460–4. [PubMed]
- Böszörményi E. Barcs, I., Domján, Gy., Bakó, K., Fodor, A., Makrai, L., Vozik, D. [Xenorhabdus budapestensis entomopathogenic bacteria cell free conditioned medium and purified peptide fraction effect on some zoonotic bacteria]. Orvosi Hetilap, Volume/Issue: Volume 156: Issue 44. https://doi.org/. 2018. In Hungarian. Brachmann AO, Forst S, Furgani GM, Fodor A, Bode HB. Xenofuranones A and B: phenylpyruvate dimers from Xenorhabdus szentirmaii. J Nat Prod. 2006, 69, 1830–2. [CrossRef] [PubMed]
- Amin A, Bilic I, Berger E, Hess M. Trichomonas gallinae, in comparison to Tetratrichomonas gallinarum, induces distinctive cytopathogenic effects in tissue cultures. Vet Parasitol. 2012, 186, 196–206. [CrossRef] [PubMed]
- Brachmann AO, Forst S, Furgani GM, Fodor A, Bode HB. Xenofuranones A and B: phenylpyruvate dimers from Xenorhabdus szentirmaii. J Nat Prod. 2006, 69, 1830–2. [CrossRef] [PubMed]
- Gualtieri M, Banéres-Roquet F, Villain-Guillot P, Pugnière M, Leonetti JP. The antibiotics in the chemical space. Curr Med Chem. 2009, 3, 390–393. [CrossRef] [PubMed]
- Gualtieri, M, Villain-Guillot, P, Givaudan, A,Pages, S. Nemaucin, an Antibiotic Produced by Entomopathogenic Xenorhabdus cabanillasii. A Patent WO2012085177A1, 28 June 2012, France.
- Fuchs SW, Sachs CC, Kegler C, Nollmann FI, Karas M, Bode HB. Neutral loss fragmentation pattern-based screening for arginine-rich natural products in Xenorhabdus and Photorhabdus. Anal Chem. 2012, 84, 6948–55. [CrossRef] [PubMed]
- Fuchs SW, Grundmann F, Kurz M, Kaiser M, Bode HB. Fabclavines: bioactive peptide-polyketide-polyamine hybrids from Xenorhabdus. Chembiochem. 2014, 15, 512–6. [CrossRef] [PubMed]
- Wenski SL, Kolbert D, Grammbitter GLC, Bode HB. Fabclavine biosynthesis in X. szentirmaii: shortened derivatives and characterization of the thioester reductase FclG and the condensation domain-like protein FclL. J Ind Microbiol Biotechnol. 2019, 46, 565–572. [CrossRef] [PubMed]
- Wenski SL, Cimen H, Berghaus N, Fuchs SW, Hazir S, Bode HB. Fabclavine diversity in Xenorhabdus bacteria. Beilstein J Org Chem. 2020, 16, 956–965. [CrossRef] [PubMed] [PubMed Central]
- Houard J, Aumelas A, Noël T, Pages S, Givaudan A, Fitton-Ouhabi V, Villain-Guillot P, Gualtieri M. Cabanillasin, a new antifungal metabolite, produced by entomopathogenic Xenorhabdus cabanillasii JM26. J Antibiot (Tokyo). 2013, 66, 617–20. [CrossRef] [PubMed]
- Dreyer J, Rautenbach M, Booysen E, van Staden AD, Deane SM, Dicks LMT. Xenorhabdus khoisanae SB10 produces Lys-rich PAX lipopeptides and a Xenocoumacin in its antimicrobial complex. BMC Microbiol. 2019, 19, 132. [CrossRef] [PubMed] [PubMed Central]
- Gualtieri M, Aumelas A, Thaler JO. Identification of a new antimicrobial lysine-rich cyclolipopeptide family from Xenorhabdus nematophila. J Antibiot (Tokyo). 2009, 62, 295–302. [CrossRef] [PubMed]
- Fuchs SW, Proschak A, Jaskolla TW, Karas M, Bode HB. Structure elucidation and biosynthesis of lysine-rich cyclic peptides in Xenorhabdus nematophila. Org Biomol Chem. 2011, 9, 3130–2. [CrossRef] [PubMed]
- Isaacson PJ, Webster JM Antimicrobial activity of Xenorhabdus sp. RIO (Enterobacteriaceae), symbiont of the entomopathogenic nematode, Steinernema riobrave (Rhabditida: Steinernematidae) . J Invertebr Pathol. 2002, 79, 146–53. [CrossRef] [PubMed]
- Pantel L, Florin T, Dobosz-Bartoszek M, Racine E, Sarciaux M, Serri M, Houard J, Campagne JM, de Figureueiredo RM, Midrier C, Gaudriault S, Givaudan A, Lanois A, Forst S, Aumelas A, Cotteaux-Lautard C, Bolla JM, Vingsbo Lundberg C, Huseby DL, Hughes D, Villain-Guillot P, Mankin AS, Polikanov YS, Gualtieri M. Odilorhabdins, Antibacterial Agents that Cause Miscoding by Binding at a New Ribosomal Site. Mol Cell. 2018, 70, 83–94. [CrossRef] [PubMed]
