Submitted:
12 November 2024
Posted:
12 November 2024
Read the latest preprint version here
Abstract
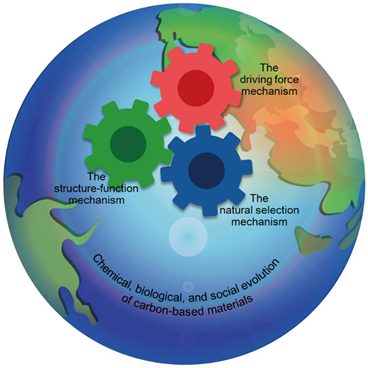
Why did simple carbon-based materials (CBMs) on Earth evolve into complex, orderly, and diverse organisms and societies? Extensive research has yet to explicitly and comprehensively answer this fundamental question in science. Here we propose the Carbon-Based Evolutionary Theory (CBET), potentially providing the first explicit and holistic answer to this question. CBET systematically integrates numerous factors critical to evolution from a bird's-eye view using mathematical logic and the novel concepts of eight hierarchies of CBMs. This integration reveals three key mechanisms: the driving force mechanism, the structure-function mechanism, and the natural selection mechanism, which hierarchically escalates the complexity of CBMs, increases the abundance, diversity, and orderliness of complex CBMs, and leads to chemical, biological, and social evolution of CBMs. CBET clarifies the intricate relationships among energy, structures, functions, and orderliness. It also uncovers the natural roots of multiple pivotal social notions, such as the balance between inclusiveness and competition and the balance between selfishness and altruism. CBET could bridge the natural and social sciences and promote the balanced, harmonious, and peaceful advancement of human societies.
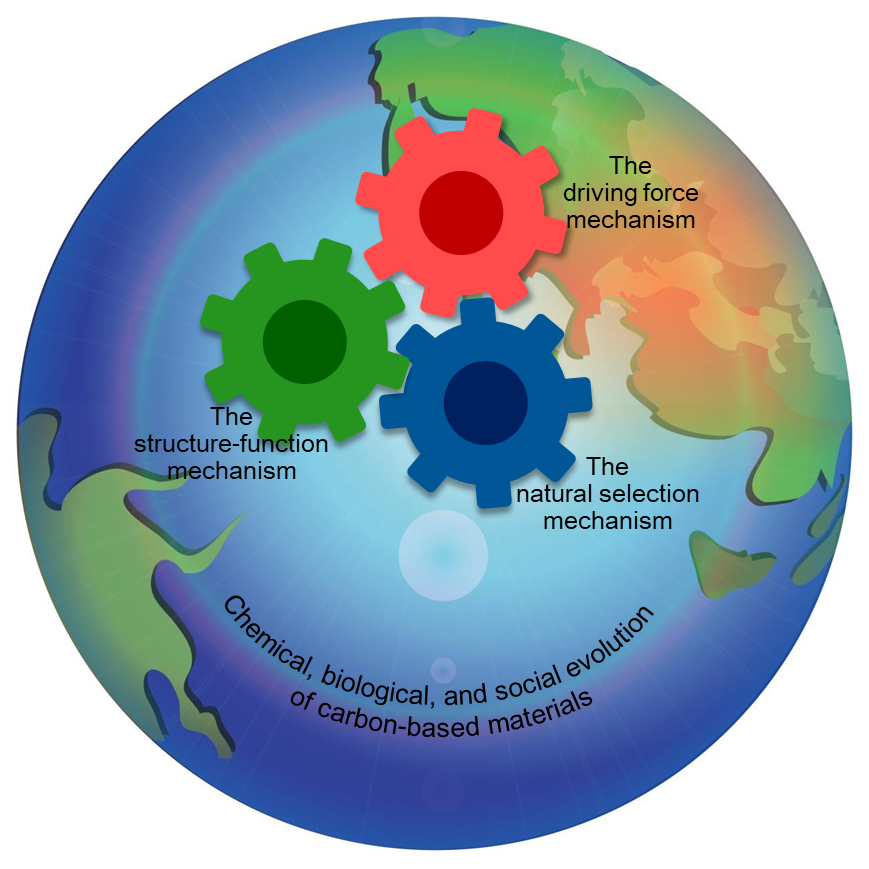
Keywords:
Graphic Abstract
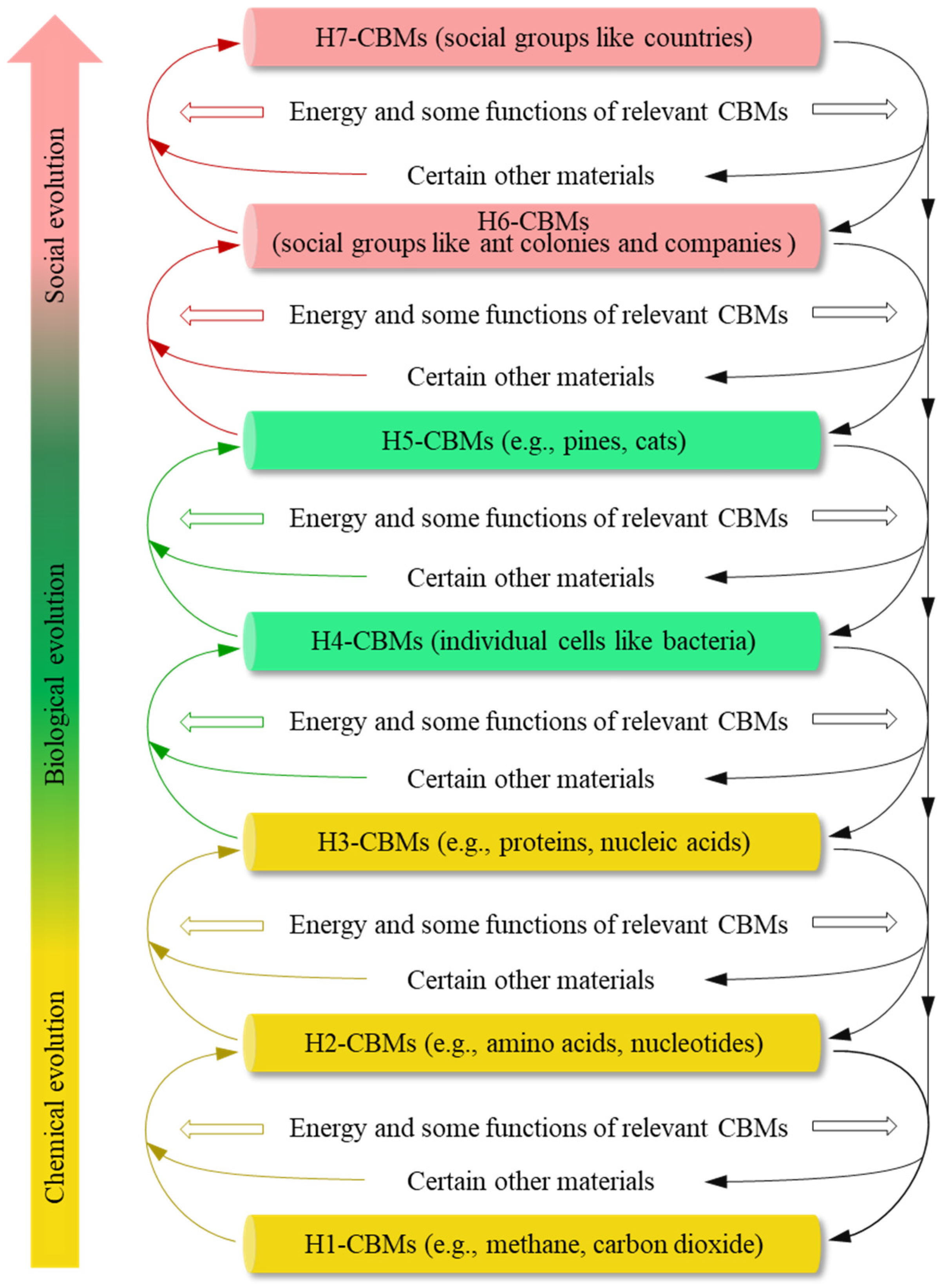
1. Methods and Definitions
2. Results
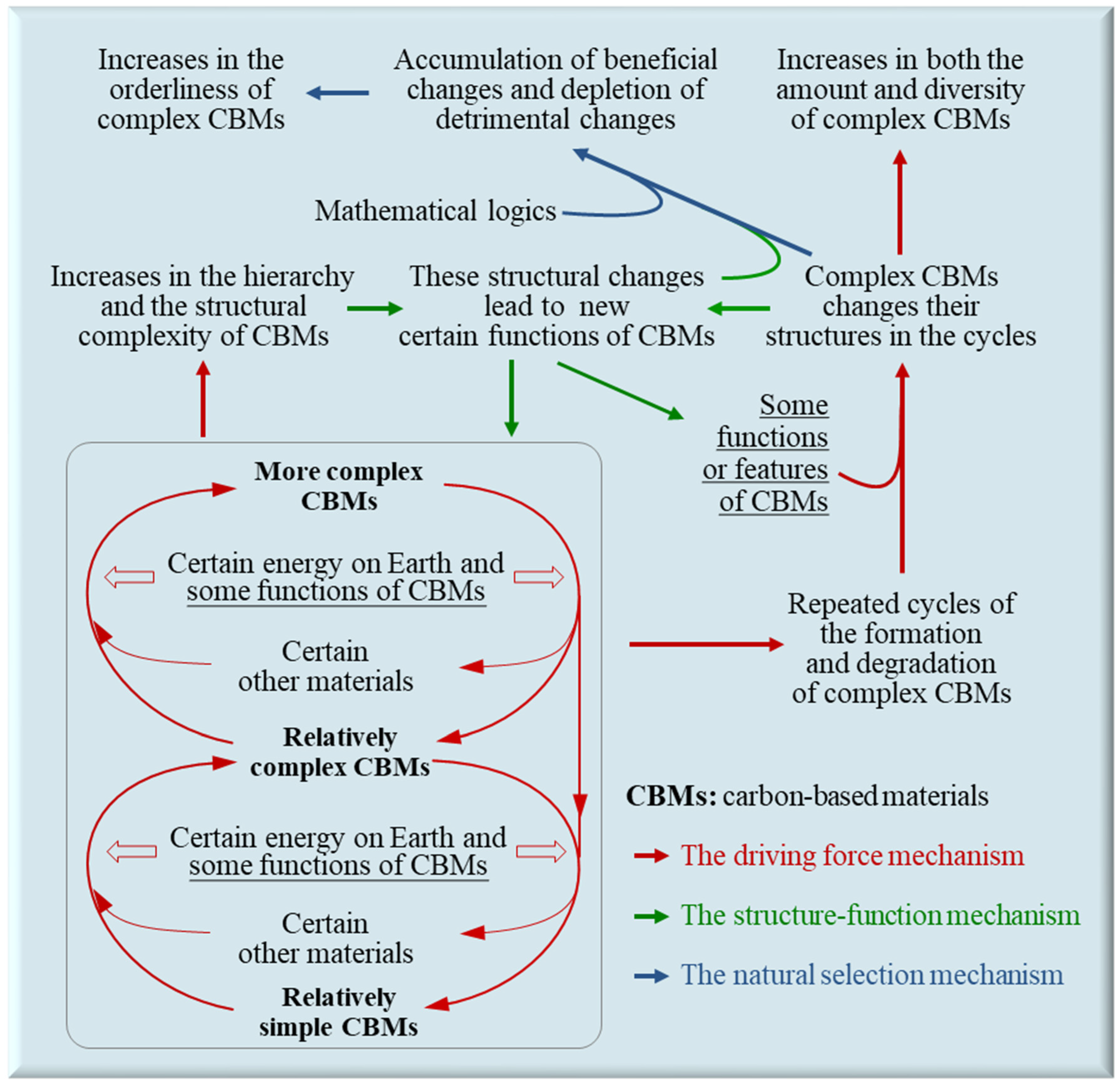
2.1. The Driving Force Mechanism
2.1.1. Origin of the Driving Force Mechanism
2.1.2. Effects of the Driving Force Mechanism
2.1.3. Repeated Cycles of the Formation and Degradation of Complex CBMs
2.2. The Structure-Function Mechanism
2.2.1. Origin of the Structure-Function Mechanism
2.2.2. Effects of the Structure-Function Mechanism
2.3. The Natural Selection Mechanism
2.3.1. Origin of the Natural Selection Mechanism
2.3.2. Features or Effects of Natural Selection
2.3.3. Other Effects of the Natural Selection Mechanism
2.4. The Synergy of the Three Mechanisms Leads to Evolution
3. Novelties of CBET
4. Credibility of CBET
5. Conclusions and Outlook
Author Contributions
Acknowledgments
Conflict of Interest
References
- Xie, P. 2014 The aufhebung and breakthrough of the theories on the origin and evolution of life. Beijing, China: Science Press.
