Submitted:
10 July 2025
Posted:
31 July 2025
Read the latest preprint version here
Abstract
Why have simple carbon-based materials (CBMs) evolved stepwise into complex, orderly, and diverse organisms and societies? To answer this fundamental unsolved scientific question, here we extend Darwinian theory into the Carbon-Based Evolutionary Theory (CBET) to schematically interpret chemical, biological, and social evolution, through holistic reasoning and empirical evaluation. CBET involves nine hierarchies of CBMs, seven reasoning steps, and four feedback loops, which are based on three fundamental principles governing the interplay of the environment, energy, driving forces, changes, structures, and functions. These principles collectively elevate CBM hierarchies stepwise and increase the CBM diversity. They lead to natural selection, which enhances the fitness and orderliness of HHCBMs according to mathematical rules. CBET corrects multiple misconceptions about natural selection and highlights the balance between competition and inclusiveness, selfishness and altruism, and other pivotal social notions. CBET supports the harmonious development of human societies and can thus bridge natural and social sciences.
Keywords:
carbon-based material
; mechanism
; chemical evolution
; biological evolution
; social evolution
; natural selection
; theory
Scholars have been captivated for centuries to unveil the mechanisms underlying the stepwise evolution of carbon-based materials (CBMs) on Earth from simple molecules to complex life and societies, which encompasses chemical, biological, and social evolution (Figure 1) [1,2,3]. Chemical evolution refers to the evolution from small carbon-containing molecules (CCMs) to large CCMs, and further to multiple-molecule aggregates (MMAs) [4,5]. Biological evolution refers to the origin and evolution of life. Social evolution refers to the origin and evolution of animal social organizations (e.g., ant colonies) and human social organizations [6,7,8]. Darwinian theory, which profoundly explains certain aspects of biological evolution, does not address chemical and social evolution. Here we extend Darwinian theory into the Carbon-Based Evolutionary Theory (CBET) to schematically interpret mechanisms underlying the entire evolution of CBMs.
Materials and Methods
Selection and classification of the core concept in CBET. The core concept of organisms in Darwinian theory is extended into CBMs in CBET. Many CBMs are classified into nine hierarchies (Figure 1), according to the rationale that higher-hierarchy CBMs (HHCBMs) are formed by lower-hierarchy CBMs (LHCBMs) and other materials through physical, chemical, biological, and/or social interaction. H1-CBMs refer to carbon atoms, which possibly originated within the interiors of giant or supergiant stars [9]. H2-CBMs refer to small CCMs, such as CH4, CO2, and HCN. H3-CBMs refer to various CCMs of intermediate size, such as lysine, gentamicin, and glucose. H4-CBMs refer to large CCMs, such as lipids, proteins, and nucleic acids. H5-CBMs refer to MMAs, such as phospholipid bilayers and cellulose microfibrils. H6-CBMs refer to individual cells, such as bacteria and cells in rats, composed of numerous H5-CBMs and other materials. H7-CBMs refer to multicellular organisms, such as pines and rabbits. H8-CBMs refer to single-level social organizations, such as ant colonies, bee colonies, and many small companies. These organizations are composed of animal or human individuals cooperating closely with different duties. H9-CBMs refer to multiple-level social organizations, such as universities, armies, and countries. H8-CBMs and H9-CBMs are often regarded as superorganisms [6]. HHCBMs and LHCBMs are defined by comparing their hierarchies. For instance, H6-CBMs are HHCBMs compared to H5-CBMs but LHCBMs compared to H7-CBMs. Notably, no clear lines separate the CBM hierarchies above. For instance, some peptides are between H3-CBMs and H4-CBMs, some viruses are between H4-CBMs and H5-CBMs. Certain CBMs that occupy intermediate positions between hierarchies can facilitate the evolution of HHCBMs into higher hierarchies.
Reasoning and evaluation of CBET. Three universal, fundamental principles were derived through inductive synthesis. These principles and three universal mathematical rules are used to deduce the mechanisms underlying natural selection and chemical, biological, and social evolution. Then this theory was refined through repeated evaluation with a series of randomly selected facts in diverse domains, until no counterevidence was identified (Text S1).