- Sarciaux M, Pantel L, Midrier C, Serri M, Gerber C, Marcia de Figureueiredo R, Campagne JM, Villain-Guillot P, Gualtieri M, Racine E. Total Synthesis and Structure-Activity Relationships Study of Odilorhabdins, a New Class of Peptides Showing Potent Antibacterial Activity. J Med Chem. 2018, 61, 7814–7826. [CrossRef] [PubMed]
- Racine E, Gualtieri M. From Worms to Drug Candidate: The Story of Odilorhabdins, a New Class of Antimicrobial Agents. Front Microbiol. 2019, 10, 2893. [CrossRef] [PubMed] [PubMed Central]
- Lanois-Nouri A, Pantel L, Fu J, Houard J, Ogier JC, Polikanov YS, Racine E, Wang H, Gaudriault S, Givaudan A, Gualtieri M. The Odilorhabdin Antibiotic Biosynthetic Cluster and Acetyltransferase Self-Resistance Locus Are Niche and Species Specific. mBio. 2022, 13, e0282621. [CrossRef] [PubMed]
- Xiao, Y.; Meng, F.; Qiu, D.; Yang, X. Two novel antimicrobial peptides purified from the symbiotic bacteria Xenorhabdus budapestensis NMC-10. Peptides 2012, 35, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Reimer D, Nollmann FI, Schultz K, Kaiser M, Bode HB. Xenortide Biosynthesis by Entomopathogenic Xenorhabdus nematophila. J Nat Prod. 2014, 77, 1976–80. [CrossRef] [PubMed]
- Esmati N, Maddirala AR, Hussein N, Amawi H, Tiwari AK, Andreana PR. Efficient syntheses and anti-cancer activity of xenortides A-D including ent/epi-stereoisomers. Org Biomol Chem. 2018, 16, 5332–5342. [CrossRef] [PubMed]
- Reimer D, Cowles KN, Proschak A, Nollmann FI, Dowling AJ, Kaiser M, ffrench-Constant R, Goodrich-Blair H, Bode HB. Rhabdopeptides as insect-specific virulence factors from entomopathogenic bacteria. Chembiochem. 2013, 14, 1991–7. [CrossRef] [PubMed]
- Bi Y, Gao C, Yu Z. Rhabdopeptides from Xenorhabdus budapestensis SN84 and Their Nematicidal Activities against Meloidogyne incognita. J Agric Food Chem. 2018, 66, 3833–3839. [CrossRef] [PubMed]
- Xi X, Lu X, Zhang X, Bi Y, Li X, Yu Z. Two novel cyclic depsipeptides Xenematides F and G from the entomopathogenic bacterium Xenorhabdus budapestensis. J Antibiot (Tokyo). 2019, 72, 736–743. [CrossRef] [PubMed]
- Zhao, L.; Kaiser, M.; Bode, H.B. Rhabdopeptide/Xenortide-like peptides from Xenorhabdus innexi with terminal amines showing potent antiprotozoal activity. Org. Lett. 2018, 20, 5116–5120. [Google Scholar] [CrossRef] [PubMed]
- Ohlendorf B, Simon S, Wiese J, Imhoff JF. Szentiamide, an N-formylated cyclic depsipeptide from Xenorhabdus szentirmaii DSM 16338T. Nat Prod Commun. 2011, 6, 1247–50. [PubMed]
- Nollmann FI, Dowling A, Kaiser M, Deckmann K, Grösch S, ffrench-Constant R, Bode HB. Synthesis of szentiamide, a depsipeptide from entomopathogenic Xenorhabdus szentirmaii with activity against Plasmodium falciparum. Beilstein J Org Chem. 2012, 8, 528–33. [CrossRef] [PubMed]
- Gualtieri M, Ogier JC, Pagès S, Givaudan A, Gaudriault S. Draft Genome Sequence and Annotation of the Entomopathogenic Bacterium Xenorhabdus szentirmaii Strain DSM16338. Genome Announc. 2014, 2, e00190–14. [CrossRef] [PubMed] [PubMed Central]
- Duchaud E, Rusniok C, Frangeul L, Buchrieser C, Givaudan A, Taourit S, Bocs S, Boursaux-Eude C, Chandler M, Charles JF, Dassa E, Derose R, Derzelle S, Freyssinet G, Gaudriault S, Médigue C, Lanois A, Powell K, Siguier P, Vincent R, Wingate V, Zouine M, Glaser P, Boemare N, Danchin A, Kunst F. The genome sequence of the entomopathogenic bacterium Photorhabdus luminescens. Nat Biotechnol. 2003, 21, 1307–13. [CrossRef] [PubMed]
Figure 1.