- Schrödinger E. 2012 What is Life. Cambridge, UK: Cambridge University Press.
- Prigogine I. 1978 Time, structure and fluctuation. Science 201, 777–785. [CrossRef]
- Dewar RC. 2010 Maximum entropy production and plant optimization theories. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 365, 1429–1435. [CrossRef]
- Oparin AI. 1969 Chemistry and the origin of life. R. Inst. Chem. Rev. 2, 1–12. [CrossRef]
- Guo X, Fu S, Ying J, Zhao Y. 2023 Prebiotic chemistry: a review of nucleoside phosphorylation and polymerization. Open Biol. 13, 2202–2234. [CrossRef]
- Sumie Y, Sato K, Kakegawa T, Furukawa Y. 2023 Boron-assisted abiotic polypeptide synthesis. Commun. Chem. 6, 89. [CrossRef]
- de Graaf R, De Decker Y, Sojo V, Hudson R. 2023 Quantifying catalysis at the origin of life. Chemistry 29, e202301447. [CrossRef]
- Nogal N, Sanz-Sánchez M, Vela-Gallego S, Ruiz-Mirazo K, de la Escosura A. 2023. The protometabolic nature of prebiotic chemistry. Chem. Soc. Rev. 52, 7359–7388. [CrossRef]
- Chieffo C, Shvetsova A, Skorda F, Lopez A, Fiore M. 2023 The origin and early evolution of life: Homochirality emergence in prebiotic environments. Astrobiology 23, 1368–1382. [CrossRef]
- Fiore M. 2022 Prebiotic chemistry and life's origin. Cambridge, UK: Royal Society of Chemistry. [CrossRef]
- Farías-Rico JA, Mourra-Díaz CM. 2022 A short tale of the origin of proteins and ribosome evolution. Microorganisms 10, 2115. [CrossRef]
- Ershov B. 2010 Natural radioactivity and chemical evolution on the early Earth. Proc. Natl. Acad. Sci. U. S. A. 107, 2763–2768. [CrossRef]
- Futuyma DJ, Kirkpatrick M. 2017. Evolution. Sunderland, USA: Sinauer Associates.
- Rudman LA, Saud LH. 2020 Justifying social inequalities: The role of social Darwinism. Pers. Soc. Psychol. B. 46, 1139–1155. [CrossRef]
- Nowak M, Tarnita C, Wilson E. 2010 The evolution of eusociality. Nature 466, 1057–1062. [CrossRef]
- Bell, J. 2019. The Earth book. Sterling, USA: Sterling.
- Razeghi, M. 2019 The mystery of carbon: An introduction to carbon materials. Bristol, UK: Institute of Physics Publishing.
- Cowan J. 2012 One of the first of the second stars. Nature 488, 288–289. [CrossRef]
- Seager S. 2013 Exoplanet habitability. Science 340, 577–581. [CrossRef]
- Borgnakke C, Sonntag RE. 2022 Fundamentals of thermodynamics. Hoboken, United States: Wiley.