Results
Derivation of Three Universal, Fundamental Principles
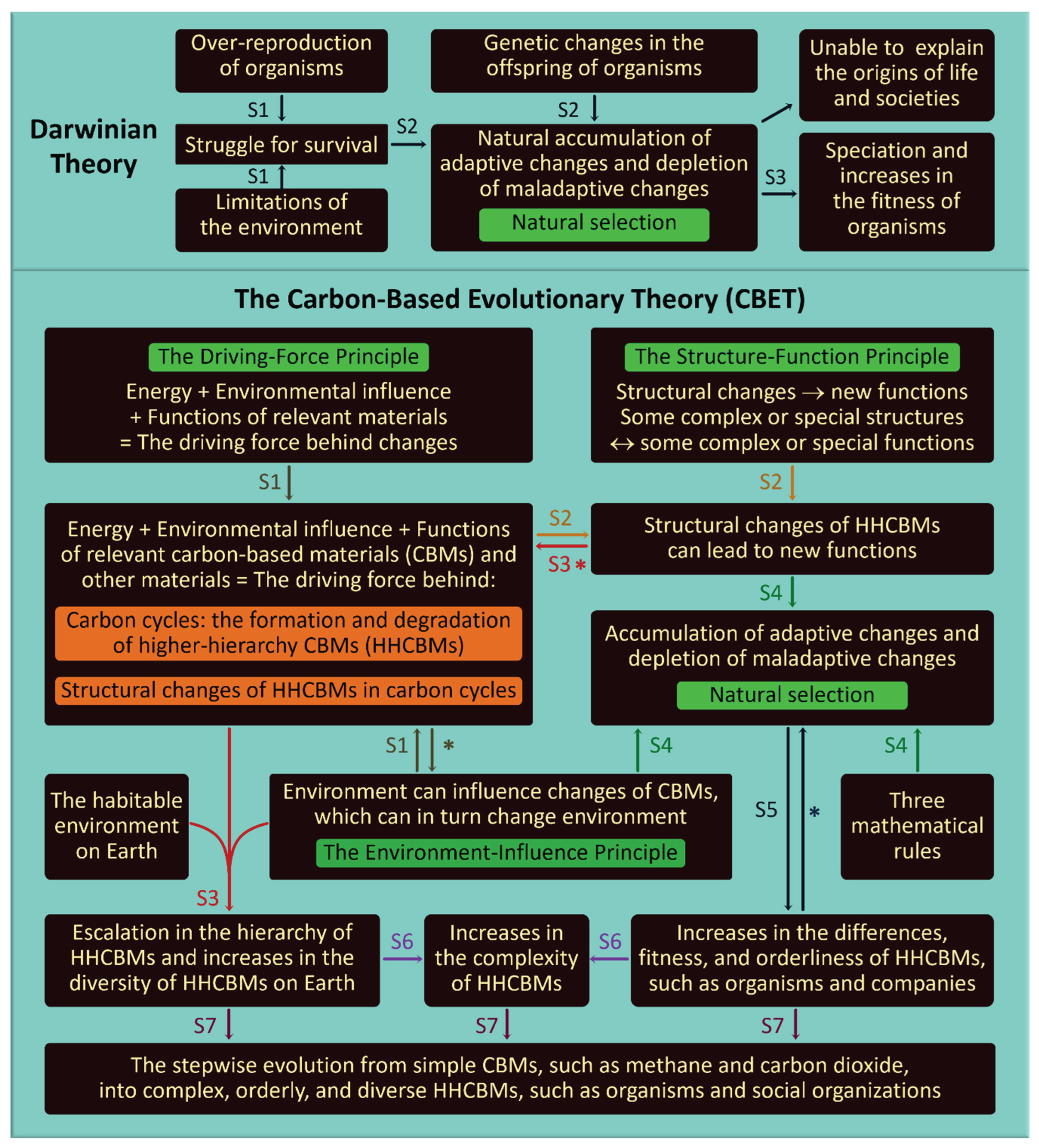
The straightforward reasoning of Darwinian theory is extended into seven complex but comprehensible reasoning steps in CBET (Figure 2). Darwinian theory claims that organisms’ offspring usually exhibit heritable changes in their traits and struggle for survival due to over-reproduction of organisms and environmental limitations, and those offspring with adaptive heritable changes are more likely to survive and reproduce than those with maladaptive heritable changes. This process, termed natural selection, leads to the emergence of new species and increases in the fitness of organisms. By contrast, the reasoning of CBET starts with the following three basic principles, which are derived from inductive synthesis of empirical evidence across physics, chemistry, biology, and social sciences (Figure 2).