Results of the in vitro experiment with EMA CFCM in anaerobic conditions. The active ingredient inhibited the growth of Clostridium perfringens cells in agar media before (a) and after (b) pepsin digestion. For details, see Materials, and Method. Photo: Cs. Pintér.
Figure 1.
Results of the in vitro experiment with EMA CFCM in anaerobic conditions. The active ingredient inhibited the growth of Clostridium perfringens cells in agar media before (a) and after (b) pepsin digestion. For details, see Materials, and Method. Photo: Cs. Pintér.
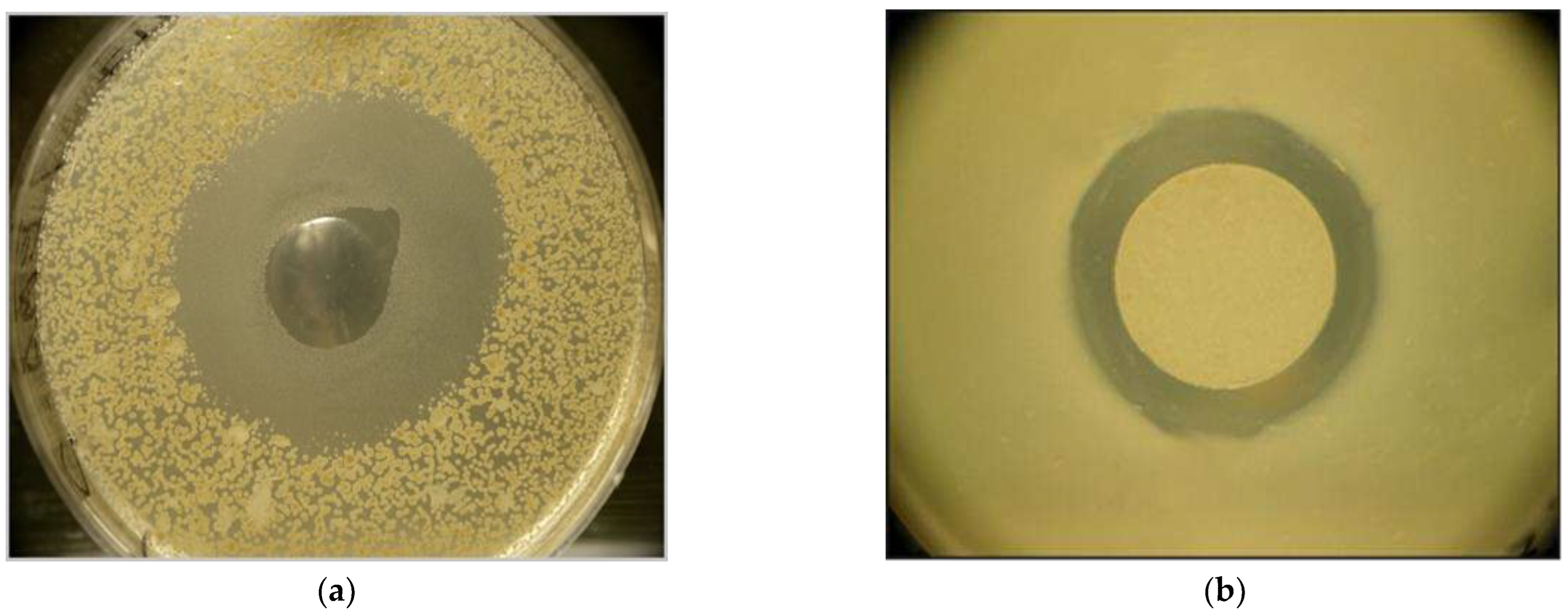
Figure 2.