- Schmitt-Kopplin P, Gabelica Z, Gougeon RD, et al. 2010. High molecular diversity of extraterrestrial organic matter in Murchison meteorite revealed 40 years after its fall. Proc. Natl. Acad. Sci. U. S. A. 107, 2763–2768. [CrossRef]
- Dodd MS, Papineau D, Grenne T, et al. 2017 Evidence for early life in Earth's oldest hydrothermal vent precipitates. Nature 543, 60–64. [CrossRef]
- Weiss MC, Sousa FL, Mrnjavac N, et al. 2016 The physiology and habitat of the last universal common ancestor. Nat. Microb. 1, 16116. [CrossRef]
- Stobart CC, Moore ML. 2014 RNA virus reverse genetics and vaccine design. Viruses 6, 2531–2550. [CrossRef]
- Hutchison CA 3rd, Chuang RY, Noskov VN, et al. 2016 Design and synthesis of a minimal bacterial genome. Science 351, aad6253. [CrossRef]
- Xie P. 2021 Who is the missing "matchmaker" between proteins and nucleic acids? Innovation (Camb) 2, 100120. [CrossRef]
- Han TM, Runnegar B. 1992 Megascopic eukaryotic algae from the 2.1-billion-year-old Negaunee iron-formation, Michigan. Science 257, 232–235. [CrossRef]
- Zhu S, Zhu M, Knoll A, et al. 2016 Decimetre-scale multicellular eukaryotes from the 1.56-billion-year-old Gaoyuzhuang formation in North China. Nat. Commun. 7, 11500. [CrossRef]
- Anderson RP, Woltz CR, Tosca NJ, Porter SM, Briggs DEG. 2023 Fossilisation processes and our reading of animal antiquity. Trends Ecol. Evol. 38, 1060–1071. [CrossRef]
- Zhao Z, Yin X, Shih C, Gao T, Ren D. 2020 Termite colonies from mid-Cretaceous Myanmar demonstrate their early eusocial lifestyle in damp wood. Natl. Sci. Rev. 7, 381–390. [CrossRef]
- Mera-Rodríguez D, Jourdan H, Ward PS, Shattuck S, Cover SP, Wilson EO, Rabeling C. 2023 Biogeography and evolution of social parasitism in Australian Myrmecia bulldog ants revealed by phylogenomics. Mol. Phylogenet. Evol. 186, 107825. [CrossRef]
- Plowers N. 2010. An introduction to eusociality. Nature Education Knowledge 3, 7.
- Majelantle TL, Ganswindt A, Hart DW, Hagenah N, Ganswindt SB, Bennett NC. 2024 The dissection of a despotic society: exploration, dominance and hormonal traits. Proc. Biol. Sci. 291, 20240371. [CrossRef]
- Li ZH, Bailleul AM, Stidham TA, Wang M, Teng T. 2021 Exceptional preservation of an extinct ostrich from the Late Miocene Linxia Basin of China. Vertebrata PalAsiatica 59, 229. [CrossRef]
- Heck PR, Greer J, Kööp L, et al. 2020 Lifetimes of interstellar dust from cosmic ray exposure ages of presolar silicon carbide. Proc. Natl. Acad. Sci. U. S. A. 117, 1884–1889. [CrossRef]
- Reichlem, DE. 2023 The global carbon cycle and climate change. Amsterdam, Netherlands: Elsevier.
- von Bertalanffy, L. 1968 General system theory: Foundations, development, applications. George Braziller.
- Lovinger GJ, Sak MH, Jacobsen EN. 2024 Catalysis of an SN2 pathway by geometric preorganization. Nature 632, 1052–105. [CrossRef]
- Stone EA, Cutrona KJ, Miller SJ. 2020 Asymmetric catalysis upon helically chiral loratadine analogues unveils enantiomer-dependent antihistamine activity. J. Am. Chem. Soc. 142, 12690–12698. [CrossRef]
- Morrell, DG. 2019 Catalysis of organic reactions. Boca Raton, USA: CRC Press.
- Kilpeläinen J, Finér L, Neuvonen S, et al. 2009 Does the mutualism between wood ants (Formica rufa group) and Cinara aphids affect Norway spruce growth? For Ecol Manage 257, 238–243.