(1) The Environment-Influence Principle states that the environment can provide energy, constituent materials, aiding factors, competitive factors, and other conditions for many structural changes of materials and can thus influence the changes, which, in turn, can influence the environment. For instance, Earth, with its abundant liquid water, CBMs, and energy flow, constitutes an extensive habitable environment conducive to CBM evolution [10].
(2) The Driving-Force Principle states that, qualitatively, the synergy of energy flow, environmental factors (as external causes), and functions of relevant materials (as internal causes) drives structural changes in CBMs and other materials (e.g., water, cars, and computers). This principle highlights the role of energy, which is a concrete and fundamental concept across physics, chemistry, biology, and social sciences, in driving changes. Change-driving energy can originate from the materials themselves, the environment, or their interaction, and its effects are influenced by the functions of relevant materials and the environment. The structures, functions, and environments in this principle can become more complex for higher hierarchies, making this principle applicable to all hierarchies (Text S1).
Figure 2.
Three reasoning steps of Darwinian theory are extended into seven reasoning steps (S1 to S7) in CBET. Asterisks denote feedback and arrows indicate causation. For CBET, three principles drive CBM structural changes (S1), which lead to new functions (S2). The new functions feedback to S1, escalating HHCBM hierarchies and increasing HHCBM diversity (S3). The above changes in HHCBMs along with environmental limitations and mathematical rules lead to natural selection (S4), which increases the fitness and orderliness of HHCBMs and feeds back into itself to intensify competition (S5). Hierarchical elevation of CBMs and orderliness increase of HHCBMs enhance HHCBM complexity (S6). The steps above drive the stepwise evolution from simple CBMs into complex, orderly, and diverse HHCBMs (S7). (3) The Structure-Function Principle states that: (a) some complex or special structures can generate complex or special functions; (b) some complex or special functions can in turn support complex or special structures, and (c) some structural changes can generate new functions. Accordingly, due to the special structures of carbon atoms, only carbon atoms can serve as the molecular backbone of many stable and water-soluble small, middle, or large CCMs [11]. These highly diverse and water-soluble CCMs are essential for the natural formation of extraordinarily complex structures of life, which are essential for the extraordinarily complex function of reproduction of organisms. Consequently, CBMs are likely the only materials capable of naturally evolving from simple structures into life [11]. Some HHCBMs are relatively stable due to their complex or special structures. For instance, some proteins have been preserved in fossils for millions of years [12], and various H3-CBMs could have been preserved for billions of years in meteorites [13], and humans can typically live for decades. Furthermore, due to their complex structures, H6-CBMs and H7-CBMs possess the information-utilization function, involving encoding, reading, translating, mutating, storing, and/or transmitting information primarily through the interaction of nucleic acids and proteins [2]. This function is crucial for organisms to reproduce relatively accurately and accumulate advantageous changes, such as those of non-random mutations. In addition to genetic information, H6-CBMs and H7-CBMs have developed various information transmission pathways, such as nerves, due to their complex structures to respond to numerous inside or outside changes, while H8-CBMs and H9-CBMs have harnessed other forms of information due to their complex structures, manifesting in scent (volatile compounds), sound (language), actions, script, and/or electromagnetic waves.
Figure 2.
Three reasoning steps of Darwinian theory are extended into seven reasoning steps (S1 to S7) in CBET. Asterisks denote feedback and arrows indicate causation. For CBET, three principles drive CBM structural changes (S1), which lead to new functions (S2). The new functions feedback to S1, escalating HHCBM hierarchies and increasing HHCBM diversity (S3). The above changes in HHCBMs along with environmental limitations and mathematical rules lead to natural selection (S4), which increases the fitness and orderliness of HHCBMs and feeds back into itself to intensify competition (S5). Hierarchical elevation of CBMs and orderliness increase of HHCBMs enhance HHCBM complexity (S6). The steps above drive the stepwise evolution from simple CBMs into complex, orderly, and diverse HHCBMs (S7). (3) The Structure-Function Principle states that: (a) some complex or special structures can generate complex or special functions; (b) some complex or special functions can in turn support complex or special structures, and (c) some structural changes can generate new functions. Accordingly, due to the special structures of carbon atoms, only carbon atoms can serve as the molecular backbone of many stable and water-soluble small, middle, or large CCMs [11]. These highly diverse and water-soluble CCMs are essential for the natural formation of extraordinarily complex structures of life, which are essential for the extraordinarily complex function of reproduction of organisms. Consequently, CBMs are likely the only materials capable of naturally evolving from simple structures into life [11]. Some HHCBMs are relatively stable due to their complex or special structures. For instance, some proteins have been preserved in fossils for millions of years [12], and various H3-CBMs could have been preserved for billions of years in meteorites [13], and humans can typically live for decades. Furthermore, due to their complex structures, H6-CBMs and H7-CBMs possess the information-utilization function, involving encoding, reading, translating, mutating, storing, and/or transmitting information primarily through the interaction of nucleic acids and proteins [2]. This function is crucial for organisms to reproduce relatively accurately and accumulate advantageous changes, such as those of non-random mutations. In addition to genetic information, H6-CBMs and H7-CBMs have developed various information transmission pathways, such as nerves, due to their complex structures to respond to numerous inside or outside changes, while H8-CBMs and H9-CBMs have harnessed other forms of information due to their complex structures, manifesting in scent (volatile compounds), sound (language), actions, script, and/or electromagnetic waves.