Destruction of LMH [42] cell monolayer caused by serially diluted EMA CFCM samples. [HM1] The scoring system is [44] shown in Table S1. (Abbreviations: EMA= Xenorhabdus budapestensis DSM16342(T) (Lengyel). CFCM= cell-free conditioned culture media). Data are given in Supplementary Material TableS2 and TableS3.
Figure 2.
Destruction of LMH [42] cell monolayer caused by serially diluted EMA CFCM samples. [HM1] The scoring system is [44] shown in Table S1. (Abbreviations: EMA= Xenorhabdus budapestensis DSM16342(T) (Lengyel). CFCM= cell-free conditioned culture media). Data are given in Supplementary Material TableS2 and TableS3.
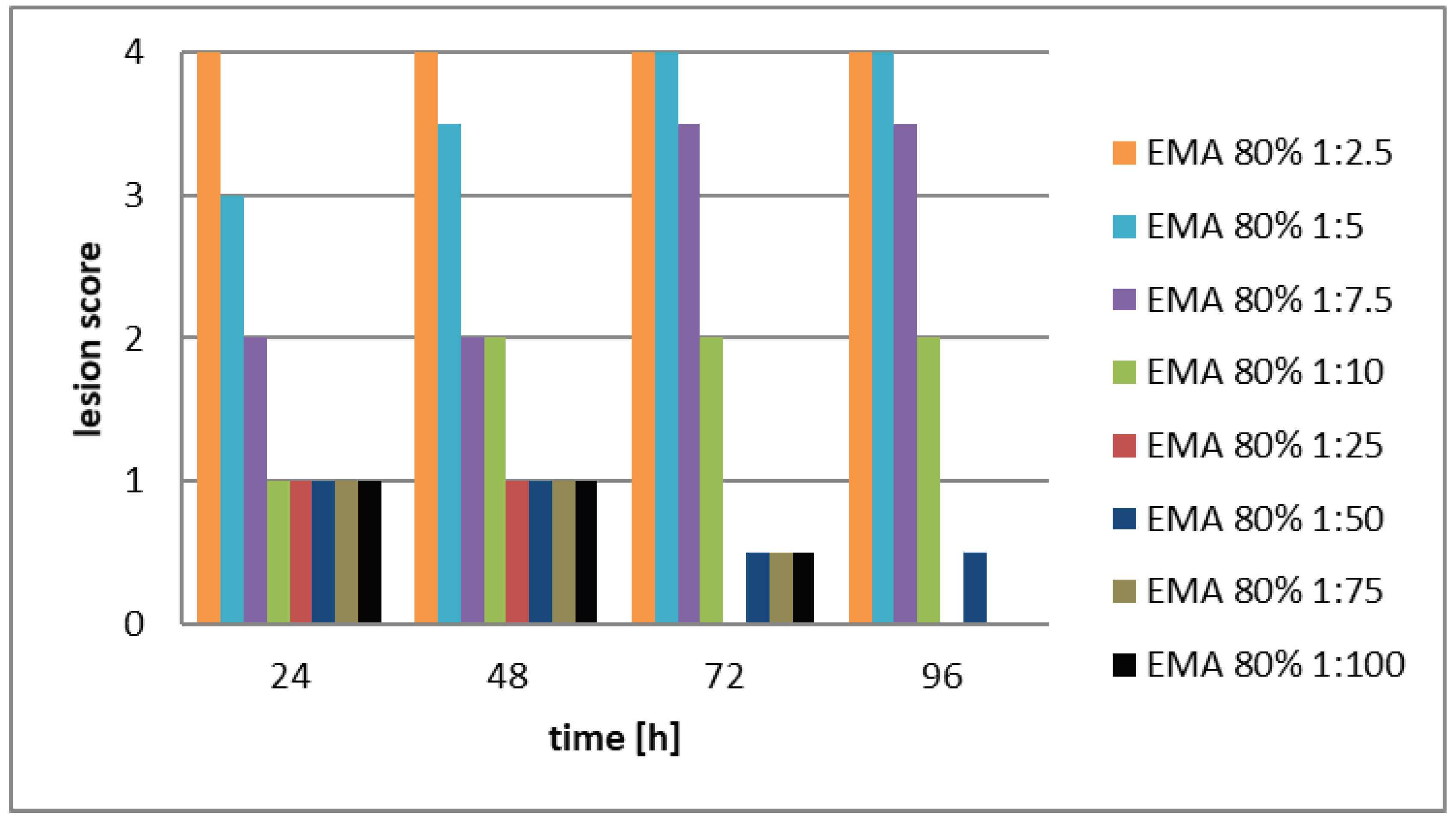
Figure 3.