- Benton MJ. 2009 The red queen and the Court Jester: species diversity and the role of biotic and abiotic factors through time. Science 323, 728–732. [CrossRef]
- Suen G, Teiling C, Li L, et al. 2011 The genome sequence of the leaf-cutter ant Atta cephalotes reveals insights into its obligate symbiotic lifestyle. PLoS Genet. 7, e1002007. [CrossRef]
- Kim E, Fang X, Fushan A, et al. 2011 Genome sequencing reveals insights into physiology and longevity of the naked mole rat. Nature 479, 223–227. [CrossRef]
- Moffett, MW. 2010 Adventures among ants. University of California Press is located in Oakland, United States: University of California Press.
- SIPRI Military Expenditure Database. Available online: https://www.sipri.org/databases/milex (accessed on 9 November 2024).
- Wilson Mantilla GP, Chester SGB, Clemens WA, et al. 2021 Earliest Palaeocene purgatoriids and the initial radiation of stem primates. R. Soc. Open Sci. 8, 210050. [CrossRef]
- Olejarz J, Iwasa Y, Knoll AH, et al. 2021 The Great Oxygenation Event as a consequence of ecological dynamics modulated by planetary change. Nat. Commun. 12, 3985. [CrossRef]
- Bejan A. 2023 The principle underlying all evolution, biological, geophysical, social and technological. Philos. Trans. A. Math. Phys. Eng. Sci. 381(2252), 20220288. [CrossRef]
- Ramstead MJD, Badcock PB, Friston KJ. 2018 Answering Schrödinger's question: A free-energy formulation. Phys. Life Rev. 24, 1–16. [CrossRef]
- Kauffman SA. 2011 Approaches to the origin of life on the Earth. Life (Basel) 1, 34–48. [CrossRef]
- Robertson MP, Joyce GF. 2012 The origins of the RNA world. Cold Spring Harb. Perspect. Biol. 4, a003608. [CrossRef]
- Chen JM, Chen JW, Zivieri R. 2024 Systematically challenging three prevailing notions about entropy and life. Qeios. [CrossRef]
- Schreiber A, Gimbel S. 2010 Evolution and the second law of thermodynamics: Effectively communicating to non-technicians. Evo. Edu. Outreach 3, 99–106. [CrossRef]
- Chen JM, Chen JW. 2000 Root of science—the driving force and mechanisms of the extensive evolution.Beijing, China: Science Press.
- Ma S, Guo Y, Liu D, et al. 2023 Genome-wide analysis of the membrane attack complex and perforin genes and their expression pattern under stress in the Solanaceae. Int. J. Mol. Sci. 24, 13193. [CrossRef]
- Li Z, Liu X, Wang C, et al. 2023 The pig pangenome provides insights into the roles of coding structural variations in genetic diversity and adaptation. Genome Res. 33, 1833–1847. [CrossRef]
- Wang Y, Li X, Feng Y. 2023 Autotetraploid origin of Chinese cherry revealed by chromosomal karyotype and in situ hybridization of seedling progenies. Plants (Basel) 12, 3116. [CrossRef]
- Fitzgerald DM, Rosenberg SM. 2019 What is mutation? A chapter in the series: How microbes “jeopardize” the modern synthesis. PLoS Genet. 15, e1007995. [CrossRef]
- Olivieri DN, Mirete-Bachiller S, Gambón-Deza F. 2021 Insights into the evolution of IG genes in amphibians and reptiles. Dev. Comp. Immunol. 114, 103868. [CrossRef]
- Sabarís G, Fitz-James MH, Cavalli G. 2023 Epigenetic inheritance in adaptive evolution. Ann. N. Y. Acad. Sci. 1524, 22–29. [CrossRef]
- Gómez-Schiavon M, Buchler NE. 2019 Epigenetic switching as a strategy for quick adaptation while attenuating biochemical noise. PLoS Comput. Biol. 15: e1007364. [CrossRef]
- McNeill JR, Pomeranz K. 2015 The Cambridge World History. Cambridge, UK: Cambridge University Press.
- Eldredge N, Pievani T, Serrelli E, Tëmkin I. 2016 Evolutionary theory: A hierarchical perspective. Chicago, USA: The University of Chicago Press.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