Seven Reasoning Steps of CBET
The Driving-Force Principle suggests that the synergy of energy flow, environmental factors, and functions of relevant materials drives many changes of CBMs and other materials (e.g., water) constitutes the driving force underlying all changes of CBMs, including the formation, degradation, and structural changes of HHCBMs in the repeated carbon cycles on Earth (Step 1) [14]. Structural changes of HHCBMs can generate new functions according to the Structure-Function Principle in Step 2. The resultant new functions can feed back to Step 1, leading to the stepwise formation of further higher-hierarchy CBMs from H1-CBMs to H9-CBMs (Step 3), because of the special functions of CBMs and the rare habitable environment of Earth in the known universe [10], as detailed in Text S1. Step 3 can also lead to the increases in the diversity of HHCBMs on Earth, because of the habitable environment of Earth and the functions of many HHCBMs: stability and changeability during the repeated carbon cycles on Earth. Step 4 shows that natural selection directly results from changes of HHCBMs, the Environment-Influence Principle, and three mathematical rules elaborated below. Natural selection is a process which favors those HHCBMs having the changes aiding the formation or sustenance of the HHCBMs and disfavors those HHCBMs having the changes harming the formation or sustenance of the HHCBMs (Step 5). Consequently, natural selection leads to the increases in the fitness and orderliness of HHCBMs. Natural selection can feed back into itself to intensify the competition in natural selection over time, particularly for predators and prey, parasites and hosts, as well as plants and herbivores [15]. The hierarchical elevation of CBMs in Step 3, coupled with the increased orderliness and fitness of HHCBMs resulted from natural selection in Step 5, leads to the increased complexity of HHCBMs (Step 6). The six steps above explicitly interpret why simple CBMs could evolve into complex, orderly, and diverse organisms and social organizations (Step 7). The steps above involve three feedback loops, which are termed evolutionary feedback loops. They denote that CBM evolution affects the environment and functions of CBMs and thus, in turn, affects itself. This constitutes the fourth evolutionary feedback loop. For instance, the evolution of human societies significantly influences which molecules (e.g., gentamicin) and organisms (e.g., chicken) thrive on Earth.
Mathematical Rules of Natural Selection
(1) The Quantity-Existence Rule: mathematically, an HHCBM exists if its cumulative formation exceeds its cumulative degradation, regardless of whether it is fitter than its ancestor or other HHCBMs. Accordingly, organisms, multicellular organisms, organisms with sexual reproduction, and endothermic animals can survive even if they are not inherently fitter than inanimate materials, unicellular organisms, organisms with asexual reproduction, or ectothermic animals, provided their overall formation exceeds their overall degradation. This rule is defined in CBET as “survival of the fit,” which is more accurate than the prejudicial phrase “survival of the fittest”, a definition of natural selection in Darwinian theory [1], since the fittest organism could become extinct in a harsh environment, and some less fit organisms or people could become prevalent in an amiable environment.
(2) The Quantity-Change Rule: mathematically, the quantity of an HHCBM is increasing (or decreasing) if its formation-to-degradation ratio is >1 (or <1).
The two rules above are self-evident and can be deduced mathematically from the following equation (Text S1):
where the current quantity of an HHCBM equals the cumulative formation minus the cumulative degradation.
(3) The Quantity-Limitation Rule: the total quality of an HHCBM is limited by its functions and the energies, materials, and other conditions in the environment. This rule is self-evident and can be deduced from the three principles above.