Destruction of LMH [42] cell monolayer caused by serially diluted EMC CFCM samples. [HM1] The scoring system is [44] shown in Table S1. (Abbreviations: EMC= Xenorhabdus budapestensis DSM16338(T) (Lengyel). CFCM= cell-free conditioned culture media). Data are given in Supplementary Material TableS2 and TableS3.
Figure 3.
Destruction of LMH [42] cell monolayer caused by serially diluted EMC CFCM samples. [HM1] The scoring system is [44] shown in Table S1. (Abbreviations: EMC= Xenorhabdus budapestensis DSM16338(T) (Lengyel). CFCM= cell-free conditioned culture media). Data are given in Supplementary Material TableS2 and TableS3.
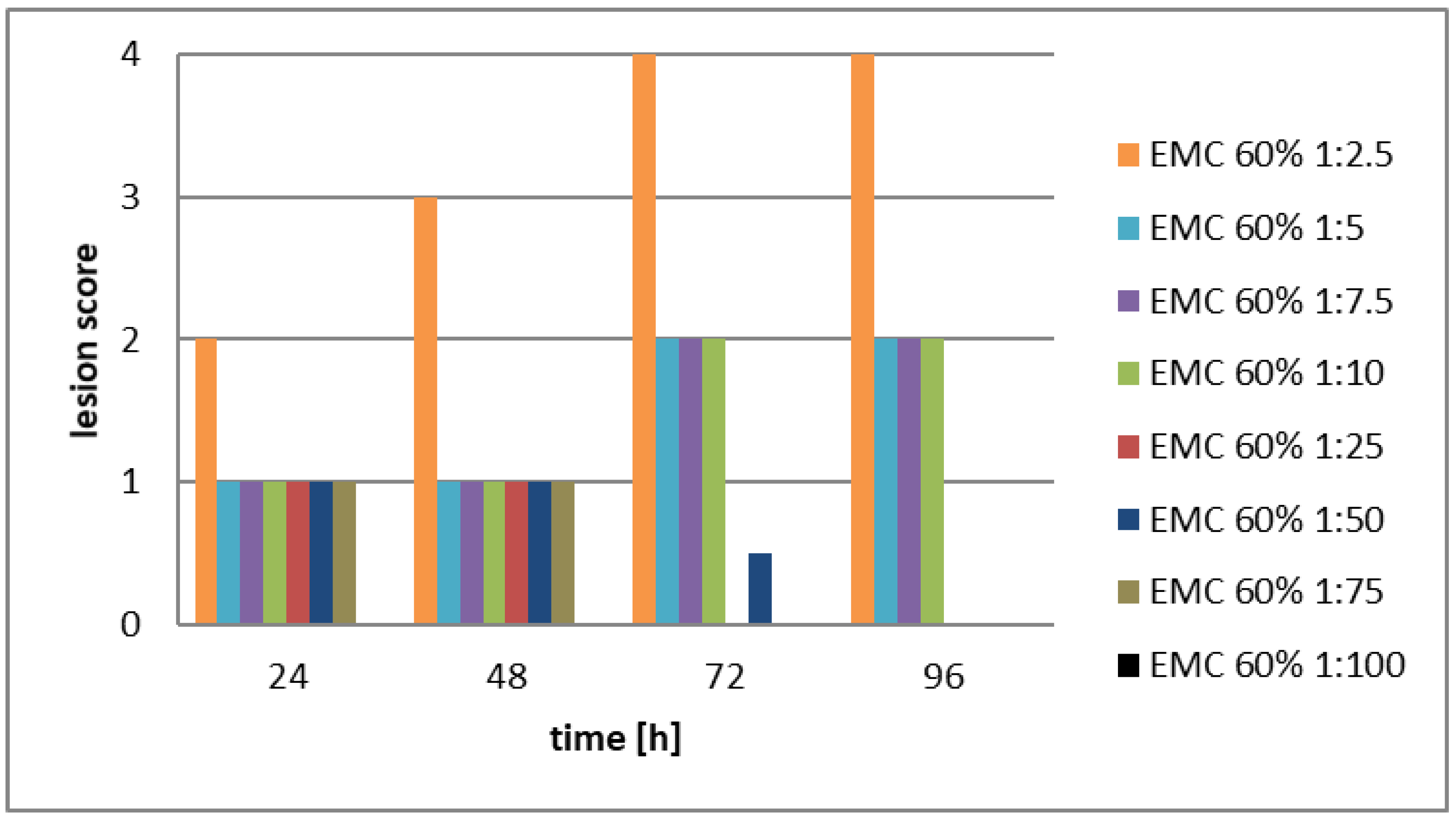
Figure 4.
Determination of viable cell count using plate counting method (Clostridium perfringens, blood agar). .
Figure 4.
Determination of viable cell count using plate counting method (Clostridium perfringens, blood agar). .

| Samples | Replicates | 24h | 48h |
|---|---|---|---|
|
M199 the original (unchanged) culture Media, in which LMH layer had been developed |
A | 0 | 0 |
| B | 0 | 0 | |
| C | 0 | 0 | |
| Fresh (199 + 15% + PKS) culture media added | A | 0 | 0 |
| A | |||
| B | 0 | 0 | |
| C | 0 | 0 | |
|
EMA CFCM (EMA had been cultured inM199) |
A | 3 | 4 |
| B | 3 | 4 | |
| C | 3 | 4 | |
|
EMC CFCM (EMC had been cultured inM199) |
A | 4 | 4 |
| B | 4 | 4 | |
| C | 4 | 4 | |
|
TT01 YELLOW CFCM (TT01 had been cultured inM199) |
A | 4 | 4 |
| B | 4 | 4 | |
| C | 4 | 4 | |
|
TT01 RED CFCM TT01 had been cultured inM199) |
A | 4 | 4 |
| B | 4 | 4 | |
| C | 4 | 4 |
1Table 1 Footnotes. The numbers in the table are the score values (see Table 1). Abbreviations: M199 Unchanged C growth control media (M199, unchanged); M199 supplemented with + 15% + PKS C growth control media (fresh M199, supplemented with 15% fibroblast growth media); EMA CFCM: cell-free conditioned culture media of X. budapestensis DSM16442 (EMA); EMC CFCM: cell-free conditioned culture media of X. szentirmaii DSM16338 (EMC); TT01 YELLOW CFCM: cell-free conditioned culture media of Photorhabdus luminescens ssp. akhurstii TT01, yellow colony-color variant (from McConkey agar media); TT01 RED CFCM: cell-free conditioned culture media of P. luminescens ssp. akhurstii TT01 RED colony-color variant (from McConkey agar media).
Table 4.
Gastro-enteral antimicrobial potential (on Clostridium (CFU) in the lower ileum CFU). The absolute, and relative weights of the immune organs (bursa of Fabricius, and spleens of XENOFOOD[HM1] .
Table 4.
Gastro-enteral antimicrobial potential (on Clostridium (CFU) in the lower ileum CFU). The absolute, and relative weights of the immune organs (bursa of Fabricius, and spleens of XENOFOOD[HM1] .
| TREATMENTS | SPLEEN | bursa of Fabricius | CLOSTRIDIUM CFU | |||||
| weight | weight | size | individual bursa/spleen | (lower ileum) | ||||
| N | mg | mg | mm | ratio | ||||
| C (Control) | 10 | 2266.6 | 2761.0 | 25.2 | 1.39 | 149.9 | ||
| T (Xenofood-fed) | 10 | 1618.0 | 3618.2 | 26.9 | 2.38 | 48.1 | ||
| t | +2.09 | +3.16 | 3.02 | 3.7 | -2.128733 | |||
| P | 0.056 | 0.006 | 0.009 | 0.006 | 0.028 | |||
| Significance | NS | ** | ** | * | * | |||
1 Table 4: The average measurements (sizes and (weights) of the spleen, as well as those of the bursas of Fabricius.
Table 5.