The three mathematical rules demonstrate that the fate of an HHCBM is determined by its overall performance of the formation and sustenance (OPFS), which can be quantitatively measured with the quantity fitness using the following equation:
where the fitness from time a to time b is calculated as the formation (rate) divided by the degradation (rate) during this interval. This equation circumvents the flaw of the traditional calculation of fitness (the number of offspring an individual leaves in the next generation [2])—namely, its inability to describe the fitness of molecules, social organizations, and worker bees, which do not possess the reproduction function.
In biology, the OPFS is determined by the collective contribution of all components, traits, and hierarchies of an HHCBM. On the other hand, certain traits of HHCBMs, such as the sprinting ability of antelopes, the cooperative behavior of wolves, or the intelligence of humans, can markedly influence the OPFS of an HHCBM. Therefore, natural selection emphasizes both all-round development and specialized development.
Correcting Misconceptions About Natural Selection
CBET corrects the widespread misconception that all biological traits (e.g., giraffes’ long necks [16]) are universally adaptive under natural selection. Natural selection in CBET allows the existence of neutral or even detrimental traits, provided the cumulative formation of an HHCBM exceeds its cumulative degradation. For instance, human infants are more fragile compared to those of some other mammals, and this generally disadvantageous trait can be offset by some advantageous traits of humans. Similarly, natural selection allows the existence of African buffaloes with strong combat abilities but slow running speeds and gazelles with weak combat abilities but fast running speeds. Natural selection also allows the prevalence of neutral mutations in genomes [2], as these mutations have minimal effects on the OPFS. Additionally, due to the Environment-Influence Principle, many traits are advantageous in some environments but detrimental in some other environments. Furthermore, a trait can be advantageous in some respects but detrimental in some other respects. For instance, a tall man is usually advantageous to a short man in fighting and can be disadvantageous in surviving famines. Together, these phenomena show a type of inclusiveness in natural selection and indicate that a trait can be generally advantageous, neutral, or detrimental in natural selection.
CBET corrects the widespread misconception that natural selection entails fierce competition rather than inclusiveness. Natural selection does represent competition and can feed back into itself to intensify the competition in natural selection over time (Figure 2). Furthermore, competition in natural selection can be intensified over time for other reasons. For instance, the increasing diversity of HHCBMs driven by the Driving-Force mechanism can result in a growing scarcity of the materials required for their formation, according to the Quantity-Limitation Rule. Similarly, some environmental changes, such as asteroid impacts, volcanoes, climate changes, and those caused by CBM evolution, can lead to difficulties for the existence of certain HHCBMs [2]. For instance, the increase in photosynthetic bacteria around 2.5 billion years ago likely resulted in a significant increase in the concentration of oxygen in the air, posing a disaster for anaerobic bacteria, although it provided opportunities for aerobic bacteria [2,17]. On the other side, inclusiveness is pervasive throughout evolution, as evident in the generally increasing diversity of molecules, organisms, and social organizations during CBM evolution, and the fact that natural selection in CBET allows neutral or even detrimental traits to persist, provided cumulative formation exceeds cumulative degradation.
CBET corrects the widespread misconception that natural selection represents selfishness rather than collaboration and altruism [1,18,19]. Selfishness is essential for animals to secure sufficient energy, resources, favorable environments, and mating opportunities to thrive on Earth. Paradoxically, collaborative and altruistic behaviors are also pervasive throughout evolution. Collaboration within HHCBMs is essential for HHCBMs to obtain advantages to compete to survive natural selection, and thus natural selection leads to the increases in internal collaboration within HHCBMs. Furthermore, many molecules act as catalysts, energy sources, or building blocks for the synthesis of other molecules within organisms [4,5,20]. In H6-CBMs (e.g., bacteria), many molecules aid in the transmission of nucleic acids to the next generation. In H7-CBMs (e.g., rats), many cells assist in the passage of reproductive cells to subsequent generations. In H8-CBMs (e.g., ant colonies), many individual animals support the reproduction of a select few within the social group. In some H9-CBMs, many soldiers, police officers, and firefighters sacrifice themselves for the good of their society or other individuals.
CBET corrects the widespread misconception that natural selection underscores only heritable factors [1,2]. Non-heritable factors, such as vaccination and education, can significantly affect the OPFS of relevant organisms and the existence of the relevant organisms. Their existence is required for the occurrence and accumulation of genetic changes. These non-heritable factors can thus participate in natural selection. Similarly, epigenetic changes, no matter whether they are inheritable and non-heritable, affect the OPFS of relevant organisms and participate in natural selection [21].