Body Weights.
| TREATMENTS | N | DAYS OF THE EXPERIMENT | ||
| 10 | 14 | 42 | ||
| C (Control) | 34 | 252.9 +/- 33.4 | 1196.8 +/- 123 | 2842.5 +/- 184 |
| T (XENOFOOD-fed) | 34 | 271.3 +/- 39.4 | 1132.7+/- 162 | 2984.2+/- 207 |
| T-value | -177 | 187 | - 1.02 | |
| P-value | 0.08 | 0.07 | 0.32 | |
| Significance | ns | ns | ns | |
1Table 5 footer: During the 42-day experiment, no animal was lost. We did not detect any significant differences either in the body weights.
Table 6.
Feed Conversion Rates of XENOFOOD – Fed Chicken and Control Chicken.
| TREATMENTS | N | GROWTH PERIODS (DAYS) | ||
| 1-10 | 11-24 | 1-24 | ||
| C (Control) | 34 | 1.06 | 1.68 | 1.53 |
| T (XENOFOOD-fed) | 34 | 0.97 | 1.71 | 1.52 |
| Significance | ns | ns | ns | |
Table 7.
A list of the non-ribosomal templates antimicrobial peptides discovered in different Xenorhabdus species. R
Table 7.
A list of the non-ribosomal templates antimicrobial peptides discovered in different Xenorhabdus species. R
| Natural NR-AMPs | Xenorhabus specoes | refeence |
|---|---|---|
| Xenofuranone A and B | X. szetirmaii (EMC) DSM(16338)T | [45] Brachmann, et al., 2006 |
| Nemaucin | X. cabanillasii | [46] Gualtieri, et al., 2009)a,[47] Gualtieri, et al., 2012 |
| Fabclavine | X. budapestensis DSM16342)T (EMA) | [48] (Fuchs et al., 2012 |
| Fabclavine, A, B | X. szentirmaii DSM(16338)T (EMC) | [49] (Fuchs et al., 2014) |
| Fabclavine, biosynthetic intermediers, derivatives, and analogs |
X. szentirmaii DSM(16338)T (EMC) All but e few Xenorhabdus |
[50] Wenski et al., 2019. [51] Wenski et al., 2020 |
|
Cabanillasin |
X. cabanillasii, X. khoisanae, SB10 |
[52] Houard et al., 2013, |
| PAX peptides | X. nematophila | [54] Gualtieri et al, 2009b |
| [55] Fuchs et al, 2011), | ||
| X. khoisanae, SB10 | [53] Dreyer, et al., 2019 | |
| Odilorhabdins | X. riobrave | [56] Isaacson and Webster, 2013 |
| [57] Pantel, et al, 2018 | ||
| [58] Sarciaux et al., 2018 | ||
| [69] Racine, and Gualtieri 2019 | ||
| [60] Lanois-Nouri, et al., 2022. | ||
| Anti-oomycete peptides |
X. budapestensis NMC-10 |
[61] (Xiao et al., 2012) |
|
Xenortide |
X. nematophila |
[62] (Reimer, 2014 |
|
Xenortide A-D |
X. nematophila |
[63] Esmati, et al., 2018 |
| Rhabdopeptide | X. nematophila | [64] Reimer at al., 2013 |
| Rhabdopeptide (with nematicide activity) | X. budapestensis SN84 | [65] (Bi et l., 2018) |
| Rhabdoopeptide/xenortide-like peptides | Xenorhabdus innexi | [66] Zhao, 2018 |
| New cyclic depsipeptide xenematide F, and G, (anti-oomycete activity) | X. budapestensis SN84 | [67] (Xi et al, 2019 |
| Szentiamide |
X. szentirmaii DSM16338T (EMC) |
[68] Ohlendorf, et al., 2011) |
| [69] (Nollmann, et al, 2012) | ||
| Genomic information: 71 NR-AMP operons in. | X. szetirmaii (EMC)T DSM16338 | [70] Gualtieri et al.,2014 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