Empirical Evaluation of CBET
All CBET’s viewpoints passed the empirical evaluation with a series of facts about molecules, organisms, and social organizations (Text S1).
Discussion
As elaborated in Text S1 and exemplified in Table 1, CBET is a coherent constitution-like theory with significant innovations across physics, biology, and the social sciences. It systematically integrates various concrete factors with robust empirical evidence to elaborate an explicit, holistic, and schematic answer to the fundamental question of how simple CBMs have evolved into complex organisms and societies. CBET corrects various misconceptions surrounding natural selection and emphasizes the balance between competition and inclusiveness, selfishness and altruism, individual and collective interests, as well as numerous other pivotal social concepts. CBET underpins the harmonious development of human societies and constructs an interdisciplinary bridge between the natural and social sciences.
While CBET provides guidance for exploring the specific details of chemical, biological, and social evolution in CBMs, it does not furnish those details itself. Moreover, CBET explains the possibility—rather than the inevitability—of evolutionary processes. Paradoxically, these limitations have significantly lowered the barrier to establishing the theory while substantially boosting its credibility.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Text S1.: Part 1. Novelties of CBET, Part 2. Mathematical Formalization of CBET. Part 3. Rationality and Empirical Evidence of CBET.
Author Contributions
Jiming Chen conceived, designed, performed, and financially supported this study; Jiwang Chen performed and financially supported this study; Jiming Chen wrote the manuscript; all authors revised and approved the manuscript.
Acknowledgments
This study was supported by the High-Level Talent Fund of Foshan University (No. 20210036). The authors thank Yiqing Chen, Meng Yang, Feng Tang, Hai Xiang, Ying Li, and many other people for their previous comments on this study. Yiqing Chen contributed greatly the mathematical work of this study.
Conflicts of Interest
The authors declare no competing interests.
References
- Darwin C. 1860. On the Origin of Species. London, UK: John Murray.
- Futuyma DJ, Kirkpatrick M. 2017. Evolution. Sunderland, USA: Sinauer Associates.
- Xie P. 2014. The aufhebung and breakthrough of the theories on the origin and evolution of life. Beijing, China: Science Press.
- Oparin AI. 1969. Chemistry and the origin of life. R. Inst. Chem. Rev. 2, 1–12.
- Fiore M. 2022. Prebiotic chemistry and life’s origin. Cambridge, UK: Royal Society of Chemistry. [CrossRef]
- Nowak M, Tarnita C, Wilson E. 2010. The evolution of eusociality. Nature 466, 1057–1062. [CrossRef]
- Birch J. 2017. The Philosophy of Social Evolution. Oxford, United Kingdom: Oxford Academic. [CrossRef]
- Chen JM, Chen JW. 2000. Root of science—the driving force and mechanisms of the extensive evolution. Beijing, China: Science Press.
- Cowan J. 2012. One of the first of the second stars. Nature 488, 288–289. [CrossRef]
- Seager S. 2013. Exoplanet habitability. Science 340, 577–581. [CrossRef]
- Razeghi M. 2019. The mystery of carbon: An introduction to carbon materials. Bristol, UK: Institute of Physics Publishing.
- Li ZH, Bailleul AM, Stidham TA, Wang M, Teng T. 2021. Exceptional preservation of an extinct ostrich from the Late Miocene Linxia Basin of China. Vertebrata PalAsiatica 59, 229. [CrossRef]
- Heck PR, Greer J, Kööp L, et al. 2020. Lifetimes of interstellar dust from cosmic ray exposure ages of presolar silicon carbide. Proc. Natl. Acad. Sci. U. S. A. 117, 1884–1889. [CrossRef]
- Reichlem DE. 2023. The global carbon cycle and climate change. Amsterdam, Netherlands: Elsevier.
- Benton MJ. 2009. The red queen and the Court Jester: species diversity and the role of biotic and abiotic factors through time. Science 323, 728–732. [CrossRef]
- Wang SQ, Ye J, Meng J, et al. 2022. Sexual selection promotes giraffoid head-neck evolution and ecological adaptation. Science 3, 376(6597): eabl8316. [CrossRef]
- Olejarz J, Iwasa Y, Knoll AH, et al. 2021. The Great Oxygenation Event as a consequence of ecological dynamics modulated by planetary change. Nat. Commun. 12, 3985. [CrossRef]
- Dawkins, R. 1976. The Selfish Gene. Oxford, United Kingdom: Oxford University Press.
- Belcher LJ, Madgwick PG, Kuwana S, et al. 2022. Developmental constraints enforce altruism and avert the tragedy of the commons in a social microbe. Proc Natl Acad Sci U S A. 119(29): e2111233119. [CrossRef]
- Guo X, Fu S, Ying J, Zhao Y. 2023. Prebiotic chemistry: a review of nucleoside phosphorylation and polymerization. Open Biol. 13, 2202–2234. [CrossRef]
- Gómez-Schiavon M, Buchler N E. 2019. Epigenetic switching as a strategy for quick adaptation while attenuating biochemical noise. PLoS Comput. Biol. 15: e1007364 (2019).
Figure 1.
The entire evolution of carbon-based materials (CBMs) from H1-CBMs to H9-CBMs and carbon cycles on Earth. CCM: carbon-containing molecules.
Figure 1.
The entire evolution of carbon-based materials (CBMs) from H1-CBMs to H9-CBMs and carbon cycles on Earth. CCM: carbon-containing molecules.
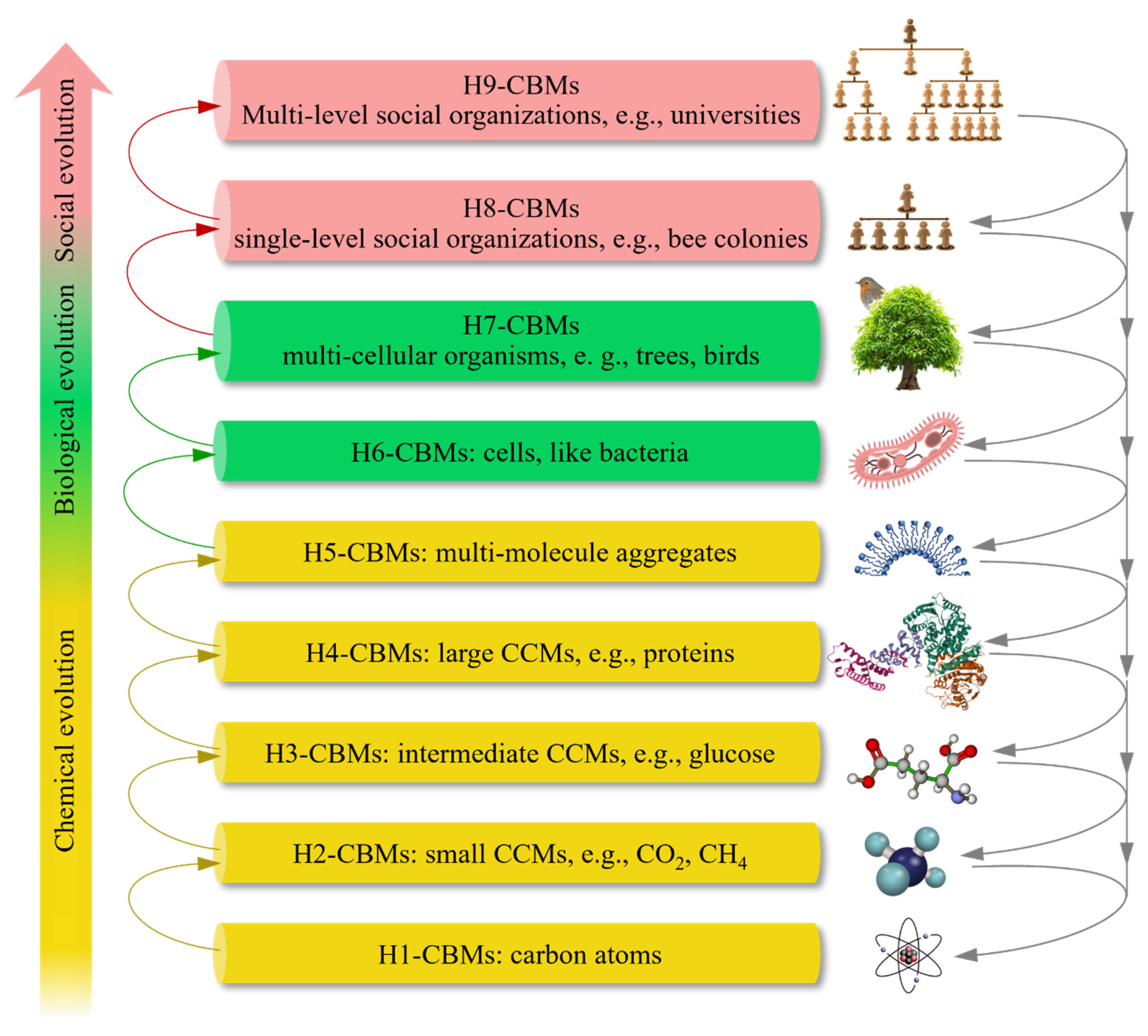

Table 1.
Differences between Darwinian theory and the Carbon-Based Evolutionary Theory (CBET).
| Aspect | Darwinian theory | CBET |
|---|---|---|
| Scope | Biological evolution | Chemical, biological, and social evolution |
| Reasoning | Three steps without feedback loops | Seven steps with feedback loops |
| Significance | Challenging creationism using natural selection | Unveiling the mechanisms shared by chemical, biological, and social evolution |
| Core concept | Organisms | Carbon-based materials (CBMs), classified into nine hierarchies from carbon atoms to social organizations |
| Driving force | Natural selection on heritable variations; no direct link to energy | The synergy of energy, environmental factors, and functions of relevant materials |
| Principles | Natural selection due to genetic variation, survival or reproduction competition, and environmental limitations | Three principles regarding the interplay of the environment, energy, driving force, changes, structures, and functions derived through inductive synthesis of innumerable facts across diverse domains |
| Mechanisms | Natural selection leading to adaptation and speciation | The three principles above leading to the hierarchical escalation of CBMs, the increases in the diversity of high-hierarchy CBMs, and natural selection, which, in turn, leads to adaptation |
| Feedback Loops | No explicit feedback mechanisms | Four evolutionary feedback loops associated with the environment, functions, natural selection, and CBM evolution |
| Tautology | Lacking explanations for the origins of the options of natural selection and thus attracting sustained criticism regarding its potential tautology | CBET is grounded in three fundamental principles derived through inductive synthesis of abundant facts and elucidates the origins of the options of natural selection, fundamentally circumventing tautological reasoning |
| Mathematical rules | Qualitative and inaccurate description of natural selection as “survival of the fittest” | Three rules accurately describing the mathematical requirements regarding the existence and quantitative changes of HHCBMs |
| Definition of fitness | Fitness was not calculated in Darwinian theory. It was defined in modern biology as the number of offspring an individual leaves in the next generation, which cannot address the fitness of worker bees | Fitness during a period is calculated as the formation (or rate) divided by the degradation (or rate) during this interval. This definition can describe the quantitative changes of molecules and social organizations and can accurately describe the fitness of worker bees |
| On biological traits | Usually assuming biological traits (e.g., giraffe’s long necks) are advantageous in natural selection | Usually assuming biological traits (e.g., giraffe’s long necks) can be generally advantageous, neutral, or disadvantageous in natural selection |
| On the origin of life | Unable to explain the origin of life | The seven reasoning steps of CBET schematically explain the possibility and the mechanisms underlying the origin of life |
| On the origin of social development | Unable to explain because it overestimates competition and selfishness. Moreover, it has been used to rationalize slavery, racism, and genocide | The seven reasoning steps of CBET explain the mechanisms underlying the origin and development of social organizations and the importance of collaboration and altruism for social organizations |
| On changes | Focuses on heritable genetic changes and neglects the role of non-heritable factors and epigenetic changes | Clarifies the effects of non-heritable factors (e.g., vaccination and education) and epigenetic changes in natural selection |
| Neutral mutations | Lacks explicit explanation for neutral or detrimental mutations | Neutral/detrimental mutations can persist if cumulative formation > cumulative degradation |
| On inclusiveness | Emphasizes “survival of the fittest” | Emphasizes “survival of the fit” and the balance between competition and inclusiveness |
| On collaboration and altruism | Emphasizes competition, selfishness, and elimination | Emphasizes the balances of competition versus inclusiveness and selfishness versus altruism |
| Explanatory features |
|
|
| Predictability | Qualitatively predicts the fate of an organism in natural selection | Qualitatively predicts various changes of CBMs from the synergy of the tree principles and natural selection |
| Falsifiability | Falsifiable, and its multiple views have been challenged | Falsifiable, and its multiple views have been refined through repeated evaluation and revision |
| Experimental evidence | Given the scientific limitations of his time, Darwin conducted crucial experiments to establish his theory | Modern science has accumulated innumerable experimental findings and abundant knowledge for the establishment and validation of CBET |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



